Открытие
сульфаниламидных препаратов и применение их в медицинской практике составили
известную эпоху в химиотерапии многих инфекционных заболеваний, в том числе
сепсиса, менингита, пневмонии, рожистого воспаления, гонореи и некоторых
других.
Однако
наибольший интерес для медицины представили различные биологически активные
вещества, полученные биосинтетическим путем, то есть соединения, образующиеся в
процессе жизнедеятельности разнообразных организмов.
В
конце XIX века русские ученые В. А. Манассеин и А. Г.
Полотебнов показали, что грибы из рода Penicillium
способны задерживать в условиях in vivo развитие возбудителей ряда кожных заболеваний
человека.
И.
И. Мечников в 1894 году обратил внимание на возможность использования некоторых
сапрофитных бактерий в борьбе с патогенными микроорганизмами.
В
1896 году Гозио из культуральной
жидкости Penicillium выделил
кристаллическое соединение - микофеноловую кислоту,
подавляющее рост бактерий сибирской язвы.
Эммерих и Лоу в
1899 году сообщили об антибиотическом веществе, образуемом Pseudomonas pyocyanea, они
назвали его пиоцианазой; препарат использовался в
качестве лечебного фактора как местный антисептик.
В
1910-1913 годах Black O. и Alsberg U. выделили из гриба рода Penicillium пенициловую
кислоту, обладавшую антимикробными свойствами.
К
сожалению, эти и некоторые другие наблюдения и открытия не получили в то время
дальнейшего развития, но они оказали огромное положительное влияние на более
поздние исследования в области изучения биологически активных продуктов
жизнедеятельности организмов.
В
1929 году Александром Флемингом был открыт новый препарат пенициллин, который
только лишь в 1940 году удалось выделить в кристаллическом виде. Это новое и
весьма эффективное химиотерапевтическое вещество было получено в результате
жизнедеятельности микроорганизма, то есть биосинтетическим путем.
Применение
пенициллина в борьбе с различными инфекционными заболеваниями и воспалительными
процессами явилось мощным стимулом для поиска новых, еще более эффективных
антибиотических веществ, образуемых различными группами микроорганизмов
(бактериями, актиномицетами), низшими растениями
(дрожжами, водорослями, плесневыми грибами, высшими грибами), высшими
растениями и животными организмами.
Настойчивые
поиски продуцентов новых антибиотиков увенчались блестящими успехами.
Так,
если проследить за динамикой роста числа описываемых антибиотиков, то можно
заметить следующую закономерность.
В
1896 году Гозио выделил микофеноловую
кислоту, в 1899 году Эммерих и Лоу
описали пиоцианазу. В 1937 году Вельш
описал первый антибиотик актиномицетного
происхождения актиномицин, в 1939 году Красильниковым и Кореняко был получен линцетин
и Дюбо-тиротрицин. Таким образом, к моменту получения
пенициллина в очищенном виде (1940) уже было известно 5 антибиотических
веществ. После этого число антибиотиков росло очень быстрыми темпами:
|
Год
|
Число антибиотиков
|
Год
|
Число антибиотиков
|
|
1856
1899
1937
1939
1940
1945
|
1
2
3
5
6
32
|
1949
1953
1964
1968
настоящее время
|
150
450
>
1600
>
2000
около 3000
|
Из
около 3 000 антибиотиков, известных к настоящему времени, лишь примерно сто
находят применение в медицинской практике: при лечении воспалительных процессов
(пневмония, перитонит, фурункулез), различных форм туберкулеза, при борьбе со
многими инфекционными заболеваниями, считавшимися ранее неизлечимыми или трудно
излечимыми и т.д. Применение этих соединений привело к резкому снижению
смертности при таких заболеваниях, как крупозное воспаление легких, сепсис,
различные формы менингита и др.
Большинство
описанных антибиотиков не находит применения в медицинской практике из-за их
токсичности, инактивации в организме больного или
других причин.
Работы
по изысканию антибиотических веществ нового поколения, эффективных при лечении
вирусных и раковых заболеваний, борьба с которыми является одной из важнейших
проблем современности, продолжаются с неослабленной энергией.
Последние
15-20 лет ежегодно различным вопросам, связанным с изучением антибиотиков,
посвящается более пяти тысяч работ.
Открытие
и изучение свойств нового антибиотика, применяемого в медицинской практике, -
это огромный труд ученых различных направлений (микробиологов, биохимиков,
генетиков, химиков, технологов, фармакологов, врачей и др.).
По
подсчетам некоторых американских ученых над открытием лишь одного такого
антибиотика широкого спектра 55 ученых непрерывно работали 2,5года. Они
обследовали более 100 тысяч образцов почвы, израсходовали на это более 4 млн долларов.
Итальянской
фармацевтической компании "Лепетит" для
производства нового противотуберкулезного антибиотика потребовалось одиннадцать
лет научно-исследовательских работ, которые обошлись в несколько миллионов
долларов.
Каковы
же основные причины столь быстрого роста числа антибиотиков, происходящего за
последние 20-25 лет, несмотря на огромные финансовые затраты и необходимость
привлечения большого числа исследователей?
Среди
них можно назвать следующие:
1.
Многие антибиотические вещества - незаменимые лечебные
препараты. Они широко применяются при лечении большого числа инфекционных
заболеваний, которые ранее, до открытия антибиотиков, считались неизлечимыми
или сопровождались высоким летальным исходом. К их числу следует отнести
некоторые формы туберкулеза, чуму, азиатскую холеру, брюшной тиф, бруцеллез,
пневмонию, различные септические процессы.
2.
Антибиотики - очень полезные вещества в сельском
хозяйстве, прежде всего как лечебные препараты в животноводстве, птицеводстве,
пчеловодстве и растениеводстве, а отдельные антибиотические вещества - и как
стимуляторы роста животных.
3.
При широком применении антибиотиков в качестве
лечебных препаратов происходит быстрое накопление резистентных к этим
соединениям форм микроорганизмов. Проблема резистентности
микроорганизмов ставит задачу замены одних антибиотиков другими, то есть поиска
все новых и новых антибиотических веществ.
4.
Некоторые из антибиотиков с успехом применяются в
пищевой и консервной промышленности в качестве консервантов
скоропортящихся продуктов (свежей рыбы, мяса, сыра, различных овощей).
5.
Антибиотические вещества - новые, ранее неизвестные по
химическому строению соединения - представляют огромный интерес для
специалистов в области химии природных соединений. Изучение структуры этих
веществ, а также синтез некоторых из них способствовали бурному развитию
указанного направления в химии, а следовательно, и самой науки об антибиотиках.
Достаточно указать, что к настоящему времени синтезированы такие антибиотики,
как пенициллины, хлорамфеникол, тетрациклины и др.
6.
Антибиотики нашли широкое применение в научных
исследованиях в качестве веществ, используемых при изучении отдельных сторон
метаболизма организмов, расшифровки тонких молекулярных механизмов биосинтеза
белка, механизма функционирования мембран и других биохимических превращений
как специфические ингибиторы определенных реакций. Например, одни антибиотики
специфически ингибируют отдельные этапы синтеза белка
на рибосомах (хлорамфеникол,
пуромицин, тетрациклин), другие - синтез на разных
уровнях нуклеиновых кислот (саркомицин подавляет
активность полимераз; актиномицин блеомицин,
рубомицин и другие нарушают функцию ДНК; и т.д.),
третьи - образование клеточных стенок
(пенициллины) и т.д.
7.
Изучение путей образования антибиотиков способствует
глубокому проникновению в механизмы синтетической деятельности продуцентов этих
биологически активных соединений, раскрытию основных этапов их метаболизма.
Таким
образом, все эти факторы способствовали и продолжают способствовать тому, что к
проблеме антибиотиков привлечено внимание огромных групп ученых различных
направлений: микробиологов, микологов, биохимиков, химиков, генетиков, цитологов, фармацевтов, врачей, технологов и т.д. изучение
антибиотиков - это типичный пример комплексного подхода к проблеме, что само по
себе способствовало прогрессу в исследовании этих биологически активных соединений.
5. Биологическая роль антибиотиков в природе.
Результаты,
получаемые в лабораторных условиях нельзя непосредственно переносить на
явления, имеющие место в естественных местах обитания организмов, как отмечал Новогрудский (1948), явления микробного антагонизма в почве
протекают своеобразно, иногда значительно отличаясь от антагонизма тех же
микробов на искусственных питательных средств. Это положение имеет особенно
важное значение при рассмотрении вопроса в биологической роли антибиотиков,
т.е. о той роли этих веществ, какую они играют в естественных местах нахождения
микроорганизмов, образующих их.
О
биологической роли антибиотиков не сущществует
единого менения. Это говорит о том. Что обсуждаемый
вопрос представляет собой не простое явление.
Рассмотрим
две противоположные точки зрения и биологической роли антибиотиков.
Первая
исходит из того, что образование антибиотиков следует рассматривать как
специфическую особенность обмена веществ организмов, возникшую и закрепленную у
них в процессе эволюционного развития. Образование и выделение антибиотиков в
окружающую среду при жизни организмов или после их отмирания - могущественный
фактор в борьбе за существования видов.
Такая
точка зрения о роли антибиотических веществ широко распространена среди ведущих
российских и зарубежных специалистов. Ее поддерживают Имженецкий,
Красильников, Гаузе, Гроссбард, Брайэн, Гаррет, Торнтон и другие ученые.
Биосинтез
антибиотиков - наследственная особенность организмов, проявляющаяся в том. Что
каждый вид (штамм) способен образовывать один или несколько вполне
определенных, строго специфичных для него антибиотических веществ.
Вместе
с тем известно, что одинаковые антибиотики могут образовываться несколькими
видами организмов. И это нисколько не противоречит мысли о наследственно
закрепленном свойстве микроорганизмов продуцировать определенные
антибиотические вещества.
Выявление
потенциальной возможности образовывать в процессе жизнедеятельности антибиотики
связано с условиями культивирования организмов. В одних условиях организм
образует антибиотик, в других условиях тот же организм при хорошем росте не
будет обладать способностью синтезировать антибиотическое вещество. Однако
такие явления наблюдаются в лабораторных условиях культивирования изучаемого
организма, в условиях ограниченного или слишком богатого выбора источников
питания.
Вторая
точка зрения состоит в том, что антибиотические вещества, образуемые
микроорганизмами, носят случайный характер, зависящий лишь от условий
культивирования. По мнению этих авторов (Ваксман и
другие), образование антибиотиков - это не закрепленное свойство организма,
проявляющееся только при развитии организма в специфической среде и при наличии
особых внешних условий. Поэтому антибиотики не имеют для продуцентов
приспособительного значения, их образование не связано с эволюцией
микроорганизмов. Эта точка зрения основывается на двух положениях:
1.
Не все микроорганизмы образуют антибиотические
вещества, что, однако, не мешает их широкому распространению в природе.
2.
Антибиотические вещества, даже самые устойчивые,
довольно быстро инактивируются в почве, в этом
естественном местообитании большинства
микроорганизмов. Только при максимальном насыщении почвы антибиотиками можно
получить соответствующий биологический эффект.
Как
пишут Ваксман и Лешевалье
(1962), антибиотики являются "лабораторными продуктами, образуемыми
растущими чистыми культурами микроорганизмов в условиях богатой питательными
веществами среды при хорошей аэрации, но они не обнаруживаются в почве".
Итак,
по мнению названных авторов, следует что антибиотики, образуемые
микроорганизмами, носят случайный характер, зависящий только от условий
культивирования. Если бы это действительно имело бы место, то мы были бы вправе
ожидать, что при изменении условий культивирования, например, для продуцента
стрептомицина, можно было бы получить образование, например, хлортетрациклина
или пенициллина. Но ведь этого никогда не бывает, как бы не менялись условия
культивирования, как бы серьезно не менялся состав среды для культивирования.
Экспериментатору
удается путем изменения условий культивирования получить больший или меньший
выход антибиотика, или создать условия, при которых антибиотик вообще не будет
образовываться. Можно также путем изменения условий культивирования продуцента
добиться преимущественного биосинтеза одного из антибиотиков, при условии
образования изучаемым организмом нескольких антибиотических веществ, или же
получить новые формы антибиотиков, но только в пределах тех соединений, которые
способны синтезироваться этим организмом. Но экспериментатору, по-видимому,
никогда не удается достичь того, чтобы продуцент стрептомицина Act. Streptomycini путем
изменения условий культивирования начал образовывать хлортетрациклин и
пенициллин. Наследственная особенность продуцента стрептомицина состоит в том,
что он может образовывать стрептомицин или гризеин,
или другие антибиотики, свойственные данному виду. И только их, но не
пенициллин, не антибиотики тетрациклиновой группы, не актиномицины
и не какие другие антибиотики.
Образование
антибиотиков нельзя считать случайным явлением в жизнедеятельности
микроорганизмов. Этот процесс обусловлен определенным характером обмена
веществ, возникшим и закрепленным в процессе эволюции организма. Однако нельзя
отрицать того факта, что в отдельных случаях проявление антагонизма у
микроорганизмов связано с образованием продуктов обмена, не являющихся
специфическими веществами их метаболизма. Такой характер имеет антагонизм у
уробактерий, обусловленный выделением аммиака при использовании мочевины, или
антагонизм некоторых молочнокислых бактерий, связанный с выделением ими
перекиси водорода и т.д. Но такие продукты жизнедеятельности микроорганизмов мы
не называем антибиотиками.
Ваксман и Лешевалье
(1962) считают, что антибиотики образуются только чистыми культурами. Однако
это положение далеко не обязательно. Многие виды микроорганизмов, в частности актиномицеты, способны образовывать антибиотические
вещества только в присутствии других организмов (Макаровская,
1956; Егоров и др., 1960; Красильников и Егорова,
1960).
Ваксман и его последователи основывают
свою концепцию на том, что не все микроорганизмы способны образовывать
антибиотики, что, однако, не мешает им быть широко распространенными в природе.
Образование
антибиотиков - лишь одна из форм антагонистических взаимоотношений,
существующих в мире микроорганизмов. В борьбе за распространение в природе
микроорганизмы "используют" не только фактор антибиотикообразования,
но и многие другие эволюционно закрепленные особенности, дающие им преимущество
в острой конкурентной борьбе с другими видами.
Образование
антибиотиков микроорганизмами при культивировании их в условиях лаборатории
действительно, по мнению многих исследователей, проявляется далеко не у всех
организмов. Так, в ряде работ отмечается, что всего лишь 40 - 70 процентов
штаммов актиномицетов обладают антибиотической
активностью, а остальные штаммы являются неактивными.
Однако
Егоров (1956) экспериментально показал, что при соответствующих условиях
культивирования все, так называемые, неактивные штаммы актиномицетов
способны в той или иной степени образовывать антибиотические вещества и в
лабораторных условиях.
Макаровская (1956) и Егоров с сотр. (1960) установили, что продукты жизнедеятельности
некоторых микроорганизмов способствуют проявлению антибиотических свойств у так
называемых неактивных штаммов актиномицетов; они
также вызывают усиление уже ранее определенных антагонистических свойств актиномицетов.
Гаузе (1968), Davies, Williams (1970) и другие авторы, применяя при поисках
продуцентов антибиотических веществ новые тест-организмы
(например, штаммы микробов с дефектом окисления, актиномицеты),
а также используя метод выращивания актиномицетов на
средах, содержащих некоторые антибиотики, получали ранее неизвестные
антибиотические вещества с ценными свойствами (например, со свойствами
антиметаболитов и др.).
Наконец,
сторонники второй концепции основываются на том, что антибиотические вещества в
почве быстро инактивируются, а поэтому не могут
играть какой-либо биологической роли.
Действительно,
многие антибиотики, искусственно внесенные в почву, довольно быстро в ней
исчезают. Известно также, что многие антибиотики при тех же условиях могут
сохраняться в почве довольно длительное время (до нескольких недель).
При
рассмотрении вопроса о биологической роли антибиотиков, образуемых
микроорганизмами в естественных местах обитания, в особенности в почве, следует
иметь ввиду, что в почве микроорганизмы расселяются не диффузно, а живут
отдельными колониями (Красильников, 1936, 1951; Новогрудский, 1936, 1949).
Существенное
значение приобретает факт адсорбции микроорганизмов и продуктов их
жизнедеятельности, в том числе и антибиотиков, на частицах почвы (Звягинцев, 1973). В естественных местах обитания (почва)
микроорганизмов происходит своеобразная иммобилизация клеток и образующихся
продуктов метаболизма (антибиотиков, ферментов, токсинов и др.), что играет
огромную роль в проявлении биологической активности у микроорганизмов.
Красильниковым было показано, что в тех местах,
где имеется больше органических остатков, обильнее развиваются микробы и
образуемые ими очаги имеют большие размеры. Обычно такое развитие
микроорганизмов наблюдается в порах между твердыми частицами почвы или на
частицах почвы.
При
развитии микробного очага, который может состоять из представителей одного или
нескольких (не антагонистических) видов, образуются продукты жизнедеятельности.
в том числе и антибиотические вещества, которые диффундируя в соседние поры,
могут играть там важную биологическую роль.
Итак,
антибиотики могут образовываться и образуются при развитии микроорганизмов в
естественных местах их обитания (почва) без внесения туда дополнительных
питательных веществ.
Образовавшиеся
в почве антибиотики в зависимости от их химического строения способны
сохраняться там определенное время и проявлять свое биологическое действие.
Положения,
из которых исходят противники активной биологической роли антибиотиков. имеют
серьезные возражения. Они не могут ни в какой степени поколебать единственно
правильный взгляд от активной биологической роли антибиотиков. развиваемый
нашими микробиологами и поддержанный рядом зарубежных исследователей,
рассматривающих образование антибиотических веществ как средство
приспособления, выработавшееся в процессе эволюционного развития организмов.
Разумеется,
биологическую роль подробно можно выяснить лишь при детальном изучении
отдельных веществ. установлено, например, что некоторые антибиотики оказываются
довольно вредными продуктами жизнедеятельности для собственных продуцентов.
Так, флавинин, образуемый грибом Aspergillus
flavipes, подавляет развитие собственного продуцента
в концентрации 1,25 мкг/мл, а антибиотик пиоцианин играет активную роль в
окислительно-восстановительном процессе бактерий, образующих это вещество.
Однако
любое детальное изучение биологической роли того или иного антибиотика может
подтвердить лишь вывод о том, что это вещество не может быть случайным
продуктом обмена, а есть результат метаболизма, появившегося в процессе
эволюции продуцента.
III. Общие сведения о
действии антибиотиков.
1. Классификация антибиотиков.
К
настоящему времени описано около 3000 антибиотических веществ. разобраться в
таком количестве антибиотиков возможно только при соответствующей
классификации, распределении их в определенном порядке.
Сложилось
несколько подходов к классификации антибиотиков, причем они определяются,
главным образом. Профессиональными интересами ученых. Так, для биологов,
изучающих организмы - продуценты антибиотических веществ, условия образования
этих соединений и другие, типичные для этой группы ученых, проблемы наиболее
приемлемой классификацией антибиотиков будет такая, в основу которой положен
принцип биологического происхождения антибиотиков. Для специалистов, изучающих
вопросы механизма физиологического действия антибиотиков, наиболее удобным
принципом классификации антибиотических веществ, естественно, будут признаки их
биологического действия. Для химиков, изучающих детальное строение молекул
антибиотиков и разрабатывающих пути их химического синтеза, приемлемой будет
классификация, основанная на химическом строении антибиотиков и т.д.
I.
Классификация антибиотиков по их биологическому
происхождению.
1.
Антибиотики, образуемые микроорганизмами, относящимися
к ряду Eubacteriales.
А.
Образуемые представителями рода Pseudomonas:
Пиоцианин - Ps. aeruginsa,
Вискозин
- Ps. viscosa
Б.
Образуемые представителями родов Micrococcus, Streptococcus, Diplocoooccus, Chromobacterium, Escherichia, Proteus:
Низин
- Str. lactis
Дипломицин - Diplococcus
X-5
Продигиозин - Chromobacterium
prodigiosum (serratia, marcescens)
Колиформин - E. coli
Протаптины - Pr. vulgaris.
В.
Образуемые бактериями рода Bacillus:
Грамицидины
- Bac. brevis
Субтилин - Bac. subtilis
Полимиксины - Bac. polymyxa
Колистатины - неиндентифицированная
споровая аэробная палочка.
2.
Антибиотики, образуемые микроорганизмами, принадлежащими к ряду Actinomycetales:
стрептомицин
- Act. streptomycini,
тетрациклины
- Act. aureofaciens, Act. rimosus,
новобиоцин - Act. spheroides,
актиномицины - Act. antibioticus и др.
3.
Антибиотики, образуемые несовершенными грибами:
пенициллин
- Penic. chrysogenum
гризеофульвин - Penic.
griseofulnum
трихоцетин - Tricholecium
roseum
4.
Антибиотики. Образуемые грибами, относящимися к классам бизидиомицетов
и аскомицетов:
термофиллин - базидомицет
Lenzites thermophila,
лензитин - Lenzites
sepiaria,
хетомин - Chaetoomium
cochloides (аскомицет).
5.
Антибиотики, образуемые лишайниками, водорослями и низшими растениями:
усниновая кислота (биан)-
лишайником,
хлореллин - Chlorella
vulgaris.
6.
Антибиотики, образуемые высшими растениями:
алмицин - Allium
sativum,
рафанин - Raphanus
sativum
фитоалексины: пизатин
в горохе (Pisum sativum), фазеолин в фасоли (Phaseolus vulgaris).
7.
Антибиотики животного происхождения:
лизоцим, экмолин,
круцин (Tripanosoma cruzi),
интерферон.
II.
Классификация антибиотиков по механизму из биологического действия.
1.
Антибиотики, ингибирующие
синтез клеточной стенки (пенициллины, тацитрацин, ванкомицин, цефалоспорин, Д-циклосерин).
2.
Антибиотики, нарушающие функции мембран (альтомиицин, аскозин,
грамицидины, кондицидины, нистатин,
трихомицин, эндомицин и
др.).
3.
Антибиотики. Избирательно подавляющие синтез (обмен)
нуклеиновых кислот:
а)
подавляющие синтез РНК (актиномицин, гризеофульвин, канамицин, неомицин, новобиоцин, оливомицин и др.).
б)
подавляющие синтез ДНК (актидион, брунеомицин,
митомицины, новобиоцин, саркомицин, эдеин и др.).
4.
Антибиотики - ингибиторы синтеза пуринов и пиримидинов
(азасерин, декоинин, саркомицин и др.).
5.
Антибиотики, подавляющие синтез белка (бацитрацин, виомицин, канамицин, неомицин, тетрациклины,
хлорамфеникол, эритромицин
и др.).
6.
Антибиотики, являющиеся ингибиторами дыхания (антимицины, олигомицины, патулин, пиацианин, усниновая кислота и др.).
7.
Антибиотики - ингибиторы окислительного фосфорилирования (валиномицин,
грамицидины, колицины, олигомицин, тироцидин и др.).
8.
Антибиотики, обладающими антиметаболитными
свойствами. Антибиотические вещества, образуемые некоторыми актиномицетами
и плесневыми грибами. Эти антибиотики выступают в качестве антиметаболитов
аминокислот, витаминов, нуклеиновых кислот.
К
числу антибиотиков-антиметаболитов относятся: фураномицин
- антиметаболит лейцина; антибиотик - антагонист метаболизма аргинина и орнитина, образуемый Act. griseovariabilis; антибиотик - антагонист метионина и тиамина, выделенный
из культуры Act. globisporus;
антибиотическое вещество - антиметаболит аргинина, лизина или гистидина, синтезируемое Act. macrosporus (термофилл).
2. Единицы биологической
активности.
Выражение
величин биологической активности антибиотиков обычно производят в условных
единицах, содержащихся в 1 мл раствора (ед/мл) или в
1 мг препарата (ед/мг). За единицу антибиотической
активности принимают минимальное количество антибиотика, способное подавить
развитие или задержать рост стандартного штамма тест-микроба в определенном
объеме питательной среды.
Единицей
антибиотической активности пенициллина считают минимальное количество
препарата, способное задерживать рост золотистого стафиллококка
штамм 209 в 50 мл питательного бульона.
Для
стрептомицина единица активности будет иной, а именно: минимальное количество
антибиотика, задерживающее рост E. сoli в одном
миллилитре питательного бульона.
После
того как многие антибиотики были получены в химическом чистом виде, появилась
возможность для ряда из них выразить условные единицы биологической активности
в единицах массы.
Установлено,
что 1 мг чистого основания стрептомицина эквивалентен 1000 единицам
биологической активности. Следовательно, одна единица активности стрептомицина
эквивалентна одному микрограмму (мкг)
чистого основания этого антибиотика. В связи с этим в настоящее время в
большинстве случаев количество стрептомицина выражают в мкг/мг
или в мкг/мл. Чем ближе число мкг/мг
в препаратах стрептомицина стоит к 1000, тем, следовательно, чище данный
препарат, тем меньше он содержит балластных веществ.
У
таких антибиотиков, как карбомицин, эритромицин, новобиоцин, нистатин, трихотецин и некоторых
других, одна единица активности эквивалентна или приблизительно эквивалентна 1 мкг вещества.
Однако
у ряда антибиотиков единица биологической активности значительно отличается от
1 мкг вещества. Например, 1 мг чистого основания неомицина содержит 300 ед. активности. Поэтому 1 единица
активности этого антибиотика эквивалентна 3,3 мкг.
Для бензилпенициллина 1 ед
активности эквивалентна примерно 0,6 мкг, так как 1 мкг антибиотика содержит 1667 ед. (оксфордских). Для фумагиллина за единицу фагоцидного
действия принято брать 0,1 мкг чистого вещества. 1
единица бацитрацина эквивалентна 20 мкг вещества.
Соотношение
единиц биологического действия (ед.) некоторых стандартных антибиотиков и
единиц их массы приведено в таблице 5.
Таблица 5.
Соотношение единиц действия некоторых антибиотиков и единиц
массы этих антибиотиков (по Герольд, 1966).
|
Антибиотик - стандарт
|
Ед/мг
|
Единица массы
|
|
Альбомицин (сульфат)
|
700000
|
Нет
|
|
Бацитрацин
|
52
|
Нет
|
|
Эритромицин (основание)
|
1000
|
1 мкг основания
|
|
Хлортетрациклин (хлоргидрат)
|
1000
|
1 мкг чистого хлоргидрата
|
|
Карбомицин (основание)
|
1000
|
1 мкг основания
|
|
Окситетрациклин (дигидрат)
|
925
|
1
мкг чистой безводной амфотерной
формы
|
|
Пенициллин (натриевая соль)
|
1667
|
0,587
мкг чистой кристаллической калиевой соли
|
|
Полимиксин В (сульфат)
|
7200
|
Нет
|
|
Саркомицин
|
12
|
Нет
|
|
Тетрациклин (тригидрат)
|
890
|
1 мкг чистой безводной амфотерной формы
|
|
Стрептомицин (сульфат)
|
800
|
1 мкг чистого основания
|
|
Биомицин (сульфат)
|
745
|
1 мкг чистого основания
|
а) Пенициллин - антибиотик,
образуемый филаментозным грибом.
Огромная
группа организмов, принадлежащих к грибам, образует большое число (около 400)
разнообразных антибиотических веществ, отдельные представители которых
завоевали всеобщее признание в качестве лечебных средств. Основная же часть
грибных антибиотиков не нашла еще практического применения главным образом в
силу своей высокой токсичности.
В
медицинской и сельскохозяйственной практиках имеют значение ограниченное число
антибиотиков, образуемых некоторыми видами грибов, а именно: пенициллин, фумагиллин и некоторые другие.
Пенициллин (Penicillin).
Известный
английский бактериолог Александр Флеминг опубликовал в 1929 г. сообщение о литическом действии зеленой плесени на стафиллококки.
Флеминг выделил гриб, который оказался Penicillium notatum, и установил, что культуральная
жидкость этой плесени способна оказывать антибактериальное действие по
отношению к патогенным коккам.
Культуральная жидкость гриба, содержащая
антибактериальное вещество, названо Флемингом пенициллином.
Попытки
Флеминга выделить активное начало, образуемое Penicillium,
не увенчалось успехом.
Несмотря
на это, Флеминг указал на перспективы практического применения обнаруженного им
фактора.
Спустя
примерно десять лет после сообщения Флеминга Е. Чейн
начал с конца 1938 г. изучать пенициллин. Он был убежден, что это вещество -
фермент. В 1940 г. Флори и Чейн
получили индивидуальное соединение пенициллина, который оказался не ферментом,
а низкомолекулярным веществом.
Об
антагонистических свойствах зеленой плесени (Penicillium)
было известно задолго до наблюдений Флеминга. Следует указать, что еще в
глубокой древности индейцы из племени майя использовали зеленую плесень,
выращенную на зернах кукурузы, для лечения ран. Философ, врач и
естествоиспытатель Абу-Али Ибн-Сина (Авиценна)
рекомендовал использовать плесень при гнойных заболеваниях.
Ибн-Сина
написал пятитомный "Канон врачебной науки", который был впервые
переведен на латинский язык и издан в Европе через 400 лет после его смерти - в
1437 г. На русском языке "Канон" издан лишь в 1960 г. Авиценна
утверждал, что заразные заболевания вызываются невидимыми для глаза живыми
возбудителями, которые могут передаваться от больного к здоровому через воздух
и воду. Заключение это сделано за 600 лет до изобретения микроскопа.
В
русской народной медицине с давних времен применялись для лечения ран присыпки,
состоящие из зеленой плесени.
В
работах русских ученых Манассеина и Полотебнова в 1871 - 1872 гг. указывалось на отношение Penicillium glancum к разным
бактериям. Полотебнов впервые в научно-клинической
обстановке изучил применение зеленой плесени, показав при этом практические
ценные результаты. Манассеин установил, что молодая
культура плесени подавляет рост некоторых бактерий. В 1877 г. русский врач
Лебединский доложил о подавлении плесенью бактерий желудочно-кишечного тракта.
Английский
физик Тиндаль описал в 1876 г. способность Penicillium подавлять бактерии, находящиеся в жидкости, но
объяснял это явление чисто физическими причинами.
Таким
образом, приведенные данные показывают, что человечество на разных уровнях
своего развития знало о целебных свойствах зеленой плесени. Однако, эти
сведения носили разрозненный характер и касались лишь воздействия самого гриба
на микроорганизмы. В то время не могло быть и речи о выделении и изучении
активного начала, образуемого плесенью.
И
лишь когда в 1940 г. Флори и Чейн
получили препараты (пенициллин) в очищенном виде, после этого появился широкий
научный интерес к этому антибиотическому веществу.
Изучение
пенициллина в Советском Союзе было начато З. В. Ермольевой.
В
1942 г. под руководством Ермольевой в лаборатории
биохимии микробов Всесоюзного института экспериментальной медицины в Москве был
получен первый отечественный пенициллин - крустозин,
сыгравший огромную роль в спасении жизней воинов Советской Армии, раненных на
полях сражений Великой Отечественной войны.
В
январе 1944 г. Москву посетила группа иностранных ученых, среди которых был
профессор Флори, привезший с собой английский штамм
продуцента пенициллина. Сравнение двух штаммов (советского и английского)
показало, что советский штамм образует 28 ед/мл, а
английский - 20 ед/мл (Ермольева,
1967).
После
того как было установлено, что пенициллин обладает мощными лечебными
свойствами, начались интенсивные поиски продуцентов этого антибиотика. В
результате большого числа работ удалось установить, что пенициллин могут
образовывать многие виды Penicillium (Penic. chrysogenum, Penic. bericompactum, Penic. nigricans, Penic. turbatum, Penic. steckii, Penic. corylophilum), а также
некоторые виды Aspergillus (Asp.
flavus, Asp. flavipes, Asp. janus, Asp. nidulans
и др.). есть указания, что пенициллин образуется также термофильным организмом Malbranchia pulchella (см. Беккре, 1963).
Первые
выделенные из естественных субстратов штаммы Penicillium
как наиболее активные продуценты пенициллина образовывали не более 20 единиц
(12 мкг) антибиотика на 1 мл культуральной
жидкости. Даже промышленное производство этого ценнейшего препарата было начато
при активности культуральной жидкости не выше 30 мкг/мл или 50 ед/мл. насколько
низка эта активность, можно судить по тому факту, что в настоящее время в
промышленных условиях получают культуральные жидкости
с содержанием пенициллина более 15000 ед/мл, а
отдельные штаммы способны синтезировать антибиотик в количестве до 25 тыс. ед/мл.
Получение
высоких выходов антибиотика достигнуто в результате изучения условий его
образования и селекции наиболее активных штаммов продуцента пенициллина.
б) Действие пенициллина на
бактерии.
Вопросу
рассмотрения антибиотической активности пенициллина в отношении ряда
микроорганизмов уделено достаточно много внимания. Установлено, что пенициллин
оказывает антимикробное действие в отношении некоторых
грамположительных бактерий (стафиллококков,
стрептококков, диплококков и некоторых других) и практически неактивен в
отношении грамотрицательных видов и дрожжей.
Высокие
концентрации пенициллина (10 мг/мл) вызывают гибель клеток гаплоидного
штамма дрожжей Saccharomyces cerevisiae
и E. coli (Lingel, oltmanns, 1963).
По
характеру действия на микроорганизмы пенициллин является бактериостатическим и
при определенных концентрациях бактерицидным антибиотиком.
Различные
типы пенициллинов обладают различной степенью биологической активности. В
особенности это различие заметно в опытах
in vivo (таблица 8).
Таблица 8.
Сравнение биологической активности различных типов
пенициллинов в отношении некоторых микроорганизмов в опытах in
vivo.
|
Тест - организм
|
Относительная активность
|
|
Бензилпенициллин (G)
|
2-пентилпенициллин (F)
|
n-гептилпенициллин (K)
|
Оксибензилпенициллин (Х)
|
|
Spirochaeta novyi
Pneumococcus типа 1
Strept. haemolitycus
Strept. pyogenes
Treponema pallidum
|
100
100
100
100
100
|
55
85
100
50
17
|
35
17
60
9
9
|
22
140
500
260
5
|
Как
следует из данных таблицы, n-гептилпенициллин менее активен, чем остальные типы
пенициллинов. Это, по-видимому, связано с тем, что n-гептилпенициллин
значительно быстрее инактивируется в организме.
Чувствительные
к пенициллину микроорганизмы относительно легко и быстро приобретают
устойчивость к антибиотику. Так, Staph. aureus прекращает развитие при концентрации пенициллина
0,05 - 0,06 ед/мл в среде, но уже при 20
последовательных пересевах с постепенно увеличивающимися концентрациями
антибиотика устойчивость стафилококка возрастает в 700 раз, т.е. для остановки
роста бактерии требуется концентрация пенициллина равная 42 ед/мл,
а после 40 пересевов его устойчивость возрастает более чем в 5500 раз.
Микроорганизмы,
приобретшие устойчивость к одному из типов пенициллина, как правило, резистентны и к другим типам пенициллина.
У
некоторых бактерий устойчивость к пенициллинам сопровождается способностью
образовывать пенициллиназу.
В
ряде случаев микроорганизмы с приобретением устойчивости к пенициллину теряют
вирулентность. Но вирулентность восстанавливается после нескольких пассажей
через животных и при этом сохраняется резистентность
к антибиотикам.
Таблица 9.
Различные типы пенициллинов и строение их радикалов
|
Название пенициллина
|
Строение радикала (R)
|
|
Общепринятое
|
условное
|
|
Бензилпенициллин
n-Оксибензилпенициллин
2-Пентенилпенициллин
n-Гептилпенициллин
n-Амилпенициллин
Феноксиметилпенициллин
Аллилмеркаптометилпенициллин
|
G
X
F
K
ДигидроF
V
O
|
|
Таблица 10.
Пенициллины,
полученные в результате смешанного (биологического и химического) синтеза
(полусинтетические пенициллины).
|
Тип пенициллина
|
Общепринятое название
|
Строение радикала
|
|
Кислотоустойчивые
препараты
|
|
a-Феноксиэтилпенициллин
|
Фенетициллин
|
|
|
a-Феноксипропилпенициллин
|
Пропициллин
|
|
|
a-Феноксибензилпенициллин
|
Фенбенициллин
|
|
|
Пенициллиназоустойчивые препараты
|
|
2-6-диметоксифенилпенициллин
|
Метициллин
|
|
|
Кислото- и пенициллиназаустойчивые
препараты
|
|
5-метил-3-фенил-4-изоксиазолилпенициллин
|
Оксациллин
|
|
|
2-этокси-1-нафтилпенициллин
|
Нафциллин
|
|
|
2-бифенилпенициллин
|
Дифенициллин
|
|
|
3-О-хлорфенил-5-метил-4-изооксазолил
|
Клоксациллин
|
|
|
Кислотоустойчивые и широкоспектровые препараты
|
|
a-d-(-)Аминобензилпенициллин
|
Ампициллин
|
|
а) Стрептомицин (Streptomycin) - аминогликозидный
антибиотик.
В
группу аминогликозидных антибиотиков включаются
биологически активные соединения, содержащие в молекулах гликозидные
связи. К этим антибиотикам относятся стрептомицины, неомицины,
канамицины, гентамицины, гигромицин и некоторые другие вещества.
Антибиотики
этой группы имеют большое практическое значение, многие из них применяются в
медицинской практике.
Актиномицет, образующий стрептомицин, впервые
выделен в лаборатории микробиологии Раттерского
университета в 1943 г.
Первое
сообщение о выделении антибиотика было сделано Шатц, Буги и Ваксманом в январе 1944 г.
Антибиотик получил название стрептомицин (от родового названия актиномицетов Streptomyces), а
организм, образующий этот антибиотик, был определен как Streptomyces
griseus.
Стрептомицин
образуют не только штаммы Act. Streptomycini,
но и другие актиномицеты - Act.
bikiniensis, Act. raneus, Act. humidus,
Act. reticuli, Act. griseocarneus, Act. mashuensis.
Актиномицет, продуцирующий стрептомицин, как и
другие виды актиномицетов, может размножаться с
помощью спор или отдельных участков мицелия.
Культура
актиномицетов вообще и Act.
streptomycini в частности, весьма вариабельны.
На изменчивость актиномицетов оказывают влияние
условия их культивирования и, в особенности, состав среды. На более богатых по
составу средах наблюдается более быстрая изменчивость актиномицетов.
В
результате изменчивости продуцента стрептомицина нередко появляются аспорогенные формы, т.е. формы, лишенные воздушного
спороносного мицелия. Как правило, эти варианты или вообще неактивны, или же
образуют незначительное количество стрептомицина.
Снижение
образования антибиотика наблюдается и у вариантов с усиленной стимуляцией.
Образующиеся
в результате изменчивости Act. streptomycini
варианты могут отличаться от исходной культуры окраской воздушного мицелия;
последний может быть белым, беловато-палевым, светло-серым, серым и
серо-зеленым. Различные варианты отмечаются друг от друга по величине и форме
колоний. Встречаются так же формы, неспособные образовывать стрептомицин.
Однако установить какие-либо цитологические различия между активными и
неактивными вариантами не удалось.
б) Антибиотические свойства
стрептомицина.
По
отношению к стрептомицину все микроорганизмы условно можно разделить на три
группы (Шемякин, Хохлов и др., 1961).
1.
Весьма чувствительные микроорганизмы, которые
подавляются в большинстве случаев при концентрации стрептомицина 10 мкг/мл. союда можно отнести
организмы, принадлежащие к следующим родам: Bacillus,
Bordetella, Brucella, Klebsiella, Mycobacterium, Bacteroidum и некоторые другие.
2.
Умеренно чувствиетльные. Для
подавления которых in vitro
необходимо иметь концентрацию стрептомицина в пределах 10 - 100 мкг/мл. К этой группе относятся многие бактерии из родов Aerobacter, Corynebacterium, Diplococcus, Proteus, Staphylococcus, Strepticoccus, Vibrio.
3.
Устойчивые формы микробов, для подавления которых
необходима концентрация антибиотика, превышающая 100 мкг/мл.
сюда относятся виды Bacteroides, Clostridium,
некоторые виды Proteus, многие виды грибов, дрожжей,
риккетсий, вирусы.
Итак,
различные организмы по-разному реагируют на присутствие в среде стрептомицина. Степень антимикробного действия антибиотика также различна в
отношении различных видов организмов (таблица 11).
Таблица 11.
Антибиотическая активность стрептомицина in
vitro.
|
Микроорганизм
|
Концентрация стрептомицина
(мкг/мл), вызывающее подавление
|
|
Наиболее чувствительные
штаммы
|
Наиболеее устойчивые штаммы
|
Большинство штаммов
|
|
Aerobact. aerogenes
Bac. anthracis
Bac. megatherium
Bac. subtilis
Candida albicans
Clostridium botulinum
Corinebact. giphtheriae
Diplococcus pneumniae
E. coli
Mycob. tuberculosis
Proteus vulgaris
Ps. aeroginosa
Bact. thyphi
Bact. dysenteriae
Bac. cereus
|
0,300
0,250
0.250
0.056
-
-
0.400
0.500
0.015
0.100
1.000
0,100
0,004
2,000
0,830
|
1000
10
4
128
-
-
200
50
>1000
12,5
>1000
1000
20
8
2
|
25
5
2
25
Устойчивы
>>
20
25
25
5
15
50
5
5
1
|
Наряду
с тем, что стрептомицин подавляет рост многих видов микроорганизмов, к нему
довольно легко появляется устойчивость, возникают формы бактерий, резистентные
к стрептомицину. По данным Прайса (Price et al.,
1947), повышение устойчивости к стрептомицину в 1 000 раз возникает у
золотистого стафилококка всего лишь через три пассажа на бульоне с
возрастающими концентрациями антибиотика, а у Bact. typhi повышение устойчивости в 22 600 раз происходило через
14 пассажей.
Образование
устойчивых форм бактерий к стрептомицину происходит также in
vivo. Приобретенная к стрептомицину устойчивость
сохраняется у организмов довольно длительное время. С возникновением
устойчивости появляются некоторые изменения в характере обмена веществ. так, у
резистентного к стрептомицину хромогенного микроорганизма происходит резкое
изменение его окраски. Стрептомициноустойчивая форма
синегнойной палочки теряет способность образовывать пигмент, изменяются и
некоторые другие стороны обмена.
Однако
у устойчивых и чувствительных к стрептомицину штаммов бактерий не наблюдается
заметных различий в вирулентности.
В
ряде случаев под действием стрептомицина в опытах in vitro возникают не только устойчивые к нему штаммы, но и
зависимые от стрептомицина формы, способные развиваться только в присутствии
данного антибиотика.
Описаны
случаи, когда штаммы менингококка, Mycob. ranae и другие микроорганизмы развиваются лишь на среде,
содержащей от 100 до 150 мкг/мл стрептомицина.
Стрептомициноустойчивые и зависимые от
стрептомицина штаммы обычно получаются из чувствительных форм микроорганизмов.
Соотношение между чувствительными, устойчивыми и зависимыми от стрептомицина
штаммами изображено на рисунке.
3. Поглощение антибиотиков клетками
микробов.
Первый
этап во взаимодействии микроорганизмов с антибиотиками - адсорбция его
клетками. Пасынский и Косторская
(1947) впервые установили, что одна клетка Staphylococcus
aureus поглощает примерно 1 000 молекул пенициллина.
В последующих исследованиях эти расчеты были подтверждены. Так, по данным Мааса
и Джонсона (1949), приблизительно 2·10-9
М пенициллина поглощается 1 мл стафилококков, причем около 750 молекул этого
антибиотика необратимо связываются одной клеткой микроорганизма без видимого
эффекта на ее рост.
Игл
с сотрудниками (1955) определил, что при связывании бактериальной клеткой 1 200
молекул пенициллина угнетения роста бактерий не наблюдается. Угнетение роста
микроорганизма на 90 % наблюдается в тех случаях, когда клеткой будет связано
от 1 500 до 1 700 молекул пенициллина, а при поглощении до 2 400 молекул на
клетку происходит быстрая гибель культуры.
Установлено,
что процесс адсорбции пенициллина не зависит от концентрации антибиотика в
среде. При низких концентрациях препарата (порядка 0,03 мкг/мл)
он может весь адсорбироваться клетками, и дальнейшее повышение концентрации
вещества не поведет к повышению количества связанного антибиотика.
Имеются
данные (Купер, 1954) о том, что фенол препятствует поглощению пенициллина
клетками бактерий, однако он не обладает способностью освобождать клетки от
антибиотика.
Пенициллин,
стрептомицин, грамицидин С, эритрин и другие
антибиотики связываются различными бактериями в заметных количествах. Причем антибиотики-полипептиды адсорбируются микробными клетками в
большей степени, чем, например, пенициллины и стрептомицин.
Булгакова
и Полин (1966) установили, что грамицидин С адсорбируется как чувствительными,
так и устойчивыми к нему бактериями. Причем адсорбция этого антибиотического
вещества бактериями происходит сразу же после внесения антибиотика в суспензию
клеток и достигает значительных величин (до 500 мкг/мг
сухой биомассы).
В
присутствии положительно заряженных ионов (Na+,
K+, NH+, Mg2+), а также при рН среды, равном 4,
поглощение грамицидина С бактериальными клетками заметно снижается.
Адсорбированный
чувствительными клетками грамицидин С прочно связывается с бактериями и
снимается с них лишь при длительной экстракции подкисленной спиртово-водной
смесью.
У
устойчивого к антибиотику штамма E. coli при
промывании клеток раствором NaCl удается удалить лишь
до 30% адсорбированного грамицидина С.
Ванкомицин, образуемый Actinomyces
orientalis, необратимо и относительно быстро
связывается клетками бактерий, чувствительными к нему. По данным Reunolds (1966), при концентрации антибиотика в среде,
равной 30 мкг/мг массы сухих бактерий, около 90% ванкомицина связывается с бактериями. Показано, что
максимальное связывание ванкомицина бактериальными
клетками достигает величин 107 молекул антибиотика на клетку.
Связанные
клеткой антибиотики могут проявлять двоякого рода действия: содной
стороны, некоторые из них могут действовать как поверхностно-активные вещества,
а с другой стороны, антибиотики, проникая вглубь клетки, нарушают отдельные
стороны метаболизма организма.
Гибель
клеток под воздействием поверхностно-активных антибиотиков может быть связана с
нарушением механизма осмотического равновесия, имеющего место на поверхности
микробной клетки; она может также наступить в результате способности этих
веществ, скапливающихся у поверхности раздела жидкая фаза - микробная клетка,
связываться с компонентами клеток.
Экспериментальные
данные в этом отношении показали, что изменения в регулировании осмотического
давления сопровождаются разрушением систем клеточной стенки микроба. Нарушение
проницаемости клеточной стенки является или результатом прямого вливания
антибиотических веществ, или же может быть результатом вторичных процессов.
Действие
антибиотиков как поверхностно-активных веществ может вызывать диссоциацию белка
с отделением от него простетических групп или
нуклеиновых кислот. Эти вещества могут также приводить к денатурации белков и,
таким образом, непосредственно влиять на энзиматические
системы, связанные с клеточной стенки (инвертазы, фосфатазы, различные дегидрогеназы,
цитохромные системы).
Таким
образом, если антибиотик обладает способностью нарушать системы, регулирующие
осмотические свойства клеточной стенки, иными словами, если антибиотик
выступает в качестве поверхностно-активного соединения, то он может оказывать
бактерицидное действие.
К
числу антибиотических веществ, механизм действия которых связан с
поверхностно-активными свойствами, следует отнести грамицидин С, тироцидин, полимиксины, а также
тетрациклины, если последние применяются в концентрациях, во много раз превышающие
бактериостатические.
Вместе
с тем, как уже отмечалось выше, антибиотики, попадая в микробную клетку, могут
нарушать отдельные этапы метаболизма организма, подавлять некоторые звенья в
цепи биохимических процессов.
Подавление
отдельных звеньев в биохимической цепи превращений может происходить как путем
необратимого связывания антибиотиком одного из компонентов реакции, так и в
результате конкурентного подавления биологически важных метаболитов клетки.
Если
в организме или в среде одновременно присутствуют два вещества - обычный для
организма субстрат S и ингибитор I, сходный по структуре с субстратом и оба они
могут вступать в связь с энзимом (Е), как показано в
уравнениях (1 и 2):
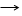
 S+E SE (1)
S+E SE (1)
I+E IE (2)
то
мы имеем типичный пример конкурентного подавления или конкурентного обмена.
В
качестве примера конкурентного обмена можно привести участие сульфаниламина как антиметаболита n-аминобензойной кислоты
в витамине фолиевой кислоты.
Как
только сульфаниламид, в случае его присутствии в клетке,
включается в фолиевую кислоту вместо ПАБК,
ферментативные функции бактериальной клетки блокируются. Это в свою очередь
приводит к нарушению механизма обмена веществ клетки и, вслед за этим, к ее
гибели.
В
качестве примера конкурентного действия антибиотика при биосинтезе белка можно
назвать пуромицин. Этот антибиотик образуется
культурой Act. alboniger.
По химическому строению пуромицин - нуклеозидное производное
6-диметил-амино-9(3'-пара-метокси-L-фенилаланил-амино-3'-дезокси-b-D-рибофуранозил)-пурина.
Пуромицину соответствует следующая формула:
Анализ
химической структуры пуромицина показал, что он
представляет собой структурный аналог 3-конечной аминоацилированной
группировки тРНК.
Суммирование
имеющихся данных по механизму биологического действия пуромицина
дает возможность заключить, что оно выступает в качестве конкурентного аналога аминоацил-тРНК, заменяя последнюю в реакции с пептидил-тРНК, что приводит к освобождению пептидила в виде пептидил-пуромицина
из рибосомы и, таким образом, прекращая синтез белка
(Спирин, Гаврилова, 1971).
Есть
указания на то, что окситетрациклин выступает в
качестве конкурентного ингибитора дифосфопиридиннуклеотида
при действии его на E. coli.
Установлено,
что актитиазовая кислота (антибиотик актиномицетного происхождения) - конкурент витамина биотина, сходного по строению с кислотой. Вместе с тем
необходимо подчеркнуть, что явление конкурентного ингибирования
не имеет широкого распространения в механизме действия антибиотических веществ.
Существует
гипотеза, впервые высказанная Ч. Кэвеллито, что
биологическая активность многих антибиотиков (бензилпенициллина,
стрептомицина, аллицина, пиоцианина и др.)
обусловлена тем, что они вступают в связь с сульфгидрильными группами (-SH) ферментов, превращая их в неактивные вещества, но
такая точка зрения на механизм действия антибиотиков не была строго
обоснованной.
Однако
появились указания (Gross, Morell,
1967) на то, что механизм биологического действия антибиотика низина
связан с взаимодействием его с сульфгидрильными группами метаболитически
важных ферментов (глютатин, ацетилкоэнзим
А).
V. Симбиотические связи бактерий с бобовыми.
Симбиотические
связи бактерий с бобовыми благодаря широкому использованию их в полевом
растениеводстве, луговодстве и, частично, в лесоводстве, изучались весьма
интенсивно. Установлено, что не все виды бобовых имеют на корнях клубеньки. При
обобщении имеющихся данных, оказалось, что из 1285 изученных бобовых (в широком
понимании этой группы) клубеньки отсутствовали у 166 (13,0%), в том числе у 77,4%
изученных цезальпиновых, у 13% мимозовых и 7% -
мотыльковых (Fabaceae) (E. Allen, O. Allen, 1961).
Отсутствие
клубеньков на корнях не всегда означает неспособность данного вида бобовых к
симбиозу с клубеньковыми бактериями, иногда это происходит из-за местных
условий, неблагоприятных для образований клубеньков, или по тому, что в почве
нет соответствующих рас клубеньковых бактерий. В то же время наличие клубеньков
на корнях бобовых не всегда указывает на активную фиксацию азота клубеньковыми
бактериями. Установлено, что многочисленные мелкие белые клубеньки на боковых
корнях травянистых бобовых образованы малоэффективной расой клубеньковых
бактерий, неспособной связывать атмосферный азот или фиксирующий его в
незначительном количестве, в то время как крупные, окрашенные в розовый цвет
клубеньки на стержневом корне обычно характеризуют энергично идущий процесс
усвоения азота.
А
процесс естественного отбора и сопряженной эволюции возникло много рас
клубеньковых бактерий, способных выступать в эффективные симбиотические
отношения с определенными видами бобовых. Значение отдельных рас клубеньковых
бактерий давно было выяснено для возделываемых видов бобовых, и, в связи с
необходимостью в ряде случаев вносить бактериальное удобрение (нитраты),
содержащие соответствующие клубеньковые бактерии, они были разделены на ряд
групп в соответствии с пригодностью для определенных видов бобовых. Принято
выделять следующие расы клубеньковых бактерий по их способности к эффективному
симбиозу со следующими определенными видами или группами видов бобовых:
1)
горох, вика, кормовые бобы;
2)
фасоль; 3) соя; 4) люпин, сераделла;
5)
вигна, магу, арахис; 6) нуж;
7) клевер;
8)
люцерна, донник, пажитник; 9) эспарцет;
10)
лядвенец (Вознесенская, 1969).
На
самом деле число рас клубеньковых бактерий значительно больше.
Выявлена
специфичность рас клубеньковых бактерий в пределах уже установленных групп, в
частности в "клеверной группе", например у клевера несходного,
клубеньки возникают лишь при участии особой расы клубеньковых бактерий.
Эта
специфичность проявляется также в значительных различиях фиксируемого ими азота
в зависимости от вида клевера, с которым они связаны. В таблице 12 показано,
что наиболее эффективные для клеверов лугового и ползучего расы клубеньковых
бактерий для клевера подземного были наименее эффективными, и наоборот. Расы,
обеспечивающие фиксацию очень значительных количеств азота при инокуляции ими
клевера подземного, в симбиозе с клевером луговым и ползучим фиксировали
незначительное количество азота. Все это обусловило выделение среди
"клеверной группы" трех подгрупп:
А - клевера ползучий, луговой, розовый, простертый, зямляничный;
В - подземный, инкармантный, скученный,
александрийский; С - несходный. Три подгруппы выделяют и среди "люцерновой
группы".
Таблица 12.
Способность отдельных рас клубеньковых бактерий фиксировать
атмосферный азот при симбиозе с различными видами клевера (содержание азота в
мг на 8 растений; по White et
al., 1953).
|
Виды клевера
|
Расы бактерий
|
|
№1
|
№16
|
№8
|
№13
|
|
Клевер луговой
Клевер ползучий
Клевер подземный
|
51,4
59,6
7,9
|
34,3
46,5
7,2
|
2,8
10,5
138,5
|
1,9
7,0
153,5
|
Возможность
фиксации атмосферного азота клубеньковыми бактериями и количество
фиксированного азота определяются также средой - отсутствием условий,
ограничивающих жизнедеятельность бактерий и бобовых растений (высокая
кислотность, высокое содержание растворимого алюминия, плохая аэрация и др.), а
также достаточной обеспеченностью фосфором, калием, кальцием, молибденом,
серой, кобальтом, водой и др. Фиксация азота клубеньковыми бактериями снижается
по мере увеличения содержания в почве растворимых форм азота, доступных для
бобовых. Большое значение имеют условия освещения, поскольку клубеньковые
бактерии получают от бобового углеводы, необходимые им как энергетический
материал для фиксации азота, и потому зависят от фотосинтеза. При затенении
резко снижается не только число клубеньков, но и их размеры, а также предельная
глубина их образования.
Клубеньковые
бактерии более экономно используют энергию, необходимую для фиксации азота,
затрагивая 3-4 г углеводов на 1 г азота, в то время как свободноживущие
азотфиксирующие бактерии затрачивают 50 - 100 и более граммов на фиксацию 1 г
азота. Это связано с тем, что у свободноживущих азотфиксаторов фиксация азота
происходит в процессе их роста, и потому большое количество энергии
потребляется на этот рост. Кроме того, в целях создания благоприятных условий
для активности нитрогеназы - фермента, участвующего в
фиксации азота, для снижения парциального давления кислорода усиливается
дыхание, что связано с затратой энергии. Эти расходы энергии отсутствуют у
клубеньковых бактерий. Поскольку фиксация азота происходит в бактероидах,
клетках, прекративших рост, а внутри клубеньков создаются благоприятные условия
для активности нитрогеназы. В том числе сниженное
содержание кислорода. Очень существенно то, что фиксируемый клубеньковыми
бактериями азот на 90 - 95% передается бобовым растениям. Бобовые, получая
связанный азот от клубеньковых бактерий, не зависят или мало зависят от
обеспечения минеральным азотом почвы и потому могут успешно произрастать
совместно с другими растениями на почвах, бедных доступными формами азота.
Количество
азота, фиксируемого клубеньковыми бактериями бобовых, сильно варьирует от
фитоценоза к фитоценозу, а в пределах конкретных фитоценозов может изменяться
от года к году. Оно определяется участием бобовых в фитоценозах, условиями
среды и эффективностью соответствующих рас бактерий. Для некоторых лугов в
Новой Зеландии с травостоями, где преобладает клевер, отмечена фиксация азотом
до 450 - 550 кг/га.
Фиксация
азота в столь больших количествах возможна лишь в условиях исключительно
благоприятного климата Новой Зеландии (равномерное распределение большого
количества атмосферных осадков, отсутствие засух, благоприятные тепловые
условия, возможность вегетации в течении всего года), когда растения в
результате применения известкования и внесения удобрений произрастают в
условиях благоприятных для них реакций почвы и обеспечены необходимыми зольными
элементами. Однако, даже в Новой Зеландии среднее количество азота,
фиксируемого клубеньковыми бактериями бобовых, входящих в состав луговых
травостоев, составляло 185 кг/га (колебания достигали 85 - 340 кг/га от луга к
лугу и в среднем от года к году 145 - 225 кг/га). В годы с более влажным и прохладным
летом оно было выше, а в годы с сухим, более теплым летом - ниже. Данные о
сходных количествах фиксируемого азота клубеньковыми бактериями бобовых
получены для Ирландии (в среднем 160 кг/га) и Южной Англии (250 - 270 кг/га) и
относятся к лугам, созданным путем посева трав. На природных лугах нашей
страны, в травостоях которых бобовые принимают меньшее участие, количество
азота, фиксируемого клубеньковыми бактериями, как правило, не более 30 - 50
кг/га. В посевах многолетних трав (бобовых или бобовых в смеси со злаками)
количество фиксируемого азота обычно не превышает 200 кг/га.
Давно
замечено, что бобовые благоприятно влияют на произрастающие совместно с ними
злаки и другие растения, в том числе увеличивают содержание азота в их органах.
На основе результатов вегетационных опытов Виртанен пришел к выводу, что такое
воздействие связано с прижизненными выделениями из корней бобовых в почву
значительных количеств азотсодержащих соединений. Проверка результатов опытов
Виртанена, проведенная в СССР, Шотландии, США, Австралии, не подтвердила его
выводов. Оказалось, что в условиях нормального фотосинтеза бобовые не выделяют
в почву сколь-либо заметных количеств азотистых
соединений. Лишь с ослаблением фотосинтеза (при сниженной интенсивности
освещения), когда растения не располагают достаточным количеством углеводов и
органических кислот для связывания всего фиксируемого клубеньковыми бактериями
азота, часть его может выделяться в почву. Благоприятное влияние бобовых на
другие растения можно объяснить поступлением в почву азота с их отмирающими
органами, у древесных растений - в основном с опадом,
у травянистых (в фитоценозах, используемых как сенокосы и пастбища) -
преимущественно с отмирающими подземными органами. На пастбищах злаки получают
азот бобовых из экскрементов скота, поедающих их. Так как бобовые, если
фиксация азота клубеньковыми бактериями идет достаточно активно, не поглощают
из почвы азот или поглощают его в незначительных количествах, на долю небобовых растений остается больше доступных форм азота, и
потому они растут лучше с бобовыми, чем в их отсутствие. Совместное
произрастание бобовых с небобовыми растениями
оказывает благоприятное влияние на фиксацию азота клубеньковыми бактериями, так
как в результате поглощения небобовыми азота из почвы
содержание его доступных форм снижается до незначительных величин, что
стимулирует фиксацию азота.
Биологическая фиксация азота.
Баланс
доступного растениям азота на Земном шаре поддерживается за счет деятельности
особой группы организмов - так называемых азотфиксаторов. В экономике природы
процессам биологической фиксации азота принадлежит исключительная роль, которая
по значению вполне равнозначна процессу фотосинтеза.
В
группу азотфиксаторов входят свободно живущие организмы, а также организмы,
способные существовать лишь в симбиозе с другими видами.
Первой
в ряду свободно живущих азотфиксаторов открыта анаэробная спороносная бактерия Clostridium pasterianum. Это
открытие принадлежит русскому ученому С. Н. Виноградскому
(1893). Через 8 лет (1901) М. Бейеринком был открыт
аэробный микроорганизм, названный азотбактером (Azotobacter).
Оба
организма являются сапрофитами. Для восстановления молекулярного азота они
используют энергию, получаемую ими при окислении глюкозы и других органических
соединений (например, маннита). На каждый грамм сброженной глюкозы азотбактер
накапливает около 15 мг связанного азота, тогда как Clostridium - не более 2 -3 мг.
Выделив
азотбактер, Бейринк обратил
внимание на большое сходство свойств этого микроорганизма со свойствами фотосинтезирующей
бактерии Chromatium. В
настоящее время установлено, что способность фиксировать азот широко
распространена у различных видов бактерий. Эта функция свойственна, в
частности, сульфатредуцирующим бактериям, развитие которых осуществляется в
анаэробных условиях. Способностью ассимилировать молекулярный азот обладает
также пурпурная бактерия Rhodospirillum rubrum (Чест и Камен).
К
свободно живущим азотфиксаторам принадлежат также синезеленые
водоросли (Nostoc, Phormidium).
Осуществляемая ими ассимиляция молекулярного азота имеет в особенности большое
значение для пресноводных бассейнов, для развития растений риса на заливаемых
водой плантациях и т.п. Благодаря своей крайне четко выраженной автотрофности синезеленые
водоросли способны заселять голые скалы; они развиваются на вулканической пыли,
пензе и т.д.
Общий
уровень азотфиксирующей активности свободно живущих организмов невысок. В
зависимости от вида и условий существования они накапливают в год от 10 до 30 -
40 кг связанного азота на гектар.
Основную
роль в пополнении убыли запасов связанного азота выполняют бактерии - симбиоты, в первую очередь Bacterium radicicola. В настоящее
время известно, что кроме бобовых имеется еще около 100 видов других растений,
на корнях которых развиваются специфические для каждого растения клубеньковые
бактерии.
Химическая
природа взаимодействия бобовых с клубеньковыми бактериями изучена неполно, в
связи с чем ряд факторов в этой области не находит еще объяснения. Например,
установлено, что клубеньковые бактерии способны восстанавливать молекулярный
азот только при взаимодействии с корнями бобовых растений. Этой способности
лишены бактерии, культивируемые в искусственных средах. Клубеньковые бактерии
развивались на корнях бобовых растений и в том случае, если последние служили
подвоем, а привоем - любое другое растение. В тех вариантах, где бобовые
служили привоем, а подвоем - различные небобовые
растения, бактерии не развивались.
Все
это показывает, что тканям бобовых должны быть свойственны определенные
особенности.
Представитель
экспериментального направления в физиологии растений. Замечательный французский
ученый Жан Батист Буссенго
первый широко использовал в своих исследованиях по питанию метод выращивания
растений в вегетационных сосудах. Точные эксперименты позволили Буссенго опровергнуть представления Либиха об азотном
питании растений. Буссенго первый отметил
специфические особенности бобовых растений как азотсобирателей,
а вслед за тем Гельригель открыл, что эту роль
бобовые выполняют в симбиозе с клубеньковыми бактериями. Большое значение имели
исследования русского ботаника М. С. Воронина, который первый сформулировал
представление о клубеньках как о болезненных образованиях,
возникающих в результате заражения корней.
Симбиоз
корней высших растений с грибами носят название микориз, симбиозы с бактериями
- бактериориз. В зависимости от формы симбиотрофизма различают микоризы экто-
и эндотрофные. Первые развиваются на поверхности корней, вторые проникают
внутрь ткани. Классическим примером бактериотрофизма
является взаимодействие бобовых растений с клубеньковыми бактериями.
Исследования
микробиологов, проведенные в последние годы, значительно расширили
представления о значении симбиотрофизма в процессах
корневого питания растений.
Число
микробных телец, приходящихся на 1 г почвы, исчисляется многими сотнями
миллионов и даже миллиардами. Некоторое представление об этом дают цифры,
заимствованные из работ Н. А. Красильникова:
|
Число микроорганизмов в 1
г почвы
|
Вес микробных тел в
пахотном слое, кг/га
|
|
Бактерии
Грибы
Актиномицеты
Водоросли
|
1´109
1´105
1´105
1´103
|
600
60
8
160
|
Почвенные
микроорганизмы сосредоточены в основном в зоне размещения корней.
Важное
значение азотфиксирующих бактерий диктует необходимость обеспечения наиболее
благоприятных условий для их развития и физиологической деятельности.
Один
из путей решения этой важной практической задачи состоит в увеличекнии
численности микробного населения почвы с помощью бактериальных удобрений.
Данное
мероприятие в равной степени оправдывает себя в отношении как клубеньковых
бактерий, так и свободно живущих азотфиксаторов.
Дело
в том, что даже при длительной культуре бобовых растений, почвы, занятые ими
содержат нередко недостаточное количество Bact. radicicola, в результате чего на корнях образуется мало
клубеньков, либо они не образуются вовсе. В этих условиях бобовые не обогащают
почву азотом, а аналогично другим растениям истощают имеющиеся в ней азотистые
соединения.
Искусственное
обогащение почвы клубеньковыми бактериями оказывает весьма благоприятное
влияние на азотный баланс почвы и на развитие бобовых и других растений
севооборота. Препарат клубеньковых бактерий, который называется нитрагином, вносят вместе с семенами
бобовых при посеве. При приготовлении нитрагина следует учитывать специфичность
клубеньковых бактерий.
VI. Воздействие внешних факторов на рост и развитие
растений.
Измерение
скорости роста, проведенное немецким физиологом Ю. Саксом (1872), позволило
установить определенные закономерности. В начальный период темпы роста, как
правило, низкие. Затем рост усиливается и идет с большой скоростью (период
большого роста), а затем снова замедляется. В результате рост (увеличение
размера) клетки, органа или организма в целом может быть изображен в виде Ы-образной кривой. Эта закономерность имеет общебиологическое
значение и справедлива по отношению к росту всех живых организмов, включая и
человека.
Анализируя
полученную кривую, можно ее разделить на 3 участка: 1) фаза, когда рост
изменяется экспоненциально (логарифмически); 2) фаза, когда рост изменяется линейно;
3) фаза торможения роста. Именно период, когда рост прямо (линенйно)
возрастает, называется периодом большого роста.
Влияние внешних условий на рост.
Внешние
условия оказывают на рост как прямое, так и косвенное влияние. Последние
связаны с тем, что скорость роста зависит от интенсивности всех остальных
физиологических процессов, воздушного и корневого питания, снабжения водой,
напряженности процессов обмена веществ и энергии. В этой связи влияние внешних
условий может сказаться на интенсивности роста через изменения любого из
указанных процессов. При этом далеко не всегда причина того или иного влияния
можно с достаточной точностью установить, поскольку в естественной обстановке
влияние отдельных факторов тесно взаимосвязано.
Температура. Рост растений возможен в сравнительно широких
температурных пределах. Растения ранневесенней флоры
растут при температуре даже несколько ниже 0°С. Есть
растения, для которых верхняя температурная граница роста несколько превышает
50°С. для каждого вида растения, в зависимости от его
особенностей и, главным образом, от географического происхождения, характерны
определенные температурные границы, в которых возможно протекание ростовых
процессов. Различают три кардинальные температурные точки: минимальная
температура, при которой рост только начинается, оптимальная - наиболее
благоприятная для ростовых процессов, максимальная, при которой рост
прекращается. С повышением температуры от минимальной до оптимальной скорость
роста резко возрастает. Так, скорость роста проростков гороха при повышении
температуры от 0 до 10°С возрастает в 9 раз, от 10
до 20°С - в 2,5 раза,
от 20 до 30°С - всего в 1,9 раза. Оптимальные температуры могут
быть неодинаковыми для роста разных органов одного и того же растения. Как
правило, оптимальная температура для роста корневых систем ниже по сравнению с
надземными органами. Для роста боковых побегов оптимальная температура ниже по
сравнению с ростом главного стебля. Установлено, что растения интенсивно растут
в ночной период суток. Для роста многих растений благоприятной является сменная
температура в течение суток - днем повышенная, а ночью - пониженная. Это
явление Ф. Вент назвал термопериодизмом. Явление термопериодизма хорошо проявляется на культуре томатов. Показано
(Н. И. Якушкина), что пониженные ночные температуры ускоряют рост корневой
системы и боковых побегов у растений. Такое влияние может быть объяснено тем,
что при понижении температуры более активно работают ферменты,
катализирующие распад крахмала на
сахара. В листьях образуются растворимые транспортные формы углеводов, легко
передвигающиеся к точкам роста корня и боковых побегов, благодаря чему их рост
усиливается.
Содержание воды. В процессе роста растения особенно чувствительны к
недостатку воды. Уменьшение содержания
воды в почве приводит, естественно, и к уменьшению содержания ее в растении, а
это, в свою очередь, резко тормозит процессы роста. Снижается деление клеток и особенно
их рост растяжением. Наибольшая насыщенность водой требуется для процессов
роста. Насыщенность клетки или ткани растений водой называют гидратурой, она выражается в процентах. Рост клеток
идет лишь в том случае, если гидратура не падает ниже
95%. Для того чтобы поддержать такую гидратуру, точки
роста надземных органов растений защищены смыкающимися листочками с хорошо
развитой кутикулой. Точки роста корня не имеют подобной защиты и поэтому
требуют повышенной влажности почвы для своего роста.
Свет. Растения используют свет двумя путями: первый, где свет
служит источников энергии для синтеза органических веществ (фотосинтез);
второй, где свет - сигнал или источник информации. Во втором случае энергия
света может быть на несколько порядков ниже, чем в первом. Свет оказывает
большое и разностороннее влияние на темпы и характер роста как отдельных
органов, так и растительного организма в целом. При этом на разные стороны
ростовых процессов влияние света проявляется неоднозначно. Так свет необходим
для протекания процесса фотосинтеза, и, поэтому, накопление массы растения без
света не идет. Вместе с тем, рост клеток растяжением может идти в темноте,
более того, на свету этот процесс тормозится. Свет оказывает большое влияние на
формообразовательные процессы. Этиолированные проростки,
выросшие в темноте, характеризуются рядом анатомических и морфологических
особенностей. В отсутствие света происходит упрощение анатомической структуры
стебля. Слабо развиваются ткани центрального цилиндра, механические ткани. Вместе
с тем, растяжение клеток в темноте идет очень интенсивно. В результате
образуются длинные, вытянутые стебли. Листья редуцированы, у двудольных
растений вместо листовой пластинки образуются лишь небольшие чешуйки.
Этиолированные проростки имеют слегка желтоватый
оттенок. По-видимому, более быстрое вытягивание стебля и корня этиолированных проростков выработалось в процессе эволюции,
так как в большинстве случаев прорастание семян в почве происходит в отсутствие
света, и эти особенности, а также отсутствие листьев, облегчают проростку
проникновение через слой почвы. Возможно, что вытягивание стебля в отсутствие
света является следствием отсутствия ингибиторов роста. В темноте образуются
много гормонов ауксинов. Нарушение соотношения ауксинов и ингибиторов вызывает несбалансированный
рост. При выходе проростков на поверхность почвы происходят их внутренние и
внешние изменения. В темноте у проростков двудольных растений гипокотиль изогнут, что защищает точку роста в почве от
повреждений. Под влиянием света этот изгиб ("крючок") выпрямляется. На
свету рост стебля тормозится, рост листьев усиливается и они принимают обычную
форму. Под влиянием света происходят анатомические изменения стебля,
дифференцируется эпидермис, появляются волоски, изменяется окраска -
синтезируется хлорофилл. Эти изменения получили название фотоморфогенеза.
Говоря
о воздействии света, необходимо остановиться на влиянии круглосуточного
освещения на процессы роста. Опыты по выращиванию растений при свете
электрических ламп (светокультура) показали, что рост многих растений при
круглосуточном освещении идет значительно интенсивнее, особенно при правильном
подборе качества света (тип ламп). Так, при выращивании сеянцев древесных
культур (дуб, сосна) на непрерывном освещении темпы их роста возрастают в 1,5 -
2 раза (В. М. Леман). Такие однолетние растения, как
горох и фасоль, также характеризуются очень интенсивным ростом в условиях
круглосуточного освещения. Однако есть растения, на рост которых круглосуточное
освещение оказывает отрицательное влияние. В некоторых случаях круглосуточное
освещение вызывает явления, сходные с теми, которые обычно являются следствием
недостатка света. Особенно вредное влияние круглосуточного освещения проявляется
при высокой ночной температуре.
Снабжение кислородом. Процессы роста требуют затрат
энергии, источником которой служит процесс дыхания. В этой связи понятна
необходимость кислорода. При снижении концентрации ниже 5% рост тормозится. Это
происходит не только из-за нарушения энергетического баланса, но и в силу
накопления продуктов анаэробного обмена (спирт, молочная кислота).
Минеральное питание. Для нормального протекания
ростовых процессов необходимо достаточное снабжение всеми необходимыми
минеральными элементами. Особенно специфична роль снабжения растений азотом. Это
связано не только с тем, что азот входит в состав белков и нуклеиновых кислот,
но и с образованием двух основных групп гормонов, регулирующих ростовые
процессы (ауксины, цитокинины).