Одной из целей
теории науки является установление структуры научного мышления, научного
поиска. Научное познание включает в себя множество компонентов. Каждый из
которых связан с одним из двух уровней эмпирическим или теоретическим.
Основными
элементами научного знания являются:
факты твердо
установленные и подтвержденные в ходе наблюдений экспериментов и проверок,
будучи зафиксированы принятыми в науке способами но превращаются в научный
факт. Они составляют эмпирический базис науки, здесь исследователь имеет дело
непосредственно с природными и социальными объектами, применяются методы
наблюдения описания, измерения, эксперимента;
закономерности,
обобщающие группы фактов – существенные, необходимые, устойчивые и
повторяющиеся связи явлений, выявляются путем абстрагирования, выведения в
обобщенном виде признаков являющихся общим для многих вещей одного класса, но
не как простая одинаковость, а как сущностная связь многих предметов,
превращающая их в аспекты единой системы. Для этого требуются понятия, которые
создаются заново или заимствуются из смежных отраслей знания. В науках которые
принято называть описательными, н-р в геологии
эмпирические обобщения исследование завершают, теория в этих науках
представляет собой эмпирическое обобщение, построенное на чрезвычайно обширном
материале
теории
представляющие собой системы закономерностей и описывающие некий фрагмент
реальности в теории происходит перестройка эмпирического материала на основе
некоторых исходных принципов, на теоретическом уровне объектом исследования
являются идеализированные объекты (теоретические абстракции, математические
модели и т. д., поэтому применяются аксиоматический метод, системный и
структурно-функциональный анализ, математическое моделирование; Теория
рассматривается как высшая форма организации научного знания, дающей целостное
представление о существенных связях и отношениях в какой-либо реальности. Так
как разработка теории связана, как правило, с введением понятий, фиксирующих
непосредственно не наблюдаемые стороны объективной реальности, проверка
истинности теории не может быть непосредственно осуществлена прямым наблюдением
или экспериментом. Это приводит к тому что возникает вопрос о том право данной
конкретной теории считаться научной, как доказать что она не является чистой
спекуляцией Проверка гипотезы строится на несколько иных принципах, В истории
науки было предложено два принципа позволяющих провести границу между научными
теориями и тем, что наукой не является. 1. принцип верификации любое понятие
или суждение имеет научный смысл если оно может быть сведено к эмпирически
проверяемой форме, или оно само не может иметь такой формы, то эмпирическое
подтверждение должны иметь ее следствия, одна принцип верификации применим
ограниченно, в некоторых областях современной науки его использовать нельзя.2.
Американский философ К. Поппер предложил другой принцип – принцип
фальсификации, в его основе лежит тот факт, что прямое подтверждение теории
часто затруднено невозможностью учесть все частные случаи ее действия, а для
опровержения теории достаточно всего одного случая с ней не совпадающего,
поэтому если теория сформулирована так, что ситуация в которой она будет
опровергнута может существовать, то такая теория является научной. Теория
неопровержимая в принципе не может быть научной
научные картины
мира, рисующие обобщенные образы реальности в целом, в теории допускающие
взаимное согласование сведены в некое системное единство.
Таким образом мы
можем выделить два взаимосвязанных но самостоятельных уровня научного
исследования: эмпирический и теоретический. Они различаются по предмету (вовтором случае он может иметь свойства, которых нет у
эмпирического объекта), средствам, методам и результатам (эмпирическое
исследование завершается выведением эмпирического обобщения, теоретическое
имеет целью выдвижение гипотезы и создание теории). Но несмотря на
методологическую ценность выделения эмпирического и теоретического уровней
исследования, разделить их в целостном процессе познания невозможно. Невозможно
так же говорить о том что какой-то из них является важнейшим ведущим для
исследования. Еще на заре развития современной науки Ф. Бекон и Р. Декарт
сформулировали две разнонаправленных программы развития науки: эмпирическую,
основанную на принципе индукции (такой способ рассуждения при котором общий
вывод делается на основе частных посылок) и рационалистическую построенную на
дедукции (выведение всей системы знания из некоторых общих положений, который
носят самоочевидный характер). Обе эти методологические программы в настоящее
время считаются неадекватными требованиям научного исследования. Эмпиризм имеет
тот недостаток, что индукция не может привести к универсальным суждениям,
поскольку в большинстве ситуаций принципиально невозможно охватить все
многообразие частных случаев, рационализм оказался беспомощен при изучении тех
областей реальности, для которых ничего самоочевидного не существует. В наше
время стандартная модель строения научного знания начинается с установления
путем наблюдения и эксперимента различных фактов, если среди них можно
установить некую регулярность, то можно говорить об обнаружении эмпирического
закона, первичного эмпирического обобщения, если отыскиваются факты, которые не
встраиваются в обнаруженную регулярность, то ученый используя свой творческий
интеллект, умение мысленно перестроить известную реальность так что бы
устранить противоречие. Эмпирически выявить эту новую схемы невозможно, ее
необходимо сотворить умозрительно в виде гипотезы, если эта гипотеза снимает
противоречие или позволяет предсказать получение новых данных, означает что она
нашла подкрепление и превратилась в теорию. Такой метод принято называть гипотетико –дедуктивным.

2.Основные
теоретические модели развития науки.
Развитие науки
определяется внешними и внутренними факторами. К первым относится влияние
государства, экономичеких, культурных, национальных
факторов и ценностных установок ученых. Вторые определяются внутренней логикой
развития науки. Развитие научного знания отличается своебразной
неровностью, причудливым переплетением медленного накопления данных и резкими измененниями связанными с внедрением радикально новых идей
вызывающим обвальное изменение всей естественнонаучной картины мира. Внутренняяя логика имеет свои особенности на каждом из
уровней исследования, эмпирическому уровню присущь
кумулятивный характер, поскольку даже отрицательный результат наблюдения или
эксперимента вносит свой вклад в накопление знаний. Теоретический уровень имеет
более скачкообразный характер, так как каждая новая теория представляет собой
качественное преобразование свей системы знания, как правило новая теория не
отрицает предшествующую полностью, а включает ее в качестве частного случаю.
Вопрос о смене научных концепций является одним из наиболее интересных в
современной методологии науки. В первой половине XX века основной структурной
единицей развития науки считалась теория, и вопрос о ее смене ставился в
зависимость от ее подтверждения или опровержения. Главной методологической
проблемой считалось сведение теоретического уровня исследования к
эмпирическому, что в конце-концов оказалось невозможным.
В 60-е Огромное
влияние на представления о логике и динамике развития науки оказала
американского историка и философа Томаса Куна. Он ввел в методоллогию
науки приннципиально новое понятие «парадигма» (от
греч. образец). Парадигма фиксирует определенный способ организации знания,
подразумевающий набор предписаний задающих видение мира, и т.о. влияющий на
определение направлений исследования, в парадигме содержатся так же обще
принятые образцы решения конкретных проблем, парадигма это не теория (хотя как
правило ее ядро составляет фундаментальная теория), она не выполняет
объяснительной функции, парадигма это способ задать систему отсчета,
предварительная предпосылка построения и обоснования различных теорий. Это
место теоретическое образование, закрепляющее дух и стиль научных исследований.
Кун определял парадигму как «признанные всеми научные достижения, которые в
течение определенного времени дают модель постановки проблем и их решения
научному сообществу». Парадигма принятая научным сообществом может на долгие
годы определить круг проблем, привлекающих внимание ученых, является
официальным подтверждением, подлинной научности получаемых ими данных. В рамках
действия одной парадигмы приращение знаний осуществляется по схеме «нормальной
науки», смена парадигмы сопровождается «научной революцией». В ходе «научной
революции выбор новой парадигмы не является однозначно детерминированной , ен носит линейного характера, выбор парадигмы из числа
возможных осуществляется в значительной случайно под воздействием совершенно
посторонних факторов, поэтому почти мгновенный акт смены парадигмы не может
быть истолкованы строго рационально, Кун даже склонен сравнивать это явление с
возникновением новой религии, как некое просветление и озарение, иррациональный
акт веры.
Данное положение
было подвергнуто критике американским философом венгерского происхождения И. Лакатосом, который предложил в качестве основной единицы
развития научного знания не нормальную науку, основанную на той или иной
парадигме, «научно-исследовательскую программу».
Структура
научно-исследовательской программы:
«жесткое ядро»,
включающее неопровержимые для сторонников программы исходные положения.
область
«негативной эвристики» - это своеобразный защитный пояс ядра программы,
состоящий из вспомогательных гипотез и допущений, снимающих противоречия с
фактами не согласующимися с ядром программы (например, расхождение между
расчетным положением и реальной ситуацией объясняется неточностью измерения,
присутствием не установленных возмущающих факторов, но фундаментальные
положения сомнению не подвергаются.
«Позитивная
эвристика» - правила указывающие как выбирать пути исследования и как по ним
следовать то есть, это ряд положений на основании которых можно изменять и
пересматривать те варианты исследовательской программы которые оказались под
угрозой опровержения, в результате исследовательская программа предстает не как
изолированная тория как серия модифицированных теорий в основе которых лежат
общие исходные принципы.
Благодаря этому
ученые работающие внутри какой-то исследовательской программы могут долгое
время обходить противоречащие программе факты, они могут ожидать что данные, не
укладывающиеся в программу в данный момент, удастся объяснить со временем, это
придает науке стабильность. Однако рано или поздно позитивная эвристика себя
исчерпывает, вытеснение одной программы другой представляет собой научную
революцию, выбор между двумя программами осуществляется рационально, на основе
того что одна из них признается прогрессирующей, а другая регрессирующей,
программу признается прогрессирующей если ее теоретический рост превышает ее
эмпирический, то есть если она успешно предсказывает новые факты, программа
регрессирует тогда когда ее теоретический рос отстает от эмпирического, то есть
она только запоздало объясняет случайные открытия или факты, открываемые
конкурирующей программой. Таким образом, источником развития науки выступает
конкуренция исследовательских программ.
3.Единство
интегративный-дифференциальных тенденций развития науки.
Важной
закономерностью процессов развития науки принято считать единство процессов диффренциации и интеграции научного знания.
В настоящее
время насчитывается не менее 15 тыс. различных научных дисциплин. Такое
усложнение структуры научного знания имеет несколько причин, во-первых, в
основе всей современной науки лежит аналитический подход к действительности, то
есть основной прием познания это расчленение изучаемого явления на простейшие
составляющие. Этот методологический прием ориентировал исследователей на
подробную детализацию изучаемой действительности. Во-вторых за последние 300
лет резко возросло число объектов доступных для научного изучения,
существование универсальных гениев, которые могли охватить все многообразие
научного знания стало сейчас физически невозможным, человек способен познать лишь
незначительную часть того что известно человечеству. Процесс формирования
отдельных научных дисциплин происходил за счет отграничения предмета каждой из
них от предметов других наук. Стержнем того или иного предмета исследования
являются объективные законы действительности.
Подобная
специализация полезна и неизбежна. Дифференциация научного знания позволяет
более глубоко изучить, отдельные аспекты реальности. Она облегчает труд ученых,
оказывает влияние на саму структуру научного сообщества. Данный процесс
продолжается и по сей день, генетика относительно молодая наука, но в ней уже
наметилось целое семейство дисциплин: эволюционная, популяционная,
молекулярная. Продолжают дробиться и более старые науки, например, в химии
появились квантовая химия, радиационная химия и т. д. Но в то же время
дифференциация научного знания несет в себе и опасность, разложения единой
научной картины мира. Отпочковавшись от системы протознания,
дисциплины оказывались в изоляции друг от друга, элементы науки (отдельные
научные дисциплины) становились самодовлеющими в своей автономности,
естественные связи между ними нарушались, структурные взаимодействия исчезали.
Это было характерно не только для отношений между крупными отраслями знания, но
и внутри отраслевых рамок отдельных наук. В результате наука из целостной
системы знания все больше превращалась в суммативную.
Взаимное размежевание наук, дифференциация изоляционистского типа была ведущей
тенденцией в сфере науки в плоть до XIX века, это привело к тому, что не смотря
на большие успехи достигнутые наукой на пути прогрессирующей специализации,
происходил рост рассогласования научных дисциплин. Возник кризис единства
науки. Но уже в рамках классического естествознания постепенно утверждается
идея принципиального единства всех явлений природы, а следовательно и
отражающих их научных дисциплин, поэтому начали возникать смежные научные
дисциплины например, физическая химия, биохимия. Границы проведенные между
оформившимися научными дисциплинами становятся все более условными, фундаментальные
науки так глубоко проникли друг в друга, что возникает проблема формирования
единой науки о природе, то есть интеграции научного знания.
Термин
интеграция (от лат. восстановление, восполнение), как правило используется для
обозначения объединения каких-то частей в единое целое, при этом
подразумевается так же преодоление дезинтегрирующих факторов ведущих к
разобщенности системы, к чрезмерному росту самостоятельности элементов или
частей, что должно повысить степень упорядоченности и организованности системы.
сейчас этот термин уже утвердился в качестве общенаучного понятия: некоторые
исследователи даже предлагают рассматривать его как философскую категорию. В
основе решения проблемы интеграции научного знания лежит философский принцип
единства мира. Поскольку мир един его адекватное отражение должно представлять
единство; системный целостный характер природы обуславливает целостность
естественнонаучного знания, в природе нет абсолютных разграничительных линий. А
есть только относительно самостоятельные формы движения материи, переходящие
друг в друга и составляющие звенья единой цепи движения и развития, по этому
науки изучающие их могут обладать не абсолютной, а только относительной
самостоятельностью.
в организации
исследований на стыке наук. Результатом являются «пограничные» науки, но такая
интеграция возможна только между смежными дисциплинами
в разработке
междисциплинарных научных методов, которые могут применяться в различных науках
(спектральный анализ, хроматография, компьютерный эксперимент, еще боле широкую
интеграцию позволяет осуществлять применение математического метода)
в поиске
объединительных теорий и принципов, к которым можно было бы свести бесконечное
разнообразие явлений природы (единая теория поля, глобальный эволюционный
синтез в биологии, физике химии и т.д.)
разработка
теорий выполняющих общеметодологические функции в естествознании. В результате
возникаю синтезирующие дисциплины объединяющие ряд далеко отстоящих друг от
друга наук. (кибернетика, синергетика).
изменении самого
принципа выделения научных дисциплин. Появился новый тип проблемных наук, они
по большей части становятся комплексными, привлекающими для решения одной
проблемы сразу несколько дисциплин (онкология и пр.).
В настоящее
время можно проследить в науке одновременно и процессы дифференциации и
процессы интеграции, но последние судя по всему пересиливают, интеграция стала
ведущей закономерностью развития научного прогресса. К настоящему времени в
науке действует множество интегрирующих факторов, которые позволяют утверждать,
что она стала целостным системным образованием, в этом отношении наука вышла из
кризиса, и проблема состоит теперь в достижении еще большей организованности и
упорядоченности. В современных условиях дифференциация наук уже не приводит к
дальнейшему разобщению, а наоборот к их взаимному цементированию. Однако
разобщение еще далеко не преодоленно, а на отдельных
участках оно даже усиливается. При этом следует учитывать что интеграция и
дифференциация не взаимоисключающие, а взаимодополняющие процессы.
4.Особенности
современных методов научного познания. Применение научных методов в
естествознании.
Структура
научного познания, о которой мы говорили выше, представляет собой способ
научного познания или научный метод. Метод это совокупность действий,
призванных помочь достижению желаемого результата. Современная наука
основывается на определенной методологии – то есть совокупности используемых
методов и учений о методе. Система методов научного исследования включает в
себя, во-первых, методы применяемые не только в науке, но и в других отраслях
знания, во-вторых, методы применяемые во всех отраслях науки и. В-третьих,
методы специфические для отдельных определенных разделов науки, отдельных
научных дисциплин.
Среди всеобщих
методов, характерных для всех сфер человеческого познания можно выделить такие
методы как:
анализ –
расчленение целостного предмета на составные части (стороны признаки отношения)
с целью их более глубокого изучения.
синтез –
соединение ранее выделенных частей предмета в единое целое.
абстрагирование
– отвлечение от ряда несущественных для данного исследования свойств и
отношений с одновременным выделением интересующих нас свойств и отношений.
обобщение –
прием мышления – в результате исследования, в результате которого общие
свойства и признаки целого класса объектов.
индукция.
дедукция.
аналогия – прием
познания при котором на основе сходства объектов по одним признакам делается заключение
об их сходстве по другим.
моделирование –
изучение объекта (оригинала) путем создания и исследования его копии (модели),
воспроизводящей оригинал с определенной точки зрения интересующей
исследователя.
классификация –
разделение всех изучаемых предметов на отдельные группы в соответствии с
каким-либо важным для исследователя признаком ( особое значение имеет в
описательных науках6 геологии, географии, некоторых разделах биологии).
Научный метод
как таковой разделяется на методы используемые на каждом уровне исследования.
Таким образом, выделяются эмпирические и теоретические методы.
Эмпирические
методы:
наблюдение –
целенаправленное восприятие явлений объективной действительности.
описание –
фиксация средствами естественного или искусственного языка сведений об
объектах.
измерение –
сравнение объектов по каким-либо сходным свойствам или сторонам.
эксперимент –
наблюдение в специально создаваемых и контролируемых условиях, он позволяет
восстановить ход явления при повторении условий.
Научные методы
теоретического уровня исследования:
формализация –
построение абстрактно-математических моделей, раскрывающих сущность изучаемых
процессов действительности.
аксиоматизацию –
построение теорий на основе аксиом (утверждений доказательства истинности
которых не требуются).
гипотетико-дедуктивный
метод – создание систем дедуктивно связанных между
собой гипотез, из которых выводятся утверждения об эмпирических фактах.
В конце
XVII-XVIII веках сначала физика в исследованиях И. Ньютона, затем химия в лице
Лавуазье встали на путь количественного исследования, затем то же произошло и в
других естественно научных дисциплинах. Применения математики, столь же
характерно для современного естествознания как применение экспериментальных
методов, логическая стройность, строго дедуктивный характер построений,
общеобязательность выводов математики, делали ее прекрасной опорой для
естествознания. Достоинство математизации естество знания чрезвычайно
многообразны. Во-первых, во многих случаях математика играет роль универсального
языка естествознания, прекрасно подходящего для лаконичной и точной фиксации
различных положений. Во-вторых, математика может служить источником моделей,
алгоритмических схим, для связей, отношений и процессов, составляющих предмет
естествознания. Разумеется любая математическая модель это своего рода
упрощение, но упрощение в данном случае не тождественно огрублению, это скорее
выявление сущностных особенностей объекта. Поскольку в математических формулах
и уравнениях воспроизведены некие общие связи и отношения, реального мира, они
могут повторяться в разных его областях. На этом построен метод естественно
научного исследования, который называют математической гипотезой, в ней не
создают математическое описание природных объектов, а пытаются готовой
математической модели подобрать соответствие в природе. Часто исходная
математическая формула заимствуется из смежной и даже не смежной области
знания, в нее подставляются значения, иной природы, а затем проверяют,
совпадение рассчитанного и реального поведения объекта. Разумеется,
применимость этого метода ограничена теми дисциплинами, которые уже накопили,
достаточно богатый математический арсенал. В целом значение математики в
современном естествознании невозможно переоценить, сейчас ни одна теоретическая
интерпретация не считается полностью завешенной, если не удается создать
математическую модель изучаемого явления. Однако не следует думать, что все
естествознание может быть сведено к математике, построение формальных систем,
моделей, алгоритмических схем, это только метод, одна из сторон научного
поиска. Развивается наука прежде всего как содержательное, неформализованное, неалгоритмизироанное знание.
Большое значение
в современной науке приобрели статистические методы, позволяющие определять средние
значения характеризующие всю совокупность изучаемых явлений или предметов.
Применение статистического метода не позволяет ученым предсказывать поведение
отдельного индивида в совокупности, можно лишь утверждать, что он будет вести скбя определенным образом с определенной вероятностью.
Статистические законы применяются только к большим совокупностям.
|
|
Валентинас Мураускас
Вильнюс
2003-10-22
Пространство -
основа или параметр осознания.
|
|
|
В XXI век
западная цивилизация вступила с громадным багажом научного знания в
технологическом порыве совершенствовать мир и наши жизни. Большинство
уверенно, что на базе научного знания построенная картина мира вполне
конструктивна и требует только уточнений и детализации. Предчувствие
неведомого и глобальных открытий стало синонимом романтики ученных и
мыслителей прошлых веков. Даже область фундаментальных исследований приобрела
прикладную или технологическую наклонность. Критерием ценности научного
поиска стало не познание, как таковое, а применимость открытий в совершенствовании
технологий.
Научная
картина мира стала неотъемлемой частью удобств и удовлетворенности собой
нашей цивилизации. По этому любая попытка критического взгляда на научную
картину мира принимается как покушение на устои западной цивилизации и ниспровергается
давлением авторитетов. Не смотря на предполагаемую реакцию, предпосылок для
внимательного рассмотрения научной парадигмы достаточно. Любой профессионал
"переднего фронта" науки сможет перечислить целый ряд застарелых
парадоксов, принципиально не решаемых в рамках существующей научной картины
мира. Другой, с точки зрения авторитетов от науки "грязный" довод,
указывающий на неполноту научной картины мира - это мистический или
религиозный опыт. Стараясь не испачкаться об многозначность и расплывчатость,
небольшая часть этого знания была принята в науку в виде трансперсональной
психологии и антропологии. Несмотря на такое "достижение"
брезгливое отношение академической науки к этим "новым" областям
научного знания стойко сохраняется. Другие области мистического опыта
бесповоротно ставящие науку в неудобное положение, такие, как парапсихология,
религиозные и оккультные доктрины просто игнорируются или бездоказательно
отрицаются.
Внешний
наблюдатель видит тупиковую ситуацию неаргументированного отрицания на фоне
полной и обоснованной уверенности в себе с обеих сторон. В это же время
представители западной науки позволяют себе демонстрировать религиозность при
отрицании религиозных доктрин, а мистики, как правило, предпочитают удобства
технологически развитых стран и, в лучшем случае, посмеиваются над научной
парадигмой. Следовательно, "де-факто" объединение столь
несовместимых взглядов на мир уже происходит. Большей частью защитные реакции
с обеих сторон вызывает страх "быть проглоченным" в модель мира
"противника".
Однако, нет
никакой необходимости противостояния. Мистическое видение мира прошло
проверку временем и списывать его на интеллектуальную недоразвитость сегодня
уже просто глупо.
Сомневаться в
правдивости научного знания тоже может только недостаточно образованный -
достижения технологий доказали ее конструктивность. Научная модель мира,
однозначно, правильная. Возникает логический парадокс: как может правильная
модель Реальности вызывать ощущение ее неполноты, порождать парадоксы и
ставить перед выбором доверия собственному опыту или его отрицания в целях
сохранения "научности".
Выход может
обнаружиться если обратить внимание на область применимости научной модели.
Что эта область не абсолютная доказывает наличие аксиоматики. Более того, эту
мысль, хоть и с трудом, признают большинство, с кем пришлось общаться.
Значит сумму
научного знания мы в праве назвать научной моделью мира. А между моделью и
миром (точнее будет назвать Реальностью) всегда существуют некоторые
начальные допущения задающие правила или логику самого моделирования. Это
аксиомы модели. Их забыли из за отождествленности с
моделью, как аксиомы, потому, что они очевидны и само собой разумеющиеся.
Исторически сложившееся полное отождествление мыслящего человека с
доминирующей моделью мира вселяет ложную уверенность, что модель Реальности -
это видимый и ощущаемый мир. (Не это ли имеет в виду из глубин веков
пришедшие традиции знания утверждающие иллюзорность видимого мира.)
Этой мысли
можем и испугаться, но если приглядеться пристальнее, то выяснится, что этим
утверждается только одно - наличие отождествленности
нашего среднестатистического западного осознания с научной картиной мира.
Предпосылки настороженности вызывают поспешные выводы. Попробуем некоторое
время их сдерживать и вернемся к аксиоматике.
Аксиоматику
мистических моделей мира определить непросто, по этому начнем с научной
модели мира. Одна из аксиом научной модели мира - пространство. Пока не будем
касаться даже свойств пространства и остановимся на самом понятии
"пространство".
Пространство
воспринимается вместилищем процессов и нас самих. Оно кажется неотъемлемым и
бесконечным атрибутом Вселенной, который никак не связан с осознанием. Это
считается очевидным и бесспорным.
Если кто и
попытается найти что-нибудь написанное о пространстве, а не о математических математических выкладках о его кривизне или мерности, то
с удивлением обнаружит, что термина "пространство" не найти в
большей части уважаемых энциклопедий, толковых словарей и справочников. Чаще
всего мы находим только "пространство и время", определенное как
Эйнштейновский континуум с несколькими историческими ссылками. Большинство
написанного о самом пространстве с небольшой погрешностью можем свести к
следующему: пространство и время "являют собой несущую конструкцию любой
известной до сих пор объяснительной картины мира". Проще говоря,
пространство и время - это фундамент, на котором строится любая модель
окружающего мира, в том числе и научная. Но фундамент модели - это допущения,
аксиомы, которые должны быть очевидны и бесспорны.
Оставим пока в
покое время и пристальнее взглянем только на одного "кита" любой
модели мира - пространство. Заметим, что любой исследователь измененных
состояний сознания отмечает изменение переживаемого пространства, но не
находит слов для однозначного определения этого изменения. И, действительно,
как же можем описать изменчивость свойств пространства если в науке они
определены однозначно.
Так с первого
шага возникают сложности. Чтобы взглянуть на фундамент научной картины мира,
необходимо - для лучшего обозрения - с него слезть. Иначе говоря, критический
взгляд на аксиоматику научной модели мира потребует встать на позиции другой
аксиоматики. Подобные попытки известны из опыта ряда более древних культур,
но заметного влияния на научные взгляды Запада "импортные" модели
мироздания не произвели. И так случилось по той простой причине, что
аксиоматика этих моделей несовместима с "очевидными" допущениями
западной науки. Когда допущения сделаны и трудами ряда поколений модель
построена, то уже поздно сомневаться в правильности допущений, ведь смена
аксиоматики грозит разрушить саму модель. Тем более, что правдивость научной
модели мира доказана бесспорным торжеством технологий, радикально изменивших
и Землю и условия жизни ее обитателей. Именно правдивость и продуктивность
научного знания внушает нам уверенность в исключительной правоте аксиоматики
научной модели мира. Так на пути желающего пристальнее взглянуть на
"очевидные и бесспорные" допущения научной модели мира возникает
трудно преодолимый психологический и образовательный барьер.
Становится
очевидным, что простая смена аксиоматики модели мира неприемлема из-за
отсутствия чего-то общего у этих моделей, чего-то позволяющего вести
конструктивный диалог. Несовместимость допущений исключает адекватность
осмысления увиденного с позиций другой модели. Разная аксиоматика - это
разные логики построения модели. Перевод между логиками невозможен по
определению. Возможна только интерпретация на основе "своей"
логики. Несовместимая аксиоматика порождает, с первого взгляда,
антагонистические модели. Как этот антагонизм проявляется в жизни, мы можем
видеть на примере доброжелательных попыток диалога между
"неиспорченным" европейцем и "неиспорченным" тибетским
горцем, или между представителем Живой Этики и "нормальным"
профессором западного университета. Конечно, исключения известны, но они
требуют многолетней подготовки одного из участников диалога. При отсутствии
этой подготовки, как правило, получаются монологи представителей
несоприкасающихся миров с несопоставимыми взглядами и ценностями. Эта
тупиковая ситуация наблюдается не менее века.
Теоретически
возможен еще один вариант решения задачи исследования аксиоматики
используемой модели мира. Это нахождение аксиоматики, превращающей
используемые модели мира в частные случаи или в элементы более общей модели
способной интегрировать уже существующие. Пусть в этом месте меня простят и
представители мистических доктрин, и ученные, уверенные в абсолютности их
моделей мира, но без допущения практической применимости оппонирующей модели
процесс интеграции знания немыслим. Истинность моделей мира доказывается их
практической применимостью и в этом, при отсутствии предвзятости, убедиться
нетрудно каждому того желающему.
Очевидно, что
удачный подход интеграции известных моделей мира должен иметь следующие
следствия:
Все
закономерности научной модели мира должны непротиворечиво
"вписаться" в более общую модель мира, как частный случай.
Все
закономерности моделей мира оккультных и религиозных доктрин тоже должны
"вписаться" в интегральную модель мира, как частный случай.
Известные как
науке, так и другим традиционным системам знания, парадоксы должны найти свое
естественное решение.
С позиций
интегральной модели должна существовать возможность конструктивного диалога
между сторонниками научной картины мира и сторонниками мистических доктрин.
Эту немыслимую
задачу можем попытаться решить изменением только одной аксиомы - пространство
не обязательно трехмерно и оно не является элементом мира (Реальности).
Пространство можем считать только фундаментом модели мира, свободно или
принудительно (например, заданной воспитанием) операционной средой
моделирования, которая становится изменчивой. Заметим, что изменчивость
пространства - это изменчивость аксиоматики модели. Размерность пространства,
в таком случае, отдельный параметр пространства, задающий предельную
сложность модели. Ведь очевидно, что увеличение на единицу мерности
пространства моделирования повысит вместимость модели в бесконечность раз. А
бесконечность - это уже не количественный, а качественный или принципиальный
скачок уровня моделирования. Так что изменение мерности модели - это ее
качественное изменение или ступенька адекватности.
Возникает
справедливый вопрос об избыточной сложности подобной модели. Нет возможности
доказать необходимость предлагаемого изменения аксиоматики, опираясь на
"старую" аксиоматику. Полезнее потерять сколько-то времени и
попытаться нечто построить на "новом" интегральном фундаменте. Если
в этом строении проявятся вышеперечисленные ожидания, это докажет хотя бы
умозрительную полезность модели.
Обратим
внимание на то, что научная модель мира, являясь суммой научного знания, есть
только модель мира. Работоспособная, продуктивная, подтверждаемая личным
опытом каждого, но все-таки модель. Мы, представители западной культуры,
настолько отождествлены с научной моделью мира, что у нас не возникает
сомнения в том, что воспринимаемый мир - это лишь модель мира или Реальности.
Такой поворот мысли может вызвать эмоциональный протест, но, отложив эмоции в
сторону, сосредоточимся на формальной стороне подхода. С этой точки зрения
нет возможности спорить с тем, что и культура целиком, и индивид в
отдельности, моделируют окружающий мир и взаимодействуют с ним, опираясь на
знание постоянно развивающейся модели.
Следовательно,
пространство является фундаментом, основой модели мира, а не элементом или
свойством самого мира (Реальности). Этот тезис сам по себе ничего особо не
меняет. Смещается только точка зрения, дарящая дополнительную свободу
взглядов. Сумма нашего знания о мире превращается в модель Реальности без
всякого пересмотра содержания. Единственное, что добавляется, это знание, что
научная модель мира - не единственно возможная. Критерий правдивости модели
то же ясен - ее практическая применимость, конструктивность. Таким образом
исчезает антагонизм к другим моделям Реальности и мы уже в готовы их
рассматривать, понимая, что они необязательно может быть неверные. Разницу
видений мира обусловливает другая аксиоматика модели. Значит дело не в
абсолютной правильности, что недостижимо в любой модели, а в выборе начальных
аксиом, в том числе и свойств пространства. Если шаман достигает намеченного
и нам очевидного результата странными или невозможными в нашем понимании
методами, это значит, что его модель мира другая, но она вполне
работоспособна. Пропадает повод для восторга или испуга и появляется
конструктивные предпосылки для исследования.
Далее, чтобы
избежать путаницы в терминах, то, что мы пытаемся моделировать, назовем
Реальностью. Реальность окружает нас, и мы сами, со всеми своими попытками
осознать и моделировать Реальность, являемся тоже Реальностью или ее частью.
Нет ничего, что не является Реальностью и все, что есть, - является
Реальностью. Будем считать это допущением или аксиомой интегральной модели,
попутно замечая, что она тоже очевидна и не противоречит ни науке, ни
здравому уму.
Этот тезис
особых возражений вызывать не должен. Тем более что мы не пытаемся приписать
Реальности никаких особых или дополнительных свойств. Единственное, что следует
из общей логики и стоит отметить, так это то, что любая, даже самая сложная
модель Реальности будет представлять упрощение Реальности. А значит,
Реальность всегда сложнее любой модели. В противном случае, учитывая
системный подход, мы признаем себя равными по сложности или даже более
сложными, чем Реальность, частью которой мы являемся. Тем более, что общая
взаимосвязанность уже достаточно доказана в разных областях научного знания
от физики до антропологии. В этом случае процесс познания сводится к освоении
существующих моделей Реальности и построению новых, более сложных или
интегральных.
Отметим, что с
позиций интегрального подхода естественно отпадает вопрос о познаваемости -
непознаваемости Реальности. Теперь имеет смысл говорить только об уровне познания
Реальности. Грубейшая градация уровня познания Реальности - это мерность
среды моделирования или мерность пространства. Наше осознание развито до
уровня трехмерного моделирования. Хотя с начала это кажется неочевидным,
анализ деятельности как бодрствующего, так и находящегося в измененных
состояниях сознания разума всегда в остается в рамках трехмерного
моделирования. Более мелкая градация сложности моделирования связана с
причинностью, как следствием времени. Но это уже отдельная тема.
В предельном
масштабе получается, что попыткой моделирования себя самой через нас
занимается сама Реальность. Пока нам кажется, что мы отдельные и
самостоятельные субъекты, нам никто не запретит так думать. Однако сегодня
менее противоречиво считать себя частью познающей себя Реальности.
И так подводим
первые итоги поиска аксиоматики для интегральной модели Реальности:
Пространство
не трехмерно и оно не является элементом Реальности.
Пространство -
это только операционная среда моделирования более сложной, чем любая ее
модель, Реальности.
Мерность
пространства моделирования - это мера предельной сложности модели, а не
атрибут Реальности.
Уровень
познания Реальности зависит от сложности применяемой модели и ее аксиоматики.
Как видим, на
первый план выступает процесс моделирования как способ познания Реальности.
Но моделирование основано на механизме осознания. Поэтому придется обратить
внимание на сам процесс осознания, изучение которого в настоящее время
фрагментарно. Если структура и функционирование органов чувств, нервной
системы, мозга на уровне морфологии изучено достаточно детально, то попытки
построения интегральных моделей нашей с вами психической активности и
познавательности находятся на уровне поиска подходов. Взгляд на процесс
осознания, как механизм отражения физической реальности явно слишком узкий
для целей интегральной модели осознания, так как находится "внутри"
научной картины мира.
В нашей
технологической цивилизации нормальным осознанием считается осознание на
основе "врожденного", т.е. в ходе воспитания впитанного алгоритма
осознания, который кажется единственным, естественным и не подлежащим
обсуждению. Касательно западной цивилизации таким образом осуществляется
преемственность научной модели мира и последовательное развитие этой модели.
Алгоритм осознания - это определенная логика или способ развертки, сортировки
и обозначения поступающих переживаний. Операционная среда, в которой
происходит упомянутая обработка переживаний, названа пространством. Это среда
построения модели Реальности, как индивидуальной, так и общекультурной. Само
пространство, сколь пустым мы его не воспринимали бы, в себе всегда прячет
определенный алгоритм осознания. Индивидуальными отклонениями алгоритма
осознания, в зависимости от их величины, занимаются или психотерапевты или
психиатры, выполняющие роль стражей культурного стандарта алгоритма осознания
Реальности.
Как отмечалось
высшее, любое доступное нам пространство моделирования - трехмерно (явь, сон,
трансы….). "Нормальные" представители человечества способны строить
не более и не менее чем трехмерные модели Реальности. Математические попытки
оперирования многомерными пространствами - всего лишь механистический перенос
логики трехмерного моделирования в область многофакторных задач. Определение
границ универсальности математического аппарата при переходе к четырехмерному
моделированию сегодня, вероятнее всего, невыполнимая задача, так как органы
чувств нашего физического тела непригодны для восприятия четырехмерной
Реальности.
Трехмерное
осознание задает определенный потолок сложности моделирования, с которым уже
столкнулись первопроходцы в разных областях науки. Число парадоксов и тупиков
в научном поиске растет с каждым годом, вызывая все крепнущее подозрение, что
в научном подходе на самом фундаментальном уровне допущена ошибка. Но если
следовать логике изложенного, это не ошибка, а предел возможностей
трехмерного моделирования на основе выбранной аксиоматики. Таким образом
можем делать вывод, что западная цивилизация приближается к пределу
возможностей трехмерного моделирования Реальности на основе общепринятой
аксиоматики. Накопленные парадоксы свидетельствует об этом. Встает вопрос
смены аксиоматики моделирования. В таком случае, потенциально, для человека
открываются немыслимые просторы познания, а доктрины традиционных или
мистических систем знания оказываются путеводными знамениями для следующего,
более глубокого, этапа познания Реальности и себя, как ее части. Не будет
странным, что приходящая смена аксиоматики моделирования Реальности приведет
к естественному появлению этической компоненты познания. Сегодняшняя научная
модель мира при всем желании не может вместить в себя ни этики, ни живого.
Эту зияющую дыру в модели мира пытаются заполнять религии и другие системы
знания пришедшие из глубины веков, но сегодня эти попытки развитому
интеллекту западного человека небезосновательно кажутся странными или просто
глупыми.
С другой
стороны, говоря о более многомерной модели Реальности, мы сами попадаем в
затруднительное положение. Зная, что способны пользоваться только трехмерным
пространством моделирования и говоря о, как минимум, четырехмерной модели
Реальности, мы оказываемся неспособными на деле ее применить. Вообразить
четвертую ортогональную плоскость в добавление к известным трем нам не
позволяет доминирующий трехмерный алгоритм осознания. Но обратим внимание на
то, что доминирование определенного алгоритма осознания - это только
доминирование, а не запрет иных и даже более сложных алгоритмов осознания. В
случае увеличения мерности алгоритма осознания получим предпосылки для
создания соответствующей мерности модели Реальности. Но, как уже отмечали,
модель увеличенной мерности будет отличаться от предыдущей качественно, а не
количественно. Бесконечность трехмерного пространства, умноженная на
бесконечность, - это уже не математическое исчисление, а переход на
качественно иной уровень моделирования. С непривычки подобное заявление
должно звучать достаточно фантастически, если не обратить внимание на
содержание традиционных систем знания. А если присмотреться повнимательнее,
да еще и позволить на себе "опробовать" их методы воздействия на
осознание, то выяснится, что суть их - учение о сознательном управлении
осознанием вплоть до освоения четырехмерного моделирования (просветление).
В западной
культуре идей о волевом управлении осознанием почти еще нет. Тем не менее,
переключение осознания возможно и происходит под воздействием психотропных
веществ, из-за отклонений физиологических функций, в случае крайней
психологической напряженности и т.п. Такие отдельные отклонения осознания
оставляют глубочайшие впечатления, которые в нашей культуре нет способа
объяснить и поэтому их "списывают", как иллюзорные или болезненные.
Однако повторяющиеся переключения могут подарить их обладателю возможность
освоить какое-то "волшебное" ремесло, а может, дать работу
психиатрам. Однако "одаренный" таким подарком индивид обычно не
понимает сути происходящего и считает случившееся даром потусторонних сил или
болезнью. Психиатры, понимая столько же, уверены в болезненности подобных отклонений
осознания и изобретают способы лечения. С точки зрения результата успешность
психиатрического лечения сомнительна, и еще более сомнительна с точки зрения
концепции психического здоровья, являющейся частью научной модели мира в
которой для психики, кроме эмпирического определения нормы, место вообще
непредусмотрено.
Так как
представители западной культуры в большей степени являются материалистами,
попутно заметим, что физиология мозга влияет на процессы осознания, но это
влияние однобоко: это изменение уровня отождествленности
с "врожденным" алгоритмом осознания. Все известные средства
приводящие к измененным состояниям сознания работают по принципу подавления
"нормального" алгоритма осознания с весьма слабым
"направляющим" действием. Ничего более радикального добиться не
получается. А это значит, что мозг, как часть модели физического мира, связан
только с одним алгоритмом осознания, представляющим нам Реальность в виде
научной модели мира (видимость физической реальности).
Попробуем
графически представить механизм осознания.
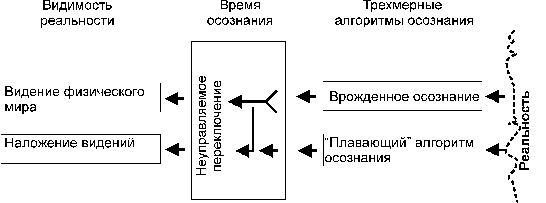
Рисунок 1.
Нормальное осознание мира и неуправляемое переключение осознания.
Средний блок
рисунков, обозначенный как "время осознания" намекает на механизм
управления осознанием, но его пока не будем разбирать и остановимся на общих
чертах механизма осознания. В случае управляемого осознания (традиционные
доктрины знания допускают таковое) возникающие иные видения реальности сильно
удивляют своими фундаментальными отличиями, но оставляют возможность при
помощи параллельного осознания опознать элементы Реальности и достичь
осознанного пребывания в конкретном осознании.
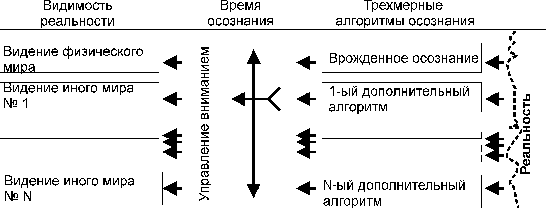
Рисунок 2.
Управляемое осознание реальности, через управление вниманием.
Управление
осознанием позволяет отличать и осознанно выбирать алгоритмы осознания
реальности, добиваться плавного наложения иного видения на обычное видение
физического мира, осознавая дополнительные связи и свойства осознаваемого.
Подобное "размытие" врожденного алгоритма осознания знакомо тем,
кто научился пользоваться интуицией или владеет каким либо
"пограничным" ремеслом. В случае овладения данным механизмом, а это
только механизм осознания, экспериментатору собственным осознанием будет
понятно происходящее с ним и он даже не попытается искать потусторонние силы,
другие миры или божий дар. Но зато у него появится чувство ответственности за
им творимое.
Из
нарисованных схем видим, что одна и та же Реальность, будучи по-разному
отображена разными алгоритмами осознания, в соответствующих операционных
средах (пространствах) осознания породит целый ряд видимых миров. Реальность
нам может предстать видимостью целого ряда совершенно непохожих между собой
миров. Тот, кто занимался измененными состояниями сознания, подтвердит их
разительные отличия. А тот, кто применяет измененные состояния сознания
(измененные алгоритмы осознания) с целью лечения, подтвердит эффективность
таких методов. Тут и напрашивается вывод, что изменение алгоритма осознания -
это вовсе не иллюзия, а просто другое видение той же самой Реальности. И в
этом месте мы приходим к поразительному выводу, что Реальность невообразимо
сложна, раз есть возможность ее осознавать столь многими способами и каждый
из этих способов осознания, овладев им, позволяет результативно
взаимодействовать с ней.
В результате
получаем контуры интегральной модели Реальности:
Мы не способны
напрямую осознать Реальность.
Для
взаимодействия с Реальностью мы строим модели Реальности и, отождествившись с
ними, через них осознаем отдельные аспекты Реальности, обусловленные
аксиоматикой конкретной модели Реальности.
Пространство -
это не более чем операционная среда конкретного осознания Реальности и
является только основой моделирования.
Возможны
разные конструктивные алгоритмы осознания, а научная модель мира является
только одним из них. Разные алгоритмы осознания - это переживание разных
пространств.
В
четырехмерной модели Реальности решаются все возможные парадоксы трехмерных
моделей Реальности, как научной, так и других.
Уровень
познания Реальности напрямую связан с максимально доступной сложностью
моделирования. Грубейшие ступени сложности модели Реальности - это мерность
среды моделирования или мерность пространства осознания.
Несколько
дополнительных следствий допущения интегральной модели осознания:
живое
проявляется во всех моделях реальности, а проявление в жизни в научной модели
мира отражает только один из ее аспектов;
возможны формы
жизни не отражающиеся в научной модели мира;
существуют не
параллельные миры, а параллельное осознание той же Реальности на основе
разных алгоритмов осознания;
истинность
алгоритма осознания доказывает практическая результативность его применения;
понятие
объективности корректно применимо только в рамках одного алгоритма осознания;
разделение
разделения
объект - субъект, материальное - идеальное является следствием научной модели
мира;
дуалистическое
мышление возможно только в трехмерных средах моделирования и не является
универсальным;
в
четырехмерных средах моделирования может появиться и трехполюсная
полярность в добавления к известным в нашей модели одно- и двухполюсным
полярностям;
время
становится параметром меняющим осознание в рамках моделей одинаковой
мерности;
……..
Еще мы можем
заметить, что при следовании предложенной логике решается большинство
"вечных" философских проблем, но появляются новые. Открывается
новый смысл беспредельности не в пространственном смысле, а в контексте
развития осознания. Становится очевидным ограничивающий характер научной
картины мира в познании Реальности и актуализируется вопрос исследования
сознания в новом контексте. Становится очевидным почему научная модель не
позволяет полномасштабного исследования феномена Жизни.
Изложенное -
это только легкое прикосновение к одному элементу аксиоматики научной картины
мира - пространству. Время тоже готово показать свои невидимые с позиций
научной картины мира свои стороны.
|
|
|
|
|
1. ОБЩИЕ
СВЕДЕНИЯ ОБ ЭКОСИСТЕМАХ И ЛАНДШАФТАХ, ИХ МОНИТОРИНГ
1.1. Экосистемы,
законы их функционирования и продуктивности
В переводе с
греческого systema - составленное из частей, т. е.
множество элементов, находящихся в отношениях и связях друг с другом,
образующих объективное единство связанных друг с другом предметов, явлений, а
также знаний. Экосистема - совокупность живых организмов в неорганической среде
их обитания, занимающих определённое пространство и способных к саморегуляции. Абиотические компоненты
экосистемы представлены минералами и горными породами, растительным опадом, остатками микроорганизмов и животных. Биотическая
часть экосистемы состоит из первичных продуцентов - автотрофов и консументов
- гетеротрофов и сапротрофов,
которые разлагают растительные и животные остатки до уровня исходных
неорганических веществ, выполняя одновременно роль вторичных продуцентов (43). Связываете
неживую и живую природу почва, которая осуществляет важнейшие многочисленные
экологические функции.
Выделяют микроэкосистемы, например ствол гниющего
дерева, мезоэкосистемы (лес, пруд и т. п.), макроэкосистемы (океан, континент и т. п.). Глобальная
экосистема одна -
биосфера (364).
Все экосистемы различаются структурами пространственно-временного разнообразия,
потоками энергии, трофическими
цепями, биогеохимическими круговоротами элементов и веществ, развитием, эволюцией и
управлением (298).
В отечественной научной литературе широко применяются термины
"биоценоз", "биогеоценоз", "биопедоценоз"
и другие, а в зарубежной преобладают "экологическая система" или
"экосистема". В 60 - 70-х годах на научно-методических и программных
совещаниях ЮНЕСКО, было рекомендовано применять единый термин - экологическая
система или экосистема (209). Однако в
России параллельно с концепцией экосистем с 1942 г. продолжает развиваться
учение о биогеоценозах (403) и с 1979 г. -
о биопедоценозах (288).
В. Н. Сукачев определил биогеоценоз как совокупность однородных природных
элементов на определённом участке поверхности Земли. По Н.Ф. Реймерсу (364) биогеоценоз -
эволюционно сложившаяся, относительно пространственно ограниченная, внутренне
однородная природная система функционально взаимосвязанных живых организмов и
окружающей их абиотической среды, характеризующаяся определённым энергетическим
состоянием, типом и скоростью обмена веществом и информацией. Он отмечал, что
понятия "экосистема" и "биогеоценоз" - синонимы.
Существует мнение (43),
что под определение биогеоценоза экосистемы не подпадают, так как им не
свойственны некоторые признаки этого определения, например, территориальная
ограниченность. Экосистема, напротив, может включать несколько биогеоценозов,
т. е. понятие "экосистема" шире, чем понятие "биогеоценоз".
Любой биогеоценоз является экологической системой, но не всякую экосистему
можно считать биогеоценозом, сугубо наземным образованием, имеющим чёткие
границы.
Биопедоценоз - это элемент территории, однородный в
геоморфологическом и гидротермическом отношениях, единый по характеру
почвенного покрова и естественной растительности (фитоценоз) с присущими им
организмами и режимами в пределах почвенного и растительного покровов (322).
К антропогенным факторам, оказывающим воздействие на экосистемы, биогеоценозы и
биопедоценозы относят сельское хозяйство, освоение
земель, вырубку лесов, горнодобывающую и перерабатывающую промышленность,
производство энергии, транспорт, строительство и др. В результате различных
антропогенных воздействий в этих природных системах происходят изменения
элементов, пищевых цепей, круговорота веществ и потоков энергии. Они
приобретает определённые особенности, свойственные конкретным типам
хозяйственной деятельности. При аграрных антропогенных воздействиях экосистема
трансформируется в агроэкосистему, биогеоценоз - в агробиогеоценоз, биопедоценоз - в
агропедоценоз, т. е. в эколого-экономические системы
соответствующего уровня.
В процессе длительной эволюции естественные системы приобрели способность к саморегуляции и самовосстановлению. При их трансформации в агроэкосистемы вещественно-энергетические и информационные
связи, в том числе и в агроландшафте, значительно
изменяются. Если такие изменения выходят за рамки допустимых пределов, то
экосистемы и ландшафты теряют способность к самовоспроизводству
основных компонентов и в итоге быстро деградируют. Природные системы обладают
рядом свойств, определяющих их отношение к внешним воздействиям. Одними из них
и очень важными для эффективного функционирования агрогенных
аналогов являются изменчивость и устойчивость.
Изменчивость - способность экосистемы переходить из одного состояния в другое
под влиянием внешних сил или факторов саморазвития.
Устойчивость - способность к самосохранению и саморегулированию в пределах, не
превышающих определённых критических величин (допустимых пределов изменений).
Эта научная категория имеет основополагающее значение для оценки систем
земледелия и практических мер по управлению агроэкосистемой.
Функционирование и продуктивность экосистем подчиняются объективным законам,
правилам и принципам, открытым учёными. Основные из них приведены ниже с
преимущественным использованием трактовок, принятых в книгах выдающегося
отечественного эколога Николая Федоровича Реймерса
"Природопользование" (1990), "Надежды на выживание человечества:
концептуальная экология" (1992) и "Экология (теории, законы, правила,
принципы и гипотезы)" (1994). В изложении сформулированных закономерностей
почти нет ссылок на литературные источники, так как многие из них были выявлены
повторно, нередко многократно. Авторство указывается лишь тогда, когда оно
"канонизировано" в учебниках и специальных словарях.
Закон хиральной чистоты Л. Пастера: живое вещество состоит
из хирально чистых структур. Хиральная
чистота, - наличие объектов, несовместимых со своим зеркальным отражением (типа
правой и левой руки, откуда и происхождение термина: с греческого "хира" - рука). Белки живого построены только из
"левых" (поляризующих свет влево) аминокислот, нуклеиновые кислоты
сложены исключительно из поляризующих свет вправо сахаров и т. п. Вещества небиогенного происхождения хирально
симметричны - "левых" и "правых" молекул в них поровну. Хиральная чистота обусловливает специфику живого, несводимость его к неживому и практическую невозможность
получения живого из неживого.
Хиральная асимметричность, свойственная живому,
объективно указывает на его физико-химическое единство, что и отражает закон
физико-химического единства живого вещества В. И. Вернадского: всё живое
вещество Земли физико-химически едино. Этот закон не
исключает количественной биогеохимической специфики видов живого. Однако
качественно жизнь едина, и потому подчиняется единым правилам.
Термодинамическое правило Вант-Гоффа - Аррениуса в
биолого-экологической модификации гласит, что подъем температуры на 10° С
приводит к 2-3-кратному ускорению химических процессов. Это правило существенно
углубляется закономерностью, формулируемой для теплокровных животных в виде
правила поверхностей: отношение продуцируемого особью животного тепла к единице
площади её тела приблизительно одинаково и варьирует около величины 1000
ккал/м2/сутки.
Первый биогеохимический принцип В.И. Вернадского: биогенная миграция атомов
химических элементов в биосфере всегда стремится к максимальному своему
проявлению.
Второй биогеохимический принцип В.И. Вернадского: эволюция видов в ходе
геологического времени, приводящая к созданию устойчивых в биосфере форм жизни,
идет в направлении, усиливающем биогенную миграцию атомов.
Третий биогеохимический принцип В.И. Вернадского: живое вещество находится в
непрерывном химическом обмене с космической средой, его окружающей, и создается
и поддерживается на нашей планете космической энергией Солнца.
Закон сохранения термодинамического состояния: энтропия и информация обратно
пропорциональны, что ведет к дифференциации отдельных функций организма и,
следовательно, специализации его частей. Это вызывает эволюционные адаптации, в
том числе ведет к развитию систем управления, соотносящих работу отдельных
реагентов и стабилизирующих их параметры. Этот закон объясняет внутренние
побудительные механизмы возникновения адаптаций, тенденцию к эволюционным
переменам. Внутренние процессы сложно взаимодействуют с процессами, вызываемыми
внешними причинами, в рамках четырёхединства
биологических систем: целостности, преобразуемости, саморегуляции и воспроизводимости.
Правило ускорения эволюции: с ростом сложности организации биосистем
продолжительность существования вида в среднем сокращается, а темпы эволюции
возрастают. Например: средняя продолжительность существования вида птиц - 2
млн. лет, млекопитающих - 800 тыс. лет, предковых форм человека - 200-500 тыс.
лет, современного человека - пока не более 50 тыс. лет.
Правило эквивалентности в развитии биосистем (Л. фон Берталамфи):
биосистемы способны достигнуть конечного (финального) состояния (фазы) развития
вне зависимости от степени нарушения начальных условий своего развития. Это
происходит лишь при сохранении минимума внешних и внутренних условий
существования биосистемы.
Правило максимального "давления жизни": организмы размножаются с
интенсивностью, обеспечивающей максимально возможное их число. "Давление
жизни" лимитировано емкостью среды жизни и многими другими
закономерностями системного мира: законом ограниченного роста, правилами взаимоприспособленности, внутренней непротиворечивости,
соответствия среды генетической предопределенности организма и пр.
Всем этим лимитирующим механизмам организмы противопоставляют возможности преадаптации. Принцип преадаптации:
способность к приспособлению у организмов заложена "изначально" и не
связана непосредственно с их взаимодействием со средой обитания.
Закон генетического разнообразия: все живое генетически различно и имеет
тенденцию к увеличению биологической разнородности. Естественно, что при
известном условии отсутствия в природе двух генетически идентичных особей
(кроме однояйцевых близнецов), варианты преадаптации неисчислимы.
Закон эволюции А.Н. Северцова гласит о чередовании
ароморфозов (арогенезов), принципиально меняющих возможности вида к адаптации и
расширению сферы обитания, и периодов аллогенезов
(идиоадаптаций) - изменений частного порядка.
Принцип внезапного усиления патогенности: живое болезнетворное начало действует
тем сильнее и разрушительнее, чем выше разница между его патогенностью и
сопротивляемостью к ней, особенно, если этот дисбаланс возникает внезапно.
Усиление патогенности возникает в следующих случаях:
- при мутации болезнетворного организма, приведшей к резкому возрастанию его
способности к поражению;
- при выработке болезнетворным организмом новой экологической ниши (обычно при
освобождении этой ниши другими организмами) в ранее не пораженном виде или иной
систематической категории организмов;
- при внедрении нового болезнетворного организма в экосистему, где нет
механизмов регуляции его численности;
- при очень резком изменении среды жизни для поражаемого организма или для
экосистемы, куда он входит, что снижает его способность к сопротивлению
болезнетворному началу;
- при неблагоприятном изменении половозрастной структуры поражаемого вида
(сильного постарения его особей или, наоборот, резкого их омоложения), в обоих
случаях приводящем к снижению иммунитета.
Мутация может привести как к ароморфозу (арогенезу) - резкому расширению сферы
обитания вида, возникновению многих близких форм, так и к аллогенезу
(идиоадаптации) - изменениям частного порядка. В первом случае возможно
катастрофическое поражение страдающего организма. Если при этом заметно
изменились условия его обитания, может произойти полное вымирание поражаемого
вида и более крупной систематической категории живого при условии, что он (она)
не имеет очень большого запаса генетических преадаптаций
для сопротивления болезнетворному началу.
Закон эколого-системной направленности эволюции: любые эволюционные изменения в
конечном итоге направлены экологическими факторами и системными особенностями
развития эволюционизирующей совокупности, т. е.
прогресс направляется как внешними, так и внутренними факторами. Внешними для
развивающейся системы живого служат все воздействия надсистем - от Вселенной до
ближайшей по иерархии, которые выступают как ограничения развития и как
механизмы, к которым развивающаяся совокупность живого или с его участием
вынуждена адаптироваться. Развитие каждой из надсистем ведет к изменению
условий существования, ужесточающих естественный отбор и одновременно
вызывающих усиление мутационного процесса. Может воздействовать также
параллельная система. Например, воздействие на организмы пестицидов приводит у
части из них к генетически закрепленной адаптации к этим ядохимикатам.
Правило происхождения новых видов от неспециализированных предков Э. Копа:
новые крупные группы организмов берут начало не от специализированных
представителей предков, а от их сравнительно неспециализированных групп. Это
правило дополняет принцип дивергенции Ч. Дарвина, или правило адаптивной
радиации Г. Ф. Осборна: филогенез любой группы сопровождается разделением её на
ряд отдельных филогенетических стволов, которые расходятся в разных адаптивных
направлениях от среднего исходного состояния.
Эти новые группы либо остаются относительно неспециализированными, либо
подчиняются правилу прогрессирующей специализации Ш. Депере,
гласящему, что группа, вступающая на путь специализации, как правило, в
дальнейшем развитии будет идти по пути все более глубокой специализации. И чем
глубже такая специализация, тем возможнее расхождение темпов адаптации
организма и скорости изменения среды его обитания. Этот факт обобщает правило
более высоких шансов вымирания глубоко специализированных форм О. Марша:
быстрее вымирают более специализированные формы, генетические резервы которых
для дальнейшей адаптации снижены.
Взаимосвязь вещественно-энергетических и информационных закономерностей
приводит в действие закон увеличения размеров (роста) и веса (массы) организмов
в филогенетической ветви Э. Копа и Ш. Депере. В. И.
Вернадский так сформулировал этот закон: "По мере хода геологического
времени выживающие формы увеличивают свои размеры (а следовательно, и вес) и
затем вымирают".
Все закономерности, характерные для живого, имеют адаптивное значение.
Биосистемы вынуждены приспосабливаться к непрерывно изменяющимся условиям
жизни. Эти изменения имеют разную шкалу времени - от эволюционной до
сиюминутной. В вечно меняющейся среде жизни каждый вид организмов по-своему
адаптирован. Это выражается правилом экологической индивидуальности (Л.Г.
Раменский): каждый вид специфичен по экологическим возможностям адаптации, двух
идентичных видов не существует.
Экологическая специфичность видов подчеркивается так называемой аксиомой адаптированности, носящей также название экологической
аксиомы или аксиомы Ч. Дарвина: каждый вид адаптирован к строго определенной,
специфичной для него совокупности условий существования - экологической нише.
Из правила экологической индивидуальности и аксиомы адаптированности
вытекает экологическое правило (С.С. Шварц): каждое изменение условий
существования прямо или косвенно вызывает соответствующие перемены в способах
реализации энергетического баланса организма; чем выше уровень систематической
категории или больше их классификационное различие, тем значительнее отличие в
энергетических процессах.
Закон относительной независимости адаптации: высокая адаптированность
к одному из экологических факторов не дает такой же степени приспособления к
другим условиям жизни.
Закон подобия части и целого: часть является миниатюрной копией целого, а
потому все части одного уровня иерархии систем похожи друг на друга. Пример:
модель атома и солнечной системы.
Закон подобия части и целого отнюдь не означает их абсолютной идентичности. Об
этом гласит аксиома системной целостности: целое больше суммы его частей, или
аксиома эмерджентности: целое всегда имеет особые
свойства, отсутствующие у его частей - подсистем и не равно сумме элементов, не
объединенных системообразующими связями. При сложении
системного целого образующаяся интеграция подчиняется иным (хотя возможно, и
подобным) законам формирования, функционирования и эволюции. Образно говоря,
одно дерево еще не сад, как и группа деревьев. Механическое сосредоточение
химических элементов, молекул органических веществ, даже тканей и органов, не
дает организма. Для сада необходимо сочетание всех его экологических
компонентов, составляющих именно его агроэкосистему,
образование круговоротов веществ, регуляция потока энергии и т. п.
Система не может формироваться из абсолютно идентичных элементов. Отсюда
вытекает закон необходимого разнообразия. Сельскохозяйственная монокультура
вообще лишь метафорическое понятие, если это не стерильная гидро-
или аэропоника одного клона растений. В любой монокультуре участвуют сотни
видов микроорганизмов, растений (сорняки) и животных (почвенные, вредные и
др.). Для каждого типа агроэкосистем необходимое
разнообразие количественно различно и часто строго фиксировано. Нижний предел -
не менее двух элементов, верхний - бесконечность.
Системные образования состоят из подсистем. Их необходимое число и разнокачественность более или менее постоянны и
определяются законом (правилом) полноты составляющих: число функциональных
составляющих системы и связей между ними должно быть оптимальным - без
недостатка или избытка в зависимости от условий среды или типа системы. Четкие
лимиты характерны для всех систем, но экологические их модификации часто не
теряют функциональных черт и при довольно большом разбросе числа составляющих.
Для них характерны естественные колебания даже количества входящих видов.
Размах колебаний ограничен законом необходимого разнообразия. При всех
колебаниях числа составляющих оно подчиняется действию закона избыточности
системных элементов при минимуме числа вариантов организации: многие
динамические системы стремятся к относительной избыточности основных своих
составляющих при минимуме вариантов организации. Избыточность числа элементов
нередко служит непременным условием существования системы, её
качественно-количественной саморегуляции и
стабилизации надежности, обеспечивает её квазиравновесное состояние. В то же
время число вариантов организации жестко лимитировано. Природа часто
"повторяется", ее "фантазия", если говорить не о числе и
разнообразии однотипных элементов, а о количестве самих типов организации,
очень ограничена. Отсюда многочисленные структурные аналогии и гомологии, однопорядковые формы организации.
Имеется и стратегия самоограничения, направленная на замену количественного
роста качественным усовершенствованием: забота о потомстве, большая его
жизненность, интенсификация производства. Принцип перехода избыточности в
самоограничение заключается в следующем: избыточность системных элементов может
быть заменена повышением качества этих составляющих (индивидуальной надежности)
или их агрегации, в том числе в функциональные надсистемы.
Фиксированное число разнокачественных элементов возникает, очевидно, под
давлением объективных причин. Всякое объединение не случайно, если оно не
вызвано антропогенными внешними обстоятельствами. Движущим механизмом служит
"выгода" большей надежности при объединении - действие правила
конструктивной эмерджентности: надежная система может
быть сложена из ненадежных элементов или из подсистем, не способных к
индивидуальному существованию. Иерархическое строение природных систем
результат действия обсуждаемого правила. И для этого случая можно
сформулировать закон (правило) перехода в подсистему, или принцип кооперативности: саморазвитие любой взаимосвязанной
совокупности, её формирование в систему приводят к включению её как подсистемы
в образующуюся или существующую надсистему: относительно однородные системные
единицы образуют общее целое. "Кооперативный эффект" проявляется на
всех уровнях организации материи, и его часто называют системным, или системообразующим, эффектом. Фундамент возникновения
кооперативного эффекта - значительный вещественно-энергетический и
информационный выигрыш.
Это преимущество постепенно растет согласно закону (принципу) увеличения
степени идеальности (Г. В. Лейбница): гармоничность отношений между частями
системы историко-эволюционно возрастает. Этот принцип практически не имеет
исключений.
Эмпирические наблюдения подводят к формулировке аксиомы, или закона системного
сепаратизма: разнокачественные составляющие системы всегда структурно
независимы. Между ними существует функциональная связь, может быть взимопроникновение элементов, но это не лишает
целостностей, входящих в систему, структурной самостоятельности при общности
"цели" - сложения и саморегуляции общей
системы. Например, организм состоит из органов. Каждый из них "не
заинтересован" в ухудшении работы другого органа или в уменьшении его
размеров. Наоборот, в составе системы организма каждый орган тесно связан с
другими.
Итоговым обобщением и развитием перечисленных закономерностей сложения систем
служит закон оптимальности: с наибольшей эффективностью любая система
функционирует в некоторых характерных для неё пространственно-временных
пределах (или: никакая система не может сужаться или расширяться до
бесконечности). Размер системы должен соответствовать выполняемым ею функциям.
Обычно такой размер называют характерным размером системы.
Для перехода к обсуждению законов внутреннего развития систем важна
формулировка правила системно-динамической комплементарности,
или закона баланса консервативности и изменчивости: любая саморазвивающаяся
система состоит из двух рядов структур (подсистем), один из которых сохраняет и
закрепляет её строение и функциональные особенности, а другой способствует
видоизменению и даже саморазрушению системы с образованием новой
функционально-морфологической специфики, как правило, соответствующей
обновляющейся среде существования системы. Чем жестче организована система, тем
сильнее в ней механизмы консервации, прямолинейней и непосредственней их
действие. При этом большее значение имеют внешние для системы, а не внутренние
для неё факторы развития.
Мир систем функционально аналогичен, поэтому помимо общих закономерностей
сложения систем существуют общие правила их развития - как эволюционного, так и
индивидуального. Прежде всего это закон вектора развития: развитие
однонаправлено. Нельзя прожить жизнь наоборот - от смерти к рождению,
невозможно в том же направлении развернуть эволюцию планеты, жизни на ней. Эта
невозвратность сформулирована в виде закона необратимости эволюции живого Л. Долло: организм (популяция, вид) не может вернуться к
прежнему состоянию, уже осуществленному в ряду его предков. Очевидно, что эта
закономерность универсальна.
Столь же всеобща тенденция всего сущего к усложнению организации и
дифференциации функций и подсистем. Изначально эта закономерность (в другой
интерпретации и в несколько других словах) была сформулирована (или стала
широко известной) в виде закона усложнения системной организации (организмов)
К.Ф. Рулье: историческое развитие природных систем
приводит к усложнению их организации путем нарастающей дифференциации функций и
подсистем, выполняющих эти функции.
Вместе с тем в рамках эволюции крупных космических систем (например,
солнечной), очевидно, действует закон неограниченности прогресса: развитие от
простого к сложному эволюционно неограничено.
Абсолютизировать эту закономерность не стоит. Прогресс неограничен лишь при
очень значительных усилиях и саморегуляции как
ведущем факторе развития. Он требует постоянных жертв, число которых также
ограничено пределами разумной достаточности, а длительность
"неограниченности" все же лимитирована эволюционными рамками. Для
Земли это время существования самой планеты.
В сфере закономерностей развития есть как бы два ряда -
эволюционно-исторический и собственного развития индивидуальных систем, т. е.
филогенетический и онтогенетический в биологической трактовке. Э. Геккель
сформулировал биогенетический закон: онтогенез всякого организма есть краткое и
сжатое повторение филогенеза данного вида, т. е. индивид в своем развитии
повторяет (в сокращенном и закономерно измененном виде) историческое
(эволюционное) развитие своего вида. Д.В. Рундквист
открыл геогенетический закон: минералогические
процессы в короткие интервалы времени как бы повторяют (в измененном виде, со
своими "акцентами") общую историю геологического развития, или,
другими словами, геологические процессы развития однонаправлены во всех
масштабах геоэволюции (общей эволюции Земли, в рамках
геологического мегацикла, тектономагматического цикла и т. п.).
Функциональные изменения следуют проверенным длинной серией проб и ошибок
путем. Они определяются законом последовательности прохождения фаз развития:
фазы развития природной системы могут следовать лишь в эволюционно и
функционально закрепленном (исторически, эволюционно, геохимически
и физиолого-биохимически обусловленном) порядке,
обычно от относительно простого к сложному, как правило, без выпадения
промежуточных этапов, но, возможно, с очень быстрым их прохождением или
эволюционно закрепленным отсутствием. Насильно убрать какую-то из фаз развития
практически невозможно. Иногда доступно ее несколько сократить во времени.
Нельзя существенно отклонить и направление развития. Можно лишь его задержать,
даже отсечь какие-то последующие фазы, но не качественно изменить. Доступно и
некоторое ускорение процесса путем регуляции внутренних взаимосвязей.
Существует общий системогенетический закон: природные
системы в индивидуальном развитии повторяют в сокращенной и нередко в
закономерно измененной и обобщенной форме эволюционный путь развития своей
системной структуры. Каждая подсистема следует за своей системой, вернее,
развитие надсистемы определяет многие ограничения в развитии входящих в нее
подсистем. Такой процесс "подталкивания", направления развития характерен
для всего системного мира как в сверхдлинных отрезках эволюционного времени,
так и в сравнительно коротких сроках индивидуального развития. Всюду есть
взаимоотношения в иерархии систем - эволюция эволюций и развитие развитий.
Процессы-организаторы и их механизмы как бы подталкивают развитие по
определенным руслам, и как в реке ниже лежащая масса воды пройдет раньше выше
лежащей, так и в процессах развития предыдущие стадии не могут идти позже за
ними следующих. Такова логическая модель действия системогенетического
закона. Принципы управления остаются все теми же - целое ограничивает число
степеней свободы своих частей, то же делают части по отношению друг к другу и к
целому.
Системогенетический закон в отличие от
биогенетического своего предшественника позволяет прогнозировать будущее
развитие. Вернее, не сам закон, а одно из его следствий. Если развитие
относительно детерминировано воздействием иерархии надсистем, а отчасти и
подсистем в прошлом (подсистемы, изменяясь, не могут не влиять на целое, пример
тому мутации), то характер процессов не изменится и в будущем, во всяком случае
ближайшем (в масштабе характерного времени систем). И хотя принцип
"развитие есть движение движений во всей иерархии значимых систем" не
позволяет создать одной безальтернативной модели, все же можно прогнозировать
вероятный ход событий. Если же рассматривается процесс, описываемый системогенетическим законом, в начальных (не в конечных)
фазах развития, точность прогноза резко возрастает: никогда бабочка не вылетит
из яйца или из гусеницы, только из куколки.
Отсечение надсистемой и межсистемными отношениями многих вероятных путей
развития подсистем, детерминированность их развития
предполагает наличие единого русла изменений, их одновременности и
сопряженности. Ж. Кювье сформулировал закон анатомической (или структурной)
корреляции. Его современное звучание: в организме, как в целостной системе, все
его части соответствуют друг другу как по строению (закон соподчинения
органов), так и по функциям (закон соподчинения функций). Изменение одной части
организма или отдельной функции неизбежно влечет за собой изменение других
частей и функций.
Но в любой системе имеется согласование частей, иначе она бы шла вразнос, не
было бы никакого единства. Очевидно, действует закон согласования строения и
ритмики (функций) частей (подсистем), или закон синхронизации и гармонизации
системных составляющих: в системе как самоорганизующемся единстве
индивидуальные характеристики подсистем согласованы между собой. Одно из
важнейших следствий этого закона в том, что выпадение одного из звеньев системы
меняет структуру и функции других, сопряженных с этим законом, или полностью
изменяет целое. Выпадение атома из молекулы делает молекулу совсем иной;
эволюционное изменение одного из функциональных органов меняет весь строй
организма и т. п.
Поскольку имеется корреляция между органами и функциями, а развитие есть
движение многих движений, в системе одновременно должны присутствовать
подсистемы бурно прогрессирующие, стабильные и регрессирующие. Система как правило,
развивается неравномерно. Дж. Хаксли сформулировал закон аллометрии
- неравномерности роста частей тела в процессе развития организма. Автор закона
аллометрии считал такую неравномерность развития
аномалией, даже патологией. Однако неравномерное развитие подсистем внутри
одной системы весьма обычно. Так развиваются - сезонно и в многолетнем ряду -
организмы в экосистеме. Н.Ф. Реймерс (1994) отмечает,
что закон неравномерности развития систем, или, лучше, закон разновременности
развития (изменения) подсистем в больших системах может быть сформулирован в
таком виде: системы одного уровня иерархии (как правило, подсистемы системы
более высокого уровня организации) развиваются не строго синхронно - в то
время, когда одни из них достигли более высокого уровня развития, другие ещё
остаются в менее развитом состоянии.
В жизни экологических систем действуют общие термодинамические принципы и
законы сохранения энергии, вещества, информации. Закон (принцип) энергетической
проводимости: поток энергии, вещества и информации в системе как целом должен
быть сквозным, охватывающим всю систему или косвенно отзывающимся в ней. Иначе
система не будет иметь свойства единства. Это закон (принцип) не следует
понимать слишком упрощенно и ограничиваться короткими интервалами времени.
Очевидно, для экологической системы определенного уровня иерархии длительность
прохождения потока энергии, вещества и информации будет специфичной. Больше
всего расчетов сделано для воды. Водообмен в
биологической особи занимает часы, влаги в атмосфере (следовательно, и в аэробиосфере) - 8 дней, свободных континентальных
поверхностных вод - от 16 дней в реках до 17 лет в озерах; подземные воды
обновляются за 1400 лет, а воды океана за 2500 лет (Реймерс,
1994).
Закон сохранения массы: сумма массы вещества системы и массы эквивалентной
энергии, полученной или отданной той же системой, постоянна. Первоначальная
формулировка этого закона - масса (вес) веществ до химической реакции равна
массе (весу) веществ после химической реакции, а более расширительно - масса
поступающего в систему вещества минус масса выходящего из системы вещества
равна массе накапливаемого в системе вещества. Если накопления или убывания
вещества в системе не наблюдается, она находится в устойчивом, гомеостатическом
состоянии. Близкий к этому закон сохранения энергии, или первый принцип (закон,
начало) термодинамики, гласит: любые изменения в изолированной системе
оставляют её общую энергию постоянной; или: при всех макроскопических процессах
энергия не создается и не исчезает, а лишь переходит из одной формы в другую.
В агроэкологии особенно значим второй принцип (начало, или закон)
термодинамики, имеющий множество формулировок и смысловых оттенков. Три
важнейших для экологии: 1) энергетические процессы могут идти самопроизвольно
только при условии перехода энергии из концентрированной формы в рассеянную; 2)
потери энергии в виде недоступного для использования тепла всегда приводят к
невозможности стопроцентного перехода одного вида энергии (кинетической) в
другую (потенциальную) и наоборот; результат - невозможно создать вечный
двигатель 2-го рода; 3) закон возрастания энтропии: в замкнутой (изолированной
в тепловом и механическом отношении) системе энтропия либо остается неизменной
(если в системе протекают обратимые, равновесные процессы), либо возрастает
(при неравновесных процессах) и в состоянии равновесия достигает максимума.
В открытых системах, согласно теореме сохранения упорядоченности в них,
сформулированной И.Р. Пригожиным (1955), энтропия не
возрастает - она в открытых системах падает до тех пор, пока не достигается
минимальная постоянная величина, всегда большая нуля. При этом в системе
вещество распределяется неравномерно и организуется таким образом, что местами
энтропия возрастает, а в других местах резко снижается. В целом же, используя
поток энергии, система не теряет упорядоченности. Деятельность же живых систем
всегда негэнтропийна, пока сохраняется их свойство
системности: таково индивидуальное развитие организмов, средообразующая
их роль в биосфере и другие процессы в открытых системах.
Все системы, с которыми приходится иметь дело агроэкологии, негэнтропийны,
упорядочены таким образом, что, по известному меткому выражению Ю. Одума, как бы "откачивают из сообщества
неупорядоченность". Это происходит до тех пор и постольку, поскольку
действует принцип Ле Шателье
- Брауна: при внешнем воздействии, выводящем систему из состояния устойчивого
равновесия, это равновесие смещается в том направлении, при котором эффект
внешнего воздействия ослабляется. Поскольку в биосфере механизм осуществления
принципа Ле Шателье -
Брауна основывается на функционировании систем живого, оно, это
функционирование, как постулировал В.И. Вернадский, служит основным регулятором
общеземных процессов. Действие принципа Ле Шателье - Брауна в рамках биосферы в наши дни глубоко
нарушено. Если в конце прошлого века еще происходило увеличение биологической
продуктивности и биомассы в ответ на возрастание концентрации углекислого газа
в атмосфере, то с начала нашего века это явление не обнаруживается. Наоборот, биота выбрасывает углекислый газ, а биомасса её
автоматически снижается. Поскольку биосфера имеет лишь одно устойчивое
состояние, единственным способом восстановить действие принципа Ле Шателье - Брауна будет
сокращение площадей антропогенно измененных земель.
Важное значение для экологических и биолого-эволюционных процессов имеет
общефизический закон минимума диссипации (рассеивания) энергии Л. Онсагера, или принцип экономии энергии: при вероятности
развития процесса в некотором множестве направлений, допускаемых началами
термодинамики, реализуется то, которое обеспечивает минимум диссипации энергии
(или минимум роста энтропии).
Тесно связан с другими энергетическими постулатами закон максимизации энергии и
информации: наилучшими шансами на само сохранение обладает система, в
наибольшей степени способствующая поступлению, выработке и эффектному
использованию энергии и информации; максимальное поступление вещества как
такового не гарантирует системе успеха в конкурентной группе других аналогичных
систем.
Первоначально закон энергии и информации был сформулирован как закон
максимизации только энергии: в соперничестве с другими системами выживает
(сохраняется) та из них, которая наилучшим образом способствует поступлению
энергии и использует максимальное её количество наиболее эффективным способом.
"С этой целью система: 1) создаёт накопители (хранилища)
высококачественной энергии; 2) затрачивает накопленную энергию на обеспечение
поступления новой энергии; 3) обеспечивает кругооборот различных веществ; 4)
создает механизмы регулирования, поддерживающие устойчивость системы и её
способность приспособления к изменяющимся условиям; 5) налаживает с другими системами
обмен, необходимый для обеспечения потребности в энергии специальных
видов" (Г. Одум, Э. Одум.
Энергетический базис человека и природы. М.: Прогресс, 1978. С. 72-73).
Закон максимизации энергии и информации имеет более обобщенную и краткую
формулировку в виде принципа максимизации мощи: системы с мощной энергетикой
как правило вытесняют системы с более низкой энергетической "мощью".
Обобщающим выводом из серии закономерностей энергетического ряда может служить
правило основного обмена: любая большая динамическая система в стационарном
состоянии использует приход энергии, вещества и информации главным образом для
своего самоподдержания и саморазвития. Таково
соотношение основного метаболизма и производимой организмом работы, таково
положение в экосистемах и хозяйстве.
Общие принципы формирования иерархии систем: 1) дублирование относительно
разнокачественных структур, составляющих в своей организованной совокупности
нечто новое, т. е. наличие свойства эмерджентности
(древние говорили: целое больше суммы его частей) и 2) определенность
функциональной цели организации в рамках связей со средой и внутренних
возможностей системы. Сам принцип иерархической организации, или принцип интегративных уровней, в биологии и экологии принимается
как аксиома или эмпирически наблюдаемый факт. Столь же аксиоматически
утверждается и проявление эмерджентности с переходом
от одного уровня иерархии к другому. Эмерджентность -
наличие у системного целого особых свойств, не присущих его подсистемам,
элементам и (несистемным) блокам, а также сумме элементов и блоков, не
объединенных системообразующими связями. Свойство
цели как функциональное состояние и закономерность построения системы,
достигаемая путем возникновения обратных связей, создает некое поле
взаимодействий. Это поле не может быть бесконечным по способу организации, так
как любая система существует в рамках ее характерного времени и пространства
(размера).
Размер системы, или характерный размер системы, - это пространственная её
протяжённость (объем, площадь) или масса, а также минимальное (максимальное)
число подсистем, позволяющее системе существовать и функционировать с
осуществлением саморегуляции и самовосстановления в
рамках своего характерного времени. Системное время (характерное, или
собственное, время системы) - это время, рассматриваемое в рамках периода
существования данной системы и/или происходящих в ней процессов. Эти процессы
ограничены термодинамикой системы, её функциональными особенностями. В то же
самое время, поскольку системы с одной и той же функциональной целью,
формируемой обратными связями, располагаются на одном уровне иерархии и поэтому
ограничены однотипным характерным временем и пространством, их построение
подчинено одному ряду внутренних закономерностей.
Ограничение числа и форм взаимодействия подсистем одного системного уровня
ограничивает "фантазию природы", укладывает её в строго
лимитированный закономерный ряд образований. Периодический закон химических
элементов Д. И. Менделеева (свойства химических элементов, проявляющиеся в простых
веществах и соединениях, находятся в периодической зависимости от заряда ядер
их атомов) был первой формализацией "третьего измерения" таблицы
"системы систем". Тот же характер имеет закон гомологических рядов и
наследственной изменчивости Н. И. Вавилова. (Родственные виды, роды, семейства
и др. систематические категории обладают закономерно возникающими гомологичными
генами и порядками генов в хромосомах, сходство которых тем полнее, чем
эволюционно ближе сравниваемые таксоны. Гомология генов у родственных форм
проявляется в сходстве рядов их наследственной изменчивости. Циклы изменчивости
проходят через все роды и виды, составляющие семейство). Менее известен и
признан периодический закон географической зональности А. А. Григорьева - М. И.
Будыко: со сменой физико-географических поясов Земли
аналогичные ландшафтные зоны и их некоторые общие свойства периодически
повторяются.
Закон периодичности строения системных совокупностей или системопериодический
закон (формулировка Н.Ф. Реймерса, 1990): принципы структурного
построения и управления однородных природных систем в иерархическом
соподчинении и особенно сложении таких же природных систем одного уровня
организации (иерархии) повторяются с некоторой правильностью в зависимости от
действия единого (комплекса) системообразующего
фактора (факторов).
Периодичность систем природы не исчерпывается приведенными выше частными
законами периодического строения элементов, гомологических рядов и
географической зональности. Она проявляется в строении космических тел,
прохождении геологических эпох, сукцессионных стадий
развития экосистем и во многих других случаях. Системопериодический
закон можно считать всеобщей закономерностью, действующей во всем системном
мире, в том числе в системах с участием живого - агроэкосистемах.
Абсолютно изолированные системы вне связи с окружающей их средой длительное
время существовать не могут в силу действия второго начала термодинамики
(закона возрастания энтропии). Самые общие закономерности взаимосвязи система -
среда обобщены в философски расширенной формулировке принципа дополнительности Нильса Бора. Изначальный смысл этого
принципа был расширен: две взаимосвязанные, но различные материальные системы
дополняют друг друга в своем единстве и противоположности. Такой "общий
принцип дополнительности" очень существенен для
функционирования агроэкологических систем.
Без относительного (динамического) равновесия в рамках "общего принципа дополнительности" взаимодействие будет
кратковременным: система разрушится. Этого не происходит в силу действия
принципа Ле Шателье -
Брауна. Следствием этого служит принцип торможения развития: в период
наибольших потенциальных темпов развития системы возникают максимальные
тормозящие эффекты. В зависимости от силы процесса они могут быть заметны или скрыты
ходом этого процесса.
Вещество и энергия для функционирования и развития системы могут поступать лишь
из окружающей эту систему среды, и только за счет этой среды может существовать
и прогрессировать любая система. Этот очевидный факт отражает закон развития
системы за счет окружающей ее среды: любая система может развиваться только за
счет использования материально-энергетических и информационных возможностей
окружающей её среды; абсолютно изолированное саморазвитие невозможно.
Энергия, вещество и информация, поступающие в систему извне и выступающие как
факторы её жизни, действуют не в "чистом" виде, а селектируются
и видоизменяются этой системой. Если они проходят предварительно через
надсистемы рассматриваемой системы, то эти процессы идут многократно и до неё
доходят в трансформированном всеми надсистемами виде. Поиск прямых связей между
очень далекими по иерархическому уровню системными образованиями, если эти
связи не настолько сильны, что проходят "транзитом" через
промежуточный ряд иерархии систем, как правило, бывает очень затруднителен.
Действует принцип преломления действующего фактора в иерархии систем. При этом
не следует сбрасывать со счета и саму рассматриваемую систему как преобразующий
фактор члена иерархии: фактор, действующий на систему, преломляется через всю
иерархию её надсистем и через функциональные особенности самой системы. В связи
с этим, как правило, воздействия надсистем не равны по силе и не совпадают по
времени с интенсивностью и моментом их возникновения.
Принцип преломления действующего фактора внутри системы: внешние для систем
воздействия, как правило, проявляются не прямо, а опосредованы механизмами
функционирования этой системы; они могут быть ослаблены её буферными свойствами
или усилены возникающими цепными реакциями, справедлив для не моментально
гибельных воздействий на систему.
В силу преломления действующего фактора в иерархии систем и наличия многих
"фильтров" этот фактор либо ослабляется, либо усиливается, а чаще
всего оказывается неравномерным по силе воздействия с ходом времени. Система
немедленно или с задержкой реагирует на возникающие флуктуации. Этот факт
констатируется законом функционально-системной неравномерности: темпы реакций и
прохождения фаз развития системы (в ответ на действие внешних факторов)
закономерно неравномерны - они то убыстряются (усиливаются), то замедляются
(ослабевают).
В развитии систем действует правило затухания процессов: насыщающиеся системы с
увеличением степени равновесности с окружающей их
средой или внутреннего гомеостаза (в случае изолированности системы)
характеризуются затуханием в них динамических процессов. Это затухание может
быть постепенным при линейности процесса или идти скачкообразно по затухающей
синусоиде, лавинообразно (экспоненциально и сверхэкспоненциально).
Такое явление характерно для насыщающихся растворов, термодинамических
процессов, темпов размножения акклиматизированных организмов и др.
Частным, но важным случаем правила затухания процессов и одновременно
следствием закона оптимальности, перекликающегося в данном случае со вторым
принципом термодинамики, служит выведенный Г.Ф. Хильми
закон растворения системы в чуждой среде или закон обеднения разнородного
живого вещества (биоты) в островных его сгущениях
(принцип организационной деградации): "индивидуальная система, работающая
в ...среде с уровнем организации более низким, чем уровень самой системы
обречена: постепенно теряя структуру, система через некоторое время растворится
в окружающей... среде". Это общесистемный закон. Он тесно связан с законом
оптимальности и в значительной мере отражает термодинамику малой системы,
находящейся в чуждой среде. Сохранить островную систему изолировано на малых
территориях при любых условиях в длительном интервале времени практически
невозможно.
Принцип экологической комплементарности (дополнительности): никакая функциональная часть экосистемы
(экологический компонент, элемент и т. п.) не может существовать без других
функционально дополняющих частей. Происходит это потому, что каждая подсистема
связана с другими согласно общему принципу дополнительности
и группе закономерностей сложения систем.
Принцип экологической конгруэнтности (соответствия): функционально дополняя
друг друга, живые составляющие экосистемы вырабатывают для этого
соответствующие приспособления, скоординированные с условиями абиотической
среды, в значительной мере преобразуемой теми же организмами (биоклимат и т.
п.). То есть наблюдается двойной ряд соответствия - между организмами и ими и
средой их обитания - внешней и создаваемой ценозом.
Принцип (закон) формирования экосистемы (функционально-пространственной
экологической целостности, связи биотоп - биоценоз): длительное существование
организмов возможно лишь в рамках экологических систем, где их компоненты и
элементы дополняют друг друга и соответственно приспособлены друг к другу. Это
обеспечивает воспроизводство среды обитаний каждого вида и относительно
неизменное существование всех экологических компонентов. Принцип формирования
экосистемы есть суммарное отражение принципа экологической комплементарности
(дополнительности) и принципа экологической
конгруэнтности (соответствия).
Закон однонаправленности потока энергии: энергия,
получаемая сообществом (экосистемой) и усваиваемая продуцентами, рассеивается
или вместе с их биомассой передается консументам
первого, второго и т. д. порядков, а затем редуцентам
с падением потока на каждом трофическом уровне в результате процессов,
сопровождающих дыхание. Поскольку в обратный поток (от редуцентов
к продуцентам) поступает ничтожное количество изначально вовлечённой энергии,
говорить о "круговороте энергии" нельзя. Существует лишь круговорот
веществ, поддерживаемый потоком энергии. Однонаправленность
потока энергии формирует в экосистемах относительно замкнутый круговорот
веществ. Закономерности распространения информации также связаны с потоком
энергии и перемещением вещества, но они пока еще мало изучены. Видимо, здесь
большую роль играют горизонтальные связи между экосистемами, хотя
информационные сети явно гуще и лучше налажены в пределах образований каждого
уровня их иерархии. Пока еще поле обобщений в области экоинформатики
почти не возделано (Реймерс, 1994).
Закон внутреннего динамического равновесия Н.Ф. Реймерса:
вещество, энергия, информация и динамические качества отдельных природных
систем (в том числе экосистем) и их иерархии взаимосвязаны настолько, что любое
изменение одного из этих показателей вызывает сопутствующие
функционально-структурные количественные и качественные перемены, сохраняющие
общую сумму вещественно-энергетических, информационных и динамических качеств
систем, где эти изменения происходят, или в их иерархии.
Справедливость закона внутреннего динамического равновесия доказывается всей
практикой и особенно характером региональных экологических катастроф. Этот
закон - является одним из основных в управлении природопользованием.
Важные для практики следствия из закона внутреннего динамического равновесия
Н.Ф. Реймерс (1990, 1994) приводит в следующем
порядке:
1. Любое изменение среды (вещества, энергии, информации, динамических качеств
экосистем) неизбежно приводит к развитию природных цепных реакций, идущих в
сторону нейтрализации произведенного изменения или формирования новых природных
систем, образование которых при значительных изменениях среды может принять необратимый
характер;
2. Взаимодействие вещественно-энергетических экологических компонентов
(энергия, газы, жидкости, субстраты, организмы-продуценты, консументы
и редуценты), информации и динамических качеств
природных систем количественно нелинейно, т. е. слабое воздействие или
изменение одного из показателей может вызвать сильные отклонения в других (и во
всей системе в целом). Например, малое отклонение в составе газов атмосферы, её
загрязнение окислами серы и азота вызвают огромные
изменения в экосистемах суши и водной среды. Именно оно привело к возникновению
кислотных осадков, а с ними к деградации и гибели лесов в Европе и Северной
Америке, обезрыбливанию озер Скандинавии, нарушению циклов развития морских
организмов, личиночные стадии которых проходят на мелководьях. Столь же
абсолютно незначительное изменение концентрации СО2 ведет к возникновению
тепличного эффекта, а комплекс малых перемен формирует массовые размножения
организмов;
3. Производимые в крупных экосистемах изменения относительно необратимы -
проходя по их иерархии снизу вверх, от места воздействия до биосферы в целом,
они меняют глобальные процессы и тем самым переводят их на новый эволюционный
уровень;
4. Любое местное преобразование природы вызывает в глобальной совокупности
биосферы и в её крупнейших подразделениях ответные реакции, приводящие к
относительной неизменности эколого-экономического потенциала, увеличение
которого возможно лишь путем значительного возрастания энергетических вложений.
Искусственный рост эколого-экономического потенциала ограничен
термодинамической (тепловой) устойчивостью природных систем.
Пока изменения среды слабы и произведены на относительно небольшой площади, они
или ограничиваются конкретным местом, или "гаснут" в цепи иерархии
экосистем. Но как только перемены достигают существенных значений для крупных
экосистем, например, происходят в масштабах больших речных бассейнов или в
размерах, ограниченных правилами одного и десяти процентов, они приводят к
существенным сдвигам в этих обширных природных образованиях, а через них,
согласно 2-му следствию из обсуждаемого закона, и во всей биосфере Земли.
Будучи относительно необратимыми (3-е вышеприведенное следствие), изменения в
природе в конечном итоге оказываются и трудно нейтрализуемыми с
социально-экономической точки зрения: их выправление требует слишком больших
материальных средств и физических усилий. Иногда возникает даже ситуация
"чем больше пустынь мы превратим в цветущие сады, тем больше цветущих
садов мы превратим в пустыни". При этом в силу нелинейности процессов
(следствие 2) опустынивание по темпам значительно опережает создание
"цветущих садов" и теоретически не может не опережать, поскольку
такое создание базируется на нарушении компонентного равновесия в экосистемах (Реймерс, 1994).
Сдвигая динамически равновесное (квазистационарное)
состояние природных систем с помощью значительных вложений энергии (например,
путем распашки и других агротехнических приёмов), люди нарушают соотношение
экологических компонентов, достигая увеличения полезной продукции (урожая) или
состояния среды, благоприятного для жизни и деятельности человека. Если эти
сдвиги "гаснут" в иерархии природных систем (от элементарных до
биосферы и экосферы планеты в целом) и не вызывают
термодинамического разлада, положение благоприятно, или, во всяком случае,
терпимо. Однако излишнее вложение энергии и возникающий в результате
вещественно-энергетический разлад ведут к снижению природно-ресурсного
потенциала вплоть до опустынивания территории.
Закон экологической корреляции: в экосистеме, как и в любом другом целостном
природно-системном образовании с участием живого, все входящие в неё виды
живого и абиотические экологические компоненты функционально соответствуют друг
другу. Выпадение одного элемента системы, например, исчезновение вида,
неминуемо ведет к исключению всех тесно связанных с этим элементом системы
других её частей и функциональному изменению целого в рамках внутреннего
динамического равновесия. При этом выпадение элемента может вести и к включению
нового блока, т. е. исключению с положительным, а не отрицательным знаком -
внедрению в экосистему нового вида или их группы всегда вместе с их консорбентами.
Действие закона экологической корреляции ведет к скачкообразным изменениям
экологической устойчивости: при достижении порога изменения функциональной
целостности происходит часто неожиданный срыв - экосистема теряет свойство
надежности. Например, многократное увеличение концентрации загрязняющего
вещества может не приводить к заметным последствиям, но затем ничтожная его
прибавка приведет к катастрофе.
Правило оптимальной компонентной дополнительности:
никакая экосистема не может самостоятельно существовать при искусственно
созданном значительном и перманентном избытке или недостатке одного из
экологических компонентов. "Нормой" экологического компонента следует
считать ту, которая обеспечивает экологическое равновесие определенного типа,
позволяющее функционировать именно той экосистеме, которая эволюционно
сложилась и соответствует балансу в природной надсистеме и всей иерархии
природных систем на данной единице пространства (в конкретном биотопе).
Принцип экологической (рабочей) надежности: эффективность экосистемы, её
способность к самовосстановлению и саморегуляции (в
пределах естественных колебаний) зависит от её положения в иерархии природных
образований (краевое, маргинальное, положение невыгодно), степени
взаимодействия её компонентов и элементов, а также от частных приспособлений
организмов (размеры, продолжительность жизни, скорость смены поколений,
отношение продуктивности к биомассе и т. п.), составляющих биоту
экосистемы.
Принципы видового обеднения:
- консорционной целостности ("никто не гибнет в
одиночку"): с исчезновением из экосистемы вида, образующего консорцию, исчезают и многие консорбенты,
не входящие в другие консорции той же или другой
территориально и функционально близкой экосистемы;
- биологического замещения ("принцип незваного гостя"): вновь
внедрившийся в экосистему вид всегда, вырабатывая свою экологическую нишу,
сужает возможности менее конкурентоспособных видов и тем ведет к их
исчезновению или лишь слегка видоизменяет экологические ниши функционально
близких видов, создавая предпосылки для сокращения их численности или,
наоборот, массового размножения; при этом меняется все сообщество, включая даже
как будто не связанные с внедрившимся видом формы (в силу смещения общего
экологического равновесия);
- смены трофических цепей ("экосистема не терпит пустоты"):
исчезнувшая трофическая цепь видов сменяется цепью эколого-энергетических
аналогов, что позволяет экосистеме усваивать и перерабатывать поступающую извне
энергию;
- неопределенности хозяйственного значения смены видов ("старый друг лучше
новых двух"): при замене трофических цепей вновь проникающие в экосистему
виды могут быть как желательными, так и нежелательными в хозяйственном и
медицинском отношении.
Принцип сукцессионного замещения: на жизнепригодном субстрате биотические сообщества
последовательно формируют закономерный ряд экосистем, ведущий к наиболее
устойчивой в данных условиях климаксовой фазе в чисто
природных условиях или узловой фазе при природно-антропогенном режиме.
Сказанное не означает, что регионально не существует мозаика климаксовых экосистем, обусловленная топографическими,
геоморфологическими и другими условиями среды. Именно такая мозаика лежит в
основе теории мозаичного климакса Р. Уиттекера: сукцессионные ряды в долине реки, в сосняках, протянувшихся
вдоль нее, в лесах склонов гор и на их верхних частях составляют достаточно
четкую мозаику. Если бы была приведена карта восстановленной растительности,
эта закономерность выступила бы еще ярче. Но это не значит, что, скажем,
климакс приречного ельника может как угодно варьировать.
Закон сукцессионного замедления: процессы, идущие в
зрелых равновесных экосистемах, находящихся в устойчивом состоянии, как
правило, проявляют тенденцию к снижению темпов.
Правило максимума энергии поддержания зрелой системы Г. Одума
и Р. Пинкертона: сукцессия идет в направлении
фундаментального сдвига потока энергии в сторону увеличения её количества,
направленного на поддержание системы.
Принцип "нулевого максимума" или минимизации прироста в зрелой
экосистеме: экосистема в сукцессионном развитии
стремится к образованию наибольшей биомассы при наименьшей биологической
продуктивности.
Принцип "сукцессионного очищения" или
стабилизации и минимизации видового состава климакса: разнообразие стремится к
пику на ранних или средних фазах сукцессии, а затем снижается в климаксе.
Правило сукцессионного мониторинга (индикации
состояния среды) или степени завершенности сукцессии: там, где разнообразие
видов недостаточно для формирования биосреды,
служащей основой нормального, естественного хода сукцессионного
процесса, а сама среда резко нарушена (по глубине или частоте нарушений),
сукцессия не достигает фазы климакса, а заканчивается узловым сообществом, параклимаксом, длительно- или даже кратковременнопроизводным
сообществом. Качество среды данной территории, суммарная напряженность
антропогенного воздействия отражается на степени завершенности сукцессионного ряда. Чем глубже нарушенность
среды какого-то пространства, тем на более ранних фазах оканчивается сукцессия.
Достижение климакса при потере одного или группы видов в результате их
уничтожения (реже вымирания, тоже, как правило, связанного с антропогенным
исчезновением местообитаний) не есть полное восстановление природной
обстановки. Фактически это новая экосистема, так как в ней возникли новые связи
и утеряны многие старые, сложилась иная "притёртость"
видов. Вернуться в старое состояние экосистема не может, так как утерянный вид
невосстановим.
Закон эволюционно-экологической необратимости: экосистема, потерявшая часть
своих элементов или сменившаяся другой в результате дисбаланса экологических
компонентов, не может вернуться к первоначальному состоянию в ходе сукцессии,
если в ходе изменений произошли эволюционные перемены в экологических элементах
(сохранившихся или временно утерянных) или один либо группа видов исчезла
навсегда или на (системно) долгий срок.
Правило объединения в популяции С.С. Четверикова:
любая популяция обладает строго определенной генетической, фенотипической,
половозрастной и другой структурой, она не может состоять из меньшего числа
индивидов, чем необходимо для обеспечения стабильной реализации этой структуры
и устойчивости популяции к факторам внешней среды. Именно в этом состоит
принцип минимального размера популяций. Такой размер не есть константа для
любых видов, он строго специфичен для каждой популяции. Выход за пределы
минимума чреват для популяции гибелью: она уже не будет в состоянии самовозобновиться. Однако её возможно восстановить
искусственно, поскольку каждый индивид содержит в себе хотя и обедненное, но
все же весьма значительное число генетических задатков вида, к которому он
принадлежит.
Если есть минимум, то возможен и максимум. Правило популяционного максимума
конкретизируют два обобщения. Первое из них известно как теория лимитов
популяционной численности, или теория X. Г. Андреварты-Л.
К. Бирча: численность естественных популяций
ограничена истощением пищевых ресурсов и условий размножения, недоступностью
этих ресурсов и слишком коротким периодом ускорения роста популяции. Второе
обобщение дополняет первое. Оно носит название теории биоценотической регуляции
численности популяции К. Фридерихса: регуляция
численности популяции есть результат комплекса воздействий абиотической и
биотической среды в местообитании вида.
Правило пищевой корреляции В. Уини-Эдвардса: в ходе
эволюции сохраняются только те популяции, скорость размножения которых скоррелирована с количеством пищевых ресурсов среды их
обитания.
Теория, или принцип Д. Лэка: среднее количество яиц,
откладываемых птицами, эволюционно ограничено возможностями выращивания
выведшихся из них птенцов. Очевидно, этот лимит связан с комплексом условий:
интенсивностью обмена веществ, длиной дня, наличием пищи, ее близостью и т. д.
В основе, безусловно, лежит энергетика системы - она отработана эволюцией вида
в конкретных условиях его обитания.
Правило колебаний (цикличности) численности: никакая популяция не находится в
состоянии абсолютной уравновешенности числа особей, обязательно помимо сезонных
изменений численности особей возникают периодические флуктуации, обусловленные
внешними по отношению к популяции факторами, и осцилляции, связанные с собственными
(внутренними) динамическими изменениями популяции.
Закон количественной константности популяционного населения или правило
максимума размера колебаний плотности популяционного населения: существуют
определенные верхние и нижние пределы для размеров популяции, которые
соблюдаются в природе или которые теоретически могли бы существовать в течение
сколь угодно длительного отрезка времени.
Отклонение от этого закона подчеркивает неблагополучие ситуации в регионе,
служит биоиндикатором разлада в его экосистемах.
Правило максимальной рождаемости (воспроизводства): в популяции имеется
тенденция к образованию теоретически максимально возможного количества новых
особей. Оно достигается в идеальных условиях, когда отсутствуют лимитирующие
экологические факторы, и размножение ограничено лишь физиологическими
особенностями вида.
Следствием правила максимальной рождаемости (воспроизводства) популяции служит
правило стабильности её возрастной структуры: любая естественная популяция
стремится к стабильной возрастной структуре, четкому количественному
распределению особей по возрастам. Для многих организмов правило стабильности
возрастной структуры популяции следует дополнить правилом стабильности
соотношения полов. Вместе эти два правила составляют общее правило стабильности
половозрастной структуры популяции.
Принцип территориальности: все особи и функциональные их объединения в
популяции обладают индивидуальным и групповым пространством, возникающим в
результате действия механизмов активного разобщения - поведенческого у высших
организмов или физико-химического (аллелопатия и др.) у низших.
Принцип скопления (агрегации) особей В. Олли:
скопление (агрегация) особей усиливает конкуренцию между индивидами, но
способствует выживанию группы в целом; следовательно, как перенаселенность, так
недонаселенность, препятствующая агрегации, могут
служить лимитирующими факторами. Принцип агрегации особей диктует необходимость
образования стай, стад, колоний, оптимальной густоты и распределения растений в
посевах и имеет другие практические следствия.
Правило топографического, или популяционного, кружева ареала (Н.Ф. Реймерс): популяция заселяет пространство неравномерно,
оставляя "пустые" места, которые непригодны для ее жизни, и
распадаясь на экологически разнородные микропопуляции,
каждая из которых приурочена к определенному местообитанию.
Принцип (теорема) В. Людвига: вопреки стабилизации экологических ниш вновь
возникшие внутри популяции генотипы могут образовывать экологические субниши, расширять использование ресурсов среды.
Теория (принцип) А. Николсона: популяции - стабильные системы, способные
противостоять факторам внешней среды и контролировать (компенсировать) эти
факторы изменением своей плотности населения.
Закона единства организм - среда (В.И. Вернадский): жизнь развивается в
результате постоянного обмена веществом и информацией на базе потока энергии в
совокупном единстве среды и населяющих её организмов. Связано это с активностью
всех биосистем. А поскольку отношения организма и его среды системны, действует
принцип экологического соответствия: форма существования организма всегда
соответствует условиям его жизни. Если рассматривать эту закономерность не отвлеченно-философски, а конкретно-биологически,
то формулируется правило соответствия условий среды жизни генетической
предопределенности организма: вид организмов может существовать до тех пор и
постольку, поскольку окружающая его среда соответствует генетическим
возможностям приспособления этого вида к ее колебаниям и изменениям.
Закон максимума биогенной энергии (энтропии) В. И. Вернадского-Э.
С. Бауэра: любая биологическая или биокосная (с
участием живого) система, находясь в подвижном (динамическом) равновесии с
окружающей её средой и эволюционно развиваясь, увеличивает своё воздействие на
среду. Давление растет до тех пор, пока не будет строго ограничено внешними
факторами (надсистемами или другими конкурентными системами того же уровня
иерархии), либо не наступит эволюционно-экологическая катастрофа.
Максимальному давлению жизни, максимизации биогенной энергии (энтропии)
противостоит действие закона давления среды жизни, или закона ограниченного
роста Ч. Дарвина, который гласит, что хотя не существует исключений из правила,
что потомство одной пары особей, размножаясь в геометрической прогрессии,
стремится заполнить весь земной шар, имеются ограничения, не допускающие этого
явления. Эти ограничивающие силы определенным образом упорядочены, что
позволило сформулировать довольно большое количество формализованных правил,
принципов и законов.
Наиболее общее значение имеет закон совокупного (совместного) действия
факторов: взаимосвязь экологических факторов и их взаимное усиление и
ослабление определяют их воздействие на организм и успешность его жизни. При
этом важны не только воздействия извне, но и физиологическое состояние
организма. Иногда закон совокупного действия факторов называют законом
физиологических взаимодействий. Обычно этот закон связывают с урожаем
сельскохозяйственных культур, поскольку он был сформулирован немецким
агрохимиком и физиологом растений Э.А. Митчерлихом
под названием "закона эффективности факторов" в приложении к
сельскохозяйственным культурам, Б. Бауле назвал этот закон "законом
совокупного действия", а А. Тинеман
дополнил и развил. Закон носит имена этих исследователей.
Совокупность факторов воздействует сильнее всего на те фазы развития
организмов, которые имеют наименьшую экологическую валентность - минимальную
способность к приспособлению.
В совокупном давлении среды выделяются факторы, которые сильнее всего ограничивают
успешность жизни организма. В наиболее общем виде эту закономерность
формулирует закон ограничивающих (лимитирующих) факторов Ф. Блэкмана:
факторы среды, имеющие в конкретных условиях пессимальное
значение, особенно затрудняют (ограничивают) возможность существования вида в
данных условиях, вопреки и несмотря на оптимальное сочетание других отдельных
условий. Очень близок к нему закон толерантности В. Шелфорда:
лимитирующим фактором процветания организма (вида) может быть как минимум, так
и максимум экологического воздействия, диапазон между которыми определяет
величину выносливости (толерантности) организма к данному фактору.
Выносливость организмов зависит от их возраста и пола. Это значимо в текущей
жизни и в процессе эволюции: женский организм более чуток к факторам среды в
ходе эволюции вида, чем мужской (а мужской - к индивидуальным факторам
воздействия). Эта закономерность известна как правило Геодекяна,
или правило меньшей эволюционно-экологической толерантности женского организма.
Закон минимума Ю.Либиха: выносливость организма определяется самым славым звеном в цепи его экологических потребностей, т. е.
жизненные возможности лимитируют экологические факторы, количество и качество
которых близки к необходимому организму или экосистеме минимуму; дальнейшее их
снижение ведёт к гибели организма или деструкции экосистемы.
Поскольку любой фактор может оказаться в минимуме, лишь их оптимальная
совокупность обеспечивает процветание. Этот факт сформулирован в виде закона
равнозначности всех условий жизни: все условия среды, необходимые для жизни,
играют равнозначную роль.
Правило взаимодействия факторов в законе минимума: организм в определенной мере
способен заменить дефицитное вещество или другой действующий фактор жизни
функционально близким веществом или фактором (например, одно вещество другим,
химически близким) - вызвало поток аналогичных постулатов. Среди них закон
относительности действия лимитирующих факторов, или закон Лундегарда
- Полетаева: форма кривой роста численности популяции (ее биомассы) зависит не
только от одного вещества с минимальной концентрацией, а от концентрации и
свойств других ионов, имеющихся в среде. Важной поправкой и дополнением служит
закон неоднозначного (селективного) действия фактора на различные функции организма:
любой экологический фактор неодинаково влияет на функции организма, оптимум для
одних процессов, например, дыхания, не есть оптимум для других, например,
пищеварения, и наоборот.
Дополнительное правило замещения дефицитных веществ закона минимума Ю. Либиха
было углублено Э. Рюбелем в виде закона (эффекта)
компенсации (взаимозаменяемости) факторов: отсутствие или недостаток некоторых
экологических факторов может быть компенсирован другим близким (аналогичным)
фактором. Например, недостаток света может быть компенсирован для растения
обилием углекислого газа. Частным случаем закона относительности действия
лимитирующих факторов Лундегарда - Полетаева и закона
компенсации факторов Рюбеля служит эффект Лундегарда - Стиберга - Якобсона:
увеличение биомассы растения при постоянном или даже пониженном содержании
какого-либо питательного элемента.
В.В. Алехин сформулировал близкое к изложенным правило замещения экологических
условий: любое условие внешней среды в некоторой степени может замещаться
другим; внутренние причины экологических явлений при аналогичном внешнем
проявлении могут быть различными.
Стремлению сузить сферу действия закона минимума противостоит закон
незаменимости фундаментальных факторов, связываемый с именем В.Р. Вильямса. Он
жестко сформулировал ограничение: полное отсутствие в среде фундаментальных
экологических (физиологических) факторов (света, воды, биогенов
и т.п.) не может быть заменено другими факторами. Совершенно очевидно, что
любой из экологических компонентов до конца не может быть заменен - при
отсутствии энергии нет жизни, то же при полном безводье и так далее. Для
элементарных потребностей это очевидно, но в более сложных ситуациях, особенно
когда речь идет о фундаментальных, но не столь известных и осознанных факторах
такая очевидность исчезает.
Пока речь шла об общем длительном воздействии на организмы. К этой группе
примыкает несколько отличное от других правило или закон фазовых реакций
("польза - вред"): малые концентрации токсиканта
действуют на организм в направлении усиления его функций (их стимулирования) ,
тогда как более высокие концентрации угнетают или даже приводят его к смерти.
Н.Ф. Реймерс (1994) указывал, что эта
токсикологическая закономерность справедлива для многих, но не для всех
ядовитых веществ и особенно спорна для малых доз радиации. Благоприятное
воздействие малых доз вредных экологических факторов называют гормезисом. Радиационное воздействие пороговых, малых и
даже запороговых доз пока однозначно не известно.
Теоретически вывод биологических систем из состояния равновесия с помощью
слабых доз токсикантов не может идти на пользу.
Особенно это важно учитывать в свете закона "всё или ничего" Г.П. Боулича: подпороговые раздражения
не вызывают нервного импульса ("ничего"), а пороговые стимулы или
суммирование подпороговых воздействий создает условия
для формирования максимального ответа ("всё"), или в общесистемном
смысле - слабые воздействия могут не вызвать у природной системы ответных
реакций до тех пор, пока, накопившись, они не приведут к развитию бурного
динамического процесса. При этом между воздействиями нет линейной
пропорциональности, и могут интегрироваться различные факторы (температура,
влажность, радиация, химические вещества и т. п.). На этом законе базируется
концепция пороговых значений системных воздействий. Наличие порогов,
несомненно, но и континуум нарастания воздействия очевиден.
Хотя слабые раздражения по принципу "ничего" не воспринимаются, чем
сильнее раздражитель, тем труднее субъективно оценить его количественно; это
положение называют законом субъективной количественной оценки раздражителя Э.
Вебера - Г. Фехнера. Чем контрастней фон, тем легче
улавливаются и оцениваются раздражения при их слабости, но сильные источники
раздражения уже могут не давать эффекта разницы в восприятии.
Закон пирамиды энергий или закон (правило) 10 % Р. Линдемана:
с одного трофического уровня экологической пирамиды переходит на другой, более
высокий её уровень (по "лестнице": продуцент - консумент
- редуцент) в среднем около 10 % поступившей на
предыдущий уровень экологической пирамиды энергии. Обратный поток, связанный с
потреблением веществ и продуцируемой верхним уровнем экологической пирамиды
энергии более низкими её уровнями, например, от животных к растениям, намного
слабее - не более 0,5 % от общего её потока, и потому говорить о круговороте
энергии в биоценозе не корректно. Если энергия при переходе на более высокий
уровень экологической пирамиды десятикратно теряется, то накопление ряда
веществ, в том числе токсичных и радиоактивных, в примерно такой же пропорции
увеличивается. Этот факт фиксирован в правиле биологического усиления. Н.Ф. Реймерс (1994) отмечал, что оно справедливо для всех
экосистем. В водных экосистемах накопление многих токсичных веществ, в том
числе хлорорганических пестицидов, коррелирует с
массой жиров (липидов), т. е. явно имеет энергетическую подоснову.
Правило "метаболизм и размеры особей" или правило Ю. Одума: при неизменном энергетическом потоке в пищевой сети
или цепи, более мелкие наземные организмы с высоким удельным метаболизмом
создают относительно меньшую биомассу, чем крупные.
Если измельчание особей, согласно правилу Ю. Одума,
ведет к производству относительно меньшего объема биомассы, то удельный её
выход с единицы площади в силу более полного заселения пространства
увеличивается. Никогда слоны не дадут той биомассы и продукции с единицы
площади, какую способны дать мелкие беспозвоночные. Этот эмпирический факт
называют законом удельной продуктивности.
Правило (принцип) экологического дублирования: исчезающий или уничтожаемый вид
живого в рамках одного уровня экологической пирамиды заменяет другой
функционально-ценотический, аналогичный, по схеме: мелкий сменяет крупного,
эволюционно ниже организованный более высокоорганизованного, более генетически
лабильный и мутабельный менее генетически
изменчивого.
Географическое дублирование реализуется согласно принципу (правилу)
эквивалентности В. Тишлера: в географически
удаленных, но экологически сходных биотопах идентичные биоценотические функции
выполняют систематически различные виды, занимающие эквивалентные экологические
ниши.
Дублирование - один из механизмов поддержания надежности экосистем. Это
наиболее мобильный способ их адаптации. Возможно межвидовое и внутривидовое
дублирование, а в сельском хозяйстве даже межсортовое. Общий "смысл"
остается тем же - максимально полное проведение и использование потока энергии,
стабилизация экосистемы в меняющихся условиях существования.
А.А. Еленкин сформулировал принцип подвижного равновесия: биотическое
сообщество сохраняется как единое целое вопреки регулярным колебаниям среды его
существования, но при воздействии необычных факторов структурно изменяется с
переносом "точки опоры" на другие растительные компоненты (группы
растений). Если необычные, нерегулярные факторы оказывают многолетнее
воздействие, то сообщество формирует иную структуру. Однако следует заметить,
что, как правило, сохраняются элементы дублирования в виде малочисленных в ценозе видов, которые могут быть мобилизованы в случае
новых резких изменений среды.
Принцип продукционной оптимизации Г. Реммерта:
отношение между первичной и вторичной продукцией (между продуцентами и консументами) соответствует принципу оптимизации -
"рентабельности" биопродукции. Как правило,
растения и другие продуценты дают биомассу достаточную, но не излишнюю, для
потребления всем биотическим сообществом (с эволюционно определенным запасом,
который обеспечивает надежность системы и обычно в 100 раз превышает
потребление в экосистемах суши). При относительном "перепроизводстве"
органического вещества экосистема становится "нерентабельным",
возникают предпосылки для массового размножения отдельных видов. После периода
автоколебаний отношение "популяция - потребление" уравновешивается,
экосистема стабилизируется, балансируются отношения
между трофическими уровнями.
Именно благодаря экологическому дублированию, сдвигу - в подвижном равновесии и
снижению "рентабельности" агроэкосистемы
возникают массовые размножения нежелательных в хозяйстве организмов.
Монокультура в сельском хозяйстве, однопородные и
одновозрастные леса с "точки зрения" природы весьма мало рентабельны,
неравновесны и поэтому "требуют
исправления" массовыми организмами.
Все перечисленные закономерности саморегуляции
экосистем обобщают в виде принципа стабильности: любая относительно замкнутая
биосистема с проходящим через неё потоком энергии в ходе саморегуляции
развивается в сторону устойчивого состояния. Этот принцип характерен не только
для экосистем нижнего уровня иерархии, но и для биосферы в целом.
Правило экосистемной надежности: надежность
экосистемы зависит от её энергетической эффективности в данных условиях среды и
возможностей структурно-функциональной перестройки в ответ на изменение внешних
воздействий (материала для дублирования, межвидового и внутривидового,
поддержания продукционной "рентабельности" и т. п.).
Принцип биоценотической прерывности: виды формируют экологически определенные
системные совокупности - сообщества и биоценозы, - отличающиеся от соседних,
хотя и сравнительно постепенно в них переходящие.
Закон действия факторов А. Тинемана: состав
сообщества и биоценоза по видам и численности особей в них определяется тем
фактором среды, который оказывается в пессимуме (наиболее неблагоприятен) для
данного сообщества.
А. Тинеман сформулировал широко известные
биоценотические принципы: принцип (правило) разнообразия условий биотопа - чем
разнообразнее условия жизни в рамках биотопа, тем больше число видов в
заселяющем его биоценозе; и принцип отклонения условий существования от нормы -
чем больше отклонения условий существования от оптимума (нормы) в пределах
биотопа, тем беднее видами становится заселяющий его биоценоз и тем
относительно больше особей имеет каждый присутствующий вид. Число особей внутри
вида и число видов в ценозе обратно пропорциональны.
Исходя из биоценотических принципов А. Тинемана, Н.Ф.
Реймерс (1994) заключил, что в оптимальных условиях внутрибиотическое межвидовое напряжение (конкуренция между
видами) усиливается: каждый вид действует на другой как лимитирующий (или
управляющий) фактор. В неблагоприятных условиях или условиях экологической
монотонности число видов мало, и межвидовая конкуренция снижается.
Первый фитоценологический принцип П. Жаккара: видовое
богатство территории пропорционально разнообразию экологических условий.
Второй фитоценологический принцип П. Жаккара:
экологическое разнообразие возрастает с увеличением рассматриваемого
пространства и падает с возрастанием однообразия условий.
Правило приспособления (специализации) к экстремальным условиям Р. Крогеруса: в биотопах с экстремальными условиями
складываются биоценозы из строго специализированных (стенотопных) видов с
относительно большим числом особей.
Принцип плотной упаковки Р. Макартура: виды,
объединенные в сообщество (биоценоз, экосистему), используют все возможности
для существования, предоставляемые средой, с минимальной (но не нулевой!)
конкуренцией между собой и максимальной биологической продуктивностью в
условиях данного конкретного места обитания (биотопа); при этом пространство заполняется
с наибольшей плотностью.
Правило обязательности заполнения экологических ниш: пустующая экологическая
ниша всегда и обязательно бывает естественно заполнена.
Число экологических ниш возрастает при пространственном переходе одного
биоценоза в другой, поскольку это случается на границах биотопов, обладающих
свойствами стыкующихся ценозов, нередко дающих не
простую сумму, а новое системное качество. В таких маргинальных полосах
возникают сгущения видов и особей, так называемый краевой эффект, или эффект
опушки. Правило экотона или краевого эффекта состоит
в том, что на стыках биоценозов увеличивается число видов и особей в них. На
таких же контактах сообществ подобное увеличение, как правило, не наблюдается,
что еще раз подчеркивает разницу между этими понятиями.
Первый биоценотический постулат В. Тишлера:
биоценотическая система возникает из сочетания готовых частей, а не в
результате дифференциации внутренних зачатков.
Второй биоценотический постулат В. Тишлера утверждает
заменяемость частей ценоза
и необязательную связь этих частей со всей системой, но лишь с общей основой их
и её существования - с биотопом.
Третий биоценотический постулат В. Тишлера: биоценоз
как система поддерживается взаимной компенсацией сил благодаря антагонизму, а
не координации составляющих его частей.
Четвертый биоценотический постулат В. Тишлера:
существует лишь количественная, а не качественная способность к регуляции
выпадающих компонентов биоценоза.
Пятый биоценотический постулат В. Тишлера:
ограничения функционирования системы обусловлены внешними условиями, а не
внутренними предпосылками.
Правила взаимоприспособленности организмов в
биоценозе К. Мебиуса - Г. Ф. Морозова: виды в биоценозе приспособлены друг к
другу настолько, что их сообщество составляет внутренне противоречивое, но
единое и взаимно увязанное системное целое.
Биоценотическое правило В.С. Ивлиева: межвидовое
напряжение гораздо сильнее, чем во внутривидовых отношениях (между особями
внутри вида).
Правило относительной внутренней непротиворечивости: в естественных экосистемах
(и прежде всего в их биотических компонентах, составляющих биоценозы)
деятельность входящих в них видов направлена на поддержание этих систем как
среды собственного обитания.
Принцип экологического высвобождения: при удалении из данного сообщества
(биоценоза) части популяции одного вида другой, близкий к нему вид,
увеличивается в числе.
Принцип экологической компрессии: увеличение числа видов лимитирует возможность
проникновения их в большое число местообитаний одновременно снижает число
особей в каждом из видов. И наоборот, множественность видов ограничена
способностью их внедрения в новые местообитания.
Закон периодического цикла: процесс уничтожения жертвы хищником нередко
приводит к периодическим колебаниям численности популяций обоих видов,
зависящим только от скорости роста популяций хищника и жертвы и от исходного
соотношения их численностей.
Закон сохранения средних величин: средняя численность популяции для каждого
вида постоянна независимо от начального уровня при условии, что специфические
скорости увеличения численности популяций, а также эффективность хищничества
постоянны.
Закон нарушения средних величин: при аналогичном нарушении популяций хищника и
жертвы средняя численность популяции жертвы растет, а популяции хищника падает.
Теорема Л. Р. Гинзбурга: успешность сосуществования двух и более видов (их
популяций) определяется не их начальной численностью, а относительными
коэффициентами конкуренции.
Правило монокультуры Ю. Одума: эксплуатируемые для
нужд человека системы, представленные одним видом, равно как и системы
монокультур, неустойчивы по своей природе.
Пагубность монокультур "учтена" природой. Более того, ею выработана
стратегия сохранения гомеостаза на основе поддержания разнообразия и взаимозависимости
членов биоценоза. Один из путей реализации такой стратегии отражает принцип
сопряженной эволюции или теория сопряженной эволюции П. Эрлиха и П. Равена, называемая также принципом коэволюции:
случайное функциональное изменение жертв (потребляемого растения) ведет к
закономерному изменению свойств хищников (потребителей), что в свою очередь
стимулирует разнообразие как первых, так и вторых.
Правило управляющего значения консументов,
растительноядные организмы, снижая массу видов-доминантов,
дают простор для видов с менее выраженными доминантными свойствами.
Правило (принцип) экологической индивидуальности видов или принцип континуума
Л.Г. Раменского - Г.А. Глизона: каждый вид
пространственно распределен в соответствии со своими генетическими,
физиологическими и другими биологическими особенностями и глубоко специфично
относится к факторам среды, в том числе к другим видам. Широкое перекрытие
экологических амплитуд и рассосредоточенность центров
распределения популяций вдоль градиента среды приводят к плавному переходу
одного сообщества в другое, поэтому не образуется строго фиксированных
сообществ.
Правило колебания границ ареала: многие виды не имеют четко отграниченной
постоянной области распространения и временами либо исчезают на границе ареала,
либо достигают исчезающе малой численности. В. Тишлер сформулировал принцип воздействия факторов, согласно
которому границы, размер и характер ареала вида или местообитания популяции
обусловлены их биологическими особенностями, и наоборот, биологические
особенности вида или популяции могут указывать (быть индикаторами) места, где
их можно найти.
Правило географической изменчивости кружева ареала: на севере вид приурочен к
приречным, низменным участкам, затем к югу заселяет более широкие пространства,
в горах обитает лишь в локальных участках, а на южном пределе распространения
вновь приурочен к приречным более влажным и прохладным местообитаниям. У
каждого широко распространенного вида характер изменения кружева ареала
специфичен, но смены происходят закономерно.
Наиболее общим объяснением причин формирования границ ареала вида служит
правило ограничивающих факторов: факторы среды, наиболее удаляющиеся от
оптимума экологических потребностей вида, лимитируют возможности его
существования в данных условиях.
Правило викариата Д. Джордана: ареалы
близкородственных форм организмов (правило установлено для животных) обычно
занимают смежные территории и существенно не перекрываются; родственные формы,
как правило, викарируют, т. е. географически замещают
друг друга. Согласно правилу представительства рода одним видом А. Монара, в однородных условиях и на ограниченной территории
систематический (таксономический) род, как правило, представлен только одним
видом.
Принцип конкурентного исключения (закон или теорема) Г. Ф. Гаузе:
два вида с близкими экологическими требованиями длительное время не могут
занимать одну экологическую нишу и, как правило, входить в одну экосистему.
Следовательно, одновременно происходит и географическое замещение видов, что
констатирует принцип видо-родового представительства
И. Иллиеса: поскольку два близкородственных вида не
могут занимать одинаковые экологические ниши в одном биотопе, и,
соответственно, в биоценозе, богатые видами роды представлены в экосистеме
единственным своим представителем.
Принцип сосуществования, или парадокс Дж. Хатчинсона: два вида в порядке
исключения могут сосуществовать в одной экологической нише, и если близкие виды
вынуждены использовать одни и те же ресурсы, то естественный отбор может
благоприятствовать их сосуществованию, и они способны эволюционировать в
близком направлении.
Правило географического оптимума: условия обитания вида наиболее оптимальны для
него в центре ареала.
Правило К. Бергмана: у теплокровных животных, подверженных географической
изменчивости, размеры тела особей статистически (в среднем) больше у популяций,
живущих в более холодных частях ареала вида.
Как бы дополнением к правилу Бергмана служит правило Р. Гессе: особи популяций
в северных районах обладают относительно большей массой сердца (при её
сравнении с массой тела), чем особи популяций, живущих в южных районах или в
более теплых местообитаниях.
Закон покровов (покрытия) тела: плотность (густота и терморегулирующая
способность) покровов тела млекопитающих и птиц достигает максимума в холодных
и засушливых областях. Уточнение к закону покровов (покрытия) тела служит
правило густоты мехового покрова: у млекопитающих холодных местообитаний мех
как правило гуще, чем у живущих в теплых географических зонах и поясах. Часто
это правило справедливо и для млекопитающих сухого климата, где густой мех
предохраняет от излишнего испарения воды с поверхности тела, а также от
перегревания.
Правило Д. Аллена: увеличение выступающих частей тела
одного вида теплокровных животных (конечностей, хвоста, ушей и т. п.) при
продвижении с севера на юг.
Для рыб существует особое правило числа позвонков А. Жордана,
согласно которому в водоемах с повышенной соленостью и более низкими
температурами в хвостовой части тела возрастает число позвонков. Правило Жордана не имеет отношения к правилу Аллена,
а связано с приспособлением к движению в более плотной среде. Подобного рода
закономерностей много у почвенных и глубоководных организмов - их удлиненность,
часто червеобразность.
Правило, или закон К. Глогера: виды животных северных
и влажных географических зон имеют более интенсивную окраску, чем обитатели
южных сухих регионов.
Принцип основателя: единичные особи-основатели новой (изолированной) колонии
или популяции несут в себе лишь часть общей генетической информации, присущей
виду. Нехватка потенциала наследственной изменчивости может лежать в основе
правила островного измельчания: популяции видов животных, обитающие на
островах, как правило, образуются более мелкими особями, чем материковые,
живущие в аналогичных условиях.
Правило (Уоллеса) увеличения видового разнообразия по мере продвижения с севера
на юг: в тропиках значительно больше абсолютное число видов, чем на Севере, и в
составе южных сообществ их также намного больше.
Принцип эколого-географического максимума (стабильности числа) видов: число
видов в составе географических зон и их экосистем относительно постоянно и
регулируется вещественно-энергетическими процессами. Это число всегда
естественно стремится к необходимому и достаточному максимуму.
Принцип взаимоисключаемости биотических комплексов:
две генетически сходные группы организмов разного происхождения, обладающие
близкими (одинаковыми) требованиями к окружающей их среде, взаимоисключают
друг друга. Совершенно очевидно, что этот принцип следует учитывать при
попытках акклиматизировать виды, относящиеся к различным, но близким
биотическим комплексам.
Правило предварения, или правило постоянства местообитания (В. В. Алехина - И.
Вальтера): плакорный вид или плакорный
биоценоз предваряется на юге или на севере в соответствующих условиях
местообитания, что связано с меньшей или большей инсоляцией склонов различной
экспозиции.
Образуются участки незональных ценозов,
соответствующие более северным или южным зонам (например, степные участки в
тайге). Обычно в этих маргинальных образованиях ценозы
беднее видами.
Видимо, именно обеднённость ценозов
в зонах контакта климатических и экологических зон формирует закономерность,
известную как закон минимума видов или эффект А. Ремане:
минимум морских и пресноводных видов животных наблюдается в солоноватых водах.
В данном случае возникает не географическая, а экологическая, функциональная маргинальность.
Принцип территориальной общности физико-географических единиц (районов, провинций
и т. п.): островные экосистемы, возникшие в результате дробления изначальных
при их нарушении человеком, составляют целостные природные образования
(надсистемы) более высокого иерархического уровня, и, соответственно,
функционально целостные физико-географические территориальные единицы
образуются лишь в том случае, если эти экосистемы расположены не как
изолированные, удаленные друг от друга участки, а как относительно компактная,
хотя и мозаичная совокупность.
Закон обеднения разнородного живого вещества (биоты)
в островных его сгущениях Г.Ф. Хильми (или принцип
организационной деградации, или закон растворения системы в чуждой среде):
"индивидуальная система, работающая в ... среде с уровнем организации
более низким, чем уровень самой системы обречена: постепенно теряя структуру,
система через некоторое время растворится в окружающей... среде".
Правило К. Дарлингтона: уменьшение площади острова в
10 раз сокращает число живущих на нем видов вдвое.
Теория биполярности Л.С. Берга: жизнь Арктики и Антарктики аналогична по
происхождению и имеет истоки в третичном времени.
Закон преломления космических воздействий: космические факторы, оказывая
воздействие на биосферу и особенно её подразделения, подвергаются изменению со
стороны экосферы планеты и потому по силе и времени
проявления могут быть ослаблены и сдвинуты или могут даже полностью утерять
свой эффект.
Биосферу, как и любую другую систему, формируют не только и не столько внешние
факторы, как внутренние закономерности. И они затем взаимодействуют с внешней
средой. Одним из важнейших свойств биосферы - слоя взаимодействия живого и
неживого - является закон биогенной миграции атомов, открытый В. И. Вернадским:
"миграция химических элементов на земной поверхности и в биосфере в целом
осуществляется или при непосредственном участии живого вещества (биогенная
миграция) или же она протекает в среде, геохимические особенности которой
обусловлены живым веществом, как тем, которое в настоящее время населяет
биосферу, так и тем, которое действовало на Земле в течение всей человеческой
истории".
Правило незамкнутости биотических круговоротов:
степень замкнутости глобальных биогеохимических круговоротов довольно высока
(для различных элементов и веществ она неодинакова), но она не абсолютна.
Закон константности количества живого вещества В.И. Вернадского: количество
живого вещества биосферы (для данного геологического периода) есть константа.
Значит, для живого вещества планеты неизбежна количественная стабильность. Она
характерна и для числа видов.
Закон сохранения структуры биосферы (информационной и соматической) или первый
закон экодинамики Ю. Голдсмита:
в живой природе наблюдается постоянное сохранение информационной и соматической
структуры, хотя она и несколько меняется с ходом эволюции.
Закон стремления к климаксу - второй закон экодинамики
Ю. Голдсмита: для сохранения структуры биосферы живое
стремится к достижению состояния зрелости или экологического равновесия.
Биосфера - высший уровень иерархии экосистем нашей планеты, и естественно, её
законы функционирования аналогово справедливы и для
ниже расположенных уровней в этой иерархии, хотя имеется и специфика - биосфера
более закрытая система, чем её подразделения.
Закон экологического мутуализма (принцип экологического порядка) - третий закон
экодинамики Ю. Голдсмита:
эволюционно возникшие в биосфере живые элементы различного геологического
возраста и первоначального географического происхождения сложно переплетены.
Системный мутуализм, т. е. взаимопомощь в рамках экологического порядка, подчеркивается
законом упорядоченности заполнения пространства и пространственно-временной
определенности: заполнение пространства внутри природной системы в силу
взаимодействия между её подсистемами упорядочено таким образом, что позволяет
реализоваться гомеостатическим свойствам системы с минимальными противоречиями
между частями внутри неё. Из этого закона следует невозможность длительного
существования "ненужных" природе случайностей, в том числе и чуждых
ей созданий человека. Нарушение природного порядка обходится людям
дополнительными вложениями средств и сил. Это нарушение имеет строгие, к
сожалению, недостаточно еще изученные пределы, за которыми мутуализм сменяется
антагонизмом, порядок нарушается, а потому ухудшаются и динамические качества
всей биосферы.
В блок правил мутуалистического системного порядка в
биосфере входит принцип системной дополнительности:
подсистемы одной природной системы в своем развитии обеспечивают предпосылку
для успешного развития и саморегуляции других
подсистем, входящих в ту же систему.
Закон самоконтроля и саморегуляции живого - четвертый
закон экодинамики Ю. Голдсмита:
живые системы и системы под управляющим воздействием живого способны к
самоконтролю и саморегулированию в процессе их адаптации к изменениям в
окружающей среде.
Правило автоматического поддержания глобальной среды обитания: живое вещество в
ходе саморегуляции и взаимодействия с абиотическими
факторами автодинамически поддерживает среду жизни,
пригодную для её развития.
Принцип Реди: живое происходит только от живого,
между живым и неживым веществом существует непроходимая граница, хотя и имеется
постоянное взаимодействие.
Экоисторический принцип: геологические процессы и
явления не оставались неизменными в течение эволюционного времени, в силу чего
процессы далекого прошлого нельзя полностью отождествлять с современными.
Принцип максимума эффекта внешней работы, закон саморазвития биосистем или
закон исторического развития биологических систем Э. Бауэра: развитие
биологических систем есть результат увеличения их внешней работы - воздействия
этих систем на окружающую среду.
Принцип (правило) катастрофического толчка: природная или
природно-антропогенная катастрофа всегда приводит к существенным эволюционным
перестройкам, которые относительно прогрессивны для биосферы, так как
адаптируют её системы к новым условиям среды. А поскольку существуют ускорения
и замедления эволюции, действует и принцип прерывности и непрерывности развития
биосферы: процесс медленного эволюционного изменения организмов закономерно
прерывается фазами бурного развития и вымирания практически без переходных
(палеонтологических) форм.
Правило константности числа видов в ходе стационарной эволюции биосферы: число
нарождающихся видов в среднем равно числу вымерших, и общее видовое разнообразие
в биосфере есть константа.
Правило усиления интеграции биологических систем И. И. Шмальгаузена:
биологические системы в процессе эволюции становятся всё более
интегрированными, с всё более развитыми регуляторными механизмами,
обеспечивающими такую интеграцию.
Закон бумеранга или закон обратной связи взаимодействия человек - биосфера П. Дансеро, иначе - четвертый закон Б. Коммонера "ничто
не даётся даром": глобальная экосистема представляет собой единое целое, в
рамках которого ничего не может быть выиграно или потеряно и которое не может
являться объектом всеобщего улучшения: всё, что было извлечено из неё
человеческим трудом, должно быть возвращено.
Закон незаменимости биосферы: сокращение естественной биоты
в объёме, превышающем пороговое значение, лишает устойчивости биосферу, которая
не может быть восстановлена за счёт никаких прогрессивных технологий.
Незаменимая биосфера до поры до времени работала в рамках принципа Ле Шателье-Брауна, что для этой
фазы эволюции сформулировано в виде закона обратимости биосферы П. Дансеро: биосфера стремится к восстановлению экологического
равновесия тем сильнее, чем больше давление на нее: это стремление продолжается
до достижения экосистемами климаксовых фаз развития.
Фактически это повторение закона стремления к климаксу в приложении к
взаимоотношениям типа природа - человек. Однако П. Дансеро
тогда же сформулировал закон необратимости взаимодействия человек - биосфера: возобновимые природные ресурсы делаются невозобновимыми
в случае глубокого изменения среды, значительной переэксплуатации,
доходящей до поголовного уничтожения или крайнего истощения, а потому
превышения возможностей их восстановления. Именно такова фаза развития системы
взаимоотношений человек - природа в наши дни.
Правило меры преобразования природных систем: в ходе эксплуатации природных
систем нельзя переходить некоторые пределы, позволяющие этим системам сохранять
свойство самоподдержания (самоорганизации и саморегуляции) и обычно ограниченные заметным изменением
природных систем трех сопряженных уровней иерархии (надсистем рассматриваемой
системы).
Из правила меры преобразования природных систем следует ряд выводов:
1. Единица (возобновимого) ресурса может быть
получена лишь в некоторый, определяемый скоростью функционирования системы (и
их иерархии), отрезок времени. В течение этого отрезка нельзя переходить рубежи
ограничений, диктуемых законами экологии;
2. Перешагнуть через фазу последовательного развития природной системы с
участием живого, как правило, невозможно;
3. Рационально проведение хозяйственных мероприятий лишь в рамках некоторых
оптимальных размеров, выход за которые в меньшую и большую стороны снижает их
хозяйственную эффективность;
4. Преобразовательная деятельность не должна выводить природные системы из
состояния равновесия путем избытка какого-то из средообразующих
компонентов, т. е., если это необходимо, требуется достаточная компенсация в
виде относительно непреобразованных природных систем,
например, оптимальная лесистость и т. п.;
5. Преобразование природы (если оно не восстановительное, "мягкое")
даёт локальный или региональный выигрыш за счет ухудшения каких-то показателей
в смежных местностях или в биосфере в целом;
6. Хозяйственное воздействие затрагивает не только ту систему, на которую оно
направлено, но и на её надсистемы. которые, согласно принципу Ле Шателье - Брауна,
"стремятся" нивелировать производимые изменения. В связи с этим
расходы на преобразование природы никогда не ограничиваются лишь затратами на
непосредственно планируемые воздействия;
7. Природные цепные реакции никогда не ограничиваются изменением вещества и
энергии, но затрагивают динамические качества систем природы;
8. Вторичное постепенно сложившееся экологическое равновесие, как правило,
устойчивее, чем первичное, но потенциальный "запас преобразований"
(т. е. будущих их возможностей) при этом сокращается;
9. Несоответствие "целей" естественно-системной регуляции в природе и
целей хозяйства может приводить к деструкции природного образования (т. е. силы
природы и хозяйственных преобразований при большей величине последних в ходе
противоборства сначала "гасят" друг друга, а затем природная
составляющая начинает разрушаться);
10. Технические системы воздействия в конечном итоге (в длительном интервале
времени) всегда менее хозяйственно эффективны, чем направляемые естественные.
Принцип естественности или правило старого автомобиля: со временем
эколого-социально-экономическая эффективность технических устройств,
обеспечивающих "жесткое" управление природными системами и
процессами, снижается, а экономические (материальные, трудовые, денежные)
расходы на их поддержание возрастают.
Закон убывающей отдачи А. Тюрго - Т. Мальтуса: повышение удельного вложения
энергии в агроэкосистему не даёт адекватного
пропорционального увеличения её продуктивности (урожайности). Н.Ф. Реймерс (1994) отмечал, что длительные споры вокруг этого
закона окончились тем, что он стал азбучной истиной агроэкологии.
Закон или правило демографического насыщения: в глобальной или регионально
изолированной совокупности количество народонаселения всегда соответствует
максимальной возможности поддержания его жизнедеятельности, включая все аспекты
сложившихся потребностей человека. Однако человечество создает давление на
среду не столько биологически, сколько техногенно. Вместо
демографического насыщения как такового возникает насыщение разрушительной
техникой, поэтому Н.Ф. Реймерс (1994) предложил
называть это правило принципом технико-социально-экономического насыщения.
Правило ускорения исторического развития: чем стремительнее под воздействием
антропогенных причин изменяется среда обитания человека и условия ведения им
хозяйства, тем скорее по принципу обратной связи происходит перемена в
социально-экологических свойствах человека, экономическом и техническом
развитии общества (знак процесса может быть положительным и отрицательным).
Правило социально-экологического равновесия: общество развивается до тех пор и
постольку, поскольку сохраняет равновесие между своим давлением на среду и
восстановлением этой среды - природно-естественным и искусственным.
Закон исторической (социально-экологической) необратимости: процесс развития
человечества как целого не может идти от более поздних фаз к начальным, т. е.
общественно-экономические формации, определенным образом взаимодействующие с
природной средой и естественными ресурсами, не могут сменяться в обратном
порядке.
Закон ограниченности (исчерпаемости) природных
ресурсов: все природные ресурсы и естественные условия Земли конечны. Эта
возникает либо в силу прямой исчерпаемости, либо в
результате возмущения среды обитания, делающейся непригодной для сложившегося
хозяйства и жизни человека.
Правило интегрального ресурса: конкурирующие в сфере использования конкретных
природных систем отрасли хозяйства неминуемо наносят ущерб друг другу тем
сильнее, чем значительнее они изменяют совместно эксплуатируемый экологический
компонент или всю экосистему в целом.
Правило (неизбежных) цепных реакций "жесткого" управления природой:
"жесткое", как правило, техническое управление природными процессами
чревато цепными природными реакциями, значительная часть которых оказывается
экологически, социально и экономически неприемлемыми в длительном интервале
времени.
Правило "мягкого" управления природой Н.Ф. Реймерса
(1994): "как дипломатические переговоры желательней войны, так и
"мягкое" управление природными процессами, системное направление их в
необходимое русло с учётом законов природы в конечном итоге эффективнее грубых
техногенных вмешательств. Такое управление построено на инициации полезных
природных цепных реакций, в том числе процессов восстановления, возобновления
ресурсов".
Увеличение наукоёмкости и энергоемкости общественного
производства приводит в действие два позитивных процесса, формулируемых в виде
закона снижения природоёмкости готовой продукции:
удельное содержание природного вещества в усредненной единице общественного
продукта исторически неуклонно снижается и закона увеличения темпов оборота
вовлекаемых природных ресурсов: в историческом процессе развития мирового
хозяйства быстрота оборачиваемости вовлеченных природных ресурсов (вторичных,
третичных и так далее) непрерывно возрастает на фоне относительного уменьшения
объёмов их вовлечения в общественное производство (относительно роста темпов
самого производства).
Наиболее общим принципом или правилом охраны природы считают закон шагреневой,
кожи: глобальный исходный природно-ресурсный потенциал в ходе истерического
развития непрерывно истощается, что требует от человечества научно-технического
совершенствования, направленного на более широкое и глубокое использование
этого потенциала.
Возможно, из закона шагреневой кожи следует выделить правило "экологичное - экономично" как один из основополагающих
принципов охраны природы и среды жизни: сохранение ресурсов в конечном итоге
выгодно в социальном и экономическом отношениях, а к тому же оставляет время
для кардинального решения демографических проблем.
Закон неустранимости отходов и/или побочных воздействий производства
(хозяйства): в любом хозяйственном цикле образующиеся отходы и возникающие
побочные эффекты неустранимы, они могут быть лишь переведены из одной
физико-химической формы в другую или перемещены в пространстве. Если бы была
реальная возможность избавиться от отходов, это было бы нарушением законов
сохранения массы и энергии. Суммарное количество отходов в виде вещества,
энергии и побочных эффектов (изменения динамических качеств природных систем -
их устойчивости, надежности и т. п.) фактически постоянно: в производственных
циклах меняется лишь место их возникновения, время образования и
физико-химическая или биологическая форма. Поэтому закон неустранимости отходов
может быть дополнен законом постоянства количества отходов в технологических
цепях.
Принцип разумной достаточности и допустимого риска: расширение любых действий
человека не должно приводить к социально-экономическим и экологическим
катастрофам, подрывающим саму возможность существования людей.
Принцип неполноты информации (принцип неопределенности): информация при
проведении акций по преобразованию и вообще любому изменению природы всегда
недостаточна для априорного суждения о всех возможных результатах таких
действий, особенно в далекой перспективе, когда разовьются все природные цепные
реакции.
Принцип инстинктивного отрицания - признания: факты и закономерности, в глубине
подсознания концептуально отрицаемые составителем машинной программы,
неосознанно исключаются им из модели, а фактам, признаваемым верными,
инстинктивно придается больший вес, чем они имеют в реальности.
Принцип удаленности события: явления, отдаленные во времени и в пространстве,
психологически кажутся менее существенными.
Принцип обманчивого благополучия или эйфории первых успехов связан с излишней
поспешностью суждений - первые успехи или неудачи в природопользовании могут быть
кратковременными: успех мероприятия по преобразованию природы или управлению ею
объективно может быть оценен лишь после выяснения хода и результатов природных
цепных реакций в пределах естественного природного цикла (от немногих лет до их
десятков) и лишь после возникновения нового уровня экологического баланса.
Закон увеличения наукоёмкости общественного развития:
человечество стало приближаться к ноосфере.
Закон ноосферы В.И. Вернадского (1944): биосфера неизбежно превратится в
ноосферу, т. е. в сферу, где разум человека будет играть доминирующую роль в
развитии системы человек - природа. Хаотичное саморазвитие, основанное на
процессах естественной саморегуляции, должно быть
заменено разумной стратегией, базирующейся на прогнозно-плановых началах, регулировании
процессов естественного развития. Это управление, безусловно, может быть лишь
"мягким". В нём можно только следовать законам природы и развития
общества. Лишь благо и заинтересованное понимание, а не насилие и волюнтаризм
могут быть в основе формирования ноосферы.
Обсуждая в научной литературе различные аспекты устойчивости экосистем,
исследователи в той или иной форме обращаются к вопросу о роли почв в этом
процессе. Именно устойчивость биогеоценотических функций почв и почвенного
покрова рассматривают как основное условие устойчивости биогеоценозов и
биосферы в целом (391).
При анализе возможностей восстановления нарушенных экосистем обращают внимание
на тот факт, что даже при очень сильных нарушениях экосистем сохраняется
возможность их восстановления, если сохранился почвенный покров (88). Поэтому в
современных экологических условиях почву нужно рассматривать не столько как
результат почвообразовательного процесса, а как полифункциональную
энергетически открытую самоорганизующуюся природную систему, обеспечивающую
устойчивый циклический характер воспроизводства жизни на планете.
Характеристика многочисленных экологических функций почв дана в работах В.А.
Ковды (207, 208); Розанова (374); Г.В.
Добровольского, Е.Д. Никитина (294) и других
учёных. Прямым доказательством важности и устойчивости эколого-биосферных
функций почв служат следующие факты ( 166):
- биомасса суши, создающаяся системой почвы-растения-животные, составляет 99,8
% всей биомассы Земли, хотя площадь продуктивных почв в несколько раз меньше
площади гидросферы (океана);
- видовое разнообразие растений на суше составляет 92 %, а животных - 93 %
всего разнообразия жизни на Земле. В мире микроорганизмов именно почва является
основной средой их обитания и видового разнообразия. Количество микроорганизмов
в 1 г почвы достигает миллиардов;
- биопродуктивность суши, т. е. почв, во много раз
выше продуктивности океана. Отсюда следует вывод: именно тончайший почвенный
покров на поверхности Земли явился той пленкой сгущения жизни по В.И.
Вернадскому, которая обеспечивает её разнообразие и эволюцию. По всем
палеонтологическим и историко-геологическим данным резкое возрастание скорости
эволюции и разнообразия жизни на Земле связано с выходом живых существ из
океана на сушу и образованием "настоящих" почв. Это время относится,
как известно, к девону-карбону, т. е. 350-400 млн. лет назад.
Жизнь выбрала для себя наиболее надежную, т. е. наиболее благоприятную и
устойчивую сферу обитания - педосферу, что является
доказательством неотъемлемости почвы как компонента наземных экосистем и
необходимости учёта её функций при определении их устойчивости. В 70-е годы
наряду с обсуждением понятия устойчивости наземных экосистем начал интенсивно
разрабатываться вопрос об устойчивости самих почв. Г.В. Добровольский (166) определил
устойчивость почв как "способность сохранять и восстанавливать свою
структуру и функционирование при изменяющихся внешних условиях". В понятии
устойчивости почв он различает два аспекта:
- способность сопротивляться воздействию внешнего фактора (соответствует
понятию "буферность"), означающему
сопротивление механическому давлению, размыванию, подкислению и т. п.;
- способность восстанавливать структуру и функции после их нарушения под воздействием
внешних факторов, т. е. способность регенерировать эти свойства, например,
восстанавливать после
деградации структуру, аккумуляцию гумуса и плодородие
в целом.
Устойчивость почв - понятие многосмысловое,
комплексное и охватывает как частные виды устойчивости - физическую,
химическую, биологическую, так и общую - интегральную.
В оценке, нормировании и отображении разных видов устойчивости почв сделано уже
немало. Разработаны критерии и группировки: геохимической устойчивости разных
почв к техногенезу, противоэрозионной, биологической
и интегральной устойчивости почв.
Особенно важны разработки принципов оценки состояния и устойчивости почв,
основанные на методах нормирования отдельных показателей с последующим их
суммированием по балльной системе (393; 340; 88).
Г.В. Добровольский (166)
отмечает, что ещё не разработаны параметры экологической оценки почв с учётом
их региональных особенностей, типов использования, видов культур. Пригодны для
этого интегральные показатели, например, почвенно-экологический индекс (ПЭИ),
методика расчёта которого предложена в Почвенном институте им. В.В. Докучаева и
рекомендована Академией сельскохозяйственных наук для использования в
производстве (262;
451).
Традиционным комплексным показателем оценки почв, как фундаментального
компонента агроэкосистем, является плодородие. При
его описании и оценке в специальной литературе чётко прослеживается стремление
к объединению косных и живых составляющих экосистем с социальной сферой (322).