Содержание
1. Характеристика цикла Крепса. Схема значения,
энергетический выход. 3
2. Укажите этапы устичной транспирации. Закон
Стефана. Явление краевой диффузии 4
3. Напишите схему и дайте характеристику цикла
Кальвена. 7
4. Охарактеризуйте этапы поступления ионов в
клетку, их значение. 11
Список литературы.. 14
1. Характеристика цикла Крепса. Схема значения,
энергетический выход
Основным циклом
производства энергии в живом организме является цикл Крепса.
Составляющие этого цикла используют кислород в митохондориях.
Если мыслить в рамках
параидальной современной биологии, то получится, что клетки погибают при
отсутствии кислорода.
Кривая сохранения
жизнеспособности была предложена американским ученым, доктором Дрингером. Эта
кривая показывает время, прошедшее с момента остановки дыхания, и процент
людей, реанимированных с помощью искусственного дыхания. Кривая, идущая снизу,
показывает процент реанимации, а кривая, идущая сверху, - процент смертности. Если
с момента прекращения дыхания прошло хотя бы 5 минут, то из 100 человек спасти
не удается 75. А по прошествии десяти минут не удается спасти почти все 100
процентов. Таким образом, остановка дыхания на 11 часов 20 минут не может быть объяснена с обычной точки
зрения.
Действительно, с точки
зрения обычной биологии все так, но, похоже, это дает нам ответ только на
вопрос, что станет, если энергия не будет производиться.
Я думаю, есть нечто, что
в будущем должно найти научное подтверждение. Это существование другого вида
энергии, которая называется праной. Мы практически приходим к выводу, что
существование праны хорошо сохраняет плоть, даже если происходит остановка
дыхания или сердечной деятельности. Мы добьемся ясности в этом вопросе, если
будем упорно стремиться к обоснованию практической стороны научными методами и
при условии повышения чувствительности современных измерительных приборов.
Праной можно назвать
особую энергию, которая возникает вследствие работы души. Например, если чья-то
душа будет приближаться к божественному состоянию, то на практике он сможет
испытать подъем праны. Или же простой человек может испытать в определенном
смысле вредное влияние праны. Например, из-за заражения вирусом СПИДа происходит
понижение наиболее негативно воздействующей праны, Апана-ваю. В результате
болезнь прогрессирует и в конце концов приводит к расстройству желудка,
бессилию, повреждению рассудка и смерти. Поэтому я полагаю, что если мы сможем
контролировать прану психически и при этом измерять ее научными методами, то
человечество поднимется еще на один уровень в своем развитии[1].
2. Укажите этапы устичной транспирации. Закон
Стефана. Явление краевой диффузии
Рассмотрим этапы устичной
транспирации на примере листа. Лист снаружи и снизу покрыт эпидермой, между
слоями которой находятся мякоть листа - мезофилл, представленный
хлорофиллоносной паренхимой, а также сосудисто-волокнистые пучки и механическая
ткань. Клетки кожицы защищают лист от высыхания, механических и других повреждений,
проникновения микроорганизмов. Они не содержат хлоропластов, покрыты кутикулой
или восковым слоем.
Устьице располагаются
преимущественно на нижней стороне листа и обеспечивает транспирацию и
газообмен. Замыкающие клетки устьиц содержат хлоропласты, и при освещении а них
начинается фотосинтез, продукты которого приводят к повышению осмотического
давления. Вследствие притока воды стенки этих клеток растягиваются и устичная
щель закрывается. В темноте и в жаркую погоду при усиленном испарении воды
устьица закрываются.
Клетки мезофилла,
содержащие большое количество хлоропластов, у большинства растений
дифференцируются на столбчатую и губчатую ткани. Столбчатая ткань примыкает к
верхней кожице, а губчатая - эпидермису нижней стороны листа. Губчатая ткань
состоит из нескольких слоев клеток округлой или извилистой формы с большими
межклетниками. Такая структура наилучшим образом обеспечивает функции
транспирации и газообмена а тканях листа. Приводящие ткани входят в состав
жилок листа. В верхней части жилки расположены сосуды ксилемы, в нижней -
флоэма, В составе жилки находится механическая ткань, которая наряду с жилками
обеспечивает упругость и эластичность листа.
Кроме того, эти ткани
выполняют функцию отражающей изоляции, действие которого описывается одним из
законов физики – законом Стефана (законом Стефана-Больцмана).
Полная энергия,
излучаемая абсолютно черным телом в 1
секунду пропорциональна четвертой степени абсолютной температуры (закон
Стефана- Больцмана). Постоянная Стефана-Больцмана была определена опытным
путем. В системе СГС она выражается в эрг/(см2 сек град). Пользуясь известным
значением, Планк впервые определил постоянную h (постоянная Планка).
Из закона Стефана-Больцмана
следует, что количество теплоты, передаваемое единицей поверхности абсолютно
черного тела, находящегося при температуре T1, в окружающую среду, имеющую
температуру T2, если среду можно рассматривать как абсолютно черное тело.
Излучение всех остальных
тел подчиняется такой же закономерности, их излучение для каждой длины волны в
раз меньше, чем для абсолютно черного тела.
Из функции Планка можно
сделать вывод о распределении излучения по длинам волн.
Иными словами, длина
волны, на которую приходится максимум интенсивности излучения с увеличением
температуры смещается в сторону коротких длин волн (первый закон Вина).
На основе закона Стефана
опишем явление краевой диффузии.
Питание бессосудистой
роговой оболочки происходит путем диффузии из краевой петлистой сети,
находящейся вокруг лимба и образованной веточками передних цилиарных артерий,
путем осмоса за счет влаги передней камеры, отчасти кислорода воздуха и слезной
жидкости. При воспалительных процессах в роговицу врастают сосуды из указанной
петлистой сети.
Чем светлее и равномернее
цвет ядра и зоны диффузии, тем работоспособнее масло. При росте мехпримесей
темнеет ядро и зона диффузии, что приводит к потере краевой зоны. При потере
присадок уменьшается зона диффузии, расширяется внешнее светлое кольцо.
На процесс краевой диффузии
влияет температура окружающей среды (чем она выше - тем выше скорость
диффузии), материал герметика, толщина уплотняющей массы и ширина зазора для
уплотнителя. По результатам многочисленных опытов установили, что наилучшей с
точки зрения диффузионной плотности является двухстадийная герметизация на
полисульфидной массе. При подобном методе герметизации количество
газа-наполнителя (аргона) снижается на 0,4 - 2 % в год, тогда как при других
решениях снижение достигает нескольких процентов.
3. Напишите схему и дайте характеристику цикла
Кальвена

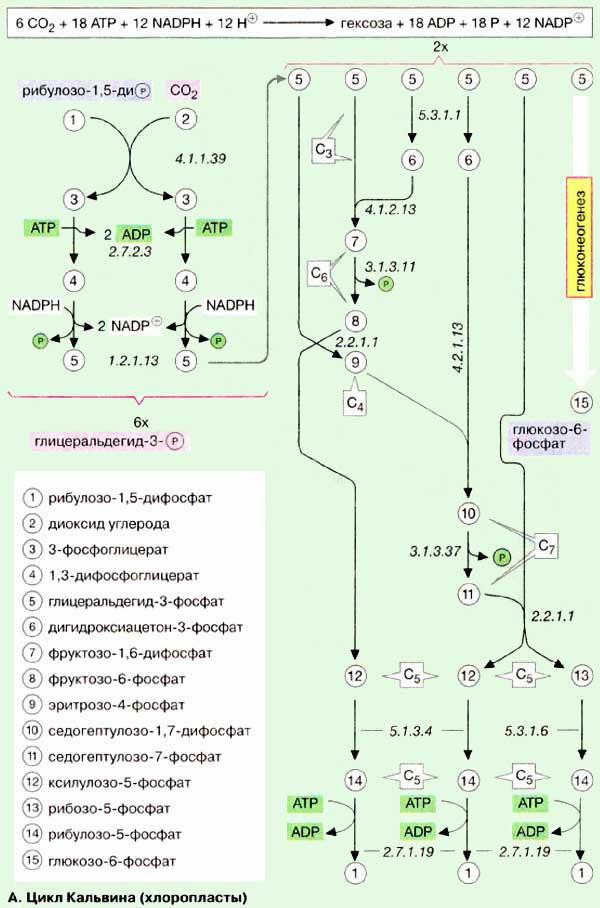
В листьях растений
присутствуют пероксисомы, метаболизирующие гликолат. Гликолат образуется в
процессе фотодыхания, охарактеризованном В.И. Чиковым в. При фотодыхании часть
углерода, ассимилированного при фотосинтезе, выделяется в виде СО2,
а часть возвращается в фотосинтетический цикл Кальвина.
В стареющих листьях
фотодыхательные пероксисомы нагружаются ферментами глиоксилатного цикла и
окисления жирных кислот, а также протеазами. Они участвуют в деградации
мембранных липидов и белков отмирающих клеток. Такие органеллы были названы
геронтосомами.
Уреидные пероксисомы,
присутствующие в клубеньках бобовых, участвуют в процессе азотфиксации в
бактероидах, окисляя продукты пуринового обмена клубенька: мочевую кислоту
(урат) и ксантин. Ксантиноксидаза обнаружена у растений и в других типах
пероксисом. Уреидные пероксисомы играют роль в интеграции метаболизма
бактероида и растения-хозяина. Особые формы пероксисом (например, окисляющие
метанол или щавелевую кислоту) имеются у грибов.
Окисление жирных кислот
сопровождается образованием пероксида водорода, который разрушается каталазой.
В этом состоит отличие пероксисомального окисления жирных кислот от
митохондриального, в котором электроны переносятся в дыхательную цепь
митохондрий, а пероксид в существенной концентрации не образуется. В растениях
митохондриальное окисление жирных кислот, хотя и происходит, но его
интенсивность низкая. Мобилизация запасных жиров наблюдается только в
глиоксисомах.
В глиоксилатном цикле,
также протекающем в глиоксисомах, образовавшийся при окислении жирных кислот
двууглеродный остаток уксусной кислоты, связанный с коферментом А (ацетил-КоА),
превращается в ходе пяти ферментативных реакций в четырехуглеродную молекулу
янтарной кислоты (сукцината). Два фермента - изоцитратлиаза и малатсинтаза -
функционируют только в глиоксилатном цикле. Три других - цитратсинтаза,
аконитаза и малатдегидрогеназа - общие с митохондриальным циклом Кребса, хотя в
глиоксилатном цикле они представлены другими молекулярными формами
(изоферментами). Все ферменты глиоксилатного цикла (возможно, за исключением
аконитазы, локализованной в цитозоле) присутствуют, вероятно, только в
глиоксисомах высших растений. При протекании глиоксилатного цикла у водорослей
и грибов в глиоксисомах выявляется только активность изоцитратлиазы и
малатсинтазы, тогда как ферменты, общие с циклом Кребса, - цитратсинтаза,
аконитаза и малатдегидрогеназа - в глиоксисомах этих организмов отсутствуют.
Глиоксилатный цикл
является необходимым звеном превращения жиров в углеводы. Этот процесс имеет
особое значение в быстрорастущих тканях: запасные липиды используются для
синтеза углеводов, идущих на образование клеточной стенки и другие процессы.
Однако этим роль глиоксилатного цикла не исчерпывается: в нашей лаборатории
показано, что он обеспечивает образование органических кислот, подкисляющих
эндосперм злаков, что тем самым делает возможным мобилизацию запасного
крахмала, и участвует в синтезе аминокислот. Процессы, связанные с
глиоксисомой, обеспечивают взаимодействие клеточных компартментов. Синтез
КоА-производных жирных кислот, вероятно, протекает на поверхности олеосомы
(жирозапасающей органеллы), окисление жирных кислот и глиоксилатный цикл - в
глиоксисоме, превращение сукцината в щавелевоуксусную кислоту (оксалоацетат) -
в митохондриях, дальнейшие стадии синтеза углеводов (обращенный гликолиз) - в
цитоплазме[2].
Гены, кодирующие ферменты
глиоксилатного цикла, имеются, вероятно, у всех организмов, но экспрессируются
не во всех тканях и не на всех стадиях онтогенеза. При старении листьев
глиоксилатный цикл обеспечивает протекание биодеградативных процессов
(например, окисление ацетил-КоА, образующегося в ходе деградации мембранных
липидов). Образование пероксида водорода и супероксидных радикалов в ходе
метаболизма пероксисом также играет важную роль в биодеградации.
Считалось общепринятым,
что у высших животных глиоксилатный цикл отсутствует. И только совсем недавно в
нашей лаборатории было показано, что в условиях голодания ферменты
глиоксилатного цикла появляются в печени (у крыс на третий день голодания),
меньшая их активность выявляется и в других тканях. Это обеспечивает
превращение запасных жиров в углеводы, поскольку иных источников углеводов в этих
условиях организм не имеет.
Пероксисомы
фотосинтезирующих тканей метаболизируют гликолевую кислоту (гликолат). Гликолат
образуется из фосфогликолевой кислоты в хлоропластах и транспортируется в
пероксисому. Метаболизм гликолата в фотосинтезирующих клетках составляет
процесс фотодыхания, подробно рассмотренный В.И. Чиковым[3].
Классическая схема
метаболизма гликолата включает согласованную работу хлоропласта, пероксисомы и
митохондрии, и на электронных микрофотографиях нередко наблюдается ассоциация
пероксисом с этими органеллами. Процесс фотодыхания связан с оксигеназной
реакцией центрального фермента фотосинтеза - рибулозобисфосфаткарбоксилазы
(Рубиско), функция которого состоит в фиксации углекислого газа. Присоединение
молекулы СО2 к субстрату данного фермента, рибулозо-1,5-бисфосфату,
- первая реакция фотосинтетического цикла углерода (цикла Кальвина).
Она приводит к образованию двух молекул фосфоглицериновой кислоты. Механизм
фотосинтетической фиксации углерода описан В.В. Климовым.
Фермент Рубиско помимо
сродства к СО2 имеет и некоторое сродство к кислороду, в результате
чего вместо двух молекул фосфоглицериновой кислоты образуются одна молекула
фосфоглицериновой кислоты (идущая в цикл Кальвина) и одна
молекула фосфогликолевой кислоты (с нее начинается фотодыхательный гликолатный
цикл). В ходе реакций фотодыхательного метаболизма поглощается кислород и
выделяется углекислый газ (в митохондриях при окислении глицина), и потому
имеется аналогия с дыханием. Отсюда название «фотодыхание», так как процесс протекает
на свету и связан с фотосинтетическим метаболизмом.
От фосфогликолевой
кислоты в хлоропластах отщепляется фосфатная группа. Образующаяся гликолевая
кислота транспортируется в пероксисому. В пероксисомах фотосинтезирующих тканей
гликолат окисляется гликолатоксидазой (с образованием глиоксилата и пероксида
водорода, разрушаемого каталазой). Глиоксилат затем аминируется с образованием
глицина, что происходит посредством переноса аминогруппы глутамата или серина.
Глицин транспортируется в митохондрию. Здесь две молекулы глицина окисляются с
образованием одной молекулы серина в реакции, катализируемой
глициндекарбоксилазным комплексом. Содержание данного комплекса может
составлять до трети и даже половины всех белков матрикса растительных
митохондрий. Реакция сопровождается выделением аммония (который рефиксируется в
хлоропласте), углекислого газа и восстановлением митохондриального НАД.
Образующийся серин
возвращается в пероксисому, где вступает в реакцию трансаминирования. В этой
реакции образуется гидроксипируват, который восстанавливается
гидроксипируватредуктазой с образованием глицериновой кислоты. Последняя
транспортируется из пероксисомы в хлоропласт, где превращается в
фосфоглицериновую кислоту, возвращающуюся в цикл Кальвина. Таким
образом, из двух молекул двууглеродной гликолевой кислоты один атом углерода
выделяется в виде СО2, а три возвращаются в цикл Кальвина
в форме фосфоглицериновой кислоты.
4. Охарактеризуйте этапы поступления ионов в
клетку, их значение
Поглощение ионов из почвы
или питательного раствора корневыми системами осуществляется разными путями, от
которых зависит вероятность поступления иона непосредственно в цитоплазму
клеток и скорость его перемещения по тканям и органам растений.
Иммобилизация ТМ в
корневой системе может происходить за счет неметаболического необратимого
связывания их ограниченным числом участков, расположенных на поверхности
клеточной стенки и вдоль симпласта корней[4]. С
одной стороны, необратимое связывание предотвращает передвижение части
токсичных ионов по клеткам и тканям растения, с другой – этот процесс
способствует установлению концентрационного градиента и позволяет в дальнейшем
аккумулировать элемент посредством диффузии.
В случае обратимого
связывания иона компонентами клеточной стенки путем обменной физико-химической
сорбции, он может пассивно диффундировать в симпласт при наличии
концентрационного градиента, что обеспечивает транспорт иона по растению.
Участки, задействованные в обменной адсорбции, не обладают селективностью. С их
участием поступают Cd, Zn, Cu, Hg и другие металлы. Накопление ионов металлов в
свободном пространстве клеточной стенки определяется величиной ионообменного
коэффициента, которая в значительной степени зависит от количества гистидильных
групп белков, а также карбоксильных групп, размещенных на поверхности пектинов.
Обменно-связанная фракция ионов металла легко вымывается из корневых систем
растений при замене питательного раствора на не содержащий данный элемент.
Проникая через клеточную
стенку, одна часть ионов связывается с реактивными компонентами апопласта,
другая поступает в цитозоль. Существуют данные, свидетельствующие, что
двухфазный характер поступления ионов металлов, например Cd, в корневые системы
отражает два вышеуказанные процесса. Причем линейный участок зависимости
поглощения иона металла от времени (для корней пшеницы – 75 мин) соответствует
связыванию Сd с компонентами апопласта, а плато (фаза насыщения) является
результатом транспорта Cd через плазматическую мембрану в клетки корней. Важно,
что в цитируемой работе линейно зависящее от времени поглощение Cd наблюдали
при крайне низких концентрациях этого элемента в растворе – от 2.2 до
56.2 мкг/л, тогда как в случае высокого (более 1 г/л) его содержания в
субстрате быстро достигался выход на плато. Отсюда следует, что микроколичества
Cd и, видимо, других ТМ могут эффективно задерживаться клеточной стенкой или
реактивными центрами апопласта. Эти же механизмы предотвращают поступление
избыточных количеств ТМ в клетки и ограничивают передвижение ионов металлов по
тканям растений. Данный факт имеет большое значение при выборе концентраций
токсиканта для исследования его генетических эффектов действия на растения.
В случае повышенных
концентраций ТМ в питательной среде указанные механизмы не могут полностью
исключить поступление ионов через плазматическую мембрану в цитозоль. Транспорт
молекул через мембраны осуществляется разными путями в зависимости от
химических свойств элементов и их биологической значимости. Небольшие
нейтральные молекулы могут проникать через мембраны за счет обычной диффузии.
Скорость диффузии вещества определяется его растворимостью в мембране,
коэффициентом диффузии в мембране и градиентом концентраций снаружи и внутри
клетки[5].
Поступившие в цитоплазму
ионы могут связываться там с биомолекулами. В этом случае хеллатный комплекс
либо выводится из клетки, либо аккумулируется в ней (чаще всего в вакуолях).
Механизмы внутриклеточной детоксикации будут подробно рассмотрены в следующем
разделе.
Оставшаяся в цитозоле в
виде свободных ионов или растворимых комплексов фракция симпластическим или
трансклеточным путем перемещается из корня в побег и далее – в листья растений
по заряженным участкам ксилемы, либо увлекается с транспирационным потоком воды.
Список литературы
1. Ботаника / Под ред. С.И. Трунева. М.:
Агороиздат.
2. Землянухин А.А., Землянухин Л.А.,
Епринцев А.Т., Игамбердиев А.У. Глиоксилатный цикл растений. Воронеж: Изд-во
ВГУ, 1986.
3. Мецлер Д. Биохимия. Химические
реакции в живой клетке. М.: Мир, 1980.
4. Перцов Л.А. Ионизирующие излучения
биосферы. М.: Атомиздат, 1973.
5. Чиков В.И. Фотодыхание // Соросовский
Образовательный Журнал. 1996. № 11.
[1] Ботаника / Под ред. С.И.
Трунева. М.: Агороиздат. С. 99-100.
[2]
Землянухин А.А., Землянухин Л.А., Епринцев А.Т., Игамбердиев А.У. Глиоксилатный
цикл растений. Воронеж: Изд-во ВГУ, 1986. С. 148.
[3]
Чиков В.И. Фотодыхание // Соросовский
Образовательный Журнал. 1996. № 11.
C. 2-8.
[4] Перцов Л.А. Ионизирующие излучения биосферы. М.:
Атомиздат, 1973. С. 287.
[5] Мецлер Д. Биохимия. Химические реакции в живой клетке. М.: Мир, 1980.
С. 47.