Содержание
1. Функции симпатической
нервной системы.. 3
2. Общая характеристика и теории боли. 6
3. Функции ретикулярной формации. 11
Список использованной литературы.. 15
1. Функции симпатической нервной
системы
Симпатическая нервная система - это часть нервной системы,
связывающая (инервирующая) все внешние и внутренние
органы от кожи до сердца. Она представлена особыми нервными «веточками», связь
между которыми осуществляется с помощью гормона норадреналина. Поэтому выброс
катехоламинов вторично активизирует симпатическую нервную систему.
Основное назначение симпатической нервной системы - это
мобилизация всех наличных энергетических, пластических и прочих резервов на
борьбу с неблапгоприятным фактором. Активизация
симпатической нервной системы резко стимулирует окислительно-восстановительные
реакции, распад гликогеновых запасов и утилизацию
жиров. При экстремальных физических нагрузках именно симпатическая нервная
система обеспечивает энергию, необходимую для мышечного сокращения. Если мышца
даже до предела утомлена, то электрическая стимуляция симпатических веточек,
иннервирующих мышцу, восстанавливает ее работоспособность. Именно симпатический
отдел вегетативной нервной системы обеспечивает утилизацию молочной кислоты в
печени, что позволяет организму бороться с утомлением.
Симпа- R=0,64
тическая р=0,008
актив-
ность
сердца
Инсулинорезинстентность, равновесный
уровень
глюкозы, плазмы
Рис. 1 Инсулинорезинстентность и активность
симпатической нервной системы
Помимо повышения энергетического потенциала как такового,
симпатическая нервная система перераспределяет энергетические ресурсы между
различными органами. К примеру, при необходимости происходит перераспределение
крови между условным «ядром» и «оболочкой» тела. Сосуды головного мозга,
сердца, печени и поек расширяются, в результате чего кровь притекает к более
жизненно важным органам за счет оттока от менее важных. Организм как бы готов
временно пожертвовать нормальным функционированием части своих внутренних
органов, чтобы сохранить самое главное - жизнь. Это один из
ключевых приспособительных механизмов, закрепившийся в процессе эволюции
человека.
Центральная часть симпатического отдела представлена
симпатическими ядрами боковых рогов спинного мозга от первых грудных до
поясничных сегментов. Аксоны этих ядер выходят из спинного мозга в виде белых
соединительных ветвей (преганглионарные волокна) и
вступают в узлы симпатического ствола. Скорость проведения возбуждения в этих
аксонах 1-20 м/с.
Периферическая часть образована чувствительными и
эфферентными нейронами симпатических ганглиев. Околопозвоночные ганглии
располагаются по обе стороны позвоночника от основания черепа до крестца в виде
цепочек, которые называются правым и левым симпатическим стволом. Со спиномозговыми нервами узлы соединены белыми и серыми
ветвями. По белой ветви в узел входит преганглионарное
волокно, которое может переключаться на эффекторный ганглионарный нейрон. Часть постганглионарных
волокон по серой соединительной ветви вновь возвращается в спиномозговой
нерв и далее следует в его составе без перерыва к эффекторному
органу. Другие волокна собираются в отдельные веточки (чревные нервы) и
направляются к органам грудной и брюшной полости, таза или к предпозвоночным узлам, а затем к исполнительным органам. Постганглионарные волокна большей частью лишены миелиновой
оболочки, поэтому имеют сероватую окраску. В ганглиях симпатического ствола
прерывается только часть преганглионарных волокон,
остальные же проходят их транзитом и переключаются на эффекторный
нейрон в превертебральных ганглиях.
Наиболее крупными превертебральными
ганглиями являются: солнечное сплетение (образованное чревным и краниальным брызжеечным узлами) и каудальный брыжеечный узел. От клеток
этих узлов начинаются постганглионарные симпатические
волокна, иннервирующие почти все органы брюшной полости. Они образуют
многочисленные нервные стволы, которые, направляясь к органам брюшной полости,
образуют ряд сплетений: желудочное, печеночное и др[1].
Эффекторами, которые снабжаются постганглионарными
симпатическими волокнами, являются гладкие мышцы всех органов (сосудов,
волосяных луковиц, зрачка, легких, органов пищеварения, выделения, потовые,
сальные и пищеварительные железы), а также клетки печени и жировой клетчатки.
Симпатическая нервная система помимо эфферентных путей имеет
собственные чувствительные пути. По локализации клеточных тел, ходу и длине
отростков они делятся на две группы. Первая группа периферических афферентных
нейронов включает клетки, тела которых локализуются в предпозвоночных
симпатических ганглиях. Один из длинных отростков направляется на периферию,
второй - в сторону спинного мозга, вступая в него в составе дорсальных
корешков. Вторая группа характеризуется тем, что длинный отросток этих
чувствительных клеток идет к рабочему органу, а короткие распределяются в самом
ганглии, контактируя с вставочными нейронами и через них с эфферентными
нейронами, образуя морфологическую основу для вегетативного периферического ганглионарного рефлекса[2].
2. Общая
характеристика и теории боли
Боль
изначально является жизненно важным защитным биологическим феноменом, в
нормальных условиях играющим роль основного физиологического механизма защиты.
Она мобилизует все необходимые для выживания организма функциональные системы,
позволяющие преодолеть вредоносные воздействия, спровоцировавшие боль, или
избежать их. Около 90% всех заболеваний связано с болью. По данным разных
исследователей, от 7 до 64% населения периодически испытывают чувство боли, а
от 7,6 до 45% страдают рецидивирующей или хронической болью.
Терапия
болевых синдромов предусматривает определение и устранение источника или
причины, вызвавшей боль, определение степени вовлечения различных отделов
нервной системы в формирование болевого ощущения и снятие или подавление самой
боли.
Начальным, или самым периферическим, звеном системы
восприятия боли является раздражение болевых рецепторов (ноцицепторов),
представляющих собой свободные нервные окончания афферентных волокон. Различают
три типа рецепторов боли: мономодальные А-d ноцицепторы, реагирующие на механическое и термическое
раздражение; полимодальные С-ноцицепторы,
раздражаемые механическими, тепловыми и химическими стимулами, и так называемые
«спящие» ноцицепторы. Эти ноцицепторы
активизируются только во время воспаления. Анатомически первый тип ноцицепторов представлен свободными нервными окончаниями,
разветвленными в виде дерева (миелинизированные
волокна). Они представляют собой быстрые А-d –
волокна, проводящие раздражение со скоростью 6–30 м/с. Эти волокна возбуждаются
высокоинтенсивными механическими и иногда термическими раздражениями кожи.
Другой
тип ноцицепторов (С-ноцицепторы
и молчащие ноцицепторы) представлен плотными некапсулированными гломерулярными
тельцами. Их афферентные волокна (немиелинизированные
С-волокна, проводящие раздражение со скоростью 0,5–2 м/с) у человека и других
приматов представлены полимодальными ноцицепторами,
поэтому реагируют как на механические, так и на температурные и химические
раздражения. С-волокна распределяются во всех тканях за исключением центральной
нервной системы, они присутствуют и в периферических нервах (nervi nervorum). Рецепторы кожи связаны с обоими видами
нервных волокон, а глубокие ткани, внутренние органы в основном иннервируются
наиболее медленно проводящими безмиелиновыми
С-волокнами. Волокна, связанные с рецепторами, воспринимающими повреждение
тканей, содержат так называемые сенсорные нейротрансмиттеры,
к которым относят субстанцию Р (СР, от английского рain – боль) и кальцитонин ген-связанный пептид
(КГСП), а волокна из внутренних органов – вазоактивный
интестинальный пептид. Возникшие при возбуждении ноцицепторов нерв ные импульсы по
периферическим отросткам попадают через узел заднего корешка (УЗК) в спинной
мозг.
Первым
центральным звеном, воспринимающим разномодальную
афферентную информацию, является нейрональная система
заднего рога спинного мозга. Она представляет собой цитоархитектонически
весьма сложную структуру, которую в функциональном отношении можно
рассматривать, как своеобразный первичный интегративный
центр сенсорной информации[3].
Выделяют
ноцицептивную, невропатическую и психогенную боль.
Ноцицептивная боль возникает
при любом повреждении ткани, вызывающем возбуждение периферических болевых
рецепторов и специфических соматических или висцеральных афферентных волокон. Ноцицептивная боль, как правило, транзиторная или острая,
болевой раздражитель очевиден, боль обычно четко локализована и хорошо
описывается больными. Исключение составляют висцеральные боли и отраженные
боли. Для ноцицептивной боли характерен быстрый
регресс после назначения короткого курса болеутоляющих средств, в том числе
наркотических анальгетиков.
Невропатическая боль обусловлена повреждением или
изменением состояния соматосенсорной (периферических и/или центральных отделов)
системы. Невропатическая боль может развиваться и сохраняться при отсутствии
явного первичного болевого раздражителя, проявляется в виде ряда характерных
признаков, часто плохо локализована и сопровождается различными нарушениями
поверхностной чувствительности: гипералгезией
(интенсивная боль при легком ноцицептивном
раздражении зоны первичного повреждения, либо соседних и даже отдаленных зон); аллодинией (возникновение болевого ощущения при воздействии
не болевых, различных по модальности раздражителей); гиперпатией
(выраженная реакция на повторные болевые воздействия с сохранением ощущения
сильной боли после прекращения болевой стимуляции); болевой анестезией
(ощущение боли в зонах, лишенных болевой чувствительности). Невропатическая
боль мало восприимчива к морфину и другим опиатам в обычных анальгетических
дозах, что свидетельствует об отличии ее механизмов от ноцицептивной
боли.
По отношению к степени вовлечения симпатической
нервной системы спонтанные боли можно разделить на симпатически независимые и
симпатически обусловленные. Симпатически независимая боль связана с активацией
первичных ноцицепторов в результате повреждения периферического
нерва и исчезает (или значительно регрессирует) после местной блокады
анестетиком поврежденного периферического нерва или пораженного участка кожи.
Болевой синдром такого типа связан с высвобождением медиаторов воспаления, в
этом его сходство с болевым синдромом, развивающимся при воспалении в
отсутствие повреждения периферического нерва.
Психогенные боли возникают в отсутствие какого–либо органического
поражения, которое позволило бы объяснить выраженность боли и связанные с ней
функциональные нарушения. Вопрос о существовании болей исключительно
психогенного происхождения является дискуссионным, однако те или иные
особенности личности пациента могут влиять на формирование болевого ощущения.
Психогенные боли могут быть одним из множества нарушений, характерных для соматоформных расстройств. Любое хроническое заболевание
или недомогание, сопровождающееся болью, влияет на эмоции и поведение личности.
Боль часто ведет к появлению тревожности и напряженности, которые сами
усиливают восприятие боли. Психофизиологические (психосоматические) механизмы,
воздействуя через кортикофугальные системы, изменяют состояние внутренних
органов, поперечнополосатой и гладкой мускулатуры, стимулируют выделение альгогенных субстанций и активацию ноцицепторов.
Возникающая боль, в свою очередь, усиливает эмоциональные нарушения, замыкая
таким образом порочный круг. Среди других форм психических расстройств наиболее
тесно связанной с хронической болью является депрессия. Возможны различные
варианты временных взаимоотношений этих нарушений – они могут возникать
одновременно или одно опережать проявления другого. В этих случаях депрессия
чаще имеет не эндогенный, а психогенный характер. Взаимоотношения боли и
депрессии довольно сложны. У больных с клинически выраженной депрессией
снижается болевой порог, и боль считается обычной жалобой у больных с первичной
депрессией. У больных с болевым синдромом, обусловленным хроническим
соматическим заболеванием, часто также развивается депрессия. Наиболее редкой
формой боли при психических заболеваниях является ее галлюцинаторная форма, что
наблюдается у больных эндогенными психозами[4].
Исследования (О.В.
Петров, 1997) показали, что ЦНС функционирует так, что сигналы от различных
органов чувств, импульсов, для нее изначально равнозначны. Важность того или
иного информационного потока определяется не типом его модальности, а тем,
какое значение этой модальности придает представление обокружающем
мире, создавшееся в ЦНС. Для описания работы ноцицептивной
системы организма, полезно введение понятие «образа боли». Это позволяет
осознать то, что ноцицептивная импульсация,
независимо от своей мощности, становиться болью тогда и только тогда, когда ЦНС
при поступлении этой информации распознает ее как «образ боли». Соответственно,
обезболивание может означать нарушение процесса распознавания образа боли. По
существу вышесказанное является краткой формулировкой информационной концепции
боли, которая определяет, что такое боль и что такое обезболивание. В настоящее
время известно три принципиально различных метода общего обезболивания. Самый
простой и наиболее распространенный - фармакологический. Он основан на
разрушении способности мозга распознавать внешние образы вообще, например
применение общих анестетиков. Менее распространен метод «перепрограммирования
образа боли». Он позволяет проводить обезболивание под гипнозом. Третий – «физиотерапевтический»
- основан на конкурентном вытеснении ноцицептивной
информации из ЦНС. При соответствующих условиях он производит ошеломляющий
эффект, например, при аудиогенной анальгезии, электроанестезии и иглорефлексотерапии.
3. Функции
ретикулярной формации
Ретикулярная
формация - состоит из многочисленных, не имеющих четких границ групп нейронов,
длинные и сильно ветвящиеся отростки которых формируют сети в дорсальной части
ствола мозга и вокруг серого вещества спинного мозга.
Впервые
описана в середине 19 века, а название этой структуре, происходящее от лат.
слова reticulum - сеточка, дал О.Дейтерс.
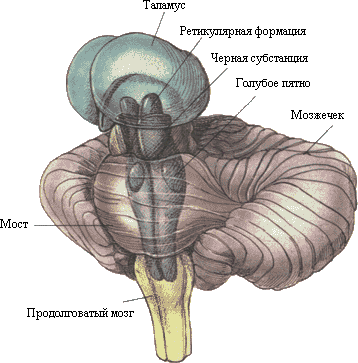
Рис. 2 Расположение ретикулярной формации
Классификация
В
ретикулярная формация ствола мозга выделяют свыше 100 ядер, которые на
протяжении от промежуточного мозга до спинного мозга объединяются в три основные
группы.
1)
Срединная группа ядер концентрируется вокруг средней линии, в основном, в
области шва моста и продолговатого мозга (ядра шва). Шов продолговатого мозга и
моста образован волокнами чувствительных проводящих путей, следующих от
спинного мозга, ядер тройничного нерва и формирующих перекрест вдоль средней
линии.
2)
Медиальная группа ядер расположена по сторонам от предыдущей. Ей принадлежат
медиальное крупноклеточное ядро, голубоватое место, нейроны центрального серого
вещества среднего мозга и др.
3)
Латеральная группа ядер находится латеральнее
медиальной и включает латеральное ретикулярное ядро, парабрахиальные
ядра и др.
Нейроны
ретикулярной формации имеют различную величину, однако в ядрах ствола мозга
имеется определенная закономерность их распределения - в срединных и медиальных
ядрах находятся крупные нервные клетки, а в латеральных - средние и мелкие
нейроны. Отростки крупных нейронов формируют длинные афферентные и эфферентные
проводящие пути, а мелкие и средние клетки являются, в основном, ассоциативными
нейронами.
Медиаторный химизм
Следует
отметить, что нейроны ретикулярной формации различаются не только величиной, но
и составом медиаторов. Так, большинство клеток в качестве передатчика нервного
импульса используют пептиды (энкефалины, нейротензин и т.д.). Вместе с тем широко представлены и
моноамины. Так, ядра шва содержат серотонинергические
нейроны, а нервные клетки клетки голубоватого места
являются норадренергическими.
Связи Ретикулярная формация
Афферентные
связи. На нейронах Ретикулярная формация заканчиваются волокна:
-от
спинного мозга, следующие по коллатералям (ответвлениям) всех чувствительных
проводящих путей, а также по спиноретикулярному
тракту
-от
ядер черепных нервов в составе коллатералей ядерно-корковых, слухового и
зрительных путей
-от
мозжечка в составе мозжечково-ретикулярного пути
-от
ядер таламуса, субталамуса и гипоталамуса
-полосатого
тела
-структур
лимбической системы
-различных
участков коры большого мозга, в том числе и по коллатералям
корково-спинномозговых и корково-ядерных путей.
Эфферентные
связи. Нейроны Ретикулярной формации имеют длинные тонкие отростки, или аксоны,
делящиеся на восходящую и нисходящую ветви, которые направляются к различным
отделам головного и спинного мозга:
-моторным
нейронам передних рогов спинного мозга и двигательным ядрам черепных нервов
ствола мозга в составе ретикуло-ядерных и ретикуло-мозжечковых путей;
-мозжечку;
-красному
ядру, черному веществу и ядрам пластинки крыши спинного мозга
-ретикулярным
ядрам таламуса
-ядрам
гипоталамуса
-опосредованно,
через ядра промежуточного мозга к полосатому телу, лимбической
системе и новой коре.
С
помощью ретикулярной формации двигательные и вегетативные ядра ствола мозга
объединяются в функциональные центры, регулирующие многие сложные: циркуляторные, дыхательные, кашлевой, глотательный, рвотный
и др.
Функции ретикулярной формации
1)
Поддерживание состояния бодрствования - участие в цикле сон/бодрствование.
Увеличивая или уменьшая поток сенсорной информации к коре больших полушарий и
подкорковым структурам, Ретикулярная формация играет роль своеобразного
вентильного механизма, регулирующего уровень сознания.
2)
Контроль рефлекторной деятельности путем стимуляции или торможения мотонейронов передних рогов серого вещества спинного мозга
и двигательных ядер черепных нервов ствола мозга. В медиальной части
продолговатого мозга расположен ингибиторный центр, стимуляция которого
понижает тонус скелетных мышц, угнетает рефлексы и препятствует прохождению к мотонейронам любых импульсов со стороны коры большого мозга
и ядер экстрапирамидной системы. Напротив, Ретикулярная формация моста и
среднего мозга активируетнервные клетки
периферических двигательных центров.
3)
Интегративная деятельность. Объединяет группы
нейронов различных отделов головного и спинного мозга, благодаря чему возможно
выполнение сложных рефлекторных актов: глотания, жевания, кашля, рвоты и т.д.
4)
Обеспечение вегетативной регуляции за счет координации сигналов в
соответствующих центрах ствола мозга. Так, сосудодвигательный и дыхательный
центры объединяют группы нейронов, ответственных за регуляцию дыхания и
кровообращения.
5)
Участие в эмоциональном воспринятии чувствительных
сигналов путем увеличения или уменьшения поступления афферентных импульсов к лимбической системе.
Регулируя
медиаторный обмен нейронов Ретикулярная формация или
модулируя активность их рецепторов с помощью определенных лекарственных
препаратов или немедикаментозных средств, можно, в
случае необходимости, продлить состояние бодрствования и активизировать
деятельность коры больших полушарий или наоборот - добиться сна. Например,
кофеин, содержащийся в кофе или чае стимулирует нервные клетки ретикулярной
формации[5].
Список
использованной литературы
1.
Алексеев
В.В. Основные принципы лечения болевых синдромов // Российский медицинский
журнал. № 5. 2003.
2.
Бабский
Е.Б. Физиология человека. М.: Просвещение, 1985.
3.
Костюк П.Г. Физиология центральной нервной системы. Киев: Высшая школа, 1971.
4.
Меринков
М.Д. Человек и лекарство // Российский медицинский журнал. № 3. 2000.
5.
Федюкович Н.И. Анатомия и физиология человека. Ростов н/Д: Феникс, 1995.
[1] Бабский Е.Б. Физиология
человека. М.: Просвещение, 1985. С. 155.
[2] Федюкович
Н.И. Анатомия и физиология человека. Ростов н/Д:
Феникс, 1995. С. 80-81.
[3] Меринков М.Д. Человек и
лекарство // Российский медицинский журнал. № 3. 2000. С. 19.
[4]
Алексеев В.В. Основные принципы лечения болевых синдромов // Российский
медицинский журнал. № 5. 2003. С. 10-12.
[5] Костюк
П.Г. Физиология центральной нервной системы. Киев: Высшая школа, 1971. С. 331.