Содержание
Введение. 3
1. Основные аспекты эволюции растительного и животного мира. 4
1.1. Организмы I-го поколения. 5
1.2. Организмы II-го поколения. 7
1.3. Организмы III-го поколения. 10
1.4. Организмы IV-го поколения. 11
2. Загадка кембрийского взрыва. 12
Заключение. 17
Глоссарий. 18
Список литературы.. 19
Введение
Целью настоящей работы является изучение процессов эволюции растительного и животного мира, изучение загадки кембрийского взрыва.
Примерно 3800 миллионов лет назад в результате химической реакции образовалось первое сложное соединение, способное самовоспроизводиться. Так было положено начало жизни на Земле.
Зарождение жизни на Земле, в целом, остается загадкой. Но с тех пор как Чарльз Дарвин впервые описал процесс эволюций растительного и животного мира, ученые придерживаются мнения, что все формы жизни находятся в процессе постоянного и непрерывного развития. С каждым новым поколением сильные стороны оттачиваются, а слабые отсеиваются. Один вид пращуров мог породить несколько форм жизни, после чего или вымирал, или находил собственную нишу в экосистеме. Судя по найденным окаменелостям, ясно: система родственных линий между существующими и исчезнувшими организмами напоминает древо, на котором с течением времени появляются новые ветви. Многие из них засыхают и отмирают (например, динозавры), но другие ветви вырастают и расцветают. Если проследить любую из них до основания, то, в конечном итоге, мы придем к единому стволу - прародителю всех когда-либо живших организмов, т.е. источнику происхождения жизни на Земле [1, с. 56].
Кембрийский эволюционный взрыв - одна из величайших загадок в истории развития жизни на Земле. Понадобилось 2,5 млрд лет, чтобы простейшие клетки развились в более сложные эукариотные клетки, и еще 700 млн лет для возникновения первых многоклеточных организмов. А затем, всего за какие-то 100 млн лет, мир оказался заселен невероятным разноооразием многоклеточных животных. С тех пор за более чем 500 млн лет на Земле не появилось ни одного нового типа (принципиально иного строения тела) животных.
1. Основные аспекты эволюции растительного и животного мира
Эволюция растительного и животного мира происходила в течение сравнительно длительного периода времени и основные её этапы хорошо известны. Вместе с тем, на всём протяжении этого развития от водорослей и бактерий до современных нам представителей флоры и фауны все процессы их образования, существования и отмирания подчинялись единым принципам системной организации Материи, действие которых распространяется на каждый организационный уровень, включая и подуровень И. Все относящиеся к нему организмы представляют собой целостные системы, структуры которых можно представить в виде определённым образом расположенных в пространстве ячеек, заполненных в качестве единиц органическими клетками.
Системы организмов имеют, как правило, подсистемы - органы, несущие ту или иную нагрузку. Структуру органов составляют ячейки с примерно однотипными алгоритмами и поэтому заполняющие их единицы - клетки имеют приблизительно одинаковое строение и, соответственно, свойства. Группы таких клеток носят название тканей. Как и в предыдущих организационных подуровнях, время существования ячеек не совпадает с периодом функционирования единиц. Поэтому все организмы имеют подсистемы, обеспечивающие доставку комплектующих элементов - различных атомов и молекул для образования новых единиц, идентичных заменяемым в ячейках отфункционировавшим единицам. При этом свойства вновь образованных клеток должны полностью совпадать свойствами заменяемых и, в конечном счёте, корреспондировать алгоритмам заполняемых ячеек. Механизмом, обеспечивающим поддержание в ячейках подсистем организмов в постоянной готовности соответствующих единиц, является митоз (деление) клеток.
Зарождение так называемой живой природы произошло в водах мирового океана, или вернее, на стыке морей и суши. Наличие всех компонентов, включая воду, а также атомов большинства химических элементов в совокупности с каждодневным постоянным источником энергии - лучистой энергией Солнца - создало идеальные предпосылки для системного конструирования различных структур ячеек, которые тут же могли заполняться необходимыми единицами. И поэтому не эпизодические грозовые разряды (бывшие лишь необходимым условием, но отнюдь не причиной) послужили толчком к зарождению сложных биоструктур (как утверждают отдельные научные гипотезы), а последовательное перебирание различных системных вариантов в сочетании с соответствующими благоприятными условиями внесистемной среды привело к созданию динамически устойчивых биосистем. Молекулы морской воды в сочетании с различными химическими элементами в виде растворов проникали сквозь оболочку новых системных формирований и заполняли в качестве единиц соответствующие ячейки их структур, при этом лучистая энергия Солнца, преобразуясь и застывая в виде энергии межмолекулярной связи, способствовала удержанию на период функционирования единиц в своих ячейках [2, с. 88].
1.1. Организмы I-го поколения
В результате длительного процесса эволюции организмов и их подсистем появилось большое количество разнообразных растений, которые мы отнесём к одной группе так называемых организмов первого поколения. Несмотря на кажущееся внешнее различие, а также наличие несхожих подсистем, всех их объединяет, и это особенно важно, единый принцип построения структур. А именно: в их ячейки в качестве единиц поступают в виде растворов представители всего набора подуровней В и Г - атомы, молекулы, ионы, радикалы и т.п., то есть элементы неорганических соединений, находящихся в почве, а точнее, в окружающей среде и соединяемых в ячейках данного вида организмов с помощью энергии Солнца в системы очень сложной организации. Синтезированные таким образом из CO2, H2O и других системных образований нижних подуровней глюкоза, аминокислоты, а затем углеводы, белки, нуклеотиды и т.д., то есть формирования более высоких подуровней заполняли в качестве единиц ячейки подсистем органических клеток, которые сами уже являлись единицами в структуре организмов растений. Клетки, системная организация которых позволяла производить синтез структур указанным образом, впоследствии стали называться автотрофными. Характерными их представителями являются клетки современных нам зелёных растений.
Основной реакцией, протекающей в организмах первого поколения, является реакция фотосинтеза:
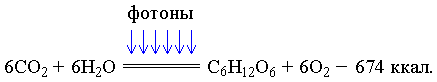
Кванты света, бомбардируя молекулярную структуру хлорофилла, передают определённое количество своей кинетической энергии части его электронов, переводя их таким образом в возбуждённое состояние. В результате этого электроны покидают свои орбитали и перескакивают в более высокие. Часть из них, присоединяясь к ионам водорода, превращает их в водород и т.д. Одновременно при этом процессе АДФ превращается в АТФ, а CO2 в глюкозу. Фотосинтез служит основой постоянно протекающего в природе великого созидательного процесса биосинтеза, в результате которого создаётся бесчисленное множество единиц, заполняющих соответствующие им ячейки в структурах различных биоорганизмов.
Организмы I-го поколения, несмотря на их относительную примитивность, имели уже довольно надёжную подсистему алгоритмозаписи, в основе которой лежит биохимическая запись генетического кодирования ДНК. В ней собирается информация практически от всех клеток, входящих в организм. По мере усложнения системной организации растений повышалась надёжность и подсистемы алгоритмозаписи, обеспечивавшей кодирование развёртывания структуры ячеек всех подсистем организма, соотнесённое с пространственно-временными интервалами. Вначале подсистему алгоритмозаписи имел практически каждый орган растения. Так до сих пор существуют растения, у которых при культивировании лишь одного органа происходит развёртывание всех остальных. К ним можно отнести лесной ландыш (корневище), тополь (часть стебля) и т.д. Однако, в конечном итоге, наиболее надёжной оказалась система записи алгоритмов, производимая в особом, специально для этого предназначенном органе растения - его семенах. Одно из главных преимуществ такой записи является возможность её реализации (алгоритмочтения) с большим интервалом как в пространстве, так и во времени [2,с. 112].
Как ни велико множество разновидностей организмов I-го поколения, функциональная нагрузка их в целом одинакова, а разница состоит лишь в структурной организации их подсистем, приспособленных под те или иные особенности биогеоценоза, в котором они территориально размещены и единицами которого они сами являются. Поэтому, исчерпав весь набор возможных функциональных приращений в структурах организмов I-го поколения, Развитие Материи перешло в новую область - к конструированию структур с новыми функциями у организмов с более высокой системной организацией, которые объединяются в следующую группу - организмы II-го поколения. Их появление было следствием существования уже достаточно развившихся организмов I-го поколения, хотя последующее одновременное функционирование и развитие организмов обоих поколений несколько затушевывает вторичность генезиса организмов II-го поколения. Но уже то, что их различает, а именно: при формировании единиц для ячеек своих подсистем в последних используются в качестве основы комплексные блоки единиц организмов I-го поколения, раскрывает очерёдность появления поколений.
1.2. Организмы II-го поколения
К II-му поколению организмов относятся все растительноядные представители животного мира. Развитие у них подсистемы ускоренного искусственного расщепления органических соединений тканевых структур растений позволило им добывать в большом количестве комплексные материальные соединения, с помощью которых они могли постоянно заполнять ячейки своих всё более усложнявшихся подсистем, что способствовало появлению ячеек с новыми свойствами и соответствовало движению Материи по ординатам качества-времени. Мы не будем подробно анализировать эволюцию организмов II-го поколения от простейших одноклеточных до современных нам хордовых класса млекопитающих травоядных животных. Отметим лишь, что главной причиной дивергенции их системной организации была потребность следования законам Развития Материи. Основой этого длительного процесса явилось усложнение морфофизиологической структуры организмов, что привело к появлению в протерозойскую эру (2 млрд. лет назад) животных с двусторонней симметрией тела с дифференцировкой его на передний конец и задний. Передний конец стал местом для развития органов чувств, нервных узлов, а в дальнейшем - головного мозга. В процессе последующей эволюции происходила, главным образом, дивергенция типов животного мира и замена первоначальных низкоорганизованных примитивных форм более высокоорганизованными путём ещё большей дифференциации строения и функций тканей и органов организмов. При этом ячейки тканей организмов II-го поколения заполнялись в качестве единиц уже лишь гетеротрофными клетками, то есть неспособными к синтезу органических соединений из неорганических. В самих клетках ещё более усовершенствовалась система генозаписи в цепочках ДНК. Характерной особенностью клеток любого органа осталось то, что в каждой из них имелся весь набор генов данного вида организмов, однако в клетках разных тканей используются лишь немногие группы генов, то есть только те из них, на которых записаны алгоритмы структурного развёртывания и функционирования структур ячеек, которые данные клетки занимают в качестве единиц.
Протекавший многие сотни миллионов лет морфофизиологический прогресс, или ароморфоз, привёл к значительным эволюционным изменениям подсистем структуры организмов II-го поколения (что выразилось в общем подъёме их организации), биологическому прогрессу, а также другим не менее важным последствиям. Сюда, прежде всего, следует отнести отрыв их систем от гумусового слоя почвы и способность свободно и автономно перемещаться по субстрату. Вследствие этого организмы получили возможность постепенно осваивать до этого пустынные районы земной поверхности в трёх сферах: на суше, в воде и воздухе, что вело к увеличению разнообразия их структур и вполне отвечало требованиям движения Материи в качестве-времени-пространстве. Приобретённая способность к передвижению в пространстве околоземной поверхности позволило организмам II-го поколения перемещаться от одного источника питания (системы организмов I-го поколения) к другому, максимально расширяя ареал своего обитания. Кроме того, в неблагоприятные моменты организм стал иметь возможность укрываться в более безопасном для себя месте. Потребление разнообразных травянистых растений увеличило набор элементов, из которых слагались единицы, заполнявшие ячейки подсистем организмов животных, при этом каждый элемент заполнял отведённую именно для него ячейку, где он мог проявить свои, присущие лишь ему свойства. Вместе с тем, как и во всех системных образованиях предыдущих подуровней, любая вновь означившаяся ячейка структуры того или иного организма безусловно требовала для своего заполнения лишь единицу, способную выполнять её набор алгоритмов. Малейшее несоответствие единицы заполняемой ячейке вело к нарушению функционирования данной подсистемы организма и к возможному прекращению существования всей его системы в целом.
В процессе эволюции новые, высшие по своей организации группы организмов возникали путём ароморфозов, идиоадаптаций и дегенераций. На одном из этапов указанного процесса развития системной организации Материи появились представители организмов третьего поколения. К ним относятся организмы, использующие в качестве строительных полуфабрикатов при синтезировании своих единиц не неорганические вещества гумусового слоя и не органические соединения размельчённых тканей отдельных органов растений, а значительно более сложные органические вещества тканей организмов II-го поколения. В результате этого, у плотоядных животных, как они впоследствии стали называться, отпала необходимость постоянно и в больших количествах потреблять отдельные органы разнообразных растений для того, чтобы заполнить ячейки своих подсистем соответствующими единицами. Им стало достаточно овладеть одним из организмов II-го поколения, чтобы получить сразу в большом количестве разнообразие многих необходимых элементов, находящихся в ячейках организма растительноядного животного и из которых они могли синтезировать единицы для подсистем своего организма. Начиная с этого времени необходимые элементы организм стал получать готовыми блоками (блок-питание), что вполне отвечало принципам построения материальных систем, предопределяющим использование в качестве единиц в структурах всех последующих ступеней организации устойчивых комплексов единиц предыдущих уровней [3,с. 144].
1.3. Организмы III-го поколения
В системной организации организмов III-го поколения произошло меньше изменений по отношению к организмам II-го поколения, чем это было между поколениями II-м и I-м. Прежде всего, значительно изменилась подсистема пищеварения, приспосабливаясь под новый вид питания, а также ещё большую значимость получила нервная подсистема. Среди организмов III-го поколения по уровню своего развития всё больше стали выделяться наземные животные. В конечном итоге, всё дальнейшее развитие животного мира стало сводиться, в основном, именно к последовательному усложнению у наземных организмов III-го поколения ЦНП, повышению интенсивности и эффективности её использования, увеличению разнообразия спектра её функций. Главным образом, это сказалось на системной организации головного мозга, который всё больше становился специализированной подсистемой умножающихся аналитических фн. центров, объединяющих анализаторы и инициаторы большинства процессов, протекавших внутри организма, и некоторых - вне его.
Несмотря на большое количество видов организмов всех трёх поколений (а их только в настоящее время насчитывается на Земле около 0,5 млн. видов растений и 1,5 млн. - животных) и их разнородность, на ординате качества-времени тем не менее всё равно наступил момент, когда всего этого разнообразия стало недостаточно для обеспечения дальнейшего Развития Материи. Выход из этого мог быть, как и прежде, найден лишь в ещё более сложной системной организации Материи путём создания очередного нового организационного уровня. Первые предпосылки перехода к нему начали возникать ещё около 30 млн. лет назад, когда в лесах палеогена и неогена появились парапитеки - животные величиной с кошку, которые жили на деревьях и питались растениями и насекомыми. От парапитека произошли современные гиббоны и орангутаны и ещё одна ветвь - вымершие древние обезьяны дриопитеки, которые дали три ветви, приведшие к шимпанзе, горилле и человеку. Чарльз Дарвин убедительно доказал, что человек представляет собой последнее, высокоорганизованное звено в цепи развития живых существ четырёх поколений и имеет общих далёких предков с человекообразными обезьянами.
1.4. Организмы IV-го поколения
Итак, результатом движения Материи по организационному уровню следует считать создание наиболее развитых организмов - организмов IV-го поколения, к коим мы причисляем только человека, система организма которого в целом к тому времени достигла стабильного совершенства. Будучи производной системой, вобравшей в себя всё лучшее от организмов II-го и III-го поколений, человек получил в качестве генетического наследства набор всех тех подсистем, которые обеспечивали его существование и надёжное функционирование в широком диапазоне окружающей среды. В качестве питания для заполнения ячеек своих подсистем его организм всё более приспосабливался к потреблению высокопитательных частей организмов I-го и II-го поколений. Так, в его рационе всё большую долю стали занимать элементонасыщенные аккумулятивные подсистемы, формируемые вокруг семян у организмов I-го поколения (плоды, ягоды, фрукты) и различные части организмов II-го поколения. Части организмов III-го поколения, то есть плотоядных животных, человек в пищу практически не потреблял и не употребляет, как этого не делают и сами плотоядные животные, ввиду невозможности их использования для заполнения ячеек подсистем его организма. Однако, опережающее развитие и специализацию в дальнейшем вплоть до наших дней в организме человека всё более стала получать подсистема, регулирующая его высшую нервную деятельность, и в первую очередь, структура его головного мозга.
2. Загадка кембрийского взрыва
Примерно 530 миллионов лет назад, в начале кембрийской эпохи, на Земле произошло уникальное событие - внезапно, быстро и почти одновременно возникло множество новых биологических форм, ставших предшественниками важнейших типов современных организмов вплоть до человека. Многим биологам до сих пор трудно примириться с мыслью о реальности этого кембрийского взрыва. В одном из стандартных учебников биологии для американских университетов можно, например, и сейчас прочесть утверждение, что "формы, жившие в тот (кембрийский) период, наверняка должны были произойти от предков, существовавших по крайней мере за сотни миллионов, а то и миллиарды лет до того". Но в том-то и загадка кембрийского взрыва, что никакие переходные, промежуточные формы не соединяли появившиеся тогда новые типы организмов с бактериями и простейшими водорослями, которые населяли земные океаны до них. И это отсутствие привычного "мостика" ставит перед биологами труднейшую задачу - объяснить, каким образом мог произойти такой загадочный эволюционный скачок.
Уникальность и загадочность особенностей Кембрийского взрыва - вот что привлекает к нему неослабное внимание биологов на протяжении последних ста пятидесяти лет.
Сложность проблемы состоит, однако, не только в этой загадочности кембрийского "биологического Биг Бэнга" и причин, его породивших. Не менее важным толчком к острым и непрекращающимся спорам вокруг него служит тот факт, что проблема Кембрийского взрыва имеет еще и прямое отношение к дарвиновской теории эволюции. Точнее говоря, попросту ей противоречит. Первым, кто это осознал, был сам Дарвин. Он же был первым, кто предложил возможный выход из этого противоречия. Однако предложенная Дарвином гипотеза не удовлетворила многих его последователей, и в результате биологи-эволюционисты разделились на два враждующих лагеря, спор между которыми длится вот уже полтора столетия. Попробуем разобраться в этом споре [4,с. 110].
Первооткрывателем Кембрийского взрыва был Роберт Мэрчисон - английский аристократ, который под влиянием своей честолюбивой жены решил заняться наукой. Изучая окаменелости древних эпох, обнаруженные в соответствующих отложениях, он обнаружил, что слои этих отложений разделены резкой границей. Ниже этой границы они крайне бедны биологическими останками и демонстрируют повсеместное распространение одних лишь простейших одноклеточных организмов - бактерий и водорослей, а затем, начиная с кембрийской эпохи, около 550 миллионов лет назад, внезапно обзаводятся невиданным богатством новых биологических форм. Будучи человеком верующим и разделяя убеждение великого Линнея, что "существует ровно столько видов, сколько их первоначально сотворил Создатель", Мэрчисон расценил открытое им явление как прямое свидетельство вмешательства Божьей руки в развитие жизни. Понятно, что такое креационистское (от слова creation - сотворение) объяснение было несовместимо с представлением о естественной эволюции биологических форм.
Мэрчисон опубликовал результаты своих исследований в тридцатые годы прошлого века. Несколько десятилетий спустя вышла в свет знаменитая работа Дарвина "Происхождение видов", в которой впервые последовательно излагалась и подробно аргументировалась теория развития жизни на Земле, основанная на представлениях о наследуемых изменениях и естественном отборе. Разумеется, Дарвин не принимал креационизма. Но он сразу же усмотрел, что Кембрийский взрыв является камнем преткновения для его теории в другом - не менее важном - аспекте.
Дело в том, что, по Дарвину, эволюция должна была происходить постепенно, плавно и непрерывно, то есть, как сегодня говорят, градуально. В своей книге он весьма недвусмысленно писал: "Естественный отбор ежедневно и ежечасно подвергает строжайшему придирчивому обследованию все происходящие в мире изменения, даже самые малейшие, отвергая то, что плохо, сохраняя и улучшая то, что хорошо... Мы не замечаем этих медленных изменений в их постепенном становлении и замечаем их лишь тогда, когда ход времени отмеряет огромные промежутки целых исторических эпох".
Понятно, что дарвиновский градуализм был несовместим с наличием столь резкого, кратковременного и стремительного явления, как Кембрийский взрыв. Градуализм покоится на убеждении, прекрасно сформулированном знаменитым популяризатором дарвинизма Т.Х.Хаксли: "Природа не терпит скачков".
Как уже сказано, Дарвин попытался найти выход из возникшей ситуации. Возможно, предположил он, Кембрийский взрыв, на самом деле, не был настоящим "взрывом"; возможно, в действительности ему предшествовал длительный период постепенного накопления эволюционных изменений и становления новых биологических форм; но Мэрчисон попросту не сумел обнаружить эти предшествующие, промежуточные формы. Это объяснение позволяло сохранить тот непрерывный и плавный характер эволюции, который Дарвин постулировал на основе собранных им эмпирических данных и который был в его глазах стержневой особенностью всего эволюционного процесса.
Некоторые биологи-эволюционисты не согласились с дарвиновским толкованием кембрийской загадки. (Уже Хаксли в своем письме Дарвину накануне публикации "Происхождения видов" предупреждал: "Вы напрасно взвалили на себя совершенно ненужную трудность, приняв, что природа не терпит скачков".) Эти дарвинисты вообще не могли принять дарвиновского градуализма. Он казался им не столько выведенным из эмпирических фактов (ведь противоречил же он фактам Мэрчисона), сколько привнесенным в биологию извне [5, с. 80].
Так случилось, что кембрийская загадка с самого начала разделила дарвинистов-эволюционистов на два противостоящих лагеря, по-разному понимающих ход биологической эволюции. По одну сторону водораздела оказались убежденные "градуалисты", по другую - столь же убежденные "катастрофисты". (Третий лагерь, противостоящий и "градуалистам", и "катастрофистам" в своем полном отрицании эволюции вообще, составляют современные "креационисты").
Сторонники дарвиновского градуализма предлагают различные возможные объяснения отсутствию докембрийских промежуточных форм. Одни утверждают, что предшествующие кембрию биологические формы не сохранились потому, что не имели скелета или наружного панциря и были мягкими, желевидными (что, кстати, в основном соответствует истине). Другие объясняют отсутствие переходных форм в до-кембрийских отложениях чисто физическими причинами, утверждая, что до-кембрийские породы подверглись такому сильному нагреву и давлению, что в них не сохранились никакие биологические останки (что не вполне соответствует истине). Третьи выдвигают предположение, что до-кембрийская жизнь развивалась в озерах, а кембрийский взрыв - попросту следствие бурной и быстрой миграции уже сформировавшихся в этих озерах биологических форм в моря и океаны (эта гипотеза получила своеобразное развитие в упомянутой выше работе Киршвинка и сотрудников). Все эти гипотезы объединяет стремление показать, что переход от до-кембрийских форм к после-кембрийским был главным и непрерывным, только следы его по тем или иным причинам пока не найдены или вообще не сохранились.
И действительно, не так давно исследователям удалось обнаружить первые виды многоклеточных организмов, непосредственно предшествовавших кембрийским. Они были найдены в отложениях вблизи австралийского поселка Эдиакара и потому получили название "эдиакарских". Почти до самого последнего времени, до восьмидесятых годов, эдиакарские организмы трактовались в духе дарвиновского градуализма - как промежуточное звено в истории постепенного усложнения, или эволюции биологических форм от до-кембрийских к после-кембрийским.
Но около пятнадцати лет назад более тщательное изучение этих останков показало, что, на самом деле, они не имеют связи с современными биологическими формами. Возможно, они вообще представляли собой некую особую, тупиковую ветвь биологической эволюции, которая не дала никакого продолжения. Некоторые биологи полагают, что эта ветвь жизни была уничтожена в ходе какой-то катастрофы, предшествовавшей Кембрийскому взрыву.
Заключение
Таким образом, по работе можно сделать следующие выводы.
В результате длительного организационного процесса, протекавшего многие миллионы лет, вначале появились простейшие одноклеточные организмы - синезелёные водоросли и бактерии, затем зелёные водоросли, грибы и другие многоклеточные растения, имевшие самое примитивное строение, но являвшиеся венцом творения Материи на тот момент её Развития. Последующее течение времени и соответствующее продвижение Материи по ординате качества требовало дальнейшего умножения функций В силу этого водоросли, попадая на сушу, стали всё более приспосабливаться к обезвоженной среде. В их организме началось расслоение подсистем, каждая из которых выполняла определённую функцию. В отдельных случаях некоторые ткани стали наделяться двумя и более функциями, то есть становились полифункциональными, отвечая тем самым законам общего Развития Материи.
В кембрийский период на Земле существовали громадные области, занятые континентальным шельфом, или материковыми отмелями. Здесь создались идеальные условия для жизни: дно, покрытое слоем мягкого ила, и теплая вода. К этому времени в атмосфере образовалось много кислорода, хотя его и было меньше, чем сегодня. Развитие твердых покровов привело к появлению новых жизненных форм, таких, как членистоногие, артроподы. Животным понадобились новые способы защиты от новых высокоорганизованных хищников. Улучшились средства их защиты - и уже хищникам пришлось вырабатывать новые методы охоты, чтобы преодолеть сопротивление жертвы. На протяжении кембрийского периода уровень моря неоднократно повышался и понижался. При этом некоторые популяции вымирали, а места их обитания занимали другие животные, которым, в свою очередь, приходилось приспосабливаться к новым условиям жизни. Животный мир становился разнообразнее, и все больше видов животных могло существовать бок о бок, не претендуя на пищевые ресурсы соседей.
Глоссарий
Термин "эволюция" (развертывание), введенный в 18 веке Шарлем Боннэ, в современном глоссарии обозначает учение об истории развития органического мира, утверждающем происхождение всех видов организмов от раннее существующих путем изменений.
Термин "эволюция" чаще всего применяется для обозначения исторического развития живых организмов, определяемого изменчивостью, наследственностью и естественным отбором; сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием сообществ организмов и биосферы в целом.
Организмы первого поколения – живые организмы, к которым относятся разнообразные виды растений
Организмы второго поколения – живые организмы, к которым относятся растительноядные представители животного мира
Организмы третьего поколения – живые организмы, к которым относятся наземные животные
Организмы четвертого поколения – человек
Докембрий (криптозой) - промежуток времени в истории Земли, предшествовавший палеозойской эре (кембрийскому периоду).
"Кембрийский взрыв" - бурное и одновременное возникновение морской фауны и множества новых биологических видов, произошедшее приблизительно 540 миллионов лет назад.
Список литературы
1. Дубнищева Т.Я. Концепции современного естествознания: Учебник / Под ред. академика РАН М.Ф. Жукова. – Новосибирск: ЮКЭА, 1997.
2. Дубнищева Т.Я., Пигарев А.Ю. Современное естествознание: Учеб. пособие. – Новосибирск: ЮКЭА, 2001 .
3. Карпенков С.Х. Концепции современного естествознания: Учебник. – М.: ЮНИТИ, 1997.
4. Потеев М.И. Концепции современного естествознания: Учебник – СПб.: Питер, 2002.
5. Горелов А.А. Концепции современного естествознания: Учебник – М.: Владос, 1999.