Механизмы реализации путей гибели клетки
с
другими группами
крови.
Соответственно
лица с AB (FV) группой
крови являются
универсальными
реципиентами,
т.е. им можно
перелить любую
группу крови.
На поверхности
эритроцитов
имеется также
резус-фактор
(Rh-фактор) - агглютиноген.
Он присутствует
у 86 % людей; у 14 %
отсутствует
(резус-отрицательные).
Переливание
резус-положительной
крови резус-отрицательному
пациенту вызывает
образование
резус-антител
и гемолиз
эритроцитов.
Агглютинация
эритроцитов
свойственна
нормальной
свежей крови,
при этом образуются
так называемые
"монетные
столбики".
Это явление
связано с потерей
заряда плаэмолеммы
эритроцитов.
Скорость оседания
(агглютинации)
эритроцитов
(СОЭ) в 1 ч у здорового
человека составляет
4-8 мм у мужчин
и 7-10 мм у женщин.
СОЭ может значительно
изменяться
при заболеваниях,
например при
воспалительных
процессах, и
поэтому служит
важным диагностическим
признаком. В
движущейся
крови эритроциты
отталкиваются
из-за наличия
на их плазмолемме
одноименных
отрицательных
зарядов. Поверхность
плазмолеммы
одного эритроцита
составляет
около 130 мкм2
крови. При
окрашивании
мазка крови
азур И-эозином
по Романовскому
- Гимзе большинство
эритроцитов
приобретают
оранжево-розовый
цвет (оксифильны),
что обусловлено
высоким содержанием
в них гемоглобина.
В небольшой
части эритроцитов
A-5 %), являющихся
более молодыми
формами, сохраняются
остатки органелл
(рибосомы,
гранулярный,
эндоплазматический
ретикулум),
которые проявляют
базофилию.
Такие эритроциты
окрашиваются
как кислыми
красителями
(эозин), так и
основными (азур
II) и называются
полихроматофилъными.
При специальной
суправитальной
окраске
(бриллиант-крезилфиолетовым)
в них выявляются
зернисто-нитчатые
структуры,
поэтому их
называют
ретикулоцитами.
Эритроциты
различаются
по степени
насыщенности
гемоглобином.
Среди них
выделяются
нормохромные,
гипохромные
и гиперхромные,
соотношение
между которыми
значительно
изменяется
при заболеваниях.
Количество
гемоглобина
в одном эритроците
называют цветным
показателем.
Электронно-микроскопически
гемоглобин
выявляется
в гиалоплазме
эритроцита
в виде многочисленных
плотных гранул
диаметром 4-5
нм.
Гемоглобин
- это сложный
белок F8 КД, состоящий
из 4 полипептидных
цепей глобина
и гема (железосодержащий
порфирин), обладающий
высокой способностью
связывать
кислород. В
норме у человека
содержится
два типа гемоглобина
- НЬА и HbF. Эти
гемоглобины
различаются
составом аминокислот
в глобиновой
(белковой) части.
У взрослых
людей в эритроцитах
преобладает
НЬА, (от англ.
adult - взрослый),
составляя 98 %.
Он содержит
две а-глобиновые
цепи и две
C-глобиновые
цепи, включающие
574 аминокислоты.
HbF, или фетальный
гемоглобин
(от англ. foetus - плод),
составляет
у взрослых
около 2 % и преобладает
у плодов. К моменту
рождения ребенка
HbF составляет
около 80 %, а НЬА
только 20 %. Эти
гемоглобины
отличаются
составом аминокислот
в глобиновой
(белковой) части.
Железо (Fe2+)
в геме может
присоединять
О2 в легких
(в таких случаях
образуется
оксигемоглобин
- НЬО2) и отдавать
его в тканях
путем диссоциации
НЬО, на кислород
(О2) и НЬ; валентность
Fe2+ не изменяется.
При ряде
заболеваний
(гемоглобинозы,
гемоглобинопатии)
в эритроцитах
появляются
другие виды
гемоглобинов,
которые характеризуются
изменением
аминокислотного
состава в белковой
части гемоглобина.
В настоящее
время выявлено
более 150 видов
аномальных
гемоглобинов.
Например, при
серповидно-клеточной
анемии имеет
место генетически
обусловленное
повреждение
в C-цепи гемоглобина
- глютаминовая
кислота, занимающая
6-е положение
в полипептидной
цепи, заменена
на аминокислоту
валин. Такой
гемоглобин
обозначается
как HbS (от англ.
sickle - серп), так как
в условиях
понижения
парциального
давления О2
он превращается
в тектоидное
тело, придавая
эритроциту
форму серпа.
В ряде стран
тропического
пояса определенный
контингент
людей являются
гетерозиготными
для серповидных
генов, а дети
двух гетерозиготных
родителей по
законам наследственности
дают либо нормальный
тип B (5%), либо бывают
гетерозиготными
носителями,
и 25 % страдают
серповидноклеточной
анемией.
Гемоглобин
способен связывать
О2 в легких,
при этом образуется
оксигемоглобин,
который транспортируется
ко всем органам
и тканям и там
отдает О2. В
тканях выделяемая
СО поступает
в эритроциты
и соединяется
с НЬ, образуя
карбоксигемоглобин.
При разрушении
эритроцитов
(старых или при
воздействии
различных
факторов - токсины,
радиация и др.)
гемоглобин
выходит из
клеток, и это
явление называется
гемолизом.
Старые эритроциты
разрушаются
макрофагами
главным образом
в селезенке,
а также в печени
и костном мозге,
при этом НЬ
распадается,
а высвобождающееся
из железосодержащего
гема железо
используется
для образования
новых эритроцитов.
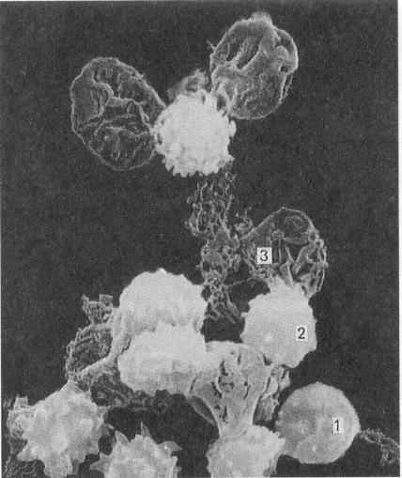
Рис 15. Электронная
микрофотография
гемолиза эритроцитов
и образование
их “теней”
В макрофагах
НЬ распадается
на пигмент
билирубин и
гемосидерин
- аморфные агрегаты,
содержащие
железо. Железо
гемосидерина
связывается
с трансферрином
- негеминовым
белком плазмы,
содержащим
железо, и захватывается
специальными
макрофагами
костного мозга.
В процессе
образования
эритроцитов
(эритропоэз)
эти макрофаги
передают трансферрин
в формирующиеся
эритроциты,
что послужило
основанием
назвать их
клетками-кормилками.
В цитоплазме
эритроцитов
содержатся
ферменты анаэробного
гликолиза, с
помощью которых
синтезируются
АТФ и НАДН,
обеспечивающие
энергией главные
процессы, связанные
с переносом
О2 и СО2, а также
поддержание
осмотического
давления и
перенос ионов
через плазмолемму
эритроцита.
Энергия гликолиза
обеспечивает
активный транспорт
катионов через
плазмолемму,
поддержание
оптимального
соотношения
концентрации
К+ и Na+ в эритроцитах
и плазме крови,
сохранении
формы и целостности
мембраны эритроцита.
НАДН участвует
в метаболизме
НЬ, предотвращая
окисление его
в метгемоглобин.
Эритроциты
участвуют в
транспорте
аминокислот
и полипептидов,
регулируют
их концентрацию
в плазме крови,
т.е. выполняют
роль буферной
системы. Постоянство
концентрации
аминокислот
и полипептидов
в плазме крови
поддерживается
с помощью
эритроцитов,
которые адсорбируют
их избыток из
плазмы, а затем
отдают различным
тканям и органам.
Таким образом,
эритроциты
являются подвижным
депо аминокислот
и полипептидов.
Сорбционная
способность
эритроцитов
связана с состоянием
газового режима
(парциальное
давление О2
и СО2 - Ро, Рсо):
в частности,
при действии
О2 наблюдаются
выход аминокислот
из эритроцитов
и увеличение
их содержания
в плазме.
2.3.2 Тромбоцитыкровяные
пластинки
Мелкие
дисковидные
двояковыпуклые
безъядерные
постклеточные
структуры
диаметром 2-4
мкм, циркулирующие
в крови. Они
образуются
в красном костном
мозге в результате
фрагментации
участков цитоплазмы
мегакариоцитов
(гигантских
клеток костного
мозга), поступают
в кровь, в которой
находятся в
течение 5-10 дней,
после чего
фагоцитируются
макрофагами,
преимущественно
в селезенке
и легком. Часть
тромбоцитов
разрушается
за пределами
сосудистого
русла, куда они
попадают при
повреждении
стенки сосудов.
Общее количество
тромбоцитов
в крови взрослого
человека 0,8-2,4
*1012; из этого
числа около
15 % обновляются
ежедневно. В
норме в крови
циркулирует
2/3 общего числа
тромбоцитов,
а 1/3 находится
вне циркуляции
в красной пульпе
селезенки. На
мазках крови
тромбоциты
вследствие
агрегации
обычно выявляются
в виде скоплений.
Концентрация
тромбоцитов
в крови равна
200-400 тыс. /мкл (200-400 *109
/л) крови.
Функции
тромбоцитов
осуществляются
как внутри
сосудистого
русла, так и
вне его. К ним
относятся:
1) Остановка
кровотечения
при повреждении
стенки сосудов
(первичный
гомеостаз) -
основная функция
тромбоцитов;
2) Обеспечение
свертывания
крови - (гемокоакуляция)
- вторичный
гомеостаз
(совместно с
эндотелием
кровеносных
сосудов и плазмой
крови);
3) Участие
в реакциях
заживления
ран (в первую
очередь, повреждений
сосудистой
стенки) и воспаления;
4) Обеспечение
нормальной
функции сосудов,
в частности,
их эндотелиальной
выстилки
(ангиотрофическая
функция).
Строение
тромбоцитов.
Тромбоцит
окружен плазмолеммой
и включает
светлую
прозрачную
наружную часть,
называемую
гиаломером,
центральную
окрашенную
часть, содержащую
аэурофильные
гранулы - грануломер.
В некоторых
случаях выявляются
небольшие
псевдоподии,
выступающие
из периферической
части гиаломера.
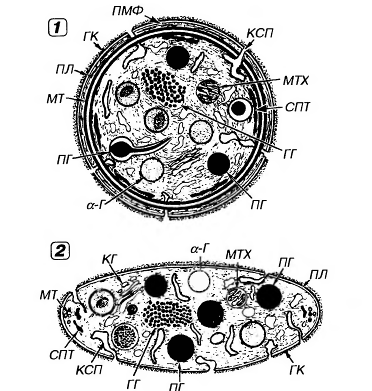
Рис.16. Ультраструктура
тромбоцита.1
- сечение в
экваториальной
плоскости, 2 -
поперечный
разрез, ПЛ -
плазмолемма,
ГК - гликокалис,
КСП - канальцы,
связанные с
поверхностью,
СПТ - система
плотных трубочек,
МТ - микротрубочки,
ПМФ - подмембранные
микрофиламенты,
ГГ - гликоген,
КГ - комплекс
Гольджи, МТХ
- митохондрия,
α-Г -
α-гранулы,
ПГ - плотные
гранулы.
Плазмолемма
тромбоцитов
покрыта снаружи
толстым (от 50
до 150-200 нм) слоем
гликокалиса
с высоким
содержанием
гликозаминогликанов
и гликопротеинов
(см. рисунок
17).
Она
содержит
многочисленные
рецепторы,
посредующие
действие веществ,
активирующих
и ингибирующих
функции тромбоцитов,
обуславливающие
их прикрепление
(адгезию) к
эндотелию
сосудов и агрегацию
(склеивание
друг с другом).
Наиболее
важными из них
в функциональном
отношении
являются рецепторные
гликопротеины
Ib (GP Ib), IIb (GP IIb), IIIa (GP IIIa), рецепторы
к АДФ, адреналину,
тромбину, фактору
Ха, фактору
агрегации
тромбоцитов
(ФАТ), коллагену.
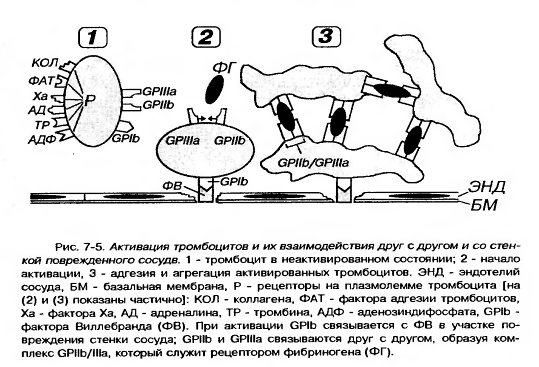
Гиаломер
содержит две
системы трубочек
(канальцев) и
большую часть
элементов
цитоскилета
(см. рисунок
17):
1. Система
канальцев,
связанных с
поверхностью
(открытая система
канальцев),
представлена
гладкими
анастомозирующими
трубочками,
которые открываются
в инвагинации,
образованные
плазмалеммой.
Функция этой
системы канальцев,
скорее всего,
связана с процессами
поглощения
и выведения
веществ, в частности,
она облегчает
экзоцитоз
содержимого
гранул тромбоцитов.
2. Система
плотных трубочек
образуется
комплексом
Гольджи мегакариоцитов.
Она представлена
узкими мембранными
трубочками
заполненными
плотным зернистым
содержимым,
которые располагаются
непосредственно
под кольцом
микротрубочек.
Или разбросаны
по цитоплазме.
Предполагают,
что они накапливают
и выделяют
Са2+, т.е. являются
аналогом
саркоплазматической
сети мышечных
клеток. Их связывают
так же с выработкой
простагландинов.
Цитоскелет
тромбоцитов
представлен
микротрубочками,
микрофиламентами
и промежуточными
филаментами.
Микротрубочки
в количестве
4-15 располагаются
по перефирии
цитоплазмы
и формируют
мощный пучек
(краевое кольцо),
служащий жестким
каркасом и
способствующий
поддержанию
формы тромбоцитов.
Микрофиламенты,
образованные
актином, многочисленны
(актин составляет
25% белка тромбоцитов),
располагаются
по всей цитоплазме
в виде коротких
нитей; в гиаломере
они концентрируются
между пучком
микротрубочек
и плазмалеммой
и образует
подмембранный
аппарат. Последний
участвует в
образовании
выпячиваний
цитоплазмы
при движении
и агрегации
тромбоцитов.
Актиновые
филаменты
связаны в единую
систему посредством
белков α
- актина, миозина
и тропомиозина,
а с плазмалеммой
- с помощью белка
филамина.
Промежуточные
филаменты
образованы
белком виментином
и располагаются
примущественно
под плазмалеммой.
Грануломер
содержит митохондрии,
частицы гликогена,
отдельные
рибосомы, единичные
короткие цистерны
грЭПС, элементы
комплекса
Гольджи и гранулы
нескольких
типов.
α
- гранулы - самые
крупные (диаметр
300-500 нм), с умеренно
плотным матриксом,
в котором
содержаться:
фибриноген,
фибронектин,
тромбоспондин
(белок сходный
с актомиозином),
тромбоглобулин,
тромбоцитарный
фактор роста
(ТРФР), ЭФР, ТФРβ,
фактор свертывания
V и фактор Виллебанда
(белок переносчик
фактора VIII свертывания),
а так же ряд
других белков.
Составляют
большую часть
гранул, окрашивающихся
азуром.
δ
- гранулы (плотные
гранулы или
тельца) - немногочисленные
(до 5) мембранные
пузырьки диаметром
250-300 нм с плотным
матриксом,
который иногда
располагается
в них эксцентрично.
Матрикс содержит
АДФ, АТФ, Са2+,
Mg2+, пирофосфат,
гистамин, серотонин.
Последний не
синтезируется
тромбоцитами,
а поглощается
ими из крови.
γ
- гранулы мелкие
(диаметр 200-250 нм)
пузырьки, содержащие
гидролитические
ферменты.
Рассматриваются
как лизосомы.
3. Иммунитет
Под иммунитетом
понимается
невосприимчивость,
слабовосприимчивость,
сопротивляемость
организма
инфекциям
и инвазиям
чужеродных
организмов
(в том числе -
болезнетворных
микроорганизмов)
и относительная
устойчивость
к вредным веществам.
В более широком
смысле это -
способность
организма
противостоять
изменению его
нормального
функционирования
под воздействием
внешних факторов.
Иммунитет - это
внутренняя
система организма,
которая обеспечивает
его защиту от
вредных воздействий
внешней среды.
Эта система
защищает как
основные как
внешние его
параметры, так
и функции
жизнедеятельности
его органов
и тканей.
У большинства
живых организмов
существуют
механизмы
неспецифической
резистентности,
которую также
называют врождённым,
конституциональным
или видовым
иммунитетом.
У челюстноротых
позвоночных
появляется
качественно
новая система
приобретённого
иммунитета,
позволяющая
реагировать
на конкретный
чужеродный
биоматериал
и запоминать
его, существенно
усиливая
интенсивность
ответной реакции
на вторжение
при повторном
контакте с этим
же материалом.
Различают два
типа иммунитета:
специфический
и неспецифический.
Специфический
иммунитет носит
индивидуальный
характер и
формируется
на протяжении
всей жизни
человека в
результате
контакта его
иммунной системы
с различными
микробами
и антигенами.
Специфический
иммунитет
сохраняет
память о перенесенной
инфекции и
препятствует
ее повторному
возникновению.
Неспецифический
иммунитет носит
видоспецифический
характер, то
есть практически
одинаков у всех
представителей
одного вида.
Неспецифический
иммунитет
обеспечивает
борьбу с инфекцией
на ранних этапах
ее развития,
когда специфический
иммунитет еще
не сформировался.
Состояние
неспецифического
иммунитета
определяет
предрасположенность
человека к
различным
банальным
инфекциям,
возбудителями
которых являются
условно патогенные
микробы.
Невосприимчивость
организма к
инфекционным
заболеваниям
бывает специфической
и неспецифической
(неспецифическая
резистентность).
Специфическая
невосприимчивость
= иммунитет.
Неспецифическая
резистентность
бывает обусловлена:
Видовой
невосприимчивостью
(например,
невосприимчивость
человека к
болезням животных);
Бактерицидными
факторами
организма
(фагоцитоз и
воспаление,
лизоцим, показатель
pH, лихорадочные
реакции)
Стерильный
иммунитет
(выздоровление
сопровождается
полным устранением
микробов)
Нестерильный
иммунитет
(формируется
т. н. носительство).
Иммуноглобулины
типа M (IgM) (выделяются
в первое время
после контакта
с инфекцией,
обусловливают
первичный
иммунный ответ).
Антитела
типа G (IgG) (защищает
организм на
протяжении
длительного
времени, участвует
во вторичном
иммунном ответе,
при первичном
заражении, пик
их концентрации
приходится
на 2 неделю
болезни).
Иммуноглобулины
типа Е (IgE) обусловливают
аллергические
реакции -
гиперчувствительность
немедленного
типа (ГНТ).
Иммуноглобулин
типа А (защищает
организм от
проникновения
инфекции через
кожу и слизистые
оболочки, в
большом количестве
содержится
в грудном молоке).
На первом
этапе столкновения
с чужеродным
антигеном
запускается
неспецифический
патологический
защитный процесс
- воспаление,
сопровождающийся
фагоцитозом,
выделением
медиаторов
воспаления
- гистамина,
серотонина,
цитокинов и
т.п. Фагоциты
(макрофаги)
поглощают
антигены и
контактируют
с лимфоцитами
Т-хелперами,
представляя
им на поверхности
антигенные
детерминанты.
Т-хелперы запускают
размножение
(выделяя специфические
белковые вещества
- интерлейкины)
специфических
для данного
антигена клонов
Т-киллеров и
В-лимфоцитов
из предсуществующих
стволовых
клеток, которые
прошли проверку
на толерантность
в эмбриональном
периоде
(клонально-селекционная
теория Бернета).
У млекопитающих
сформировались
два типа иммунитета:
клеточные и
гуморальный.
Это происходит
из-за того, что
у млекопитающих
развивается
2 типа лимфоцитов
- Т - и В-клеток.
Эти лимфоциты
образуются
из стволовых
клеток-предшественников
в костном мозге.
Специфический
и неспецифический
иммунитет.
Неспецифический
(врожденный)
иммунитет
обуславливает
однотипные
реакции на
любые чужеродные
антигены. Главным
клеточным
компонентом
системы неспецифического
иммунитета
служат фагоциты,
основная функция
которых - захватывать
и переваривать
проникающие
извне агенты.
Для возникновения
подобной реакции
чужеродный
агент должен
иметь поверхность,
т.е. быть частицей
(например, заноза).
Если же вещество
молекулярно-дисперсное
(например: белок,
полисахарид,
вирус), и при
этом не токсичное
и не обладает
физиологической
активностью
- оно не может
быть нейтрализовано
и выведено
организмом
по вышеописанной
схеме. В этом
случае реакцию
обеспечивает
специфический
иммунитет. Он
приобретается
в результате
контакта организма
с антигеном;
имеет приспособительное
значение и
характеризуется
формированием
иммунологической
памяти. Его
клеточными
носителями
служат лимфоциты,
а растворимыми
- иммуноглобулины
(антитела).
Первичный и
вторичный
иммунный ответ.
Специфические
антитела
продуцируются
специальными
клетками -
лимфоцитами.
Причем для
каждого вида
антител существует
свой тип лимфоцитов
(клон).
Первое взаимодействие
антигена (бактерии
или вируса) с
лимфоцитом
вызывает реакцию,
названную
первичным
иммунным ответом,
в ходе которого
лимфоциты
начинают развиваться
(пролиферировать)
в виде клонов,
претерпевая
затем дифференцировку:
некоторые из
них становятся
клетками памяти,
другие превращаются
в зрелые клетки,
продуцирующие
антитела. Главные
особенности
первичного
иммунного
ответа - существование
латентного
периода до
появления
антител, затем
выработка их
лишь в небольшом
количестве.
Вторичный
иммунный ответ
развивается
при последующем
контакте с тем
же самым антигеном.
Основная особенность
- быстрая пролиферация
лимфоцитов
с дифференцировкой
их в зрелые
клетки и быстрая
выработка
большого количества
антител, которые
высвобождаются
в кровь и тканевую
жидкость, где
могут встретиться
с антигеном
и эффективно
побороть болезнь.
Естественный
и искусственный
иммунитет.
К факторам
естественного
иммунитета
относят иммунные
и неиммунные
механизмы. К
первым относятся
гуморальные
(система комплемента,
лизоцим и др.
белки). Ко вторым
относятся
барьеры (кожа,
слизистая),
секрет потовых,
сальных, слюнных
желез (содержит
разнообразные
бактерицидные
вещества), желез
желудка (соляная
кислота и
протеолитические
ферменты), нормальная
микрофлора
(антагонисты
патогенных
микроорганизмов).
Искусственный
иммунитет
вырабатывает
при введении
в организм
вакцины или
иммуноглобулина.
Активный и
пассивный
иммунитет.
Активная
иммунизация
стимулирует
собственный
иммунитет
человека, вызывая
выработку
собственных
антител. Вырабатывается
у человека в
ответ на возбудитель.
Образуются
специализированные
клетки (лимфоциты),
которые продуцируют
антитела к
конкретному
возбудителю.
После инфекции
в организме
остаются "клетки
памяти", и в
случае последующих
столкновений
с возбудителем
начинают снова
(уже быстрее)
продуцировать
антитела.
Активный иммунитет
может быть
естественным
и искусственным.
Естественный
приобретается
в результате
перенесенного
заболевания.
Искусственный
вырабатывается
при введении
вакцин.
Пассивный
иммунитет: в
организм вводятся
уже готовые
антитела
(гамма-глобулин).
Введенные
антитела в
случае столкновения
с возбудителем
"расходуются"
(связываются
с возбудителем
в комплекс
"антиген-антитело"),
если встречи
с возбудителем
не произошло,
они имеют некий
период полужизни,
после чего
распадаются.
Пассивная
иммунизация
показана в тех
случаях, когда
необходимо
в короткие
сроки создать
иммунитет на
непродолжительное
время (например,
после контакта
с больным).
Когда ребенок
появляется
на свет, он обычно
имеет иммунитет
(невосприимчивость)
к некоторым
инфекциям. Это
заслуга борющихся
с болезнями
антител, которые
передаются
через плаценту
от матери к
будущему
новорожденному.
Передаются
антитела против
возбудителей
тех болезней,
которыми мать
переболела
или против
которых была
иммунизирована.
Впоследствии,
вскармливаемый
грудью младенец
постоянно
получает
дополнительную
порцию антител
с молоком матери.
Это естественный
пассивный
иммунитет. Он
также носит
временный
характер, угасая
к концу первого
года жизни.
Стерильный
и нестерильный
иммунитет
После заболевания
в некоторых
случаях иммунитет
сохраняется
пожизненно.
Например корь,
ветряная оспа.
Это стерильный
иммунитет. А
в некоторых
случаях иммунитет
сохраняется
только до тех
пор, пока в организме
есть возбудитель
(туберкулез,
сифилис) - нестерильный
иммунитет.
4. Гемопоэз
4.1 Стволовые
клетки крови
Стволовыми
клетками называют
клетки-предшественники,
из которых
образуются
при необходимости
все другие типы
клеток, составляющие
различные
органы и ткани
человека. Термин
"стволовая
клетка" впервые
ввел в 1908 году
русский гематолог
из Санкт-Петербурга
А. Максимов.
Значительный
объем исследований
стволовых
клеток проведен
биологами А.
Фриденштейном
и И. Чертковым
в России, в 60-х
годах прошлого
века. Именно
они открыли
мезенхимальные
стволовые
клетки (МСК) в
костном мозге,
обладающие
уникальной
регенерационной
способностью.
Отличие эмбриональных
и мезенхимальных
стволовых
клеток заключается
в том, что первые
могут быть
получены на
ранней стадии
развития эмбриона
человека (из
внутренней
массы бластоцисты
- оплодотворенной
яйцеклетки
- или из зачатков
половых органов
на самых ранних
этапах развития,
буквально в
первые дни), а
вторые встречаются
в течение всей
жизни человека
во всех его
органах и тканях.
Эмбриональные
СК значительно
активнее
мезенхимальных,
обладают более
высокой способностью
размножения,
большим потенциалом
дифференцировки.
Помимо мезенхимальных
СК выделяют
еще гемопоэтические
клетки - предшественники
клеток крови.
Они встречаются
в кровеносном
русле в отличие
от мезенхимальных,
которые в крови
циркулируют
только при
серьезных
повреждениях
организма.
Стволовые
клетки способны
восстанавливать
кроветворение
у облученных
животных
(радиозащитное
действие), длительно
поддерживать
кроветворение
и образовывать
колониеобразующие
единицы селезенки
(двенадцатидневные
селезеночные
колонии), дающие
начало гранулоцитарным,
моноцитарным,
эритроидным,
мегакариоцитарным
и лимфоидным
колониям. Все
клетки гемопоэтического
происхождения
образуются
из примитивных
стволовая
кроветворная
клеток (пСКК),
локализованных
в костном
мозге и дающих
начало клеткам
четырех основных
направлений
дифференцировки:
эритроидного
(эритроциты),
мегакариоцитарного
(тромбоциты),
миелоидного
(гранулоциты
и моноядерные
фагоциты)
лимфоидного
(лимфоциты).
Дивергенция
общего стволового
элемента происходит
на самом раннем
этапе костномозговой
дифференцировки.
Антигенпрезентирующие
клетки в основном,
но не исключительно,
развиваются
из миелоидных
клеток-предшественников.
Клетки миелоидного
и лимфоидного
ряда наиболее
важны для
функционирования
иммунной системы.
Лимфопоэтическая
своловая клетка
определяет
две самостоятельные
линии развития,
приводящие
к образованию
Т-клеток
и В-клеток.
Первая
образующаяся
из ГСК клетка-предшественник
представляет
собой колониеобразующуюся
единицу (КОЕ)
, которая определяет
линии развития,
приводящие
к образованию
гранулоцитов,
эритроцитов,
моноцитов
и мегакариоцитов.
Созревание
этих клеток
происходит
под влиянием
колониестимулирующих
факторов (КСФ)
и ряда интерлейкинов,
в том числе
ИЛ-1,
ИЛ-3,
ИЛ-4,
ИЛ-5
и ИЛ-6.
Все они играют
важную роль
в положительной
регуляции
(стимуляции)
гемопоэза и
продуцируются,
главным образом,
стромальными
клетками костного
мозга, но также
и зрелыми формами
дифференцированных
миелоидных
и лимфоидных
клеток. Другие
цитокины
(например, ТРФ-бета)
могут осуществлять
понижающую
регуляцию
(подавление)
гемопоэза).
У всех клеток
как лимфоидного,
так и миелоидного
ряда время
жизни ограничено,
и все они непрерывно
образуются.
У
млекопитающих
в период внутриутробного
развития ГСК
присутствуют
в желточном
мешке, печени,
селезенке
и костном
мозге. Во взрослом
организме
гемопоэтические
стволовые
клетки находятся
в основном в
костном мозге,
где они в норме
довольно редко
делятся, производя
новые стволовые
клетки (самообновление).
Животное можно
спасти от последствий
облучения в
летальных дозах
введением
клеток костного
мозга, которые
заселяют его
лимфоидную
и миелоидную
ткани.
Плюрипотентные
стволовые
клетки дают
начало коммитированным
клеткам-предшественницам,
которые уже
необратимо
определились
как предки
кровяных клеток
одного или
нескольких
типов. Полагают,
что коммитированные
клетки делятся
быстро, но
ограниченное
число раз, при
этом делятся
они под воздействием
факторов
микроокружения:
соседних клеток
и растворимых
или мембраносвязанных
цитокинов.
В конце такой
серии делений
клетки эти
становятся
терминально
дифференцированными,
обычно больше
не делятся и
погибают через
несколько дней
или недель.
Плюрипотентные
стволовые
клетки малочисленны,
их трудно
распознавать,
и все еще неясно,
как они выбирают
свой путь среди
разных вариантов
развития.
Программирование
клеточных
делений и выведение
клеток на
определенный
путь дифференцировки
(коммитирование),
видимо, включают
в себя и случайные
события. Стволовая
клетка плюрипотентна,
т.к. дает начало
многим видам
терминально
дифференцированных
клеток. Что
касается клеток
крови, то эксперименты
показывают,
что все классы
клеток крови
- и миелоидных
и лимфоидных
- происходят
от общей гемопоэтической
стволовой
клетки.
Гемопоэтическая
стволовая
клетка развивается
следующим
образом. У эмбриона
гемопоэз
начинается
в желточном
мешке, но по
мере развития
эта функция
переходит к
печени
плода и, наконец,
к костному
мозгу, где и
продолжается
в течение всей
жизни. Гемопоэтическая
стволовая
клетка, дающая
начало всем
элементам
крови, плюрипотентна
и заселяет
другие гемо
- и лимфопоэтические
органы и
самовоспроизводится,
превращаясь
в новые стволовые
клетки. Животное
можно спасти
от последствий
облучения
в летальных
дозах введением
клеток костного
мозга, которые
заселяют его
лимфоидную
и миелоидную
ткани.
Во
взрослом организме
гемопоэтические
стволовые
клетки находятся
главным образом
в костном
мозге, где они
в норме довольно
редко делятся,
производя новые
стволовые
клетки (самообновление).
Клетку-предшественницу,
дающую в культуре
клеток начало
колонии эритроцитов,
называют
колониеобразующей
единицей эритроидного
ряда, или КОЕ-Э,
и она дает начало
зрелым эритроцитам
после шести
или даже меньшего
числа циклов
деления. КОЕ-Э
еще не содержит
гемоглобин.
Гемопоэзом
(haemopoesis) называют
развитие крови.
Различают
эмбриональный
гемопоэз, который
происходит
в эмбриональный
период
и приводит
к развитию
крови как ткани,
и постэмбриональный
гемопоэз, который
представляет
собой процесс
физиологической
регенерации
крови. Развитие
эритроцитов
называют
эритропоэзом,
развитие гранулоцитов
- гранулоцитопоэзом,
тромбоцитов
- тромбоцитопоэзом,
развитие моноцитов
- моноцитопоэзом,
развитие лимфоцитов
и иммуноцитов
- лимфоцито - и
иммуноцитопоэзом.
Эмбриональный
гемопоэз.
В развитии
крови как ткани
в эмбриональный
период можно
выделить 3 основных
этапа, последовательно
сменяющих друг
друга:
1) мезобластический,
когда начинается
развитие клеток
крови во внезародышевых
органах - мезенхиме
стенки желточного
мешка, хориона
и стебля (с 3-й
по 9-ю неделю
развития зародыша
человека) и
появляется
первая генерация
стволовых
клеток крови
(СКК);
2) печеночный,
который начинается
в печени с 5-6-й
недели развития
плода, когда
печень становится
основным органом
гемопоэза, в
ней образуется
вторая генерация
СКК.
Кроветворение
в печени достигает
максимума через
5 месяцев и
завершается
перед рождением.
СКК печени
заселяют тимус
(здесь, начиная
с 7-8-й недели,
развиваются
Т-лимфоциты),
селезенку
(гемопоэз начинается
с 12-й недели) и
лимфатические
узлы (гемопоэз
отмечается
с 10-й недели);
3) медуллярный
(костномозговой)
- появление
третьей генерации
СКК в костном
мозге, где гемопоэз
начинается
с 10-й недели и
постепенно
нарастает к
рождению, а
после рождения
костный мозг
становится
центральным
органом гемопоэза.
Кроветворение
в стенке желточного
мешка. У человека
оно начинается
в конце 2-й - начале
3-й недели эмбрионального
развития. В
мезенхиме
стенки желточного
мешка обособляются
зачатки сосудистой
крови, или кровяные
островки. В них
мезенхимные
клетки округляются,
теряют отростки
и преобразуются
в стволовые
клетки крови.
Клетки, ограничивающие
кровяные островки,
уплощаются,
соединяются
между собой
и образуют
эндотелиальную
выстилку будущего
сосуда. Часть
СКК дифференцируется
в первичные
клетки крови
(бласты), крупные
клетки с базофильной
цитоплазмой
и ядром, в котором
хорошо заметны
крупные ядрышки.
Большинство
первичных
кровяных клеток
митотически
делится и
превращается
в первичные
эритробласты,
характеризующиеся
крупным размером
(мегалобласты).
Это превращение
совершается
в связи с накоплением
эмбрионального
гемоглобина
в цитоплазме
бластов, при
этом сначала
образуются
полихроматофильные
эритробласты,
а затем оксифилъные
эритробласты
с большим содержанием
гемоглобина.
В некоторых
первичных
эритробластах
ядро подвергается
кариорексису
и удаляется
из клеток, в
других ядро
сохраняется.
В результате
образуются
безъядерные
и ядросодержащие
первичные
эритроциты,
отличающиеся
большим размером
по сравнению
с нормоцитами
и поэтому получившие
название мегалоцитов.
Такой тип
кроветворения
называется
мегалобластическим.
Он характерен
для эмбрионального
периода, но
может появляться
в постнатальном
периоде при
некоторых
заболеваниях
(злокачественное
малокровие).
Наряду с мегалобластическим
в стенке желточного
мешка начинается
нормобластическое
кроветворение,
при котором
из бластов
образуются
вторичные
эритробласты;
сначала они
превращаются
в полихроматофильные
эритробласты,
далее в нормобласты,
из которых
образуются
вторичные
эритроциты
(нормоциты);
размеры последних
соответствуют
эритроцитам
(нормоцитам)
взрослого
человека. Развитие
эритроцитов
в стенке желточного
мешка происходит
внутри первичных
кровеносных
сосудов, т.е.
интраваскулярно.
Одновременно
экстраваскулярно
из бластов,
расположенных
вокруг сосудистых
стенок, дифференцируется
небольшое
количество
гранулоцитов
- нейтрофилов
и эозинофилов.
Часть СКК остается
в недифференцированном
состоянии и
разносится
током крови
по различным
органам зародыша,
где происходит
их дальнейшая
дифференцировка
в клетки крови
или соединительной
ткани. После
редукции желточного
мешка основным
кроветворным
органом временно
становится
печень.
Кроветворение
в печени. Печень
закладывается
примерно на
3-4-й неделе эмбриональной
жизни, а с 5-й недели
она становится
центром кроветворения.
Кроветворение
в печени происходит
экстраваскулярно,
по ходу капилляров,
врастающих
вместе с мезенхимой
внутрь печеночных
долек. Источником
кроветворения
в печени являются
стволовые
клетки крови,
из которых
образуются
бласты, дифференцирующиеся
во вторичные
эритроциты.
Процесс их
образования
повторяет
описанные выше
этапы образования
вторичных
эритроцитов.
Одновременно
с развитием
эритроцитов
в печени образуются
зернистые
лейкоциты,
главным образом
нейтрофильные
и эозинофильные.
В цитоплазме
бласта, становящейся
более светлой
и менее базофильной,
появляется
специфическая
зернистость,
после чего ядро
приобретает
неправильную
форму. Кроме
гранулоцитов,
в печени формируются
гигантские
клетки - мегакариоциты.
К концу внутриутробного
периода кроветворение
в печени прекращается.
Кроветворение
в тимусе. Тимус
закладывается
в конце 1-го месяца
внутриутробного
развития, и на
1-8-й неделе его
эпителий начинает
заселяться
стволовыми
клетками крови,
которые дифференцируются
в лимфоциты
тимуса. Увеличивающееся
число лимфоцитов
тимуса дает
начало Т-лимфоцитам,
заселяющим
Т-зоны периферических
органов иммунопоэза.
Кроветворение
в селезенке.
Закладка
селезенки
происходит
в конце 1-го месяца
эмбриогенеза.
Из вселяющихся
сюда стволовых
клеток происходит
экстраваскулярное
образование
всех видов
форменных
элементов
крови, т.е. селезенка
в эмбриональном
периоде представляет
собой универсальный
кроветворный
орган. Образование
эритроцитов
и гранулоцитов
в селезенке
достигает
максимума на
5-м месяце эмбриогенеза.
После этого
в ней начинает
преобладать
лимфоцитопоэз.
Кроветворение
в лимфатических
узлах. Первые
закладки
лимфатических
узлов человека
появляются
на 7-8-й неделе
эмбрионального
развития. Большинство
лимфатических
узлов развивается
на 9-10-й неделе.
В этот же период
начинается
проникновение
в лимфатические
узлы стволовых
клеток крови,
из которых на
ранних стадиях
дифференцируются
эритроциты,
гранулоциты
и мегакариоциты.
Однако формирование
этих элементов
быстро подавляется
образованием
лимфоцитов,
составляющих
основную часть
лимфатических
узлов. Появление
единичных
лимфоцитов
происходит
уже в течение
8-15-й недели развития,
однако массовое
"заселение"
лимфатических
узлов предшественниками
Т - и В-лимфоцитов
начинается
с 16-й недели, когда
формируются
посткапиллярные
венулы, через
стенку которых
осуществляется
процесс миграции
клеток. Из
клеток-предшественников
дифференцируются
лимфобласты
(большие лимфоциты),
а далее средние
и малые лимфоциты.
Дифференцировка
Т - и В-лимфоцитов
происходит
в Т - и В-зависимых
зонах лимфатических
узлов.
Кроветворение
в костном мозге.
Закладка костного
мозга осуществляется
на 2-м месяце
эмбрионального
развития. Первые
гемопоэтические
элементы появляются
на 12-й неделе
развития; в это
время основную
массу их составляют
эритробласты
и предшественники
гранулоцитов.
Из СКК в костном
мозге формируются
все форменные
элементы крови,
развитие которых
происходит
экстраваскулярно.
Часть СКК сохраняется
в костном мозге
в недифференцированном
состоянии, они
могут расселяться
по другим органам
и тканям и являться
источником
развития клеток
крови и соединительной
ткани. Таким
образом, костный
мозг становится
центральным
органом, осуществляющим
универсальный
гемопоэз, и
остается им
в течение
постнатальной
жизни. Он обеспечивает
стволовыми
кроветворными
клетками тимус
и другие гемопоэтические
органы.
Постэмбриональный
гемопоэз.
Постэмбриональный
гемопоэз представляет
собой процесс
физиологической
регенерации
крови (клеточное
обновление),
который компенсирует
физиологическое
разрушение
дифференцированных
клеток.
Миелопоэз
происходит
в миелоидной
ткани (textus myeloideus),
расположенной
в эпифизах
трубчатых и
полостях многих
губчатых костей.
Здесь развиваются
форменные
элементы крови:
эритроциты,
гранулоциты,
моноциты, кровяные
пластинки,
предшественники
лимфоцитов.
В миелоидной
ткани находятся
стволовые
клетки крови
и соединительной
ткани.
Предшественники
лимфоцитов
постепенно
мигрируют и
заселяют такие
органы, как
тимус, селезенка,
лимфатические
узлы и др.
Лимфопоэз
происходит
в лимфоидной
ткани (textus lymphoideus), которая
имеет несколько
разновидностей,
представленных
в тимусе, селезенке,
лимфатических
узлах. Она выполняет
основные функции:
образование
Т - и В-лимфоцитов
и иммуноцитов
(плазмоцитов
и др.).
Миелоидная
и лимфоидная
ткани являются
разновидностями
соединительной
ткани, т.е. относятся
к тканям внутренней
среды. В них
представлены
две основные
клеточные линии
- клетки ретикулярной
ткани и гемопоэтические.
Ретикулярные,
а также жировые,
тучные и остеогенные
клетки вместе
с межклеточным
веществом
(матрикс) формируют
микроокружение
для
гемопоэтических
элементов.
Структуры
микроокружения
и гемопоэтические
клетки
функционируют
в неразрывной
связи. Микроокружение
оказывает
воздействие
на дифференцировку
клеток крови
(при контакте
с их рецепторами
или путем выделения
специфических
факторов).
Для миелоидной
и всех разновидностей
лимфоидной
ткани характерно
наличие
стромальных
ретикулярных
и гемопоэтических
элементов,
образующих
единое функциональное
целое. В тимусе
имеется сложная
строма, представленная
как соединительнотканными,
так и ретикулоэпителиальными
клетками.
Эпителиальные
клетки секретируют
особые вещества
- тимозины,
оказывающие
влияние на
дифференцировку
из СКК Т-лимфоцитов.
В лимфатических
узлах и селезенке
специализированные
ретикулярные
клетки создают
микроокружение,
необходимое
для пролиферации
и дифференцировки
в специальных
Т - и В-зонах Т
- и В-лимфоцитов
и плазмоцитов.
СКК являются
плюрипотентными
(полипотентными)
предшественниками
всех клеток
крови и относятся
к самоподдерживающейся
популяции
клеток. Они
редко делятся.
Впервые представление
о родоначальных
клетках крови
сформулировал
в начале XX в.А.
А. Максимов,
который считал,
что по своей
морфологии
они сходны с
лимфоцитами.
В настоящее
время это
представление
нашло подтверждение
и дальнейшее
развитие в
новейших
экспериментальных
исследованиях,
проводимых
главным образом
на мышах. Выявление
СКК стало возможным
при применении
метода колониеобразования.
Экспериментально
(на мышах) показано,
что при введении
смертельно
облученным
животным (утратившим
собственные
кроветворные
клетки) взвеси
клеток красного
костного мозга
или фракции,
обогащенной
СКК, в селезенке
появляются
колонии клеток
- потомков одной
СКК. Пролиферативную
активность
СКК модулируют
колониестимулирующие
факторы (КСФ),
интерлейкины
(ИЛ-3 и др.). Каждая
СКК