Основные проблемы генетики и механизм воспроизводства жизни
самцов
частота проявления
рецессивного
признака выше,
чем у самок,
наводил на
мысль, что
рецессивный
аллель, определяющий
белоглазость,
находится в
X- хромосоме,
а Y- хромосома
лишена гена
окраски глаз.
Чтобы проверить
эту гипотезу,
Морган скрестил
исходного
белоглазого
самца с красноглазой
самкой из F
1.
В потомстве
были получены
красноглазые
и белоглазые
самцы и самки.
Из этого Морган
справедливо
заключил, что
только X- хромосома
несет ген окраски
глаз. В Y –хромосоме
соответствующего
локуса вообще
нет. Это явление
известно под
названием
наследования,
сцепленного
с полом.
2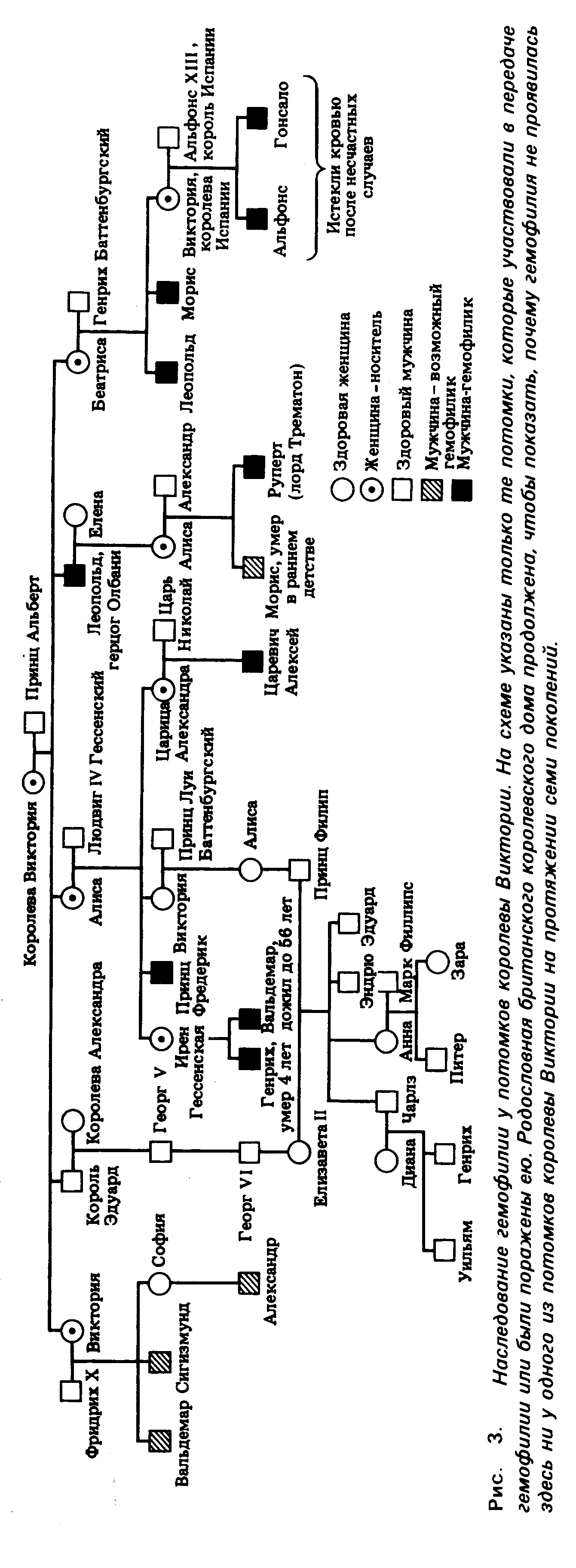 .8.
Наследование,
сцепленное
с полом
.8.
Наследование,
сцепленное
с полом
Гены,
находящиеся
в половых
хромосомах,
называют сцепленными
с полом. В X-хромосоме
имеется участок,
для которого
в Y- хромосоме
нет гомолога.
Поэтому у особей
мужского пола
признаки,
определяемые
генами этого
участка, проявляются
даже в том случае,
если они рецессивны.
Это особая
форма сцепления
позволяет
объяснить
наследование
признаков,
сцепленных
с полом, например,
цветовой слепоты.
Раннего облысения
и гемофилии
у человека.
Гемофилия –
сцепленный
с полом рецессивный
признак, при
котором нарушается
образование
фактора VIII, ускоряющего
свертывание
крови. Ген,
детерминирующий
синтез фактора
VIII, находится
в участке X-
хромосомы, не
имеющем гомолога,
и представлен
двумя аллелями
– доминантным
и рецессивным
мутантным.
Один
из наиболее
хорошо документированных
примеров
наследования
гемофилии мы
находим в
родословной
потомков английской
королевы Виктории.
Предполагают,
что
 ген
гемофилии
возник в результате
мутации у самой
королевы Виктории
или у одного
из ее родителей
(рис. 5).
ген
гемофилии
возник в результате
мутации у самой
королевы Виктории
или у одного
из ее родителей
(рис. 5).
Возможны
следующие
генотипы и
фенотипы:
|
Генотип
|
Фенотип
|
|
XHXH
|
Нормальная
женщина |
|
XHXh
|
Нормальная
женщина (носитель) |
|
XHY
|
Нормальный
мужчина |
|
XhY
|
Мужчина
– гемофилик |
|
XhXh
|
Женщина
– гемофилик.
Редчайший
случай, возможный
лишь в случае,
если отец –
гемофилик,
а мать – гемофиличка
или носитель.
|
2.9.
Неполное
доминирование
Известны
случаи, когда
два или более
аллелей не
проявляют в
полной мере
доминантность
или рецессивность,
так что в гетерозиготном
состоянии ни
один из аллелей
не доминирует
над другим.
Это явление
неполного
доминирования,
или кодоминантность,
представляет
собой исключение
из описанного
Менделем правила
наследования
при моногибридных
скрещиваниях.
К счастью, Мендель
выбрал для
своих экспериментов
признаки, которым
не свойственно
неполное
доминирование;
в противном
же случае оно
могло бы сильно
осложнить его
первые исследования.
Неполное
доминирование
наблюдается
как у растений,
так и у животных.
В большинстве
случаев гетерозиготы
обладают фенотипом,
промежуточным
между фенотипами
доминантной
и рецессивной
гомозигот.
2.10. Изменчивость
Изменчивостью
называют всю
совокупность
различий по
тому или иному
признаку между
организмами,
принадлежащими
к одной и той
же природной
популяции или
виду. Поразительное
морфологическое
разнообразие
особей в пределах
любого вида
привлекло
внимание Дарвина
и Уоллеса во
время их путешествий.
Закономерный,
предсказуемый
характер передачи
таких различий
по наследству
послужил основой
для исследований
Менделя. Дарвин
установил, что
определенные
признаки могут
развиваться
в результате
отбора, тогда
как Мендель
объяснил механизм,
обеспечивающий
передачу из
поколения в
поколение
признаков, по
которым ведется
отбор.
Мендель
описал, каким
образом наследственные
факторы определяют
генотип организма,
который в процессе
развития проявляется
в структурных,
физиологических
и биохимических
особенностях
фенотипа. Если
фенотипическое
проявление
любого признака
обусловлено
в конечном
счете генами,
контролирующими
этот признак,
то на степень
развития
определенных
признаков
может оказывать
влияние среда.
И зучение
фенотипических
различий в
любой большой
популяции
показывает,
что существует
две формы
изменчивости
– дискретная
и непрерывная.
Для изучения
изменчивости
какого-либо
признака, например,
роста у человека,
необходимо
измерить этот
признак у большого
числа индивидуумов
в изучаемой
популяции. На
представлены
типичные результаты,
получаемые
при таких
исследованиях,
и они наглядно
демонстрируют
различие между
дискретной
и непрерывной
изменчивостью.
зучение
фенотипических
различий в
любой большой
популяции
показывает,
что существует
две формы
изменчивости
– дискретная
и непрерывная.
Для изучения
изменчивости
какого-либо
признака, например,
роста у человека,
необходимо
измерить этот
признак у большого
числа индивидуумов
в изучаемой
популяции. На
представлены
типичные результаты,
получаемые
при таких
исследованиях,
и они наглядно
демонстрируют
различие между
дискретной
и непрерывной
изменчивостью.
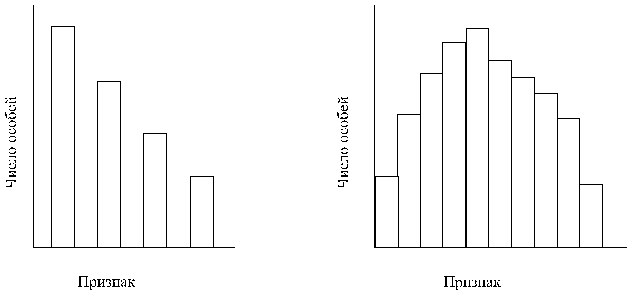
 А
Б
А
Б
Рис. 7. Гистограммы,
отражающие
распределение
частот в случае
прерывистой
(А) и непрерывистой
(Б) изменчивости.
2.11.
Влияние среды
Главный
фактор, детерминирующий
любой фенотипический
признак, - это
генотип. Генотип
организма
определяется
в момент оплодотворения,
но степень
последующей
экспрессии
этого генетического
потенциала
в значительной
мере зависит
от внешних
факторов,
воздействующих
на организм
во время его
развития. Так,
например,
использованный
Менделем сорт
гороха с длинным
стеблем обычно
достигал высоты
180 см. Однако для
этого ему необходимы
были соответствующие
условия – освещение,
снабжение
водой и хорошая
почва. При
отсутствии
оптимальных
условий (при
наличии лимитирующихфакторов) ген
высокого стебля
не мог в полной
мере проявить
свое действие.
Эффект взаимодействия
генотипа и
факторов среды
продемонстрировал
датский генетик
Иоганнсен. В
ряде экспериментов
на карликовой
фасоли он выбирал
из каждого
поколения
самоопылявшихся
растений самые
тяжелые и самые
легкие семена
и высаживал
их для получения
следующего
поколения.
Повторяя эти
эксперименты
на протяжении
нескольких
лет, он обнаружил,
что в пределах
«тяжелой» или
«легкой»
селекционной
линии семена
мало отличались
по среднему
весу, тогда
как средний
вес семян из
разных линий
сильно различался.
Это позволяет
считать, что
на фенотипическое
проявление
признака оказывает
влияние как
наследственность,
так и среда.
На основании
этих результатов
можно определить
непрерывную
фенотипическую
изменчивость
как «кумулятивный
эффект варьирующих
факторов среды,
воздействующих
на вариабельный
генотип». Кроме
того, эти результаты
показывают,
что степень
наследуемости
данного признака
определяется
в первую очередь
генотипом. Что
касается развития
таких чисто
человеческих
качеств, как
индивидуальность,
темперамент
и интеллект,
то, судя по
имеющимся
данным, они
зависят как
от наследственных,
так и от средовых
факторов, которые,
взаимодействуя
в различной
степени у различных
индивидуумов,
создают фенотипические
различия между
индивидуумами.
Мы пока еще не
располагаем
данными, которые
твердо указывали
бы на то, что
влияние каких-то
из этих факторов
всегда преобладает,
однако среда
никогда не
может вывести
фенотип за
пределы, детерминированные
генотипом.
2.12. Источники
изменчивости
Необходимо
ясно себе
представлять,
что взаимодействие
между дискретной
и непрерывной
изменчивостью
и средой делает
возможным
существование
двух организмов
с идентичным
фенотипом.
Механизм репликации
ДНК при митозе
столь близок
к совершенству,
что возможности
генетической
изменчивости
у организмов
с бесполым
размножением
очень малы.
Поэтому любая
видимая изменчивость
у таких организмов
обусловлена
воздействиями
внешней среды.
Что же касается
организмов,
размножающихся
половым путем,
то у них есть
широкие возможности
для возникновения
генетических
различий.
Практически
неограниченными
источниками
генетической
изменчивости
служат два
процесса,
происходящие
во время мейоза:
Реципкорный
обмен между
хроматидами
гомологичных
хромосом, который
может происходить
в профазе 1 мейоза.
Он создает
новые группы
сцепления,
т.е. служит важным
источником
генетической
рекомбинации
аллелей.
Ориентация
пар гомологичных
хромосом
(бивалентов)
в экваториальной
плоскости
веретена в
метафазе I
мейоза определяет
направление,
в котором каждый
член пары будет
перемещаться
в анафазе I.
Эта операция
носит случайный
характер. Во
время метафазы
II пары хроматид
опять-таки
ориентируются
случайным
образом, и этим
определяется,
к какому из
двух противоположных
полюсов направится
та или иная
хромосома во
время анафазыII.
Случайная
ориентация
и последующее
независимое
расхождение
(сегрегация)
хромосом делают
возможным
большое число
различных
хромосомных
комбинаций
в гаметах; число
это можно
подсчитать.
Третий
источник
изменчивости
при половом
размножении
– это то, что
слияние мужских
и женских гамет,
приводящее
к объединению
двух гаплоидных
наборов хромосом
в диплоидном
ядре зиготы,
происходит
совершенно
случайным
образом (во
всяком случае,
в теории); любая
мужская гамета
потенциально
способна слиться
с любой женской
гаметой.
Эти
три источника
генетической
изменчивости
и обеспечивают
постоянную
«перетасовку»
генов, лежащую
в основе все
время происходящих
генетических
изменений.
Среда оказывает
воздействие
на весь ряд
получающихся
таким образом
фенотипов, и
те из них, которые
лучше всего
приспособлены
к данной среде,
преуспевают.
Это ведет к
изменениям
частот аллелей
и генотипов
в популяции.
Однако эти
источники
изменчивости
не порождают
крупных изменений
в генотипе,
которые необходимы,
согласно
эволюционной
теории, для
возникновения
новых видов.
Такие изменения
возникают в
результате
мутаций.
2.13.
Мутации
Мутацией
называют изменение
количества
или структуры
ДНК данного
организма.
Мутация приводит
к изменению
генотипа, которое
может быть
унаследовано
клетками,
происходящими
от мутантной
клетки в результате
митоза или
мейоза. Мутирование
может вызывать
изменения
каких-либо
признаков в
популяции.
Мутации, возникшие
в половых клетках,
передаются
следующим
поколениям
организмов,
тогда как мутации,
возникшие в
соматических
клетках, наследуются
только дочерними
клетками,
образовавшимися
путем митоза
и такие мутации
называют
соматическими.
Мутации,
возникающие
в результате
изменения
числа или
макроструктуры
хромосом, известны
под названием
хромосомных
мутаций или
хромосомных
аберраций
(перестроек).
Иногда хромосомы
так сильно
изменяются,
что это можно
увидеть под
микроскопом.
Но термин «мутация»
используют
главным образом
для обозначения
изменения
структуры ДНК
в одном локусе,
когда происходит
так называемая
генная, или
точечная, мутация.
Представление
о мутации как
о причине внезапного
появления
нового признака
было впервые
выдвинуто в
1901 г. голландским
ботаником Гуго
де Фризом, изучавшим
наследственность
у энотеры Oenothera
lamarckiana. Спустя
9 лет Т.Морган
начал изучать
мутации у дрозофилы,
и вскоре при
участии генетиков
всего мира у
нее было идентифицировано
более 500 мутаций.
2.14. Генные
мутации
Внезапные
спонтанные
изменения
фенотипа, которые
нельзя связать
с обычными
генетическими
явлениями или
микроскопическими
данными о наличии
хромосомных
аберраций,
можно объяснить
только изменениями
в структуре
отдельных
генов. Генная,
или точечная
(поскольку
она относится
к определенному
генному локусу),
мутация – результат
изменения
нуклеотидной
последовательности
молекулы ДНК
в определенном
участке хромосомы.
Такое изменение
последовательности
оснований в
данном гене
воспроизводится
при транскрипции
в структуре
иРНК и приводит
к изменению
последовательности
аминокислот
в полипептидной
цепи, образующейся
в результате
трансляции
на рибосомах.
Существуют
различные типы
генных мутаций,
связанных с
добавлением,
выпадением
или перестановкой
оснований в
гене. Это дупликации,
вставки, делеции,
инверсии или
замены оснований.
Во всех случаях
они приводят
к изменению
нуклеотидной
последовательности,
а часто – и к
образованию
измененного
полипептида.
Например, делеция
вызывает сдвиг
рамки.
Генные
мутации, возникающие
в гаметах или
в будущих половых
клетках, передаются
всем клеткам
потомков и
могут влиять
на дальнейшую
судьбу популяции.
Соматические
генные мутации,
происходящие
в организме,
наследуются
только теми
клетками, которые
образуются
из мутантной
клетки путем
митоза. Они
могут оказать
воздействие
на тот организм,
в котором они
возникли, но
со смертью
особи исчезают
из генофонда
популяции.
Соматические
мутации, вероятно,
возникают
очень часто
и остаются
незамеченными,
но в некоторых
случаях при
этом образуются
клетки с повышенной
скоростью
роста и деления.
Эти клетки
могут дать
начало опухолям
– либо доброкачественным,
которые не
оказывают
особого влияния
на весь организм,
либо злокачественным,
что приводит
к раковым
заболеваниям.
Эффекты
генных мутаций
чрезвычайно
разнообразны.
Большая часть
мелких генных
мутаций фенотипически
не проявляется,
поскольку они
рецессивны,
однако известен
ряд случаев,
когда изменение
всего лишь
одного основания
в определенном
гене оказывает
глубокое влияние
на фенотип.
Одним из примеров
служит серповидноклеточная
анемия – заболевание,
вызываемое
у человека
заменой основания
в одном из генов,
ответственных
за синтез
гемоглобина.
Молекула
дыхательного
пигмента гемоглобина
у взрослого
человека состоит
из четырех
полипептидных
цепей (двух -
и двух –
цепей), к которым
присоединены
четыре простетические
группы гема.
От структуры
полипептидных
цепей зависит
способность
молекулы
гемоглобина
переносить
кислород. Изменение
последовательности
оснований в
триплете, кодирующем
одну определенную
аминокислоту
из 146, входящих
в состав -
цепей, приводит
к синтезу
аномального
гемоглобина
серповидных
клеток (HbS).
Последовательности
аминокислот
в нормальных
и аномальных
-цепях
различаются
тем, что в одной
точке аномальных
цепей гемоглобина
S глутамидовая
кислота замещена
валином.В
результате
такого, казалось
бы, незначительного
изменения
гемоглобин
S кристаллизуется
при низких
концентрациях
кислорода, а
это в свою очередь
приводит к
тому, что в венозной
крови эритроциты
с таким гемоглобином
деформируются
(из округлых
становятся
серповидными)
и быстро разрушаются.
Физиологический
эффект мутации
состоит в развитии
острой анемии
и снижении
количества
кислорода,
переносимого
кровью. Анемия
не только вызывает
физическую
слабость, но
и может привести
к нарушениям
деятельности
сердца и почек
и к ранней смерти
людей, гомозиготных
по мутантному
аллелю. В гетерозиготном
состоянии этот
аллель вызывает
значительно
меньший эффект:
эритроциты
выглядят
нормальными,
а аномальный
гемоглобин
составляет
только около
40 %. У гетерозигот
развивается
анемия лишь
в слабой форме,
а зато в тех
областях, где
широко распространена
малярия, особенно
в Африке и Азии,
носители аллеля
серповидноклеточности
невосприимчивы
к этой болезни.
Это объясняется
тем, что ее
возбудитель
- малярийный
плазмодий -
не может жить
в эритроцитах,
содержащих
аномальный
гемоглобин.
2.15. Летальные
мутацииИзвестны
случаи, когда
один ген может
оказывать
влияние на
несколько
признаков, в
том числе и на
жизнеспособность.
Летальные
мутации вызывают
такие изменения
в развитии,
которые несовместимы
с жизнедеятельностью.
Доминантные
летальные гены
трудны для
изучения, и
сведения о них
ограничены.
Напротив, гены
с рецессивным
летальным
действием
изучены гораздо
лучше. Известно
множество
рецессивных
мутаций у различных
организмов,
которые никак
себя не проявляют
фенотипически.
Существует
также очень
много доминантных
мутаций, имеющих
в гетерозиготном
состоянии
четко отличающийся
фенотип, которые
в гомозиготном
состоянии
вызывают летальный
эффект. Фаза
летального
действия, т.е.
время, когда
мутантный ген
реализуется,
существенно
варьирует: от
самых первых
этапов эмбрионального
развития до
периода полового
созревания.
В некоторых
случаях летальные
гены могут
иметь более
одной фазы
летального
действия. Это
означает, что
ген или его
продукты могут
иметь несколько
раз активно
работать и
использоваться
в ходе онтогенеза.
Летальный
эффект одних
мутантных
генов проявляется
всегда, другие
показывают
существенную
зависимость
от условий
среды. У человека
и у других
млекопитающих
определенный
рецессивный
ген вызывает
образование
внутренних
спаек легких,
что приводит
к смерти при
рождении. Другим
примером служит
ген, который
влияет на
формирование
хряща и вызывает
врожденные
уродства, ведущие
к смерти новорожденного.
Воздействие
летального
гена ясно видно
на примере
наследования
окраски шерсти
у мышей. У диких
мышей шерсть
обычно серая,
типа агути; но
у некоторых
мышей шерсть
желтая. При
скрещивании
между желтыми
мышами в потомстве
получаются
как желтые
мыши, так и агути
в отношении
2:1. Единственное
возможное
объяснение
таких результатов
состоит в том,
что желтая
окраска шерсти
доминирует
над агути, и
что все желтые
мыши гетерозиготны.
Атипичное
менделевское
отношение
объясняется
гибелью гомозиготных
желтых мышей
до рождения.
При вскрытии
беременных
желтых мышей,
скрещенных
с желтыми же
мышами, в их
матках были
обнаружены
мертвые желтые
мышата. Если
же скрещивались
желтые мыши
и агути, то в
матках беременных
самок не оказывалось
желтых мышат,
поскольку при
таком скрещивании
не может быть
потомства,
гомозиготного
по гену желтой
шерсти.
Мутации,
характеризующиеся
в гомозиготном
состоянии
летальным
эффектом, далеко
не всегда
фенотипически
проявляются
у гетерозигот.
К их числу относится
комплекс рецессивных
t- мутаций
у мышей, локализованных
в аутосоме.
Одной из самых
ранних мутаций
у млекопитающих,
является мутация
t12, вызывающая
гибель гомозигот
уже на стадии
морулы (~20-30 клеток).
Гетерозиготные
животные
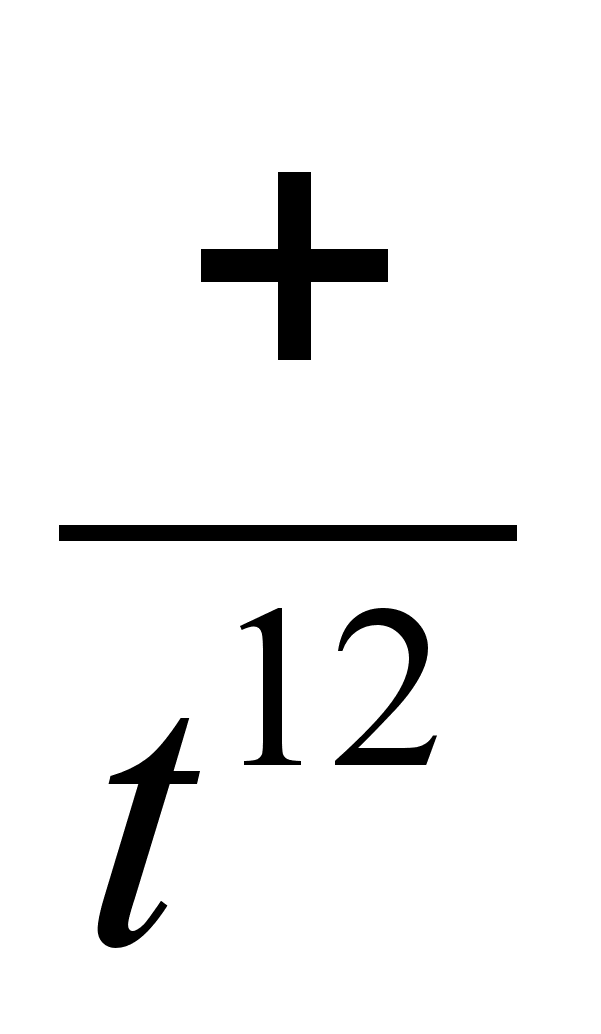 имеют нормальный
фенотип и
жизнеспособность.
имеют нормальный
фенотип и
жизнеспособность.
Летальные
мутации обнаруживаются
не только у
животных. Наглядный
пример, иллюстрирующий
летальное
действие генов
у растений, -
явление хлорофильных
мутаций. У
гомозиготных
по хлорофильной
мутации растений
нарушен синтез
молекулы хлорофилла.
Такие растения
развиваются
до тех пор, пока
запасы питательных
веществ в семени
не иссякают,
поскольку они
не способны
к фотосинтезу.
2.16. Значение
мутаций
Хромосомные
и генные мутации
оказывают
разнообразные
воздействия
на организм.
Во многих случаях
эти мутации
летальны, так
как нарушают
развитие; у
человека, например,
около 20 % беременностей
заканчиваются
естественным
выкидышем в
сроки до 12 недель,
и в половине
таких случаев
можно обнаружить
хромосомные
аномалии. В
результате
некоторых
хромосомных
мутаций определенные
гены могут
оказаться
вместе, и их
общий эффект
может привести
к появлению
какого-либо
«благоприятного»
признака. Кроме
того, сближение
некоторых
генов друг с
другом делает
менее вероятным
их разделение
в результате
кроссинговера,
а в случае
благоприятных
генов это создает
преимущество.
Генная мутация
может привести
к тому. Что в
определенном
локусе окажется
несколько
аллелей. Это
увеличивает
как гетерозиготность
данной популяции,
так и ее генофонд,
и ведет к усилению
внутрипопуляционной
изменчивости.
Перетасовка
генов как результат
кроссинговера,
независимого
распределения,
случайного
оплодотворения
и мутаций может
повысить непрерывную
изменчивость,
но ее эволюционная
роль часто
оказывается
преходящей,
так как возникающие
при этом изменения
могут быстро
сгладиться
вследствие
«усреднения».
Что же касается
генных мутаций,
то некоторые
из них увеличивают
дискретную
изменчивость,
и это может
оказать на
популяцию
более глубокое
влияние. Большинство
генных мутаций
рецессивны
по отношению
к «нормальному»
аллелю, который,
успешно выдержав
отбор на протяжении
многих поколений,
достиг генетического
равновесия
с остальным
генотипом.
Будучи рецессивными,
мутантные
аллели могут
оставаться
в популяции
в течение многих
поколений,
пока им не удастся
встретиться,
т.е. оказаться
в гомозиготном
состоянии и
проявиться
в фенотипе.
Время от времени
могут возникать
и доминантные
мутантные
аллели, которые
немедленно
дают фенотипический
эффект.
Глава 3. Современные
возможности
и задачи генетики
и генной инженерии
3.1.
Химеры
Широкие
возможности
глубже понять
роль генов в
дифференцировке
клеток и в регуляции
взаимодействий
между клетками
в процессе
развития дают
химерные и
трансгенные
животные. Развитие
экспериментальных
методов в последнее
время сделало
возможным
получать совершенно
необычных
животных, которые
несут гены не
только одного
отца и одной
матери, но и
большего количества
предков.
Химерные
животные – это
генетические
мозаики, образующиеся
в результате
объединения
бластомеров
от эмбрионов
с разными
генотипами.
Получение
таких эмбрионов
осуществляется
во многих
лабораториях.
Принцип получения
химер сводится
главным образом
к выделению
двух или большего
числа ранних
зародышей и
их слиянию. В
том случае,
когда в генотипе
зародышей,
использованных
для создания
химеры есть
отличия по
ряду характеристик,
удается проследить
судьбу клеток
обоих видов.
С помощью химерных
мышей был, например,
решен вопрос
о способе
возникновения
в ходе развития
многоядерных
клеток попречнополосатых
мышц. Изучение
химерных животных
позволило
решить немало
трудных вопросов,
и в будущем
благодаря
применению
этого метода
появится возможность
решать сложные
вопросы генетики
и эмбриологии.
3.2.
Трансгенные
организмы
Развитие
генной инженерии
создало принципиально
новую основу
для конструирования
последовательностей
ДНК, нужную
исследователям.
Успехи в области
экспериментальной
биологии позволили
создать методы
введения таких
искусственно
созданных
генов в ядра
яйцеклеток
или сперматозоидов.
В результате
возникла возможность
получения
трансгенных
животных, т.е.
животных, несущих
в своем организме
чужеродные
гены.
Одним
из первых примеров
успешного
создания трансгенных
животных было
получение
мышей, в геном
которых был
встроен гормон
гена роста
крысы. Некоторые
из таких трансгенных
мышей росли
быстро и достигали
размеров,
существенно
превышавших
контрольных
животных.
Первая в мире
обезьяна с
измененным
генетическим
кодом появилась
на свет в Америке.
Самец по кличке
Энди родился
после того,
как в яйцеклетку
его матери был
внедрен ген
медузы. Опыт
проводился
с макакой-резусом,
которая гораздо
ближе по своим
биологическим
признакам к
человеку, чем
любые другие
животные, до
сих пор подвергавшиеся
экспериментам
по генетической
модификации.
Ученые говорят,
что применение
этого метода
поможет им при
разработке
новых способов
лечения таких
болезней, как
рак груди и
диабет. Однако,
как сообщает
ВВС, этот эксперимент
уже вызвал
критику со
стороны организаций
по защите животных,
которые опасаются,
что эти исследования
приведут к
страданиям
множества
приматов в
лабораториях.
Создание
гибрида человека
и свиньи. Из
человеческой
клетки извлекается
ядро и имплантируется
в ядро яйцеклетки
свиньи, которую
предварительно
освободили
от генетического
материала
животного. В
результате
получился
эмбрион, который
прожил 32 дня,
пока ученые
не решили его
уничтожить.
Исследования
проводятся
как всегда
ради благородной
цели: поиска
лекарств от
заболеваний
человека. Несмотря
на то, что попытки
клонировать
человеческие
существа не
одобряются
многими учеными
и даже теми,
кто создал
овечку Долли,
подобные
эксперименты
будет трудно
остановить,
так как принцип
техники клонирования
уже известен
многим лабораториям.
В
настоящее
время интерес
к трансгенным
животным очень
велик. Это
объясняется
двумя причинами.
Во-первых, возникли
широкие возможности
для изучения
работы чужеродного
гена в геноме
организма-хозяина,
в зависимости
от места его
встраивания
в ту или иную
хромосому, а
также строения
регуляторной
зоны гена.
Во-вторых,
трансгенные
сельскохозяйственные
животные могут
представлять
в будущем интерес
для практики.
3.3. Немного
о клонировании
Термин "клон"
происходит
от греческого
слова "klon", что
означает - веточка,
побег, черенок,
и имеет отношение
прежде всего
к вегетативному
размножению.
Клонирование
растений черенками,
почками или
клубнями в
сельском хозяйстве,
в частности
в садоводстве,
известно уже
более 4-х тыс.
лет. При вегетативном
размножении
и при клонировании
гены не распределяются
по потомкам,
как в случае
полового
размножения,
а сохраняются
в полном составе
в течение многих
поколений.
Однако у животных
есть препятствие.
По мере роста
их клеток, они
в ходе клеточной
специализации
- дифференцировки
- теряют способность
реализовывать
всю генетическую
информацию,
заложенную
в ядре. Возможность
клонирования
эмбрионов
позвоночных
впервые была
показана в
начале 50-х годов
в опытах на
амфибиях. Опыты
с ними показали,
что серийные
пересадки ядер
и культивирование
клеток in vitro в
какой-то степени
увеличивает
эту способность.
Уже в начале
90-х была решена
и проблема
клонирования
эмбриональных
клеток млекопитающих.
Реконструированные
яйцеклетки
крупных домашних
животных, коров
или овец сначала
культивируют
не in vitro, а in vivo - в
перевязанном
яйцеводе овцы
- промежуточного
(первого) реципиента.
Затем их оттуда
вымывают и
трансплантируют
в матку окончательного
(второго) реципиента
- коровы или
овцы соответственно,
где их развитие
происходит
до рождения
детеныша.
Впервые
клонированное
животное (овца
по кличке Долли)
появилось
в результате
использования
донорского
ядра клетки
молочной железы
взрослой
овцы. У этого
первого успешного
эксперимента
есть существенный
недостаток
- очень низкий
коэффициент
выхода живых
особей (0,36 %). Однако
он доказывает
возможность
полноценного
клонирования,
(или получения
копии взрослого
человека). Остаётся
лишь разрешить
технические
и этические
вопросы.
Но вернёмся
к клонированию
человека. Существует
и достаточно
изящный способ
обойти этические
проблемы. Вспомним,
что наиболее
близки к человеку
по строению
внутренних
органов, как
ни странно,
свиньи.
В марте 2000 г.
PPL Therapeutics
объявила о
том, что в их
исследовательском
центре родились
пять клонированных
поросят. Клонирование
свиньи более
сложная операция,
чем клонирование
овец или коров,
так как для
того, чтобы
поддерживать
одну беременность
необходимо
несколько
здоровых плодов.
Органы свиньи
наиболее подходят
к человеку по
размерам. Свиньи
легко размножаются
и известны
своей неприхотливостью.
Но самой большой
проблемой
остается отторжение
органа животного,
который человеческий
организм не
принимает за
свой. Именно
в этом направлении
будут развиваться
дальнейшие
исследования
ученых. Ученые
видят один из
возможных
путей решения
этой проблемы
в том, чтобы
генетически
"замаскировать"
органы животного,
для того, чтобы
человеческий
организм не
мог распознать
их как чужие.
Еще одной темой
для исследования
является попытка
"очеловечить"
генетическим
путем органы
свиньи, для
того чтобы
значительно
снизить риск
отторжения.
Для этого
предполагается
вводить человеческие
гены в хромосомы
клонируемых
свиней.
Той же задачей,
но без применения
клонирования,
занимаются
и другие институты.
Например, компания
"Imutran", расположенная
в Кембридже,
смогла получить
целое стадо
свиней, в генетическом
наборе которых
уже отсутствует
одна из ключевых
характеристик,
ответственная
за отторжение
чужеродных
тканей. Как
только будет
получена пара
мужской и женской
особи, они будут
готовы производить
на свет "генетически
чистое потомство",
с органами,
которые можно
будет использовать
для трансплантации.
Ещё один шаг
к бессмертию
- искусственное
изменение ДНК.
В июне 2000 года
и случилось
то, чего так
долго ждали
и чего некоторые
так боялись.
Появилось
сообщение, что
ученым из уже
знаменитой
своей овцой
Долли шотландской
фирмы PPL
Therapeutics удалось
получить успешные
клоны овечек
с измененной
ДНК. Шотландские
ученые смогли
осуществить
клонирование,
при котором
генетический
материал клона
был "подправлен"
с лучшую сторону.
Существует
и уже узаконенный
путь обхода
запрета на
клонирование
человека, который
называется
"терапевтическое"
клонирование
человеческих
существ. Речь
идет о создании
ранних эмбрионов
- своего рода
банка донорских
тканей для
конкретных
индивидуумов.
Для этого
используются
стволовые
клетки (упрощенно
- клетки ранних
человеческих
зародышей).
Потенциал
роста стволовых
клеток просто
фантастический
- достаточно
вспомнить, что
триллионноклеточный
организм
новорожденного
человека образуется
из одной-единственной
клетки всего
лишь за 9 месяцев!
Но еще больше
впечатляет
потенциал
дифференцировки
- одна и та же
стволовая
клетка может
трансформироваться
в любую(!) клетку
человека, будь
то нейрон головного
мозга, клетка
печени или
сердечный
миоцит. "Взрослым"
клеткам такая
трансформация
не по силам.
Но одно уникальное
свойство этих
клеток превращает
их поистине
в надежду
человечества
– они отторгаются
гораздо слабее,
чем пересаженные
целые органы,
состоящие из
уже дифференцированных
клеток. Это
означает, что
в принципе
можно выращивать
в лабораторных
условиях
предшественники
самых разных
клеток (сердечных,
нервных, печеночных,
иммунных и
др.), и затем
трансплантировать
их тяжело больным
людям вместо
донорских
органов.
А в январе
2001 года появилась
информация
об открытии,
которое может
сделать клонирование
просто не нужным.
Удалось повернуть
вспять биологические
часы внутри
человеческой
клетки, заставив
ее вернуться
к состоянию,
в котором она
находилась
на момент
образования
в эмбрионе.
3.4.
Лечение и
предупреждение
некоторых
наследственных
болезней человека
Повышенный
интерес медицинской
генетики к
наследственным
заболеваниям
объясняется
тем, что во многих
случаях знание
биохимических
механизмов
развития позволяет
облегчить
страдания
больного. Больному
вводят не
синтезирующиеся
в организме
ферменты. Так,
например,
заболевание
сахарным диабетом
характеризуется
повышением
концентрации
сахара в крови
вследствие
недостаточной
(или полного
отсутствия)
выработки в
организме
гормона инсулин
поджелудочной
железой. Это
заболевание
вызывается
рецессивным
геном. Еще в
19 веке это заболевание
практически
неизбежно
приводило к
смерти больного.
Получение
инсулина из
поджелудочных
желез некоторых
домашних животных
спасло жизни
многим людям.
Современные
методы генной
инженерии
позволили
получать инсулин
гораздо более
высокого качества,
абсолютно
идентичный
человеческому
инсулину в
масштабах,
достаточных
для обеспечения
каждого больного
инсулином и
с намного меньшими
затратами.
Сейчас известны
сотни заболеваний,
в которых механизмы
биохимических
нарушений
изучены достаточно
подробно. В
некоторых
случаях современные
методы микроанализов
позволяют
обнаружить
такие биохимические
нарушения даже
в отдельных
клетках, а это,
в свою очередь,
позволяет
ставить