Поступление и превращение азота в растениях
Поглощение
азота растением
Формы
азота, используемые
растением. Азот
входит в состав
важнейшей части
живого организма,
а именно в состав
запасных белков
и белков цитоплазмы.
В составе золы
азота нет, так
как при сжигании
растений он
образует газообразные
окислы. В сухом
веществе растения
содержится
в среднем 1,5%
азота.
Добывание азота
представляет
для растения
наибольшие
трудности, так
как азот не
входит в состав
минералов и
его накопление
и превращение
в почве полностью
связано с
жизнедеятельностью
организмов.
В почве
доступный для
растения азот
находится в
основном в
форме нитратов
аммонийных
солей.
Восстановление
нитратов растениями.
Нитраты
представляют
собой окисленную
форму азота
и должны быть
восстановлены
растением до
NH2, после
чего они могут
войти в состав
аминокислот,
а затем белка.
Можно считать,
что восстановление
нитратов идет
двумя путями:
1)
восстановление
за счет химической
энергии дыхания
и 2)
фотохимическое
восстановление
в хлоропластах.
Восстановление
нитратов идет
этапами: сначала
до азотистой
кислоты HNO2,
затем до гидрокисламина
NH2OH и,
наконец, до
аммиака NH3.
Восстановление
нитратов до
NH3-и NH2-гpyпп
осуществляется
с помощью фермента
нитратредуктазы,
в состав кофермента
которой входит
молибден.
Восстановленный
азот нитратов
или непосредственно
поглощенный
ион аммония,
соединяясь
с продуктами
превращения
углеводов,
образует
аминокислоты,
а затем белки.
Аммиак, реагируя
с некоторыми
органическими
кислотами,
может образовать
аминокислоты.
Так, например,
аммиак, реагируя
с пировиноградной
кислотой, образует
аминокислоту
аланин:
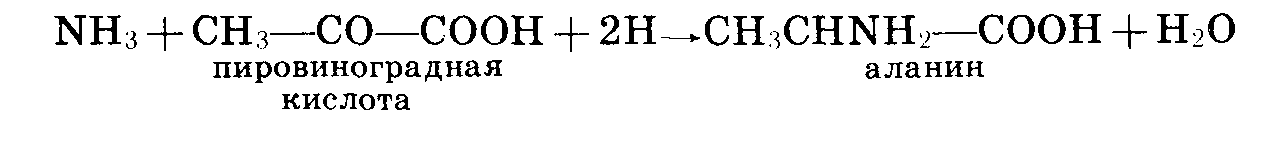
Образовавшиеся
белковые вещества
подвергаются
превращениям
в теле растения.
Животный организм
все время выводит
азот из своего
тела в виде
мочевины и
отчасти мочевой
кислоты. В отличие
от животных
растение очень
бережно относится
к азоту, не теряя
его.
При прорастании
семян расщепляются
запасные белки,
а количество
конституционных
белков не только
не уменьшается,
а все время
увеличивается.
Затем происходит
накопление
белков в связи
с переходом
растения к
автотрофному
питанию.
Роль
амида, аспарагина,
глютамина и
мочевины в
растении. При
восстановлении
нитратов, а
также при
дезаминировании
аминокислот
(т.е. отщеплении
от них аммиака)
в растениях
может накопляться
аммиак, который
ядовит для
большинства
из них. В растении
аммиак обезвреживается,
так как он
связывается
аспарагиновой
или глутаминовой
кислотой, образуя
соответственные
амиды (аспарагин,
глутамин). У
многих низших
растений образуется
мочения:
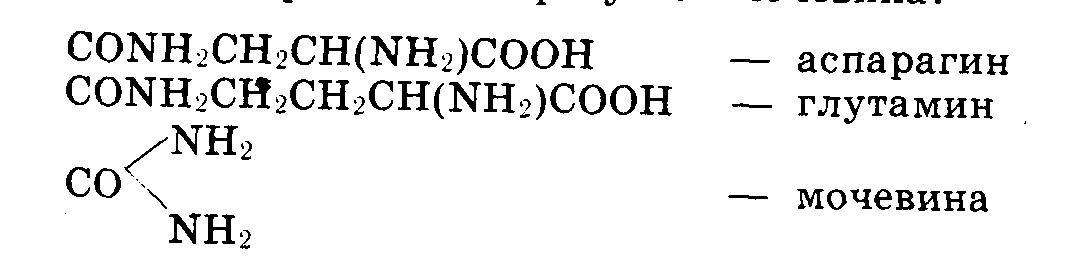
Доказан
и прямой синтез
мочевины из
углеводов и
аммиака у многих
грибов (дождевики,
шампиньоны).
Содержание
мочевины у
дождевиков
доходит до
10,7% от сухого
вещества. Таким
образом, аспарагин,
глутамин и
мочевина играют
большую физиологическую
роль, так как
являются
соединениями,
обезвреживающими
ядовитое действие
аммиака, а также
представляют
собой резерв
аминогрупп
NH2
в растении для
синтеза аминокислот.
Подводя
итоги, можно
отметить два
типа синтеза
белков: первичный
и вторичный.
В обоих этих
синтезах аммиак
играет большую
роль, что и дало
возможность
Д.Н. Прянишникову
сказать, что
аммиак есть
альфа и омега
(первая и последняя
буквы греческого
алфавита), т.е.
начало и конец,
превращения
белков в растениях.
При первичном
синтезе из
аммиака и углеводов
строится белок
(левая часть
схемы). При распаде
белка образуются
аминокислоты,
от которых при
дезаминировании
отщепляется
аммиак, связывающийся
в аспарагин
или глютамин.
При вторичном
синтезе белков
(правая и нижняя
части схемы)
происходит
отщепление
аммиака от
аспарагина
и образование
аминокислот
из углеводов
(вернее, из продуктов
их превращения)
и аммиака. Все
эти представления
можно объединить
в следующую
схему Прянишникова:
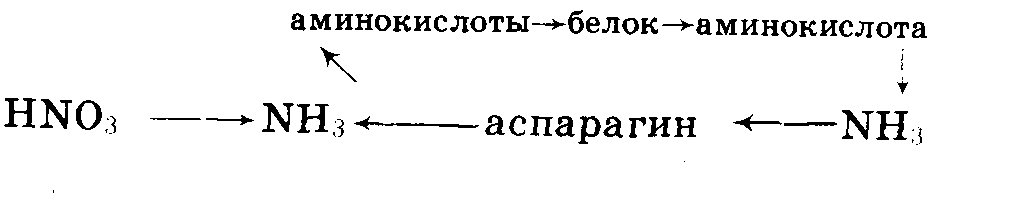
Усвоение
органических
форм азота
Стерильные
культуры
покрытосеменных
растений
Долгое
время оставался
нерешенным
вопрос о возможности
усвоения корневой
системой растений
органических
форм азота.
Вопрос этот
можно было
решить только
в стерильных
культурах, так
как в нестерильных
условиях развились
бы бактерии,
которые своими
ферментами
разложили бы
органический
азот и превратили
бы его в минеральные
формы. Корневая
система высших
растений находилась
в простерилизованном
питательном
растворе, содержащем
органический
азот. Семена
растений
стерилизовались
бромной водой
или раствором
сулемы.
Опыты
показали, что
хотя аминокислоты
и могут быть
усвоены зеленым
растением, но
это усвоение
идет крайне
медленно, и
растения, выращенные
на этих соединениях,
всегда отстают
в росте от растений,
получивших
минеральные
формы азота.
Насекомоядные
растения. Большой
интерес представляют
высшие растения
со своеобразным
типом азотистого
питания. Сюда
относятся
некоторые
сапрофиты,
паразиты,
полупаразиты
и, наконец,
насекомоядные
растения. Своеобразие
азотистого,
а у некоторых
форм и углеродного
питания возникло
в процессе
эволюции под
влиянием условий
существования
и естественного
отбора. Таким
образом, в отличие
от грибов и
бактерий, где
гетеротрофное
питание азотом
имеет первичный
характер, у
этих растений
оно возникло
вторично. Наиболее
интересную
группу растений,
питающихся
органическим
азотом, составляют
насекомоядные
растения.
К
насекомоядным
растениям
принадлежит
примерно 500 видов
растений. Все
они обитатели
болот. Несмотря
на богатство
болотных почв
органическим
веществом,
находящийся
в этих почвах
органический
азот недоступен
для растений.
Болотные почвы
также очень
бедны и минеральными
солями (фосфор,
калий и др.). Все
насекомоядные
растения имеют
хлорофилл, т.е.
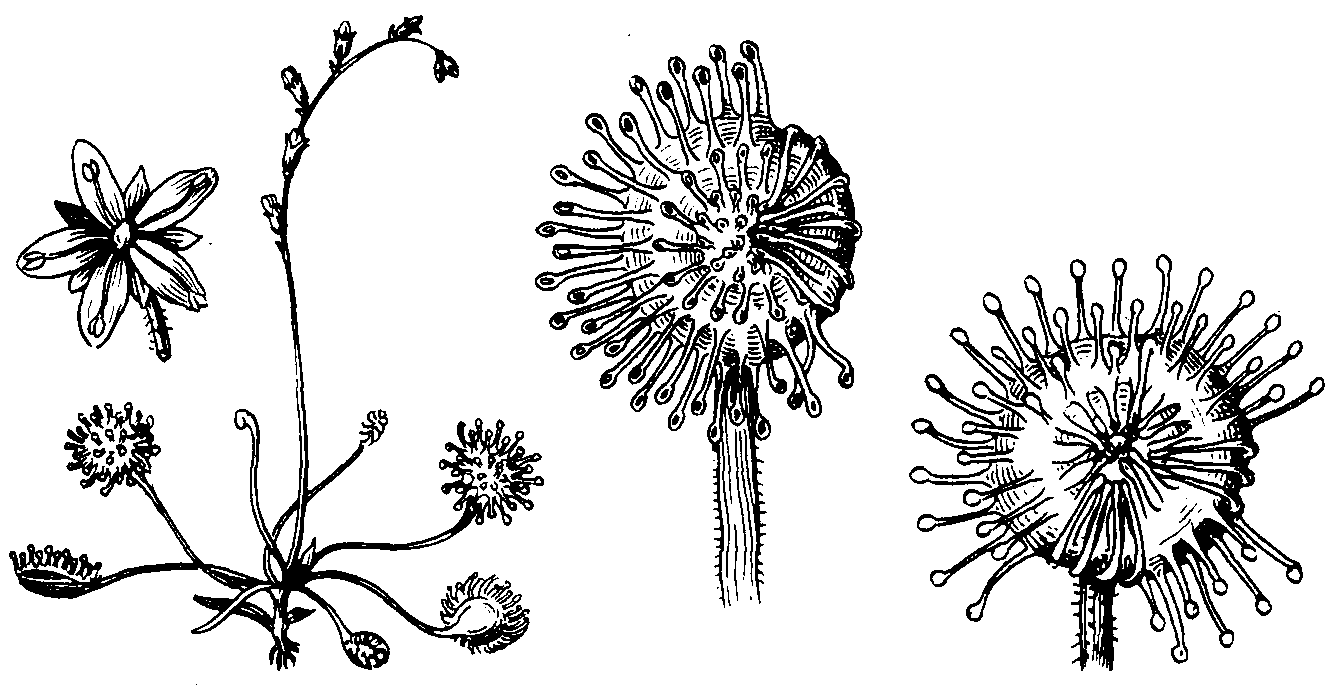
Ознакомимся
с некоторыми
представителями
насекомоядных
растений.
Росянка
- многолетнее
растение, растет
на сфагновых
болотах. Каждый
год на поверхности
мха образуется
новая розетка
листьев росянки.
Листья снабжены
многочисленными
железистыми
волосками (их
часто называют
щупальцами),
выделяющими
липкую жидкость,
к которой и
прилипают
мелкие насекомые
- комары и мелкие
мухи. При прилипании
насекомого
пластинка листа
свертывается.
Особенно хорошо
это свертывание
пластинки видно
у вида росянки
длиннолистной.
После того как
насекомое
прилипнет к
листу, в растении
начинается
выделение
гидролитических
ферментов -
протеаз, разлагающих
белки, и муравьиной
кислоты. Кислота
способствует
работе фермента
протеазы, и,
кроме того, она
действует как
яд на бактериальную
флору. Последнее
очень важно,
так как пышное
развитие флоры
гнилостных
бактерий могло
бы сказаться
отрицательно
на самом растении.
Ботаник
Фрэнсис Дарвин,
сын Чарльза
Дарвина, выяснил
благоприятное
влияние питания
росянки насекомыми.
Он взял шесть
сосудов с растениями
росянки и разделил
каждый из них
перегородкой.
По одну сторону
перегородки
росянки получали
мясо, а по другую
сторону им его
не давали.
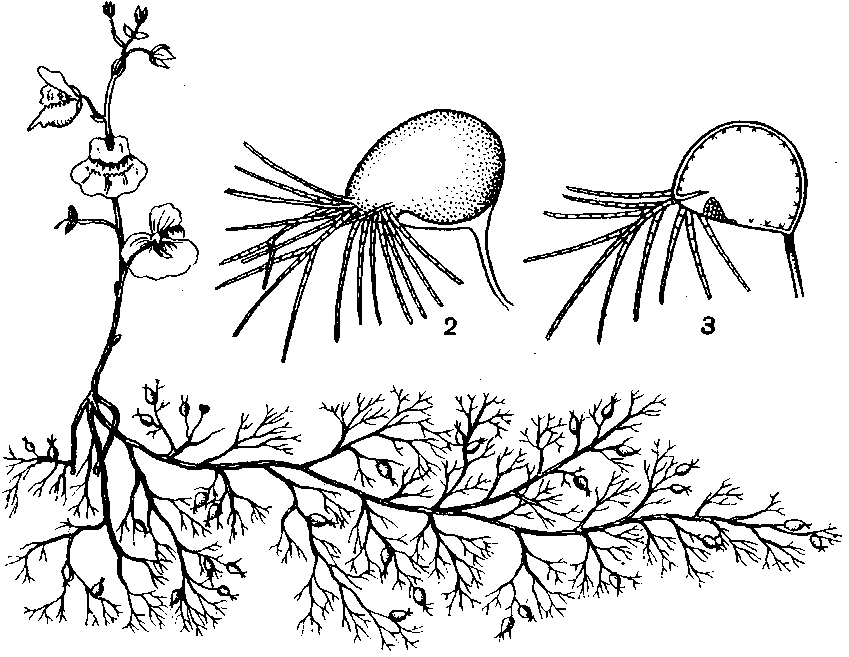
В
конце опыта
выяснилось,
что на 100 цветков
у контрольной
группы, которые
не получали
мяса, приходится
165 цветков у
получавших
мясо. Иными
словами, репродуктивная
способность
растений росянки,
питавшихся
мясом, сильно
возрастала.
Большой
интерес представляет
обитающая в
болотистых
водоемах пузырчатка.
Помимо рассеченных
листьев, она
несет еще характерные
пузыревидно
измененные
листья. В такой
пузырек проникают
мелкие рачки
и уже не могут
выбраться
наружу, так как
створка, сквозь
которую проник
рачок, открывается
в одну сторону.
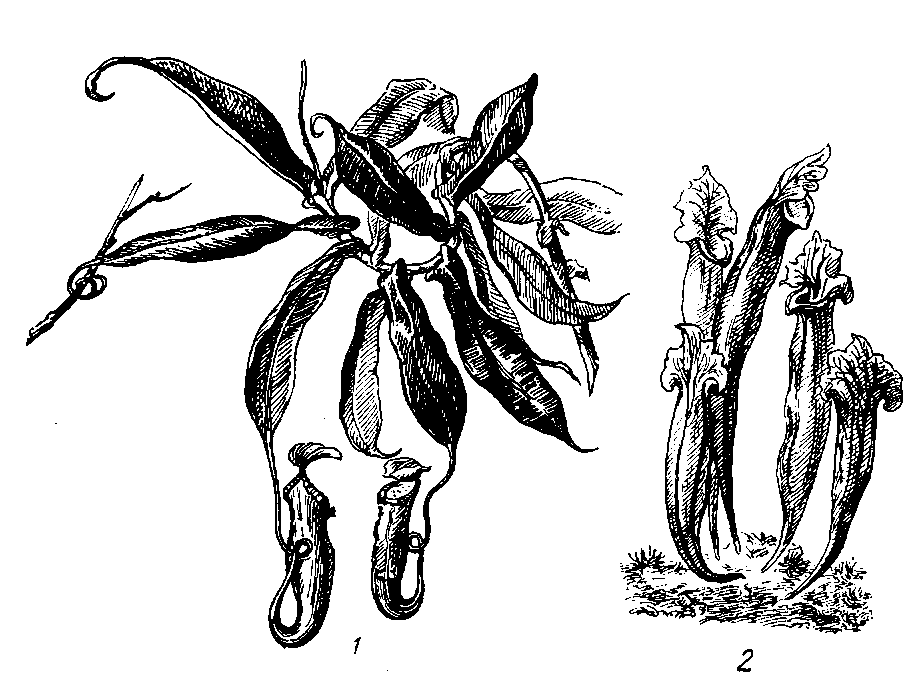
Из
других насекомоядных
растений можно
отметить кувшиноносы
(непентес)
из тропиков
Мадагаскара
и Явы: австралийский
цефалотус и
американскую
сарацению,
листья
которых имеют
вид кувшинов,
куда и попадают
насекомые. Эти
растения также
выделяют
гидролизирующие
белки-ферменты
и переваривают
насекомых. У
цефалотуса
выделения
ферментов не
происходит.
Многие
насекомоядные
растения привлекают
к себе насекомых
яркой окраской
листьев, а некоторые
выделяют сладкий
сок. Так, например,
у кувшиноноса
в верхней части
кувшинчиков
имеются железки,
выделяющие
сладкий сок.
Особенно
интересна
венерина
мухоловка,
растущая
на болотах
штата Каролина
в Северной
Америке. Это
небольшое
растение активно
захлопывает
створки листьев,
когда насекомое
заденет чувствительный
волосок его
листа.

Симбиоз
и паразитизм.
Особую группу
покрытосеменных
растений составляют
сапрофиты.
Встречаются
они на богатой
органическими
веществами
почве, в лесах,
среди разлагающейся
лесной подстилки.
К ним относятся
такие растения,
как подъельник
и орхидея гнездовка.
Оба эти растения
бесцветны.
Правда, в листьях
гнездовки
содержится
небольшое
количество
хлорофилла
а, хлорофилла
б у нее совсем
не найдено.
Подъельник
- растение-сапрофит,
лишенное хлорофилла.
По-видимому,
гриб снабжает
подъельник
углеводами
и азотистыми
веществами
из малодоступного
для растения
перегноя, очевидно,
получая от
растения
физиологически
активные вещества
(витамины), а
также, возможно,
и аминокислоты.
Выращивая сосну
в стерильных
условиях и
затем заражая
ее определенным
видом гриба
(эктотрофная
микориза), удалось
экспериментально
доказать наличие
связи между
высшим растением
и грибом. При
наблюдениях
за растениями
и грибами в
природной
обстановке
выявилась
тесная связь
между определенными
грибами и высшими
растениями.
По меткому
выражению
одного ученого,
гриб кортинариус
следует за
березой, как
"дельфин за
кораблем".
Большинство
наших съедобных
грибов образуют
эктотрофную
микоризу и
тесно связано
с определенными
деревьями. Это
давно отмечено
в названиях
грибов.
В
настоящее
время показано,
что семена
орхидеи содержат
очень незначительные
количества
витамина РР
(никотиновой
кислоты). Грибок
снабжает семена
орхидеи никотиновой
кислотой, после
чего они и начинают
прорастать.
Кроме того,
синтез витамина
Bi
также
несколько
затруднен у
орхидей, и снабжение
семян этими
веществами
способствует
их прорастанию
и росту корней
и надземной
массы.
Большинство
травянистых
дикорастущих
и культурных
растений также
содержат эндотрофную
микоризу, вызываемую
низшими грибами,
имеющими
неразделенный
перегородками
мицелий. Отмечено,
что при неблагоприятных
условиях, например
при сильном
увлажнении,
гриб часто
становится
паразитом
растения.
По-видимому,
и в случае
эндотрофной
микоризы гриб
снабжает растение
азотом, добывая
его из перегноя,
а от растения
получает углеводы,
а также физиологически
активные вещества.
К последней
группе растений,
отклоняющихся
в своем азотистом
питании, относятся
полупаразиты
и паразиты.
По-видимому,
путь к паразитизму
у высших растений
лежал и лежит
через полупаразитизм.
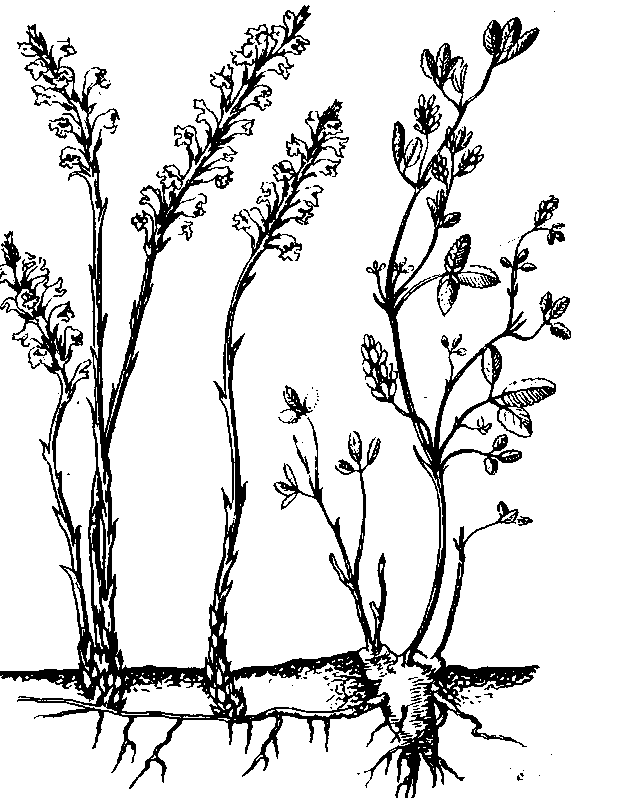
Много
полупаразитов
встречается
в семействе
норичниковых.
Среди полупаразитов
из норичниковых
можно отметить
характерные
растения лугов:
погремок, очанку
и др. Растения
эти присасываются
своими корнями
к корням других
растений. Одни
из них сохраняют
более или менее
нормальную
зеленую окраску,
а другие уже
значительно
меньше содержат
хлорофилла
(как, например,
погремок). Основной
причиной перехода
этих растений
к паразитическому
образу жизни
является слабое
развитие корневой
системы, вследствие
чего они не
могут свести
своего водного
баланса.
Из
полных паразитов
можно упомянуть
о видах заразихи,
поражающей
подсолнечник,
тыквенные и
ряд дикорастущих
растений. Мелкие
семена заразихи
прорастают,
стимулируемые
подкислением
субстрата
корневыми
выделениями.
Основным мероприятием
по борьбе с
заразихой
является создание
невосприимчивых
(иммунных) сортов.
Паразит
повилика
в
отличие от
заразихи, которая
поражает корни,
обвивает растение
и присасывается
к его стеблю.
У повилики
имеется очень
незначительное
количество
хлорофилла.
Проросток
повилики совершает
круговое движение,
свойственное
всем растениям,
но у повилики
оно проявляется
особенно резко.
Если при этом
повилика не
встретит растения,
вокруг которого
она может обвиться,
то она погибает.
Усвоение
молекулярного
азота микроорганизмами
Клубеньковые
бактерии.
Способность
бобовых растений
использовать
атмосферный
азот была доказана
опытами немецких
ученых Г. Гельригеля
и Г. Вильфарта
в 1886 г. Им удалось
показать, что,
посеянные в
прогретый
песок, в котором
убиты все бактерии,
бобовые растения,
не образующие
в этом случае
клубеньков,
не усваивают
(не фиксируют)
атмосферный
азот, а растут
лишь при наличии
его в виде
сортветственных
солей в песке.
Впоследствии
бактерии были
выделены в
чистую культуру
и названы
клубеньковыми
бактериями.
Оказалось, что,
прекрасно
развиваясь
на питательных
средах, клубеньковые
бактерии обычно
не фиксируют
при этом атмосферного
азота. Усвоение
азота воздуха
идет у них
беспрепятственно
только в симбиозе
(сожительстве)
с бобовыми
растениями.
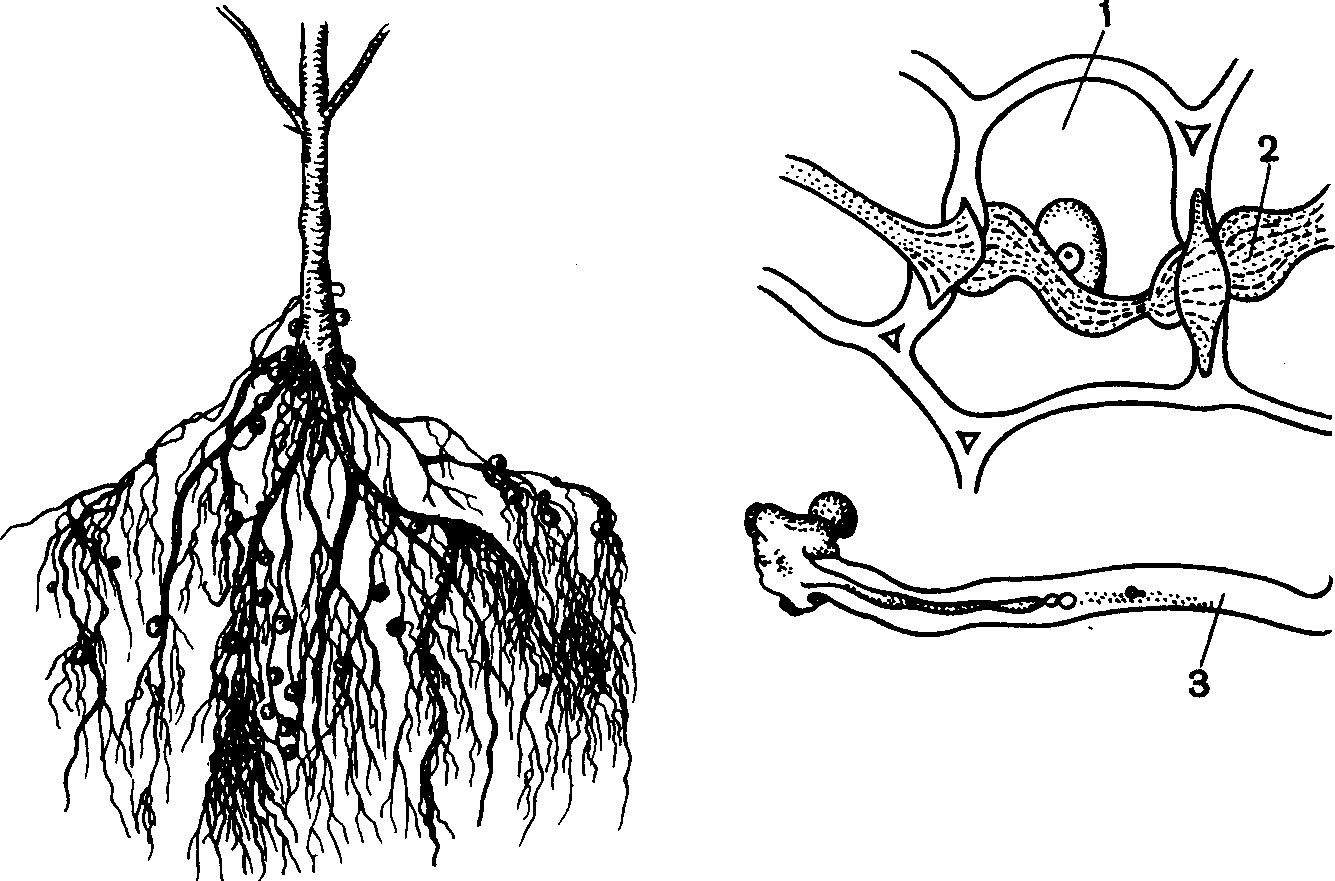
Характер
симбиоза. Находящиеся
в почве клубеньковые
бактерии проникают
в корень бобового
растения и
здесь начинают
размножаться,
образуя сплошной
тяж бактерий,
идущий через
ряд клеток.
Бактерии интенсивно
делятся и заполняют
клетки корня.
Бобовое растение
не остается
инертным по
отношению к
проникшей
бактерии, а
реагирует
усиленным
делением клеток,
разрастающихся
в виде клубеньков
или желваков.
Клубеньковые
бактерии приносят
растению пользу,
снабжая его
азотом.
Специфичность
клубеньковых
бактерий.
Клубеньковые
бактерии,
поселяющиеся
на корнях клевера,
не заражают
никакой другой
бобовой культуры.
Клубеньковые
бактерии,
развивающиеся
на горохе, могут,
кроме гороха,
заражать вику,
чечевицу, чину
и конские бобы.
Иными словами,
клубеньковые
бактерии образуют
специфические
расы, заражающие
только определенные
виды бобовых
растений.
Вирулентность
клубеньковых
бактерий.
Вирулентностью
бактерий называется
их способность
заражать данное
растение. Очень
часто клубеньковые
бактерии оказываются
маловирулентными,
т.е. не заражают
или плохо заражают
бобовые растения.
Активность
клубеньковых
бактерий. Помимо
вирулентности,
важное значение
имеет и активность
данной расы
бактерий. Раса
клубеньковой
бактерии может
быть очень
вирулентной,
но в то же время
неактивной,
т.е. она может
давать много
клубеньков,
но не усваивать
атмосферного
азота.
Бактериальное
удобрение
нитрагин. Фактически
очень часто
даже на землях,
где десятилетиями
культивировались
мотыльковые
растения, на
корнях образуется
очень небольшое
число клубеньков
или даже их
совсем не образуется.
Для того чтобы
обеспечить
наличие активных
клубеньков,
мотыльковые
растения перед
посевом можно
заразить
бактериальным
препаратом,
состоящим
обычно из нескольких
рас клубеньковых
бактерий. Такой
бактериальный
препарат получил
название нитрагин.
Другие
азотфиксирующие
симбиотические
организмы.
Помимо
клубеньковых
бактерий, в
природе встречаются
и другие аналогичные
симбиозы. На
корнях ольхи
образуются
большие деревянистые
вздутия (клубеньки),
в которых находятся
актиномицеты,
фиксирующие
атмосферный
азот.
Свободноживущие
азотфиксаторы.
Помимо
клубеньковых
бактерий, в
почве встречаются
еще и другие
виды, способные
усваивать
атмосферный
азот. Выделить
подобную бактерию
удалось С.Н.
Виноградскому
в 1893 г. на специальной
среде для
азотфиксирующих
бактерий. Для
этой цели он
взял среду,
содержащую
глюкозу и некоторые
соли, но абсолютно
не содержащую
связанного
азота ни в
органической,
ни в минеральной
форме. Таким
образом, в этой
среде могли
развиваться
только те бактерии,
которые усваивают
азот из воздуха.
Кроме того,
опыт был поставлен
в анаэробных
условиях, т.е.
без доступа
кислорода. В
этих условиях
удалось выделить
бактерию, вызывающую
масляно-кислое
брожение, хорошо
фиксирующую
атмосферный
азот, - клостридиум
пастерианум.
Свое
видовое название
бактерия получила
в честь Пастера,
а родовое - от
латинского
слова "клострум"
- веретено.
Клостридиум
является сравнительно
крупной палочкой,
в 3 - 4 мкм длины,
дающей споры.
Во время спорообразования
клетка клостридиума
вздувается
в виде веретена.
Клостридиум
имеет жгутики,
расположенные
по всей
поверхности
тела, и может
сравнительно
быстро перемещаться.
В лабораторных
условиях клостридиум
фиксирует
атмосферный
азот, хотя и в
небольших, но
заметных количествах
от 1 до 5 мг азота
на 1 г использованного
сахара. Клостридиум
- очень широко
распространенная
бактерия,
встречающаяся
в самых разнообразных
почвах - кислых,
нейтральных
и щелочных.
Азотобактер.
Другой азотфиксирующей
бактерией
является азотобактер,
открытый в 1901
г. Азотобактер
в отличие от
клостридиума
- форма аэробная,
развивающаяся
при широком
доступе кислорода.
Азотобактер
имеет характерную
форму удлиненного
кокка, делящегося
не путем появления
поперечной
перегородки,
а перетяжкой
(Рис.60). Клетки
азотобактера
довольно крупные.
Размер их колеблется
от 1 до 10 мкм. Клетки
окружает слизистая
капсула. Форма
азотобактера
не остается
без изменения.
В молодом возрасте
он имеет форму
очень толстой
палочки, затем
эллиптическую,
а часто и совсем
округлую форму.
Фиксация азота
азотобактером
более интенсивна,
чем у клостридиума,
а именно от 2
до 12 и даже до
20 мг азота на
1 г сахара. Азотобактер
очень чувствителен
к реакции среды.
Оптимум для
его развития
будет при рН
= 7,0 или 7,2, максимум
- при рН = 9,0. В почвах,
имеющих рН ниже
5,6, он обычно не
встречается.
Механизм
фиксации азота
не может считаться
до сего времени
полностью
выясненным.
Наиболее вероятное
предположение
заключается
в том, что водород
при брожении
у клостридиума
и при дыхании
у азотобактера
выделяется
не в молекулярном
(Нг) виде, а в форме
атомного водорода
(2Н). Вот этот-то
активный атомный
водород и способен
связывать
молекулярный
азот атмосферы
в виде аммиака.
В последнее
время, применяя
тяжелый азот
(l5N2),
удалось
показать значительную
достоверность
этой точки
зрения.
Установлено,
что многие
сине-зеленые
водоросли также
фиксируют
атмосферный
азот.
Азотобактерин.
Существует
препарат азотобактера
для заражения
семян, названный
азотобактерином.
Азотобактерин
готовится на
аграрной среде
в бутылках. Для
заражения
порции семян
на 1 га требуется
этого препарата
всего 10 - 15 г. Многочисленные
опыты дали
очень неустойчивые
результаты
при применении
азотобактерина.
Лучше всего
на азотобактерин
реагируют
некоторые
овощные культуры.
Величины
фиксации азота
бактериями.
Фиксация азота
азотфиксирующими
бактериями
достигает
значительных
величин. Клевер
за счет бактерий
накапливает
ежегодно в
среднем 150-160 кг
азота на 1 га,
люцерна - около
300 кг, люпин - до
160 кг. Однолетние
бобовые фиксируют
значительно
меньшие количества
азота. Так, например,
соя фиксирует
из воздуха в
год около 100, вика
- 80, горох - около
60, фасоль - около
70 кг.
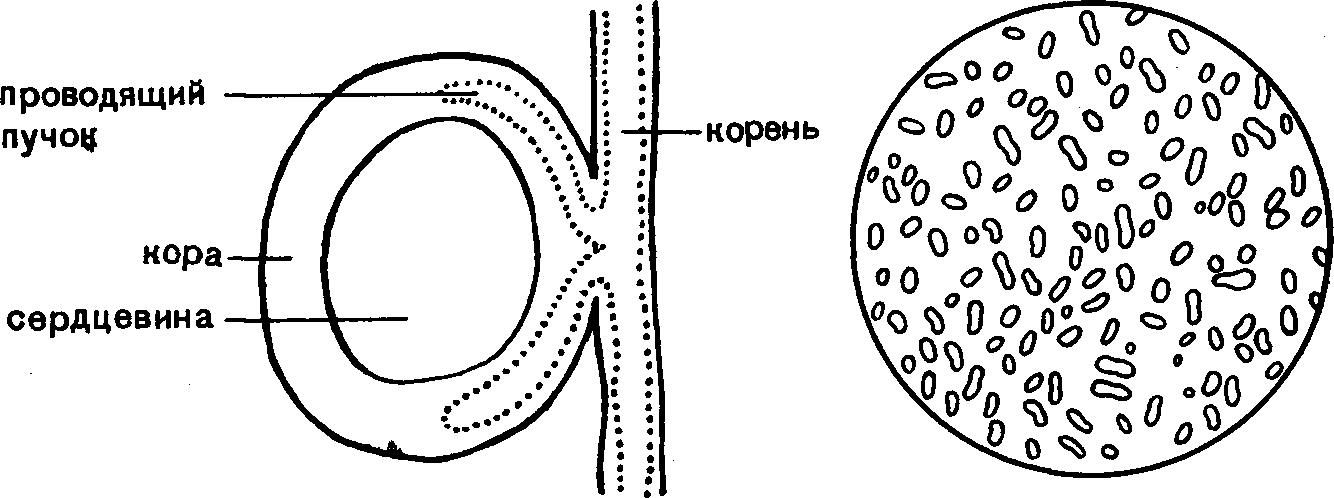
Бактерии в
почве и их роль
в круговороте
веществ в природе
Число
бактерий в
почве. В
почве содержится
огромное число
бактерий. Раньше
их число измерялось
сотнями тысяч
на один грамм
почвы. С.Н. Виноградский
(1924) разработал
метод непосредственного
микроскопического
подсчета бактерий
в почве путем
их окраски.
После этого
стало ясно, что
число бактерий
измеряется
сотнями миллионов
в 1 г. В бедных
тундровых или
песчаных почвах
пустыни их
насчитывается
до J500 миллионов,
в слабоподзолистых
почвах - до одного
миллиарда, а
в богатых
органическим
веществом
(чернозем) - до
двух миллиардов
и выше.
Два миллиарда
бактерий в 1 г
почвы составляют
около 3% сухой
массы почвы.
Такое большое
число бактерий
позволяет
считать, что
большинство
процессов,
происходящих
в почве, носит
биологический
характер, т.е.
связано с
жизнедеятельностью
бактерий.
Если бы
процесс накопления
азота, так же
как и углерода,
шел только в
одну сторону,
то жизнь стала
бы скоро на
Земле невозможной
из-за обилия
неразложившихся
органических
остатков. Мы
уже знаем, что
жизнедеятельность
бактерий является
причиной разложения
белковых веществ.
Разложение
белков бактериями.
Бактерии,
разлагающие
белковые вещества
на более простые
составные
части, называются
гнилостными
бактериями
или аммонификаторами,
так как в результате
разложения
белков в среде
накапливается
аммиак. Разлагая
сложные белковые
вещества на
простые минеральные
соединения,
бактерии сами
питаются продуктами
разложения
и размножаются.
Однако образуемая
ими масса тел
составляет
лишь ничтожную
долю от разложившегося
вещества. В
этой минерализующей
деятельности
и заключается
та огромная
полезная роль
гнилостных
бактерий, которую
они играют в
природе.
Процесс
гниения протекает
как в анаэробных,
так и в аэробных
условиях. Особенно
быстро он проходит
в аэробных
условиях.
В факультативно-анаэробных
условиях гниение
белков осуществляется
целым рядом
бактерий. Из
них можно отметить
кишечную палочку
и протея.
В аэробных
условиях разложение
белков производит
сенная палочка
и другие спорообразующие
формы. Из
неспоро-образующих
форм можно
упомянуть
небольшую
палочку (1-2 мкм)
- псевдомонас.
При гниении
образуются
вода, углекислый
газ, аммиак,
сероводород,
метилмеркаптан
(CH3SH).
Очень характерными
продуктами
анаэробного
расщепления
белков являются
дурно пахнущие
продукты индол
и скатол, возникающие
в результате
частичного
разрушения
аминокислоты
триптофана
в анаэробных
условиях.
Высушенное
белковое вещество
не разлагается
бактериями
и может сохраняться
очень долго.
Сушеное или
прокопченное
мясо, сухой
яичный порошок
не портятся,
если их хранить
в сухом
месте.
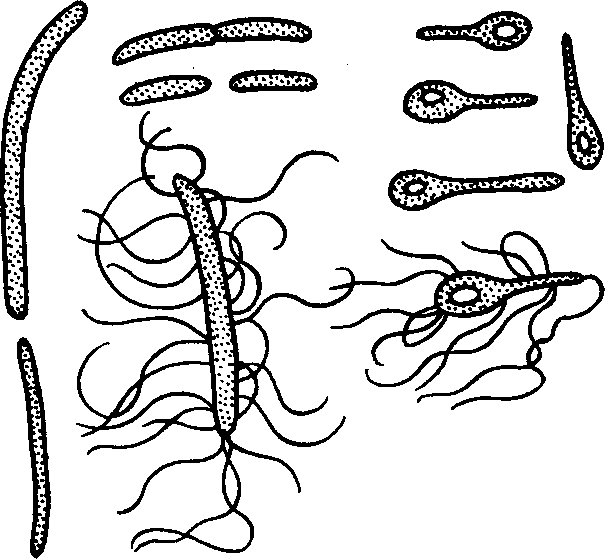
Разложение
мочевины. Одной
из специальных
групп аммонификаторов
являются бактерии,
разлагающие
мочевину. Мочевина
- главная составная
часть мочи
человека и
большинства
животных. Человек
выделяет бактерии,
разлагающие
в день от 30 до
50 г мочевины.
Под влиянием
бактерий мочевина
разлагается,
образуется
карбонат аммония.
Последний
быстро распадается
на воду, аммиак
и углекислый
газ.
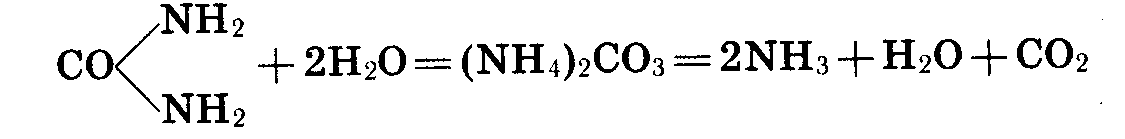
Процесс
нитрификации.
Образовавшийся
в результате
аммонификации
аммиак или
используется
высшими растениями,
или нитрифицируется.
Процесс нитрификации
заключается
в окислении
аммиака до
азотной кислоты.
Первая фаза
нитрификации
вызывается
микробом, окисляющим
аммиак до азотистой
кислоты. Он
получил название
нитрозомонас.
Вторая фаза
вызывается
бактерией
нитробактер,
окисляющей
азотистую
кислоту до
азотной. В почве
азотистая
кислота не
накапливается,
так как обе эти
бактерии встречаются
всегда вместе,
находясь в
своеобразном
симбиозе.
Нитрозомонас
представляет
собой снабженную
жгутиком шарообразную
бактерию, а
нитробактер
неподвижен
и является
маленькой
палочкой. На
первом этапе
нитрификации
выделяется
больше энергии,
чем на втором.

В
первой фазе
нитрификации
выделяется
663,6 Дж (или 158 кал):
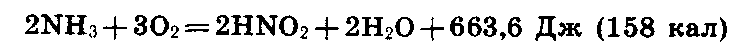
Во
второй фазе
нитрификации
энергии освобождается
значительно
меньше:
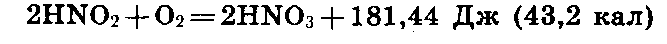
Нитрификаторы
синтезируют
органическое
вещество путем
хемосинтеза
за счет энергии
окисления
аммиака в азотистую
кислоту, а азотистой
кислоты в азотную.
Нитрификаторы,
так же как и
зеленые растения,
используют
для питания
углекислый
газ.
С.Н.
Виноградский
обнаружил очень
высокую чувствительность
нитрификаторов
к органическому
веществу, которое
действует на
них как яд, причем
нитрозомонас
более чувствителен
к органическому
веществу, чем
нитробактер.
Малые концентрации
органического
вещества задерживают
рост бактерий,
а несколько
большие окончательно
его останавливают.
Нитрификация
в почве. Нитрификация
в почве идет
несколько
отлично от
нитрификации
в лабораторной
обстановке.
В первую очередь
это касается
влияния на этот
процесс органического
вещества. Если
в лабораторной
обстановке
нитрификаторы
проявляют очень
высокую чувствительность
к органическому
веществу и в
его присутствии
не растут, то
в природной
обстановке
наблюдается
как раз обратная
картина. Наличие
органического
вещества способствует
процессу
нитрификации,
так как является
источником
образования
аммиака.
Процесс
денитрификации.
С круговоротом
азота в природе
связан также
и процесс
денитрификации,
обратный по
своей сути
процессу
азотфиксации.
Денитрификацией
называется
процесс восстановления
нитратов до
свободного
азота.
Процесс
денитрификации,
в отличие от
нитрификации
и азотфиксации,
вызывается
целым рядом
малоспецифических
микроорганизмов,
относящихся
к неспороносным
палочкам.
Денитрифицирующие
бактерии являются
факультативными
анаэробами.
В условиях
широкого доступа
кислорода они
денитрификации
не производят.
Стоит им, однако,
попасть в анаэробные
условия, как
при наличии
нитратов и
доступного
им органического
вещества начинается
процесс денитрификации.
При нехватке
кислорода
микроорганизмы
начинают отнимать
его от нитратов,
восстанавливая
их. Одновременно
при этом окисляется
усвояемое ими
органическое
вещество - сахара
или соли органических
кислот. Наилучшими
условиями для
протекания
процесса
денитрификации
являются анаэробные
условия, наличие
нитратов и
подходящего
для микроорганизмов
органического
вещества.
Круговорот
азота в природе.
Подведем итоги
по круговороту
азота в природе.
Высшее растение
синтезирует
белок в своем
теле из связанного
минерального
азота и углеводов.
Растения поедаются
животными,
которые сами
не в состоянии
синтезировать
белки из углеводов
и минерального
азота. Отмирая,
животные и
растения становятся
пищей гнилостных
бактерий, разлагающих
белки до аммиака,
эти же бактерии
разлагают и
белки, находящиеся
в навозе. Аммиак
усваивается
растением или
нитрифицируется.
Азотфиксаторы
связывают
атмосферный
азот и переводят
опять в белковый,
который в дальнейшем
может разлагаться
гнилостными
бактериями.
Здесь следует
еще упомянуть
о связывании
азота электрическими
разрядами в
атмосфере,
который в виде
азотной кислоты
с дождем попадает
в почву. Так
происходит
круговорот
азота в природе;
он переходит
из одной формы
в другую, подтверждая
великий закон
природы - закон
сохранения
вещества, открытый
М.В. Ломоносовым.