Замечательный комплект с ответами на вопросы
по психофизиологии для студентов специальности 020400 (психология).
Созданы на основе учебника «Психофизиология»
Н.Н.Даниловой.
Рязанский Государственный педагогический университет им.С.А. Есенина
Баранов Святослав, 2003 г.
lache.weak@laposte.net
ICQ: 1611030
2.
Принципы кодирования информации в нервной системе.
Одним из простых способов
кодирования информации признается специфичность рецепторов, избирательно реагирующих
на определенные параметры стимуляции
Для каждой модальности эволюция нашла свое более адекватное
решение проблемы передачи информации.
Способ передачи информации
получил название частотного кода. Наиболее явно он связан с
кодированием интенсивности раздражения. Однако позже закон Фехнера был
подвергнут серьезной критике. С. Стивене на основании своих психофизических
исследований, проведенных на людях с применением звукового, светового и
электрического раздражения, взамен закона Фехнера предложил закон степенной
функции. Этот закон гласит, что ощущение пропорционально показателю степени
стимула, при этом закон Фехнера представляет лишь частный случай степенной
зависимости.
В качестве альтернативного
механизма к первым двум принципам кодирования — меченой линии и частотного
кода — рассматривают также паттерн ответа нейрона (структурную организацию
ПД во времени). Устойчивость временного паттерна ответа — отличительная черта
нейронов специфической системы мозга. Система передачи информации о стимулах с
помощью рисунка разрядов нейрона имеет ряд ограничений. В нейронных сетях, работающих
по этому коду, не может соблюдаться принцип экономии, так как он требует
дополнительных операций и времени по учету начала, конца реакции нейрона,
определения ее длительности. Кроме того, эффективность передачи информации о
сигнале существенно зависит от состояния нейрона, что делает данную систему
кодирования недостаточно надежной.
На роль ансамбля нейронов в кодировании информации указал Д.
Хебб. Он считает, что ни один нейрон не может пересылать никакой информации
другим нейронам и что она передается исключительно через возбуждение группы
нейронов, входящих в состав соответствующих ансамблей. Д. Хебб предложил
рассматривать ансамбль нейронов в качестве основного способа кодирования и
передачи информации. Различные наборы возбужденных нейронов одного и того
же ансамбля соответствуют разным параметрам стимула, а если ансамбль находится
на выходе системы, управляющей движением, — то и разным реакциям. Данный
способ кодирования имеет ряд преимуществ. Он более надежен, так как не зависит
от состояния одного нейрона. К тому же не требует дополнительно ни операций,
ни времени. Однако для кодирования каждого типа стимулов необходим свой
уникальный набор нейронов.
Особый принцип обработки информации вытекает из детекторной
теории. Он получил название принципа кодирования информа-
31
ции номером детектора (детекторного канала). Передача информации по номеру канала (термин предложен
Е.Н. Соколовым) означает, что сигнал следует по цепочке нейронов, конечное
звено которой представлено нейроном-детектором простых или сложных признаков, избирательно
реагирующим на определенный физический признак или их комплекс.
Идея о том, что информация кодируется номером канала, присутствовала
уже в опытах И.П. Павлова с кожным анализатором собаки. Вырабатывая условные
рефлексы на раздражение разных участков кожи лапы через «касалки», он установил
наличие в коре больших полушарий соматотопической проекции. Раздражение
определенного участка кожи вызывало очаг возбуждения в определенном локусе
соматосенсорной коры. Пространственное соответствие места приложения стимула и
локуса возбуждения в коре получило подтверждение и в других анализаторах:
зрительном, слуховом. Тонотопическая проекция в слуховой коре отражает пространственное
расположение волосковых клеток кортиевого органа, избирательно чувствительных
к различной частоте звуковых колебаний. Такого рода проекции можно объяснить
тем, что ре-цепторная поверхность отображается на карте коры посредством
множества параллельных каналов — линий, имеющих свои номера. При смещении
сигнала относительно рецепторной поверхности максимум возбуждения перемещается
по элементам карты коры. Сам же элемент карты представляет локальный детектор,
избирательно отвечающий на раздражение определенного участка рецепторной
поверхности. Детекторы локальности, обладающие точечными рецептивными полями и
избирательно реагирующие на прикосновение к определенной точке кожи, являются
наиболее простыми детекторами. Совокупность детекторов локальности образует
карту кожной поверхности в коре. Детекторы работают параллельно, каждая точка
кожной поверхности представлена независимым детектором.
Сходный механизм передачи сигнала о стимулах действует и тогда,
когда стимулы различаются не местом приложения, а другими признаками.
Появление локуса возбуждения на детекторной карте зависит от параметров
стимула. С их изменением локус возбуждения на карте смещается. Для объяснения
организации нейронной сети, работающей как детекторная система, Е.Н. Соколов
предложил механизм векторного кодирования сигнала.
Принцип векторного кодирования информации впервые был
сформулирован в 50-х годах шведским ученым Г. Йохансоном, который и положил
начало новому направлению в психологии — векторной психологии. Г. Йохансон
основывался на результатах
детального изучения восприятия движения. Он показал, что если две
точки на экране движутся навстречу друг другу — одна по горизонтали, другая по
вертикали, — то человек видит движение одной точки по наклонной прямой. Для
объяснения эффекта иллюзии движения Г. Йохансон использовал векторное представление.
Движение точки рассматривается им как результат формирования двухкомпонентного
вектора, отражающего действие двух независимых факторов (движения в
горизонтальном и вертикальном направлениях). В дальнейшем векторная модель
была распространена им на восприятие движений корпуса и конечностей человека,
а также на движение объектов в трехмерном пространстве. Е.Н Соколов развил
векторные представления, применив их к изучению нейронных механизмов сенсорных
процессов, а также двигательных и вегетативных реакций.
Векторная психофизиология — новое направление,
ориентированное на соединение психологических явлений и процессов с векторным кодированием
информации в нейронных сетях.
3. Вызванные потенциалы
Сенсорные стимулы вызывают изменения в суммарной электрической
активности мозга, которые выглядят как последовательность из нескольких
позитивных и негативных волн, которая длится в течение 0,5-1 с после стимула.
Этот ответ получил название вызванного потенциала (evoked potential). Его нелегко выделить из фо-16
новой ЭЭГ. В 1951 г. Дж. Даусон (G. Dawson) разработал технику когерентного
накопления или усреднения ответов. Использовалась процедура синхронизации ЭЭГ
относительно момента предъявления стимула, который поэтому многократно
повторялся. Сначала использовалась суперпозиция — наложение нескольких реакций
(участков ЭЭГ, следующих за стимулом). Обычно это выполнялось на фотопленке,
что позволяло выявить наиболее устойчивые части реакции на стимул. Затем
процедура суперпозиции была заменена на суммацию участков ЭЭГ и получение усредненного
вызванного потенциала (average
evoked potential) (Шагас Ч., 1975; Рут-ман Э.М.,
1975).
Эффективность этой процедуры была наглядно продемонстрирована при
выявлении звуковых стволовых вызванных потенциалов (ВП). Из-за их очень малой
амплитуды требуется просуммировать и усреднить несколько тысяч единичных
ответов. На рис. 2 представлены основные группы компонентов звукового усредненного
ВП. По латентному периоду компоненты делятся на три группы: потенциалы ствола
мозга (с латенцией до 10—12 мс), средне-латентные (до 50 мс) и длиннолатентные
(более 100 мс) потенциалы. Звуковые стволовые потенциалы состоят из 7
отклонений. Волна I зависит
от реакции волокон слухового нерва улитки. Волна II с латенцией 3,8 мс возникает в том случае, если импульсы
слухового нерва достигают ствола мозга. Волна III отражает реакцию верхней оливы на уровне моста. Волна IV с латенцией около 4,5 мс связана с
активностью латеральных лемнисков. Волна V имеет латенцию около 5,2 мс и отражает активность нижнего
двухолмия. Фазы VI— VII — распространение сигналов по
таламо-кортикальной радиации, они совпадают с медленной негативностью,
предшествующей корковому ответу. Ранние компоненты нечувствительны к сну,
наркозу. Они вызываются звуковыми тонами частотой 2000—4000 Гц. Звуки на
частоте ниже 2000 Гц вызывают только волну V.
Стволовые потенциалы — высокочувствительный инструмент
для тестирования слуховой функции. Они позволяют определить сохранность
слухового анализатора на периферическом и стволовом уровнях. Особенно это
важно при обследовании слуха у детей, в том числе у новорожденных, когда словесные
реакции не могут быть использованы. Значение этого теста возрастает в связи с
тем фактом, что даже незначительная потеря слуха в раннем детстве может
привести к существенной задержке развития речи. Стволовые звуковые потенциалы
применяют также в клинике для выявления опухолей, определения коматозного
состояния, обследования пациентов с демиелинизацией волокон. Если стволовые потенциалы
полностью отсутствуют, можно говорить о смерти мозга.
17
2 - 6384
Среднелатентные и длиннолатентные компоненты отражают функционирование
кортикального уровня слухового анализатора. Среднелатентные компоненты (Н0,
П(), На, Па, Н6) регистрируются от
первичной слуховой коры, имеют "малую амплитуду, более лабильны, чем
стволовые потенциалы, чувствительны к сну, наркозу. Максимальная их амплитуда
вызывается звуковыми тонами речевого диапазона. Длиннолатентные ответы
включают компонент Н,с латенцией пика в 100 мс. Потенциал характеризуется 18
полимодальностью и чувствительностью к активации. Кроме того, на
него может накладываться другой потенциал — негативность рассогласования (HP), которую связывают с процессами
пред-внимания (см. главу «Внимание»). Компонент П2 имеет специфические
и неспецифические составляющие. Волна Н2 также включает несколько
компонентов.
Позже техника усреднения ВП была применена для выявления
потенциалов, связанных с движением. Участки ЭЭГ усреднялись относительно не
стимула, а начала движения. Это дало возможность исследовать моторные
потенциалы и потенциалы готовности, предшествующие движению. Для
обозначения всех групп потенциалов был введен общий для них термин —
«потенциалы, связанные с событиями» (ПСС), объединяющий ВП, моторный потенциал
и др.
На основе многоканальной регистрации ЭЭГ был разработан метод картирования
биотоков мозга (brain
mapping). Картирование дает
представление о пространственном распределении по коре любого выбранного
показателя электрической активности мозга. Это может быть ВП, один из его
компонентов или альфа-ритм (или другие частотные полосы спектра ЭЭГ). Значения мощности
выбранного показателя подразделяются на уровни. В одном варианте каждому
уровню приписывается свой цвет и изменение локу-са активности выглядит как
перемещение определенного цвета по карте. В другом варианте значения
показателя, принадлежащие одному уровню, соединяются изолиниями, как на
топографических картах, на которых можно видеть возвышенности и впадины. Рассматриваются
карты, полученные в разное время и в разных условиях. Этот метод позволяет
выявить фокусы активности мозга. Используется процедура вычитания одной карты
потенциалов из другой, что позволяет связать паттерн ЭЭГ-активности с той или
другой когнитивной операцией. На рис. 3 приведен пример картирования мозговой
активности по основным ритмам ЭЭГ для двух состояний взрослого испытуемого
(открытые и закрытые глаза). Измерялась мощность распределения для каждого
ритма (дельта, тета, альфа, бета-1, бета-2) в процентах. Показаны карты максимального
различия и сходства для сравниваемых двух состояний. Открытые глаза, создающие
условия для перцептивной активности, вызывают усиление бета-2 с фокусом в
теменно-затылочной области правого полушария, отвечающего за
конкретно-образное мышление и сенсорно-пространственные преобразования. Второй
фокус активности бета-2 локализован в левой фронтальной коре, функции которой
связаны с управлением выполняемой деятельности и рабочей памятью. Одновременно
открытые глаза усиливают мощность бета-1 в теменно-центральных отведениях обоих
полушарий.
Чтобы сжать информацию, содержащуюся в карте с изолиниями, делают
следующий шаг: рассчитывают некоторый источник тока — диполь, эквивалентный
реальному источнику тока в мозге. Определяют его локализацию, ориентацию,
длину. Таким диполем обычно можно объяснить до 80-90% потенциалов, зарегистрированных
от поверхности черепа. Процедура определения диполя включает построение новой
карты распределения потенциалов, исходя из характеристик первично
рассчитанного диполя. Затем рассчитанную карту сравнивают с исходной картой
потенциалов.
5.
нейронные механизмы восприятия. Детекторы, векторы, гештальты, гностич. единиц.
Сведения, накопленные за
последние десятилетия о нейронах сенсорных систем, подтверждают детекторный
принцип нейронной организации самых разных анализаторов. Для зрительной коры
были описаны нейроны-детекторы, избирательно отвечающие на элементы фигуры,
контура — линии, полосы, углы (рис. 6). Американские ученые Д. Хьюбел и Т.
Визель (Hubel D., Wiesel
T.) разработали классификацию
нейронов-детекторов зрительной коры, селективно чувствительных к различной
ориентации линий и их размеру, связав их с простыми, сложными и сверхсложными
рецептивными полями. За эти работы, а также за открытие сенситивного
(критического) периода, в течение которого под влиянием сенсорных воздействий
происходит закрепление запрограммированных свойств нейронов-детекторов и их
изменение за счет избирательной сенсорной депривации, в 1981 г. исследователи
получили Нобелевскую премию.
Нейроны-детекторы звуковысотного слуха были изучены Я. Ка-цуки(1 Katsuki). Крупным событием явились работы С. Зеки (S. Zeki),показавшего существование
детекторов цвета, селективно настроенных на различные оттенки цветов. В
слуховой коре летучих мышей Н. Суга (N. Suga) открыл нейроны-детекторы, которые осуществляют
локацию окружающей среды с помощью отраженных ультразвуковых сигналов,
излучаемых самой мышью. Обладая избирательной чувствительностью к отраженным
звуковым сигналам, они реагируют на определенную локализацию и величину
объектов.
Важным шагом в развитии теории сенсорных систем явилось открытие константных
нейронов-детекторов, учитывающих, кроме зрительных сигналов, сигналы о
положении глаз в орбитах. В теменной коре реакция константных
нейронов-детекторов привязана к определенной области внешнего пространства,
образуя константный экран (Пигарев И.Н., Родионова Е.Н., 1985). Другой
тип константных нейронов-детекторов, кодирующих цвет, открыт С. Зеки в
экстрастриарной зрительной коре, в поле V4. Их реакция на определенные отражательные свойства цветовой поверхности
объекта не зависит от условий освещения.
Д. Хьюбел и Т. Визель провели опыты с частичной деприваци-ей
зрения у котят в сенситивный период. Для этого они временно закрывали один или
оба глаза или же перерезали наружные мышцы одного глаза. После того как веки
котенка вновь открывались, тестирование свойств нейронов показало уменьшение
числа нейронов, отвечающих на возбуждение, поступающее через ранее
депревированный глаз. При повторении аналогичного опыта на детенышах обезьян
были получены примерно такие же результаты: лишь 15% клеток предпочитают
реагировать на сигналы от ранее зашитого глаза, в то время как у интактных
животных было 50% таких клеток. Поведенчески животные были слепы на депревированный
глаз. Если при этом закрывали глаз, который не подвергался депривации,
животные падали со стола, натыкались на ножки стула.
У кошки период пластических перестроек нейронов, благодаря
которому зрительная депривация приводит к корковому дефекту, длится от 4-й
недели по 4-й месяц после рождения. У обезьян этот период начинается раньше, с
момента рождения, и длится дольше, постепенно заканчиваясь к году.
Чувствительность к депривации особенно высока в первые недели сенситивного
периода. Отключение глаза у взрослых животных независимо от продолжительности
не вызывало никаких вредных последствий.
После работ Д. Хьюбела и Т. Визеля во многих лабораториях было
проведено множество экспериментов с разными видами зрительной депривации. В
1970 г. К. Блейкмор и Дж.Ф. Купер
35
(К. Blackmor, G. Cooper) из Кембриджского университета
ежедневно показывали новорожденным котятам в течение нескольких часов
чередующиеся черные и белые вертикальные полосы, а в остальное время содержали
их в темноте. Результатом было сохранение корковых клеток, реагирующих на
вертикальные полосы, и резкое уменьшение числа клеток, предпочитающих другие
ориентации. X. Хирш и Н. Спинели (Н. Hirsch, N. Spineli) использовали очки, позволявшие котенку видеть одним глазом
только вертикальные, а другим — только горизонтальные контуры. В результате
получилась кора, содержащая клетки с предпочтением вертикалей и клетки с
предпочтением горизонталей, но очень мало клеток, отвечающих на наклонные
линии. Кроме того, клетки, реагирующие на горизонтальные линии, возбуждались
лишь через тот глаз, который подвергался ранее воздействию горизонтальных
линий, а на клетки, возбуждаемые вертикальными линиями, влиял лишь глаз,
подвергавшийся воздействию вертикальных линий. В опытах других исследователей
животное содержали в темноте, лишь иногда подавая ему яркий импульс света
(один или несколько раз в секунду). Он позволял животному увидеть, где оно
находится, но сводил к минимуму восприятие любого движения. Результатом этих
экспериментов было уменьшение числа нейронов, чувствительных к движению. Таким
образом, отсутствие естественных изображений на сетчатке в раннем периоде
жизни ведет к глубоким и стойким изменениям свойств нейронов-детекторов.
Изучение вертикальных и горизонтальных связей нейронов-детекторов
различного типа привело к открытию общих принципов нейронной архитектуры коры.
В. Маунткасл (V. Mountcastle) — ученый из медицинской школы Университета Джонса Гопкинса — в
60-х годах впервые описал вертикальный принцип организации коры больших
полушарий. Исследуя нейроны соматосенсорной коры у наркотизированной кошки, он
нашел, что они по модальности сгруппированы в вертикальные колонки. Одни
колонки реагируют на стимуляцию правой стороны тела, другие — левой, а два
других типа колонок различались тем, что одни из них избирательно реагировали на
прикосновение или на отклонение волосков на теле (т.е. на раздражение
рецепторов, расположенных в верхних слоях кожи), другие — на давление или на
движение в суставе (на стимуляцию рецепторов в глубоких слоях кожи). Колонки
имели вид трехмерных прямоугольных блоков разной величины и проходили через все
клеточные слои. Со стороны поверхности коры они выглядели как пластины размером
от 20-50 мкм до 0,25-0,5 мм. Позже эти данные подтвердились и на
наркотизированных обезь-36
янах. Другие исследователи уже на ненаркотизированных животных
(макаках, кошках, крысах) также представили дополнительные доказательства
колончатой организации коры.
В. Маунткасл (1981. С. 26), формулируя основные положения своей
теории о колончатой организации коры, отмечал, что «основной единицей
активности в новой коре служит вертикально расположенная группа клеток с
множеством связей между этими клетками по вертикальной оси и малым их числом в
горизонтальном направлении». Кортикальная колонка представляет собой обрабатывающее
устройство со входом и выходом. Расположение в виде колонок делает возможным
картирование одновременно нескольких переменных на двухмерной матрице
поверхности коры. Между корковыми колонками и их группами существуют специфические
связи.
Благодаря работам Д. Хьюбела и Т. Визеля сегодня мы более детально
представляем колончатую организацию зрительной коры. Исследователи используют
термин «колонка», предложенный В. Маунткаслом, но отмечают, что наиболее
подходящим был бы термин «пластина». Говоря о колончатой организации, они подразумевают,
что «некоторое свойство клеток остается постоянным во всей толще коры от ее
поверхности до белого вещества, но изменяется в направлениях, параллельных
поверхности коры» (Хьюбел Д., 1990. С. 122). Сначала в зрительной коре (поле
17) были обнаружены группы клеток (колонок), связанных с разной глазодоминантностью,
как наиболее крупные. Было замечено, что всякий раз, когда регистрирующий
микроэлектрод входил в кору обезьяны перпендикулярно ее поверхности, он
встречал клетки, лучше реагирующие на стимуляцию только одного глаза. Если же
его вводили на несколько миллиметров в сторону от предыдущего, но также
вертикально, то для всех встречающихся клеток доминирующим был только один
глаз — тот же, что и раньше, или другой. Если же электрод вводили с наклоном и
как можно более параллельно поверхности коры, то клетки с разной глазодоминантностью
чередовались. Полная смена доминантного глаза происходила примерно через
каждый 1 мм.
Структурная организация нейронов коры была уточнена гистохимическими
методами. Открытие нового метода окраски нейронов с применением микроинъекций фермента
пероксидазы хрена в нейроны ЛКТ позволило проследить путь отдельных
аксонов, приходящих из ЛКТ, и распределение их окончаний в коре. Метод основан
на использовании явления аксонного транспорта вещества, введенного в клетку,
которое окрашивает ее, но не влияет на ее структуру. Было установлено, что
каждый аксон из ЛКТ прохо-
37
дит через нижние слои и оканчивается разветвлениями в
IV слое. Здесь отдельные веточки
одного аксона образуют скопления си-наптических окончаний шириной 0,5 мм,
отделенные друг от друга промежутками той же ширины. Волокна от одного глаза
оканчиваются в одних участках, а от другого — в промежутках между ними (рис.
7). Таким образом, каналы передачи зрительной информации от разных глаз в IV слое коры, так же как и в ЛКТ таламу-са, не
пересекаются.
Однако клетки с бинокулярными свойствами, реагирующие на сигналы
от обоих глаз, в коре присутствуют. При этом они составляют более половины
нейронов, расположенных в слоях выше и ниже IV слоя. В верхних слоях коры при переходе от колонки одной
глазодоминантности к другой существуют промежуточные зоны, где смена
доминантности происходит не скачком, а постепенно, проходя через промежуточные
стадии. В промежуточных зонах находятся бинокулярные клетки.
Второй метод, который был применен для изучения колонок
глазодоминантности во всей толщине коры, связан с использованием меченой
дезоксиглюкозы. Метод предложен в 1976 г. Л. Соко-лоффым (L. Sokoloff) в Национальном институте
здоровья в Бе-тезде. Дезоксиглюкоза по химической структуре близка к обычной
глюкозе, которая интенсивно поглощается в качестве источника энергии
возбужденными нейронами. Однако меченая дезоксиглюкоза не может быть полностью
расщеплена нейроном. Она накапливается в нем и может быть обнаружена с помощью
радиоавтографии. Чтобы получить радиоавтограф, делают горизонтальный
срез коры и покрывают его фотоимульсией, на которой после длительной экспозиции
возникает карта распределения радиоактивной дезоксиглюкозы. При стимуляции
одного глаза меченая дезоксиглюкоза будет накапливаться в колонках зрительной
коры, связанных с этим глазом. На рис. 8 представлено распределение меченой
дезоксиглюкозы в срезе коры, взятой параллельно ее поверхности от левого
полушария мозга обезьяны после опыта со стимуляцией одного глаза. Во время
опыта на ненаркотизированной макаке в центр зрительного поля ее правого глаза
в течение 45 мин экспонировали сложный стимул, напоминающий мишень с радиальными
линиями. Другой глаз был закрыт. Предваритель-
39
но животному была сделана инъекция радиоактивной
дезоксиг-люкозы. Перед тем как сделать срез, корковую ткань растянули и
заморозили. На радиоавтографе полукруглые линии стимула отображаются в коре
вертикальными полосками, а радиальные линии — горизонтальными. Пунктирный
характер каждой полоски на срезе обусловлен тем, что в опыте стимулировался
только один глаз, т.е. возбуждались колонки только одной глазодоминантнос-ти,
связанной с правым глазом. Колонки глазодоминантности имеются у кошек,
некоторых низших обезьян, шимпанзе и человека. У грызунов их нет.
Кроме колонок глазодоминантности, в зрительной коре разных
животных (обезьяна, кошка, белка) обнаружены ориентаци-онные колонки. При
вертикальном погружении микроэлектрода через толщу зрительной коры все клетки
в верхних и нижних слоях (кроме IV слоя)
избирательно реагируют на одну и ту же ориентацию линии. При смещении
микроэлектрода картина остается той же, но меняется предпочитаемая ориентация,
т.е. кора разбита на колонки, предпочитающие свою ориентацию. Радиоавтографы,
взятые со срезов коры после стимуляции глаз полосками, определенным образом
ориентированными, подтвердили результаты электрофизиологических опытов.
Соседние колонки нейронов выделяют разные ориентации линий.
В коре обнаружены также колонки, избирательно реагирующие на
направление движения или на цвет. Ширина цветочувстви-тельных колонок в
стриарной коре около 100—250 мкм. Колонки, настроенные на разные длины волн,
чередуются. Колонка с максимальной спектральной чувствительностью к 490-500 нм
сменяется колонкой с максимумом цветовой чувствительности к 610 нм. Затем
снова следует колонка с избирательной чувствительностью к 490-500 нм.
Вертикальные колонки в трехмерной структуре коры образуют аппарат многомерного
отражения внешней среды.
В зависимости от степени сложности обрабатываемой информации в
зрительной коре выделено три типа колонок. Микроколонки реагируют на
отдельные градиенты выделяемого признака, например на ту или другую ориентацию
стимула (горизонтальную, вертикальную или другую). Макроколонки объединяют
микроколонки, выделяющие один общий признак (например, ориентацию), но
реагирующие на разные значения его градиента (разные наклоны — от 0 до 180°). Гиперколонка,
или модуль, представляет локальный участок зрительного поля и отвечает на
все стимулы, попадающие на него. Модуль — вертикально организованный участок
коры, выполняющий обработку самых разнообразных характеристик стимула
(ориентации, цвета, глазодоминантности и др.). Мо-
40
дуль собирается из макроколонок, каждая из которых
реагирует на свой признак объекта в локальном участке зрительного поля (рис.
9). Членение коры на мелкие вертикальные подразделения не ограничивается
зрительной корой. Оно присутствует и в других областях коры (в теменной,
префронтальной, моторной коре и др.).
В коре существует не только вертикальная (колончатая) упорядоченность
размещения нейронов, но и горизонтальная (послойная).
R k'OnOHk'P nfVKP ГТ МЫ С71ЛТГЯ ПП nRlllPVIV ППЬШ-mw
Л ГПГ>Ы дуль собирается из макроколонок, каждая из которых
реагирует на свой признак объекта в локальном участке зрительного поля (рис.
9). Членение коры на мелкие вертикальные подразделения не ограничивается
зрительной корой. Оно присутствует и в других областях коры (в теменной,
префронтальной, моторной коре и др.).
В коре существует не только вертикальная (колончатая) упорядоченность
размещения нейронов, но и горизонтальная (послойная).
н kTinni-ij^p nft"KPпuuu (птга пп
г\Ян1р\л\; nni/ni-iQL'A? Л
пппы
дуль собирается из макроколонок, каждая из которых
реагирует на свой признак объекта в локальном участке зрительного поля (рис.
9). Членение коры на мелкие вертикальные подразделения не ограничивается
зрительной корой. Оно присутствует и в других областях коры (в теменной,
префронтальной, моторной коре и др.).
В коре существует не только вертикальная (колончатая) упорядоченность
размещения нейронов, но и горизонтальная (послойная), Нейроны в колонке
объединяются по общему признаку. А слои объединяют нейроны, выделяющие разные
признаки, но одинакового уровня сложности. Нейроны-детекторы, реагирующие на
более сложные признаки, локализованы в верхних слоях.
Таким образом, колончатая и слоистая организации нейронов коры
свидетельствуют, что обработка информации о признаках объекта, таких, как
форма, движение, цвет, протекает в параллельных нейронных каналах. Вместе
с тем изучение детекторных свойств нейронов показывает, что принцип дивергенции
путей обработки информации по многим параллельным каналам должен быть дополнен
принципом конвергенции в виде иерархически организованных нейронных сетей. Чем
сложнее информация, тем более сложная структура иерархически организованной
нейронной сети требуется для ее обработки.
7.
Характеристики и виды внимания.
К характеристикам внимания
относят его селективность, объем, устойчивость, возможность распределения и
переключения. Селективность, или избирательность, внимания
характеризует его направленность на любой аспект стимула: на его физическую
или лингвистическую характеристику. Изменение селективности внимания подобно
изменению фокуса проектора, создающего либо точечное изображение, либо
расплывчатое пятно, которое не зависит от физических характеристик стимула.
Поэтому внимание может характеризоваться либо узкой, либо широкой настройкой
селективности. Под влиянием стресса фокус внимания обычно сужается. Объем
внимания измеряется количеством одновременно отчетливо осознаваемых
объектов и характеризует ресурсы внимания человека. Объем внимания близок
объему кратковременной памяти и составляет 7—9 стимулов, или элементов.
Эффективность выполнения второго задания часто используют в качестве меры
запасов ресурсов внимания, не востребованных первым, основным заданием. Распределение
внимания предполагает распределение его ограниченных ресурсов для
одновременного выполнения двух или более заданий. Оно часто измеряется в
экспериментах с дихотическим прослушиванием. Устойчивость внимания определяется
по длительности выполнения задания, требующего непрерывного внимания или
«бдительности», например, оператора при обнаружении им редких и слабых
сигналов на экране локатора. Под
переключением внимания понимается возможность более или
менее легкого перехода от одного вида деятельности к другому.
Психологи выделяют три вида внимания. Произвольное, или активное,
внимание характеризуется направленностью субъекта на сознательно выбранную
цель. Описанные выше характеристики внимания в основном рассмотрены на примерах
произвольного внимания. Однако существует форма и пассивного, или непроизвольного,
внимания. Оно выражается в переключении внимания на неожиданное изменение
физических, временных, пространственных характеристик стимулов или на
появление значимых сигналов. Третий вид внимания называется постпроизвольным
вниманием, которое появляется в процессе освоения деятельности и увлеченности
выполняемой работой. Оно не требует усилий воли, так как поддерживается
интересом к ней.
Непроизвольное внимание оперирует только в настоящем времени,
тогда как произвольное внимание может быть обращено как в прошлое, так и в
будущее. Например, мы можем направить свое внимание на след в эхоической
памяти, т.е. на те стимулы, которые звучали несколько секунд назад, — случай,
когда мы пытаемся восстановить смысл слов, только что сказанных нашим собеседником,
которого мы невнимательно слушали. Другой пример обращения внимания в прошлое —
попытка извлечь определенную информацию из долговременной памяти. Примером
внимания, направленного в будущее, является антиципирующее внимание, когда
человек ожидает появления определенного сигнала, события, на которое он должен
отвечать некоторой реакцией. Функция антиципирующего внимания состоит в
облегчении и ускорении опознания цели. Информация, содержащаяся в
предупреждающем сигнале о месте появления императивного сигнала — стимула, на
который нужно реагировать, влияет на время реакции. Если предупреждающий
сигнал правильно указывает место появления императивного, то латентное время
реакции сокращается. При формировании предупреждающим сигналом ошибочной
установки на место появления императивного стимула время реакции на него удлиняется.
Правильное ожидание стимула гарантирует «успех», ошибочное ведет к потерям при
обнаружении релевантного стимула.
Большинство процессов в
мозге, связанных с обработкой информации, протекают автоматически и не требуют
от субъекта специального внимания. Такое мнение сложилось уже в 70-80-х годах,
когда чрезвычайно популярной стала проблема автоматичности восприятия.
Исследователи искали ответы на следующие вопросы: какой тип обработки
сенсорной информации протекает автоматически, каковы пределы этой
автоматичности, какие задания могут выполняться параллельно и в какой степени
обрабатывается незначимая сенсорная информация?
М. Познер и К. Снайдер (Posner
M.I., Snyder
C.R., 1975) выделили три основных критерия отличия автоматических
процессов от контролируемых:
• автоматические процессы
характеризуются непроизвольностью и протекают без усилий субъекта, не
требующих обращения к ресурсам организма, возможности которых достаточно
ограничены;
• они не достигают уровня
сознания;
• не взаимодействуют (не
интерферируют) с какой-либо текущей психической (интеллектуальной)
деятельностью, т.е. не нарушают ее.
Авторы утверждают, что большое количество сложных, но привычных
мыслительных процессов могут протекать автоматически. В качестве примера они
приводят эффект Струппа, который демонстрирует, что субъект не может избежать
обработки тех свойств, которые он и хотел бы проигнорировать. Эффект Струппа
показывает, что обработка сенсорной и семантической информации может
протекать параллельно и автоматически. Испытуемому предъявляются слова —
названия цветов, написанные чернилами разного цвета. Его задача — называть цвет
чернил. Устное выполнение задания замедляется, если семантический компонент
слова не совпадает с перцептивным, и ускоряется, если название цвета и цвет
чернил совпадают. Таким образом, семантическая информация обрабатывается
независимо от установки субъекта, параллельно анализу перцептивных свойств
стимула. Интерференция возникает не между процессами обработки сенсорной и
семантической информации, а на этапе запуска исполнительной реакции: между
речевой реакцией, которую требует задание, и импульсом, стимулирующим
непроизвольное речевое воспроизведение прочитанного слова — названия цвета. При
несовпадении одновременно инициируемых речевых реакций возникает задержка в
исполнении.
Автоматическая обработка семантической информации характерна не
только для эффекта Струппа. Она описана также для ди-хотического прослушивания.
Хорошо известно явление автоматического распространения активации на следы
семантически связанных стимулов, хранящихся в долговременной памяти. Эффекты
иррадиации возбуждения из первой сигнальной системы во вторую и обратно,
обнаруженные и исследованные И.П. Павловым и
сю учениками, также являются автоматически протекающими
процессами. К автоматическим процессам относят операции сравнения стимула со
следами в кратковременной памяти, которые были выявлены при изучении объема
кратковременной памяти и ее влияния на время реакции (парадигма Стенберга и
зрительного поиска). Обработка сенсорной информации нейронными детекторными
системами, создающими базу сенсорных данных для формирования соответствующих
образов и их следов в памяти, также протекает автоматически. Р. Наатанен
связывает эти процессы с досознательным этапом обработки информации — пред-вниманием.
В составе вызванных потенциалов оно представлено негативностью рассогласования
(см. раздел «Непроизвольное внимание»). Предвнимание обнаружено не только
в состоянии бодрствования, но и во сне. Еще один пример автоматических процессов
— это перевод сенсорной информации в сенсорную память (эхоическую,
иконическую). Известно, что цифры, поступающие через игнорируемый канал, могут
быть воспроизведены не позже чем через 5 с — время, в течение которого след от
стимула сохраняется в сенсорной памяти.
Деление когнитивных операций на автоматические и контролируемые
хорошо согласуется с разделением внимания на непроизвольное и
произвольное.
8.
Непроизвольное внимание и ориентировочный рефлекс. Н. модель стимула Соколова
Непроизвольное внимание относится к феномену переключения
внимания на стимул, который ранее не привлекал внимания. Оно протекает
автоматически и не требует специальных усилий. Основу непроизвольного внимания
составляет ориентировочный рефлекс (ОР). Его появление автоматически
включает произвольное внимание, которое и обеспечивает дальнейшую и контролируемую
обработку стимула.
Ориентировочный, или исследовательский, рефлекс был открыт И.П.
Павловым и назван им рефлексом «Что такое?». Он описал его как комплекс
двигательных реакций (поворот головы, глаз, настораживание ушей) в направлении
нового стимула. Ю. Конор-
61
ский (1970) этому комплексу соматических реакций дал
название рефлекса прицеливания, назначение которого — обеспечить лучшее
восприятие нового стимула.
Однако сначала наиболее подробно была исследована тормозная
функция ОР, который всякий раз прерывал текущую деятельность собаки, подавлял
ее условные рефлексы и вызывал общее торможение животного. Представление об ОР
как о многокомпонентной реакции активации сложилось много позже, когда была
создана и получила распространение техника полиграфической регистрации ЭЭГ,
вегетативных и двигательных реакций человека и животных, хотя И.П. Павловым уже
были получены первые данные о зависимости формирования условного рефлекса от
наличия ориентировочного, который обеспечивал необходимую для обучения
активацию мозга.
Е.Н. Соколов (1960), а затем Р. Линн (R. Lynn, 1966) описали систему
компонентов ОР. На уровне неокортекса ОР представлен ЭЭГ-реакцией активации
(блокадой альфа-ритма, усилением высокочастотных колебаний ЭЭГ), коррелирующей
с ростом возбудимости, реактивности и лабильности у нейронов коры. К вегетативным
компонентам ОР относят расширение зрачков, увеличение кожной проводимости,
снижение ЧСС, изменение дыхания, рост дыхательной аритмии в частотном спектре
ритмограммы сердца, расширение сосудов головы и сужение сосудов рук. В моторной
сфере ориентировочный рефлекс, как уже отмечалось, представлен поворотом
головы, движением глаз, настораживанием ушей в направлении нового стимула и
увеличением мышечного тонуса, обеспечивающего повышенную готовность к моторным
реакциям. Выделен сенсорный компонент ОР, который состоит в увеличении
чувствительности анализатора и может быть измерен по субъективным ответам
испытуемого.
Первоначально новый стимул вызывает генерализованный ОР, связанный
с возбуждением ретикулярной формации ствола мозга. Он характеризуется
ЭЭГ-активацией, которая охватывает всю кору в течение достаточно длительного
времени (тоническая форма ОР, часто возникающая параллельно
генерализованному ОР). Генерализованный ориентировочный рефлекс быстро
угасает, обычно после 10—15 применений индифферентного стимула. Это позволяет
выявить локальный ориентировочный рефлекс, который более устойчив к
угасанию и требует 30 и более применений стимула. Его генерацию связывают с
активацией неспецифического таламуса. При локальном ОР реакция ЭЭГ-активации
сохраняется в сенсорной коре, соответствующей модальности повторяемого
раздражителя. Так, свет продолжает вызывать блокаду альфа-ритма, но только
62
в зрительной коре; звук — блокаду тау-ритма, регистрируемого от
нисочной области; тактильное и проприоцептивное раздражение при пассивном
подъеме пальца руки вызывает депрессию мю-рит-ма (роландического ритма)
локально в сенсомоторной коре. Длительность реакции сокращается, она
становится физической, появляясь только на включение и выключение
стимула (фазическая реакция пробуждения). Продолжая повторять стимул, можно добиться
полного угасания ОР. Ориентировочный рефлекс с полным набором компонентов
возникает не у всех испытуемых, что выражает их индивидуальные различия.
Отличительной особенностью ОР является его угасание, развивающееся
с повторением стимула. Однако любое изменение в повторяющемся стимуле
восстанавливает ОР. Оно может быть вызвано изменением модальности,
интенсивности, пространственных и временных характеристик, включая длительность
стимула и меж-стимульные интервалы. Изменения в сложных стимулах или в их
комплексах, например, такие, как пропуск одного из его элементов или изменение
их порядка, также восстанавливают ОР. Величина его восстановления
пропорциональна числу одновременно измененных параметров и степени их
изменения.
Специфическим стимулом, на который всякий раз возникает ОР,
является его абсолютная или относительная новизна. Д. Бер-лайн (D. Berlyne) добавляет к факторам,
вызывающим безусловный ОР, удивление, неопределенность, двусмысленность, сложность,
необычность. Однако, по мнению Р. Линна (Lynn R., 1966), их также можно
интерпретировать как разные формы новизны. Удивление может быть вызвано
появлением чего-либо нового в последовательности стимулов, а другие качества
могут быть связаны с появлением новизны в паттерне стимулов.
Таким образом, ОР, обладая свойством безусловного рефлекса,
возникающего на новизну, вместе с тем обнаруживает сходство и с условным
рефлексом. Оно проявляется в его способности к угасанию. Угасание — это особая
форма обучения, получившая название негативного научения, ведущего к
потере способности данного стимула вызывать ОР.
Для объяснения механизма появления качества новизны у стимула,
вызывающего ОР и последующее его угасание в результате многократного повторения
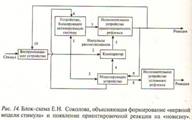
стимула, Е.Н. Соколовым (1963) была предложена теория «нервной модели стимула»
(рис. 14). В его концепции ключевая роль принадлежит блоку нервной модели, которая
формируется в мозге только за счет повторения стимула и без специального
подкрепления. В ней отражаются все параметры повторяющегося стимула (3). Нервная
модель стимула многомер-
63
на, что убедительно доказано во многих работах
восстановлением ОР при изменении самых разных физических характеристик стимула.
Другой ключевой структурой является наличие компаратора — устройства,
выполняющего операцию сопоставления сформированной модели с сигналами
сенсорной системы от предъявляемого раздражителя (2). При их несовпадении
возникает сигнал рассогласования (5), активирующий исполнительное устройство
ориентировочного рефлекса (7). Нервная модель стимула по мере формирования
усиливает свои тормозные влияния на систему неспецифической активации в момент
действия стимула, модель которого уже сформирована (6). При этом второй путь
вызова ОР не работает, так как импульсы рассогласования отсутствуют. Появление
ОР оказывает активирующее влияние на воспринимающее устройство (10).
Предполагается, что при выработке условного рефлекса модель индифферентного
стимула (будущего условного) связывается с исполнительным устройством
условного рефлекса (8). Сигнал от сенсорной системы параллельно поступает к
структурам, где формируется модель (3), на компаратор (2) и к неспецифической
системе (1), где может развиваться привыкание — ослабление реакции активации.
Возникает вопрос: в каких структурах мозга может формироваться
нервная модель стимула? Сначала Е.Н. Соколов указывал на роль коры в
формировании энграмм стимулов. Однако после открытия О.С. Виноградовой (1975) в
гиппокампе двух типов ней-64
ронов, связанных с ОР, он стал подчеркивать роль гиппокампа в
формировании нервной модели стимула. Нейроны В и Т в поле САЗ гиппокампа
отвечают на новый стимул противоположными реакциями. Нейроны В возбуждаются,
нейроны Т тормозятся. Но обе группы нейронов обнаруживают свойства ОР. Е.Н.
Соколов предположил, что нервная модель стимула представлена параллельными
матрицами потенцированных синапсов от корковых нейронов, избирательно отвечающих
на физические свойства, конфигурацию сенсорного стимула, на нейронах новизны
(В-нейро-ны) и тождества (Т-нейроны) гиппокампа. Реципрокные реакции нейронов
новизны и тождества гиппокампа противоположно влияют на активирующую и
инактивирующую системы мозга. Совместное действие обеих групп нейронов
увеличивает отношение стимула к шуму, повышая тем самым надежность системы,
детектирующей новизну. Реакция активации на новый раздражитель определяется
возбуждением активирующей и торможением инак-тивирующей систем мозга. Привычный
стимул перестает возбуждать активирующую систему и тормозить инактивирующую.
Возникновение ОР на изменение привычной стимуляции объясняется тем, что оно
активирует новые, еще не потенцированные (не подвергшиеся пластическим перестройкам)
синапсы нейронов новизны и тождества в гиппокампе (Соколов Е.Н., 1995).
Процесс формирования нервной модели стимула отвечает всем
критериям, предложенным М.И. Познером и К.Р. Снайдером для выделения
автоматически протекающих процессов. Он характеризуется непроизвольностью и
протекает без усилий, не осознается1 и не влияет на текущую
деятельность. В процессе формирования модели информация обо всех физических
свойствах стимула обрабатывается параллельно, без ограничений их количества.
Это происходит и тогда, когда внимание субъекта отвлечено от стимула и
направлено на какую-то деятельность. Операции сличения действующего
раздражителя с моделями памяти также протекают. автоматически. Таким образом,
безусловный ОР — конечное звено серии операций, представляющих автоматическую
обработку информации.
Концепция Е.Н. Соколова о нервной модели стимула и экспериментальные
данные, подтверждающие ее, хорошо согласуются с теориями внимания — ранней и
поздней селекции.
9.
Произвольное внимание. Модель Канемана.
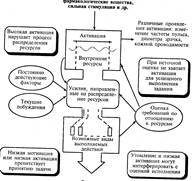
В
модели Канемана внимание регулирует ресурсы, которые понимаются как
недифференцированные энергетические активационные возможности организма. Их
взаимосвязь показана волнистой линией внутри блока активации, символизирующей
определенный параллелизм в изменениях общей активации
и той, которая может быть использована произвольным и непроизвольным
вниманием. Ограниченность ресурсов внимания как части от общей активации
показана сплошной горизонтальной линией в блоке активации. Изменение общей
активации сопровождается соответствующими изменениями энергетических ресурсов,
доступных вниманию. Общая активация организма зависит от многих факторов:
эмоций (тревога, страх, гнев и др.), мышечного напряжения, сенсорных воздействий,
мотивации и др. (стрелка вверху). Но главная детерминанта активации — это
оценка субъектом требований, предъявляемых к энергетическим ресурсам
выполняемой деятельностью. Она показана стрелкой к блоку активации от системы
оценки. Центральным в модели является блок распределения ресурсов. Их
распределение зависит от соотношения сигналов, поступающих от четырех блоков,
потребляющих ресурсы. Текущие побуждения, намерения (мотивация) определяют
направленность произвольного внимания и потребляют соответствующие
энергетические ресурсы. Постоянно действующие факторы — значимые и новые
стимулы — вызывают непроизвольное внимание и соответствующие затраты ресурсов.
Оценка требований, предъявляемых деятельностью к ресурсам внимания, наиболее
сильно влияет на их распределение. Кроме того, существует отрицательное
влияние стресса, потребляющего много ресурсов и при этом дезорганизующего
внимание. Низкая мотивация привлекает недостаточное количество ресурсов, что
снижает эффективность деятельности. Оба эффекта показаны стрелкой от блока
активации к центральному блоку. Оптимальное выполнение задания требует
определенного количества внимания (усилия). Разные виды деятельности потребляют
разное количество внимания. При превышении допустимого уровня ресурсов суммарным
запросом часть видов деятельности прекращается (показано столбиками внизу).
При выполнении каждой задачи используется вход от системы ресурсов (пунктир).
В своей модели, представляющей
процессы обработки информации во время безусловного ориентировочного рефлекса,
Д. Канеман рассматривает факторы новизны и значимости как главные источники ОР.
Концепцию Д.Канемана отличают три
важных положния, определяющие взаимоотношение ОР и произвольного внимания:
1.ОР выполняет функцию запуска
произвольного внимания.
2.Обращение произвольного
внимания на новый стимул возникает ступенчато и рекурсивно, после того, как
стимул уже вызвал ОР..
3. Реакция расширения зрачка и
возрастание кожной проводимости рассматриваются как объективные показатели
произвольного внимания.
В овалах
обозначены факторы, расходующие ресурсы внимания.
В прямоугольниках содержатся комментарии
10.
Модулирующая система мозга. Три субсистемы активации.
Модулирующая система мозга реализует свои функции через
особый класс функциональных систем, регулирующих процессы активации в составе
различных видов деятельности. Она регулирует цикл бодрствование — сон, стадии
и фазы сна, уровни и специфику функциональных состояний во время
бодрствования, а также процессы внимания благодаря ее способности создавать как
локальные, так и генерализованные эффекты активации и инактивации в нервной
системе.
Модулирующая система мозга
представлена многими активирующими и инактивирующими структурами, находящимися
в сложных взаимоотношениях друг с другом и локализованными на разных уровнях
ЦНС. Среди них выделяют неспецифическую систему (ретикулярную формацию)
среднего мозга, которая регулирует состояние и вызывает генерализованные и
тонические реакции активации, а также активирующую—инактивирующую систему
неспецифического таламуса, причастную к возникновению локальных и фазических
реакций активации. К структурам с тормозными функциями относят
синхронизирующий центр Моруц-ци в срединной части варолиевого моста ствола
мозга, преопти-ческую область латерального гипоталамуса. Тормозные функции
выявлены и у фронтальной коры. Важную роль в регуляции цикла бодрствование—сон,
формировании парадоксального сна с быстрыми движениями глаз (КЕМ sleep), а также ортодоксального сна (NREM sleep) играет комплекс синего пятна (locus coeruleus) и ядер шва (n.raphe). Синее пятно — скопление
нейронов в покрышке ствола мозга, в котором осуществляется синтез НА. Последний
в составе переднемозгового пучка доставляется в кору и передний мозг. Ядра шва,
локализованные в средней части продолговатого мозга, моста и среднего мозга,
являются центром синтеза серото-нина, который по системе длинных волокон
достигает многих структур мозга: коры, латерального гипоталамуса, гиппокампа и
др.
Гетерогенность модулирующей системы
Ранее предполагалось, что по динамике любой из физиологических
реакций (ЭЭГ, ЭМГ, диаметр зрачка, кожное сопротивление, ЧСС), испытывающих
влияние неспецифической системы, можно предсказать изменение всех остальных
реакций. Такой подход основывался на концепции Д. Линдсли (D. Lindsley) о единстве и синергизме
влияний от восходящих и нисходящих неспецифических систем мозга,
обусловливающих параллелизм всех электроэнцефалографических, вегетативных и
двигательных реакций. Предполагалось, что активация в ЭЭГ (проявление действия
восходящей активирующей неспецифической системы) развивается параллельно
эффектам нисходящей неспецифической системы в виде соматических и вегетативных
реакций (изменения тонуса мышц, движения глаз, изменения частоты дыхания и
сердцебиения, КГР, изменения диаметра сосудов). Однако в дальнейшем
между ними были найдены очень низкие коэффициенты корреляции. На
основе обзора литературных данных Р. Лазарус (R. Lazarus) приходит к выводу, что
максимальные значения коэффициентов корреляции не превышали 0,5.
Кроме того, при определенных условиях была обнаружена диссоциация
между поведенческой и электроэнцефалографической активациями. Так, введение
атропина собаке вызывало ЭЭГ-кар-тину медленного сна, при этом животное поведенчески
продолжало бодрствовать. Аналогичные данные были получены и на кошках. С
помощью физостигмина у них можно было вызвать активацию на ЭЭГ, но
поведенчески животное могло находиться в состоянии дремоты. Дж. Моруцци
показал, что если перерезать ствол мозга выше варолиевого моста, но ниже
ретикулярной формации, то в результате прерывания синхронизирующих влияний,
поступающих из него в кору, в ЭЭГ развивается десинхронизация. В это время
животное может спать.
Более того, в 1957 г. Дж. Лейси (J. Lacey) впервые обнаружил реципрокные
отношения между вегетативными и электроэнцефалографическими показателями
активации. В его опытах появление депрессии альфа-ритма сочеталось не с ростом
ЧСС, а с ее снижением. Этот комплекс реакций в дальнейшем был связан с ориентировочным
рефлексом. Полученные данные легли в основу концепции Дж. Лейси о
«дирекционной фракционности активации», согласно которой неспецифическая
активация мозга обеспечивается несколькими субсистемами активации. Каждая из
них проявляется в своих вегетативных, моторных и электроэнцефалографических
реакциях.
П.К. Анохину принадлежит
концепция «специфичности неспецифической активации». Она утверждает, что
каждый тип мотивации обеспечивается возбуждением собственной неспецифической
активирующей системы, обладающей особой химической специфичностью. В опытах на
кроликах он выделил и описал различные паттерны ЭЭГ для оборонительных,
пищевых и ориентировочных мотивационных состояний. Применяя различные фармакологические
вещества, он продемонстрировал возможность избирательной блокады каждого из
них. На необходимость выделять две системы активации — лимбическую и
ретикулярную формации с их относительным антагонизмом — указывал А.
Роуттен-берг (Routtenberg А.). По его мнению,
активирующая ретикулярная формация обеспечивает энергетическую базу исполнения
реакций, тогда как лимбическая система связана с подготовительными фазами
поведения и преимущественно имеет отношение к вегетативной активации.
Субсистемы активации
Изучение биохимических основ активационных процессов в мозге и
роли в этом основных медиаторов позволяет выделять три главные системы
активации, причастные к регуляции уровня бодрствования и реакции активации.
Это стволово-таламо-кортикаль-ная система, базальная холинергическая система
переднего мозга и каудо-таламо-кортикальная система.
Стволово-таламо-кортикальная система
Открытие Г. Мэгуном и Дж. Моруцци в 1949 г. в среднем мозге
неспецифической, ретикулярной формации (РФ) сыграло огромную роль в раскрытии
механизмов реакции активации. В опытах с электрической стимуляцией РФ и
отключением ее хирургическими и фармакологическими методами они установили,
что уровень активности коры больших полушарий находится под непрерывным
контролем ретикулярной формации. Высокочастотная электрическая стимуляция РФ
пробуждает животное от сна и поддерживает его бодрствование. У бодрствующей
обезьяны электрическое раздражение РФ сокращает время реакции на сигнальный
стимул и уменьшает дифференцировочный временной порог — критический интервал различения
следующих друг за другом вспышек света. Противоположный успокаивающий эффект
имеет электрическая стимуляция на низкой частоте. Перерезка ствола мозга,
отсекающая активирующую РФ (препарат cervea
isole), вызывала на ЭЭГ картину сна,
характерную для спящего животного.
РФ долгое время рассматривалась как главная система активации с
восходящими проекциями, облегчающими сенсорные процессы, и нисходящими путями,
облегчающими моторную активность. Позже неспецифическая система активации была
найдена в таламусе. Для изучения ее функций Г. Джаспер (Н. Jasper) использовал реакцию вовлечения '(recruting response). Эта реакция возникает в ответ на ритмическую электрическую
стимуляцию неспецифических ядер таламуса и состоит в воспроизведении биопотенциалами
коры ритма электрического раздражения. Реакция вовлечения использовалась для
моделирования ЭЭГ медленновол-нового сна. Применение сенсорных раздражителей
разной модальности на этом фоне вызывало блокаду волн вовлечения, что рассматривалось
как аналог реакции активации. На такой модели впервые было показано
существование локальных реакций активации, они были модально-специфичны и
возникали в коре того анали-
затора, для которого модальность сенсорного стимула была адекватной.
В 1955 г. Джаспер изложил свою теорию о диффузно-проек-ционной таламической
системе, согласно которой неспецифические ядра таламуса через линии своих
проекций на кору вызывают на ЭЭГ реакцию десинхронизации — разрушение
регулярной мед-ленноволновой активности.
Таламическое происхождение реакций активации в коре в дальнейшем
получило подтверждение при параллельной регистрации ЭЭГ коры и нейронной
активности неспецифических ядер таламуса. Показано, что сенсорные раздражения
(звуковое, кожное и др.) вызывают у нейронов неспецифического таламуса реакцию
десинхронизации в виде разрушения пачек спайков и замены их одиночными
потенциалами действия (ПД) (Данилова Н.Н., 1968, 1985). Нейронная таламическая
реакция десинхронизации соответствует появлению на ЭЭГ реакции активации,
которая выглядит как блокада медленных ритмов ЭЭГ или появление
гиппокампаль-ного тета-ритма. На рис. 21 показана связь импульсной активности
двух нейронов таламуса с электрической активностью коры и подкорковых структур
у кролика. Видны усиление пачечной активности нейронов во время медленных волн
и замена их одиночными спайками во время реакции ЭЭГ-активации.
Реакция десшхронизации нейронов неспецифического таламуса обладает
свойствами ОР. С повторением стимула она ослабевает и восстанавливается после
экстрараздражителя. Рис. 22 демонстрирует динамику одного из таких нейронов,
реакция которого оценивалась по продолжительности нейронной реакции
десинхрониза-ции и по частоте одиночных спайков во время нее.
Таламическая неспецифическая система определяет появление в коре локальных
форм активации. Из-за стратегической позиции одного из ядер
неспецифического таламуса — ретикулярного ядра
(n.Ret) — предполагают его ключевую роль в происхождении локальной
активации; оно является воротами для сенсорной информации, поступающей в кору.
Однако ретикулярное ядро не имеет прямого выхода на кору. Вместе с тем оно
получает входы от коры, ствола мозга, а также от всех других ядер таламуса. При
этом его собственный выход может выборочно тормозить или не тормозить те или
другие специфические ядра таламуса. Это единственное ядро, которое не имеет
своих проекций в коре, и единственное ядро с тормозным выходом, по-видимому,
выполняющее функцию внут-риталамического регулятора.
Кроме того, возможно, что это ядро через неспецифический таламус
регулирует паттерны активации, необходимые для произвольных движений.
Ретикулярное ядро находится под контролем РФ, которая может перекрывать
селективное торможение, создаваемое n.Ret, и запускать генерализованный ОР.
В работах норвежских исследователей из Гетеборгского университета
(Осло) П. Андерсена (P. Anderssen) с сотрудниками была изучена
роль нейронов специфических ядер таламуса в происхождении локальных корковых
реакций активации и инактивации. Они показали таламическое происхождение сонных
и барбитуровых веретен коры и определили размеры участков коры и таламуса,
образующих линии проекции, по которым из таламуса распространяются на кору
синхронизирующие и десинхронизирующие влияния. Двигая один из подкорковых
электродов, они определяли корреляцию электрической активности обеих структур.
Высокая корреляция была найдена между корковыми и таламическими волнами для
всех специфических ядер таламуса (латерального и медиального коленчатых тел,
вентро-базального комплекса). Участки с высокой корреляцией в коре имели
площадь диаметром 0,8 мм, в таламусе — 100-150 мкм. На основании расчета
исследователи предположили, что в специфическом таламусе существует около
25000— 30000 локальных пейсмекеров медленноволновой активности, что совпадает
с предполагаемым количеством колонок в коре одного полушария кошки. Согласно их
теории локальных пейсмекеров специфического таламуса каждый из таких
пейсмекеров контролирует одну из корковых колонок через те же нейроны, которые
передают в кору специфическую информацию. В зависимости от режима работы
локального пейсмекера (одиночные спайки или пачечные разряды) в кору локально
приходят либо активирующие, либо тормозные посылки.
Генерализованная активация охватывает обширные зоны коры. Ее
происхождение связывают с функцией РФ ствола мозга. Генерализованная активация
более длительна и быстрее угасает по сравнению с локальной активацией
таламического происхождения. Принято считать, что функция стволовой
неспецифической системы состоит в поддержании в мозге определенного
функционального состояния, тогда как неспецифическая система в неспецифических
и специфических ядрах таламуса связана с селективным вниманием.
Как показало изучение биохимических основ ЭЭГ-реакции активации,
кора получает широкие проекции от четырех основных медиаторных систем —
ДА-ергической, НА-ергической, се-ротонинергической и холинергической, однако
только последняя причастна к ее активации. Кортикальная активация, вызванная
электрической стимуляцией РФ ствола, сопровождается высвобождением
ацетилхолина (АХ) в коре, т.е. РФ действует на кору через холинергические
афференты. Конечное звено корковой активации холинергично и представлено
мускариновыми (М-) рецепторами нейронов коры, чувствительными к АХ. Локальное
наложение АХ на кору возбуждает примерно до 50% ее нейронов. Атропин, избирательно
угнетающий М-рецепторы, блокирует этот эффект. Сенсорные раздражения разной
модальности увеличивают количество высвобождающегося АХ в коре, а
деафферентация животного сопровождается падением свободного АХ. Реакция
ЭЭГ-активации коррелирует с ростом количества АХ в жидкости, омывающей
поверхность коры. Наркоз снижает выход АХ, одновременно блокируя
ЭЭГ-активацию.
Во время парадоксального сна активация также имеет
холи-нергическую природу, которая развивается в коре на фоне редукции
норадренергической и серотонинергической модуляций из-за снижения активности
нейронов синего пятна и ядер шва. По данным ПЭТ, во время парадоксального сна
активацию находят в покрышке варолиева моста, в лимбической системе: миндалине,
парагиппокампальной коре, передней поясной коре и тесно связанной с
лимбическими структурами медиальной префронталь-ной коре при одновременной
деактивации дорзолатеральной пре-фронтальной коры, в которой представлен
аппарат рабочей памяти (см. главу «Память»). Такой паттерн возбуждения во
время парадоксального сна создает уникальные условия для обработки
исключительно внутренней информации, функционально изолированной от сенсорного
входа и от выхода (через дорзолатераль-ную префронтальную кору) относительно
внешнего мира.
Однако до сих пор остается неясным, через какие пути РФ среднего
мозга распространяет свою активацию на кору. Утверждение, что РФ имеет прямой
выход на кору, основано на косвенных доказательствах, полученных в
поведенческих и физиологи-
ческих опытах. Никогда не были точно показаны анатомические связи
стволовой РФ с корой. Более детальное изучение этой проблемы показало, что
выход стволовой РФ на кору осуществляется, по-видимому, через базальную
холинергическую систему переднего мозга, которая моносинаптически
проецируется на кору. Кроме того, она имеет вход от РФ и один из ее выходов на
кору проходит через таламус.
Базальная холинергическая
система
В базальном переднем мозге (Basal Forebrain) найдены магно-целлюлярные
нейроны, содержащие АХ и моносинаптически проецирующиеся на кору. Главная
холинергическая структура — ядро Мейнерта (n.basalis — NB). Электрическая стимуляция NB, а также SI (substancia innominata) у анестезированной крысы высвобождает в коре АХ. Эффект можно
блокировать атропином — антагонистом мускариновых рецепторов. Параллельно
электрическая стимуляция увеличивает кровоток в коре, что указывает на холинергическую
природу расширения сосудов мозга.
Холинергическая система переднего мозга принимает участие как в
регуляции цикла сон—бодрствование, так и в вызове корковой реакции активации.
В составе холинергической системы мозга различают активирующие и тормозные
структуры, взаимодействие которых определяет уровень активации коры.
Активирующие структуры (NB и др.)
поддерживают бодрствование. В них выделены нейроны, связанные с
бодрствованием. Уровень их активности меняется параллельно циклу
бодрствование—сон, увеличиваясь во время бодрствования и парадоксального сна (Szymusiak R., 1995). Частота их спайков особенно растет во время движения
животного. У этих нейронов, так же как у нейронов таламуса, режим одиночных
спайков в бодрствовании сменяется пачечной активностью во время медленного сна.
«Нейроны, связанные с бодрствованием», находятся под контролем
неспецифической системы среднего мозга и моста (через глутаматовые рецепторы) и
заднего гипоталамуса, который также имеет свою самостоятельную проекцию на кору
и принимает участие в регуляции ЭЭГ и поведенческой активации. На нейроны
бодрствования переднего мозга конвергируют также серотонинер-гические нейроны
ядер шва и клетки синего пятна, вызывающие их инактивацию.
Тормозные структуры базальной холинергической системы представлены
магноцеллюлярным преоптическим ядром (МСР) и ядром диагонального пучка,
связанным с его горизонтальной ветвью (НОВ), и находятся в реципрокных
отношениях с активирующими структурами. Кроме того, в контроле сна принимает
участие внешняя гипногенная система: медиальная преоптическая область и
передний гипоталамус, оканчивающиеся на активационных хо-линергических
структурах переднего мозга тормозными синапсами.
Холинергическая система переднего мозга является не только
источником активации, поддерживающей бодрствование. Она также обеспечивает
высвобождение АХ в локальных участках коры, которые реагируют на стимулы,
связанные с подкреплением (локальную реакцию на значимые стимулы). При этом
большая часть нейронов переднего мозга реагирует на условные сигналы награды и
не отвечает на условные стимулы наказания. Предполагают, что в базальной
холинергической системе представлен механизм избирательного внимания к
значимым стимулам, который и обеспечивает их обработку в соответствующих
областях коры.
Опыты с фармакологическим отключением базального переднего мозга
у животных показали нарушение приобретенных навыков, включая прохождение
лабиринтов, заученное пассивное и активное избегание.
Холинергическая активирующая система находится под контролем
коры, несмотря на то что кортикофугальные проекции на базальный передний мозг
сравнительно ограничены. Количество таких проекций, идущих от первичных
сенсорных и моторных областей коры невелико. Наиболее заметный путь к
переднему мозгу образован волокнами от орбитофронтальной, инсулярной и особенно
от префронтальной коры. Кортикофугальные влияния достигают холинергической
системы через стриатум и прилегающее ядро после многих переключений. Создавая
локальную активацию в коре, холинергические нейроны переднего мозга
взаимодействуют с таламо-кортикальной неспецифической системой. Показан их
дополнительный выход на кору через таламические структуры, установлены их
проекции к некоторым ядрам таламуса (MD, AV, AM, n.Ret). Предполагают, что, влияя на
кору прямо и через тала-мус, они могут модулировать ее реакции активации в
соответствии со значимостью стимула.
Холинергические нейроны переднего мозга принимают участие и в
реакции активации, которая у животных представлена усилением гиппокампального
тета-ритма. Их активирующие влияния из медиального септального ядра (MS) и ядра диагонального пучка, связанного с
вертикальной ветвью (VDB),
достигают гиппокампа, вызывая тета-ритм. Усиление гиппокампального тета-ритма
в ответ на электрическую стимуляцию РФ реализуется через путь РФ — ба-зальная
Холинергическая система переднего мозга-гиппокамп.
Каудо-таламо-кортикальная
система
Базальные ганглии, образующие стриопаллидарную систему, представлены
комплексом нейрональных узлов, центрально расположенных в белом веществе
больших полушарий головного мозга. Их основные функции связаны с управлением движений,
реализацией врожденного поведения и приобретенных навыков. Кроме того,
стриопаллидарная система участвует в формировании избирательной активации
неокортекса, обеспечивающей готовность афферентного, ассоциативного и
эфферентного звеньев для выполнения конкретного целенаправленного поведения
(Суворов Н.Ф., Таиров О.П., 1985). Именно она распределяет актива-ционные
ресурсы произвольного внимания в соответствии с требованиями, выдвигаемыми
задачей или поведением, которое должно быть реализовано.
Ключевыми структурами базальных ядер являются неостриатум, или
хвостатое ядро (n.caudatus), и прилегающее ядро (n.accumbens). Предполагают, что
хвостатое ядро связано с хранением моторных программ — главным образом
двигательных автоматизмов, выученных движений, навыков, тогда как прилегающее
ядро — более древняя структура — причастна к хранению врожденных поведенческих
программ. Стриопаллидарная система находится под контролем лимбической системы
и неокортекса. Прилегающее и хвостатое ядра фильтруют сигналы соответственно от
лимбической системы (эмоциональный мозг) и от церебральной коры (когнитивный
мозг), используя механизм активации ДА-ергических' нейронов. Активация
ДА-ергических нейронов обеспечивает актуализацию врожденных и прибретенных
форм поведения. Это достигается за счет торможения ГАМК-ергических
интернейронов, тонически тормозящих нейронные сети, в которых хранятся программы
действий.
Наиболее изучены функции хвостатого ядра (неостриатума). Оно
получает проекции практически от всех зон коры. Кроме того, уровень его
активности контролируется системами подкрепления. На нем конвергируют проекции
ДА-ергических и НА-ергических структур ствола мозга, соответственно
представляющие системы положительного и отрицательного подкрепления.
Стриатум участвует в распределении восходящей в кору неспецифической
активации. Это достигается через таламус — систему, определяющую локальную
активацию. Под влиянием стриатума картина распределения активации в таламусе
начинает соответствовать мотивационному возбуждению и кортикофугальным
сигналам, поступающим на стриатум. На уровне коры эта картина трансформируется
в паттерн активации, который отвечает требованиям, поставленным задачей,
или реализации целенаправленного поведения.
Все три системы активации, реализуя
свои специфические функции, вместе с тем имеют общие входы и выходы. Создавая
свои локальные эффекты активации, все системы имеют выход на кору через
таламус. Стволово-таламо-кортикальная система использует таламус для локальной
активации как компонента ОР. Базаль-ная холинергическая система переднего
мозга через таламус реализует корковую активацию на значимые стимулы. Каудо-таламо-кортикальная
система также использует таламус для создания в коре картины распределения
локусов активации, необходимой для выполнения той или иной деятельности. Кортикофугальные
влияния, главным образом из префронтальной коры, достигают холи-нергической
активационной системы через стриопаллидарную (хвостатое и прилегающее ядра). Генерализованная
реакция активации, вызываемая сенсорными стимулами вследствие возбуждения
РФ среднего мозга, опосредована холинергической системой переднего мозга, si Последняя
имеет прямое отношение к регуляции цикла бодрствова- | ние—сон, что сближает механизм
генерализованного ориентировочного рефлекса с функциональным состоянием
бодрствования.
11, 13. Виды памяти. Проблема
классификации памяти.
полученных А.С. Батуевым на обезьянах, у которых вырабатывались
пищевые инструментальные отсроченные двигательные реакции. Исследователь
обнаружил одновременную активацию «нейронов памяти» в лобной и теменной коре.
Они реагировали, изменяя частоту или паттерн активности, только в период
отсрочки (от 5 до 20 с) между условным сигналом (вспышкой света) и двигательной
реакцией — нажимом на рычаг, который открывал кормушку с пищей. У обезьяны
вырабатывались дифференцировка условных сигналов по месту вспышки света (слева
— справа) и правильный выбор рычага для нажима.
Между префронтальной корой и височно-теменными областями
неокортекса существуют не только прямые, но и обратные связи. Наличие обратных
связей из префронтальной коры в теменную и нижневисочную подтверждается
появлением в них ответов на электрическую стимуляцию префронтальной коры.
Совместная активация префронтальной и теменной коры показана с
помощью авторадиографического метода (с введением 2-дезоксиглюкозы). После
выполнения теста на запоминание места нахождения фигуры и выполнения
отсроченного ответа у обученной обезьяны уровень метаболической активности в
префронтальной коре и структурах, с которыми она связана, — гиппокампе, нижней
части теменной коры и таламусе — увеличен. При выполнении обычного
инструментального рефлекса без необходимости в течение некоторого времени
удерживать в памяти целевой стимул метаболическая активность этих структур
заметно слабее.
Только наличием обратных связей от префронтальной коры к теменной
и височной можно объяснить результаты опытов со «зрительным поиском», когда
обезьяна научается выделять с помощью глазодвигательной реакции (саккады)
определенную фигуру на экране. Это означает, что образ целевого стимула в
результате тренировки животного приобретает более четкие очертания, т.е.
происходит подчеркивание, усиление признаков стимула. Это возможно только в
том случае, если нейроны памяти префронтальной коры в зависимости от
результатов подкрепления корректируют следы памяти в височной и теменной коре.
Перезапись информации через обратные связи из префронтальной коры
автоматически функционирует постоянно, обеспечивая приобретение и непрерывную
коррекцию наших знаний.
Существование обратных связей от префронтальной коры к основным
хранилищам памяти позволяет по-новому взглянуть на механизм ретроградной
амнезии. Амнестические агенты действуют на актуализированные энграммы, когда
они используются в режиме рабочей памяти. Это касается как старых следов,
активиронанных для оперативного использования, так и новых, только что
приобретенных. В результате нарушения работы нейронов памяти префронтальной коры
вносится искажение в содержание информации, которая перезаписывается через
обратные связи для длительного хранения, что приводит к избирательной потере
той памяти, которая предварительно была активирована.
Один из вопросов, который продолжает волновать психологов и
психофизиологов, — почему мы, как правило, не помним или очень быстро забываем
свои сновидения? Частичный ответ на этот вопрос дают недавние исследования
метаболической активности структур мозга методом ПЭТ в цикле бодрствование—сон.
Во время парадоксального сна выявлена сильная инактивация префронтальной
(дорзолатеральной и орбитальной) коры — структуры, ответственной за рабочую
память. Одновременно мощная активация охватывает лимбическую систему (особенно
миндалину) и часть ассоциативной зрительной и слуховой коры. Во время парадоксального
сна параллельно инактивации префронтальной коры отмечена редукция высвобождения
норадреналина и серотонина (из синего пятна и ядер шва) — медиаторов,
ответственных за обучение на отрицательном и положительном подкреплении (Hobson J.E., Stickgold R., Pace-Schott
E.F., 1998). Новые комбинации образов, которыми так насыщены
сновидения, вследствие, блокады норадренергической и серотонинергической систем
не получают подкрепления и из-за инактивации механизма рабочей памяти не
переписываются на место постоянного хранения.
15. роль
миндалины в памяти, эмоциях, условных рефлексах.
В 1937 г. чикагские исследователи Г. Клювер и П. Бьюси опубликовали
результаты опытов с удалением у обезьян обеих височных долей вместе с
миндалиной и гиппокампом. После операции у обезьян наблюдалось странное
поведение. У них пропала всякая агрессивность, в том числе та, которая
необходима для самозащиты и поддержания своего статуса во взаимоотношениях с
другими особями. Дикие и агрессивные обезьяны после такой операции становились
спокойными и доверчивыми. При этом они стали менее осторожными, их сексуальная
активность повысилась и стала неупорядоченной. Оперированные животные без
разбору исследовали все объекты, даже опасные, забирая их в рот (психическая
слепота). Кроме того, обезьяны утратили страх, перестали бояться
змей, хотя перед операцией при виде их приходили в ужас. Они как
будто перестали отличать хорошую пищу от плохой, пригодного полового партнера
от непригодного, опасные предметы и сигналы от безопасных. Подобный комплекс
нарушений наблюдался и у больных с повреждением височных долей мозга. Он получил
название «синдром Клювера — Бьюси».
Позже было доказано, что исчезновение эмоций страха прежде всего
связано с нарушением функций миндалины и ее связей с иижневисочной корой, где
локализованы гностические единицы, реагирующие на эмоциональную экспрессию.
Связь миндалины с отрицательными эмоциями и особенно со страхом в 50-х годах
получила подтверждение в опытах нейропсихолога из Оксфорда Л. Вейскранца,
который производил избирательное разрушение миндалины у обезьян.
Миндалина у человека — сложное комплексное образование,
включающеее несколько групп ядер, расположенных в глубине нисочной доли и
имеющих многочисленные связи со многими структурами мозга. Наиболее часто в
миндалине выделяют дорзо-медиальную (или центральное ядро) и базолатеральную
части (или латеральное ядро) (рис. 28). Центральное ядро представляет выход
миндалины к исполнительным механизмам. Электрическая стимуляция миндалины
вызывает у животного эмоцию страха и оборонительные поведенческие реакции.
Критической структурой для получения подобного эффекта у собак является
центральное ядро (Fonberg
E., 1986). Его электрическая
стимуляция вызывает вегетативные и двигательные проявления страха. Аксоны
нейронов центрального ядра после бифуркации достигают латерального гипоталамуса
и центрального серого вещества. Гипоталамус выполняет функцию интеграции
вегетативных, соматических и гуморальных механизмов. Электрическое раздражение
малых участков гипоталамуса вызывает у животного различные типы поведенческих
реакций (реакции страха, агрессии, бегства, пищевые, половые,
терморегуляционные реакции). Присутствие соматического, вегетативного и
гуморального компонентов в составе каждой поведенческой реакции достигается за
счет того, что латеральный гипоталамус (ЛГ) — структура с плохо дифференцированными
ядрами — имеет эфферентные связи с вегетативными и соматическими ядрами ствола
и спинного мозга. Латеральный гипоталамус регулирует гуморальные процессы через
медиальный гипоталамус (МГ) — структуру с хорошо дифференцированными ядрами, с
которыми он имеет двусторонние связи. Именно МГ служит промежуточным звеном
между нервной и эндокринной системами.
Миндалина ответственна не только за безусловные, но и за
условнорефлекторные реакции страха. Отмечено возрастание активности ее
нейронов под влиянием сочетания условного сигнала с безусловным раздражением,
вызывающим страх. При этом ее нейроны не реагировали на изолированное
предъявление условного или безусловного стимула, а также на их случайное
чередование. Удаление или разрушение миндалины устраняет ранее выработанные
условные рефлексы страха и делает невозможным выработку новых.
Миндалина получает сигналы от всех сенсорных систем. Более сложные
поступают от коры, более простые — от специфического таламуса, двухолмия
(верхнего и нижнего). По кортикофугальным путям к ней также приходят сигналы от
нижневисочной коры, где
находятся нейроны, избирательно реагирующие на лица определенных
людей и на различные паттерны эмоциональной лицевой экспрессии. Все сенсорные
сигналы разных модальностей и разного уровня сложности приходят в латеральное
ядро миндалины. Оно является сенсорным интерфейсом по отношению к центральному
ядру.
В основе формирования условного рефлекса страха лежат пластические
изменения синапсов афферентного входа к латеральному ядру миндалины и синапсов
на нейронах центрального ядра, запускающих вегетативные и двигательные реакции
страха. Первое звено пластичности обеспечивает сенсорную специфичность, второе
— моторную специфичность условного рефлекса страха.
Миндалина причастна к формированию не только процедурной
эмоциональной памяти, но и декларативной. Она играет критическую роль в эмоциональном
и социальном поведении высших млекопитающих, так как принимает участие в
декодировании эмоциональных сигналов, посылаемых другими особями, что позволяет
строить поведение в соответствии с их смыслом. После двустороннего удаления
миндалины у приматов нарушается социальное внутригрупповое поведение, так как
животные не могут дать социальную оценку сигналам, главным образом поступающим
через зрительный, слуховой и обонятельные каналы и свидетельствующим об
эмоциях и намерениях партнеров, которая так необходима для группового
поведения. Оперированные животные также не могут связать эту информацию с их
собственными эмоциональными состояниями, определяющими их внутригруп-повые
симпатии и антипатии. Поведенческие расстройства, вызванные удалением миндалин,
связаны с нарушением двусторонней передачи информации между височными долями и
гипоталамусом, которая у интактного животного опосредована миндалиной,
В опытах на животных доказано, что миндалина вовлекается в процесс
лицевого восприятия, в ней выявлены нейроны, так же как и в нижневисочной коре,
реагирующие на определенные лица независимо от их ракурса и ориентации, как и
на паттерны лицевой экспрессии эмоций.
Разрушение миндалины у человека нарушает понимание эмоциональных
сигналов, исходящих от других людей. Анализ клинического случая с частичным
билатеральным разрушением миндалевидного комплекса (пациентка D.R. в возрасте 51 года перенесла
операцию) показал, что больная плохо распознает паттерны лицевой эмоциональной
экспрессии. Если же одно и то же лицо выражало разные эмоции, у больной
возникали трудности при идентификации лица человека. Кроме того, после
операции у нее ухудшилась память на новые лица, она не могла интерпретировать
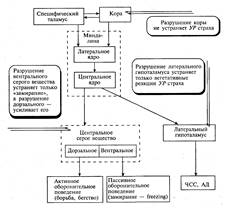
направление взгляда, интонацию речи (Young A.W. et al., 1996; Scott S.K. et al., 1997). Таким образом, разрушение миндалины создает проблему чтения эмоциональных сигналов, важных для социальной коммуникации. Исследователи полагают, что у больной отсутствуют знания о том, какие паттерны эмоциональных сигналов с какими эмоциями связаны, т.е. у нее нарушены условнореф-лекторные связи между сложными сенсорными стимулами и эмоциональными состояниями, которые могут быть индуцированы соответствующим безусловным рефлексом. Из-за разрушения миндалины страдает эмоциональная память — не функционируют и не образуются связи сенсорных сигналов с определенными эмоциональными состояниями.
При этом особенно сильно нарушается распознавание лицевой экспрессии отрицательных эмоций и прежде всего страха. Измерение методом ПЭТ метаболической активности миндалины у человека, когда он рассматривал фотографии лиц, переживающих счастье или страх, выявило избирательную активацию левой миндалины только при восприятии негативной эмоции— страха. С возрастанием интенсивности отрицательной эмоции уровень метаболической активности нейронов увеличивался. Реакция при восприятии «счастливого лица» была противоположной. Активность миндалины подавлялась, и чем сильнее лицо человека выражало эмоцию счастья, тем ниже был уровень метаболической активности в миндалине.
Как формируется декларативная эмоциональная память? Предполагают, что сенсорный сигнал от внешнего мира, поступающий из височных областей коры, и сигналы о состоянии внутренней среды, определяемые гипоталамусом, конвергируют на нейронах миндалины, изменяя их синаптические связи. Это обеспечивает формирование стабильных и длительно сохраняющихся следов эмоциональной памяти.
Миндалина обеспечивает быстрое и прочное запечатление в памяти эмоциональных событий часто после одноразового обучения. По данным Р.Н. Крутикова, удаление миндалины у животного особенно сильно нарушает однократное обучение (пассивное избегание) на отрицательном подкреплении, вызывающем оборонительную реакцию и эмоцию страха. Эффект эмоционального запоминания положительно коррелирует с активностью миндалины. Измерение расхода глюкозы методом ПЭТ во время свободного воспроизведения содержания аверсивных клипов через 3 недели после запоминания показало, что существует линейная зависимость (г = 0,93) между активностью глюкозы в миндалине и числом клипов, сохранившихся в памяти испытуемого. Эта зависимость
характеризует только память на отрицательные эмоции, она не проявляется при воспроизведении эмоционально нейтральных клипов. Эмоциональная память зависит от силы НА-ергических модулирующих влияний. Инъекция в миндалину НА, воздействие электрошоком, увеличивающее также выброс НА, улучшают долговременную эмоциональную память крысы. Прием по поводу ишемической болезни сердца наиболее распространенных лекарств, блокирующих симпатическую систему (пропранолола), приводит к снижению метаболической активности миндалины, необходимой для формирования эмоциональной памяти. Такие больные демонстрируют избирательное нарушение памяти, связанной с эмоциональными эпизодами, при сохранности памяти на нейтральные, неэмоциональные события.
14. роль гиппокампа в памяти.
Гиппокамп — древний отдел мозга. Он тесно связан с височными долями. У приматов гиппокамп прижат к миндалине в височной доле. Гиппокамп имеет мощные входные и выходные связи с перегородкой (septum) в виде толстого пучка волокон (свода). Мощный вход в гиппокамп представлен волокнами из энториаль-ной коры, куда поступают сенсорные сигналы от нейронов-детекторов и гностических единиц. Волокна энториальной коры достигают гиппокампа либо прямо, либо через зубчатую фасцию, оказывающую тормозное влияние на его нейроны. Другой вход в гиппокамп берет начало в поясной извилине — одной из структур лимбической системы.
По мере изучения гиппокампа менялось представление о его функциях. Сначала он рассматривался как кора обонятельного мозга. Затем широко распространилась точка зрения, что гиппокамп ответствен за формирование долговременной памяти. Первые свидетельства о связи гиппокампа с памятью были получены при нейрохирургических операциях на мозге.
В последнее время получены новые данные о причастности гиппокампа к процессам памяти. Они касаются способности гиппокампа различать комплексные раздражители. Оказалось, что удаление гиппокампа лишает животное способности обнаруживать общие элементы в различных стимульных комплексах. Крысы, лишенные гиппокампа, по сравнению с интактным животным быстрее и легче вырабатывают дифференцировки на комплексы в виде последовательности запахов, различающихся лишь одним компонентом. Известно, что интактное животное, чтобы научиться различать комплексы стимулов, должно пройти через стадию, когда стимулы, имеющие общий элемент, смешиваются. Это различие объясняется тем, что у оперированных животных происходит слияние (фузия) всех компонентов последовательности в единый комплекс, что облегчает различение самих комплексов, но не позволяет выявлять отдельные их составляющие. Полагают, что у интактного животного гиппокамп осуществляет диффузию элементов комплекса. Это свойство гиппокампа позволяет ему оживлять все следы ДП (как ранние, так и более поздние) и работать с большим их набором, что делает память обученного животного более гибкой. Удаление гиппокампа делает реакции животного стереотипными. На новые раздражители оно реагирует, как на старые. Оперированное животное трудно переучивается. Функция гиппокампа — оживлять следы памяти — тесно связана с его способностью инициировать ориентировочные реакции.
Огромный объем экспериментальных фактов накоплен в отношении связи гиппокампа с ориентировочным рефлексом. В вентральном гиппокампе найдены «нейроны новизны» двух типов: с возбуждающими и тормозными реакциями (Виноградова О.С., 1975). Они обнаруживают все свойства ориентировочного рефлекса — при повторении одного и того же стимула их реакция угасает, она может быть расторможена экстрараздражителем или длительной паузой в применении стимула либо изменением его параметров.
Причастность гиппокампа к реакции новизны получила подтверждение и в поведенческих опытах с регистрацией гиппокам-пального тета-ритма. У крыс во время принюхивания, исследования обстановки тета-ритм усиливается, и исчезает параллельно с ослаблением ориентировочного рефлекса или ориентировочно-исследовательского поведения.
По данным О.С. Виноградовой, пластичные перестройки в гиппокампе при привыкании локализованы на синапсах, образуемыми мшистыми волокнами зубчатой фасции на пирамидах гиппо-. кампа в САЗ и СА4. Механизм привыкания можно отключить с помощью введения в зубчатую фасцию кролика соответствующих антител, предварительно выработанных у другого животного. Антитела блокируют функцию мшистых волокон, что и приводит к исчезновению привыкания у нейронов гиппокампа.
Как возникает гиппокампальный тета-ритм? Сенсорный сигнал достигает нейронов гиппокампа в СА1 и СА2 через энтори-альную кору, а затем через зубчатую фасцию и нейронов САЗ и СА4. Гиппокамп связан двусторонними связями с перегородкой (септумом). Реакция гиппокампа активирует латеральный септум и через него ретикулярную формацию (РФ), которая в свою очередь стимулирует медиальный септум, инициирующий генерацию
iпа-ритма. Появление гиппокампального тета-ритма связано с по-яилением реакций у нейронов новизны. С повторением стимула ричвивается привыкание за счет подключения тормозной функции чубчатой фасции, куда, так же как и в гиппокамп, приходит сенсорный сигнал из энториальной коры.
Привыкание в гиппокампе специфично по отношению к повторяющему стимулу. Это обусловлено специфичностью процесса долговременной потенциации и долговременной депрессии, которая также развивается на нейронах гиппокампа (см. раздел «Научение»).
Хорошо известен эффект растормаживания ранее приобретенных условных связей под влиянием действия экстрараздражителя. Гло связывают с возникновением ориентировочного рефлекса. 11олагают, что во время ориентировочной реакции гиппокамп актуализирует следы памяти, что позволяет ее гибко использовать в поведении.
По-видимому, гиппокамп непричастен к формированию ни декларативной, ни процедурной памяти, а только к манипуляции следами памяти. Гиппокамп, скорее, менеджер долговременной памяти.
16. Виды научения и нейронные феномены пластичности ….
Существует много разновидностей научения. Они могут быть разделены на несколько групп. Наиболее часто выделяют простое научение, к которому относят привыкание, сенситизацию, ассоциативное научение, включающее выработку классического условного рефлекса, инструментального (или оперантного) рефлекса, одномоментное обучение (на аверсивном подкреплении), и сложное научение (импринтинг, латентное обучение, обучение на основе подражания, когнитивное обучение: формирование декларативной памяти).
Привыкание как простейшая форма научения выражается в ослаблении поведенческой реакции при повторных предъявлениях стимула. От утомления и истощения привыкание отличается тем, что реакцию вновь можно вызвать простым изменением стимула.
Э. Кэндел и его сотрудники исследовали привыкание оборонительной реакции у аплизии. Защитная реакция морского моллюска в виде втягивания сифона и жабры вызывалась тактильным раздражением этих органов струей холодной воды. Многократное применение такой стимуляции уменьшало величину рефлекторной реакции втягивания. Эффект ослабления реакции мог быть кратковременным, но при достаточно большом числе повторений снижение реактивности оборонительной реакции могло сохраняться в течение нескольких недель (долговременное привыкание).
Ярким примером поведенческого привыкания является угасание безусловного ориентировочного рефлекса (или его отдельных компонентов). С повторением стимула теряется новизна, что и приводит к привыканию. Привыкание в системе ориентировочного рефлекса получило название негативного научения, состоящего в том, что стимул по мере его повторения теряет способность вызывать ту реакцию, которую он ранее вызывал. Привыкание или угасание ориентировочного рефлекса связывают с формированием «нервной модели стимула» — его многомерной энграммы, которая и тормозит систему активации ориентировочного рефлекса (см. главу «Внимание»).
Сенситизация— другая форма простейшего научения, выражающаяся в усилении рефлекторной реакции под влиянием сильного или повреждающего постороннего стимула. Сенситизация — это не просто противоположность привыкания. Она является результатом активации модулирующей системы мозга, возникшей на сильный побочный раздражитель. Усиление рефлекса вызвано изменением функционального состояния организма.
Нейронные феномены пластичности
I
Пластичность— фундаментальное свойство клетки, которое проявляется в относительно устойчивых модификациях реакций нейрона и во внутриклеточных его преобразованиях, обеспечивающих изменение эффективности и направленности межнейронных связей.
Свойство пластичности нейрона лежит в основе процессов научения и памяти целостного организма, проявляющихся на поведенческом уровне. Выделяют несколько основных феноменов пластичности: привыкание, сенситизацию, клеточные аналоги ассоциативного обучения, явления долговременной потенциации и долговременной депрессии, пластичность пейсмекерного механизма клетки.
Привыкание нейрона выражается в постепенном ослаблении его реакции на повторяющийся раздражитель. Восстановление реакции происходит в результате изменения стимула или применения нового, а также после прекращения привычной стимуляции. Время восстановления реакции (секунды — недели) зависит от стимуляции и изучаемого объекта.
Сенситизация нейрона— временное усиление его реакции или появление ее на ранее неэффективный стимул, возникающее в результате какого-либо сильного воздействия (например, электрического тока). Время ее сохранения — от нескольких секунд до дней и недель. Сенситизацию связывают с активацией модулирующих нейронов, вызываемой сильным биологически значимым воздействием.
Долговременная потенциация (long-term-potentiation, LTP) впер-ные описана для нейронов гиппокампа. В 1973 г. Т. Блис и Т. Лемо, работая в лаборатории П. Андерсена в Осло (Норвегия), впервые показали, что нейроны гиппокампа обладают выраженными пластическими свойствами, нужными для обучения. Применив высокочастотную электрическую стимуляцию, чтобы вызвать короткую серию ПД в пресинаптическом нейроне, они получили эффект усиления ответов постсинаптического нейрона на последующие одиночные импульсы, приходящие от пресинапти-ческой клетки. Эффект мог сохраняться в течение часов—недель. Они назвали такое усиление синаптической проводимости долговременной потенциацией, предположив, что постсинаптическая клетка выделяет ретроградный мессенджер, который способен проникать через мембрану пресинаптической клетки. Этим мессенджером, возможно, является окись азота, которая определяет пресинаптическое облегчение у гиппокампальных нейронов млекопитающих.
Долговременная потенциация (ДВП) в гиппокампе удовлет-норяет выдвинутому Д. Хеббом критерию обучения — совпадениюпресинаптической и постсинаптической активности нейрона. Этот принцип был сформулирован Д. Хеббом в 1949 г. в книге «Организация поведения» в качестве условия, необходимого для образования новых нейронных ансамблей в процессе обучения.
В опытах на срезах гиппокампа П. Андерсен и его сотрудники (Andersen P. et al., 1977) показали, что долговременная потенциа-ция обусловлена усиленным выделением медиатора из пресинап-тических окончаний. С помощью электронного микроскопа была подтверждена связь долговременной потенциации с увеличением шипиков на дендритах нейронов гиппокампа и изменением структуры синапсов.
Позже ДВП была обнаружена в других структурах мозга — миндалине, мозжечке. ДВП относится к определенному синаптичес-кому контакту. Она выявлена у идентифицированного синапса в простой нервной системе моллюска.
Долговременную потенциацию (ДВП) можно обнаружить по увеличению амплитуды и крутизны ВПСП, повышению вероятности генерации ПД, а также по увеличению числа спайков на один тестирующий стимул. Долговременная потенциация часто используется как удобная модель для изучения механизмов научения.
Долговременная депрессия (long-term-depression, LTD ) состоит в длительном снижении проводимости через синапс. Сравнительно недавно долговременная депрессия (ДВД) обнаружена в коре мозжечка у клеток Пуркинье. Ее можно вызвать совместной низкочастотной стимуляцией (1—4 Гц) двух входов к клеткам Пуркинье: афферентного входа через параллельные волокна к ее дендри-там и входа от лазящих волокон из нижней оливы, посылающей сигналы о совершаемых двигательных рефлексах.
Сигнал от лазящего волокна увеличивает приток ионов Са2+ к клеткам Пуркинье. Это вызывает в нейроне каскад преобразований, где главная роль принадлежит процессам фосфорилирования-дефос-форилирования. Предполагают, что ДВД возникает вследствие того, что ионы Са2+ активируют белок фосфатазу, который обеспечивает дефосфорилирование рецепторов на дендритах клеток Пуркинье, снижая тем самым их синаптическую проводимость. Не исключается возможность и второго механизма торможения клеток Пуркинье через долговременную потенциацию тормозного входа к ним от корзин-чатых интернейронов. ДВД возникает только на тех синапсах параллельных волокон к клеткам Пуркинье, возбуждение которых по времени совпадает с сигналом, приходящим из нижней оливы.
Таким образом, долговременная депрессия в клетках Пуркинье — тот механизм пластических изменений в нейроне, благодаря которому в мозжечке формируются условные рефлексы.
Клеточные аналоги ассоциативного обучения можно получить во многих структурах мозга. Так, 40% нейронов гигтпокампа способны к обучению (Котляр Б.И., 1986). Процедура формирования условного рефлекса у нейрона требует, чтобы к нему последовательно, с небольшой задержкой приходили сигналы от условного и безусловного стимулов. Условным стимулом может быть любое сенсорное или электрическое раздражение, которое вызывает лишь подпороговую реакцию (ВПСП). Безусловный раздражитель должен вызывать у нейрона спайковые разряды (ПД). Под влиянием сочетания «индифферентного» раздражителя с безусловным нейрон, способный к ассоциативному обучению, начинает отвечать на условный стимул реакцией, которая ранее возникала только на безусловное раздражение.
У отдельного нейрона можно выработать не только классический условный рефлекс, но и инструментальный. Впервые это показали исследователи из Мичигана Джеймс и Мариана Олдс в 1961 г., а затем и другие исследователи. В опытах Э. Фетца и М.Э. Бейкер (Fetz E., Baker M., 1973) с помощью вживленных электродов регистрировали активность нейронов в двигательной коре у бодрствующей обезьяны. Когда частота ПД случайно превышала некоторый уровень, шкала прибора перед обезьяной освещалась и животное получало положительное подкрепление — несколько капель фруктового сока. В результате такой процедуры активность нейрона возрастала до того уровня, который подкреплялся. С отменой подкрепления частота ПД возвращалась к исходному, фоновому значению (угасание инструментального рефлекса). Параллельная регистрация ЭМГ и механограммы мышцы задней стопы показала, что возрастание частоты спайков нейрона двигательной коры, вызванное обучением, несколько опережает начало электрической и механической активности мышцы. Это позволило утверждать, что ' инструментальный условный рефлекс был выработан у нейрона, который и запускал двигательную реакцию, а не наоборот.
Многие исследования были посвящены формированию аналогов условного рефлекса на различных модельных системах: срезах мозга, изолированной нервной системе моллюска, синапти-чески связанных нейронах. Особенно впечатляющими оказались результаты изучения клеточных механизмов пластичности на полностью изолированной соме нейрона. Этот препарат впервые был получен у аплизии и применен Р. Баумгартеном с соавторами (Baumgarten R.I. et al., 1971).
На виноградной улитке ассоциативное обучение у полностью изолированного нейрона было исследовано Т.Н. Греченко (1979). Сому нейрона, изолированную от отростков, получают при помо-щи обработки ганглиев виноградной улитки протеомитическим ферментом — трипсином с последующим выдерживанием в термостате при температуре 37—38°С в течение 15—20 мин. После такой процедуры нейроны лишаются отростков. Изолированный нейрон может состоять только из сомы или из сомы с культей аксона. Он сохраняет присущую ему фоновую ативность, тип хемочувствительности.
В качестве условного стимула (УС) использовались микроаппликация медиаторов (ацетилхолина или серотонина) или прямая деполяризация током через внутриклеточный электрод. Все условные стимулы вызывали лишь ВПСП и были подпороговыми для ПД. В качестве безусловного стимула (БС) применялся раздражитель большой силы, который вызывал устойчивую генерацию ПД. Интервал времени между УС и БС 5-500 мс, интервал между парами стимулов — от 40 с до 3 мин. Через 10—15 сочетаний условный стимул начинал вызывать реакцию, характерную для БС. Условный ответ у разных нейронов сохранялся в течение 5—40 мин. Дополнительное обучение увеличивало время сохранения следового эффекта. Неподкрепление вело к постепенному угасанию условной реакции. 80% нейронов (из 503) продемонстрировали феномен отсроченного обучения. Условный ответ после прекращения процедуры обучения появлялся не сразу, а спустя некоторое время. Интервал от момента прекращения обучения до достижения максимального следового эффекта для разных нейронов составлял от 1-3 до 60 мин и более (Греченко Т.Н., Соколов Е.Н., 1987).
Был выявлен локальный характер обучения на одном нейроне. Условные и безусловные раздражители действуют через отдельные локусы мембраны, и условный ответ формируется только между теми участками мембраны, к которым направлены УС и БС. Поэтому на одном изолированном нейроне можно выработать параллельно несколько различных условных рефлексов, включая диф-ференцировку, а также получить их селективное угасание. Эти результаты невозможно объяснить механизмом пресинаптической
пластичности: полностью изолированные нейроны лишены каких-либо контактов с другими нейронами. Они указывают на роль постсинаптического и, вероятно, молекулярных механизмов в становлении следов памяти.
19. Различные определения эмоций, классификация, функции.
Обычно эмоцию определяют как особый вид психических процессов, которые выражают переживание человеком его отношения к окружающему миру и самому себе. Особенность эмоций состоит в том, что они в зависимости от потребностей субъекта непосредственно оценивают значимость действующих на индивид объектов и ситуаций. Эмоции выполняют функции связи между действительностью и потребностями.
Однако, как отмечает П.В. Симонов (1997), несмотря на успехи, достигнутые в изучении мозговых механизмов психических процессов, в настоящее время не существует единой общепринятой научной теории эмоций. Нет также точных данных о том, в каких центрах и каким образом эти эмоции возникают и каков их нервный субстрат.
По классификации эмоциональных явлений А.Н. Леонтьева выделяется три вида эмоциональных процессов: аффекты, собственно эмоции и чувства.
Аффекты — это сильные и относительно кратковременные эмоциональные переживания, сопровождающиеся резко выраженными двигательными и висцеральными проявлениями. У человека аффекты вызываются как биологически значимыми факторами, затрагивающими его физическое существование, так и социальными, например мнением руководителя, его отрицательной оценкой, принятыми санкциями. Отличительной особенностью аффектов является то, что они возникают в ответ на уже фактически наступившую ситуацию.
Собственно эмоции в отличие от аффектов представляют собой более длительно текущее состояние, иногда лишь слабо проявля-
164
ющееся во внешнем поведении. Они выражают оценочное лично-с гное отношение к складывающейся или возможной ситуации, поэтому способны в отличие от аффектов предвосхищать ситуации и события, которые реально еще не наступили. Собственно •моции возникают на основе представлений о пережитых или вд-ображаемых ситуациях.
"'•"•
Третий вид эмоциональных процессов — это так называемые предметные чувства. Они возникают как специфическое обобщение эмоций и связаны с представлением или идеей о некотором объекте, конкретном или отвлеченном (например, чувство любви к человеку, к родине, чувство ненависти к врагу и т.д.). Предметные чувства выражают устойчивые эмоциональные отношения.
По П.В. Симонову (1981), чувства — это эмоции, которые возникают на базе социальных и духовных потребностей. Учитывая роль вероятности удовлетворения потребности в происхождении •>моций, он рассматривает тревогу как реакцию на низкую вероятность избегания нежелательного воздействия. В отличие от тревоги фрустрация — это реакция на низкую вероятность достижения желаемой цели.
Особое место среди эмоциональных явлений занимают так называемые общие ощущения. Так, П. Милнер считает, что, хотя и принято отличать эмоции (гнев, страх, радость и т.п.) от так назы-наемых общих ощущений (голод, жажда и т.д.), тем не менее в них обнаруживается много общего и их разделение достаточно условно. Одна из причин, по которой их различают, — разная степень спязи субъективных переживаний с возбуждением рецепторов. Так, переживание жары, боли субъективно связывается с возбуждением определенных рецепторов (температурных, болевых). На этом основании подобные состояния обычно и обозначают как ощущения. Состояние же страха, гнева трудно связать с возбуждением каких-либо рецепторных поверхностей, поэтому их относят к эмоциям. Другая причина, по которой эмоции противопоставляются общим ощущениям, состоит в нерегулярном их появлении. Эмоции часто возникают спонтанно и зависят от случайных внешних факторов, тогда как голод, жажда, половое влечение следуют с определенными интервалами.
В настоящее время внимание исследователей привлекает еще одна категория эмоциональных явлений — настроение. Что такое настроение? У настроения нет специфического целевого объекта, как у эмоций, так же как и каких-либо специфических реакций. Следовательно, оно менее специфично, чем эмоция. К тому же субъективные переживания, связанные с настроением, по сравнению с эмоциями менее интенсивны.
Исследователи, ориентированные на когнитивную теорию эмоций, понимают настроение как некоторое состояние чувствования, возникающее в результате автоматически протекающих когнитивных процессов, о которых мы не осведомлены. Поэтому мы часто не осознаем, что находимся под впечатлением определенного настроения. Таким образом, настроение действует как контекст, подспудно трансформируя наши реакции на события.
По определению А.Изена (Isen А.), настроение— это течение или поток гедонически ориентированных идей, мыслей и образов, извлекаемых из памяти. Их объединяет общий гедонический тон (положительный или отрицательный).
К. Прибрам (Pribram К.) подчеркивает оценочную функцию настроения, рассматривая его как некий мониторинг окружающих жизненных обстоятельств. Функция настроения — информировать об общем состоянии и потребностях индивида. Настроение стимулирует субъекта к поведению, которое улучшило бы его гедонический тон. Поэтому одна из функций настроения — это его способность к саморегуляции.
Многочисленные экспериментальные данные говорят о том, что настроение — результат действия как реальных, так и воображаемых событий или информации, извлекаемой из эмоциональной памяти. Клинические исследования указывают на ключевую роль гормональных и биохимических факторов в генезе настроения.
Когда настроение достигает некоторого порога, оно привлекает внимание. Попав в фокус внимания, оно становится осознанным и может быть объяснено, включая его причины. Это может послужить толчком к трансформации настроения в эмоцию. Настроение, как осознаваемое, так и неосознанное, влияет на аффективные и когнитивные процессы, а также на поведение. Более сильное влияние оказывает осознанное настроение. При этом интенсивность аффекта зависит от уровня внимания, которое будет уделено настроению.
Одно и то же явление одновременно может вызывать как эмоцию, так и настроение, которые могут сосуществовать, влияя друг на друга. Если какая-либо эмоциональная реакция развивается во времени стремительно, то созданное ею настроение может сохраняться часами, днями и неделями.
Действия человека небеспристрастны. Поэтому эмоция как субъективное переживание присутствует в составе каждой деятельности, каждого рефлекса (Соколов Е.Н., Данилова Н.Н., 1975). В структуре поведения, как в функциональной системе, эмоции принадлежит ключевая роль. Выделяют ведущие и ситуативные эмоции (Вилюнас В.К., 1986). Они связаны с разными фазами поведе-
166
кия. Ведущие эмоции сигнализируют человеку о неудовлетворенности его потребностей и побуждают к поиску целевого объекта, < гимулируя определенное поведение. Побудительной силой обла-/шст и эмоциональная память об успешных действиях в прошлом, исправленных на удовлетворение аналогичной потребности. Положительные эмоции, связанные с удовлетворением потребностей, пключаются в акцептор результатов действия. С их помощью цель и <• родства удовлетворения потребности конкретизируются (Судаков К.В., 1997). В структуре поведенческого акта формирование акцептора результатов действия (цели) и эфферентного синтеза (предполагаемых способов действия) опосредовано содержанием иодущих эмоций. Ситуативные эмоции, возникающие в результате опенок отдельных этапов или поведения в целом, побуждают 1'убъекта действовать либо в прежнем направлении, либо менять поведение, его тактику, способы достижения цели.
ФУНКЦИИ ЭМОЦИЙ
Исследователи, отвечая на вопрос о том, какую роль играют >моции в жизнедеятельности живых существ, выделяют несколько регуляторных функций эмоций: отражательную (оценочную), побуждающую, подкрепляющую, переключательную, коммуникативную.
Отражательная функция эмоций выражается в обобщенной оценке событий. Эмоции охватывают весь организм и представляют почти мгновенную и интегральную оценку поведения в целом, что позволяет определить полезность и вредность воздействующих на человека факторов еще до того, как будет определена локализация вредного воздействия. Примером может служить поведение человека, получившего травму конечности. Ориентируясь на боль, он немедленно находит такое положение, которое уменьшает болевые ощущения.
Эмоция как особое внутреннее состояние и субъективное переживание выполняет функцию оценки обстоятельств ситуации на основе возникшей потребности и интуитивного представления о возможностях ее удовлетворения. Эмоциональная оценка отличается от осознанных когнитивных оценочных операций ума, она выполняется на чувственном уровне. В самом деле, мы никогда не оцениваем истинную пищевую потребность в количестве белков, жиров, углеводов, витаминов, солей и т.д. Возникновения чувства голода уже достаточно.
Оценочная, или отражательная, функция эмоции непосредственно связана с ее побуждающей функцией. Согласно Оксфорд-
скому словарю английского языка, слово «эмоция» произошло от французского глагола «mouvoir», означающего «приводить в движение». Его начали употреблять в XVII
в., говоря о чувствах (радость, желание, боль и т.д.) в отличие от мыслей. С.Л. Рубинштейн отмечал, что эмоция уже в себе самой заключает влечение, желание, стремление, направленное к предмету или от него. Эмоция выявляет зону поиска, где будет найдено решение задачи, удовлетворение потребности. Эмоциональное переживание содержит образ предмета удовлетворения потребности и свое пристрастное отношение к нему, что и побуждает человека к действию. Два класса эмоций — ведущие и ситуативные, или производные (Вилюнас В.К., 1986), — несут функцию побуждения. Ведущее эмоциональное переживание, направленное на предмет — цель поведения, удовлетворяющего потребность, инициирует само адаптивное поведение. Ситуативные эмоциональные переживания, возникающие в результате оценок результатов отдельных этапов поведения, также побуждают субъект действовать в прежнем направлении или менять тактику поведения и само поведение, средства достижения цели.
Анализируя ситуативные эмоции у человека, В.К. Вилюнас (1986) выделяет класс эмоций успеха—неуспеха с тремя подгруппами: 1) констатируемый успех—неуспех; 2) предвосхищающий успех—неуспех; 3) обобщенный успех—неуспех. Эмоции, констатирующие успех—неуспех, отвечают за смену проб в поведении, обобщенная эмоция успеха—неуспеха возникает в результате оценки деятельности в целом. Предвосхищающие эмоции успеха—неуспеха формируются на основе констатирующих в результате ассоциации их с деталями ситуации. При повторном столкновении с ситуацией эти эмоции позволяют предвосхищать, предвидеть события и побуждают субъекта действовать в определенном направлении.
Предвосхищающие эмоции успешно изучались в составе мыслительной деятельности при решении творческих задач (шахматных). Эмоции предвосхищения связаны с появлением переживания догадки, идеи решения, которая еще не вербализована.
П.В. Симонов (1987) выделяет у эмоций подкрепляющую функцию. Известно, что эмоции принимают самое непосредственное участие в процессах обучения и памяти. Значимые события, вызывающие эмоциональные реакции, быстрее и надолго запечатлеваются в памяти. Так, у сытой кошки нельзя выработать условные пищевые рефлексы. Для успешного обучения необходимо наличие мотивационного возбуждения, в данном случае отражающегося в чувстве голода. Однако соединения индифферентного раздражителя с голодовым возбуждением еще недостаточно для выработки условных пищевых рефлексов. Требуется третий компонент — воз-
действие фактора, способного удовлетворить существующую потребность, т.е. пищи. В опытах Т.Н. Ониани, который сочетал внешний стимул с электрическим раздражением лимбических структур мозга, вызывающим у сытой кошки потребность в еде, удалось выработать только условную реакцию избегания и страха. А условг ные пищедобывательные рефлексы не формировались. Главная причина в том, что электрическая стимуляция лимбической структуры, использованная в качестве подкрепления, не содержала награды — удовлетворения потребности. :
Не удается также выработать условнорефлекторный поиск пищи, если сочетать индифферентные обстановочные сигналы с состоянием, вызываемым пищевой депривацией. У такого животного на обстановку эксперимента вырабатывается не поисковое пищевое поведение, а реакция страха и избегания, т.е. индифферентный стимул связывается с реакцией избегания, которой животное реагирует на ситуацию длительного голодания, так как эта реакция уменьшает страх.
Таким образом, реальным подкреплением для выработки условного рефлекса (классического и инструментального) является награда. Наградой голодному животному может быть пища. Само болевое раздражение не является наградой, ее дает лишь освобождение, избегание наказания. Получение награды ассоциируется с возникновением положительных эмоций. Поэтому «только интеграция голодового возбуждения с возбуждением от фактора, способного удовлетворить данную потребность, т.е. механизм, генерирующий положительную эмоцию, обеспечивает выработку условного рефлекса» (Симонов П.В., 1987. С. 79).
Подкрепляющая функция эмоций наиболее успешно была исследована на экспериментальной модели «эмоционального резонанса», предложенной П.В. Симоновым. Было обнаружено, что эмоциональные состояния животных, подвергнутых воздействию электрокожного раздражения, влияют на эмоциональные реакции других животных. Эта модель воспроизводит типичную для социальных взаимоотношений ситуацию распространения отрицательных эмоциональных состояний в сообществе и позволяет изучать функции эмоций в наиболее чистом виде без непосредственного действия болевых раздражителей. В опытах Л.А. Преображенской, в которых собака-жертва подвергалась наказанию электрическим током на глазах у собаки-наблюдателя, у последней возрастала частота сердцебиения и увеличивалась синхронизация гиппокам-пального тета-ритма. Эти реакции указывают на появление у животного состояния негативного эмоционального напряжения. В таких условиях собака-наблюдатель способна выработать избегатель-
ный инструментальный рефлекс (в виде подъема лапы), прекращающий подачу тока собаке-жертве. При этом у нее снижается ЧСС и устраняется гиппокампальный тета-ритм, что означает исчезновение отрицательного эмоционального состояния. Следовательно, наградой, на которой и вырабатывается данная инструментальная реакция, является избавление от отрицательного эмоционального напряжения.
Переключательная функция эмоций состоит в том, что они часто побуждают, человека к изменению своего поведения.
В естественных условиях поведение человека и животных определяется многими потребностями разного уровня. Их взаимодействие выражается в конкуренции'мотивов, которые проявляются в эмоциональных переживаниях. Оценки через эмоциональные переживания обладают побуждающей силой и могут определять выбор поведения. Как было показано ранее на примере интеллектуальных эмоций, «эмоциональное предчувствие решения задачи» побуждает к завершению действий в выбранной зоне поиска, а «предчувствие противоречия» — к изменению этой зоны. По существу, эмоции успеха—неуспеха обладают способностью побуждать человека к переключению от одних действий к другим.
Переключательная функция эмоций наиболее ярко обнаруживается в экстремальных ситуациях, когда возникает борьба между естественным для человека инстинктом самосохранения и социальной потребностью следовать определенной этической норме. Конфликт потребностей переживается в форме борьбы между страхом и чувством долга, страхом и стыдом. Исход зависит от силы побуждений, от личностных установок субъекта.
Важной функцией эмоций является коммуникативная функция. Мимика, жесты, позы, выразительные вздохи, изменение интонации являются «языком человеческих чувств» и позволяют человеку передавать свои переживания другим людям, информировать их о своем отношении к явлениям, объектам и т.д.
Сравнивая проявление эмоций у человека и у домашних животных, он пришел к заключению, что движения мышц, при помощи которых человек выражает свои эмоции (радость и печаль, гнев и страх, интерес и отвращение, стыд и презрение), произошли от аналогичных двигательных актов наших предков. Согласно Ч. Дарвину полярные эмоции выражаются противоположными движениями.
Современные исследователи согласны с Ч. Дарвином в том, что экспрессивные движения и особенно мимика выполняют важную адаптивную функцию. В ходе эволюции мимические сигналы развились в систему, позволяющую передавать информацию о состоянии и «намерениях» индивида. Так, выражение испуга сигнализирует об опасности и о намерении индивида спасаться бегством, тогда как эмоция гнева воспринимается как выражение намерения вступить в борьбу. С помощью «языка чувств» регулируются социальные отношения у животных. Например, сигналы угрозы, исходящие от вожака, повышают бдительность или вызывают поведение «подчинения» у других особей.
Существуют генетически заданные универсальные комплексы поведенческих реакций, с помощью которых выражаются основные фундаментальные эмоции.
20.
Потребностно-информационная теория эмоций Симонова.
К некогнитивным факторам, вызывающим эмоции, П.В. Симонов относит потребность и оценку возможности (вероятности) се удовлетворения, которую субъект совершает непроизвольно и часто неосознанно.
Им разработана потребностно-информационная теория эмоций, согласно которой правило возникновения эмоций выражается в следующей структурной формуле:
Э = [П, (Ин - Ис), ...],
где Э — эмоция, ее степень, качество и знак; П — сила и качество актуальной потребности; (Ин — Ис) — оценка вероятности удовлетворения потребности на основе врожденного и онтогенетического опыта; Ин — информация о средствах и времени, прагматически необходимых для удовлетворения потребности; Ис — информация о существующих средствах и времени, которыми реально располагает субъект в данной ситуации.
Согласно информационной теории эмоций положительные эмоции возникают в ситуации избытка прагматической информации по сравнению с ранее существующим прогнозом (при «мгновенном срезе») или в ситуации возрастания вероятности достижения цели (если генез эмоций рассматривать в его динамике). Отрицательные эмоции представляют реакцию на дефицит информации или на падение вероятности достижения цели в процессе деятельности субъекта. Многообразие эмоций согласно концепции определяется многообразием потребностей. Эмоций столько сколько потребностей. У человека витальные потребности (голод, жажда и др.) дополнены социальными и идеальными потребностями. Среди последних выделяют идеологические потребности — производные от социальных и идеальных. Объединение витальных с социальными потребностями создает этнические потребности, которые также могут быть источником эмоций. Социальные потребности вместе с потребностью для других формируют духовные потребности (совесть). Классификация потребностей по П.В. Симонову показана на рис. 39.
В 1984г. американские исследователи Д. Прайс и Дж. Баррелл (Price D., Barrell
Y., 1984) провели психологические исследования с целью проверки потребностно-информационной теории эмоций. Для этого они предложили испытуемым мысленно представить событие, которое у них было связано с эмоциональными переживаниями. Затем на специальных шкалах испытуемый отмечал силу эмоционального переживания, предполагаемую вероятность достижения цели и силу желания разрешить проблему, которая вызвала эмоции. Количественная обработка полученных данных подтвердила зависимость трех переменных: эмоции, силы потребности и вероятности ее удовлетворения. Исследователи назвали эту зависимость общим законом человеческих эмоций.
В 1988 г. канадский психолог Ж. Годфруа (1992) воспроизвел зависимость эмоции от количества информации, выразив это следующим образом: «Эмоция = Необходимая информация — Имеющаяся информация».
П.В. Симонов полагает, что функция прогнозирования вероятности удовлетворения потребности (вероятности подкрепления) поделена между двумя информационными структурами мозга — фронтальными отделами новой коры и гиппокампом. Фронтальная кора ориентирует поведение на высоковероятные события в отличие от гиппокампа, реагирующего на сигналы маловероятных событий. Известно, что чем выше у интактного животного вероятность подкрепления, тем быстрее вырабатывается условный рефлекс. После разрушения фронтальной коры формирование пище-iiom условного рефлекса при низкой вероятности подкрепления заметно ускоряется по сравнению с интактным животным. Этот результат объясняют тем, что после разрушения одного из двух cwiokob информационной системы, состоящей из фронтальной коры и гиппокампа, мозг перестает дифференцировать высоко- и низковероятные события. Поэтому сигналы, подкрепляемые с разной пероятностью, становятся одинаково эффективными.
Гиппокамп выделяет низковероятные события, обеспечивает выработку условного рефлекса на низковероятном подкреплении условного сигнала. Крысы после удаления гиппокампа неспособны вырабатывать условные рефлексы при низковероятном подкреплении (в 25 и 30% случаев). Хотя при подкреплении пищей в 100 и 50% случаев гиппокампэктомированные крысы и отстают от иктактных, но все же с задачей справляются. Следует сказать, что необходимость различать высокую и низкую вероятность подкрепления всегда является трудной задачей даже для интактного животного и требует много времени.
Для проверки функции гиппокампа, состоящей в выделении низковероятных событий, исследователи использовали экспериментальную модель Э.А. Асратяна в виде выработки условнореф-лекторного переключения разнородных условных рефлексов, когда один и тот же сигнал (звук) утром подкреплялся пищей с высокой вероятностью и болевым раздражителем с малой вероятностью, а вечером отношения менялись на противоположные. В течение 10 дней интактная крыса безуспешно пыталась выработать ус-ловнорефлекторное переключение. Однако после двустороннего удаления гиппокампа у нее сформировалось устойчивое условно-рефлекторное переключение и она реагировала на один и тот же стимул утром пищевым, а вечером оборонительным рефлексом в соответствии с тем, какое подкрепление наиболее часто применялось. Хирургическая операция не только облегчала выработку рефлексов, но и устраняла признаки эмоционального напряжения (по показателю частоты сердечных сокращений), которое обычно наблюдается в таких опытах. Результаты этих исследований объясняют тем, что оперированные животные не измеряют
(игнорируют) низковероятные события (подкрепление), что упрощает их задачу.
Крысы ориентируются лишь на высоковероятное подкрепление пищей утром и
высоковероятное наказание вечером.
21. Лицевая экспрессия.
Люди обычно обмениваются многими невербальными сигналами, используя интонацию голоса, лицевую экспрессию, контакты глазами, жесты. С помощью этих средств передается важная информация. Сходным образом обмениваются информацией с сородичами животные в стае, используя выразительные движения, особые звуки. Ч. Дарвин (1872) был первым, кто предположил, что лицевая экспрессия играет особенно важную роль в кооперации, объединении членов сообщества, так как позволяет 'сообщать об эмоциях и намерениях действовать. Наиболее важным каналом эмоционального, невербального общения является зрительный, через который принимается информация, содержащаяся в выразительных жестах и экспрессивных реакциях лица. То, что лицо — наиболее важный источник информации в процессе общения, подтверждается исследованиями, выполненными психофизическими методами. Для сравнения возможностей зрительного и слухового невербальных каналов в передаче информации об эмоциях У. Хелл, А. Керрес и К. Шерер (Hells U., Karras
A., Scherer K.R., 1988) применили факторный анализ к результатам ранжирования видеокадров (лица человека, изображающего улыбку или эмоцию гнева), а также к записям человеческого голоса, переживающего те же эмоции (рис. 40). Для обеих модальностей суммарно авторы получили двухфакторное решение: 45% информации об эмоциях передается зрительными сигналами и 17,6% — слуховыми.
Роль мимики по сравнению с позой, жестом, с фило- и онтогенетическим развитием возрастает. У птиц, в частности у домашних цыплят, положение головы несет важную информацию о социальном статусе особи. В ряду позвоночных лицевая мускулатура претерпевает изменения от полного отсутствия у низших животных до хорошо развитой системы лицевых мышц у приматов и становится предельно дифференцированной у человека. У макаки резуса наиболее важным компонентом в передаче информации является пантомимика, в частности поза, у человекообразной обезьяны мимика становится более независимой от позы и локомо-
i>;м(жирования кадров отдельно для записи голоса человека, переживающею радость и гнев, и отдельно для видеозаписей лица человека, выра-•|тощего эмоцию радости и гнева, которые сопровождались текстом, про-п июсимым без интонации и эмоций. Факторизация только зрительных i
,|дров выделяет два фактора: лицевой экспрессии и текста. При фактори- • лции кадров с голосом получено три фактора: интонации, текста и каче-• пш голоса (с текстом). Видно, что через зрительный канал суммарно передается больше информации (о гневе и радости), чем через слуховой
канал (по Hells U. et al., 1988).
Л — лицо; Т — текст; И — интонация; Т+Л — текст и лицо; Т+К — текст и
качество голоса.
пни. Исследователями выделены различные выразительные двига-н-льные комплексы у макаки резуса, которые используются для н-монстрации угрозы, беспокойства, подчинения, испуга, миро-'иобия и важны для создания иерархии социального доминирования. У человека экспрессивные реакции лица приобретают само-i тигельное значение в передаче эмоциональных сигналов. В со-11 пильном поведении взаимодействие лиц играет огромную роль. У < мсциалистов оно получило даже специальное название — «пове-дение лицо к лицу». «Лицо — это центр передачи и приема социальных сигналов, которые являются решающими для развития индивида» (Изард К., 1980. С. 72).
В ходе эволюции мимические реакции развивались из движений — намерений, относящихся к подготовительной фазе деятельности (подготовка к нападению или, наоборот, к защите). Это предопределило одну из функций лицевой экспрессии — стать средством невербального общения, способом передачи информации о намерениях и состояниях человека, которые должны привлечь внимание другого субъекта.
Кросскультурные исследования распознавания лицевой экспрессии показали, что основные эмоции (гнев, страх, счастье, удивление, печаль, отвращение) одинаково проявляются и узнаются в обществах с различной культурой и традициями. Даже в условиях, затрудняющих наблюдение, например на большом удалении, проявляется универсальность механизма распознавания эмоций. Это доказывает, что лицевая экспрессия основных эмоций обеспечивается врожденными нейронными программами. Процент совпадения оценок лицевых эмоциональных паттернов у лиц разных национальностей — около 80%, несколько меньше он у японцев (65%).
Генетическая обусловленность реакций лицевых мышц, выражающих эмоциональные переживания, подтверждается ранним созреванием их двигательных комплексов. Все мышцы лица, необходимые для выражения эмоций, формируются у плода уже к 15—18-й неделе жизни. А к 20-й неделе у эмбриона можно наблюдать мимические реакции. К моменту рождения ребенка механизм лицевой экспрессии уже полностью сформирован и может быть использован в общении. На врожденный характер мимики указывает и ее сходство у слепого и зрячего младенцев. Но с возрастом у слепорожденного ребенка реактивность лицевых мышц угасает.
Лицо человека и даже его схематическое изображение — значимый стимул для новорожденного. Об этом можно судить по длительности его фиксации глазами, по частоте обращения внимания на него, по появлению вегетативного компонента ориентировочного рефлекса (снижению ЧСС). Ребенок предпочитает рассматривание человеческого лица любому другому стимулу (шахматной доске, изображению различных животных).
22. Корреляция активности лицевых мышц и эмоций. Кодирование и
декодирование л.э.
П. Экман и У. Фризен исследовали связь активности мышц лица с
различными эмоциями, когда испытуемые просматривали фильмы приятного (щенок,
играющий цветком, горилла в зоопарке, океан) и неприятного (травматический
инцидент в цеху) содержания. После просмотра каждого фильма испытуемые должны
были ранжировать свои эмоции по 9 шкалам (счастья, гнева, страха и др.). Была
обнаружена положительная корреляция переживания «счастья» только с активностью
большой скуловой мышцы (m. zygomaticus major). Чем сильнее активность этой мышцы, тем выше уровень
субъективной оценки переживаемого «счастья» при просмотре приятного фильма. При
этом активность других мышц лица и даже тех, которые принимают участие в улыбке,
не коррелировала с ощущением «счастья». Авторы пришли к выводу, что по
активности большой скуловой мышцы можно предсказывать появление положительного
эмоционального переживания. Чем выше была ее активность, тем о более
интенсивном позитивном переживании сообщал человек.
П. Экман и У. Фризен попытались найти паттерн двигательных реакций
лица, соответствующий искренней улыбке. Они исходили из идеи, впервые
высказанной Даченом (Duchenn), а затем
и Дарвиным о том, что улыбка радости морфологически должна отличаться от
других типов улыбок: социальной, политической, маскирующей. Исследовав
активность мышц лица во время различных улыбок, они установили, что искреннее
выражение счастья, радости сочетается с комбинированной активацией двух мышц:
большой скуловой мышцы и круговой мышцы глаза (m. orbicularis oculi), которая с трудом подчиняется волевому
контролю. Исследователи подтвердили, что большинство людей действительно
неспособно произвольно сокращать круговую мышцу глаз и что во время позитивных
переживаний обязательно активируются большая скуловая мышца, поднимающая
уголки губ кверху, к скулам, и наружная часть круговой мышцы глаза, которая
поднимает щеку и собирает кожу вовнутрь глазной впадины. Они назвали улыбку
искренней радости улыбкой Дачена (Duchene Smile). Только улыбка Дачена
положительно коррелирует с субъективным отчетом о переживании счастья и ростом
ЭЭГ-активации в левой фронтальной коре (признак появления положительной
эмоции). 10-месячный младе-нец часто реагирует улыбкой Дачена на приближение
матери и реже на приближение незнакомца. У взрослых юмор вызывает улыбку
Дачена. После успешного курса психотерапии у пациентов усиливается улыбка
Дачена, но не другие типы улыбок. У взрослого человека при просмотре забавного
фильма искренняя улыбка появляется чаще, чем другие типы улыбок (социальная,
политическая, маскирующая).
Лица, у которых обычно обнаруживается малая активность большой
скуловой мышцы, как правило, сообщают о меньших переживаниях счастья по
сравнению с теми, у кого активность этой мышцы усилена. Применение системы FACS показало, что во время отрицательных эмоций
(гнев, страх, отвращение, печаль) активируется около 41% всех мышц лица.
Субъекты, демонстрирующие низкую активность мышц лица во время просмотра
фильма с эпизодом, в котором рабочий получает увечье в цеху, характеризуются
более низкими рангами отрицательных переживаний по сравнению с теми, у кого
лицевая экспрессия более выражена. При этом лица, ареактивные (по ЭМГ) в
отношении отрицательных эмоциональных стимулов, во время просмотра «фильма ужаса»
имеют тенденцию к усилению активности тех мышц, которые связаны с переживанием
«счастья» или нейтральных состояний. Кроме того, выделены три мышцы,
активирующиеся при отвращении: одна поднимает центральную часть верхней губы,
другая поднимает и напрягает крылья носа, а третья усугубляет носогуб-ную
складку. Их активность хорошо коррелирует с переживанием отвращения.
В работах Г. Швартца (Schwartz
G.E.) и его коллег из Йельского университета штата Коннектикут была изучена
связь депрессии с особенностями активности лицевых мышц. У больных отмечены
ослабление паттернов ЭМГ-активности (по m.zygomaticus), которые связаны с
эмоцией «счастья», а также усиление активности мышц, реагирующих на переживание
«печали». По наблюдению авторов, эффективное лечение таких больных
(медикаментозными средствами или с применением плацебо) сопровождается общей
нормализацией мышечной активности лица. Отмечено также усиление активности
мышц, возбуждающихся при переживании «счастья». Установление связи депрессии с
активностью лицевых мышц чрезвычайно важно для выявления в нормальной популяции
групп риска — лиц, склонных к развитию депрессии.
Женщины в целом показывают более интенсивную лицевую экспрессию по
сравнению с мужчинами. При этом у них мышца нахмуривания более активирована как
во время представления эмоциональных ситуаций с отрицательным знаком, так и в
со-
стоянии покоя. Эти результаты согласуются с тем фактом, что
женщины
составляют основной контингент пациентов с депрессией.
Знак эмоционального переживания можно контролировать по
соотношению активности двух мышц: большой скуловой и мышц наххмуривания (m.corrugator), так как активность
скуловой мышцы положительно коррелирует с интенсивностью переживания «стья», а
мышцы нахмуривания — с состоянием «печали».
По паттерну трех лицевых мышц: m.zygomaticus (Z), m.corrugator (С) и жевательной мышцы — m.masseter (M) можно дифференцировать
4 эмоции (печаль, гнев,
радость, страх). Схематично эти и итерны реакций можно представить следующим
образом: печаль </1, СТ), гнев (Zl, СТ, МТ),
страх (Zl, Cl), радость (Zt, Cl, М). Таким образом, все
отрицательные эмоции сочетаются с по-iтлением
активности большой скуловой мышцы и ее усилением по время радости. Активность
мышцы нахмуривания возрастает во время гнева и печали и падает при страхе и
радости. Жевательная мышца активируется во время гнева и радости и не реагирует
во время печали и страха.
Как уже отмечалось, лицевая экспрессия зависит от социально-ю
контроля, определяемого принятыми нормами поведения. Он ' пижает корреляцию
эмоций с лицевыми реакциями. Требование в ' ч и,1 те контролировать
мимику также влияет на соотношение субъек-i инных оценок эмоций и выраженности лицевой экспрессии. Их корреляция
падает. В исследовании Р. Краута (Kraut
R.E., 1982) испытуем ые оценивали качество 12 запахов. В одной серии
— сразу после и пихания воздуха, содержащего тот или другой аромат; в другой '
грии испытуемый должен был сообщать экспериментатору об отношении к запаху,
изменяя свою позу. Усилия испытуемого сообщить информацию о запахах
увеличивают эмоциональные лицевые реакции на обонятельные стимулы. Однако при
этом субъективные оценки возрастают непропорционально увеличению мышечных
реакций лица (на 1/15 от изменений лицевой экспрессии). Важно то,
по корреляция субъективных оценок и лицевой экспрессии на за-пахи, когда не
требовалось передавать дополнительную информацию, была намного выше (0,5 и
0,43 против 0,1 и 0,16 для приятных и неприятных запахов соответственно).7.4.5.
Кодирование и декодирование лицевой экспрессии
Некоторые аспекты социального поведения человека обеспечиваются
биологическими программами. В частности, способность человека передавать
эмоциональные сигналы и принимать их имеет биологическую природу. Процесс
посылки эмоциональной информации при общении начинается с
мотивационно-эмОциональ-ного состояния, которое трансформируется в
соответствующее выражение лица. Это осуществляется с помощью специального кодирующего
механизма, существование которого доказывается соответствием паттернов
активности лицевых мышц определенным
эмоциям, хотя первичный паттерн лицевой экспрессии корректируется
«правилами поведения», которые были усвоены субъектом в результате обучения,
стимулируемого культурными и национальными традициями.
С эволюционной точки зрения внешняя экспрессия эмоций не была бы
полезной, если бы члены общества (группы) не могли декодировать эти сигналы и,
следовательно, адекватно реагировать на них. Это означает, что лицо,
принимающее сигналы (реципиент), должно быть подготовлено к их восприятию,
т.е. иметь специальный механизм для их декодирования. При этом демонстрация
гнева должна легко ассоциироваться с аверсивными явлениями и их следствиями —
возникновением эмоции страха. Наоборот, положительная экспрессия, например
выражение счастья на лице человека, должна вызывать положительные эмоциональные
реакции.
Механизм декодирования экспрессивной информации должен уметь
дифференцировать паттерны лицевой экспрессии, а также идентифицировать их как
сигналы определенных эмоциональных состояний. Биологическая природа этого
механизма проявляется во врожденной боязни человека большого открытого пространства
или змей и в той легкости, с которой он вырабатывает условные рефлексы на
отрицательном подкреплении, когда в качестве условных сигналов используются
аверсивные стимулы, вызывающие негативные эмоции. Для объяснения этих явлений
М. Селиг-мен (Seligman М.) в 70-х годах ввел
термин «биологическая прего-товность», полагая, что индивиды готовы отвечать
соответствующей адекватной эмоциональной реакцией на ситуации и стимулы,
которые могут иметь высокую биологическую цену.
Механизм декодирования эмоциональной экспрессии специально был
исследован У. Димбергом (Dimberg
U., 1988) из Университета в
Уппсала (Швеция), установившим, что лицевая экспрессия в зависимости от знака
эмоции по-разному влияет на эмоциональное состояние и условнорефлекторные
реакции страха партнеров. При этом эмоциональная экспрессия может воздействовать
на уровне подсознания, когда человек не отдает себе отчета о событии и факте
его воздействия. В своих опытах У. Димберг демонстрировал студентам фотографии
со «счастливыми» и «гневными» лицами. Он нашел, что оба типа стимулов нельзя
дифференцировать по вегетативным реакциям, которые одинаково вызывали снижение
ЧСС и увеличение кожной проводимости. Но при этом рассматривание счастливых лиц
сопровождалось большей активацией скуловой мышцы при тенденции к снижению
активности у мышцы нахмуривания. Лицо, выражающее гнев, вызывало противоположный
паттерн активности у тех же мышц.Паттерны реакций мышц лица, связанные с
эмоциями, возникают не только на социальные сигналы, но и на эмоциональные
ситуации в целом. Стимулы, вызывающие страх (изображение змеи), активируют
мышцу нахмуривания, а картины нейтрального содержания усиливают активность
большой скуловой мышцы. Подтвердив результаты, полученные параллельно другими
исследователями о корреляции между лицевой экспрессией и переживанием эмоций,
У. Димберг исследовал влияние специфики условного сигнала на выработку
условнорефлекторной реакции страха, измеряемой по величине кожного
сопротивления. В своих опытах он вырабатывал у человека две условные
оборонительные вегетативные реакции. В одном случае условным сигналом было
лицо с выражением счастья, в другом — лицо, выражающее гнев. В обоих случаях
подкреплением служил ток средней силы, приложенный к пальцу, и реакция на него
состояла в увеличении кожной проводимости. Перед выработкой условных рефлексов
проводилась серия опытов с привыканием к условным стимулам: фотографии с
лицами предъявлялись до тех пор, пока они не теряли способность вызывать
ориентировочный рефлекс в виде роста кожной проводимости. Признаком выработки
условного рефлекса считали появление вегетативной реакции на условный стимул.
Условнорефлек-торное повышение проводимости кожи было больше и реакция не
угасала дольше, если в роли условного сигнала выступала фотография лица с
выражением гнева, а не счастья.
Оборонительная условнорефлекторная реакция усиливалась, если в
условном сигнале увеличивали число аверсивных признаков. Например, у лица с
выражением гнева изменяли направление взгляда так, чтобы он был направлен прямо
на субъекта, а не в сторону. С помощью усиления аверсивных признаков в лице можно
задержать угасание ранее выработанной условной оборонительной реакции. Чтобы
получить этот эффект, У. Димберг сначала вырабатывал условную оборонительную
вегетативную реакцию на предъявление лица с выражением счастья, подкрепляемую
током. Затем исследовал процесс его угасания, подменяя условный сигнал на лицо
того же человека, но с выражением эмоции гнева. Это задерживало процесс
угасания оборонительной реакции по сравнению с условиями, когда подмены не
было. При обратной подмене условного сигнала («гнева» на «счастье») угасание
оборонительной вегетативной реакции, резко ускорялось, т.е. созерцание
счастливого лица успокаивало человека и убыстряло темп угасания оборонительного
рефлекса.
У. Димберг доказал, что влияние лицевой экспрессии на величину
условной вегетативной оборонительной реакции осуществ-
ияется автоматически и не зависит от процессов сознания. Чтобы
выяснить, контролируется ли влияние лицевой экспрессии когнитивными
процессами, он провел серию опытов с обратной маскировкой. Вырабатывалась
дифференцировка: лицо с выраженном гнева подкреплялось током, а с выражением
счастья — нет. Опыт включал три фазы: привыкание, обучение (формирование
условного вегетативного рефлекса — изменение кожной проводимости) и его
угасание. Обратную маскировку применяли только па стадии угасания выработанного
условного рефлекса, силу которого оценивали по величине кожной проводимости.
На стадии угасания условный сигнал (выражение гнева) экспонировали в течение 30
мс, затем заменяли лицом с выражением нейтрального состояния, которое
демонстрировали также в течение 30 мс. Такая процедура не позволяла опознать
первый стимул, испытуемый сообщал о появлении только второго. Маскировку
применя-||и к обоим стимулам: «гневному лицу», подкрепляемому током, и
«счастливому» — дифференцировочному сигналу, не подкреп-миемому током. Главный
результат — величина условного оборонительного вегетативного рефлекса на
применение стимула «гне-ма» при его обратной маскировке была больше по
сравнению с условнорефлекторным ответом, возникающим на дифференци-ровочный
стимул «счастья», который также маскировался. Таким образом, хотя человек не
осознавал предъявление условного сиг-пала «гнева», поведенчески он реагировал
адекватно, отвечая на лот стимул большей величиной аверсивной реакции. Это
означает, что лицевая экспрессия может вызывать эмоциональные эффекты,
которые влияют на исполнительное поведение. При этом моздействие протекает как
автоматический процесс на неосозна-наемом уровне (рис. 42).
Лицевые паттерны особенно сильно действуют на людей, про-янляющих
социальный страх. Субъекты, отобранные по признаку боязни публичных
выступлений, более бурно вегетативно реагируют даже на фотографию лица с
нейтральным состоянием по сравнению с изображением неодушевленных предметов
(грибы). При этом они переживают и оценивают свое восприятие нейтральных лиц
как негативное. Испытуемые с высоким социальным страхом оценивают фотографию
лица с выражением гнева как шачительно более эмоционально-отрицательное, а лицо
с выражением счастья — как менее положительное по сравнению с испытуемыми с
низким социальным страхом. Другими словами, они усиливают признаки
отрицательных и ослабляют признаки поло-* тельных эмоций на лице человека.
Таким образом, восприятие лицевой экспрессии — это не просто восприятие
некоторогоФительного паттерна, а восприятие значимого сигнала, влияющего на
состояние и поведение партнера, как это выявилось по отношению к
условнорефлекторной реакции страха.
Реакция человека на лицевую экспрессию партнера включает шемент
воспроизведения его мимики, что выражается в непроиз-польном изменении
активности мышц лица, зависящем от получаемой эмоциональной информации. Этот
процесс подобен «эмоциональному заражению, или резонансу». Лицевые мышцы как
бы имитируют, воспроизводят эмоцию, выраженную на лице партнера. Явление «эмоционального
резонанса» было описано П.В. Симоновым применительно к вегетативным реакциям.
Сопереживание с собакой-жертвой, выражающееся в появлении отрицательных
эмоциональных реакций (рост ЧСС и усиление в ЭЭГ тета-ритма), побуждает
собаку-наблюдателя вырабатывать инструментальный условный рефлекс, прекращающий
подачу тока собаке-жертве.
Для распознавания и идентификации
паттернов лицевых движений человек использует два канала. Один канал —
зрительный. Он производит опознание лицевой экспрессии с помощью гностических
нейронов нижневисочной коры, избирательно реагирующих на эмоциональное
выражение лиц. Другой канал оценивает паттерн собственной лицевой экспрессии,
созвучный эмоции партнера. С этих позиций обратная лицевая связь не только
выполняет функцию модуляции собственной эмоции, но и облегчает декодирование
эмоциональной мимики партнера.
23. Асиметрия мозга и эмоции. Индивидуальные различия.
Имеются многочисленные доказательства того, что восприятие эмоциональных сигналов находится под контролем правого полушария. Правосторонние корковые разрушения делают невозможным декодирование эмоционального настроения собеседника, нарушают распознавание лицевой экспрессии эмоций. Разрушения в правом полушарии (но не в левом) сопровождаются потерей способности выразить или передать свое переживание интонацией голоса.
Правое полушарие более тесно, чем левое, связано с вегетативными и телесными реакциями. Из клинической практики известно, что пациенты более осведомлены о нарушениях и отклонениях в реакциях на левой стороне тела. Человеку обычно легче постукивать синхронно в такт с сердечными ударами левой рукой.
Одно время казалось, что исключительно правое полушарие ответственно за обработку эмоциональных стимулов. Это дало основание рассматривать левое полушарие как «неэмоциональнуюструктуру» (Tucker D.M., 1981). Однако более поздние исследования показали, что оба полушария вносят свой вклад в эмоциональные переживания. Наиболее вероятной представляется точка зрения, согласно которой правое полушарие в большей степени связано с негативными эмоциями. Это заключение в значительной степени основано на клинических наблюдениях. При ослабленной функции левого полушария больные беспокойны, пессимистически настроены, часто плачут. При снижении активности правого полушария возникает беспричинная эйфория, индифферентное настроение или частый смех.
Т.А. Доброхотова и Н.Н. Брагина установили, что больные с поражениями левого полушария тревожны, озабочены. Правостороннее поражение сочетается с легкомыслием, беспечностью. Эмоциональные состояния благодушия, безответственности, беспечности, возникающие под влиянием алкоголя, связывают с его преимущественным воздействием на правое полушарие мозга.
По данным В.Л. Деглина, временное выключение левого полушария электросудорожным ударом тока вызывает сдвиг в эмоциональной сфере «правополушарного» человека в сторону отрицательных эмоций. Настроение ухудшается, он пессимистически оценивает свое положение, жалуется на плохое самочувствие. Выключение таким же способом правого полушария вызывает противоположный эффект — улучшение эмоционального состояния. В.Л. Деглин считает, что положительные эмоциональные состояния коррелируют с усилением альфа-активности в левом полушарии, а отрицательные эмоциональные состояния — с усилением альфа-активности в правом и усилением дельта-колебаний в левом полушарии.
Демонстрация фильмов разного содержания с помощью контактных линз раздельно в правое или в левое поле зрения показала, что правое полушарие быстрее реагирует на слайды с выражением печали, а левое — на слайды радостного содержания. При этом правое полушарие быстрее опознает эмоционально выразительные лица независимо от качества эмоции.
Распознавание мимики в большей степени связано с функцией правого полушария. Оно ухудшается при его поражении. Повреждение височной доли, особенно справа, нарушает опознание эмоциональной интонации речи. При выключении левого полушария независимо от характера эмоции улучшается распознавание эмоциональной окраски голоса.
Большинство исследователей склонны объяснять эмоциональную асимметрию полушарий головного мозга как вторичную эмоциональную специализацию (Симонов П.В., 1987). Согласно
Л.Р. Зенкову, выключение левого полушария делает ситуацию непонятной, невербализуемой и, следовательно, эмоционально-отрицательной. При выключении правого полушария ситуация оценивается как простая, ясная, понятная, что вызывает преобладание положительных эмоций. Следовательно, нарушение информационных процессов после отключения одного из полушарий нгорично сказывается на эмоциогенных механизмах.
По данным последних публикаций, локальные мозговые поражения, вызывающие изменения в эмоциональной сфере, находят но фронтальных областях коры. Это неудивительно, так как между фронтальной корой и лимбической системой существуют рецип-рокные отношения. По мнению Р. Давидсона и его коллег, именно фронтальные полюса регулируют аффективные переживания. Регистрация ЭЭГ у пациентов с депрессией выявляет особый тип функциональной асимметрии, характерный для этих больных: у них фокус активации находится во фронтальной и центральной областях правого полушария.
По результатам исследований Р. Давидсона примерно 50% самооценок состояния «счастья» определяется доминированием активности во фронтальных областях левого полушария. У 10-месячных младенцев восприятие лица человека с выражением счастья сочеталось с большей ЭЭГ-активацией в левом полушарии (Davidson R.J., Fox
N.A., 1982). В другом исследовании новорожденным давали пробовать сладкий сироп или раствор лимонной кислоты. Проглатывание сладкой жидкости вызывало интерес на лице ребенка и ЭЭГ-активацию в левой фронтальной коре. Гримаса отвращения и активация в правой фронтальной коре были реакциями на кислый сок.
Асимметрия ЭЭГ-активации исследована у 10-месячных младенцев в зависимости от типа улыбки. Анализировались улыбки на при-Г)лижение матери и другого человека. На мать ребенок реагировал улыбкой, при которой активировались большая скуловая мышца и круговая мышца глаза (улыбка Дачена). На приближение незнакомца ребенок также улыбался, однако круговая мышца глаза не реагировала, активация возникала только в m.zygomaticus. Первый тип улыбки (искренний) коррелировал с относительно большей активацией в левой фронтальной коре, второй тип улыбки — с активацией правой фронтальной коры (Davidson R.J., Fox N.A., 1982).
Н. Фокс и Р. Дэвидсон предложили модель, объясняющую знак эмоций в зависимости от межполушарных отношений. Согласно их концепции левая и правая фронтальная кора — анатомический субстрат соответственно для выражения тенденции «приближения» (approach) и «отказа* (withdrawal). Противостояние этих двух тен-
денций и определяет знак эмоции. Доминирование тенденции «приближения» сочетается с активацией левой фронтальной коры и появлением положительных эмоций. Р. Дэвидсон и В. Геллер (Davidson R.J., Heller W.) полагают, что знак эмоций зависит от соотношения активности левой (ЛФК) и правой (ПФК) фронтальной коры. Это правило В. Геллер (Heller W., 1993) представила в виде двух неравенств:
ЛФК > ПФК = положительные эмоции ПФК > ЛФК = отрицательные эмоции
Комментируя эти данные, П.В. Симонов (1997) отмечает, что в соответствии с потребностно-информационной теорией эмоций можно связать ПФК с прагматической информацией, приобретенной ранее и хранящейся в памяти, а ЛФК — с информацией, только что поступившей. Когда доминирует активность левого фронтального неокортекса, субъект располагает только новой информацией, которая не сопоставляется с ранее приобретенной. Поэтому никаких проблем не возникает и все эмоции имеют положительный знак. При доминировании активности правого фронтального неокортекса субъект располагает прежними знаниями, но понимает, что не может учитывать новую информацию, и поэтому страдает.
ИНДИВИДУАЛЬНЫЕ РАЗЛИЧИЯ И ЭМОЦИИ
Индивидуальные различия, связанные с особенностями функциональной асимметрии мозга, а также со свойствами темперамента, являются одним из факторов, определяющих специфику и силу эмоционального переживания. К измерениям темперамента, наиболее тесно связанным с эмоциями, относятся тревожность, сила — слабость процессов возбуждения, экстраверсия — интро-версия, импульсивность, эмоциональность.
Экстраверты имеют более низкий порог в отношении социальных стимулов с положительным знаком и поэтому более часто реагируют эмоциями радости и интереса. Они более чувствительны к награде, а интроверты — к наказанию. Измерение локального кровотока в мозге у интровертов в отличие от экстравертов выявило усиление кровотока в височных областях. Это различие объясняют более сильными связями коры интровертов с лимбической системой (миндалиной) — структурой, ответственной за реакции страха. Известно, что разрушение миндалины или височной коры уменьшает эмоциональные реакции страха и стимулирует ориентировочно-исследовательское поведение, в норме подавляемое оборонительными реакциями.
Лица с высокой личностной тревожностью предпочитают бо-• лее часто реагировать даже на нейтральные стимулы беспокойством, тревожностью, страхом. Повышенная тревожность субъекта коррелирует с высокой ЧСС и подавлением сосудистых и дыхательных ритмических модуляций RR-интервала в сердечном ритме, т.е. с вегетативным паттерном, указывающим на преобладание симпатической регуляции, обычно наблюдаемой во время стресса (Данилова Н.Н. и др., 1995).
Показано существование генетической предрасположенности к социальному страху, в частности проявляющемуся в боязни аудитории. Дж. Каган (Kagan J., 1989), ориентируясь на поведенческие характеристики, различает заторможенный и расторможенный типы темперамента. Дети с расторможенным типом поведения отличаются устойчивой социабельностью, говорливостью, спонтанно возникающими эмоциями. Дети с заторможенным темпераментом пугливы, застенчивы, ведут себя тихо. Их отличает низкий порог страха на новизну в социальном окружении. Устойчивость комплекса тгих поведенческих характеристик в одном исследовании была прослежена до возраста 21 мес., в другом до 7,5 лет. У сильно заторможенных детей обнаружено более высокое по сравнению с нормальными детьми содержание в крови кортизола — гормона стресса и негативных эмоциональных реакций. Уровень кортизола выше нормального сохранялся у них даже во сне.
Изучение реакций младенцев на боль и социальную изоляцию выявило устойчивые индивидуальные различия в выраженности отрицательных эмоций — боли, гнева, печали. Индивидуальные особенности эмоциональных реакций сохранялись в течение всего периода наблюдения до 4-месячного возраста.
На основании изучения поведения обезьян исследователи пришли к выводу, что степень готовности животного отвечать страхом на внешние стимулы генетически запрограммирована. Животные различаются порогами активации системы страха. Лабораторные животные в отличие от своих диких сородичей не обнаруживают страха при виде змеи, однако быстро обучаются реагировать страхом, глядя на других обезьян. Но среди них встречаются обезьяны, которых невозможно научить пугаться при виде змеи.
Эмоциональное общение в значительной степени зависит от индивидуальной способности передавать и принимать эмоциональные сигналы собеседника через мимику, позу, жесты, интонацию речи, т.е. от индивидуальных проявлений механизма кодирования и декодирования эмоций. П. Салавей и Дж. Майер (Salovey P., Mayer J.D., 1990) в своей социально ориентированной концепции эмоционального ума (emotional intelligence) настаивают на необхо…
24. Индивидуальная
активация по Закерману.
В работах М. Закермана индивидуальный уровень активации исследовался как тенденция человека к поиску или избеганию новых, сложных и интенсивных переживаний, а также как стремление к физическому и социальному риску. Эта тенденция получила название поиск ощущений (sensation seeking — SS). М. Закер-ман подчеркивает, что SS содержит в себе нечто, имеющее отношение к свойству активированности, которое проявляет себя в ответах на стимулы умеренной и большой интенсивности. Чтобы измерить индивидуальную потребность в стимуляции и активности, М.Закерман разработал специальный опросник, который позволяет оценивать тенденцию к «поиску ощущений». Пятая версия теста М. Закермана включает общую шкалу и четыре субшкалы, входящие в состав общей шкалы: 4 субшкалы были выделены на основе факторного анализа. Среди них:
• Фактор TAS (Thrill and Adventure Seeking) — поиска нервного волнения и авантюрных приключений — отражает стремление человека к опасным видам спорта, различным видам физической активности с риском для жизни;
• Фактор ES (Experience Seeking) — стремления к разнообразным впечатлениям, которые связаны с путешествиями, музыкой, искусством, общением с друзьями;
• Фактор DIS (Disinhibition) — расторможенности, характеризующий поиски социальной и гедонической стимуляции с помощью вечеринок, через сексуальное разнообразие, потребление алкоголя, участие в играх;
• Фактор BS (Boredom Susceptibility) — чувствительности к скуке — отражает индивидуальную неустойчивость к однообразным и шаблонным ситуациям и людям, а также нетерпеливость, когда не удается избежать таких ситуаций;
• Фактор G (General) — представляет суммарную оценку по 4 факторам (субшкалам), или шкалу поиска ощущений в целом (Sensation Seeking Scale — SSS).
В ряде работ показатели SS исследовались одновременно с различными компонентами ориентировочного рефлекса в виде измерения кожного сопротивления и частоты сердечных сокращений (ЧСС). Наиболее интересные результаты получены в отношении ЧСС, которая позволяет дифференцировать ориентировочный рефлекс от оборонительного. При ориентировочном рефлексе наблюдается снижение ЧСС сразу после стимула в течение нескольких секунд, тогда как появление оборонительного рефлекса выражается в кратковременном фазическом росте ЧСС. Было установлено, что лица с низким уровнем SS на звук высокой интенсивности отвечают появлением оборонительных реакций в виде фазичес-кого учащения ЧСС. Субъекты с высоким уровнем SS на ту же интенсивность звука реагируют по ориентировочному типу — кратковременным снижением ЧСС. Получены данные о том, что различные факторы SS по-разному связаны с ориентировочным и оборонительным рефлексами. Общий (генеральный) фактор — G и TAS-фактор коррелируют с ориентировочным ответом на новизну, а DIS-фактор различно связан с ориентировочными и оборонительными реакциями на интенсивную стимуляцию.
Согласно М. Закерману индивидуальный уровень SS имеет свои биохимические механизмы. Величина SS отрицательно коррелирует с содержанием в мозге фермента моноаминоксидазы (МАО), а также эндорфинов (эндогенных морфоподобных пептидов) и уровнем половых гормонов.
Лица с высокой величиной по шкале SSS имеют низкий уровень МАО, а у субъектов со слабо выраженным стремлением к поиску ощущений и переживаний уровень МАО в нейронах велик.
Сходные результаты получены и на обезьянах. Особи с высоким содержанием МАО больше времени проводят в одиночестве,. .
25. Нейроанатомия эмоций.
Наиболее убедительные данные о нейроанатомии эмоций получены в отношении тех мозговых структур, которые определяют знак эмоций. Опыты на животных с электрической стимуляцией и разрушением мозга, а затем и с самостимуляцией мозговых структур показали существование двух типов центров, возбуждение которых вызывает эмоции с противоположным знаком (приятные и неприятные). Впервые об этом заговорили в 1954 г. на Международном физиологическом конгрессе в Монреале (Канада). П. Мил-нер (P. Milner) и Дж. Олдс (J. Olds) продемонстрировали эффект самораздражения у крыс. Вживив электроды в медиальный пере-днемозговой пучок в области латерального гипоталамуса, они поместили крысу в ящик, где находился рычаг, при нажиме на который в мозг крысы посылался удар тока. Перемещаясь в клетке, крыса случайно задевала рычаг, производя таким образом само-
раздражение этого участка мозга. Со временем крыса все больше времени проводила у рычага И все чаще производила нажимы на него, создавая постоянное самораздражение, когда частота нажима на рычаг достигала до 7 тысяч за один час.
Стимуляция зон самораздражения вызывает положительные эмоциональные реакции и служит более сильным подкреплением, чем даже пищевое. На основе этого подкрепления легко вырабатываются прочные условные рефлексы.
Однако, существуют и другие центры, раздражение которых вызывает реакцию избегания. Так, крыса с вживленными в эту область электродами после нажима на рычаг избегает находиться рядом с ним. Эти зоны расположены в перевентрикулярных отделах промежуточного и среднего мозга. Области мозга, раздражение которых ведет к подкреплению или избеганию, получили соответственно название центров удовольствия и неудовольствия.
Центры неудовольствия найдены в центральном сером веществе, и в гипоталамусе. Структурой мозга, критически связанной со страхом, является миндалина (см. главу «Память»). По мнению П.В. Симонова, вместе с гипоталамусом миндалина образует мо-тивационно-эмоциональную систему, так как одной из ее функций является выбор доминирующей мотивации. В концепции американского невропатолога Дж. Пейпеца (J. Papez) о мозговом субстрате эмоций подчеркивается роль поясной извилины. Изучая эмоциональные расстройства у больных с поражением гиппокам-па и поясной извилины, он выдвинул гипотезу о существовании единой системы, объединяющей ряд структур мозга и образующей мозговой субстрат для эмоций. Эта система представляет зам-кнутую цепь и включает: гипоталамус — передневентральное ядро таламуса — поясную извилину — гиппокамп — мамиллярные ядра гипоталамуса. Она получила название круга Пейпеца. Позднее П. МакЛин (P. McLean) в 1952 г., учитывая, что поясная извилина как бы окаймляет основание переднего мозга, предложил называть ее и связанные с ней другие структуры мозга лимбической системой (limbus — край). Источником возбуждения в этой системе является гипоталамус. Сигналы от него следуют в средний мозг и нижележащие отделы для инициации вегетативных и моторных эмоциональных реакций. Одновременно нейроны гипоталамуса через коллатерали посылают сигналы в передневентральное ядро и таламусе. По этому пути возбуждение передается к поясной извилине коры больших полушарий.
Поясная извилина, по Дж. Пейпеду, является субстратом осознанных эмоциональных переживаний и имеет специальные входы для эмоциональных сигналов, подобно тому как зрительная кора
имеет входы для зрительных сигналов. Далее сигнал из поясной извилины через гиппокамп вновь достигает гипоталамуса в области его мамиллярных тел. Так нервная цепь замыкается. Путь от поясной извилины связывает субъективные переживания, возникающие на уровне коры, с сигналами, выходящими из гипоталамуса для висцерального и моторного выражения эмоций.
Однако сегодня гипотеза Дж. Пейпеца приходит в противоречие со многими фактами. Под сомнением оказалась роль гиппо-кампа и таламуса в возникновении эмоций. У человека стимуляция гиппокампа электрическим током не сопровождается появлением эмоций (страха, гнева и т.п.). Субъективно пациенты испытывают лишь спутанность сознания. П.В. Симонов (1987) относит гиппокамп к «информационному», а не эмоциональному образованию.
Из всех структур круга Пейпеца наиболее тесную связь с эмоциональным поведением обнаруживают гипоталамус и поясная извилина. Кроме того, оказалось, что и многие другие структуры мозга, не входящие в состав круга Пейпеца, обладают сильным влиянием на эмоциональное поведение. Среди них особую роль играют лобная и височная кора головного мозга, а также миндалина.
Поражение лобных долей приводит к глубоким нарушениям эмоциональной сферы человека. Преимущественно развиваются два синдрома: эмоциональная тупость и растормаживание примитивных эмоций и влечений. При этом в первую очередь страдают высшие эмоции, связанные с деятельностью, социальными отношениями, творчеством. Билатеральное удаление у обезьян височных полюсов ведет к подавлению их агрессивности и страха. Эффект сходен с разрушением миндалины. Передняя лимбическая кора контролирует эмоциональные интонации, выразительность речи у человека и звуковых сигналов у обезьяны. После двустороннего кровоизлияния в этой зоне речь пациента становится эмоционально невыразительной.
Согласно современным данным поясная извилина имеет двусторонние связи со многими подкорковыми структурами (перегородкой, верхними буграми четверохолмия, синим пятном и др.), а также с различными областями коры в лобных, теменных и височных долях. Ее связи, по-видимому, более обширны, чем у какого-либо другого отдела мозга. Г. Шеперд (1987) предполагает, что поясная извилина выполняет функцию высшего координатора различных систем мозга, вовлекаемых в организацию эмоций.
В составе эмоции принято выделять собственно эмоциональное, субъективное переживание и его соматическое и висцеральное выражение с их самостоятельными механизмами. Диссоциа-ция эмоционального переживания и его выражения в двигательных и вегетативных реакциях обнаружена при некоторых поражениях ствола мозга. Она выступает в так называемых псевдоэффектах: интенсивные мимические и вегетативные реакции, характерные для плача или смеха, могут протекать без соответствующих субъективных ощущений.
Таким образом, эмоция — сложное образование, предполагающее интеграцию соматических, вегетативных и субъективных компонентов. Нейронные сети, представляющие эмоции, распределены по многим структурам мозга. О них еще мало что известно. Наиболее полная информация получена о структурах, определяющих знак эмоции.
Центры самораздражения, активность которых связывают с появлением положительных эмоций, содержат ДА-ергические нейроны. Эффективность самораздражения соответствует плотности ДА-ергической инервации. К ДА-ергическим структурам относится медиальный переднемозговой пучок, связывающий гипоталамус и лимбическую систему с лобными долями. ДА-ергические нейроны содержатся также в вентральной тегментальной области покрышки (ВТО), прилегающем ядре (n.accumbens). Через ВТО и прилегающее ядро инициируются и реализуются основные биологические мотивации (нападение, поедание пищи и др.).
Черная субстанция и хвостатое ядро (ХЯ) также относятся к ДА-ергической системе. Но они в большей степени обеспечивают реализацию приобретенных форм поведения и причастны к выработке условных рефлексов на положительном подкреплении. В черной субстанции обезьян обнаружены нейроны, реагирующие на пищевое подкрепление соком, а после выработки условного пищевого рефлекса — и на его условные сигналы. Они составили 25% от всех обследованных нейронов этой структуры. Кроме того, эти же нейроны отвечают на новые, неожиданные раздражители. С упрочением условного пищевого рефлекса их реакция на пищевое подкрепление ослабевает, но сохраняется на условный сигнал пищи. Связь активации данной группы нейронов с подкреплением наиболее явно видна при отставлении подкрепления, когда сок подается через 500 мс после реакции животного. Нейроны активировались в момент предъявления пищевого подкрепления, но только после правильно выполненной реакции. В случае неправильных ответов, которые не подкреплялись, отвечали противоположной реакцией — торможением активности в то время, когда обычно давалось подкрепление.
Таким образом, на нейронах черной субстанции сходятся активирующие влияния, совпадающие с действием пищевого подкрепления
или его условного сигнала, и тормозные влияния, которые соответствуют отмене, отсутствию пищевого подкрепления.
В отличие от нейронов префронтальной коры, которые разряжаются в течение всего времени отставления, ДА-ергические нейроны черной субстанции своими фазическими реакциями выделяют основные сигналы, связанные с пищевым подкреплением. В то же время они получают информацию и об отсутствии ожидаемого подкрепления, что проявляется в подавлении их активности.
ДА-ергические структуры мозга обладают повышенной чувствительностью к естественным и искусственным опиатам. Это указывает на сложный биохимический механизм центров положительного подкрепления, использующих как медиаторную, так и пеп-тидергическую регуляцию. Так, микроинъекция морфина в ВТО облегчает самостимуляцию мозга, а введение опиатных антагонистов нарушает самовведение героина. Выработанная реакция самостимуляции нарушается и при повреждении ВТО, сопровождающемся снижением уровня ДА. Снижение содержание ДА в n.accumbens или повреждение каиновой кислотой его клеток-мишеней для ДА-ергических терминалей нарушает самовведение кокаина.
Эффект положительного подкрепления, возникающий при активации n.accumbens, обусловлен совместным действием на его нейронах ДА-ергических терминалей от клеток ВТО и опиатного входа. Взаимодействие опиатов с моноаминами на уровне клетки создает качественно новый эффект (Сахаров Д.А., 1992). Опиоиды открыты и у беспозвоночных. У виноградной улитки ритмическое открывание и закрывание дыхальца определяется совместным действием ДА и энкефалина. Совместное действие энкефалина и другого медиатора — серотонина — не только обусловливает появление у клетки ритмической активности, но и синхронизацию ее в ансам-Оле соседних нейронов. Почти все жизненно важные функции организма находятся под контролем опиоидных пептидов (Сахаров Д.А., 1992).
П.В. Симонов и его коллеги, исследуя механизм самораздражения латерального гипоталамуса и латеральной преоптической области крысы, выделили два типа нейронов, противоположно связанных с мотивацией и положительными эмоциями. «Мотива-ционные» нейроны проявляли максимум активности при стимуляции зон самораздражения силой тока, вызывающей мотиваци-онное поведение, и тормозили свою активность со снижением уровня естественной пищевой и водной мотивации. «Подкрепляющие» нейроны достигали максимума активности при стимуляции силой тока, вызывающей самораздражение. Они также возбуждались при естественном насыщении животного. Авторы полагают, что "подкрепляющие» нейроны в центрах самораздражения латерального
гипоталамуса и преоптической латеральной области представляют систему «истинной» награды со свойственной ей положительной эмоцией, которая позволяет вырабатывать активные формы условных инструментальных рефлексов, когда животное полностью предотвращает действие болевого раздражителя (рефлекс избегания).
П.В. Симонов (1997), рассматривая формирование инструментальных оборонительных рефлексов, различает разные формы награды — в виде ослабления наказания и получения собственно положительного подкрепления, когда активируются центры награды и вовлекается механизм положительных эмоций. Чтобы сформировать инструментальный рефлекс избегания, в результате которого крыса перестает получать наказание током, в качестве подкрепления требуется только награда. Данная форма инструментального рефлекса сочетается с активностью позитивно-подкрепляющих нейронов латерального гипоталамуса, тогда как для выработки классического оборонительного рефлекса или реакции избавления достаточно отмены отрицательного подкрепления. Их осуществление сопровождается усилением активности мотиваци-онных нейронов и подавлением импульсаций — подкрепляющих.
При высокой пищевой мотивации активируются нейроны латерального гипоталамуса, а при насыщении — вентромедиального. В латеральном гипоталамусе выявлены глюкозочувствительные нейроны, которые в ответ на электрофоретическое подведение к ним глюкозы пропорционально дозе тормозят свою активность без изменения сопротивления мембраны. В вентромедиальном гипоталамусе найдены глюкозорецептивные нейроны. На глюкозу они реагируют противоположно — в соответствии с величиной дозы увеличивают частоту разрядов.
Глюкозочувствительные нейроны латерального гипоталамуса у обезьяны реагируют учащением спайков на вид и запах пищи, на вид шприца с глюкозой, на скорлупу ореха. Их активность модулируется пищевой мотивацией. Только у голодной обезьяны эти нейроны реагируют на пищевые стимулы или их сигналы. Введение глюкозы снижает их активность. По данным Е. Роллса (Е. Rolls), сначала возникает реакция нейронов, реагирующих на вид пищи (через 15—200 мс), а затем включаются нейроны, отвечающие на прием пищи (300 мс). Обнаружены нейроны, отвечающие либо на вид пищи, либо на вид воды.
Глюкозочувствительные нейроны вовлекаются в процесс пи-щедобывательного поведения. У обезьян их активность резко возрастает за 2—0,8 с перед нажимом на рычаг для получения пищи и подавляется во время движения и следующего за ним подкрепления. Эти нейроны одновременно чувствительны к НА и опиатам.
В латеральном гипоталамусе, кроме того, имеются глюкозоне-чувствительные нейроны. Они также активируются во время пище-добывательного поведения, однако их реакции совпадают с движениями, а не предшествуют им. Эти нейроны принимают участие в реализации движений и, кроме того, распознают ключевые эк-стероцептивные стимулы. Они чувствительны к ДА. Предполагают, что глюкозонечувствительные нейроны интегрируют внешнюю информацию, тогда как глюкозочувствительные участвуют в интеграции внутренней информации и подкрепления.
Животные, как и человек, имеют тенденцию принимать стимуляторы (кокаин, амфетамин) и опиаты (героин, морфин). Действие этих веществ можно сравнить с действием электрического раздражения на центры положительного подкрепления. Наркотики действуют не только на центры награды, стимулируя их. Параллельно они тормозят центры отрицательного подкрепления (наказания). Поведенчески частично это проявляется в изменении структуры защитных реакций: усиливается роль активно-оборонительного поведения и подавляется пассивная форма защиты, снимаются страх, тревожность. При прекращении введения наркотика возникает отрицательное состояние, которое также стимулирует применение наркотика, чтобы снять это состояние.
Состояние эйфории, вызываемое наркотиком, является той положительной эмоцией, которая работает как сильное положительное подкрепление и обеспечивает быстрое формирование устойчивых условных связей данного состояния с различными внешними стимулами, обстановкой и др. Наркотическая эйфория подобна эффекту, вызываемому активацией ДА-ергических центров положительного подкрепления. Ее привлекательность определяется не только ее положительным гедоническим тоном, но и тем, что человек переживает подъем умственной и физической активности, его работоспособность повышается, он мыслит ясно и четко.
Необходимым условием для формирования активной формы поведения — инструментального рефлекса — является наличие положительной эмоции в качестве подкрепления. Связь наркотика с положительной эмоцией объясняет механизм инициации поведения «поиска наркотика». Однажды испытав положительный эффект от приема наркотика или электрического раздражения мозга, животное образует ассоциации с сигналами, которые сопутствуют приему наркотика. Так, крысы ищут место, которое ассоциировано с введением наркотика или раздражением положительного центра. Крысы, не имеющие опыта введения опиатов, легко начинают осуществлять его самовведение аналогично тому, как это происходит при самораздражении мозга.
Стимулы, сочетаемые с действием морфина, сами могут вызывать ряд реакций, присущих наркотику как безусловному раздражителю. Таким образом, условные раздражители (привычная обстановка) могут привести к активации наркотического состояния и при возвращении к прежней обстановке восстановить потребность в наркотиках.
Применение наркотиков и электрическая стимуляция системы положительного подкрепления характеризуются эффектом «разогрева». После подачи электрического стимула возникает стремление усилить самораздражение. А прием однократной дозы наркотика сопровождается увеличением его самовведения у крысы.
Эффект самораздражения мозга обнаружен и у улитки. Поведение моллюска изучали при его свободном перемещении по шару, погруженному в воду. При касании улиткой стержня, связанного с электродом, замыкалась цепь подачи тока на определенную группу нервных клеток. Когда стержень не подключен к току, улитка время от времени с некоторой фиксированной частотой касается стержня. В случае если касание стержня замыкает цепь и ток подается на группу клеток оборонительного поведения париетального ганглия, после одного-двух касаний улитка, оставаясь активной, избегает стержня и частота касания падает до нуля.
Если ток пропускается через клетки мезоцеребрума, связанные с половым поведением, улитка начинает касаться стержня все чаще и чаще. Одновременно раздражение клеток мезоцеребрума подавляет ответ в клетках оборонительного поведения, что указывает на реципрокные отношения между центром полового «удовольствия» и центром «неудовольствия».
Использование в качестве положительного подкрепления прямого раздражения нейронов полового поведения увеличивает вероятность появления подкрепляемой реакции. Так, сочетая спонтанное закрытие дыхальца с раздражением мезоцеребральных клеток, можно увеличить длительность закрытого состояния дыхальца. Сочетание открытия дыхальца с раздражением тех же клеток приводило к увеличению времени пребывания его в открытом состоянии. Раздражение нейронов, запускающих оборонительное поведение, может использоваться в качестве отрицательного подкрепления.
28. Структура двигательного акта в терминах функциональной
системы П.К. Анохина.
Взаимодействие человека и животных с окружающей средой осуществляется через целенаправленную деятельность или поведение. Двигательный акт как элемент поведения воспроизводит основные звенья его структуры. Ведущим системообразующим фактором целенаправленного поведения, так же как и отдельного двигательного акта, является полезный для жизнедеятельности организма приспособительный результат.
Следует различать такие понятия, как «двигательный акт» и «элементарное движение». Двигательный акт реализуется с помощью моторной программы, а в простейшем случае через возбуждение командного нейрона, управляющего согласованной работой группы мотонейронов и соответствующих мышечных единиц. Элементарное движение представлено сокращением/расслаблением мышечных единиц, вызываемых стимуляцией мотонейрона. Одна и та же двигательная программа может быть выполнена с помощью разного набора элементарных движений мышц. Например, левой или правой рукой.
С позиции принципа системного квантования процессов жизнедеятельности двигательный акт может быть соотнесен с отдельным квантом поведения. Это наиболее очевидно при иерархическом квантовании, когда удовлетворение ведущей потребности значительно отставлено во времени и для достижения конечного результата необходимо выполнить ряд предварительных действий. Например, при конструировании человеком определенного изделия, когда для создания конечного продукта необходимо решить ряд промежуточных задач со своими конкретными результатами (Судаков К.В., 1997).
Наиболее совершенная модель структуры поведения изложена в концепции функциональных систем П.К. Анохина. Определяя функциональную систему как динамическую, саморегулирующуюся организацию, избирательно объединяющую структуры и процессы на основе нервных и гуморальных механизмов регуляции для достижения полезных системе и организму в целом приспособительных результатов, П.К. Анохин распространил содержание этого понятия на структуру любого целенаправленного поведения
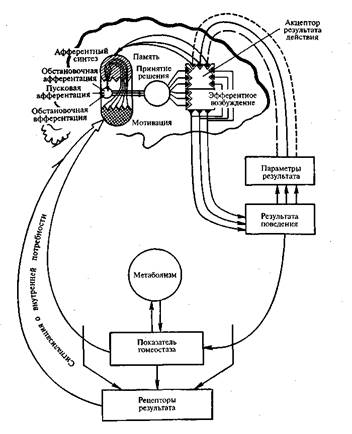
(Анохин П.К., 1968). С этих позиций может быть рассмотрена и структура отдельного двигательного акта.
Центральная архитектоника функциональных систем, определяющих целенаправленные поведенческие акты различной степени сложности, складывается из следующих последовательно сменяю-щих друг друга стадий: афферентный синтез, принятие решения, акцептор результатов действия, эфферентный синтез, формирование действия и, наконец, оценка достигнутого результата (рис. 51).
Поведенческий акт любой степени сложности начинается со стадии афферентного синтеза. Возбуждение, вызванное внешним стимулом, действует не изолированно. Оно непременно вступает во взаимодействие с другими афферентными возбуждениями, имеющими иной функциональный смысл. Мозг непрерывно обрабатывает все сигналы, поступающие по многочисленным сенсорным каналам. И только в результате синтеза этих афферентных возбуждений создаются условия для реализации определенного целенаправленного поведения. Содержание афферентного синтеза определяется влиянием нескольких факторов: мотивационного возбуждения, памяти, обстановочной и пусковой афферентации.
Мотивационное возбуждение появляется вследствии той или другой витальной, социальной или идеальной потребности. Специфика мотивационного возбуждения определяется особенностями, типом вызвавшей его потребности. Оно — необходимый компонент любого поведения. Важность мотивационного возбуждения для афферентного синтеза вытекает уже из того, что условный сигнал теряет способность вызывать ранее выработанное пище-добывательное поведение (например, побежку собаки к кормушке для получения пищи), если животное уже хорошо накормлено и, следовательно, у него отсутствует мотивационное пищевое возбуждение.
Роль мотивационного возбуждения в формировании афферентного синтеза определяется тем, что любая поступающая информация соотносится с доминирующим в данный момент мотиваци-онным возбуждением, которое действует как фильтр, отбирающий наиболее нужное для данной мотивационной установки.
Нейрофизиологической основой мотивационного возбуждения является избирательная активация различных нервных структур, создаваемая прежде всего лимбической и ретикулярной системами мозга. На уровне коры мотивационное возбуждение представлено специфическим паттерном возбуждения.
Хотя мотивационное возбуждение является очень важным ком-понетом афферентного синтеза, оно не единственный его компонент. Внешние стимулы с их разным функциональным смыслом по отношению к данному, конкретному организму также вносят свой вклад в афферентный синтез. Выделяют два класса стимулов с функциями пусковой и обстановочной афферентации.
Условные и безусловные раздражители, ключевые стимулы (вид ястреба — хищника для птиц, вызывающего поведение бегства, и
др.) служат толчком к развертыванию определенного поведения или отдельного поведенческого акта. Этим стимулам присуща пусковая функция. Картина возбуждения, создаваемая билогически значимыми стимулами в сенсорных системах, и есть пусковая аф-ферентация. Однако способность пусковых стимулов инициировать поведение не является абсолютной. Она зависит от той обстановки и условий, в которых они действуют.
Зависимость формирования условного рефлекса от обстановки опыта была описана уже И.П. Павловым. Неожиданное изменение обстановки может разрушать ранее выработанный условный рефлекс. Однако обстановочная афферентация, хотя и влияет на появление и интенсивность условнорефлекторной реакции, сама неспособна вызывать эти реакции.
Влияние обстановочной афферентции на условный рефлекс наиболее отчетливо выступило при изучении явления динамического стереотипа. В этих опытах животное тренировали для выполнения в определенном порядке серии различных условных рефлексов. После длительной тренировки оказалось, что любой случайный условный раздражитель может воспроизвести все специфические эффекты, характерные для каждого раздражителя в системе двигательного стереотипа. Для этого лишь необходимо, чтобы он следовал в заученной временной последовательности. Таким образом, решающее значение при вызове условных рефлексов в системе динамического стереотипа приобретает порядок их выполнения. Следовательно, обстановочная афферентация включает не только возбуждение от стационарной обстановки, но и ту последовательность афферентных возбуждений, которая ассоциируется с этой обстановкой. Обстановочная афферентация создает скрытое возбуждение, которое может быть выявлено, как только подействует пусковой раздражитель. Физиологический смысл пусковой афферентации состоит в том, что, выявляя скрытое возбуждение, создаваемое обстановочной афферентацией, она приурочивает его к определенным моментам времени, наиболее целесообразным с точки (рения самого поведения.
Решающее влияние обстановочной афферентации на условно-рефлекторный ответ было показано в опытах И.И. Лаптева — сотрудника П.К. Анохина. В его экспериментах звонок утром подкреплялся едой, и тот же звонок вечером сопровождался ударом электрического тока. В результате было выработано два разных условных рефлекса: утром — слюноотделительная реакция, вечером — оборонительный рефлекс. Животное научилось дифференцировать два комплекса раздражителей, различающихся только временным компонентом.Афферентный синтез включает также использование аппарата памяти. Очевидно, что функциональная роль пусковых и обстановочных стимулов уже обусловлена прошлым опытом животного. Это и видовая память, и индивидуальная, приобретенная в результате обучения. На стадии афферентного синтеза из памяти извлекаются именно те фрагменты прошлого опыта, которые полезны, нужны для будущего поведения.
Таким образом, на основе взаимодействия мотивационного, обстановочного возбуждения и механизмов памяти формируется готовность к определенному поведению. Но для того чтобы она трансформировалась в целенаправленное поведение, необходимо воздействие пусковых стимулов. Пусковая афферентация — последний компонент афферентного синтеза.
Процессы афферентного синтеза, охватывающие мотивацион-ное возбуждение, пусковую и обстановочную афферентацию, аппарат памяти, реализуются с помощью специального модулирующего механизма, обеспечивающего необходимый для этого тонус коры больших полушарий и других структур мозга. Этот механизм регулирует и распределяет активирующие и инактивирующие влияния, исходящие из лимбической и ретикулярной систем мозга. Поведенческим выражением роста уровня активации в нервной системе является возникновение ориентировочно-исследовательских реакций и поисковой активности животного.
Стадия афферентного синтеза сменяется стадией принятия решения, которая определяет тип и направленность поведения. Принятие решения реализуется через формирование аппарата акцептора результатов действия. Этот аппарат программирует результаты будущих событий. В нем актуализирована врожденная и индивидуальная память животного и человека в отношении свойств внешних объектов, способных удовлетворить возникшую потребность, а также способов действия, направленных на достижение или избегание целевого объекта. Нередко в этом аппарате запрограммированы все варианты поиска во внешней среде соответствующих раздражителей.
Представление об акцепторе результатов действия как о предвидении будущих результатов легло в основу концепции об опережающем отражении действительности, которая позволила на психофизиологическом уровне подойти к изучению проблемы цели. Ожидаемые результаты, формируемые в акцепторе результатов действия, выполняют функцию целевого объекта для поведения и двигательного акта.
До того как целенаправленное поведение начнет осуществляться, развивается еще одна стадия поведенческого акта — стадия
программирования действия, или эфферентного синтеза. На этом этапе осуществляется интеграция соматических и вегетативных возбуждений. Для данной стадии характерно, что действие уже сформировано, но еще не претворено в жизнь.
Следующая стадия — реальное выполнение программы действия под влиянием эфферентного возбуждения, достигающего исполнительных механизмов.
Благодаря аппарату акцептора результатов действия и эфферентному синтезу, организм имеет возможность сравнить ожидаемые результаты с поступающей афферентной информацией о реальных результатах и параметрах совершаемого действия. Именно результаты сравнения определяют последующее построение поведения: оно либо корректируется, либо прекращается, как в случае достижения конечного результата. И тогда поведенческий акт (действие) завершается последней санкционирующей стадией-удовлетворением потребности.
Таким образом, определяющим моментом функциональных систем, формирующих разнообразные формы поведения животных и человека, является не само действие, а полезный для системы и организма в целом результат поведения. Инициативная роль в формировании целенаправленного поведения принадлежит исходным потребностям и соответствующим им мотивациям, которые мобилизуют генетически детерминированные или индивидуально приобретенные программы поведения (Судаков К.В., 1990).
Намерение выполнить определенное движение проявляется в ЭЭГ в виде потенциала готовности. Этот потенциал отражает антиципацию (предвидение) будущих событий. Впервые явление антиципации в лабораторных условиях было изучено И.П. Павловым в виде рефлекса на время, когда у собаки слюноотделение заранее нарастало к привычному моменту приема пищи.
Чтобы выявить потенциал готовности, антиципацию, связанную с движением, испытуемому дают инструкцию: регулярно, через постоянный промежуток времени, воспроизводить какое-либо движение. В этих условиях началу движения предшествует медленная негативная волна, которая и представляет потенциал готовности. Возникая за 800 мс до начала движения, потенциал готовности за 90 мс перед выполнением движения сменяется быстрым потенциалом, связанным с посылкой управляющей команды. Потенциал готовности развивается биполярно в премоторной коре и отражает формирование замысла движения. Быстрый потенциал возникает в контралатеральном полушарии и отражает работу нейронов моторной коры.
Потенциал готовности часто трудно отделить от волны ожида-ния. Последнюю легко обнаружить в потенциалах мозга в ситуации, когда один стимул служит сигналом к появлению второго, значимого раздражителя. На интервале между предупреждающим и императивным стимулами в лобных и центральных отделах коры развивается медленно нарастающая негативность, которая достигает пика к моменту нанесения второго, императивного стимула.
Эта медленная негативная волна впервые была обнаружена Греем Уолтером (1966) — известным английским физиологом из Берденовского неврологического института в Бристоле. Он дал ей название «волна ожидания», или «волна Е». Сегодня чаще используют термин «условное негативное отклонение». Волна ожидания, которую можно записать, используя усилитель постоянного тока или переменного с большой (5—8 с) постоянной времени, связана с ожиданием сенсорной информации (рис. 52).
В управлении двигательным поведением различают стратегию и тактику. Стратегию движения определяет конкретная мотивация (биологическая, социальная и др.). Именно на ее основе определяется цель поведения, т.е. то, что должно быть достигнуто. В структуре поведенческого акта цель закодирована в акцепторе результатов действия. В отношении двигательного акта это выглядит как формирование двигательной задачи, того, что следует делать. Под тактикой понимают конкретный план действий, т.е. то, как будет достигнута цель поведения, с помощью каких двигательных ресурсов и способов действия. Тактическое планирование движения непосредственно представлено в блоке программ. При построении двигательной программы учитывают множество факторов, в том числе общую стратегию, пространственно-временные характеристики среды, сигнальную значимость ее стимулов, прошлый жизненный опыт.
30.
Механизм инициации двигательного акта.
Изучение процессов, определяющих выполнение моторных программ, привело к представлению о двух системах инициации движения. Одна из них — это лимбическая система мозга, по терминологии Ю. Конорского, «эмоциональный мозг». С помощью згой системы осуществляется «трансляция мотивации в действие», т.е. в действия, которые связаны с утолением голода, устранением страха и удовлетворением других биологических потребностей. По данным Г. Могенсона (Mogenson G. et al., 1980), эта трансляция достигается за счет особого пути передачи сигналов от лимбичес-ких структур к базальным ганглиям. При этом важным звеном, связывающим лимбическую систему с моторной, является прилегающее адро (n.accumbens). Его возбуждение сигналами из лимби-ческой системы, по-видимому, через механизм активации ДА-ер-i ических путей ведет к актуализации врожденных поведенческих программ.
С появлением ассоциативной коры у высших животных увеличивается роль когнитивных процессов. Появляется вторая система инициации движения — «когнитивный мозг» (Конорский Ю., 1970).Он обеспечивает запуск различных специфических движений в соответствии с инструкцией, установкой на ответ, прошлым опытом и обучением. Приход сигнала от ассоциативной коры в нео-стриатум базальных ганглиев, который имеет обширные сенсорные проекции, растормаживает его командные нейроны и тем самым актуализирует двигательные программы, главным образом двигательные автоматизмы, выученные движения. Эти программы через таламус также достигают моторной коры. Таков путь, через который «когнитивные команды» инициируют программы движений, сохраняемые в ЦНС. Прилегающее и хвостатое ядра фильтруют сигналы соответственно от лимбической системы («эмоционального мозга») и от церебральной коры («когнитивного мозга»). Хотя в процессе филогенетического развития роль «когнитивного мозга» в инициации двигательных ответов возрастает, «эмоциональный мозг» и «когнитивный мозг» обычно действуют совместно.
По Г. Могенсону, все процессы управления движением можно представить тремя блоками: блоком инициации движения, включающим лимбическую систему с прилегающим ядром (n.accum-bens) и ассоциативную кору; блоком программирования движения, включающим мозжечок, базальные ганглии, моторную кору, таламус как посредника между ними, а также спинальные и стволовые генераторы; исполнительным блоком, охватывающим мотонейроны и двигательные единицы. Следует иметь в виду, что управление движением включает наряду с командами по прямым связям обширную проприоцептивную и экстероцептивную информацию по обратным связям.
«Эмоциональный мозг» (лимбическая система) действует через n.accumbens и далее через другие структуры базальных ганглиев. «Когнитивный мозг» (ассоциативная кора) оказывает влияние на хвостатое ядро базальных ганглиев и параллельно на мозжечок. Моторная кора получает сигналы от мозжечка и базальных ганглиев через таламус.
Подобная схема объясняет не только инициацию непроизвольных движений, врожденных форм двигательного поведения и автоматизмов, но и произвольных движений. Рефлексы и произвольные движения не противоречат друг другу. Произвольные движения также подчиняются рефлекторному принципу. Это особенно очевидно в случае инструментальных реакций, которые являются простейшей формой произвольных движений.
Что же отличает произвольное движение от непроизвольного? По определению шведского нейрофизиолога Р. Гранита (Granit R.), которое он приводит в своей книге «Целенаправленный мозг», произвольным в произвольном движении является его цель. Цели
произвольных движений могут быть бесконечно разнообразными. B.C. Гурфинкель также определяет произвольное движение в связи с его целью. Рассматривая профессиональные движения стрелков-спортсменов, например прицеливание, он отмечает, что особенностью хорошо управляемого движения у стрелка является точное удержание цели. Пистолет неподвижен, тогда как многие части тела двигаются, т.е. положение руки стрелка в пространстве стабилизируется всякого рода рефлекторными механизмами: вестибу-лоокулярной системой, вестибулоспинальной системой и др.
Произвольные движения человека — это сознательно регулируемые движения. У человека они тесно связаны с речью. Роль речевого опосредования в превращении непроизвольных импульсивных движений у детей в произвольные и сознательно управляемые изучали А.Р. Лурия и его сотрудники. Они установили, что активация детской речи — предварительное планирование в речевой форме предпринимаемых действий — ведет ребенка к овладению своим поведением, тогда как задержка в речевом развитии приводит к снижению уровня произвольной или волевой регуляции двигательного поведения ребенка. Произвольные движения, вызываемые инструкцией или внутренним побуждением человека, опосредованы внутренней речью, претворяющей замысел (цель) во внутренний план действий.
Ведущая роль в процессе управления движением принадлежит префронтальной коре. Выполняемая ею актуализация следов памяти позволяет префронтальной коре корректировать внутреннюю модель внешнего мира в соответствии с оперативно поступающей сенсорной информацией, в том числе от выполняемого движения (Гольдман-Ракич П.С., 1992). С функцией префронтальной коры связывают способность мысленно проектировать будущую траекторию движущейся цели. У макак резусов с повреждением соответствующих участков в дорзальной части префронтальной коры, где расположены глазодвигательные центры, возникают расстройства мысленной экстраполяции траектории движения.
Таким образом, управление и контроль за движением — достаточно сложный процесс. Он включает обработку информации, получаемой через прямые и обратные связи между префронтальной корой, моторной корой, таламусом, мозжечком, базальными ганглиями, а также стволом мозга и спинным мозгом. Важная роль принадлежит проприоцептивной и экстероцептивной афферента-ции. Двигательная система организована по иерархическому принципу с постепенным увеличением сложности сенсомоторной интеграции. На каждом ее уровне имеется своя «ведущая афферента-ция» и собственный тип регулируемых движений.
31. Развитие
речи. Функции речи.
Слово становится «сигналом сигналов» не сразу. У ребенка раньше
всего формируются условные пищевые рефлексы на вкусовые и запаховые
раздражители, затем на вестибулярные (покачивание) и позже на звуковые и
зрительные. Условные рефлексы на словесные раздражители появляются лишь во
второй половине первого года жизни. Общаясь с ребенком, взрослые обычно
произносят слова, сочетая их с другими непосредственными раздражителями. В
результате слово становится одним из компонентов комплекса. Например, на слова
«Где мама?» ребенок поворачивает голову в сторону матери только в комплексе с
другими раздражениями: кинестетическими (от положения тела), зрительными
(привычная обстановка, лицо человека, задающего вопрос), звуковыми (голос,
интонация). Стоит изменить один из компонентов комплекса, и реакция на слово
исчезает. Лишь постепенно слово начинает приобретать ведущее значение, вытесняя
другие компоненты комплекса. Сначала выпадает кинестетический компонент, затем
теряют свое значение зрительные и звуковые раздражители. И уже само слово
вызывает реакцию.
Показ предмета и его называние постепенно приводят к формированию
их ассоциации, затем слово начинает заменять обозначаемый им предмет. Это
происходит к концу первого года жизни и началу второго. Однако слово сначала
замещает лишь конкретный предмет, например данную куклу, а не куклу вообще. На
этом этапе развития слово выступает как интегратор первого порядка.
Превращение слова в интегратор второго порядка, или в «сигнал
сигналов», происходит в конце второго года жизни. Для этого необходимо, чтобы
на него был выработан пучок связей (не менее 15 ассоциаций). Ребенок должен
научиться оперировать различными предметами, обозначаемыми одним словом. Если
число выработанных связей меньше, то слово остается символом, который замещает
лишь конкретный предмет.
Между третьим и четвертым годами жизни формируются понятия — интеграторы
третьего порядка. Ребенок уже понимает такие слова, как «игрушка», «цветы»,
«животные». К пятому году жизни понятия усложняются. Так, ребенок пользуется
словом «вещь», относя его к игрушкам, посуде, мебели и т.д.
В процессе онтогенеза взаимодействие двух сигнальных систем
проходит через несколько стадий. Первоначально условные рефлексы ребенка
реализуются на уровне первой сигнальной системы: непосредственный раздражитель
вступает в связь с непосредственными вегетативными и двигательными реакциями.
По терминологии А.Г. Иванова-Смоленского, это связи типа Н—Н (непосредственный
раздражитель — непосредственная реакция). Во втором полугодии ребенок начинает
реагировать на словесные раздражители непосредственными вегетативными и
соматическими реакциями, следовательно, добавляются условные связи типа С—Н
(словесный раздражитель — непосредственная реакция). К концу первого года жизни
(после 8 мес.) ребенок уже начинает подражать речи взрослого так, как это
делают приматы, используя отдельные звуки для обозначения предметов,
происходящих событий, а также своего состояния. Позже ребенок начинает произносить
отдельные слова. Сначала они не связаны с каким-либо предметом. В возрасте
1,5—2 лет часто одним словом обозначается не только предмет, но и действия и
связанные с ним переживания. Лишь позже происходит дифференциация слов на
категории, обозначающие предметы, действия, чувства. Появляется новый тип
связей Н—С (непосредственный раздражитель — словесная реакция). На втором году
жизни словарный запас ребенка увеличивается до 200 слов и более. Он уже может
объединять слова в простейшие речевые цепи и строить предложения. К концу третьего
года словарный запас достигает 500—700 слов. Словесные реакции вызываются не
только непосредственными раздражителями, но и словами. Появляется новый тип
связей С—С (словесный раздражитель — словесная реакция), и ребенок научается
говорить.
С развитием речи у ребенка в возрасте 2—3 лет усложняется
интегративная деятельность мозга: появляются условные рефлексы на отношения
величин, весов, расстояний, окраски предметов. В возрасте 3—4 лет
вырабатываются различные двигательные и некоторые речевые стереотипы.
ФУНКЦИИ РЕЧИ
Исследователи выделяют три основные функции речи: коммуникативную,
регулирующую и программирующую. Коммуникативная функция обеспечивает
общение между людьми с помощью языка. Речь используется для передачи информации
и побуждения к действию. Побудительная сила речи существенно зависит от ее эмоциональной
выразительности.
Через слово человек получает знания о предметах и явлениях
окружающего мира без непосредственного контакта с ними. Система словесных
символов расширяет возможности приспособления человека к окружающей среде,
возможности его ориентации в природном и социальном мире. Через знания,
накопленные человечеством и зафиксированные в устной и письменной речи,
человек связан с прошлым и будущим.
Способность человека к общению с помощью слов-символов имеет свои
истоки в коммуникативных способностях высших обезьян.
Л.А. Фирсов и его сотрудники предлагают делить языки на первичные
и вторичные. К первичному языку они относят поведение животного и человека,
различные реакции: изменение формы, величины и цвета определенных частей тела,
изменения перьевого и шерстного покровов, а также врожденные коммуникативные
(голосовые, мимические, позные, жестикуляторные и др.) сигналы. Таким образом,
первичному языку соответствует допонятий-ный уровень отражения действительности
в форме ощущений, восприятий и представлений. Вторичный язык связан с понятийным
уровнем отражения. В нем различают стадию А, общую для человека и животного
(довербальные понятия). Сложные формы обобщения, которые обнаруживают
антропоиды и некоторые низшие обезьяны, соответствуют стадии А. На стадии Б
вторичного языка (вербальные понятия) используется речевой аппарат. Таким
образом, первичный язык соответствует первой сигнальной системе, а стадия Б
вторичного языка — второй сигнальной системе. Согласно Л.А. Орбели,
эволюционная преемственность нервной регуляции поведения выражается в
«промежуточных этапах» процесса перехода от первой сигнальной системы ко
второй. Им соответствует стадия А вторичного языка.
Язык представляет собой определенную систему знаков и правил их
образования. Человек осваивает язык в течение жизни. Какой язык он усвоит как
родной, зависит от среды, в которой он живет, и условий воспитания. Существует
критический период для освоения языка. После 10 лет способность к развитию
нейронных сетей, необходимых для построения центров речи, утрачивается.
Маугли — один из литературных примеров потери речевой функции.
Человек может владеть многими языками. Это означает, что он
использует возможность обозначать один и тот же предмет разными символами как
в устной, так и в письменной форме. При изучении второго и последующих языков,
по-видимому, используются те же нервные сети, которые ранее были сформированы
при
овладении родным языком. В настоящее время известно более 2500
живых развивающихся языков.
Языковые знания не передаются по наследству. Однако у человека
имеются генетические предпосылки к общению с помощью речи и усвоению языка. Они
заложены в особенностях как центральной нервной системы, так и
речедвигательного аппарата, гортани. Амбидексы — лица, у которых функциональная
асимметрия полушарий менее выражена, обладают большими языковыми способностями.
Регулирующая функция речи реализует себя в высших психических
функциях — сознательных формах психической деятельности. Понятие высшей
психической функции введено Л.С. Выготским и развито А.Р. Лурия и другими
отечественными психологами. Отличительной особенностью высших психических
функций является их произвольный характер.
Предполагают, что речи принадлежит важная роль в развитии
произвольного, волевого поведения. Первоначально высшая психическая функция
как бы разделена между двумя людьми. Один человек регулирует поведение другого
с помощью специальных раздражителей («знаков»), среди которых наибольшую роль
играет речь. Научаясь применять по отношению к собственному поведению стимулы,
которые первоначально использовались для регуляции поведения других людей,
человек приходит к овладению собственным поведением. В результате процесса
интериоризации — преобразования внешней речевой деятельности во внутреннюю
речь, последняя становится тем механизмом, с помощью которого человек
овладевает собственными произвольными действиями.
А.Р. Лурия и Е.Д. Хомская в своих работах показали связь регулирующей
функции речи с передними отделами полушарий. Ими установлена важная роль конвекситальных
отделов префронталь-ной коры в регуляции произвольных движений и действий, конструктивной
деятельности, различных интеллектуальных процессов. Больной с патологией в
этих отделах не может выполнять соответствующие действия, следуя инструкции. Показано
также решающее участие медиобазальных отделов лобных долей в регуляции
избирательных локальных форм активации, необходимых для осуществления
произвольных действий. У больных с поражениями этих отделов мозга угасание
сосудистого компонента ориентировочного рефлекса на индифферентный
раздражитель не нарушается. Однако восстановления ориентировочного рефлекса
под влиянием речевой инструкции, придающей стимулам сигнальное значение, не
происходит. У них же не может удерживаться в качестве компонента произвольного
внимания тонический ориентировочный рефлекс в виде длительной ЭЭГ-активации,
хотя тонический ориентировочный рефлекс продолжает возникать на непосредственный
раздражитель. Таким образом, высшие формы управления фазическим и тоническим
ориентировочными рефлексами, так же как и регулирующая функция речи, зависят
от сохранности лобных долей.
Программирующая функция речи выражается в построении
смысловых схем речевого высказывания, грамматических структур предложений, в
переходе от замысла к внешнему развернутому высказыванию. В основе этого
процесса — внутреннее программирование, осуществляемое с помощью внутренней
речи. Как показывают клинические данные, оно необходимо не только для речевого
высказывания, но и для построения самых различных движений и действий.
Программирующая функция речи страдает при поражениях передних отделов речевых
зон — заднелобных и премоторных отделов полушария.
Клинические данные, полученные при изучении поражений мозга, а
также результаты его электрической стимуляции во время операций на мозге
позволили выявить те критические структуры коры, которые важны для способности
говорить и понимать речь. Методика, позволяющая картировать области коры,
связанные с речью, с помощью прямого электрического раздражения, была
разработана в 30-х годах У. Пенфильдом в Монреале в Институте неврологии для
контроля за хирургическим удалением участков мозга с очагами эпилепсии. Во
время процедуры, которая проводилась под местным наркозом, больной должен был
называть показываемые ему картинки. Речевые центры выявлялись по афази-ческой
остановке (потере способности говорить), когда на них попадало раздражение
током.
Наиболее важные данные об организации речевых процессов получены
при изучении локальных поражений мозга. Согласно взглядам А.Р. Лурия, выделяют
две группы структур мозга с различными функциями речевой деятельности. Их
поражение вызывает две категории афазий: синтагматические и
парадигматические. Первые связаны с трудностями динамической организации
речевого высказывания и наблюдаются при поражении передних отделов левого
полушария. Вторые возникают при поражении задних отделов левого полушария и
связаны с нарушением кодов речи (фонематического, артикуляционного,
семантического и т.д.).
К передним отделам речевых зон коры относится и центр Бро-ка. Он
расположен в нижних отделах третьей лобной извилины, у большей части людей в
левом полушарии. Эта зона контролирует осуществление речевых реакций. Ее
поражение вызывает эффе-
265рентную моторную афазию, при которой страдает
собственная речь больного, а понимание чужой речи в основном сохраняется. При
эфферентной моторной афазии нарушается кинетическая мелодия слов за счет
невозможности плавного переключения с одного элемента высказывания на другой.
Больные с афазией Брока большую часть своих ошибок осознают. Говорят они с
большим трудом и мало.
Поражение другой части передних речевых зон (в нижних отделах
премоторной коры) сопровождается так называемой динамической афазией, когда
больной теряет способность формулировать высказывания, переводить свои мысли в
развернутую речь (нарушение программирующей функции речи). Протекает она на
фоне относительной сохранности повторной и автоматизированной речи, чтения и
письма под диктовку.
Центр Вернике относится к задним отделам речевых зон коры.
Он расположен в височной доле и обеспечивает понимание речи. При его поражении
возникают нарушения фонематического слуха, появляются затруднения в понимании
устной речи, в письме под диктовку (сенсорная афазия). Речь такого
больного достаточно беглая, но обычно бессмысленная, так как больной не
замечает своих дефектов. С поражением задних отделов речевых зон коры связывают
также акустико-мнестическую, оптико-мнестическую афазии, в основе
которых лежит нарушение памяти, и семантическую афазию — нарушение
понимания логико-грамматических конструкций, отражающих пространственные
отношения предметов.
Продолжая давнюю традицию изучения речи у больных с локальными
поражениями мозга, Антониу и Анна Дамазиу (1992) предположили, что речь можно
рассматривать как трехкомпонен-тную систему: образование слов, формирование
понятий и промежуточные процессы, играющие роль посредника между первыми
двумя компонентами. Рассматривая нарушения цветового зрения, они выделили
несколько типов аномалий. Поражение зрительной коры в зонах VI и V4 приводит к ахроматопсии, когда человек теряет способность
воспринимать цвет. У таких больных страдает и представление цвета. Люди с
ахроматопсией обычно видят мир в оттенках серого. Пытаясь вызвать цветовой
образ, они видят форму, движение, текстуру, но не цвет. Когда они думают о
траве, то представляют ее не зеленой, а кровь не красной. Поражение этой части
мозга приводит к нарушению понятий и, следовательно, к дефектам мышления. Ни в
каком другом участке мозга повреждение не приводит к такому результату.
Другой тип цветовой аномалии связан с поражением височной области.
При этом страдают не понятия цветов, а их называние.
Больные хорошо различают цвета, могут их сортировать по образцам.
Но они говорят «синий» или «красный», когда им показывают зеленый или желтый
цвет, при этом они безошибочно кладут зеленый квадрат радом с рисунком луга и
желтый — рядом с изображением банана. Больной не только не может назвать
показанный ему цвет, но и слыша название цвета, не может указать на него, т.е.
у него нарушена связь между восприятием, представлением цвета и его словесным
обозначением.
А.З. Дамазиу и А. Дамазиу показали, что функция называния в
отношении различных категорий объектов выполняется различными областями мозга.
Они описали поведение больных А.Н. и Л.Р. с поражениями в передней и
средневисочной коре. У больных полностью сохранилась понятийная система. Они
безошибочно узнают, что за объекты находятся перед ними. Могут определить их
функциональное назначение, среду, в которой они существуют, ценность объекта.
Но они с трудом называют многие хорошо знакомые предметы. При этом они делают
меньше ошибок при назывании инструментов, чем при назывании животных, овощей и
фруктов. Они правильно называют части тела, но с трудом называют знакомые
музыкальные инструменты. Кроме того, пациенты А.Н. и Л.Р. испытывали трудности,
когда их просили называть своих друзей, родственников, известных популярных
деятелей.
Авторы рассматривают структуры мозга, обеспечивающие функцию называния,
как систему посредника, связывающую структуры, в которых представлены понятия,
со структурами, формирующими слова и предложения. По их данным, функция называния
для общих понятий локализована в задних левых височных областях, а для более
специальных — в передних, вблизи левого височного полюса. По существу авторы
расширяют представление о функции заднеречевой системы, куда входит и центр
Вернике. Они полагают, что задняя речевая система в левом полушарии хранит
слуховые и кинестетические записи фонем и их последовательностей, составляющих
слова. Поражение задней речевой области не нарушает ритма человеческой речи и
ее скорости. Не страдает и синтаксическая структура предложений.
Задняя речевая система сообщается с моторной и премоторной зонами
коры как непосредственно, так и через подкорковый путь. Последний включает
левые базальные ганглии и ядра передней части таламуса. Через эти пути
осуществляется двойной контроль произнесения звуков речи. Подкорковый путь
активируется при приобретении и исполнении речевого навыка. Корковый путь
связан с более осознанным контролем речевого акта. Похоже, что во время
речевого акта корковая и подкорковая системы действу-
267ют параллельно. При заучивании ребенком слова «желтый» одновременно
активируются область, ответственная за цветовые понятия, система
словообразования и двигательного контроля (через корковый и подкорковый пути).
Со временем устанавливается прямой путь между понятийной системой и базальными
ганглиями, и тогда роль структуры посредника уменьшается. Последующее заучивание
нового названия цвета на иностранном языке снова потребует участия системы
посредника для установления соответствия слуховых, кинестетических и
двигательных фонем.
На рис. 57 в обобщенном виде представлена схема распределения
системы, ответственной за речь, по Г. Шеперду (1987). Она основана на
результатах электростимуляции речевых центров у нейрохирургических больных и
анатомического изучения мозга обезьян и человека. Показаны структуры и их
связи, с помощью которых выполняется функция называния. Зрительная
информация сначала поступает в поле 17, затем она обрабатывается в полях 18 и
19. Отсюда перцептивный образ объекта передается в обширную заднюю речевую
зону, в состав которой наряду с центром Вернике входит поле 39 (в теменной
доле). Оно посылает информацию о зрительном образе предмета полю 22, где
хранится его слуховой образ. Из поля 22 информация передается в речевую зону
Брока, в которой находятся двигательные программы речи. Нужная программа
считывается в моторную кору, которая и управляет речевой мускулатурой,
обеспечивая сложную пространственно-временную
координацию работы соответствующих мыщц, необходимую для того,
чтобы мы могли назвать увиденный предмет.
Спереди от роландовой (центральной) борозды находится область,
ответственная за ритм речи и грамматику, — так называемая дополнительная (или
добавочная) моторная область (ДМО). Больные с поражением этой области говорят
без интонации, делают большие паузы между словами, путаются в грамматике, пропускают
союзы, местоимения, нарушают грамматический порядок слов. Им легче
пользоваться существительными, чем глаголами. Поражение данной области
нарушает грамматическую обработку как произносимой, так и слышимой речи, что
наводит на мысль о том, что здесь происходит «сборка» целых фраз.
На рис. можно видеть картину локального мозгового кровотока во
время устной речи и его отличие от активации мозга придвижении или только при
его воображении — ритмического сжимания и разжимания правой руки, а также в
состоянии покоя. Видно, что речь активирует как заднюю, так и переднюю речевые
зоны. При представлении движения появляются очаги активации в лобной, теменной
и височной коре. Однако в моторной коре (вдоль центральной борозды) активность
пока незначительна. При выполнении движения фокус активации смещается в
область моторной коры. В состоянии покоя можно видеть очаги активации в лобных
долях, по-видимому, отражающие течение когнитивных про-цесов, не контролируемых
заданием.
Левые базальные ганглии — составная часть передней и задней систем
речи. Известно, что базальные ганглии объединяют компоненты сложных движений в
единое целое. По-видимому, сходную функцию они выполняют и в отношении речевых
реакций, связывая слова в предложения.
Передняя речевая область коры, похоже, связана с мозжечком,
осуществляющим точное временное кодирование двигательных реакций. При
поражении мозжечка возникает моторная и когнитивная дисметрия — плохое
выполнение точных действий, включая когнитивные. Это указывает на причастность
мозжечка к выполнению речевых и мыслительных операций.
32. Асимметрия мозга и речь.
Нейроанатомы и нейропатологи, занимавшиеся посмертным изучением мозга, длительное время не замечали анатомическую межполушарную асимметрию мозга. Только в 1968 г. Н. Гешвинд и У. Левицкий из Гарвардского университета обратили внимание на значительную разницу в размерах правой и левой височных долей. В большинстве случаев участок коры около верхнего края височной извилины, уходящий глубоко в силь-виеву ямку (латеральную борозду) в левом полушарии значительно больше. Именно в этом участке находится центр Вернике — часть задней речевой зоны. Это означает, что у большинства людей речевые структуры локализованы в левом полушарии, а не в правом. Такое утверждение справедливо примерно для 99% правшей и 2/3 левшей.
Дополнительные данные о речевых функциях полушарий были получены в опытах Р. Сперри на больных «срасщепленным мозгом». После рассечения комиссуральных связей двух полушарий у таких больных каждое полушарие функционирует самостоятельно, получая информацию только справа или слева. У них в каждое полушарие поступает только половина зрительной информации:в левое полушарие — от правой половины зрительного поля, в правое — от левой.
Если больному «с расщепленным мозгом» в правую половину зрительного поля предъявить какой-либо предмет, то он может его назвать и отобрать правой рукой. То же самое со словом: он может его прочесть или написать, а также отобрать соответствующий предмет правой рукой. Таким образом, если используется левое полушарие, такой больной не отличается от нормального человека. Дефект проявляется, когда стимулы возникают на левой стороне тела или в левой половине зрительного поля. Предмет, изображение которого проецируется в правое полушарие, больной назвать не может. Однако он правильно выбирает его среди других, хотя и после этого назвать его по-прежнему не может, т.е. правое полушарие не обеспечивает функции называния предмета, но способно его узнавать.
Хотя с лингвистическими способностями преимущественно связано левое полушарие, тем не менее правое полушарие также обладает некоторыми языковыми функциями. Так, если зрительно предъявить название предмета, то больной не испытывает затруднений в нахождении левой рукой соответствующего предмета среди нескольких других, т.е. правое полушарие может понимать письменную речь.
В опытах Дж. ЛеДу и М. Газзанига на больном С.П., перенесшем комиссуротомию, у которого правое полушарие обладало значительно большими лингвистическими способностями, чем обычно, было показано, что с помощью правого полушария больной может не только «читать» вопросы, но и «отвечать» на них левой рукой, составляя слова из букв, нанесенных на карточки. Таким же способом больной С.П. мог называть предметы, предъявляемые ему зрительно в правое полушарие, а точнее, «писать» с помощью правого полушария, составляя слова из букв левой рукой.
Функциональную специализацию левого и правого полушарий головного мозга долгое время считали присущей только человеку, связывая ее с появлением речи. Однако латеральная специализация наблюдается уже у животных, обнаруживая сходство со специализацией полушарий человека.
Левое полушарие птиц, грызунов, хищников и приматов имеет преимущественное отношение к коммуникативным функциям, мученным формам поведения, тонким двигательным манипуляциям. Правое полушарие связано с эмоционально-окрашенным и зрительно-пространственным поведением. Левое полушарие важно для ориентировочно-исследовательской деятельности животного, которая ухудшается после удаления левой коры. «Когнитивные» медиаторы — ДА, АХ, ГАМК — преобладают в левом полушарии, а медиаторы, наиболее тесно связанные с мотивационно-эмоциональным поведением, — серотонин, норадреналин — в большем количестве находят в правом полушарии.
Речь могла появиться и в результате совершенствования системы, использующей звуковые сигналы в коммуникативных целях, которая преимущественно связана с функциями левого полушария.Таким образом, функциональная асимметрия полушарий формируется на этапах эволюции еще до человека. И, по-видимому, она объясняется различием способов обработки информации, представленных в разных полушариях.
33. Структура процесса мышления. Вербальный и невербальный
интеллект… .
Мышление представляет собой процесс познавательной деятельности, при котором субъект оперирует различными видами обобщений, включая образы, понятия и категории.
Появление речи в процессе эволюции принципиально изменило функции мозга. Мир внутренних переживаний, намерений приобрел качественно новый аппарат кодирования информации с помощью абстрактных символов. Это не только обусловило возможность передачи информации от человека к человеку, но и сделало качественно иным процесс мышления. Мы лучше осознаем, понимаем мысль, когда облачаем ее в языковую форму. Вне языка мы переживаем неясные побуждения, которые могут быть выражены лишь в жестах и мимике. Слово выступает не только как средство выражения мысли: оно перестраивает мышление и интеллектуальные функции человека, так как сама мысль совершается и формируется с помощью слова.
Суть мышления — в выполнении некоторых когнитивных операций с образами во внутренней картине мира. Эти операции позволяют строить и достраивать меняющуюся модель мира. Благодаря слову картина мира становится более совершенной, дифференцированной, с одной стороны, и более обобщенной — с другой. Присоединяясь к непосредственному образу предмета, слово выделяет его существенные элементарные или комплексные признаки, которые непосредственно недоступны субъекту. Слово пе-
реводит субъективный смысл образа в систему значений, что делает его более понятным как самому субъекту, так и его партнеру.
С позиции теории функциональных систем П. К. Анохина основные этапы мыслительного процесса могут быть сопоставлены с этапами структуры поведенческого акта. Направленность процесса мышления определяется доминирующей мотивацией субъекта. Афферентный синтез выбирает зону поиска решения проблемы. Поступающая информация анализируется и сопоставляется со знаниями, извлекаемыми из памяти, содержание которых существенно определяется доминирующей мотивацией. Этапу принятия решения соответствует выбор наиболее вероятной гипотезы для ее последующей проверки и доказательств. В акцепторе результатов действия в соответствии с принятой гипотезой формируются некоторые представления о том, что прежде всего следует подтвердить, доказать или опровергнуть. Эфферентный синтез содержит замыслы доказательств и проверок. Выполнение конкретного доказательства, которое подтверждает справедливость выдвинутого предположения, эквивалентно этапу осуществления реального действия. В случае неудачи активируется ориентировочно-исследовательская деятельность субъекта. Она приводит к изменению содержания акцептора результатов, а также эфферентного синтеза. Возникают новые замыслы, идеи и, возможно, привлекаются иные способы доказательств.
У человека различают два основных вида мышления: наглядно-образное и словесно-логическое. Последнее функционирует на базе языковых средств и представляет собой наиболее поздний период филогенетического и онтогенетического развития мышления.
ВЕРБАЛЬНЫЙ И НЕВЕРБАЛЬНЫЙ ИНТЕЛЛЕКТ
На основании соотношения первой и второй сигнальных систем И.П. Павлов предложил классификацию специально человеческих типов высшей нервной деятельности, выделив художественный, мыслительный и средний типы.
Художественный тип характеризуется преобладанием функций первой сигнальной системы. Люди этого типа в процессе мышления широко пользуются чувственными образами. Они воспринимают явления и предметы целиком, не дробя их на части. У мыслительного типа, у которого усилена работа второй сигнальной системы, резко выражена способность отвлечения от действительности, основанная на стремлении анализировать, дробить действительность на части, а затем соединять части в целое. Для среднего типа свойственна уравновешенность функций двух сигнальных систем.И.П. Павлов в работе «Двадцатилетний опыт» писал: «Жизнь отчетливо указывает на две категории людей: художников и мыслителей. Между ними резкая разница. Одни — художники во всех их родах: писателей, музыкантов, живописцев и т.д. — захватывают действительность целиком, сплошь, сполна, живую действительность, без всякого дробления, без всякого разъединения. Другие — мыслители — именно дробят ее и тем как бы умерщвляют ее, делая из нее какой-то временный скелет, и затем только постепенно как бы снова собирают ее части и стараются их таким образом оживить, что вполне им все-таки так и не удается» (Полн. собр. соч. М.-Л.: АН СССР, 1951. Т. 3. Кн. 2. С. 214).
Большинство людей принадлежат к среднему типу. По мнению И.П. Павлова, крайние типы — «художественный» и «мыслительный» — служат поставщиками нервных и психиатрических клиник.
Для диагностики специально человеческих типов высшей нервной деятельности — «художественного» и «мыслительного» — в работах школы Б.М. Тегаюва широко применяется методика М.Н. Борисовой. Создавая свой метод, М.Н. Борисова исходила из описания И.П. Павловым особенностей отражения действительности у «художников» и «мыслителей». Для первых характерно непосредственное, целостное отражение, для вторых — отражение аналитическое, опосредованное словом. Для своей методики она выбрала два вида деятельности — узнавание и словесное описание зрительно запоминаемых объектов (изображения наборов листьев), полагая, что узнавание зрительно запоминаемых рисунков требует функций первой сигнальной системы, так как по своей природе оно более синтетично, свернуто. Словесное описание тех же изображений развернуто, аналитично и в большей мере предполагает использование второй сигнальной системы. По результатам зрительного и вербального способов запоминания методика позволяет определять соотношение двух сигнальных систем.
Сопоставление показателей соотношения первой и второй сигнальных систем (по М.Н. Борисовой) и теста Д. Векслера, измеряющего вербальный и невербальный интеллект, выполненное методом факторного анализа, выявило связь невербального интеллекта с преобладанием у испытуемых зрительного способа запоминания (лучшие показатели при узнавании, чем при описании), а вербального интеллекта — с описанием по М.Н. Борисовой (Голубева Э.А., 1993).
В один из двух выделенных факторов (образно-действенный) с наибольшими весами вошли результаты исполнения невербальных субтестов (кубики Кооса, сюжетные картинки, сложение частей, недостающие детали, символы), их суммарная оценка, а
также «узнавание» по М.Н. Борисовой. К другому фактору (словесно-логическому) с наибольшими факторными нагрузками отнесены исполнение вербальных субтестов (словарного, сходства, общей осведомленности), их суммарная оценка и «описание» по М.Н. Борисовой.
Два специально человеческих типа высшей нервной деятельности, диагностируемые по тесту Векслера, методике Борисовой, а также Коссова, тяготеют к разным типам темперамента.Темпе-рамент по И.П. Павлову определяется как сочетание трех основных свойств нервной системы: силы, уравновешенности, подвижности процессов возбуждения и торможения. Свойства нервной системы диагностировались по ЭЭГ-методикам, по коэффициенту «в», характеризующему наклон кривой зависимости времени реакции от интенсивности стимула, и критической частоте мельканий. Установлено, что испытуемые с меланхолическим темпераментом (со слабыми нервными процессами, их инертностью и преобладанием торможения над возбуждением) имеют более высокие показатели вербального интеллекта (по Векслеру) и по соотношению сигнальных систем относятся к «мыслительному» типу. Флегматики, сангвиники и холерики по сравнению с меланхоликами примерно одинаково тяготеют к художественному типу. Однако меланхоликам в большей степени противостоят холерики. Таким образом, черты темперамента и когнитивные особенности специально человеческих типов высшей нервной деятельности образуют своего рода различные эмоционально-познавательные комплексы (Пе-ченков В.В., 1997).
Интеллектуальные особенности «мыслительного» типа сочетаются с повышенной тревожностью и пессимизмом меланхолического темперамента. Особенности «художественного» типа могут сочетаться с любым из трех других типов темперамента, для которых в целом характерен более оптимистический эмоциональный настрой по сравнению с меланхолическим темпераментом.
На основе многолетних комплексных исследований специально человеческих типов высшей нервной деятельности, темперамента, общих и специальных способностей Э.А. Голубева и ее коллеги (1993,
1997) выделяют два основных комплекса признаков, характеризующих два типа мышления, группирующихся вокруг общих либо коммуникативных, либо познавательных способностей (см. табл., с. 276). В таблице показана схема дихотического представления интеллекта в структуре индивидуальности. Наиболее высокий уровень невербального интеллекта (по Векслеру) характерен для «художников» (по методикам Борисовой, Коссова), которые обладают коммуникативными способностями: музыкальными,.. .
34.
Асимметрия мозга и мышление. Половые различия и интеллект
В норме оба полушария работают в тесном взаимодействии, дополняя друг друга. Различие между левым и правым полушариями можно выявить, не прибегая к хирургическому вмешательству — рассечению комиссур, связывающих оба полушария. Для этого может быть использован метод «наркоза полушарий». Он был создан в клинике для выявления полушария с речевыми функциями. По этому методу в сонную артерию на одной стороне шеи вводят тонкую трубку для последующего введения раствора барбитуратов (амиталнатрия). В связи с тем что каждая сонная артерия снабжает кровью лишь одно полушарие, снотворное, введенное в нее, попадает в одно полушарие и оказывает на него наркотическое действие. Во время теста больной лежит на спине с поднятыми руками и считает от 100 в обратном порядке. Через несколько секунд после введения наркотика можно видеть, как бессильно падает одна рука пациента, противоположная стороне инъекции. Затем наблюдается нарушение в счете. Если вещество попадает в речевое полушарие, остановка счета в зависимости от введенной дозы длится 2-5 мин, если в другое полушарие, задержка составляет всего несколько секунд. Таким образом, применение этого метода дает возможность на время выключать любое полушарие и исследовать изолированную работу оставшегося.
Использование методик, с помощью которых можно избирательно воздействовать только на одно полушарие, позволило исследователям продемонстрировать значительные различия в умственных способностях двух полушарий. Полагают, что левое полушарие участвует в основном в аналитических процессах; это полушарие — база для логического мышления. Преимущественно оно обеспечивает речевую деятельность — ее понимание и построение, работу со словесными символами. Обработка входных сигналов осуществляется в нем, по-видимому, последовательно. Правое полушарие обеспечивает конкретно-образное мышление и имеет дело с невербальным материалом, отвечая за определенные навыки в обращении с пространственными сигналами, за структурно-пространственные преобразования, способность к зрительному и тактильному распознаванию предметов. Поступающая к нему информация обрабатывается одномоментно и целостным способом.
Правое полушарие лучше, чем левое, справляется с различением ориентации линий, кривизны, многоугольников неправильных очертаний, пространственного расположения зрительных сигналов, глубины в стереоскопических изображениях. Однако левое полушарие обнаруживает большие способности в отношении других аспектов зрительно-пространственного восприятия. Оно лучше дифференцирует нарисованные лица, если они различаются только одной чертой. Правое полушарие лучше различает их, когда они отличаются не одной, а многими чертами. Предполагают, что левое полушарие превосходит правое, когда задача состоит в выявлении немногих четких деталей, а правое доминирует при интеграции элементов в сложные конфигурации. Это различие согласуется с клиническими данными. При патологиях правого полушария рисунки больных утрачивают целостность общей конфигурации. При поражении левого полушария основная конфигурация объекта обычно воспроизводится, но рисунок обеднен деталями. «Пространственное» правое и «временное» левое полушарие вносят каждое важный вклад в большинство видов когнитивной деятельности. По-видимому, у левого полушария больше возможностей во временной и слуховой областях, а у правого — в пространственной и зрительной.
Следует отметить, что каждое полушарие, функционируя изолированно, предпочитает формировать целостное изображение. Это особенно очевидно у больных с «расщепленным мозгом». Когда такому больному, фиксирующему точку на экране, предъявляли
рисунки-химеры (изображения, составленные из половинок двух разных объектов) и спрашивали, что он видит, он называл объект, соответствующий правой части химерного рисунка, проецирующейся в левое полушарие. Неудивительно, что он называл правый «полуобъект», так как у подавляющего большинства правшей за речь ответственно левое полушарие. Однако «говорящее» левое полушарие совершенно «не осознавало», что ему предъявляется только половина стимула. Когда же после этого опыта испытуемому в условиях свободного зрения (без фиксации определенной точки) предъявляли целые изображения тех же объектов и просили показать, какой из них он видел раньше, он почти всегда выбирал тот предмет, который раньше находился слева и воспринимался правым полушарием (Леви Д., 1995). Не умея «говорить», правое полушарие без слов демонстрировало, что воспринимает половину стимула как целый объект.
Нормальные люди в этих условиях сразу видят необычную, составную природу рисунков. При выполнении других тестов — с абстрактными фигурами, цветовыми стимулами — получены аналогичные результаты: у человека с «расщепленным мозгом» объект -воспринимается одним полушарием и всегда нерасчлененным.
Таким образом, другое полушарие в этих экспериментах ведет себя так, как будто оно «ничего не видит». С каким полушарием это может произойти, зависит от решаемой задачи. При этом неполнота стимула не означает такой же неполноты восприятия. Отвечающее полушарие (как левое, так и правое) интерпретирует изображение как целое, хотя предъявляется только его половина. Мозг строит модели целостного мира, и когда нет полной информации, как у больных с «расщепленным мозгом», они создаются на основе интеграции сенсорных данных с информацией, извлекаемой из памяти, со знаниями, которые не позволят объекту расщепляться на две половинки.
Существует точка зрения, согласно которой в функциях различных полушарий представлены различные способы познания. Функции левого полушария отождествляются с осознанными, логическими процессами мышления. Функция правого полушария — это интуитивное мышление. По мнению Р. Орнстейна (R. Ornstein), сегодня принятая система образования строится исключительно на развитии у детей способностей левого полушария, т.е. языкового и логического мышления, а функции правого полушария специально не развиваются. Невербальному интеллекту не уделяется должного внимания.
Интересную гипотезу развивает Д. Кимура (Kimura D., 1992). Исходя из того, что речевая функция левого полушария связана с движениями ведущей правой руки, она предполагает, что речевая специализация левого полушария является следствием не столько асимметричного развития символических функций, сколько развития определенных двигательных навыков, которые помогают в общении. Язык появился потому, что левое полушарие оказалось приспособленным для некоторых видов двигательной активности.
Связь левого полушария с определенными типами движения хорошо известна в клинике. Рука, соответствующая полушарию с речевым центром (чаще правая), обнаруживает большие способности к тонким движениям, чем рука, связанная с недоминантным полушарием. Больные с повреждением левого полушария и без правостороннего паралича тем не менее испытывают затруднения в воспроизведении сложной последовательности движений рук и сложных позиций пальцев. У глухонемых поражение левого полушария сопровождается распадом языка жестов, что сходно с распадом речи у нормально говорящих людей.
Д. Кимура полагает, что в эволюционном плане именно развитие руки как органа языка жестов, ее манипулятивных способностей и привело к формированию особых функций левого полушария. Способность руки к тонким манипуляциям была передана артикуляционным органам. В ее коммуникативной теории развития речи ведущая роль принадлежит не звуковым сигналам, а выразительным жестам.
Стремление понять, в чем состоит своеобразие взаимоотношений двух половин мозга в процессе творческой деятельности, побудило ученых к изучению особенностей организации полушарий у людей искусства. Была высказана гипотеза о повышенной способности творческих личностей к интеграции функций обоих полушарий (Леви Д., 1995). Исследования действительно подтвердили, что для обычных людей характерна более строгая латерали-зация функций полушарий при большей их билатеральное™ у художников. У художников-профессионалов на протяжении их творческой жизни каждая половина мозга (а не только правая) развивает структуры, формы и методы, необходимые для художественного творчества. Поэтому в случае повреждения одного из полушарий в зрелом возрасте второе сохраняет как свои врожденные художественные способности, так и приобретенные на основе взаимодействия с другим полушарием.
Исследование музыкантов тоже наводит на мысль о более двустороннем представительстве у них функций, важных для музыкальных способностей, по сравнению с не столь одаренными людьми. Восприятие мелодии включает образное представление громкости и высоты тонов, специфического звучания аккордов,
284
темпа и ритма. Доминирование того или другого полушария зависит от того, какому аспекту мелодии уделяется больше внимания. Так, хорошо знакомые мелодии могут кодироваться в виде целостного образа (гештальта), тогда как незнакомые мелодии требуют аналитического подхода. Установлено, что локализация активации в полушарии при прослушивании музыкальных произведений зависит от музыкальной грамотности слушателя. Более образованные в музыкальном отношении испытуемые, использовавшие аналитическую стратегию и умеющие обнаруживать сходство и различие звуков в аккордах, по данным ПЭТ, показывают большее потребление глюкозы левым полушарием. У лиц, не имеющих музыкального образования, прослушивание музыки усиливало метаболическую активацию (по глюкозе), особенно в теменных и затылочно-височных областях правого полушария.
Вместе с тем у одаренных музыкантов двустороннее представительство музыкальных способностей встречалось чаще, чем обычно бывает у менее талантливых музыкантов. Сведения о музыкантах с односторонним поражением мозга подтверждают, что у них, так же как у художников, соответствующие способности сохраняются лучше, чем у обычных людей. Известны случаи, когда после левостороннего инсульта композиторы продолжали успешно заниматься своей профессиональной деятельностью. Русский композитор В.Я. Шебалин успешно сочинял музыку и после левостороннего инсульта, вызвавшего у него тяжелую форму афазии. У обыкновенных людей различные аспекты их музыкальных способностей связаны с разными полушариями и неодинаково страдают при одностороннем поражении мозга. Немузыканты склонны воспринимать мелодии «глобально», т.е. в основном правым полушарием.
Музыканты-профессионалы, по-видимому, различаются между собой по степени использования способностей правого и левого полушарий, однако асимметрия в восприятии тонов, силы звука, аккордов, темпа и ритма у них, похоже, значительно меньше, чем у обычных людей. Накопленный у людей искусства творческий опыт усиливает структурно-функциональное сходство и взаимодействие их полушарий.
ПОЛОВЫЕ РАЗЛИЧИЯ
И ИНТЕЛЛЕКТУАЛЬНЫЕ ФУНКЦИИ
Поведенческие, неврологические и биохимические исследования проливают свет на те процессы, которые порождают половые различия в организации мозга. Половые гормоны начинают влиять на мозг в таком раннем возрасте, что реакция на воздействие внешней среды по-разному смонтированного мозга у мальчиков и девочек существенно различается почти сразу после рождения. Влияние пола на интеллектуальные функции проявляется скорее в характере умственных способностей, а не в общем уровне интеллекта, измеряемого по IQ.
Мужчины лучше ориентируются в пути, следуя по какому-либо маршруту. Им требуется меньше времени на запоминание маршрута, они совершают меньше ошибок. Но после того как маршрут заучен, женщины помнят большее число дорожных ориентиров, чем мужчины. По-видимому, они склонны больше пользоваться наглядными ориентирами и в повседневной жизни.
Мужчины, как правило, лучше, чем женщины, решают пространственные задачи. Они лучше выполняют тесты, в которых требуется мысленно вращать предмет или каким-либо образом манипулировать им. Они превосходят женщин в тестах, требующих математических рассуждений (Кимура Д., 1992). У мужчин обнаруживаются большие способности к формированию точных двигательных навыков прицеливания, метания, перехвата различных снарядов.
Женщины, как правило, превосходят мужчин в быстроте идентификации сходных предметов, в арифметическом счете, у них лучше развиты речевые навыки. Женщины быстрее справляются с некоторыми мануальными заданиями, где требуются точность, ювелирность движений.
Поскольку генетический материал у мужчин и женщин, за исключением половых хромосом, одинаков, скорее разное качество умственных способностей у мужчин и женщин отражает различие гормональных влияний на развивающийся мозг. Разделение полов происходит на ранних этапах эмбрионального развития под влиянием эстрогенов (женских половых гормонов) и андрогенов (мужских половых гормонов, главный из которых — тестостерон).
Тестостерон вызывает маскулинизацию, способствует формированию мужских половых органов, а также уже на ранних этапах жизни формирует стереотипы мужского поведения. Половые гормоны изменяют функционирование мозга только во время критического периода развития человека. Введение тех же самых гормонов в более поздние периоды жизни таких эффектов не вызывает. Лишение новорожденных самцов тестостерона путем кастрации или введение новорожденным самкам андрогенов приводит в зрелом возрасте к полному изменению специфических для данного пола форм поведения на противоположный. Крысы-самки, которым вводят андрогены, ведут себя как самцы. Они становятся более агрессивными, склонными к играм с элементами драки, пред--
почитают грубый физический контакт. Кастрированные самцы ведут себя, как самки. При этом у них обнаруживают характерную /1ля самок склонность к использованию наглядных ориентиров при выполнении заданий, связанных с пространственным обучением.
Изучение особенностей поведения и способностей девочек, которые в пренатальной или неонатальной жизни подверглись избыточному воздействию андрогеном (мужским половым гормоном) из-за врожденной гиперплазии надпочечников их матерей, показало, что, взрослея, они проявляли более выраженное мальчиковое поведение и большую агрессивность. Эффект был необратимым и не корректировался медикаментозной терапией. У них, так же как и у мужчин, лучше развиты пространственные функции. Они лучше выполняют тесты на пространственную манипуляцию, вращение предметов. Однако никаких различий в других перцептивных или вербальных тестах, требующих логических рассуждений, между двумя группами девочек — с гормональными нарушениями и без них — выявлено не было.
Многие ученые полагают, что в основе различия мужского и женского ума лежит менее выраженная у женщин асимметрия полушарий по сравнению с мужчинами. У женщин повреждение одного мозгового полушария чаще, чем у мужчин, вызывает меньший дефект, такая же травма у мужчин сопровождается более выраженными последствиями. Есть данные о том, что задняя часть мозолистого тела у женщин больше. Это должно указывать на более полное взаимодействие полушарий у женщин по сравнению с мужчинами.
Установлено, что у крыс-самцов кора правого полушария толще левого. Это согласуется с другими данными о том, что раннее влияние мужского полового гормона (андрогена) ведет к подавлению роста коры левого полушария.
Наблюдение за эмбрионами человека также показало, что у будущих мальчиков кора правого полушария толще левого. Однако, как вытекает из результатов обследования мужчин и женщин с поврежденным правым полушарием, способность к пространственному вращению предметов, которая лучше выражена у мужчин, не обусловлена половыми функциональными различиями правого полушария. Повреждение правого полушария не вызывало у мужчин более выраженных нарушений пространственного вращения, чем у женщин, как это можно было ожидать, если бы такие способности у мужчин определялись большей развитостью правого полушария.
Сходное предположение о более выраженной асимметрии мозга у мужчин высказано и в отношении речи. При этом исходили из того факта, что афазия чаще встречается у мужчин после травмы левого полушария. На этом основании и было сделано заключение, что у женщин оба полушария принимают большее участие в организации речи. Однако полученные некоторыми авторами данные опровергают это мнение: среди женщин с повреждением правого полушария афазия встречается столь же часто, как и среди мужчин с аналогичной травмой.
Д. Кимура (1992), исследуя нарушения речевых функций у мужчин и женщин в результате поражений мозга, установила, что у женщин организация речи и связанные с ней двигательные функции локализованы в левой лобной коре.
У мужчин центр с аналогичными функциями находится в задних отделах того же полушария. После повреждения лобной части мозга у женщин афазия развивается чаще, чем у мужчин. При повреждениях же задних отделов мозга (обычно эта травма встречается чаще, чем поражения в передних отделах) речевые функции женщин страдают реже не потому, что у них менее выражена асимметрия мозга, а потому, что у них реже подвергается разрушению центр организации речевых движений, локализованный в передних областях мозга. У мужчин же система выбора и программирования речевых движений находится в задних отделах полушария.
По мнению Д. Кимура, специфика левого полушария — это не только программирование и выбор речевых реакций, но и организация сложных движений рта, рук, участвующих в общении людей. Эти функции у женщин представлены в передних областях, а у мужчин — в задних отделах полушария.
У женщин система «праксиса», обеспечивающая выбор надлежащих движений руки, находится в топографической близости к расположенной сразу же позади нее моторной коре, что может объяснять способность женщин формировать более тонкие двигательные навыки. Напротив, у мужчин лучше формируются движения типа прицеливания, т.е. направленные на объекты, находящиеся на некотором расстоянии. Для этих навыков необходимо тесное взаимодействие со зрительной системой, локализованной в задних участках полушарий.
По данным Д. Кимура, передняя система контроля моторики женщин выявляется даже в тестах, требующих одновременного участия зрительной информации (построение по зрительной модели фигуры из кубиков). У женщин при выполнении данного теста обнаруживаются большие нарушения, когда повреждаются передние, а не задние отделы полушарий. У мужчин наблюдается обратная зависимость.
Хотя функциональная асимметрия мозга, кажется, не влияет на организацию речи и движений, а.также на способность к про-
288
странственному вращению, однако похоже, что от нее зависит
выполнение некоторых абстрактных вербальных заданий. На выполнение теста для
оценки словарного запаса у женщин влияли повреждения обоих полушарий, а у
мужчин только левого. Другими словами, женщины при осмысливании слов в большей
степени, чем мужчины, используют оба полушария. В то же время двигательные
навыки мужчин в меньшей степени зависят от левого полушария, так как среди них
чаще встречаются левши. Среди правшей женщины отличаются большей праворукостью,
чем мужчины: они чаще, чем мужчины, предпочитают пользоваться правой рукой.
Таким образом, асимметрия мозга, связанная с половыми различиями,
в зависимости от функции может выражаться в доминировании разных полушарий.
Поэтому не всегда более «асимметричным» является какой-то один пол. Так, у
женщин успешное выполнение вербальных заданий связано с активностью доминантного
левого полушария. То же можно сказать и о большем проценте среди них
праворукости. У мужчин двигательные навыки находятся в меньшей зависимости от
левого полушария.
Как следует из приведенных данных, организация головного мозга у
мужчин и женщин с самого раннего возраста идет по разному пути. Эту
дифференцировку развития направляют половые гормоны, что и формирует различные
когнитивные способности у представителей разного пола. Когнитивные операции
сохраняют свою чувствительность к половым гормонам на протяжении всей жизни.
Уровень эстрогенов, меняющийся в течение менструального цикла, влияет на
когнитивные процессы. Высокий уровень этих гормонов сочетается с относительным
ухудшением пространственных способностей и улучшением двигательных и
артикуляционных навыков. У мужчин существуют сезонные колебания тестостерона.
При некотором оптимальном его уровне в крови мужчины проявляют максимальную
способность к решению пространственных задач. Наилучшие результаты у них
отмечаются весной, когда уровень тестостерона ниже.
35.
механизмы творческой деятельности.
Многие представители творческих профессий — ученые, изобретатели,
писатели — отмечают, что важные, критические этапы в их деятельности носят
интуитивный характер. Решение проблемы приходит внезапно, а не в результате
логических рассуждений. Творчество в своей основе представлено механизмами
сверхсознания (Симонов П.В., 1975). Если сознание вооружено речью,
математическими формулами и образами художественных произведений, то язык
сверхсознания — это чувства, эмоции. Творческий процесс приводит не только к
расширению сферы знания, но и к преодолению ранее существовавших, принятых
норм.
Выделяют три основных этапа творческого процесса: замысел,
рождение догадки; генерацию различных гипотез, включая самые фантастические,
для объяснения данного явления; критический анализ и отбор наиболее
правдоподобных объяснений, которые протекают на уровне сознания.
Озарение, открытие, нахождение пути решения проблемы возникают в
виде переживания, чувствования, что выбранное направление — то самое, которое
заслуживает внимания. И здесь решающая роль принадлежит чувству, интуиции — языку
сверхсознания. Многие изобретатели отмечают, что догадка возникает в виде
расплывчатого образа, который только еще должен быть выражен словами. Однако
внезапность появления догадки, озарения — кажущаяся, так как она является
следствием интенсивной мыслительной работы человека, поглощенного увлекающей
его проблемой или художественным произведением.
По мнению Р.А. Павлыгиной и П.В. Симонова, доминанта имеет
отношение к явлениям озарения, инсайта, составляющим центральное звено
творческого процесса. Внезапное отключение доминантного состояния может
приводить к внезапному замыканию ассоциаций (установлению неожиданных
связей). В опытах на кроликах показано, что при голодной доминанте,
создаваемой естественной пищевой депривацией, любое побочное воздействие,
включая обдувание глаза воздухом, вызывает не только мигательную, но и
жевательную реакцию. Если голодному животному непосредственно после обдувания
воздухом глаза дать пищу и тем самым снять доминантное состояние, то это
приводит к формированию стабильного инструментального рефлекса. При повторном
воспроизведении такой же доминанты кролик стремится регулировать свое
состояние, демонстрируя мигательную реакцию, которая только один раз
сопровождалась устранением доминанты.
Другое явление, которое также имеет отношение к творческому
мышлению, — это установление ассоциации между подпороговы-ми раздражителями.
Совмещение подпороговой стимуляции лапы и круговой мышцы глаза приводило к
формированию связи между мигательной реакцией и движением лапы (Павлыгина Р.А.,
1990). Ее можно было выявить, заменив подпороговые раздражения над-пороговыми:
стимуляция лапы вызывала мигательную реакцию, а раздражение глаза
сопровождалось двигательной реакцией конечности (двусторонняя связь, по Э.А.
Асратяну).
Таким образом, доминанта сильно напоминает мотивацион-пое
состояние, во время которого на основе видового и приобре-юнного опыта
актуализируются ассоциации между стимулами, а 1акже между стимулами и
реакциями. В процессе анализа этой информации могут быть выявлены ранее
скрытые (подпороговые) связи, которые приведут к новому видению проблемы. Явление
внезапного формирования устойчивых ассоциаций в результате устране-пия
доминантного возбуждения исследователи рассматривают как нейрофизиологический
механизм творческого озарения.
Творчество— создание нового из старых элементов во внутреннем
мире. Создание нового продукта вызывает положительную эмоциональную
реакцию. Это положительное эмоциональное состояние служит наградой творческому
процессу и стимулирует человека действовать в том же направлении.
Выявление нового аспекта в когнитивных процессах обусловлено
работой детекторов новизны, которые способны улавливать но-иое не только во
внешнем, но и во внутреннем мире — новые мыс-пи, новые образы. Ориентировочная
реакция возникает при этом не па изменение внешнего сигнала, а на
преобразование внутреннего образа. При этом она сопровождается положительным
эмоциональным переживанием и сама является эмоциональным подкреплением.
Детекторы новизны обладают высокой чувствительностью, они сразу же фиксируют
факт появления новой мысли еще до того, как она будет оценена. Осознание
появления новой мысли сопровождается творческим волнением, которое стимулирует
умственную работу. И только после появления эмоциональной реакции мысль
начинает критически оцениваться. Таким образом, неосознанное сопоставление
разного рода информации, содержащейся в памя-i и, порождает новую мысль. Ее последующая оценка осуществляется
путем сравнения этой мысли с другими, ранее уже осознанными. Следовательно,
продукция нового осуществляется главным образом в подсознании, а его оценка —
на уровне сознания.
Процессы творческого мышления могут быть рассмотрены с юч ки
зрения взаимоотношения ориентировочного и оборонительного рефлексов. Известно,
что стресс с высоким уровнем напряженности выражает защитную, оборонительную
реакцию, которая дезорганизует когнитивные функции человека. Согласно закону
Йеркса—Додсена существует так называемое оптимальное /функциональное
состояние, определяющее наиболее высокую эффек-i инность деятельности. Изучение механизма оптимизации функционального
состояния приводит к представлению о его связи с ориен-i ировочным рефлексом. Наличие интереса, увлеченность работой — i с предпосылки, которые определяют уровень ее успешности.
Творчество связано с развитием потребности в познании, в получении
новой информации, которая достигается в процессе ориентировочно-исследовательской
деятельности. Последнюю можно рассматривать как цепь ориентировочных рефлексов.
Каждый из ориентировочных рефлексов обеспечивает получение определенной порции
информации.
Творческое мышление — это ориентировочно-исследовательская
деятельность, обращенная к следам памяти в сочетании с поступающей актуальной
информацией.
Ориентировочный рефлекс как выражение потребности в новой
информации конкурирует с оборонительным рефлексом, который является
выражением агрессии или страха, тревоги (Gracham F.K., 1997; Данилова Н.Н., 1995).
Особыми формами оборонительного поведения являются депрессия и
тревожность, которые, тормозя ориентировочно-исследовательскую деятельность,
снижают творческие возможности человека. Депрессия и тревожность могут
возникать под влиянием длительного неуспеха в преодолении конфликтных ситуаций.
Развиваясь, они ведут к соматическим нарушениям, которые, образуя контур
обратной положительной связи, еще больше углубляют депрессию и тревожность.
Разорвать этот круг самоусиления пассивно-оборонительного поведения,
приводящего к снижению творческих возможностей человека, можно лишь путем
устранения конфликтов и оказания психотерапевтической помощи. В качестве
основы «креативной психотерапии» можно рассматривать создание у индивида
творческой установки, усиление его ориентировочно-исследовательской
деятельности, которые обычно тормозят оборонительную доминанту, способствуя
раскрытию творческих способностей. Такая креативная установка может быть элементом
процесса непрерывного образования человека, в силу того, что она стимулирует
его заинтересованность в получении новой информации.
Ориентировочный рефлекс находится в реципрокных отношениях не
только с пассивно-оборонительной, но и с активно-оборонительной формой
поведения — аффективной агрессией. Длительные психологические конфликты могут
вызывать функциональные изменения, выражающиеся в понижении порога аффективной
агрессии. В результате незначительные воздействия провоцируют агрессивное
поведение. Такое снижение порога агрессивного поведения иногда наблюдается в
период полового созревания в результате нарушения медиаторного баланса. Одним
из радикальных способов снижения агрессивности может быть стимуляция ориентировочно-исследовательской
деятельности.
Таким образом, стимуляцию ориентировочно-исследовательской
активности можно рассматривать как основу развития творческого потенциала
человека и психотерапевтический способ подавления депрессии, тревожности и
агрессивности — основных факторов, препятствующих творческому самовыражению
человека.
Реципрокный характер взаимоотношений ориентировочного и
оборонительного рефлексов наиболее отчетливо проявляется в реакциях сердечного
ритма. Тип реакции в виде редукции мощности сосудистой и дыхательной модуляций
СР, роста ЧСС, индекса напряжения (ИН), уменьшения величины стандартного
отклонения RR-интервала выражает
оборонительный рефлекс. Противоположный тип реакции характеризует
ориентировочный рефлекс: одновременно с падением ЧСС (одним из компонентов
ориентировочной реакции) усиливается дыхательная и сосудистая модуляция RR-интервалов. Параллельно падает ИН, а
стандартное отклонение, характеризующее вариабельность СР, растет.
Появление ориентировочного или оборонительного рефлекса в составе
когнитивной деятельности зависит от типа информационной нагрузки и ее
сложности, а также от индивидуальных особенностей субъекта и прежде всего от
его личностной тревожности, устойчивой индивидуальной склонности более или
менее часто отвечать на различные события как на потенциально опасные.
Изменение СР по оборонительному типу преобладает при решении арифметических
задач. Последние часто создают стрессовую ситуацию, так как многими людьми
процедура выполнения арифметического теста связывается с оценкой их
интеллектуальных способностей. Лица с высоким уровнем личностной тревожности
реагируют даже на стандартную информационную нагрузку преимущественно
оборонительными реакциями в виде редукции сосудистой и дыхательной модуляций СР
совместно с ростом ЧСС и ИН. Стресс, тревожность тормозят ориентировочную
реакцию и усиливают оборонительные, что приводит к подавлению творческой
активности человека (Данилова Н.Н., 1995).
Рассматривая нейроанатомические основы творческого мышления, П.В.
Симонов связывает его с функциями следующих структур мозга. Ядра миндалины выделяют
доминирующую мотивацию, которая стимулирует поиск недостающей информации, необходимой
для решения определенной проблемы. Другая структура лим-6 и чес кой системы — гиппокамп—
обеспечивает расширенную актуализацию следов, извлекаемых из памяти и
служащих материалом для формирования гипотез. У человека гиппокамп
доминантного полушария вовлечен в анализ следов словесных сигналов, а правого
полушария — в обработку следов от невербальных стимулов.
Предполагается, что сами гипотезы генерируются в лобных отделах
неокортекса. В правом полушарии происходит их первичная эмоционально-интуитивная
оценка, при этом исключаются заведомо нереальные предположения. Левые лобные
доли выступают в роли критика, который выполняет отбор гипотез, наиболее достойных
внимания. Взаимодействие правых и левых лобных долей обеспечивает тот
диалог двух голосов— фантазирующего и критического, который знаком почти всем
творческим личностям. «Функциональная асимметрия двух полушарий мозга, в
сущности, служит сегодня наиболее приемлемой нейробиологической основой взаимодействия
осознаваемых и неосознаваемых компонентов творческого процесса» (Симонов П.В.,
1993. С. 75).
Механизмы интуиции при решении различного рода когнитивных задач
с учетом межполушарного взаимодействия были исследованы Н.Е. Свидерской
(1997). Используя метод компьютерной топоскопии синхронных биотоков мозга при
одновременном отведении ЭЭГ от 48 электродов, она определяла фокусы максимальной
активности во время решения задач, требующих различных способов обработки
информации: симультантного и сукцес-сивного. Симультантный метод служит
для одновременного анализа множества элементов информации. Его связывают с
функциями правого полушария. Сукцессивный метод представляет поэтапную
обработку информации и относится в основном к деятельности левого полушария.
Оказалось, что при решении вербальных и невербальных задач фокус активности
определяется не качеством или содержанием информации, а способом ее анализа.
Если задание требовало сукцессивного метода, фокус активности возникал в
передних областях левого полушария, а при выполнении симультантных заданий он
был локализован в задних зонах правого полушария. При решении нестандартных
задач, при незнании их алгоритма, когда требуется использовать интуитивные
формы мышления, активация доминирует в задних отделах правого полушария.
Такую же картину можно было видеть у испытуемых, правильно описывающих характер
и условия жизни человека по его портрету или местности по ее отдельным
фрагментам. Успешное выполнение такого задания возможно только на основе интуитивной
оценки. У субъектов, которые давали неправильное описание человека и
местности, фокус активности возникал в передних областях левого полушария.
Автор связывает правополушарный фокус активации с симультантным способом
обработки как осознаваемой, так и неосознаваемой информации.
Вместе с тем симультантный способ обработки, позволяющий
оперировать одновременно большим число элементов — целостным
294
представлением объекта, более адекватен для работы с неосознаваемой
информацией. Установлено, что при автоматизации навыка (обучении цифровым кодам
ЭВМ), т.е. при переходе от осознаваемого уровня анализа к неосознаваемому,
фокус активации из передних областей левого полушария смещается в задние зоны
правого.
Снижение уровня осознания болевой стимуляции, вызванной
гипнотической аналгезией, коррелирует с падением активности в передних зонах
левого полушария. Левополушарный фокус активности указывает на
сукцессивный способ обработки информации, предполагающий анализ материала на
осознаваемом уровне.
Совместная деятельность обоих полушарий, каждое из которых
использует свои методы обработки информации, обеспечивает наиболее высокую
эффективность деятельности. С усложнением задачи необходимо объединение усилий
обоих полушарий, в то время как при решении простых задач латерализация фокуса
активности вполне оправдана. При решении нестандартных, творческих задач
используется неосознаваемая информация. Это достигается совместной
деятельностью обоих полушарий при хорошо выраженном фокусе активности в задних
отделах правого полушария.
36. Понятие сознания.
Стадии восстановления сознания.
О том, что представляет собой сознание, высказывается большое
разнообразие мнений. Можно определять сознание как субъективно переживаемую
последовательность событий, противопоставляемых бессознательным процессам,
например таким, которые возникают в вегетативной нервной системе. Часто
сознание связы-нают с осведомленностью человека о том, что с ним происходит или
что он воспринимает. Осведомленность не является общим свойством всех процессов
обработки информации. Принято считать, что сознание отсутствует во время
медленного сна и у глубоко анестезированного животного. Однако парадоксальный
сон ряд исследователей относят к некоторым особым формам сознания. Хотя во
время сна когнитивные процессы не полностью нормальны и долговременная память
не функционирует, тем не менее сновидения, по-видимому, обладают какими-то
свойствами сознания (Crick
F., Koch Ch., 1990). При легкой анестезии
или состояниях,
вызываемых современными лекарственными препаратами,
подавляющими чувствительные к ним рецепторы, кажется, что сознание
также частично присутствует.
Среди разных позиций следует выделить два принципиально
различающихся подхода к определению сознания. Одно из них возникло в философии.
Сознание рассматривается как совокупность некоторых когнитивных операций,
связанных с субъективным переживанием своих мыслей, чувств, впечатлений и
возможностью передать их другим с помощью речи, действий или продуктов
творчества. Этот взгляд разделяется П.В. Симоновым, рассматривающим сознание
как совместное знание. Он пишет: «Сознание есть знание, которое с помощью
слов, математических символов и обобщающих образов художественных произведений
может быть передано, стать достоянием других членов общества, в том числе
других поколений, в виде памятников культуры» (Симонов П.В., 1993а. С. 213).
Э.А. Костандов в своих экспериментальных многолетних исследованиях
пришел к сходному заключению. Он подчеркивает, что «...решающим звеном в
структурно-функциональной системе мозга человека, организующей физиологическую
основу осознания раздражителей внешней среды, согласно предлагаемой нами
гипотезе, является активация временных связей между воспринимающими и
гностическими участками коры больших полушарий с двигательной речевой областью»
(Костандов Э.А., 1994. С. 902). Неосознаваемые процессы обработки информации, о
влиянии которых субъект не отдает себе отчета, принято относить к категории
бессознательного. П.В. Симонов (1987) предлагает выделять три группы проявления
бессознательного. Первая группа — досоз-нательное. Оно охватывает наши
биологические потребности, выражающиеся в безусловных рефлексах и врожденных
формах поведения (инстинктах), а также в генетически заданных свойствах темперамента.
Вторая группа бессознательного — подсознание. К ней относится
все то, что ранее уже было осознано и вновь может стать осознаваемым в
определенных условиях. Это различные автоматизированные навыки, стереотипы
автоматизированного поведения. К ним же относятся неосознаваемые побудители
деятельности (мотивы, смысловые установки), глубоко усвоенные человеком нормы
поведения, вытесненные из сферы сознания мотивационные конфликты. В процессе
эволюции подсознание возникло как средство защиты сознания от лишней работы и непереносимых нагрузок.
Оно предохраняет человека от излишних энергетических трат, защищает от
стресса. К подсознанию П.В. Симонов относит и те проявления интуиции, которые
не связаны с порождением новой информации, но предполагают лишь использование ранее
накопленного опыта. Примером может служить правильный диагноз, поставленный
опытным врачом при беглом осмотре пациента. Нередко врач не может объяснить,
какие именно симптомы побудили его прийти к такому заключению.
Указывая на различия сознания и подсознания, П.В. Симонов
отмечает: «Если сознание вооружено речью, символикой математических формул и
образным строем художественных произведений, неосознаваемое психическое
сообщает сознанию о результатах своей деятельности переживанием чувств, т.е.
эмоцией» (Симонов П.В., 1993а. С 215). Такая форма психического отражения, как
эмоция, не относится к категории осознаваемых явлений. Для перевода на уровень
сознания требуются дополнительные операции, связанные с обработкой символьной
информации.
Одной из наиболее характерных черт подсознания является его
консерватизм. В работах Э.А. Костандова экспериментально продемонстрирована
трудность угасания условных реакций, выработанных при подкреплении
неосознаваемых и эмоционально окрашенных стимулов (Костандов Э.А., 1983). Они
плохо поддаются воздействию тех психотропных веществ (диазепаму), которые влияют
на осознаваемые условные рефлексы (Арзуманов Ю.Л. и др., 1990). Усвоенные на
уровне подсознания условные рефлексы приобретают императивность, жесткость, присущую
безусловным рефлексам. Отсюда возникает иллюзия врожденности некоторых проявлений
бессознательного («голос крови», «классовый инстинкт»). Подсознательное из-за
своего консерватизма лишено творческого начала, которое предполагает
преодоление устоявшихся норм, правил (Симонов П.В., 19936).
Третью группу бессознательных явлений составляет сверхсознание,
или интуиция, связанная с процессами творчества, которые не контролируются
сознанием. Термин «сверхсознание» заимствован у К.С. Станиславского и идентичен
термину «подсознательное». Сверхсознание — источник новой информации,
гипотез, открытий. Под сверхсознанием понимается высший этап творческого
процесса. Его нейрофизиологическая основа — трансформация следов памяти и
порождение из них новых комбинаций, создание новых временных связей,
порождение аналогий. За сознанием остается функция отбора гипотез на основе их
логического анализа. Сверхсознание (интуиция) всегда «работает» на удовлетворение
потребности, устойчиво доминирующей в иерархии мотивов субъекта. В отличие от
подсознания деятельность сверхсозна-
ния не осознается ни при каких условиях. Сознание оценивает лишь
результаты деятельности сверхсознания. «Если подсознание защищает сознание от
излишней работы и психологических перегрузок, то неосознаваемость творческой
интуиции есть защита от преждевременного вмешательства сознания, от давления
ранее накопленного опыта» (Симонов П.В., 1993а).
Другое определение сознания имеет своим истоком экспериментальную
физиологию. Оно рассматривает сознание как определенное состояние
бодрствующего мозга или как уровень реактивности мозга. Данная точка
зрения предполагает существование различных уровней сознания — от глубокой
комы до полного бодрствования. В медицинской практике именно это содержание
вкладывается в понятие сознания. Появление движений глаз является одним из
признаков выхода пациента из комы. В НИИ нейрохирургии им. акад. Бурденко
выделяют 7 последовательных стадий восстановления сознания и психических
функций по поведенческим показателям:
1) открывание глаз; 2) фиксация взора и слежение', 3)
различение близких; 4) восстановление понимания обращенной речи; 5)
восстановление собственной речи; 6) амнестическая спутанность сознания;
7) восстановление формально ясного сознания.
Изучение угасания, выключения и восстановления сознания по
параметрам ЭЭГ показало высокую информативность среднего уровня когерентности
ЭЭГ для определения степени нарушения сознания (Добронравова И.С., 1996;
Болдырева Г.Н. и др., 1993). Выявлено несколько уровней угнетения сознания по
показателям среднего уровня когерентности ЭЭГ, по типу электрической
активности мозга и характеру ЭЭГ-реакций на сенсорные раздражители.
При угнетении сознания на стадии глубокой комы при общем снижении
среднего уровня когерентности ЭЭГ наблюдается преобладание правополушарной
асимметрии по показателю когерентности ЭЭГ преимущественно в центральной зоне
и в диапазоне тета-ритма (5 Гц). В условиях развития терминальной комы преобладает
левополушарная асимметрия ЭЭГ с большей представлен-ностью в лобной области и в
диапазоне медленных дельта-волн. Левополушарная асимметрия когерентности ЭЭГ в
диапазоне альфа-ритма преимущественно в височно-центральных отделах —
предвестник восстановления речевой функции. На разных стадиях угнетения
сознания наиболее сильным активирующим эффектом, благотворно влияющим на
процесс восстановления сознания, обладают эмоционально значимые раздражители
(слова, обращенные к пациенту). При летальном исходе сначала исчезает реакция
на свет, затем на звук и последней — на боль. С выходом из комы
восстановление ЭЭГ-реакций идет в обратном порядке.
Интересная гипотеза о сне, как особой форме сознания, принадлежит
И.Н. Пигареву (Pigarev I.N., 1994). Он полагает, что мозг
во сне не отдыхает, так же как не отдыхают и наши внутренние органы. И во сне
мозг продолжает выполнять стандартные операции над входными сигналами.
Пачечная активность нейронов, которую мы наблюдаем во сне, по мнению автора,
отражает их ответы на интероцептивную стимуляцию. Регистрируя электрическую
активность одиночных нейронов коры спящих обезьяны и кошки, он нашел, что
нейроны зрительной сенсорной и ассоциативной коры (VI, V2, V4), а также сенсомоторной области (поле 5) отвечают на
электрическую стимуляцию желудка и других внутренних органов. Таким образом,
структуры мозга, которые в бодрствовании обрабатывают информацию, поступающую
от экстероцептив-ных стимулов, во сне настраиваются на восприятие и обработку
интероцептивной информации. Обработка информации от эксте-роцептивных воздействий
и интероцепции осуществляется в одних и тех же структурах мозга, но не
одновременно. Информация о внешнем мире обрабатывается днем, а о состоянии
внутренней среды, внутренних органов — во сне.
Много исследований посвящено изучению взаимодействия коры и
внутренних органов. Во многих из них сообщается о вызванных потенциалах коры и
таламуса на электрическую стимуляцию внутренних органов, механические,
химические раздражения инте-роцепторов. Первичные ответы на висцеральную
стимуляцию (с латенцией около 30 мс) были получены в некоторых участках
соматосенсорной и ассоциативной коры. Вторичные ответы с большей латенцией (до
100 мс) практически можно записать от всей кортикальной поверхности. Однако все
реакции коры на раздражение внутренних органов были получены в острых опытах
на животных с анестезией. И их не удается зарегистрировать у неанестезированного
животного.
Таким образом, в бодрствовании результаты обработки
эксте-роцептивной информации достигают сознания и используются для организации
поведения. Во сне мозг обрабатывает сигналы от ин-тероцепторов и принимает
участие в гомеостатической регуляции (метаболизме, росте, регенерационных
процессах, иммунных ответах и др.). Однако эта активность мозга не достигает
того уровня сознания, когда мы это осознаем. И.Н. Пигарев предполагает, что
существует специальный механизм, который в бодрствовании блокирует обработку
висцеральных сигналов, а во сне — экстероцеп-тивных. Такая структура
аналогичным образом должна действовать
на моторные реакции. И такая блокада действительно известна как
мышечная атония во сне.
Кроме того, на анестезированных животных показано, что электрическая
стимуляция коры вызывает реакции в висцеральных органах. Они исчезают сразу же,
как только животное выходит из-под наркоза. Следовательно, управляющие
кортико-фугальные сигналы коры во сне направляются не к моторной системе, а к
внутренним органам. С пробуждением они меняют свою направленность,
переключаясь на регуляцию поведения и тесно связанную с ним моторную систему. Механизм,
который переключает эксте-роцептивные и интероцептивные потоки сигналов для
обработки информации, скорее всего должен быть локализован в центрах сна ствола
мозга (Pigarev I.N., 1997).
Сознание может быть широким или узким. Широту
сознания определяет количество каналов распространения локальной активации,
модулирующей информационные сети нейронов. Чем больше задействовано локальных
модуляторов, тем шире сознание. Частичное выключение локальных модуляторов
приводит к сужению поля сознания. Это можно наблюдать при гипоксии, когда в ЭЭГ
лобных отделов коры локально усиливаются медленные волны, сопровождающиеся
эйфорией и нарушением операций планирования.
В процессе развития сна могут быть выделены различные уровни
сознания. Исследование активности ассоциативных зон коры у кошки и обезьяны,
когда они выполняли сложное задание — опознание целевого стимула с отсроченной
реакцией, показало, что сон развивается в коре парциально, охватывая прежде
всего ассоциативную кору (Pigarev
I.N., 1997). В этих опытах животному в центре экрана демонстрировали
одну линию, после паузы 0,5— 1,5 с давали тестовый стимул, содержащий набор
линий, среди которых могла быть или не быть линия-образец, которая перед этим
была показана. Животное должно было сообщать о присутствии или отсутствии
образца в составе тестового стимула, нажимая на одну из двух педалей.
Правильный ответ подкреплялся небольшой порцией детского молочного питания.
Иногда животное впадало в дремотное состояние, особенно если получало много
молочного пищевого подкрепления, однако при этом оно продолжало правильно
выполнять задания. У нейронов ассоциативной зрительной коры (в области V4) такого животного можно было наблюдать пачечный тип активности,
характерный для сна, и ослабление или даже полное исчезновение их реакций на
зрительные стимулы, с которыми животное работало, решая задачу. В отличие от
нейронов ассоциативной коры нейроны первичной зрительной коры (в об-
ласти VI)
сохраняли нормальные ответы на зрительные стимулы даже при появлении ЭЭГ-признаков
сна в ассоциативной коре.
Характер изменений нейронной активности в ассоциативной коре
позволяет предположить существование локального сна. Последний прежде всего
захватывает нейроны, рецептивные поля которых находятся на периферии
зрительного поля. Таким образом, даже в пределах зрительной коры сон не
развивается синхронно. Процесс распространяется в направлении от периферии к
центру ассоциативной коры, возможно, возникая в отдельных модулях.
При развитии локального сна в ассоциативной коре обезьяна
продолжала выполнять задание на зрительный поиск с отставленной реакцией.
Таким образом, даже при высокой поведенческой активности возможен частичный
сон, особенно в ассоциативных корковых областях. По-видимому, это может быть
причиной возможного нарушения когнитивных процессов и внимания при наличии
большой потребности в сне.
Е.Н. Соколов (1997) предлагает объединить оба подхода к определению
сознания и рассматривать сознание как специфическое состояние мозга,
позволяющее осуществлять определенные когнитивные операции. Что касается
возможности передавать информацию другому, это требование не является
непременной характеристикой сознания. Но это не значит, что язык не обогащает
сознание, и возможность передать свои знания другому не отражает наиболее
высокий его уровень.
37. Теории
сознания. Осознавание ощущений по схеме Иваницкого.
Существует множество теорий о механизмах сознания, в которых
предпринимается попытка сформулировать необходимые и достаточные условия для
возникновения сознания. Их можно разделить на структурные, когда акцент
делается на рассмотрение роли отдельных структур или нейронных сетей мозга, и
на функциональные, которые определяют сознание через специальные когнитивные
операции — мышление, воображение, запоминание и желание. На деле эти два
подхода не исключают друг друга, так как специальные операции, связанные с
сознанием, реализуются с участием особых нейронных сетей, находящихся в
конкретных структурах мозга.
Изучая процессы концентрации и индукции возбуждения, их
распределение по коре, И.П. Павлов создал теорию сознания, которая получила
название теории светлого пятна. Он связывал сознание с фокусом
возбуждения, светлым пятном, областью повышенной возбудимости, которая может
перемещаться по коре.При этом сознание не обязательно связано с символьными
операциями — второй сигнальной системой и речевой деятельностью. По И.П.
Павлову, сознание есть у человека и у животных.
Изучая поведение собак в условиях свободного их перемещения в
манеже, И.С. Бериташвили выявил эффект сенсорного обучения. Он нашел,
что образ целевого объекта — пищи и места ее нахождения — формируется быстро,
фиксируется и легко извлекается из памяти, когда нужно удовлетворить
биологическую потребность. В связи с тем что эти результаты не укладывались в
простую рефлекторную схему, он ввел понятие психонервной деятельности, которая
оперирует образами. Он предположил, что внешняя среда всегда сравнивается с ее
отражением — репрезентациями, хранящимися в памяти. Это характерно как для
человека, так и для животных. Поведение строится на основе психонервных
механизмов мозгового отображения. Формирование образов он связывает с функцией
звездчатых клеток. Они не посылают свои аксоны за пределы коры, так как
относятся к внутренним клеткам коры и имеют самостимулирующую обратную связь.
Возбуждение может циркулировать по цепочке звездчатых нейронов. Такая локальная
реверберация возбуждения рассматривается как основа для формирования сложного
образа. Звездчатые нейроны создают материальный субстрат для сознания и служат
механизмом для психонервной деятельности, когда нельзя разделить психическое и
физиологическое.
Теория сознания Дж. Экклса (Eccles J., 1994) исходит из особой
функции дендритов пирамидных клеток коры. Наличие системы пирамидных нейронов
является характерной чертой неокор-текса. Находясь в нижних слоях коры, они на
уровне IV слоя коры собираются в
дендритный пучок, который достигает I слоя. Дж.
Экклс предположил, что субъективный (психический) феномен, выявляемый
интроспективными методами, которому он дал название «психон», связан с
пучком дендритов пирамидных клеток, идущим к поверхности коры. Пучок дендритов
от группы 70—100 соседних крупных и средних пирамид был назван «дендро-ном».
Каждый дендрон обладает множеством синапсов, на которых оканчиваются бутоны
терминалей аксонов. Следует подчеркнуть, что открытия, сделанные в последнее
время, указывают на особые, сложные функции, выполняемые дендритами. В шипиках
дендритов обнаружен свой собственный аппарат транскрипции — синтеза белков,
важных не только для восстановления функций нейрона, но и для его пластических
модификаций. Информационная РНК переносится из ядра к определенному
постсинаптичес-кому участку дендрита, где осуществляется синтез новых белков и
встраивание их в мембрану. Дж. Экклс приписал дендрону функцию носителя единицы
сознания. Отдельному дендрону соответствует отдельный психон как единица
локального ощущения. Множество нсихонов представляет все разнообразие
субъективных явлений. Сознание есть следствие психонов, генерируемых
соответствующими дендронами. Его теория сознания сводится к установлению
связей между психонами и отдельными дендронами. Его дуалистическая позиция
выражается в том, что он признает существование нематериального начала, которое
может воздействовать на денд-риты. Нематериальная передача осуществляется за
счет выброса кванта медиатора. В концепции сознания Дж. Экклса предполагается
активное влияние психического феномена на поведенческие акты в виде «свободы
воли». Влияния реализуются через управление вероятностью высвобождения квантов
медиатора.
Многие теории сознания исходят из принципа повторного входа
возбуждения в систему, образующую материальную основу сознания. Широкую
известность получила теория повторного входа, которую предложил и
теоретически разработал Дж. Эдельман, (см.: Эдельман Дж., Мауткастел В., 1981).
Позже эта теория получила экспериментальное и теоретическое развитие в работах
A.M. Иваницкого (1997).
Дж. Эдельман предположил, что элементарные процессы сознания
являются прерывистыми, т.е. требуют циклического повторения некоторой
последовательности событий при наличии повторного входа (reetrening) в одну и ту же группу клеток.
Это означает, что система сознания построена таким образом, что сигнал,
возникший внутри некоторой системы, входит в нее повторно, как если бы он был
внешним сигналом. Распространяя этот принцип на работу нейронных сетей, автор
предположил, что сенсорная информация от внешней и внутренней среды после ее
предварительной обработки в первичной нейрональной группе повторно входит в
нее, возвращаясь после дополнительной обработки в других группах клеток. При
этом параллельно в эту же группу нейронов поступает информация в виде
ассоциированных сигналов из долговременной памяти. Соединение этих двух потоков
информации и составляет один цикл, или повторяющуюся фазу, активности
сознания. Обсуждая возможную продолжительность каждого цикла, он предполагает,
что она равна 100 мс. Важной чертой теории сознания Дж. Эдельмана является не
только положение о повторном входе возбуждения. Он подчеркивает связь сознания
с операциями обращения к долговременной памяти. Осознание возникает в
результате обращения групп нейронов высокого порядка к мультимодальным
ассоциативным структурам, заложенным в долговременной памяти на основе
прошлого опыта.
303А.М. Иваницкий (1997) в своей теории «информационный синтез
как основа ощущений» исходит из результатов собственных исследований связи
волн вызванного потенциала (ВП) с субъективными показателями восприятия
стимула. Количественная оценка восприятия в соответствии с теорией обнаружения
сигнала производилась по двум независимым переменным: показателю сенсорной
чувствительности (сГ) и критерию принятия решения, зависящего от мотивации
субъекта. Было показано, что амплитуда ранних компонентов ВП положительно
коррелирует с показателем d', а
поздние компоненты — с критерием принятия решения. Промежуточные волны ВП с
латенцией 140—180 мс коррелировали с обоими показателями восприятия, причем
отношения были выявлены только для проекционной коры. Факты двойной корреляции
этих среднелатентных компонентов ВП — с сенсорными свойствами стимула (d') и его значимостью (принятие решения) — послужили основой для
концепции о кольцевом движении нервных импульсов с «центральной станцией» в
проекционной коре. Сначала возбуждение от проекционной коры, вызванное
сенсорным стимулом, поступает к ассоциативной. Для зрительной модальности
ассоциативные отделы находятся в височной коре. Затем сигнал поступает к структурам
лимбико-гиппокампального комплекса и подкорковым центрам эмоций и мотиваций.
Оттуда возбуждение вновь возвращается в кору, включая ее проекционные отделы,
по системе диффузных проекций. Затем к обработке информации подключается и
лобная кора. На рис. 59 схематично представлен путь кольцевого движения
возбуждения при возникновении зрительного ощущения. Ведущее звено в этой схеме
— синтез информации о физических и сигнальных (значимых) свойствах стимула на
нейронах проекционной коры. Информация о значимости стимула извлекается из
памяти с помощью мотивационно-эмоциональной системы. Синтез двух видов
информации — наличной и извлекаемой из памяти — определяет появление ощущения.
Движение возбуждения по кольцу составляет психический мониторинг происходящих
изменений во внешней и внутренней среде. Этот процесс осуществляется с периодом
квантования приблизительно 100— 150 мс. У человека возникшее ощущение затем
опознается, кате-горизируется, что требует участия лобных отделов коры, включая
вербальные центры. Реакция последних возникает примерно на 200 мс позже
ощущения. Теории повторного входа возбуждения, отказываясь от понятия рефлекса,
трансформируют его в «кольцо», внутри которого циркулирует возбуждение.
«Прожекторная теория сознания» предложена Ф. Криком (Crick F., Koch
Ch., 1990). Ф. Крик — один из
авторов расшифров
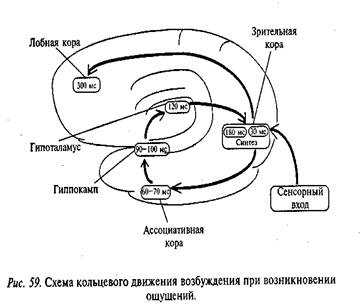
ки структуры ДНК, лауреат Нобелевской премии. В его теории идея
И.П. Павлова о сознании как светлом пятне получила дальнейшее развитие. Он
предположил наличие специального аппарата, создающего «луч прожектора», связав
его с особой формой внимания и гамма-осцилляциями в электрической активности
мозга. Кроме того, он указал на зависимость сознания от механизма некоторых
форм кратковременной памяти.
Модель сознания Крика построена на анализе работы зрения. С его
точки зрения, зрительная кора — наиболее удобный объект для изучения сознания.
У человека и животных она имеет сходную структуру.
Различные отделы зрительной коры отвечают на разные признаки
зрительных объектов. Нейроны-детекторы первичной зрительной коры (VI) реагируют на простые характеристики стимулов,
например на определенным образом ориентированную полоску. Нейроны кортикальных
областей более высокого порядка отвечают на более сложные признаки (на
определенные черты лица). Нейроны области V5 реагируют главным образом на движение зрительного стимула и не
отвечают на цвет и форму. В области V4 находят
нейроны, отвечающие на цвет и форму предметов, нейроны области 7а — на
положение стимула в пространстве относительно головы и тела. Но как возникают
целостные зрительныеобразы, сцены, которые мы видим перед собой? Отвечая на
этот вопрос, Ф. Крик предположил, что в любой момент определенные нейроны,
расположенные в разных кортикальных зонах, могут кооперироваться, чтобы сформировать
некоторый вид глобальной активности. Ее появление и соответствует зрительному
осознанию.
В качестве предполагаемого механизма, связывающего нейроны в
общую единую систему, он рассматривает одновременное появление у них коррелированных
разрядов с частотой гамма-колебаний (35—70 Гц). Он обратил внимание на то,
что нейроны, избирательно реагирующие на один и тот же стимул, обнаруживают
сходные гамма-осцилляции без фазового сдвига. Кроме того, корреляция их
гамма-активности при появлении в их рецептивных полях одного и того же объекта
была больше, чем на появление различных объектов. Все это позволило ему
утверждать, что синхронизация нейронной активности является механизмом объединения
клеток в ансамбль. Нейроны связываются в ансамбль за счет синхронизации их
активности на какое-то время. Они могут переключаться с одного ансамбля на
другой. Синхронизация активности нейронов зрительной коры постулируется как
способ пространственного связывания признаком.
Включение и выключение гамма-осцилляции зависят от механизма
последовательного (серийного) внимания, которое иногда называют прожектором
внимания. Оно характеризуется последовательным перемещением фокуса внимания с
одного места в зрительном поле на другое. По мению Ф. Крика, это движение более
быстрое, чем движение глаз, которое представляет другую форму — более
медленного внимания. Механизм внимания помогает группе взаимодействующих
нейронов разряжаться когерентно на частоте 35—70 Гц. В результате создается
глобальная единица активности, охватывающая нейроны в различных частях мозга.
38.
Асимметрия мозга и сознание.
Открытие функциональной асимметрии мозга оказало огромное влияние
на понимание мозговых механизмов сознания. С позиции вербальной теории сознания
его материальная база представлена доминантным речевым полушарием. Вместе с тем
имеются
основания выделять две функции сознания: чувство «Я» и
коммуникативную функцию сознания (Иваницкий A.M., 1997). Правое полушарие
связывают с самооценкой, с самосознанием (чувством «Я»), с мотивационной сферой
(Симонов П.В., 1993а). Его поражение ведет к нарушению личности, тогда как при
нарушении функций левого полушария личность остается сохранной, но речь
страдает. Специализацию левого полушария видят в его связи с той формой
сознания, которая вооружена речью и иными средствами передачи информации другим
членам общества. Она существенна прежде всего для коммуникации с другими
людьми. П.В. Симонов, подчеркивая коммуникативное происхождение сознания,
рассматривает возможность мысленного диалога с самим собой как свойство левого
полушария.
Вместе с тем было бы неоправданным приурочивать сознание,
связанное с речью, исключительно к левому (доминантному) полушарию у правшей.
Скорее всего начальные этапы формирования речевого высказывания (на родном
языке) связаны с правым полушарием. А завершается этот процесс в левом
полушарии.
Исследование становления речи у детей показало, что первоначально
обработка речевых сигналов осуществляется обоими полушариями и доминантность левого
формируется позже. Если у ребенка, научившегося говорить, возникает поражение
речевой области левого полушария, то у него развивается афазия. Однако примерно
через год речь восстанавливается. При этом центр речи перемещается в зону
правого полушария. Такая передача речевой функции от левого полушария правому
возможна лишь до 10 лет. Специализация правого полушария в функции ориентации в
пространстве возникает также не сразу: у мальчиков — в возрасте после 6 лет, а
у девочек — после 13 лет.
Данные о лингвистических способностях правого полушария, а также
сходство функций обоих полушарий на ранних этапах онтогенеза скорее
свидетельствуют о том, что оба полушария, обладая первоначально достаточно
сходными, симметричными функциями, в процессе эволюции постепенно
специализировались, что и привело к появлению доминантного и субдоминантного
полушарий.
Левое полушарие превосходит правое также и в способности понимать
речь, хотя эти различия не так сильно выражены. Согласно моторной теории
восприятия речи главным компонентом распознавания речевых звуков являются
кинестезические сигналы, возникающие в мышцах речевого аппарата при восприятии
речевых сигналов. Особая роль в этом принадлежит моторным системам левого
полушария.
Речевые функции у правшей преимущественно локализованы в левом
полушарии. По данным разных авторов, лишь у 1—5% лиц речевые центры находятся в
правом полушарии. У 70% леворуких центр речи, так же как и у праворуких,
расположен в левом полушарии, у 15% леворуких — в правом полушарии.
Существует мнение, что левое полушарие осуществляет функции
«интерпретатора» того, что делает правый мозг. Благодаря этой способности и при
отсутствии достаточной информации человек может логически сформулировать
некоторые гипотезы о причинах своих реакций, поступков, хотя они будут далеки
от истины. Это убедительно было показано Дж. ЛеДу (J. LeDu) и М. Газза-нигой
(М. Gassaniga) на их уникальном больном С.П. с «расщепленным мозгом» в опыте «с
цыпленком и лопатой для снега». Больному слева от точки фиксации предъявляли
картинку — зимний дворик с неубранным снегом, которая, следовательно,
проецировалась в правое полушарие. Одновременно справа от фиксационной точки
помещали картинку с изображением лапы цыпленка, т.е. эта картинка
проецировалась в левое полушарие. Больной получал задание отобрать левой и
правой рукой из серии картинок те, которые ему были показаны. Он успешно решил
эту задачу, правой рукой выбрав «цыпленка», а левой — «лопату». На вопрос
«Почему он это сделал?» ответил: «Я видел лапу и выбрал цыпленка, а вы должны
вычистить курятник лопатой». Анализируя эти данные, Дж. ЛеДу и М. Газзанига
пришли к заключению, что вербальные механизмы не всегда знают источник наших
действий и могут приписывать действиям причину, которой на самом деле не
существует.
Левое полушарие, не располагая информацией правого,
интерпретирует, находит логическое объяснение выбору левой рукой «лопаты».
Интерпретационная способность левого полушария при правостороннем поражении
мозга проявляется в изменении эмоциональных переживаний человека. Такие люди
становятся беспечными, легкомысленными, с явным преобладанием положительных
эмоций над отрицательными. Противоположные изменения эмоциональной сферы
сопровождают нарушения левого полушария. У таких больных возникает склонность к
отрицательным эмоциональным переживаниям, повышенной тревожности.
Нарушение сознания при избирательном поражении одного из полушарий
зависит от особенностей функциональной асимметрии мозга субъекта. Картина
нарушения психической деятельности зависит от правшества и левшества
(Доброхотова Т.А., Браги-на Н.Н., 1993). При однополушарном поражении мозга
были выявлены существенные различия нарушения поведения у правшей
и левшей, что послужило основанием к заключению о различии у них
сознания. Исследователи вводят новый термин — «асимметрия сознания», отражающая
индивидуальные свойства субъекта, связанные с функциональной асимметрией мозга.
При одностороннем поражении полушарий нарушения сознания протекали по типу
эпилептических припадков: наступали внезапно, быстро обрывались с последующим
восстановлением ясного сознания. Синдромы, вызываемые односторонним поражением
у правшей, хорошо согласуются с представлением о связи правого полушария с
обработкой пространственной информации и способности к зрительному и
гаптическому распознаванию объекта, а левого — с обработкой символьной
информации, управлением и программированием поведения.
При поражении в правом полушарии у правшей во время приступа
наблюдалось игнорирование левого пространства (больной не видел, не слышал
врача, стоящего слева). При этом могли появиться «вспышки пережитого» (больной
заново переживал события далекого прошлого, например в возрасте 7 лет).
Поражение левого полушария у правшей вызывает нарушение речи (не говорит и не
понимает), провалы в памяти, неадекватные поступки с последующей амнезией,
сумеречное состояние сознания, когда поведение определяется галлюцинациями или
больной автоматически продолжает начатую деятельность. Нарушение сознания у
левшей менее определенно, более разнообразно и не так жестко зависит от стороны
поражения.
Как показало обследование больных с «расщепленным мозгом», каждое
полушарие может выносить самостоятельные суждения, которые в ряде случаев могут
не совпадать. От согласованности оценок, выносимых полушариями, зависит
настроение человека. При совпадении суждений больной спокоен, легок в общении,
приятен. Если же оценки расходятся, поведение больного становится трудно
управляемым, проявляются гиперактивность, агрессивность.
Ввиду того что каждое полушарие может иметь собственную независимую
систему оценок значения событий, Р. Сперри (R. Sperry) говорит об удвоении
сознания у таких больных. Однако у нормального человека ведущая роль в
вынесении суждений принадлежит речевому, доминантному полушарию, хотя
определенные лингвистические способности присущи и субдоминантному полушарию.
В заключение следует подчеркнуть, что в сознании представлены
только конечные результаты процессов обработки информации, которые мы осознаем
в виде последовательных картин, эпизодов. Операции, лежащие в основе создания
самих эпизодов, в
сознании и интроспекции не представлены. Их можно вычленить с
помощью только специальных экспериментальных процедур. Но чтобы понять
сознание, нужно знать его механизмы, т.е. элементарные операции, которые лежат
в основе формирования субъективного образа и сами реализуются на уровне
подсознания. Чтобы понять, что такое сознание, нужно выйти за его пределы.
Прогресс в этом отношении достигается с помощью таких неинвазив-ных методов,
как электроэнцефалография, магнитоэнцефалогра-фия, измерение локального
мозгового кровотока, позитронно-эмиссионная и магнитно-резонансная томография.
В комбинации с записью одиночных нейронов эти методы открывают новые
перспективы в изучении сознания.
39. Педагогическая
психофизиология. Схема двухконтурного управления пр. обучения.
Проблема оптимизации обучения — центральная для педагогики.
Многочисленные школы и концепции обучения отражают непрерывный поиск и усилия,
предпринимаемые педагогами и психологами в этом направлении. Однако существует
чисто психофизиологический аспект решения данной проблемы. В настоящее время
управление процессом обучения осуществляется главным образом с учетом
результатов успеваемости, на основе тестирования успешности обучения. При этом
вне поля внимания остается вопрос о том, является ли выбранный педагогом режим
обучения оптимальным с точки зрения биологических критериев. Другими словами,
учитывается ли «биологическая цена», которую ученик платит за усвоение,
приобретение знаний, т.е. те энергетические затраты, которые сопровождают
процесс обучения и которые могут быть оценены через изменения функционального
состояния учащегося.
Большинство людей достаточно осведомлены о влиянии стресса и его
отрицательных последствиях. Мы хорошо знаем, что острый и хронический стресс
любой этиологии не только снижает результаты производственной и учебной
деятельности, но и влияет на развитие и протекание когнитивных процессов,
вызывает задержку в развитии ребенка, влияет на становление его как личности,
на формирование его индивидуального стиля деятельности.
Компьютеризация школ создает наиболее благоприятные условия для
контроля обучения по физиологическим параметрам. Эта задача может быть решена с
помощью мониторинга функциональ-
i~t/-t
Л
ного состояния учащегося, а также через компьютеризованную оценку
его индивидуальных психофизиологических характеристик. Особое внимание должны
привлекать те индивидуальные особенности, которые наиболее тесно связаны с
механизмами регуляции состояний и, в частности, влияют на развитие неоптимальных
состояний и стресса.
Сильное отрицательное влияние на обучение оказывают высокая
индивидуальная тревожность, повышенная реактивность симпатической системы.
Учащиеся с этими свойствами чаще других испытывают стресс и связанное с ним
нарушение когнитивной деятельности. Особенно очевидным это становится во время
экзаменов. В целом ситуация экзамена для большинства студентов является
стрессогенной, однако многие из них способны преодолевать этот стресс.
Исследование психофизиологических характеристик, которые могли бы
предсказывать развитие экзаменационного стресса, было проведено Дж. Спинксом (J. Spinks) на студентах психологического
факультета в Гонконге. Исследователи установили, что у лиц с более высокой
симпатической активацией, которую измеряли по скорости привыкания
электродермальной реакции к повторяющемуся звуковому тону, а также по уровню
потоотделения на ладони, в период экзаменационной сессии отмечалось подавление
функций иммунной системы.
У студентов, у которых привыкание электродермальной реакции на
звук, тестируемое во время летних каникул, происходило медленно (это указывало
на сильную активацию симпатической системы), в течение учебного года и во время
экзаменационной сессии в слюне снижалось содержание иммуноглобулина, что говорило
о наличии стресса. Была получена значимая положительная корреляция между
скоростью угасания электродермальной реакции и содержанием иммуноглобулина. Это
позволило по психофизиологической реакции — скорости привыкания реакции в виде
изменения кожной проводимости — предсказывать появление экзаменационного
стресса у студента. Индекс потоотделения менее коррелировал с состоянием
иммунной системы. Однако при разделении группы, состоящей из 250 студентов, на
две (по медиане) между ними были выявлены значительные различия. У лиц с
высокими значениями потоотделения (индекс симпатической активности) содержание
иммуноглобулина было снижено. Связь между вегетативной (симпатической) и
иммунной системами, которая осуществляется, по-видимому, через эндокринную
систему, объясняет, почему стресс и неоптимальные функциональные состояния
приводят к нарушениям иммунной системы и как след-В общем виде идея
двухконтурного управления процессом обучения представлена на рис. 64. Видны
два контура с обратными связями: слева — блок стимуляции, справа — сложная
живая система, обрабатывающая информацию. Верхняя часть рисунка дает
представление об управлении обучением по результатам приобретенных знаний.
Существуют различные способы оценки знаний. Однако в последние годы в
психофизике наметилось новое перспективное направление, которое дает
возможность на основе построения семантических пространств получать информацию
о структуре знаний ученика по конкретному материалу, а не только оценку по
уровню знаний (Терехина А.Ю., 1986). В нижней части 
рисунка показан контур управления по психофизиологическим параметрам
состояния.
Накопление физиологических данных в компьютере позволяет
осуществлять продолжительный контроль за эффективностью обучения. ФС сильно
зависит от подкрепления. В процессе обучения субъект сталкивается со сложными
задачами. Если в условиях выработки тонкой дифференцировки сигналов субъект
получает только негативное подкрепление, могут возникнуть невротические
реакции. Результатом этого может быть появление негативного отношения к
специфическому предмету или обучению как таковому.
Если требования, предъявляемые процессом обучения, оказываются
выше информационной способности учащегося, то в силу реципрокных отношений
между ориентировочным и оборонительным рефлексами происходит подавление
ориентировочного рефлекса за счет замещения его оборонительным. Пассивная
форма оборонительного рефлекса характеризуется депрессией или потерей
интереса. Активная форма оборонительного рефлекса выража-
ется в различных формах агрессивного поведения. В случае появления
оборонительных реакций вся ситуация обучения может стать сигналом для развития
негативных эмоций. Подавление негативного отношения к процессу обучения может
быть достигнуто лишь путем стимуляции творческой активности, которая связана с
позитивным эмоциональным тоном. Обучение, которое строится на
исследовательской деятельности, является наиболее эффективным обучением.
Оптимальный уровень активации мозга сопровождается вызовом
фазических ориентировочных реакций, отражающихся в ЭЭГ-ре-акциях активации,
расширении сосудов лба, падении сопротивления кожи и снижении частоты
сердечных сокращений. Учебный материал, вызывающий ориентировочный рефлекс,
хорошо запоминается. Привыкание ориентировочного рефлекса в результате
повторного предъявления материала может быть устранено через произвольный
ориентировочный рефлекс, закладывающий основу для творческой инициативы.
Могут быть дифференцированы два типа активации: продуктивная
активация, базирующаяся на ориентировочном рефлексе, и непродуктивная
активация, связанная с тревожностью и агрессией как формами
оборонительного рефлекса. Роль компьютерной полиграфии в обучении состоит в
том, что она позволяет выявлять неоптимальные функциональные состояния:
непродуктивную, высокую активацию, связанную со стрессом, тревожностью, агрессией,
или сниженный уровень активности за счет отсутствия мотивации или отказа от
работы из-за утомления. Манипуляция ФС учащегося позволяет вести обучение в
коридоре продуктивной активации, связанной с ориентировочным рефлексом.
Контроль за активацией ориентировочного типа может быть построен
на оценке частоты сердечных сокращений (ЧСС). Ориентировочный рефлекс
выражается снижением ЧСС, которому соответствуют ЭЭГ-активация и высвобождение ацетилхолина
в коре, что позволяет использовать фазические реакции снижения ЧСС в качестве
индекса ацетилхолиновой модуляции кортикальных нейронов, необходимой для
процессов обработки информации. Ацетилхолиновая сенситизация кортикальных
нейронов облегчает процессы внимания и научения. Базальная холинергическая
система переднего мозга, регулирующая высвобождение АХ в коре для поддержания
бодрствования и избирательной активации к значимым стимулам, рассматривается
как необходимый компонент произвольного ориентировочного рефлекса и
устойчивого внимания. Косвенным индексом возрастания холинергической активации
коры является коактивация сосудистой и дыхательной модуляций сердечного ритма,
которая положительно коррелирует со снижением ЧАС и реакциями внимания.
40. Социальная психофизиология.
Социальная психофизиология изучает поведение как результат
взаимодействия биологических и социальных факторов. Поведение
человека формируется в течение жизни на основе обучения и приобретения определенных
навыков под влиянием социальной среды, культуры и традиций общества, к которому
принадлежит субъект. Вместе с тем оно имеет и свои биологические корни,
психофизиологические механизмы, которые являются промежуточной переменной
между воздействием социума и поведением человека.
Биологические факторы, влияющие на социальное поведение человека,
выявляются уже при изучении зоосоциального поведения животных. Существует
группа зоосоциальных рефлексов, которые могут быть реализованы только через
взаимодействие с другими особями своего вида. Они составляют основу для
полового, родительского и территориального поведения. В значительной мере они
определяют и формирование групповой иерархии, где каждая особь выступает в роли
брачного партнера, родителя или детеныша, лидера или подчиненного, хозяина
территории или пришельца (Симонов П.В., 1987).
Одной из важных биологических потребностей, определяющих
зоосоциальные отношения животных, является потребность в привязанности, в
социальном контакте, в общении. Наиболее фундаментальная социальная связь —
это отношения матери и ребенка.
Они складываются по-разному в зависимости от того, как удовлетворяется
потребность в привязанности.
Характер контактов, общение животных в раннем детстве существенно
влияют на их социализацию в зрелом возрасте. Первые опыты с сепарацией
детенышей обезьян от матери и других особей, которая производилась сразу же
после рождения, были выполнены X. Харлоу
(Н. Harlow). Поведение изолированных детенышей
указывает на стремление животного компенсировать отсутствие общения с матерью
через реакции самоконтакта. Животное часто принимает позу, обхватывая лапами
свое туловище и голову.
Ранняя сепарация животных проявляется в нарушениях игрового,
полового и родительского поведения у взрослых животных. Такие животные плохо
идут на контакт с другими особями. У щенков отрицательное влияние сепарации,
совпадающей с их критическим периодом раннего возраста, впоследствии
проявляется в усилении отрицательных эмоций. Будучи уже взрослыми, они часто
реагируют на новизну (на незнакомые предметы) состоянием дистресса или страхом.
У животных, подвергшихся сепарации, биохимические сдвиги в мозге сходны с теми,
которые наблюдаются у человека при депрессии. Критический период первичной
социализации у собак соответствует возрасту 3—10 нед. Наиболее оптимальный
период для формирования у них навыков общения — 6—7-я неделя.
При воздействии отрицательных эмоциональных факторов (голод,
страх, боль) усиливается потребность в социальном контакте. На этом фоне ускоряются
процесс социализации и формирование привязанности. Существование
самостоятельной потребности в привязанности подтверждается также тем, что ее
удовлетворение может служить положительным подкреплением (наградой). У собак
можно выработать сложные формы поведения на социальном подкреплении (ласке).
У ребенка потребность в контакте со взрослым прежде всего
проявляется в его повышенном интересе к лицу человека. Лицо — наиболее
привлекательный стимул для новорожденного, которое он предпочитает рассматривать,
игнорируя другие объекты.
Формы проявления привязанности с возрастом меняются. Трехнедельный
младенец уже обнаруживает взгляд смотрящего на него человека. Первая
двигательная реакция, которая формируется у ребенка по направлению к матери, —
это фиксация глазами ее лица. В возрасте 4 мес. ребенок выделяет лицо своей
матери среди других и не отводит от него взгляда, пока она находится в поле его
фения. Когда мать удаляется, он начинает плакать, тем самымпоказывая, что
желает ее возвращения. В 6-месячном возрасте ребенок не только плачет, но и
пытается следовать за ней. Возвращение матери обычно вызывает бурную радость.
Ребенок улыбается, протягивает руки, издает радостные звуки. К 9-месячному
возрасту данная реакция становится устойчивой. Кроме того, если ребенок
пугается или огорчается, он стремится прижаться к матери. Начиная с конца 1 -го
года жизни ребенок уже может предвосхищать намерение матери уйти и реагирует
на это как на ее реальный уход.
Для формирования привязанности ребенка к матери важна не только
зрительная стимуляция, включающая лицо, но и кожные, оральные контакты. Особую
роль играют улыбка, разговор с ребенком, его поглаживание. X. Харлоу на макаках убедительно показал, что стремление
прикоснуться, ощутить телесную связь выражает насущную биологическую
потребность детеныша.
Привязанность, которая формируется между ребенком и матерью,
существенно влияет на его будущее эмоциональное и социальное развитие и
определяет его коммуникативные способности. Дети-первенцы, которые, как правило,
окружены большей заботой и вниманием родителей, имеют более сильную
потребность принадлежать какой-либо социальной группе, чем их младшие братья и
сестры.
Социальная потребность человека принадлежать к определенной
социальной группе и занимать в ней определенное место, пользоваться
вниманием окружающих имеет глубокие биологические корни в виде потребности
привязанности и боязни одиночества, которые испытывают животные.
Право принадлежать к какой-либо группе и получать от нее поддержку
налагает на особь обязанность соблюдать правила и нормы поведения,
установленные в данной группе. Последние диктуются иерархической организацией,
присущей каждой группе, в которой тон задают лидеры, доминантные особи.
Наличие иерархии стабилизирует и организует групповое поведение. Порядок в
стаде обезьян поддерживают доминирующие особи. Всякое нарушение норм поведения,
характерных для данного вида, вызывает реакцию агрессии со стороны других
членов сообщества и прежде всего особей высокого ранга. Обычно нарушитель изгоняется
из группы. У обезьян гамадрил наблюдали случаи, когда доминирующий самец
убивал собственного наследника, если тот чрезвычайно отвлекал внимание и
дестабилизировал группу.
Врожденные механизмы иерархического поведения сложны и остаются
малоизученными. Высокий ранг особи в группе обеспечивает преимущество в
получении пищи, мест для отдыха и в репро-
дуктивном поведении. Исследования показывают, что доминирование
особи зависит как от природных факторов, так и от социальных, средовых
влияний. Позицию доминирующей особи укрепляют не только ее личные притязания и
способности, но и проявление признаков подчинения субдоминантных членов
группы.
Как осуществляется взаимодействие социальных и природных факторов,
в совокупности определяющих высокий ранг особи, ее доминирующее положение? В
свете этой проблемы представляет интерес исследование, выполненное группой
ученых из Стэнфор-дского университета. Они исследовали социальную регуляцию
агрессивного поведения и репродуктивной функции у африканской рыбы (Haplochromis burtoni). У этого вида рыб функция воспроизводства монополизирована
небольшой частью самцов. Одновременно они отличаются агрессивностью, так как
защищают территорию выведения потомства от посягательств других рыб. Эти
территориальные самцы (Т-самцы) составляют около 10% всего количества самцов.
Остальные самцы (не-Т-самцы) находятся в подчиненном положении: не принимают
участия в размножении и в драках по защите территории. Кроме того, доминирующие
самцы (Т-самцы) отличаются яркой окраской, имеют высокий уровень циркуляции
андрогена (мужского полового гормона) и большого размера семенники.
На замороженных срезах мозга рыб (Т-самцов и не-Т-самцов) у
нейронов преоптической области вентрального гипоталамуса исследователи
определяли содержание гормона, выполняющего функцию релизинг-фактора для
гонадотропина — гормона, выделяемого гипофизом. Гонадотропин регулирует
половую функцию, определяет ее интенсивность. Они установили, что у Т-самцов в
гипоталамусе содержится большее количество фактора, высвобождающего
гонадотропин, а нейроны, в которых он содержится, имеют значительно большие
размеры, чем аналогичные нейроны у не-Т-самцов. Исследователи предположили, что
Т-самцы подавляют половую функцию не-Т-самцов, и это сопровождается изменениями
на нейронном уровне в гипоталамусе. Другими словами, социальные влияния на
половую функцию самцов данного вида рыб опосредованы нейронами преоптической
области вентрального гипоталамуса. Чтобы проверить эту гипотезу, они
исследовали случаи, когда не-Т-самцы превращались в Т-самцов. А это случалось
всякий раз, когда какая-либо территориальная зона для размножения оказывалась
вакантной и ее быстро осваивал не-Т-самец. Оказалось, что такая особь быстро
меняла свое поведение: оно станови-иось агрессивным. Рыба меняла свою окраску
на более яркую, при ном у нее происходили мощные гормональные изменения,
кото-рые усиливали ее репродуктивные функции. Параллельно с изменением
поведения у нового Т-самца увеличивался размер нейронов, содержащих
высвобождающий фактор для гонадотропина, так же как и его количество в этих
нейронах. Увеличивался и размер семенников. С переходом по каким-либо причинам
Т-самца в группу не-Т-самцов гормональные и поведенческие изменения шли в обратном
направлении.
Авторы полагают, что преоптическая область вентрального гипоталамуса
является непосредственной мишенью для социальных стимулов, которые могут
регулировать половую активность и функцию защиты территории, формируя либо
доминирующую, либо подчиненную особь. Подобные механизмы регуляции репродуктивной
функции, по-видимому, существуют и у других видов позвоночных, так как
ключевые нейроэндокринные компоненты оси гипоталамус — гипофиз — половые органы
принадлежат сильно консервативной системе, сохраняющейся у всех позвоночных. У
нейронов преоптической области гипоталамуса обнаруживается высокая
пластичность, хорошая способность к обучению. Возможно, они представляют то
звено, через которое реализуется взаимодействие социальных сигналов с
нейроэндокринным механизмом, обеспечивающим доминантное положение особи в
группе. Можно предположить, что некоторые биологические механизмы, обладающие
повышенной чувствительностью к воздействию социальных факторов, могут усиливать
половое и агрессивное поведение, делать их доминирующим видом поведенческой
активности, что приводит к существенным изменениям в зоосоциальных отношениях.
Одно из направлений социальной психофизиологии— изучение
связи индивидуальных различий и социальных процессов. Известно, что в силу
определенных индивидуальных различий одни субъекты проявляют ярко выраженную
склонность к доминированию, другие предпочитают позицию подчинения. Эти
качества легко могут быть выявлены с помощью психологических тестов. Однако
измерение индивидуальных различий по психофизиологическим параметрам
предоставляет значительно большие возможности для изучения природных,
биологических детерминант индивидуальных особенностей социального поведения.
Сравнительное изучение ЭЭГ на близнецах, сибсах и родственниках
показало сильную зависимость ее рисунка от генетического фактора. Установлена
высокая степень сходства ЭЭГ, особенно у монозиготных близнецов. Сходство ЭЭГ у
близнецов сохраняется до старости. Паттерны ЭЭГ у взрослых идентичны даже у
близнецовых пар с довольно разным эмоциональным прошлым, например у
близнецов, один из которых страдал тяжелым неврозом.
Близнецовые и семейные исследования показывают, что на-* следуются
не только ЭЭГ с хорошо выраженным альфа-ритмом, но и другие типы ЭЭГ
бодрствования, например низковольтная Э'-)Г с плохо выраженной или
отсутствующей альфа-активностью в затылочной области, а также ЭЭГ, где
альфа-волны замещены высокочастотной бета-активностью (16—19 в 1 с), которая,
подобно альфа-ритму, в ответ на стимул реагирует депрессией.
Таким образом, тип ЭЭГ, характерный для конкретного человека и
дающий представление о его индивидуальном уровне актива-пии, имеет сильную
генетическую детерминацию. Паттерн ЭЭГ отражает не только общий уровень
активности мозга, но и связанное с мим течение информационных процессов,
поэтому различные варианты ЭЭГ сцеплены с определенными типами
интеллектуальной к-ятельности, а также с различными личностными особенностями.
Результаты многочисленных исследований подтверждают это.
Так, лица с хорошо выраженным и регулярным альфа-ритмом часто проявляют
себя активными, стабильными и надежными нодьми. Для них характерны высокая
спонтанная активность и упорство, точность в работе, особенно в условиях
стресса, хорошая кратковременная память. Однако переработка информации у них
протекает не очень быстро. Другой паттерн ЭЭГ в виде низкоамплитудной
активности, наблюдаемой в широком диапазоне ча-i ют, сочетается со склонностью к экстраверсии и ориентированностью
на людей. Лица, обладающие данным типом ЭЭГ, отличаются хорошей
пространственной ориентацией. Для людей, у которых обнаруживают тип ЭЭГ с
диффузно распространенными бета-вол-мами, характерны низкие показатели тестов,
оценивающих концентрацию внимания и аккуратность. Они делают много ошибок м|)и
низкой скорости работы. Их устойчивость к стрессу мала.
В работах Н.Е. Свидерской показано, что индивиды, отличающиеся
друг от друга по психологическим признакам, имеют и раз-шчпую локализацию в
коре фокусов максимальной синхронизации потенциалов. Было выделено три типа их
расположения. Из О обследованных здоровых испытуемых 30% составили группу с
Фокусом максимальной синхронизации в передних отделах мозга, >0"г) с
фокусом в центральной области и 20 % лиц имели макси-мльиую синхронизацию в
теменно-затылочной области. Обследо-п.шие этих испытуемых по тесту Кеттела
показало, что индивиды • фокусом максимальной синхронизации в передних областях
характеризуются большой доминантностью, независимостью, само-перетюстью,
критичностью. Лица с фокусом максимальной син-ронпости потенциалов в задних
корковых областях, наоборот, имеют по этим же психологическим показателям
низкие баллы.Субъекты существенно различаются по чувствительности механизмов
кодирования и декодирования невербальных эмоциональных сигналов. Это
проявляется в их способности к эмпатии — постижению эмоционального
состояния, переживаний другого человека, сопереживанию ему.
Аналогичное явление обнаружено и у животных. Сигналы эмоционального
состояния другой особи того же вида — вокализация, специфические запахи,
особенности поведения партнера — обычно сильно влияют на поведение других
животных. Крысы и обезьяны уменьшают число нажатий на рычаг для получения пищи,
если эти реакции сопровождаются болевыми раздражениями другой особи того же
вида. Вместе с тем у животных существуют большие индивидуальные различия в
способности реагировать на сигналы оборонительного возбуждения, исходящие от
другой особи. Это явление П.В. Симонов назвал эмоциональным резонансом, в
котором проявляется способность животного строить и изменять свое поведение,
учитывая эмоциональные сигналы партнера.
В опытах П.В. Симонова (1987) из 247 белых крыс-самцов 77 (31%)
сравнительно быстро вырабатывали условную реакцию — избегать действий, которые
ведут к наказанию партнера током. 111 крыс (45%) вырабатывали эту же реакцию
только после того, как их несколько раз использовали в качестве жертвы. Но у
24% крыс даже после подобных испытаний не удалось выработать рефлекс избегания
криков боли партнера. Они не обращали внимания на страдания «жертвы». Таким
образом, на одного альтруиста приходится два эгоиста. Справедливость
этого соотношения была доказана для крыс, кошек, собак, обезьян и человека.
Хорошо известны опыты социального психолога С. Милгрэма (S. Milgram). Он сообщал своим испытуемым, что
хочет исследовать влияние наказания на заучивание слогов, при этом испытуемый,
выполняя роль «учителя», должен следовать правилу — наказывать «ученика» за
каждую новую ошибку все более сурово. Затем он помещал «учителя» перед рядом
кнопок, нажатие на которые вызывало удары тока напряжением от 15 до 450 В.
«Ученик» находился в соседнем помещении, окруженный проводами. Реально ток не
подавался, а «ученик», им был актер, демонстрировал свое переживание боли
криками и просьбами прекратить наказание. Результаты оказались следующими: 2/3
«учителей» — миролюбивые люди, отцы семейств — посылали «ученику», несмотря на
его стоны и мольбу, удары тока до 300 В только потому, что экспериментатор,
сидевший рядом с «учителем», настаивал продолжать «обучение», не обещая при
этом ни награды, ни наказания. И только 1/3 «учителей» отказалась продолжать
опыт.
Утех животных, у которых при виде страданий партнера появляется
эффект эмоционального резонанса, развивается отрицательное эмоциональное
состояние с признаками симпатической активации. Это заставляет его что-то
предпринять, чтобы устранить возникшую у него отрицательную эмоцию. Ради этого
животное вырабатывает сложный инструментальный навык, который и избавляет
жертву от ударов тока. Избавление от отрицательной эмоции служит
положительным подкреплением, которое и обеспечивает обучение.
Наиболее чувствительны к крикам жертвы доминирующие особи. 75%
крыс-доминантов вырабатывают условные рефлексы избавления. С жизненным опытом
чувствительность к страданиям других возрастает. Меньше всего реагируют на
страдание жертвы крысы самого низкого ранга. Животные, наиболее чувствительные
к страданиям других, одновременно обладают большей стрессоустойчи-востью.
Таким образом, эволюция поддерживает альтруистов.
Не менее важной проблемой социальной психофизиологии является
изучение влияния социальных факторов на здоровье. Социальный стресс — одна
из распространенных причин ухудшения состояния здоровья. В то же время
ограничение социальных контактов также может быть источником серьезных
заболеваний, в частности сердечно-сосудистых. Взаимоотношение
сердечно-сосудистых заболеваний и социальной среды наиболее изучено при
негативном влиянии социальных факторов. Вместе с тем следует ожидать, что существуют
и какие-то социальные факторы, способные предохранять от заболеваний и
сохранять здоровье.
Это предположение подтверждается работами, в которых показано,
что экспериментатор во время интервью с ипытуемым может снижать уровень его
симпатической активности, усиленной в результате развития лабораторного
стресса. По данным одних авторов, присутствие партнера на опыте может снижать
симпатическую активность (по электродермальному показателю и ЧСС), вызванную
физическим стрессом (болевой стимуляцией); по данным других, присутствие
посторонних лиц ухудшает состояние человека. Изучение влияния социального
присутствия на функциональное состояние человека показало, что результат
существенно зависит от характеристик наблюдателя. Снижение уровня
симпатической активности в присутствии наблюдателя возникает лишь в том случае,
если он хорошо знаком с испытуемым, имеет ранг того же уровня и при этом
ситуация не требует оценки деятельности испытуемого.
В некоторых работах исследовалось влияние касания на сердечнососудистую
активность. Установлено, что касание спины человека, находящегося в
спокойном состоянии, вызывает у него снижение ЧСС. Во время процедуры
обследования пациента отмеча-лись случаи, когда пальпация устраняла даже
желудочковую аритмию сердца. У женщин, которые до операции имели более частые
контакты с другими пациентами в виде касания плеч и спины, в послеоперационный
период обнаруживались лучшие показатели артериального давления.
В опытах Т. Камарка (Т. Kamarck) было
исследовано влияние касания на вегетативные реакции — систолическое и
диастолическое давление и ЧСС, когда испытуемый выполнял сложное арифметическое
задание, которое обычно вызывает увеличение кровяного давления и ЧСС. В одной
серии испытуемый выполнял задание в отсутствие наблюдателя, в другой — в
присутствии друга, которого просили время от времени касаться спины
испытуемого, демонстрируя ему тем самым свою поддержку. При этом друг не мог
подсказывать решение задачи, так как испытуемый был в наушниках.
Главный результат: присутствие друга снижало симпатические
реакции, вызываемые арифметическим тестом. При повторении этих опытов через 4
нед. были получены сходные результаты. Автор обращает внимание на то, что
эффект снижения симпатической активности не сопровождался какими-либо
изменениями в самочувствии субъекта. Тест Спильбергера, с помощью которого
измеряют состояние тревожности, гнева и любознательности, не фиксирует
каких-либо изменений. Отсутствие связи между состоянием вегетативной нервной
системы и самооценкой эмоционального состояния, которое получено в этих опытах,
может означать лишь то, что во время установления социальных контактов
социальные стимулы могут воздействовать на нервную систему на неосознаваемом
уровне и в зависимости от их особенностей вызывать тот или другой тип эффекта.
Положительное или отрицательное влияние социальных отношений на физиологические
процессы зависит от характера этих отношений. Дружеская поддержка снимает напряжение.
Социальное присутствие, предполагающее оценку действий человека, эквивалентно
стрессовому фактору.
Существуют различия в индивидуальной чувствительности к социальному
присутствию. Боязнь новизны в социальном окружении генетически
детерминирована. Она выявлена у некоторых новорожденных и проявляет себя как
устойчивая индивидуальная характеристика в течение жизни. Ее физиологической основой
является высокое содержание кортизола — гормона стресса. У младенцев с социальным
страхом высокий уровень кортизола обнаружен даже во сне. Негативное влияние
социального присутствия (аудитории, руководителя, партнера) на эмоции и
функциональное состояние субъекта может быть причиной затруднений, возникающих
при деловом общении.
Перспективным направлением социальной психофизиологии является
изучение психофизиологических коррелятов группового поведения. К. Хагфорс (К. Hagfors) из университета Юваскула (Финляндия)
осуществил групповую регистрацию кожно-гальваничес-кой реакции (КГР) во время
просмотра фильмов в кинозале. КГР одновременно регистрировали у 80 субъектов
при просмотре 8 коммерческих фильмов. Испытуемые имели задание — нажимать на
кнопку всякий раз при появлении эмоционального переживания любого типа и
держать ее нажатой до тех пор, пока эмоция не исчезнет. Индивидуальные записи
КГР преобразовывались в суммарную кривую, так же как и субъективные реакции,
фиксирующие моменты появления и длительность эмоционального переживания. Была
установлена высокая положительная корреляция между групповой записью КГР и
кривой субъективных оценок и выявлен общий закон изменения КГР как группового
показателя во время просмотра пятиминутных фильмов. Суммарная кривая КГР группы
испытуемых содержала два периода возрастания активности: в начале просмотра и
в конце. Эта динамика отсутствовала в индивидуальных кривых записи КГР во
время просмотра фильма, что дает основание рассматривать динамику суммарной КГР
как выражение группового эффекта. Периоды роста активности в суммарной КГР
рассматривались как отражение максимального вовлечения аудитории в общую
поведенческую активность, включающую смех, переживание успеха, ликование.
По мнению автора работы, регистрация КГР и получение кривой
групповой электродермальной активности могут быть использованы для оценки
привлекательности телевизионных и других фильмов, а также рекламы.