Содержание
3.
Клеточные органоиды, их строение и роль. 3
21) Закон
независимого расщепления признака и его цитологическая основа 9
34)
Хромосомные болезни человека, связанные с нерасхождением половых хромосом,
методы их диагностики. 10
Графическая
задача. 12
Заполнение
таблицы.. 14
Задача. 15
Список
литературы.. 16
3. Клеточные органоиды,
их строение и роль
Органоиды
клетки –
постоянно присутствующие в цитоплазме структуры, выполняющие определенные
функции в клетке. Органоиды общего значения (митохондрии,рибосомы,
эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, цитоскелет и
клеточный центр) имеются в любых клетках и их присутствие необходимо для жизни
клетки. Специальные органоиды (реснички,жгутики, микроворсинки, миофибриллы, акросомы)
есть лишь в некоторых клетках и необходимы для выполнения ими
специализированных функций. Специальные органоиды образуются как производные
органоидов общего значения. Органоиды клетки также подразделяются на мембранные
(окружены мембраной, состоящей из липидного двойного слоя, содержащего белки) и
немембранные (мембрана отсутствует).
Синтетический
аппарат клетки (рибосомы, эндоплазматическая сеть, комплекс Гольджи)
располагается в цитоплазме и включает органеллы, участвующие в процессах
синтеза различных веществ. Рибосомы – немембранные
органеллы, диаметр 15-30 нм, состоят из двух субъединиц, в состав которых
входят молекулы более чем 80 белков и четыре молекулы рибосомной
РНК, обеспечивают синтез белка по матрице информационной РНК. Рибосомы могут
находиться непосредственно в цитоплазме, в свободном состоянии или могут быть
прикреплены к мембранам ЭПС (эндоплазматической сети). Сборка рибосом из двух
субъединиц происходит в начале синтеза белка, а по окончании синтеза рибосома
обратимо диссоциирует на большую и малую субъединицы[1].
Эндоплазматическая
сеть –
мембранный органоид, синтезирующий углеводы, липиды и белки; также в ЭПС
происходят некоторые посттрансляционные изменения
белков. ЭПС – система уплощенных трубочек и пузырьков, образующих в цитоплазме
непрерывную трехмерную сеть. Мембрана ЭПС содержит больше белка, чем
плазмолемма, потому что мембрана ЭПС содержит многочисленные ферментные
системы. Шероховатая ЭПС – ЭПС, на мембране которой находятся рибосомы. Функция
шероховатой ЭПС – синтез всех мембранных белков и белков, экспортируемых
клеткой, гликозилирование и посттрансляционные
изменения белковых молекул. В полости ЭПС находятся продукты синтеза. Гладкая
ЭПС – система трубочек,цистерн и пузырьков, на
поверхности которых нет рибосом. Её функции – синтез липидов, гликогена,холестерина, накопление ионов Ca2+.
Комплекс Гольджи – мембранный органоид, образованный стопкой уплощенных
цистерн, пузырьками, вакуолями. Цистерны – это изогнутые диски диаметром до 5
мкм, образующие стопки из 3-30 элементов. Выпуклой стороной стопка обычно
обращена к ядру. Комплекс Гольджи осуществляет синтез
полисахаридов и гликопротеинов, включает углеводные компоненты в гликопротеины
из ЭПС, осуществляет фосфорилирование, ацилирование и сульфатирование
белковых молекул. Также в мембрану комплекса Гольджи
упаковываются секреторные продукты и в процессе секреции эта мембрана
встраивается в плазмолемму. Осуществляется транспорт белков из комплекса Гольджи, причем направление транспорта определяется
особенностями гликозилирования, фосфорилирования
и сульфатирования данного белка. Сортировка
происходит с помощью специфических мембранных рецепторных белков, которые
распознают сигнальные участки макромолекул и направляют их в соответствующие
пузырьки: 1) в гидролазные пузырьки, 2) в окаймленные
пузырьки и затем в плазмолемму, 3) в секреторные гранулы[2].
Аппарат
внутриклеточного переваривания – система мембранных пузырьков с низким
внутренним pH (эндосомы) и лизосом,
которые регулируют расщепление клеткой макромолекул внутри- и внеклеточного
происхождения. Эндосомы переносят макромолекулы с
поверхности клетки в лизосомы и во время переноса частично или полностью их гидролизуют.
Энергетический
аппарат клетки. Митохондрии – полуавтономные мембранные органеллы, обеспечивающие клетку
энергией,запасаемой в виде фосфатных связей АТФ.
Митохондрии синтезируют стероиды, окисляют жирные кислоты и участвуют в синтезе
нуклеиновых кислот. Митохондрии имеют эллиптическую, сферическую, палочковидную
или другую форму и состоят из наружной и внутренней мембран,разделенных
межмембранным пространством, содержат митохондриальный
матрикс, в который обращены складки внутренней мембраны – кристы.
В митохондриальном матриксе находятся митохондриальные рибосомы, митохондриальная
ДНК, обеспечивающая синтез 5-6% митохондриальных белков,в основном эти белки – ферменты электронтранспортной
цепи и некоторые ферменты синтеза АТФ. Митохондрии живут около 10 суток,после чего разрушаются путем аутофагии,
а новые митохондрии формируются путем перешнуровки существующих.
Хлоропласты. У растений фотосинтез
протекает в специализированных внутриклеточных органеллах - хлоропластах.
Хлоропласты
доставляют энергию для метаболизма только в дневные часы; ночью они прекращают
синтез высокоэнергетических метаболитов, и в это время суток растения получают
АТР в результате деятельности митохондрий, которые у них очень сходны с
митохондриями животных клеток.
Хлоропласты
содержат три мембраны—наружную, внутреннюю и тилакоидную,
которые делят органеллу на три внутренних компартмента:
межмембранное пространство, строму и тилакоидное
пространство. В тилакоидной мембране находятся все
энергетические системы хлоропласта. Наружная мембрана высокопроницаемая, в
отличие от внутренней, в которую встроены специальные транспортные белки. Эти
две мембраны разделены узким межмембранным пространством. Внутренняя мембрана
окружает большую центральную область—так называемую строму, представляющую
собой аналог митохондриального матрикса и содержащую
разнообразные ферменты, рибосомы, РНК и ДНК. Внутренняя мембрана хлоропластов
не образует крист и не содержит в себе цепи переноса
электронов, в отличие от соответствующей мембраны митохондрий.
Фотосинтезирующая поглощающая свет система, электрон-транспортная цепь и АТР-синтетаза находятся в третьей мембране.
Гены
хлоропластов наиболее изучены у растений и зеленых водорослей, у которых эти
органеллы очень сходны. Геном хлоропласта представляет собой кольцевую молекулу
ДНК; что гены хлоропластов этих очень отдаленно родственных высших растений
практически идентичны. Помимо четырех рибосомных РНК
эти геномы кодируют около 20 рибосомных белков,
некоторые субъединицы хлоропластной РНК-полимеразы, несколько белков, входящих в состав фотосистем I и II, субъединицы АТР-синтетазы,
части ферментных комплексов электрон-транспортной цепи, одну из двух субъединиц
рибулозобисфосфат-карбоксилазы и 30 тРНК . Все известные белки, кодируемые в хлоропластах,
входят в состав больших ферментных комплексов, которые содержат также одну или
несколько субъединиц, кодируемых ядерным геномом. Геномы хлоропластов и
бактерий сходны по своей организации.
Цитоскелет – система микротрубочек, микрофиламентов,
промежуточных филаментов. Все эти органоиды немембранные. Микротрубочки состоят из димеров
белков α-тубулина и β-тубулина, представляют
собой полые цилиндры диаметром 24-25 нм, в поперечном сечении которых находятся
13 субъединиц белка тубулина. Функции микротрубочек:
поддержание формы клетки и распределения ее компонентов, обеспечение
внутриклеточного транспорта, движения ресничек и хромосом в митозе, образование
основы центриолей и ресничек. Микротрубочки могут располагаться в цитоплазме 1)
отдельно, 2) в пучках, где они связаны тонкими поперечными мостиками или 3)
частично сливаясь друг с другом с формированием дублетов ( в аксонеме ресничек и жгутиков) или триплетов ( в базальном
тельце и центриоли). Обычно один конец микротрубочки закреплен, а на втором
конце происходит полимеризация/деполимеризация микротрубочки.
Микрофиламенты – спирально закрученные двойные белковые нити
диаметром 5-7 нм, состоящие из белка актина расположенные в цитоплазме
одиночно, в виде сети или пучками. Для большинства клеток характерна
кортикальная сеть – зона сгущения микрофиламентов под
плазмолеммой. Функция кортикальной сети – предотвращение резкой деформации
клетки при механических воздействиях и осуществление постепенного изменения
формы клетки. Актин из своей мономерной формы в
присутствии цАМФ и Ca2+ полимеризуется
в фибриллярный актин. Актиновые филаменты
образуют основу микроворсинок.
Промежуточные
филаменты—белковые нити диаметра 10 нм. Располагаются в
клетке в виде трехмерных сетей в цитоплазме, окружают ядро,входят
в состав десмосом и полудесмосом. Основная функция –
структурная и опорная, обеспечение
распределения органелл в цитоплазме. Подразделяются на классы—кератиновые, десминовые, виментиновые, нейрофиламенты, глиальные филаменты и ламины (образуют кариоскелет).
Ядро
клетки—самый
крупный органоид клетки, оно содержит генетический аппарат клетки – всю
хромосомную ДНК, которая упакована в нити хроматина благодаря ассоциации с
белками-гистонами. Функции ядра—хранение генетической информации в молекулах
ДНК в хромосомах, реализация генетической информации, воспроизведение и
передача генетической информации. В ядре интерфазной
клетки выделяются ядерная оболочка, хроматин, ядрышко и кариоплазма. Кариолемма состоит из двух мембран, разделенных перинуклеарным пространством и смыкающихся в области
ядерных пор, через которые содержимое ядра сообщается с цитоплазмой. Наружная
мембрана является продолжением мембраны гранулярной ЭПС, на ее поверхности
находятся рибосомы. Внутренняя мембрана гладкая, ее интегральные белки связаны
с ламиной—слоем промежуточных филаментов,
образующих кариоскелет. Ламина
поддерживает форму ядра, участвует в упорядоченной укладке хроматина,
организации поровых комплексов,формировании кариолеммы при делении клеток.
Ядрышко—видимая в световом
микроскопе структура интерфазного ядра, состоящая из
генов, кодирующих рРНК, молекул рРНК
и их предшественников, ферментов, участвующих в процессинге
рРНК, ядрышковых
рибонуклеопротеидных комплексов, белков рибосомных
субъединиц и субчастиц рибосом. Расположение всех
этих компонентов в пределах ядрышка обеспечивает быструю и точную сборку
рибосом. В диплоидных клетках человека гены рРНК
собраны в 10 кластеров, каждый из которых расположен в приконцевом
районе одной из пяти пар различных хромосом. Во время митоза ядрышко
разрушается, после митоза концевые районы 10 хромосом объединяются и ядрышко
формируется вновь. Предполагается, что размер ядрышка отражает количество
рибосом, образующихся в клетке. Размер ядрышка различен в разных типах клеток и
может изменяться в одной и той же клетке, достигая 25% от объема ядра. В
ядрышке также происходит процессинг малых и
транспортных РНК и собираются различные РНК-белковые
комплексы[3].
21)
Закон независимого расщепления признака и его цитологическая основа
Закон
независимого расщепления признака - биологический закон, согласно которому гены
одной аллельной пары распределяются в мейозе независимо от генов других пар и
комбинируются в процессе образования гамет случайно, что ведет к разнообразию
вариантов их сочетаний в зиготе.
Цитологическая
основа закона независимого расщепления признака — то все процессы, происходящие
в клетках при гаметогенезе во время мейоза и сочетание новых комбинаций генов в
зиготе при оплодотворении. Условием для выполнения закона о независимом
наследовании признака и расщепления является расположение аллельных генов
разных гомологичных хромосомах, иначе речь идет о сцепленном наследовании[4].
34)
Хромосомные болезни человека, связанные с нерасхождением половых хромосом,
методы их диагностики
Анеуплоидия по половым хромосомам или по аутосомам
возникает в результате неправильного расхождения хромосом в мейозе. При слиянии
нормальной гаметы с Х-хромосомой с гаметой, где половая хромосома отсутствует,
получается зигота, из которой развивается девочка с синдромом
Шерешевского-Тернера. Синдром Клайнфельтера может
возникать при слиянии нормальной гаметы с У-хромосомой
и гаметы с двумя Х-хромосомами, также как и при слиянии гаметы типа ХУ с
нормальной гаметой Х. Неправильное расхождение хромосом может происходить и в
митозе. В результате возникают мозаики—особи, у которых не все клетки несут
одинаковый генотип. Степень мозаицизма определяется
тем, при каком по счету клеточном делении зиготы произошло неправильное
расхождение хромосом. Среди носителей мозаичных фенотипов чаще всего
встречаются люди с клетками типов ХУ и ХУУ. Фенотипическое отклонение от нормы
в этих случаях зависит от стадии развития, на которой произошло неправильное
расхождение хромосом[5].
При
анеуплоидии по половой хромосоме (наличии в клетках одной Х-хромосомы при
отсутствии Y-хромосомы) у человека развивается синдром
Шерешевского-Тернера. Впервые синдром описан в 1925 г. Шерешевским,
подробнее был изучен Тернером в 1938г. Это единственный у человека случай моносомии. Страдающие синдромом Тернера стерильны. Фенотипически это женщины с почти атрофированными яичниками
и слабо развитыми вторичными половыми признаками. Другими характерными
признаками этого синдрома являются низкий рост, деформация грудной клетки,
крыловидная шея. Синдром Тернера обычно не сопровождается умственной
неполноценностью. Встречается это заболевание с частотой примерно 1 на 5000
новорожденных.
С большей
частотой (1:700) встречается наличие одной лишней Х-хромосомы или нескольких
Х-хромосом у женщин. Как правило, это особи с генотипом ХХХ, однако известны
случаи, когда появлялись 2 и более лишних Х-хромосом. Для женщин с трисомией по Х-хромосоме характерны сниженная плодовитость
и умственная отсталость.
При три-,
тетра-, пента- и гексасомии по Х-хромосоме у мужчин
формируется синдром Клайнфельтера. Обычно это
кариотип ХХУ, но встречаются кариотипы ХХХУ и ХХХХУ. Признаки особей с
синдромом Клайнфельтера: высокий рост,несколько
женское телосложение, увеличение пористости костей, тестикулярная
атрофия, увеличение молочных желез. У некоторых носителей кариотипа наблюдается
умственная отсталость, но у большинства коэффициент интеллектуальности в
пределах нормы.
Другой случай
анеуплоидии у человека — трисомия типа ХУУ,
встречается с частотой 1 на 1000. Большинство людей с таким кариотипом — нормальные
мужчины, несколько выше среднего роста. Интересен тот факт, что среди
заключенных и пациентов психиатрических больниц данный кариотип встречается в
20 раз чаще, чем среди населения в целом.
Диагносцировать данные заболевания можно, проанализировав
кариотип пациента и установив наличие дополнительных или отсутствие одной из
Х-хромосом.
Для пренатальной диагностики врожденных заболеваний обычно
используется метод, называемый амниоцентез. На 14-16
неделе беременности с помощью хирургического шприца берется проба амниотический
жидкости объемом 10-15 мл, после центрифугирования пробы выращивают культуру
осажденных клеток плода и проводят их кариотипический
анализ—то есть определяют число хромосом. Надосадочную
жидкость исследуют биохимическими методами, направленными на обнаружение
различных отклонений в метаболизме плода[6].
Графическая задача
33) Изобразите схему метафазы и анафазы
редукционного деления мейоза, если 2n=4(AaBb)
Мейоз—это
процесс деления клеточного ядра с образованием четырех дочерних ядер, каждое из
которых содержит вдвое меньше хромосом, чем исходное ядро. Его называют также
редукционным делением (от лат. reductio-уменьшение): число хромосом в клетке
уменьшается с диплоидного (2n) до гаплоидного (n).
Мейоз включает два последовательных деления клеточного ядра: первое — редукционное
и второе — митотическое деление мейоза, при котором каждое из дочерних ядер
делится митотическим путем, и сестринские хроматиды расходятся. Перед коньюгацией хромосом каждый из гомологов подвергается
удвоению, образуя пару тесно связанных сестринских хроматид, что похоже на
события, происходящие при обычном клеточном делении[7].
Специфические
особенности мейоза проявляются уже после завершения репликации ДНК. Вместо
того, чтобы отделиться друг от друга, сестринские хроматиды ведут себя как одно
целое (как будто дупликация хромосом не произошла): каждый дуплицированный
гомолог коньюгирует с партнером, образуя структуру,
состоящую из четырех хроматид и называется эта структура бивалентом. Бивалент
располагается на экваторе веретена деления, и в анафазе дуплицированные
гомологи (каждый состоит из двух сестринских хроматид) отделяются друг от друга
и расходятся к противоположным полюсам, причем в каждом из них две сестринские
хроматиды остаются соединенными. Таким образом, при первом делении мейоза
каждая дочерняя клетка наследует две копии одного из двух гомологов и поэтому
содержит диплоидное количество ДНК, но гаплоидный набор генов (1n2с).
Отличие от обычных диплоидных клеток состоит в 1) обе копии ДНК каждой хромосомы
происходят лишь от одной из двух гомологичных хромосом, 2) эти две копии клетка
получает в виде тесно связанных сестринских хроматид, составляющих единую
хромосому.
Таким
образом, редукционное деление мейоза состоит из следущих
стадий[8]:
Профаза I—спирализация хромосом, коньюгация
гомологичных хромосом, кроссинговер (обмен участков гомологичных хромосом)
Метафаза I—размещение
хромосом в плоскости экватора (образование метафазной пластинки) и образование
веретена деления, кинетохорные нити которого прикрепляются
к центромерам одним концом, а другим – к центриолям.
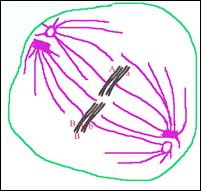 Анафаза I—расхождение
гомологичных хромосом к противоположным полюсам клетки (каждая хромосома
состоит из двух хроматид, хроматиды соединены в районе центромеры),
образуется гаплоидный набор.
Анафаза I—расхождение
гомологичных хромосом к противоположным полюсам клетки (каждая хромосома
состоит из двух хроматид, хроматиды соединены в районе центромеры),
образуется гаплоидный набор.
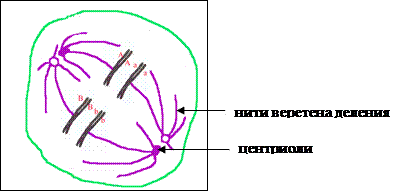
Телофаза I—образование
оболочки ядра и двух новых клеток, каждая из которых впоследствии проходит
второе деление мейоза.
Заполнение
таблицы
Таблица 7) Установите соответствие[9]:
|
1.Малярийный плазмодий
|
б) эукариотическая клетка
|
|
2.Возбудитель туберкулеза
|
в) прокариотическая клетка
|
|
3.Эритроциты человека
|
г) эукариотическая клетка
|
|
4.Возбудитель пневмонии
|
д) прокариотическая
клетка
|
|
5.Возбудитель гриппа
|
а) доклеточная форма жизни
|
Задача
24) В участок полипептида (белка) входит 52 аминокислоты. Сколько
нуклеотидов в составе экзонов гена, кодирующего этот
полипептид?
Перенос информации от мРНК
к белку основан на принципе правильного спаривания комплементарных
оснований, так же как и перенос генетической информации от ДНК к ДНК или РНК.
Однако для того, чтобы правильно расположить молекулы тРНК
на мРНК необходимо участие рибосом. Это рРНК-белковые комплексы, участвующие в трансляции. Рибосома
связывается со специальным участком молекулы мРНК и
таким образом определяет рамку считывания и первую аминокислоту, которая будет
находиться на N-(амино-) конце белка,
после чего рибосома движется по молекуле мРНК и
каждому кодону (триплету) ставит в соответствие одну аминокислоту,
присоединяемую к растущей полипептидной цепи. Достигнув конца кодирующей части
матрицы, рибосома и новосинтезированный карбоксильный
конец белка отсоединяются от 3'-конца мРНК и
переходят в цитоплазму клетки[10].
То есть, 52 аминокислоты соответствуют
52*3=156 нуклеотидов кодирующей части гена. Плюс один стоп-кодон,
который не кодирует аминокислоту, но является сигналом остановки трансляции. В
некоторых случаях первая аминокислота-метионин, кодируемый старт-кодоном
AUG отрезается, тогда в мРНК было 162
нуклеотида и первая аминокислота не метионин. Ответ: 159 нуклеотидов, если
первая аминокислота в полипептиде метионин или 162 нуклеотида, если первая
аминокислота другая, а метионин был удален.
Список литературы
1. В. Л. Быков Цитология и
общая гистология, С-Петербург, СОТИС, 2000г.
2. Б. Албертс
и др. Молекулярная биология клетки, Москва, Мир 1994г.
3. Спирин Структура и биосинтез
нуклеиновых кислот, 1990 г.
4. Б. Льюин
Гены, 1987г.
5. И. Ф. Жимулев
Общая и молекулярная генетика, Новосибирск, 2002 г.
6.
Ф. Айала,
Дж. Кайгер. Современная генетика, 1988г.
7. Догель В. А. Зоология
беспозвоночных. М.: Высшая школа, 1981.
[1] В. Л.
Быков Цитология и общая гистология, С-Петербург, СОТИС, 2000. – с. 67.
[2] Б. Албертс и др. Молекулярная биология клетки, Москва, Мир
1994. – с. 78.
[3] В. Л.
Быков Цитология и общая гистология, С-Петербург, СОТИС, 2000. – с. 89.
[4] И. Ф.
Жимулев Общая и молекулярная генетика, Новосибирск, 2002 г. – с. 76.
[5] Ф. Айала, Дж. Кайгер. Современная генетика, 1988г. – с. 122.
[6] И. Ф.
Жимулев Общая и молекулярная генетика, Новосибирск, 2002 г. – с. 79.
[7] Б. Албертс и др.
Молекулярная биология клетки, Москва, Мир 1994г. – с. 188.
[8] Б. Албертс и др.
Молекулярная биология клетки, Москва, Мир 1994г. – с. 177.
[9] Догель В. А. Зоология беспозвоночных. - М.: Высшая школа,
1981. – с. 170.
[10] Спирин Р.Ж. Структура и
биосинтез нуклеиновых кислот 1990 г. – с. 80.