Содержание
Введение. 3
1. Ферменты - биологические катализаторы: общие положения. 4
2. Классификация ферментов. 5
3. Характеристика лигаз (синтетаз) и механизм их действия. 7
4. Свойства ферментов микробиологического синтеза. 10
5. Строение ферментов микробиологического синтеза. 14
6. Иммобилизованные ферменты.. 19
Заключение. 21
Список литературы.. 22
Введение
В
основе всех жизненных процессов лежат тысячи химических реакций. Они идут в
организме без применения высокой температуры и давления, т. е. в мягких
условиях. Вещества, которые окисляются в клетках человека и животных, сгорают
быстро и эффективно, обогащая организм энергией и строительным материалом. Но
те же вещества могут годами храниться как в консервированном (изолированом от
воздуха) виде, так и на воздухе в присутствие кислорода. Возможность быстрого
переваривания продуктов в живом организме осуществляется благодаря присутствию
в клетках особых биологических катализаторов - ферментов.
Ферменты - это специфические белки,
входящие в состав всех клеток и тканей живых организмов играющие роль
биологических катализаторов. О ферментах люди узнали давно. Еще в начале
прошлого века в Петербурге К.С.Кирхгоф выяснил, что проросший ячмень способен
превращать полисахарид крахмал в дисахарид мальтозу, а экстракт дрожжей
расщеплял свекловичный сахар на моносахариды - глюкозу и фруктозу. Это были
первые исследования в ферментологии. Хотя на практике применение ферментативных
процессов было известно с незапамятных времен (сбраживание винограда,
сыроварение и др.).
Целью данной работы является
исследование ферментов микробиологического синтеза.
Поставленная цель конкретизируется
рядом задач:
1. рассмотреть общие
понятия о ферментах
2. рассмотреть
классификацию ферментов
3. рассмотреть
свойства и механизм функционирования ферментов микробиологического синтеза
4. рассмотреть
свойства лигаз
5. рассмотреть
строение лигаз
6. рассмотреть
иммобилизованные ферменты
1. Ферменты - биологические катализаторы: общие
положения
Ферменты (от лат. fermentum -
брожение, закваска), специфические белки, присутствующие во всех живых клетках
и играющие роль биологических катализаторов. Через их посредство реализуется
генетическая информация и осуществляются все процессы обмена веществ и энергии
в живых организмах. Ферменты бывают простыми или сложными белками, в состав
которых наряду с белковым компонентом (апоферментом) входит небелковая часть -
кофермент. Эффективность действия ферментов определяется значительным снижением
энергии активации катализируемой реакции в результате образования промежуточных
фермент-субстратных комплексов. Присоединение субстратов происходит в активных
центрах, которые обладают сходством только с определенными субстратами, чем
достигается высокая специфичность (избирательность) действия ферментов. Одна из
особенностей ферментов - способность к направленному и регулируемому действию.
За счёт этого контролируется согласованность всех звеньев обмена веществ. Эта
способность определяется пространственность структурной молекулы ферментов. Она
реализуется через изменение скорости действия ферментов и зависит от
концентрации соответствующих субстратов и кофакторов, рH среды, температуры, а
также от присутствия специфических активаторов и ингибиторов (например,
адениловых нуклеотидов, карбонильных, сульфгидрильных соединений и др.).
Некоторые ферменты помимо активных центров имеют дополнительные, т.н.
аллостерические регуляторные центры. Биосинтез ферментов находится под
контролем генов. Различают конститутивные ферменты, постоянно присутствующие в
клетках, и индуцируемые ферменты, биосинтез которых активируется под влиянием
соответствующих субстратов. Некоторые функционально взаимосвязанные ферменты
образуют в клетке структурно организованные полиферментные комплексы. Многие
ферменты и ферментные комплексы прочно связаны с мембранами клетки или её
органоидов (митохондрий, лизосом, микросом и т.д.) и участвуют в активном транспорте веществ
через мембраны[4,с .188].
Известно более 20000 различных ферментов, из которых многие выделены из
живых клеток и получены в индивидуальном состоянии. Первый кристаллический
фермент (уреаза) выделен американским биохимиком Д.Самнером в 1926 г. Для ряда
ферментов изучена последовательность аминокислот и выяснено расположение
полипептидных цепей в трёхмерном пространстве. В лабораторных условиях
осуществлен искусственный химический синтез фермента рибонуклеазы. Ферменты
используют для количественного определения и получения различных веществ, для
модификации молекул нуклеиновых кислот методами генной инженерии, диагностики и
лечения ряда заболеваний, а также в ряде технологических процессов, применяемых
в лёгкой, пищевой и фармацевтической промышленностях.
2. Классификация ферментов
По первой в истории изучения ферментов классификации их делили на две
группы: гидролазы, ускоряющие гидролитические реакции, и десмолазы, ускоряющие
реакции негидролитического распада. Затем была сделана попытка разбить ферменты
на классы по числу субстратов, участвующих в реакции. В соответствии с этим
ферменты классифицировали на три группы[5, с. 156].
1. Катализирующие превращения двух субстратов одновременно в обоих
направлениях: А+В)С+D. 2. Ускоряющие превращения двух субстратов в прямой реакции
и одного в обратной: А+В)С. 3. Обеспечивающие каталитическое видоизменение
одного субстрата как в прямой, так и в обратной реакции: А)В.
Одновременно развивалось направление, где в основу классификации
ферментов был положен тип реакции, подвергающейся каталитическому воздействию.
Наряду с ферментами, ускоряющими реакции гидролиза (гидролазы), были изучены
ферменты, участвующие в реакциях переноса атомов и атомных групп (феразы), в
изомеризации (изомеразы), расщеплении (лиазы), различных синтезах (синтетазы) и
т. д. Это направление в классификации ферментов оказалось наиболее
плодотворным, так как объединяло ферменты в группы не по надуманным, формальным
признакам, а по типу важнейших биохимических процессов, лежащих в основе
жизнедеятельности любого организма. По этому принципу все ферменты делят на 6
классов.
1. Оксидоредуктазы - ускоряют реакции окисления - восстановления. 2.
Трансферазы - ускоряют реакции переноса функциональных групп и молекулярных
остатков. 3. Гидролазы - ускоряют реакции гидролитического распада. 4. Лиазы -
ускоряют негидролитическое отщепление от субстратов определенных групп атомов с
образованием двойной связи (или присоединяют группы атомов по двойной связи).
5. Изомеразы - ускоряют пространственные или структурные перестройки в пределах
одной молекулы. 6. Лигазы - ускоряют реакции синтеза, сопряженные с распадом
богатых энергией связей. Эти классы и положены в основу новой научной
классификации ферментов[3, с. 155].
К классу оксидоредуктаз относят ферменты, катализирующие реакции
окисления - восстановления. Окисление протекает как процесс отнятия атомов Н
(электронов) от субстрата, а восстановление - как присоединение атомов Н
(электронов) к акцептору.
В класс трансфераз входят ферменты, ускоряющие реакции переноса
функциональных групп и молекулярных остатков от одного соединения к другому.
Это один из наиболее обширных классов: он насчитывает около 500 индивидуальных
ферментов. В зависимости от характера переносимых группировок различают
фосфотрансферазы, аминотрансферазы, гликозилтрансферазы, ацилтрансферазы,
трансферазы, переносящие одноуглеродные остатки (метилтрансферазы,
формилтрансферазы), и др. Например, амидазы ускоряют гидролиз амидов кислот. Из
них важную роль в биохимических процессах в организме играют уреаза, аспарагиназа
и глутаминаза.
Уреаза была одним из первых белков-ферментов, полученным в
кристаллическом состоянии. Это однокомпонентный фермент (М=480000), молекула
его глобулярна и состоит из 8 равных субъединиц. Уреаза ускоряет гидролиз
мочевины до NН3 и СО2.
3. Характеристика лигаз (синтетаз) и механизм их действия
Характерные черты действия ферментов класса лигаз (синтетаз) выявлены
совсем недавно в связи со значительными успехами в изучении механизма синтеза
жиров, белков и углеводов: Оказалось, что старые представления об образовании
этих соединений, согласно которым они возникают при обращении реакций
гидролиза, не соответствуют действительности. Пути их синтеза принципиально
иные.
Главная их особенность - сопряженность синтеза с распадом веществ,
способных поставлять энергию для осуществления биосинтетического процесса.
Одним из таких природных соединений является АТФ. При отрыве от ее молекулы в
присутствии лигаз одного или двух концевых остатков фосфорной кислоты
выделяется большое количество энергии, используемой для активирования
реагирующих веществ. Лигазы же каталитически ускоряют синтез органических
соединений из активированных за счет
распада АТФ исходных продуктов. Таким образом, к лигазам
относятся ферменты, катализирующие соединение друг с другом двух молекул,
сопряженное с гидролизом пирофосфатной связи в молекуле АТФ или иного
нуклеозидтрифосфата[1,с .165].
Механизм действия лигаз изучен еще недостаточно, но, несомненно, он
весьма сложен. В ряде случаев доказано, что одно из участвующих в основной
реакции веществ сначала дает промежуточное соединение с фрагментом
распадающейся молекулы АТФ, а вслед за этим указанный промежуточный продукт
взаимодействует со вторым партнером основной химической реакции с образованием
конечного продукта.
Лигазы - это ферменты, катализирующие синтез сложных молекул из более
простых за счет энергии распада аденозинтрифосфата (АТФ) и его аналогов. К
классу лигаз принадлежит большая группа ферментов. Ниже приведены отдельные
наиболее простые реакции, катализируемые лигазами. Например, синтез амидов
аспарагина катализируется ферментом аспарагинсинтетазой:
аспарагинова кислота + АТФ + NН3  аспарагин
+ АДФ + Н3РО4
аспарагин
+ АДФ + Н3РО4
Глутамнисинтетаза катализирует синтез глутамина:
глутаминова кислота + АТФ + NН3 глютамин +
АДФ + Н3РО4
Пируваткарбоксилаза катализирует синтез щавелевоуксусной кислоты
из пировиноградной кислоты и углекислоты:
пировиноградна кислота + АТФ + CO2 щавелевоуксусна
кислота + АДФ +H3PO4
Cинтетазы играют очень важную роль в процессе синтеза белка. Есть
белки, от которых требуется колоссальная точность работы. Это, например,
аминоацил-тРНК синтетазы, от точности работы которых зависит точность синтеза
белков. Поэтому они не могут позволить себе ошибаться чаще, чем однажды на
много тысяч синтезов аминоацил-тРНК, тогда как опознавание аминокислоты
специфическим субстрат-связывающим карманом допускает порядка 1% ошибок. Для
устранения этих ошибок используется эффект "двойного сита" (рис. 1).
Этот эффект обеспечивается специальной конструкцией фермента.
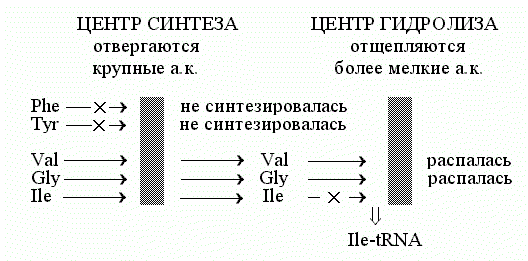
Рис.1. Схема "двойного сита" для механизма
редактирования работы изолейцил-тРНК синтетазы [1, 4 и др.]
У фермента аминоацил-тРНК синтетазы два активных центра: центр синтеза и
центр гидролиза. Причем гидролиз аминоацил-тРНК на аминоацил-тРНК синтетазе не
есть просто реакция, в точности обратная синтезу: обе эти реакции идут с
выделением свободных фосфатов, т.е. обе - с понижением свободной энергии. При
этом субстрат-связывающий карман центра гидролиза меньше, чем карман центра
синтеза аминоацил-тРНК. Принцип сита - не пропускать частицы, большие, чем
ячейки сита. На первом сите при синтезе аминоацил-тРНК, тРНК заряжается
"правильной" аминокислотой и некоторым числом
"неправильных", более мелких (более крупные аминокислоты отвергаются
все, а большая часть "неправильных" мелких все же отвергается по
причине различий в гидрофобности/ гидрофильности, но отбор по гидрофобности/
гидрофильности не столь жесток, как отбор по возможности втиснуться в карман
данного размера). Итак, на выходе этапа синтеза, много аминоацил-тРНК с
"правильной" аминокислотой и некоторое количество аминоацил-тРНК с
"неправильными" аминокислотами.
На втором активном центре, на втором "сите",
субстрат-связывающий карман которого поменьше, идет гидролиз образовавшихся
аминоацил-тРНК, - но только тех, где аминокислота меньше, чем
"правильная". То есть "правильная" аминокислота отвергается
карманом гидролизного центра, этим "вторым ситом", а все остальные
аминоацил-тРНК гидролизуются и распадаются. В сумме, два эти сита и
обеспечивают выход только "правильно заряженных" аминоацил-тРНК.
4. Свойства ферментов микробиологического синтеза
Будучи белками, ферменты обладают всеми их свойствами. Вместе с тем
биокатализаторы характеризуются рядом специфических качеств, тоже вытекающих из
их белковой природы. Эти качества отличают ферменты от катализаторов обычного
типа. Сюда относятся термолабильность ферментов, зависимость их действия от
значения рН среды, специфичность и, наконец, подверженность влиянию активаторов
и ингибиторов.
Термолабильность ферментов объясняется тем, что температура, с одной
стороны, воздействует на белковую часть фермента, приводя при слишком высоких
значениях к денатурации белка и снижению каталитической функции, а с другой
стороны, оказывает влияние на скорость реакции образования фермент-субстратного
комплекса и на все последующие этапы преобразования субстрата, что ведет к
усилению катализа[4, с. 176].
Зависимость каталитической активности фермента от температуры выражается
типичной кривой. До некоторого значения температуры (в среднем до 5О°С)
каталитическая активность растет, причем на каждые 10°С примерно в 2 раза
повышается скорость преобразования субстрата. В то же время постепенно
возрастает количество инактивированного фермента за счет денатурации его
белковой части. При температуре выше 50°С денатурация ферментного белка резко
усиливается и, хотя скорость реакций преобразования субстрата продолжает расти,
активность фермента, выражающаяся количеством превращенного субстрата, падает.
Детальные исследования роста активности ферментов с повышением
температуры, проведенные в последнее время, показали более сложный характер
этой зависимости, чем указано выше: во многих случаях она не отвечает правилу
удвоения активности на каждые 10°С в основном из-за постепенно нарастающих
конформационных изменений в молекуле фермента.
Температура, при которой каталитическая активность фермента максимальна,
называется его температурным оптимумом.
Температурный оптимум для различных ферментов неодинаков. В общем для
ферментов животного происхождения он лежит между 40 и 50°С, а растительного -
между 50 и 60°С. Однако есть ферменты с более высоким температурным оптимумом,
например, у папаина (фермент растительного происхождения, ускоряющий гидролиз
белка) оптимум находится при 8О°С. В то же время у каталазы (фермент,
ускоряющий распад Н2О2 до Н2О и О2) оптимальная температура действия находится
между 0 и -10°С, а при более высоких температурах происходит энергичное
окисление фермента и его инактивация.
Зависимость активности фермента от значения рН среды была установлена
свыше 50 лет назад. Для каждого фермента существует оптимальное значение рН
среды, при котором он проявляет максимальную активность. Большинство ферментов
имеет максимальную активность в зоне рН поблизости от нейтральной точки. В
резко кислой или резко щелочной среде хорошо работают лишь некоторые ферменты.
Переход к большей или меньшей (по сравнению с оптимальной) концентрации
водородных ионов сопровождается более или менее равномерным падением активности
фермента.
Влияние концентрации водородных ионов на каталитическую активность ферментов
состоит в воздействии ее на активный центр. При разных значениях рН в
реакционной среде активный центр может быть слабее или сильнее ионизирован,
больше или меньше экранирован соседними с ним фрагментами полипептидной цепи
белковой части фермента и т.п. Кроме того, рН среды влияет на степень ионизации
субстрата, фермент-субстратного комплекса и продуктов реакции, оказывает
большое влияние на состояние фермента, определяя соотношение в нем катионных и
анионных центров, что сказывается на третичной структуре белковой молекулы.
Последнее обстоятельство заслуживает особого внимания, так как определенная
третичная структура белка-фермента необходима для образования
фермент-субстратного комплекса.
Специфичность - одно из наиболее выдающихся качеств ферментов. Эго
свойство их было открыто еще в прошлом столетии, когда было сделано наблюдение,
что очень близкие по структуре вещества - пространственные изомеры расщепляются
по эфирной связи двумя совершенно разными ферментами.
Таким образом, ферменты могут различать химические соединения,
отличающиеся друг от друга очень незначительными деталями строения, такими,
например, как пространственное расположение метоксильного радикала и атома
водорода при 1-м углеродном атоме молекулы метилглюкозида.
По образному выражению, нередко употребляемому в биохимической
литературе, фермент подходит к субстрату, как ключ к замку. Это знаменитое
правило было сформулировано Э. Фишером в 1894 г. исходя из того, что
специфичность действия фермента предопределяется строгим соответствием
геометрической структуры субстрата и активного центра фермента.
В 50-е годы нашего столетия это статическое представление было заменено
гипотезой Д. Кошланда об индуцированном соответствии субстрата и фермента.
Сущность ее сводится к тому, что пространственное соответствие структуры
субстрата и активного центра фермента создается в момент их взаимодействия друг
с другом, что может быть выряжено формулой “перчатка - рука”. При этом в
субстрате уже деформируются некоторые валентные связи и он, таким образом,
подготавливается к дальнейшему каталитическому видоизменению, а в молекуле
фермента происходят конформационные перестройки. Гипотеза Кошланда, основанная
на допущении гибкости активного центра фермента, удовлетворительно объясняла
активирование и ингибирование действия ферментов и регуляцию их активности при
воздействии различных факторов. В частности, конформационные перестройки в
ферменте в процессе изменения его активности Кошланд сравнивал с колебаниями
паутины, когда в нее попала добыча (субстрат), подчеркивая этим крайнюю
лабильность структуры фермента в процессе каталитического акта.
В настоящее время гипотеза Кошланда постепенно вытесняется гипотезой
топохимического соответствия. Сохраняя основные положения гипотезы
взаимоиндуцированной настройки субстрата и фермента, она фиксирует внимание на
том, что специфичность действия ферментов объясняется в первую очередь
узнаванием той части субстрата, которая не изменяется при катализе. Между этой
частью субстрата и субстратным центром фермента возникают многочисленные
точечные гидрофобные взаимодействия и водородные связи.
Важнейшим свойством ферментов является преимущественное одной из
нескольких теоретически возможных реакций. В зависимости от условий ферменты
способны катализировать как прямую так и обратную реакцию. Это свойство
ферментов имеет большое практическое значение.
Другое важнейшее свойство ферментов - термолабильность, т. е. высокая
чувствительность к изменениям температуры. Так как ферменты являются белками,
то для большенства из них температура свыше 70 C приводит к денатурации и
потере активности. При увелечении температуры до 10 С реакция ускоряется в 2-3
раза, а при температурах близких к 0 С скорость ферментативных реакций
замедляется до минимума.
Следующим важным свойством является то, что ферменты находятся в тканях и
клетках в неактивной форме (проферменте). Классическими его примерами являются
неактивные формы пепсина и трипсина. Существование неактивных форм ферментов
имеет большое биологическое значение. Если бы пепсин вырабатывался сразу в
активной форме, то пепсин "переваривал" стенку желудка, т. е. желудок
"переваривал" сам себя.
5. Строение ферментов микробиологического синтеза
По строению ферменты могут быть однокомпонентными, простыми белками, и
двухкомпонентными, сложными белками. Во втором случае в составе фермента
обнаруживается добавочная группа небелковой природы.
В разное время возникли различные наименования белковой части и
добавочной группы в двухкомпонентных ферментах. Все они до сих пор
употребляются в литературе, например[4]:
Фермент в целом Белковая
часть Добавочная группа
Симплекс Ферон (носитель) Агон
(активная группа)
Холофермент Апофермент Кофермент
Добавочную группу, прочно связанную, не отделяемую от белковой части,
называют простетической группой; в отличие от этого добавочную группу, легко
отделяющуюся от апофермента и способную к самостоятельному существованию,
обычно именуют коферментом.
Химическая природа важнейших коферментов была выяснена в 30-е годы нашего
столетия благодаря трудам О. Варбурга, Р. Куна, П. Каррера и др. Оказалось, что
роль коферментов в двухкомпонентных ферментах играют большинство витаминов (Е,
К, Q, В1, В2, В6 В12, С, Н и др.) или соединений, построенных с участием
витаминов (коэнзим А, НАД+ и т. п.). Кроме того, функцию коферментов выполняют
такие соединения, как НS-глутатион, многочисленная группа нуклеотидов и их
производных, фосфорные эфиры некоторых моносахаридов и ряд других веществ.
Характерной особенностью двухкомпонентных ферментов является то, что ни
белковая часть, ни добавочная группа в отдельности не обладают заметной
каталитической активностью. Только их комплекс проявляет ферментативные
свойства. При этом белок резко повышает каталитическую активность добавочной
группы, присущую ей в свободном состоянии в очень малой степени; добавочная же
группа стабилизирует белковую часть и делает ее менее уязвимой к денатурирующим
агентам. Таким образом, хотя непосредственным исполнителем каталитической
функции является простетическая группа, образующая каталитический центр, ее
действие немыслимо без участия полипептидных фрагментов белковой части
фермента. Более того, в апоферменте есть участок, характеризующийся
специфической структурой, избирательно связывающий кофермент. Это так
называемый кофермент связывающий домен; его структура у различных апоферментов,
соединяющихся с одним и тем же коферментом, очень сходна. Таковы, например,
пространственные структуры нуклеотидсвязывающих доменов ряда дегидрогеназ.
Иначе обстоит дело у однокомпонентных ферментов, не имеющих добавочной
группы, которая могла бы входить в непосредственный контакт с преобразуемым
соединением. Эту функцию выполняет часть белковой молекулы, называемая
каталитическим центром. Предполагают, что каталитический центр
однокомпонентного фермента представляет собой уникальное сочетание нескольких
аминокислотных остатков, располагающихся в определенной части белковой
молекулы.
Чаще всего в каталитических центрах однокомпонентных ферментов
встречаются остатки сер, гис, три, арг, цис, асп, глу и тир. Радикалы перечисленных
аминокислот выполняют здесь ту же функцию, что и кофермент в составе
двухкомпонентного фермента.
Аминокислотные остатки, образующие каталитический центр однокомпонентного
фермента, расположены в различных точках единой полипептидной цепи. Поэтому
каталитический центр возникает в тот момент, когда белковая молекула
приобретает присущую ей третичную структуру. Следовательно, изменение третичной
структуры фермента под влиянием тех или иных факторов может привести к
деформации каталитического центра и изменению ферментативной активности.
Кроме каталитического центра, образованного сочетанием аминокислотных
радикалов или присоединением кофермента, у ферментов различают еще два центра:
субстратный и аллостерический.
Под субстратным центром понимают участок молекулы фермента, ответственный
за присоединение вещества (субстрата), подвергающегося ферментативному
превращению. Часто этот участок называют “якорной площадкой” фермента, где, как
судно на якорь, становится субстрат. Во многих случаях прикрепление субстрата к
ферменту идет за счет взаимодействия с -аминогрулпой радикала лиз,
расположенного в субстратном центре. Эту же роль может выполнять СООН-группа
глу, а также НS-группа цис. Однако работы последних лет показали, что гораздо
большее значение здесь имеют силы гидрофобных взаимодействий и водородные
связи, возникающие между радикалами аминокислотных остатков субстратного центра
фермента и соответствующими группировками в молекуле субстрата.
Понятие о каталитическом и субстратном центре не следует абсолютизировать.
В реальных ферментах субстратный центр может совпадать (или перекрываться) с
каталитическим центром. Более того, каталитический центр может окончательно
формироваться в момент присоединения субстрата. Поэтому часто говорят об
активном центре фермента, представляющем сочетание первого и второго. Активный
центр у ферментов располагается на две щели при двухъядерной структуре,
например у лизоцима и рибонуклеазы, или на дне глубокой впадины, как у
химотрипсиногена.
Аллостерический центр представляет собой участок молекулы фермента, в
результате присоединения к которому определенного низкомолекулярного (а иногда
- и высокомолекулярного) вещества изменяется третичная структура белковой
молекулы. Вследствие этого изменяется конфигурация активного центра, сопровождающаяся
либо увеличением, либо снижением каталитической активности фермента. Это
явление лежит в основе так называемой аллостерической регуляции каталитической
активности ферментов.
Значения молекулярных масс ферментов колеблются в широких пределах: от
нескольких тысяч до нескольких миллионов. В природе насчитывается несколько
десятков ферментов, обладающих сравнительно небольшими молекулами (до 50 тыс.).
Однако большинство ферментов представлено белками более высокой молекулярной
массы, построенными из субъединиц. Так, каталаза (М=25200) содержит в молекуле
шесть протомеров с М=42000 каждый. Молекула фермента, ускоряющего реакцию
синтеза рибонуклеиновых кислот (РНК-полимераза, М = 400000), состоит из 6
неравных субъединиц. Полная молекула глутаматдегидрогеназы, ускоряющей процесс
окисления глутаминовой кислоты (М=336000), построена из 6 субъединиц с М=56000.
Способы компоновки протомеров в мультимеры разнообразны. Крайне важно,
что достроенный из субъединиц фермент проявляет максимальную каталитическую
активность именно в виде мультимера: диссоциация на протомеры резко снижает
активность фермента. Не все ферменты-мультимеры построены исключительно из
каталитически активных протомеров. Наряду с каталитическими в их составе
отмечены регуляторные субъединицы, как, например, у
аспартат-карбамилтрансферазы.
Среди ферментов-мультимеров безусловно преобладают димеры и тетрамеры (их
несколько сотен), в меньшей мере распространены гексамеры и октамеры (несколько
десятков) и необыкновенно редко встречаются тримеры и пентамеры.
Молекулы ферментов-мультимеров в ряде случаев составлены из субъединиц
двух типов, обозначаемых условно как субъединицы типа А и В. Они сходны друг с
другом, но отличаются по некоторым деталям первичной и третичной структур. В
зависимости от соотношения протомеров типа А и В в мультимере последний может
существовать в виде нескольких изомеров, которые называют изозимами. Так, при
четырех субъединицах возможны 5 изозимов:
I II
III IV V
AAAA AAAB AABB ABBB BBBB
В настоящее время интерес к изозимам резко повысился. Оказалось, что
кроме генетически детерминированных изозимов существует большая группа
ферментов, обладающая множественными формами, возникающими в результате их
посттрансляционной модификации. Множественные формы ферментов и изозимы в
частности используются сейчас для диагностики болезней в медицине,
прогнозирования продуктивности животных подбора родительских пар при
скрещивании для обеспечения максимального гетерозиса в потомстве и т. п.
Значение пространственной организации ферментов особенно ярко выявляется
при изучении строения так называемых мультиэнзимов, т.е. ферментов, обладающих
способностью ускорять одновременно несколько химических реакций и осуществлять
сложные превращения субстрата. Примером может служить мультиэнзим, ускоряющий
реакцию окислительного декарбоксилирования пировиноградной кислоты. Этот многоферментный комплекс с М=4500000 состоит
из трех видов ферментов. Первый из них (E1) ускоряет реакцию декарбоксилирования
пировиноградной кислоты. В состав комплекса входит 12 димерных молекул этого
фермента (К=19200). Второй и третий ферменты, катализирующие
окислительно-восстановительные процессы при окислении пировиноградной кислоты,
сосредоточены внутри мультиэнзимного комплекса. Один из них (Е3) представлен
шестью димерными молекулами (М=112 000), другой (Е2) - 24 протомерами (М=70000)[3,с
.15].
В тех случаях, когда мультиэнзимный комплекс обслуживает единый,
многоступенчатый процесс биохимических превращений, его называют метаболоном
(от слова метаболизм - обмен веществ). Таковы метаболоны гликолиза, биосинтеза
ряда аминокислот, цикла дикарбоновых и трикарбоновых кислот и др.
В результате слаженного во времени и пространстве действия всех трех
видов входящих в его состав ферментов мультиэнзим с огромной скоростью
осуществляет превращение пировиноградной кислоты. Именно в кооперативном
характере каталитического процесса и кроется главное отличие биокатализаторов
от катализаторов неорганической природы, именно поэтому интенсивность
биокатализа в десятки, сотни и тысячи раз превосходит мощность действия
неорганических катализаторов.
Сравнительно недавно выявлена еще одна своеобразная черта в строении
ферментов: некоторые из них являются полифункциональными, т.е. обладают
несколькими энзиматическими активностями, но всего лишь одной полипептидной
цепью. Дело в том, что эта единая цепь при формировании третичной структуры
образует несколько функционально и стерически обособленных глобулярных участков
- доменов, каждый из которых характеризуется своей каталитической активностью.
6. Иммобилизованные ферменты
Биотехнологические процессы с
использованием микроорганизмов и ферментов уже на современном техническом
уровне широко применяют в пищевой промышленности. Промышленное выращивание
микроорганизмов, растительных и животных клеток используют для получения многих
ценных соединений — ферментов, гормонов, аминокислот, витаминов, антибиотиков,
метанола, органических кислот (уксусной, лимонной, молочной) и т. д. С помощью
микроорганизмов проводят биотрансформацию одних органических соединений в
другие (например, сорбита во фруктозу). Широкое применение в различных
производствах получили иммобилизованные ферменты. Для выделения биологически
активных веществ из сложных смесей используют моноклональные антитела. А. С.
Спириным в 1985-88 разработаны принципы бесклеточного синтеза белка, когда
вместо клеток применяются специальные биореакторы, содержащие необходимый набор
очищенных клеточных компонентов. Этот метод позволяет получать разные типы
белков и может быть эффективным в производстве. Многие промышленные технологии
заменяются технологиями, использующими ферменты и микроорганизмы. Таковы
биотехнологические методы переработки сельскохозяйственных, промышленных и
бытовых отходов, очистки и использования сточных вод для получения биогаза и
удобрений. В ряде стран с помощью микроорганизмов получают этиловый спирт,
который используют как горючее для автомобилей (в Бразилии, где топливный спирт
широко применяется, его получают из сахарного тростника и других растений). На
способности различных бактерий переводить металлы в растворимые соединения или
накапливать их в себе основано извлечение многих металлов из бедных руд или
сточных вод.
Заключение
В ходе выполнения работы была
достигнута ее основная цель и решены все задачи, поставленные во введении. В
заключении сделаем несколько выводов по работе.
Ферменты - это специфические белки,
входящие в состав всех клеток и тканей живых организмов играющие роль
биологических катализаторов. О ферментах люди узнали давно.
Лигазы - ускоряют реакции синтеза, сопряженные с распадом богатых
энергией связей.
Характерные черты действия ферментов класса лигаз (синтетаз) выявлены
совсем недавно в связи со значительными успехами в изучении механизма синтеза
жиров, белков и углеводов.
Механизм действия лигаз изучен еще недостаточно, но, несомненно, он
весьма сложен. В ряде случаев доказано, что одно из участвующих в основной
реакции веществ сначала дает промежуточное соединение с фрагментом
распадающейся молекулы АТФ, а вслед за этим указанный промежуточный продукт
взаимодействует со вторым партнером основной химической реакции с образованием
конечного продукта.
Главная их особенность - сопряженность синтеза с распадом веществ,
способных поставлять энергию для осуществления биосинтетического процесса.
Одним из таких природных соединений является АТФ. При отрыве от ее молекулы в
присутствии лигаз одного или двух концевых остатков фосфорной кислоты
выделяется большое количество энергии, используемой для активирования
реагирующих веществ. Лигазы же каталитически ускоряют синтез органических
соединений из активированных за счет
распада АТФ исходных продуктов. Таким образом, к лигазам
относятся ферменты, катализирующие соединение друг с другом двух молекул,
сопряженное с гидролизом пирофосфатной связи в молекуле АТФ или иного
нуклеозидтрифосфата.
Список литературы
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник.- М.:
Медицина, 1990.- 234 с.
2. Основы биохимии: Учебник для студ. биол. спец. ун-тов/под ред. А.А.
Анисимова.- М.: Выс.шк., 1986. – 245 с.
3. Фёршт Э. Структура и механизм действия ферментов.- М.: Мир., 1980.-
450 с.
4. Кочетов Г.А. Практическое руководство по энзимологии.- М., 1989
5. Диксон М., Уэбб Э. Ферменты: пер.англ.- М.: Мир, 1982.- т.1.- 478 с.
6. Асатиани В.С. Биохимическая
фотометрия.- М.: Изд. АН СССР, 1957.- 432 с.