Содержание
Вопрос 1. 3
Вопрос 2. 5
Вопрос 3. 8
Вопрос 4. 12
Вопрос 5. 13
Вопрос 6. 16
Список литературы.. 17
Вопрос 1
Охарактеризовать вторичные изменения химического состава и свойств оболочек и объяснить биологическое значение этих процессов.
Для многих клеток отложение новых слоев оболочки прекращается с прекращением роста клетки. У других клеток отложение оболочки изнутри продолжается и по достижении окончательного размера. При этом толщина клеточной стенки увеличивается за счет наложения, а объем полости клетки сокращается. Такой процесс носит название вторичного утолщения клеточной стенки, а сама оболочка называется вторичной[1]. Вторичная оболочка выполняет главным образом механическую функцию. Наиболее обычна она в клетках опорных тканей. Химический состав вторичной оболочки иной, чем у первичной. В ней содержится меньше воды, а количество целлюлозы достигает 40-50 % от массы сухого вещества.
Вторичная оболочка иногда откладывается неравномерно. У части сосудов она имеет вид отдельных колец или спиралей. Это позволяет клеткам сохранить способность к растяжению в длину. Стенки клеток, имеющие вторичные утолщения, часто одревесневают вследствие отложения в их матриксе лигнина.
Виды вторичного изменения клеточной оболочки[2]
1. Состав клеточной оболочки молодых растущих клеток, как правило, целлюлозно-пектиновый (процент целлюлозы варьирует). Такие оболочки очень прочны и эластичны за счет распределения микро- и макрофибрилл клетчатки.
Клетчатка как запасное вещество (ее используют в пищу только некоторые крупные животные) не используется растением для жизнедеятельности, в запасе может откладываться полуклетчатка или гемицеллюлоза. Пример - наличие ее в плодах фиников под кожицей. Целлюлозно-пектиновые клеточные оболочки не изменяются в крепких кислотах, даже в «Царской водке». Обнаружить такого состава клеточные оболочки можно специальным реактивом — хлор-цинк-йод.
Хлористый цинк переводит клетчатку в амилоидное состояние, не меняя ее молекулярного состава, а затем йод окрашивает клетчатку (как и все другие углеводы) в грязно-синий цвет.
Растения, имеющие в большом количестве ткани с преобладанием клетчатковых оболочек, относят к текстильным (прядильным): волоски в коробочках хлопчатника, стебли льна, кенафа, кендыря, рами, конопли, крапивы и т. д. Лучшие из них – хлопчатник, лен.
2. Одревеснение - наиболее распространенное видоизменение клеточной оболочки. На первичную клетчатковую оболочку откладывается лигнин - это стойкое вещество придает оболочке большую прочность, но хрупкость на излом.
Лигнин может откладываться на первичную оболочку с внутренней части клетки ровными слоями (путем аппозиции) или внедряться между мецеллами клетчатки первичной оболочки (путем интуссепции), формируя вторичную оболочку.
Клетки с одревесневшими (лигниновыми) оболочками составляют ткани, несущие в растении основную опорную функцию.
В таких вторичных оболочках могут быть простые щелевидные или ветвистые поровые каналы.
Характерный реактив на одревесневшую оболочку флороглюцин (таннид — дубильное вещество из бурых водорослей) плюс концентрированная соляная (или серная) кислота. Оболочки приобретают красновато-малиновый цвет.
Сернокислый анилин одревесневшие оболочки окрашивает в ярко-лимонный цвет.
3. Опробковение наблюдается в большинстве случаев у древесных растений, эти клетки входят в комплекс тканей, выполняющих защитную функцию.
Клетка продуцирует суберин на внутреннюю первичную оболочку, занимая всю ее полость. Содержимое клетки быстро отмирает, она становится целиком непроницаема для газа и воды.
Опробковевшие клетки, в отличие от одревесневших, не имеют поровых каналов, плотно прилегают друг к другу, изолируют внутренние части органа от внешних условий.
Характерные реакции на опробковение:
а) судан III окрашивает в ярко-коричневый цвет;
б) фенол - в грязно-серый.
Вопрос 2
Распределение в теле растений меристем, их цитологическая характеристика и функции.
Меристемы (от греч. Meristos - делимый), или образовательные ткани, обладают способностью к активному делению и образованию новых клеток, занимая ничтожный объем в теле растения (примерно 0,1 % общей массы). Меристемы формируют все прочие ткани и определяют длительный (в течение всей жизни) рост растения. Архитектура размещения тканей всего растения устанавливается на ранних этапах меристематической деятельности. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Инициальные клетки меристем задерживаются на эмбриональной стадии развития в течение всей жизни растения, а их производные постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Тело наземных растений производное относительно немногих инициальных клеток.
Существуют два основных типа меристем — апикальные, или верхушечные, и латеральные, или боковые. Апикальные меристемы располагаются на верхушках побегов и корней (рис. 1), обеспечивая нарастание их в длину. Такой рост получил название первичного, а сами меристемы — первичных. При этом часть растения, образованная первичными тканями, возникшими из первичных меристем, — это его первичное тело.
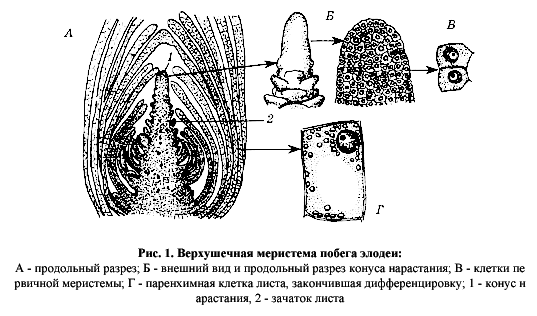
 К
первичным меристемам, помимо апикальных, относят и их непосредственные
производные, несколько отстоящие от верхушек органов. У этих производных
способность к делению в определенной степени сохранена. Речь идет прежде
всего о протодерме, прокамбии и основной меристеме. В первичном теле растения
они дают еще в ходе эмбриогенеза три первичные системы тканей — покровную (из
протодермы), проводящую (из прокамбия) и систему основных тканей (из основной
меристемы).
К
первичным меристемам, помимо апикальных, относят и их непосредственные
производные, несколько отстоящие от верхушек органов. У этих производных
способность к делению в определенной степени сохранена. Речь идет прежде
всего о протодерме, прокамбии и основной меристеме. В первичном теле растения
они дают еще в ходе эмбриогенеза три первичные системы тканей — покровную (из
протодермы), проводящую (из прокамбия) и систему основных тканей (из основной
меристемы).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, нередко образуя цилиндры, на поперечных срезах имеющие вид колец. Главнейшие латеральные меристемы — камбий и феллоген. Эти меристемы обеспечивают нарастание стеблей и стволов в толщину, образуя вторичные ткани и формируя вторичное тело растения. Камбий дает начало вторичным проводящим тканям — вторичным ксилеме и флоэме (рис. 2), а феллоген — перидерме. Помимо апикальных и латеральных меристем, нередко встречаются меристемы еще двух типов.
Интеркалярные, или вставочные, меристемы чаще всего первичны и сохраняются в виде отдельных участков в зонах активного роста, например у оснований междоузлий и в основаниях листьев злаков.
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллусу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающих место поражения. Каллусообразовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллусообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержащие мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым Микроскопом обычно незаметны. Оргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало, и они мелкие.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуются парехимные элементы проводящих тканей, а из прозенхимных — проводящие элементы (рис. 31).
В целом же расположение клеток и характер их деления в разных меристемах различен.
Если межклеточная перегородка закладывается под прямым углом к поверхности органа, такое деление называется антиклинальным. Путем антиклинального деления формируются меристемы пластинчатого
типа, которые образуют структуры, форма которых подобна пластинке. В частности, таким образом формируются плоские листовые пластинки покрытосемянных.
Деления, при которых образующиеся между клетками перегородки параллельны ближайшей поверхности органа, — периклинальные. Они особенно характерны для латеральных меристем. За счет таких делений чаще всего и происходит утолщение осевых органов. Деление с заложением перегородки касательно окружности (тангентально) определяют как тангенциальное. Клеточные деления при этом происходят, по сути, во всех плоскостях, и в результате образуется массив меристемы, напоминающий сфероид. Из такого меристематического массива, в частности, образуется эндосперм семян.
В процессе дифференцировки клеток меристем в постоянные ткани возможны два основных типа роста. Первый — симпластический, при котором рост оболочек смежных клеток происходит согласованно и их связь через плазмодесмы не нарушается. Такой тип обычен в органах удлиняющихся в период первичного роста. Второй тип роста — интрузивный, или интерпозиционный. Он характеризуется отсутствием согласованности в росте отдельных клеток и внедрением одних клеток в пространство, образовавшееся между другими клетками. Так растут камбиальные инициали, склеренхимные волокна, трахеиды и т.д.
Вопрос 3
Дать сравнительную характеристику первичного строения стебля однодольного и двудольного растений, сопроводив текст рисунками.
|
Стебель двудольных растений (первичное строение) |
Стебель однодольных растений |
|
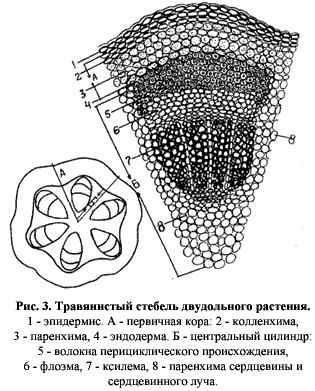
Верхушечная меристема побега двудольных довольно рано дифференцируется на несколько групп клеток (рис. 3)[3]. 1. Протодерма – самая поверхностная. Из нее в дальнейшем формируется первичная покровная ткань – эпидерма. 2. Первичная кора. 3. Инициальное кольцо – оно затем преобразуется в прокамбий или в прокамбий и перицикл (образуется из периферической части) – они дают начало центральному цилиндру. 4. Сердцевина. |
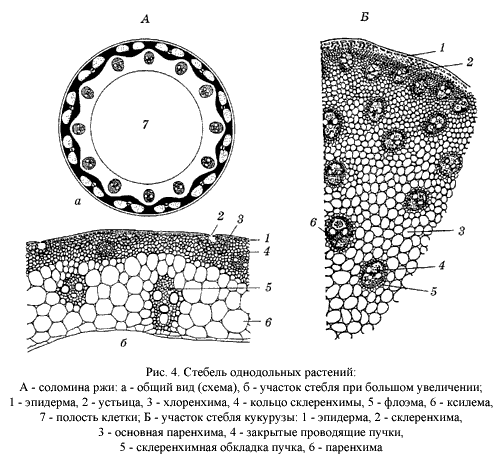
Верхушечная меристема побега однодольных не дифференцируется на первичную кору, центральный цилиндр и сердцевину, инициальное кольцо отсутствует (рис. 4). |
|
Проводящая система стебля формируется из прокамбия, из которого на периферии образуется первичная флоэма (развивается центростремительно), а ближе к центру – первичная ксилема (развивается центробежно), следовательно, образующиеся проводящие пучки будут открытыми (также прокамбий может закладываться в виде сплошного кольца, а не пучков – тогда радиальные паренхимные лучи между пучками будут отсутствовать). |
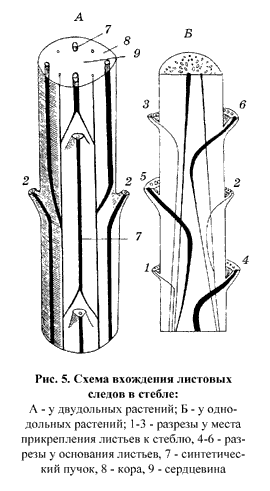
Проводящая система стебля формируется за счет проводящей системы листьев, поэтому все пучки стебля – это индивидуальные листовые следы. В силу особенностей вхождения листовых следов в стебель на поперечном срезе они расположены неупорядоченно. Пучки друг с другом не сливаются (см. рис. 5). Проводящие пучки закрытые. |
|
Тип стелы – эустела (непучкового или пучкового строения) (рис. 6). |
Тип стелы – атактостела (рис. 6). |
|
Утолщение стебля двудольных связано с деятельностью вторичных меристем, но в некоторых случаях наблюдаются первичные утолщения, называемые ростом усиления. Он реализуется за счет изменений в сердцевине (медуллярное утолщение) и отчасти в первичной коре (кортикальное утолщение) и состоит в периклинарных делениях и растяжении клеток (пример – кактусы, клубни картофеля). |
Утолщение стебля всегда носит характер роста усиления и осуществляется не за счет камбия, а благодаря образованию дополнительных паренхимных клеток по периферии стебля, возникающих в результате активности особых внепучковых периферических меристем (меристемы первичного утолщения). |

Вопрос 4
Рассмотреть процессы мегаспорогенеза и формирования женского гаметофита цветковых растений.
 Мегаспорогенез
осуществляется в мегаспорангии, которым является часть семязачатка, освобожденная
от покровов (интегументов). У цветковых растений в мегаспорангии из клеток
археспория обычно закладывается одна материнская клетка мегаспоры, имеющая,
как и все растение, диплоидный набор хромосом. Затем путем мейотического
деления из нее возникает тетрада гаплоидных мегаспор. Чаще всего мегаспоры
располагаются линейно. Обычно нижняя, находящаяся ближе кхалазе, увеличивается
в размерах и функционирует, а остальные отмирают. Из функционирующей мегаспоры
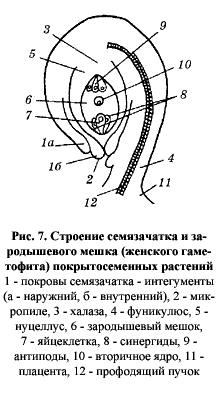
формируется женский гаметофит, называемый зародышевым мешком (рис. 7).
Мегаспорогенез
осуществляется в мегаспорангии, которым является часть семязачатка, освобожденная
от покровов (интегументов). У цветковых растений в мегаспорангии из клеток
археспория обычно закладывается одна материнская клетка мегаспоры, имеющая,
как и все растение, диплоидный набор хромосом. Затем путем мейотического
деления из нее возникает тетрада гаплоидных мегаспор. Чаще всего мегаспоры
располагаются линейно. Обычно нижняя, находящаяся ближе кхалазе, увеличивается
в размерах и функционирует, а остальные отмирают. Из функционирующей мегаспоры
формируется женский гаметофит, называемый зародышевым мешком (рис. 7).
Формирование зародышевого мешка осуществляется путем трех последовательных митотических делений. В результате первого деления мегаспоры образуются два ядра, которые расходятся к полюсам сильно удлиняющейся мегаспоры, а между ними появляется крупная вакуоль. Далее каждое из этих ядер синхронно делится еще два раза; в результате у каждого полюса возникают по четыре ядра. На этой восьмиядерной стадии процесс деления женского гаметофита заканчивается. От каждой из двух четверок ядер в центральную часть зародышевого мешка отходит по одному ядру. Эти так называемые полярные ядра сближаются в центральной части и обычно сливаются, образуя диплоидное вторичное ядро. Далее в зародышевом мешке вокруг каждого из ядер формируются клеточные оболочки. У микропилярного полюса гаметофита образуются три клетки яйцевого аппарата, одна из которых (более крупная) является яйцеклеткой, а две другие — вспомогательными клетками-синергидами. Три клетки халазального полюса получили название антипод. Так возникает наиболее распространенный тип зародышвого мешка, который сильно редуцирован по сравнению с гаметофитом голосемянных. У покрытосемянных нет архегониев.[4]
Вопрос 5
Дать характеристику апокарпных плодов, иллюстрируя текст рисунками.
Плоды-апокарпии образуются из цветков, имеющих апокарпный гинецей. Каждому отдельному, свободно сидящему на цветоножке простому пестику в зрелом плоде соответствует свободный плодик. Апокарпии всегда возникают из цветков с верхней завязью. Эволюционно апокарпии — по-видимому, наиболее архаичные плоды. Главные морфологические типы апокарпиев показаны на рис. 8 на стр. 15. Среди вскрывающихся апокарпиев следует упомянуть многолистовку, а среди невскрывающихся — многоорешек, разновидностями которого являются цинародий (плод шиповника) и земляничина, или фрага. К сочным апокарпиям относится много костянка. Иногда встречается и сочная многолистовка.
Многолистовки образованы двумя-многими, обычно сухими плодиками-листовками, вскрывающимися по брюшному шву. Довольно редкий тип плода — сочная многолистовка, — как правило, не вскрывается, но на брюшной стороне ее плодиков явственно виден шов от срастания краев плодолистиков. Плоды-многолистовки довольно обычны для примитивных магнолиид, розид, дилле-ниид и ранункулид. К ним относятся плоды пионов (Paeonia), купальницы (Trollius), калужницы (Caltha), магнолии (Magnolia).
У лимонника китайского {Schi-sandra chinensis) — дальневосточной лианы — плод — сочная многолистовка. При созревании плода лимонника коническое цветоложе, усаженное свободными пестиками, начинает удлиняться, в результате чего образуется подобие веточки, на которой сидят красные «ягоды», каждая из них — сочный плодик-листовка.
Многоорешек всегда бывает сухим и отличается от многолистовки невскрывающимися односемянными плодиками-орешками. Классический пример многоорешка — плоды видов лютиков (Ranunculus), а также адонисов (Adonis), лапчаток (Potentilla) и др. Плод лотоса орехоносного (Nelumbo nucifera) называется погруженным многоорешком. Каждый из отдельных орешков сидит в углублении дисковидного губчатого разросшегося цветоложа. Другое видоизменение многоорешка — земляничина, или фрага. У этого многоорешка плодики сидят на мясистом разросшемся цветоложе, хорошо всем известном под названием ягод земляники и клубники (виды рода Fragaria). Наконец, цинародий — многоорешек, плодики которого сидят внутри кувшинчатого сочного гипантия, хорошо знакомого на примере плодов шиповника (виды рода Rosa). Плоды видов рода рубус (Rubus) — малина, ежевика, костяника, морошка — имеют плод многокостянку. Многокостянка — это апокарпий, состоящий из двух-многих плодиков — костянок. Мезокарпий каждого такого плодика сочный, а эндокарпий — твердый, склерифицированный[5].

Вопрос 6
Рассмотреть особенности насекомоядных растений местной флоры в связи с их способом питания.
Семейство росянковые — Droseraceae включает 4 рода и около 100 видов. Преимущественно это небольшие, наземные, розеткообразующие болотные травы, распространенные почти по всему миру с наибольшим разнообразием в Австралии и Новой Зеландии. Встречаются среди росянковых и небольшие полукустарники, например обитающий в Испании, Португалии и Марокко росолист лузитанский (Drosophyllum lusitanicum), а один из представителей семейства — альдрованда пузырчатая (Aldrovanda vesiculosa) — свободноплавающее погруженно-водное растение. Этот редчайщий вид, обитающий обычно в крупных стоячих или медленно текущих хорошо прогреваемых водоемах, можно встретить и у нас в европейской части России и на Дальнем Востоке. Несравнимо более обычны росянки круглолистная (Drosera rotundifolia, рис. 277) и английская (D. anglica), являющиеся непременным элементом верховых сфагновых болот севера Евразии. Характерной чертой росянковых является переход их к на-секомоядности. У росянок и росоли-ста листья несут клейкие железистые волоски, к которым и прилипают мелкие насекомые. Затем лист обычно изгибается так, что многочисленные соседние волоски окружают жертву и переваривают ее своими выделениями. Их мелкие, акти-номорфные, обоеполые цветки с двойным пятичленным околоцветником обычно собраны в конечные колосовидные соцветия. Ценокарпный гинецей образован 3 или 5 плодолистиками. Завязь верхняя со свободными столбиками. Плод — коробочка, а очень мелкие семена распространяются обычно водой.
Список литературы
1. Ботанико-фармакогностический словарь / Под ред. Блиновой К. – М.: Высшая школа, 1990. – 272 с.
2. Гладкова З.И. Анатомия вегетативных органов покрытосеменных. – Новосибирск: НГУ, 2000. – 35 с.
3. Гладкова З.И. Клетка и ткани покрытосеменных. Методическое пособие. – Новосибирск: НГУ, 2002. – 36 с.
4. Челомбитько В.А. Ботаника. Учебник для вузов. – СПб.: СпецЛит, 2001. – 448 с.
5. Яковлев Г.П.. Учебник для вузов. – СПб.: СпецЛит, 2001. – 647 с.
[1] Яковлев Г.П. Челомбитько В.А. Ботаника. Учебник для вузов. – СПб.: СпецЛит, 2001. – с. 67.
[2] Гладкова З.И. Клетка и ткани покрытосеменных. Методическое пособие. – Новосибирск: НГУ, 2002. – с. 11-15.
[3] Гладкова З.И. Анатомия вегетативных органов покрытосеменных. – Новосибирск: НГУ, 2000. – с. 21.
[4] Яковлев Г.П. Челомбитько В.А. Ботаника. Учебник для вузов. – СПб.: СпецЛит, 2001. – с. 198-199.
[5] Яковлев Г.П. Челомбитько В.А. Ботаника. Учебник для вузов. – СПб.: СпецЛит, 2001. – с. 216.