Молекулярные основы эволюции, дифференцировки развития и старения
Известно,
что некоторые
фрагменты ДНК
могут перемещаться
с одного места
на другое в
пределах одной
хромосомы или
встраиваться
в другую хромосому.
Существование
прыгающих генов
впервые было
показано
Б. Мак-Клинток
при изучении
генетики кукурузы.
Она выяснила,
что элементы
регуляторного
гена перемещаются
в геноме с одного
места на другое
и влияют на
экспрессию
генов, приводя
к появлению
фенотипических
вариаций. Эта
работа не привлекала
внимания в
течение 30 лет,
пока исследования
на E. Coli
не подтвердили
способность
генов к передвижению
и внедрению
в другое место
в геноме, а
Б. Мак-Клинток
была присуждена
Нобелевская
премия.
Способные
к перемещению
последовательности
ДНК получили
название транспозоны
(Tn элементы)
или прыгающие
гены.
Изменение
положения
какого-то сегмента
ДНК относительно
окружающих
его последовательностей
происходят
повсеместно
в разных вариантах,
однако большинство
из них наблюдается
относительно
редко. Многие
геномные перестройки
происходят
в результате
гомологичной
рекомбинации
между аллельными
последовательностями
и не затрагивают
соседних сегментов
ДНК. Вообще же
говоря, структура
генома вполне
стабильна. Это
и не может быть
иначе, поскольку
необходимо
поддерживать
жизнеспособность
особей и видов.
Многие
геномные перестройки
не запрограммированы,
они не связаны
с каким-то
специфическим
влиянием на
экспрессию
генов и в них
есть элемент
случайности.
Случайными
могут быть
частота таких
событий, сами
сегменты ДНК
или то и другое.
Примерами таких
довольно редких
событий служит
транспозиция
последовательностей
ДНК из одного
геномного
локуса в другой
или дупликация
и последующая
амплификация
сегментов ДНК.
Однако сходные
транспозиции
и амплификации
могут быть
сопряжены также
с неслучайными,
запрограммированными
изменениями.
Такие запрограммированные
события играют
ключевую роль
в регуляции
экспрессии
некоторых генов
во время дифференцировки
и развития
определенных
типов клеток.
Типы
мобильных
элементов
Различают
несколько типов
мобильных
ДНК-элементов,
но все они обладают
следующими
общими свойствами:
несут
ген (или несколько
генов), необходимый
для транспозиции
(ген транспозазы
– фермента
участвующего
в перемещении);
во-вторых,
на концах содержат
специфические
взаимно инвертированные
повторяющиеся
последовательности,
также необходимые
для транспозиции
сами
транспозирующиеся
элементы не
кодируют никаких
существенных
для организма
функций, однако
часто содержат
специфические
гены, например
ген устойчивости
к антибиотикам.
Транспозиция
этих элементов,
как правило,
сопровождается
сильными мутагенными
эффектами.
К транспозирующимся
элементам
относят те,
транспозиция
которых протекает
без участия
обратной
транскрипции
Транспозирующиеся
элементы прокариот
Инсерционные
последовательности
(IS – insertion
sequences – последовательности
вставки).
Это сегменты‚
способные как
целое перемещаться
из одного участка
локализации
в другой (рис
73 Коничев).
Ип содержат
лишь те гены‚
которые необходимы
для их собственного
перемещения
– транспозиции.
на обоих
концах каждого
из них всегда
имеются инвертированные
повторы (рис. 10.4),
которые также
необходимы
для транспозиции.
Нуклеотидные
последовательности
этих повторов
различны для
разных IS
и варьируют
по длине от 10
до 40 п. н.
Их перемещения
в новые геномные
локусы часто
приводят к
мутациям,
заключающимся
в прерывании
регуляторных
и кодирующих
участков‚ а
промоторные
элементы внутри
самих IS
могут влиять
на экспрессию
близлежащих
генов.
Частота
транспозиций
у разных элементов
неодинакова
и составляет
10–5–10–7 на поколение.
При
транспозиции
IS в новое
положение
исходный IS
– элемент остается
на прежнем
месте; т.е., инсерция
сопровождается
точным синтезом
второй копии
и зависит от
репликативных
функций хозяина.
2. Транспозоны.
– Т. называют
сегменты ДНК‚
обладающие
теми же свойствами‚
что и ИП‚ но
содержащие
также гены‚
не имеющие
отношение
непосредственно
к транспозиции
а) сложные
транспозоны
– Часто
IS – элементы
длиной примерно
1 т. п. н. входят
в состав более
сложных транспозонов
(прозрачка 1).
– Их длина
может достигать
тысяч пар оснований,
а центральные
области могут
содержать
разнообразные
гены, (например‚
гены устойчивости
к антибиотикам‚
гены токсинов
или гены дополнительных
ферментов
клеточного
метаболизма)
– С обеих
сторон эта
центральная
область фланкируется
одинаковыми
IS элементами
(от IS 1 до IS
10) – IS – L
и IS – R с левой
и правой сторон
соответственно.
– Вся
информация,
необходимая
для перемещения
сложного транспозона,
содержится
в его IS –
части; это та
самая информация,
которая используется
IS элементом
как таковым.
б) Простые
транспозоны.
– это
транспозоны
не связаны ни
с какими IS
– элементами:
информация,
необходимая
для транспозиции,
закодирована
в них самих.
В качестве
примера можно
привести транспозон
ТnЗ (прозрачка
3), который содержит
инвертированные
концевые повторы
длиной 38 п. н., I
R-L и I
R-R, необходимые
для транспозиции.
Между
этими повторами
находятся три
гена: два из
них кодируют
транспозиционные
белки, третий
– β-лактамазу
(ген атр, не связан
с транспозицией).
Кроме
того, имеется
некодирующий
участок длиной
170 п.н., содержащий
промоторы
генов транспозиционных
белков, а также
специфический
участок, называемый
res и тоже
необходимый
для транспозиции.
Сайты-мишени
Это сегменты
ДНК, в которые
встраиваются
Мэ
– Одни
транспозирующиеся
элементы довольно
разборчивы
и охотнее
встраиваются
в короткие
геномные сегменты,
гомологичные
концам самого
мобильного
элемента.
– Другие
менее капризны
и не отдают
явного предпочтения
никаким сайтам-мишеням,
хотя наблюдается
некоторая
тенденция к
встраиванию
их в АТ-богатые
участки.
– Независимо
от типа Мэ. его
встраивание
в новый генетический
локус обычно
сопровождается
дупликацией
короткого
участка ДНК
в в сайте-мишени.
– Эти
дуплицированные
сегменты затем
фланкируют
встроившийся
элемент.
– Почти
обязательная
дупликация
сайтов-мишеней
указывает на
то, что при различиях
в механизме
большинство
вставок происходит
с образованием
в потенциальных
сайтах-мишенях
смещенных
одноцепочечных
разрывов, как
это показано
на рис. 10.1. Сайт-мишень
в геноме, содержащий
мобильный
элемент, условно
называют
«заполненным»,
а не содержащий
такового –
«свободным».
Типы
транспозиций:
Для простоты
рассмотрим
транспозицию
из одного генома
в другой (например,
из плазмиды
в бактериальный
геном или обратно).
Внутримолекулярная
транспозиция
протекает более
сложно.
Коинтеграционная
или репликативная
транспозиция
(поскольку
происходит
полная дупликация
элемента.)
донорный
геном, который
несет транспозирующийся
элемент, сливается
с реципиентной
молекулой ДНК
(прозрачка 4).
Образовавшийся
коинтеграт
содержит всю
донорную и
реципиентную
ДНК, а также
по одной копии
транспозирующегося
элемента в
местах сочленения
этих ДНК.
Коинтеграция
включает разрыв
исходных
фосфодиэфирных
связей и образование
новых, дупликацию
всего элемента
и дупликацию
сайта-мишени.
Коинтеграт
может затем
разрешаться
с образованием
двух исходных
ДНК, каждая из
которых несет
копию транспозированного
элемента. Для
разделения
коинтеграа
необходимодействие
продукта гена
tnp R‚
называемого
резолвазой
(от англ. Resolution
– разрешение)‚
которая разрезает
коинтеграт
на исходные
репликаторы.
При
транспозиции
путем коинтеграции
используется
не только
информация,
закодированная
в самом элементе,
но и репликативные
функции клетки.
Простое
встраивание
или консервативное
(или нерепликативным,
поскольку
дупликации
как таковой
не происходит)
Транспозирующийся
элемент перемещается
в новый геномный
локус, при этом
никаких других
перестроек,
кроме дупликации
сайта-мишени,
не происходит
(прозрачка 4).
Некоторые
транспозирующиеся
элементы, например
ДНК фага Мu,
участвуют как
в коинтеграции,
так и в простом
встраивании.
3. Существует
еще одна весьма
привлекательная
модель, согласно
которой:
на основе
общей промежуточной
структуры
осуществляется
транспозиция
любого из двух
типов (рис. 10.9).
В соответствии
с этой моделью
может происходить
транспозиция
ДНК фага Мu
и других мобильных
элементов.
На рисунке,
иллюстрирующем
модель, и ДНК
донора, и ДНК
реципиента
представлены
в кольцевой
форме, поскольку
в экспериментах
in vitro,
поставленных
для проверки
справедливости
данной модели,
использовалась
кольцевая
плазмидная
ДНК;
мобильным
элементом в
донорной ДНК
служила модифицированная
ДНК фага Мu.
In vivo доноры
и реципиенты,
например плазмиды
и хромосомы
Е.соli, также
находятся в
кольцевой
форме.
Согласно
данной модели,
транспозиция
начинается
с образования
одноцепочечных
разрывов в
кольцевых
донорной и
реципиентной
ДНК.
У донора
такие разрывы
происходят
с обоих З'-концов
элемента, у
реципиента
– в сайте-мишени
с образованием
5'-выступов.
Затем
разрезанные
концы мобильного
элемента соединяются
с концами
реципиента,
и образуется
общий промежуточный
продукт.
Если в
цепях, противоположных
по отношению
к разрезанным
первоначально,
никаких дополнительных
разрывов не
происходит,
то имеет место
простое встраивание.
В результате
заполнения
бреши и лигирования
происходит
дупликация
сайта-мишени.
Таким
способом может
осуществляться
нерепликативная
транспозиция
Тn 10.
Эту реакцию
катализирует
транспозаза
Тn 10, которая
ускоряет разрезание
на концах Тn
10, а также, по-видимому,
воссоединение
разорванных
концов.
При
репликативной
транспозиции
общий промежуточный
продукт претерпевает
совсем другие
превращения.
интермедиат
по своей структуре
аналогичен
кольцевой ДНК
с двумя репликативными
вилками.
Если
репликация
начинается
в каждой вилке,
то конечный
продукт представляет
собой коинтеграт-кольцо,
содержащее
и донорную, и
реципиентную
ДНК, а также
копии транспозирующегося
элемента,
реплицированные
полуконсервативным
путем.
При разрешении
коинтеграта
в конце концов
осуществляется
транспозиция
новой копии
мобильного
элемента в
другой сайт-мишень.
Разрешение
может происходить
путем гомологичной
рекомбинации
между двумя
копиями транспозирующегося
элемента с
участием
рекомбинационного
аппарата Е.соli.
(прозрачка 6).
Генетическая
изменчивость
бактерий при
транспозиции
Мобильные
элементы вызывают
генетическую
нестабильность
поблизости
от участка
своей локализации,
особенно в
процессе
репликативного
механизма
транспозиций.
В зависимости
от того, как
внесены разрывы
в ДНК-мишень,
получится либо
делеция (Выпадение
участка хромосомы
из ее внутренней
области), либо
инверсия
(перестановка)
генетического
материала между
местом расположения
транспозона
и мишенью его
перемещения.
В связи с этим
интересно
отметить, что
хромосомы
родственных
видов бактерий
отличаются
друг от друга
многочисленными
перестройками
именно этого
типа. По-видимому,
мобильные
элементы сыграли
существенную
роль в дивергенции
и видообразовании
бактерий.
1) Встраивание
IS элементов
поблизости
от молчащего
гена может
приводить к
его активации
за счет транскрипции
с промотора
IS – элемента,
т.е. изменяется
регуляция
бактериального
гена (пр. 8а).
2) Очень
важно, что мобильные
элементы служат
подвижными
участками
гомологии,
гомологическая
рекомбинация
между которыми
может приводить
к дупликациям
генов (рис. 76
Коничев). Считается,
что дупликация
– один из основных
путей эволюционного
возникновения
новых функций.
Действительно,
«лишняя» копия
гена выходит
из-под давления
естественного
отбора и получает
возможность
накапливать
изменения. Чаще
всего это приведет
к утрате какой
бы то ни было
функции, но
иногда может
получиться
ген с новыми
функциями.
3) Нельзя
забывать и тот
факт, что клетка
может получить
селективное
преимущество
за счет приобретения
в составе транспозона
гена, который
сам по себе
способен оказаться
выгодным для
бактерии в
определенных
условиях.
Действительно,
на транспозонах
«путешествуют»
гены устойчивости
к различным
бактериальным
ядам, в том числе
к тяжелым металлам
и антибиотикам,
гены дополнительных
метаболических
путей, позволяющие
использовать
необычный
источник углерода,
наконец, гены
некоторых
токсинов, делающие
бактерии патогенными
и позволяющие
им тем самым
существенно
изменить образ
жизни. Сказанное
в равной степени
относится и
к плазмидам,
поскольку
большинство
полезных для
клетки-хозяина
плазмидных
генов находится
в составе
транспозонов.
4) Если
элемент встроен
в один из генов
полицистронного
оперона, то
встроенный
элемент может
влиять на экспрессию
последующих
генов, либо
останавливая
транскрипцию
в сайте терминации,
находящемся
внутри его
самого (прозрачка
8,6), либо подавляя
трансляцию
дистальных
кодирующих
последовательностей
мРНК (прозрачка
8, в)
5) Когда
один и тот же
геном служит
и донором, и
реципиентом
при образовании
коинтеграта
(внутримолекулярная
транспозиция),
способность
мобильных
элементов
вызывать перестройки
в соседних
участках ДНК
приводит к
другим мутационным
эффектам. Все
происходящие
при этом события,
включая делеции
и инверсии,
связаны с соединением
одного или двух
концов мобильного
элемента с
новыми последовательностями
ДНК (рис. 10.13); детали
этих процессов
до конца не
установлены.
Подвижные
генетические
элементы у
эукариот
Существенную
часть генома
эукариот (10–30%)
составляют
повторяющиеся
последовательности,
имеющие определенную
структурную
организацию
и способные
перемещаться
в геноме как
в пределах
одной хромосомы,
так и между
хромосомами.
Они получили
название подвижных
генетических
элементов.
Элементы включают
в свой состав
от тысячи до
десятков тысяч
нуклеотидных
пар.
Наибольшее
количество
подвижных
элементов
обнаружено
в геноме растений
(до 50%). Подвижные
генетические
элементы обычно
рассеяны по
геному, но могут
концентрироваться
в отдельных
участках хромосом.
Виды
мобильных
элементов
эукариот
Различают
два (по признаку
молекулярных
механизмов
перемещения)
основных класса
подвижных
генетических
элементов:
1. Транспозоны
Эти элементы
ограничены
инвертированными
повторами
(последовательностями,
направленными
навстречу друг
другу), как и
некоторые
транспозоны
прокариот.
Примерами их
могут служить
Р-элемент дрозофилы
и Ас-элемент
кукурузы (рис. 92
Коничев). В геномах
этих организмов
насчитывается
по 30 – 50 копий
таких элементов.
Полные копии
(часть копий
дефектна, так
как имеет внутренние
делеции) содержат
открытые рамки
считывания,
кодирующие
транспозазу
(рис. 92).
Схема
перемещения
транспозона
показана на
рис. 93. Инвертированные
повторы необходимы
для перемещения
элемента, которое
осуществляется
благодаря их
сближению друг
с другом и узнаванию
транспозазами.
Инвертированные
повторы сближаются
и точно отрезаются
от соседних
участков ДНК
хозяина. Вырезанный
транспозон
внедряется
в район вносимого
транспозазой
разрыва в
молекуле-мишени
и сшивается
с ДНК хозяина
в новом месте.
Разрыв и зашивание
осуществляются
транспозазой
и вспомогательными
белками. Транспозаза
может кодироваться
как самим подвижным
элементом,
который будет
перемещаться,
так и другой
копией элемента,
локализованной
в том же геноме
в отдалении.
Р-элементы
обычно встраиваются
в определенные
сайты, консенсусная
последовательность
которых такова:
5'-ГГЦЦАГАГ При
встраивании
Р-элемента этот
сайт-мишень
дуплицируется.

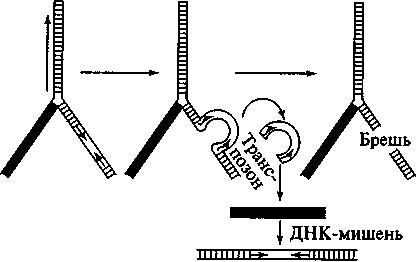
Встраивание
Рис. 93
Перемещение
транспозона
Брешь
в ДНК после
вырезания
транспозона
может заделываться
либо с помощью
гомологичного
отрезка ДНК,
либо за счет
сшивания разорванных
концов (это
особенно характерно
для транспозонов
растений).
Эффект
от перемещения
транспозона
эукариот
Р-элемент
содержится
только в некоторых
линиях D.
melanogaster.
Скрещивание
самок без Р-элемента
с самцами, несущими
Р-элементы,
приводит у
гибридов к
транспозициям
Р-элемента,
которые наблюдаются
только в клетках
зародышевого
пути. Совокупность
наблюдаемых
эффектов называют
гибридным
дисгенезом.
Дисгенные
мухи часто
оказываются
стерильными,
а если они фертильны,
то у их потомков
обнаруживается
множество
мутаций и эти
признаки передаются
следующим
поколениям.
Поэтому линии
с Р-элементом
и без него выглядят
как репродуктивно
изолированные,
по крайней мере
частично.
Биологическая
изоляция играет
огромную роль
в процессе
эволюции. Изоляция
линий вызвана
активацией
транспозиций
Р-элемента,
присутствующего
в одной из них.
Причина, почему
транспозиции
Р-элемента
ограничены
зародышевыми
клетками, объясняется
тем, что только
в клетках-предшественниках
гамет осуществляется
такой тип сплайсинга,
который приводит
к образованию
непрерывной
открытой рамки
трансляции,
кодирующей
транспозазу
(см. рис. 92).
Ограничение
транспозиции
зародышевыми
клетками имеет
определенный
смысл, поскольку
обеспечивает
выживание
особей, несущих
гаметы, в которых
произошли
геномные перестройки
вследствие
транспозиции
Р-элемента.
Подобный «геномный
шок», сопровождающийся
высокой частотой
мутагенеза,
может обеспечить
большую степень
геномной
изменчивости,
которая послужит
материалом
для отбора в
процессе эволюции.
Перемещения
Ас-элемента
происходят
и в соматических
клетках кукурузы.
За ними можно
следить по
распределению
мутантных
участков ткани,
например лишенных
пигмента вследствие
инактивации
гена, определяющего
пигментацию.
Потомство
клетки, содержащей
только инактивированные
гены, также
будет лишено
пигмента. Вырезание
мобильного
элемента приводит
к реактивации
гена. В связи
с этим был сделан
вывод о регуляторной
функции перемещающихся
элементов и
они были названы
контролирующими
(Б. Мак-Клинток).
В
целом подвижные
элементы эукариот
представляют
собой чрезвычайно
разнородную
популяцию.
Существует
мнение о том,
что они не влияют
на фенотип
организма и
размножаются
в геноме лишь
благодаря
особенностям
своей структуры,
в результате
чего постепенно
заселяют геном.
Предполагается,
что они составляют
часть так называемой
эгоистичной
(см. с. 156)
ДНК, размножение
которой ограничивается
естественным
отбором. Отбор
устраняет те
случаи, когда
внедрение
эгоистичной
ДНК сопровождается
вредными
последствиями.
На определенных
стаднях эволюции
эти элементы
могут использоваться
для целей, которые
в дальнейшем
обеспечат
селективные
преимущества
организму.
2.
Ретротранспозоны.
А)
с длинными
концевыми
повторами
– Сходны
по своей структуре
с проретровирусами,
которые внедряются
в геном, используя
механизмы
обратной
транскрипции.
– Эти
элементы содержат
«тело» размером
5–8 тыс. н.п., ограниченное
прямыми длинными
концевыми
повторами (ДКП,
или LTR
– от англ. 1опg
terminal
repeats),
обычно
содержащими
по 300–400 н. п. (рис. 94).
– Число
копий этих
элементов,
принадлежащих
к одному семейству,
достаточно
постоянно для
вида, но варьирует
от нескольких
копий до сотен
тысяч копий
в зависимости
от типа ретротранспозона.
– В
составе «тела»
элемента
обнаруживаются
открытые рамки
считывания
для обратной
транскриптазы
и нуклеазы
(интегразы).
– Разные
копии одного
семейства могут
иметь вставки
и делеции как
в ДКП, так и в
самом элементе.
– По
флангам ДКП
находятся
короткие прямые
повторы.
Способ
перемещения
ретротранспозонов
с длинными
концевыми
повторами
– предполагает
его транскрипцию
с помощью
РНК-полимеразы
П.
– В
составе ДКП
имеются сайты
инициации
транскрипции
и сигналы
полиаденилирования.
ДКП могут служить
активными
промоторами.
– Транскрипция
начинается
в одном ДКП
(условно левом,
5'-ДКП), а заканчивается
в другом (правом,
3'-ДКП).
– Синтезируемая
молекула РНК
транслируется
с образованием
ферментов,
необходимых
для синтеза
ДНК и внедрения
ее в геном.
Эта
схема полностью
повторяет схему
образования
и интеграции
провируса
(рис. 95).
Остается открытым
вопрос: произошли
ли ретровирусы
из ретротранспозонов
или, напротив,
ретротранспозоны
возникли из
вирусов в результате
потери способности
к заражению?
Подавляющая
часть ретротранспозонов
(при сравнении
их с ретровирусами)
либо потеряла
ген оболочки
вируса, либо
еще не приобрела
его. Таким образом,
ретротранспозоны
представляют
собой внутригеномные,
неинфекционные
элементы, способные
лишь к самовоспроизведению
и «подзаражению»
того же генома.
Внедрение
ретротранспозона
внутрь гена
или около гена
вызывает разные
эффекты:
– происходит
инактивация
гена (рис. 96
а).
– при
интеграции
в район промотора
на 5'-фланге гена
ретротранспозон
может резко
активировать
экспрессию
гена, обеспечивая
транскрипцию
с собственного
промотора
(рис. 96
б1). Если в этом
случае подвижный
элемент оказался
около протоонкогена,
то результатом
может быть
сверхпродукция
белка и злокачественное
перерождение
клетки.
– это
же возможно
при воздействии
энхансеров,
привносимых
мобильным
элементом
(рис. 96
б2).
– Особые
возможности
для перенесения
и приобретения
регуляторных
сигналов возникают
тогда, когда
элемент (ретротранспозон)
удаляется за
счет гомологичной
комбинации
между ДКП с
идентичными
повторами (см.
(рис. 96
в). В результате
сохраняется
лишь один ДКП
на месте внедрения
ретротранспозона.
Представленные
случаи внедрения
элемента
сопровождаются
мутациями с
самыми разными
фенотипическими
проявлениями
обусловленными
подавлением
образования
или, наоборот,
гиперпродукцией
белка.
Перемещение
элементов по
геному способствует
– распространению
регуляторных
сигналов (сайтов
инициации
транскрипции,
сигналов
полиаденилирования,
или энхансеров),
что делает
значительной
роль мобильных
элементов в
эволюции систем
регуляции.
Б)
Ретротранспозоны,
которые не
несут длинных
концевых повторов.
Механизм внедрения
их
– несколько
иной,
– но
также осуществляется
с помощью обратной
транскрипции.
– К
ним относятся
представители
семейства L1
населяющие
геном человека.
– Репликация
элемента без
ДКП сопряжена
непосредственно
с районом будущего
внедрения
ретротранспозона
(рис. 97).
– РНК,
образовавшаяся
при транскрипции
элемента,
перемещается
к достаточно
случайному
месту разрыва
ДНК-мишени,
– часто
сшивается с
одной из нитей
ДНК.
– Сюда
устремляются
и необходимые
для интеграции
белки – обратная
транскриптаза
и интеграза.
– Другая,
комплементарная
нить ДНК служит
затравкой для
копирования
РНК-копии элемента
с участием
обратной
транскриптазы.
– Фермент
копирует сначала
небольшой
участок ДНК-мишени,
а затем меняет
матрицу и копирует
РНК,
– после
чего РНК удаляется
и образуется
вторая комплементарная
нить ДНК.
Ретротранспозоны
без ДКП участвуют
в сохранении
концов хромосом
в ряду поколений.
Подобную
функцию у многих
организмов
выполняет
теломераза.
У ряда насекомых,
в том числе и
у дрозофилы,
отсутствует
теломеразная
активность.
Концы ДНК у них
удлиняются
за счет перемещений
ретротранспозонов
и в этом случае
выступают как
компоненты
генома, спасающие
хромосому от
укорачивания.
Ретротранспозоны
перемещаются,
образуя повторяющуюся
структуру, в
которой элементы
соединены друг
с другом по
типу «голова
к хвосту». Сначала
на РНК-транскрипте
как на матрице
с помощью обратной
транскрипции
строится
комплементарная
нить ДНК, а затем
после удаления
РНК-матрицы
достраивается
другая. Таким
образом, геном
хозяина приспособил
эти элементы
для сохранения
концевых участков
хромосом, для
спасения хромосомы
от потери генов.
3.
Ретропозоны
В
геноме млекопитающих,
птиц, амфибий
и насекомых
обнаруживаются
ретропозоны
– внедрившиеся
в геном ДНК-копии,
синтезированные
на разных типах
клеточных РНК,
как на матрицах.
Структура
ретропозонов
позволяет с
уверенностью
говорить об
участии обратной
транскрипции
в процессе их
образования.
Таким образом,
наряду с переносом
информации
от ДНК к РНК
осуществляется
и обратный
процесс – возвращение
ее в геном в
виде ретропозонов.
У млекопитающих
ретропозоны
составляют
более 10% ДНК,
следовательно,
мощность встречного
потока информации
от РНК к ДНК
может быть
существенной,
по крайней мере
при оценке его
во временном
эволюционном
масштабе.
Различают
разные типы
ретропозонов,
среди них:
А)
Псевдогены.
Это копии генов,
лишенные интронов
и обладающие
последовательностью
поли-дА-поли-дТ
на 3'-фланге.
Матрицей при
их копировании
послужила
«процессированная»
полиаденилированная
мРНК. Рамки
трансляции
таких «генов»
часто «испорчены»
стоп-кодонами
и делециями.
Они, как и проретровирусы,
ограничены
короткими
прямыми повторами,
представляющими
собой дупликацию
геномной
последовательности
в области сайта
внедрения
ДНК-копии,
образованной
на мРНК.
Б)
Ретрогены. Если
ретропозон
правильно
процессирован,
ДНК-копия содержит
открытую рамку
считывания
и возможна его
экспрессия,
то такие ретропозоны
называют ретрогенами.
Большую
массу повторяющихся
элементов
позвоночных
представляют
ДНК-копии клеточных
РНК, кодирующих
белки неизвестной
природы, а также
аномально
процессированных
клеточных
транскриптов
тРНК, 7S
РНК и UPНК.
Они представляют
собой длинные
и короткие
повторы. Семейства
повторяющихся
последовательностей
часто получают
свои названия
по расщепляющим
их рестриктазам
(например, короткие
Alu повторы
или длинные
Крn повторы
в геноме человека).
Каждый
вид млекопитающих
обладает собственным
главным семейством
длинных повторов,
отличающих
его от других
видов. Предполагают,
что разные
семейства
длинных повторов
возникли сравнительно
недавно, уже
после эволюционной
дивергенции
видов млекопитающих.
Короткие
повторы, например
повторы Alu
– семейства
у человека,
составляют
3 • 105, что соответствует
5 – 6% массы ДНК
клетки. Их
нуклеотидная
последовательность
гомологична
последовательности
отдельных
участков 7S
РНК. Такие повторы
рассеяны по
геному, их
обнаруживают
в интронах, на
5'-флангах генов,
в составе
З'-нетранслируемых
участков гена.
Между особями
одного вида
иногда наблюдаются
различия
(полиморфизм)
в структуре
генов и их ближайшего
окружения,
обусловленные
внедрением
коротких или
длинных повторов.
В геноме существует
множество
неактивных
дефектных копий
этих элементов,
они не способны
кодировать
транспозазу
или обратную
транскриптазу.
С другой стороны,
они сохраняют
способность
к перемещениям,
если в случае
транспозонов
имеются инвертированные
повторы, узнаваемые
транспозазой,
а в случае
ретротранспозонов
сохранены
промотор и
возможность
транскрипции
элемента. Множество
таких дефектных
копий начнут
перемещаться,
если ферменты,
ответственные
за перемещение,
будут кодироваться
другими полноценными
элементами.
В геноме человека
источником
активной обратной
транскриптазы
является
ретротранспозон
L1, число
копий которого
достигает 100
тыс. Из них активно
перемещаются
30 – 60 тыс., остальные,
поврежденные,
перемещаться
не могут.
Мобильные
генетические
элементы могут
принимать
участие в
перестройках
хромосом. Наличие
в хромосомах
нескольких
одинаковых
по нуклеотидной
последовательности
копий подвижного
элемента позволяет
в редких случаях
осуществить
рекомбинацию
по районам их
локализации.
В результате
неравного
кроссинговера
возникают
делеции отдельных
участков или,
наоборот, дупликации
(рис. 98).
Неравный
кроссинговер
по районам
локализации
транспозона
mariner
(моряк) у человека
приводит к
делеции участка
в коротком
плече 17 й
хромосомы. Если
это событие
происходит
в зародышевой
клетке при
созревании
гамет, то хромосома
с делецией
будет передана
потомкам. Это
приводит к
наследственным
заболеваниям
нервной системы
– невропатиям
и параличам.
Внутрихромосомная
рекомбинация
между двумя
элементами
приводит к
инверсии –
повороту участка
хромосомы на
180°. Инверсия
может быть
вредна для
организма, а
может способствовать
эволюции генома,
поскольку
помогает передать
потомству
случайно сложившееся
благоприятное
сочетание
генов, препятствуя
кроссинговеру.
Ретротранспозоны
с ДКП участвуют
в ликвидации
двунитевых
разрывов ДНК.
Обычно такой
разрыв залечивается
с помощью
гомологичной
молекулы ДНК,
например сестринской,
только что
регошцированной
нити. Участие
реплицирующейся
ДНК ретро-транспозонов
с ДКП в процессе
заживления
двунитевых
разрывов обнаружено
у дрожжей (рис. 99).
Брешь в двунитевой
спирали, заделанная
ретротранспозоном,
сохраняет
целостность
хромосомы, но
изменяет ее
нуклеотидную
последовательность.
Если район
хромосомы, где
была брешь, не
содержит
существенного
гена, то клетка
остается
жизнеспособной.
Подвижные
элементы не
стоит рассматривать
только как
«эгоистическую»
ДНК, паразитирующую
на ДНК клетки-хозяина.
Они приспособлены
и для нужд генома
клетки: способствуют
сохранению
структурной
целостности
хромосомной
ДНК; внедряясь
в гены, не только
инактивируют,
вызывая мутации,
но и меняют
характер их
экспрессии.
В связи с этим
подвижные
генетические
элементы, будучи
важными факторами
изменчивости
генов и участвуя
в перестройках
структуры
хромосом, имеют
огромное значение
в процессах
эволюции геномов.