Аффинные иммуносорбенты. Использование радиоактивных изотопов
Реферат
на тему:
Аффинные
иммуносорбенты.
Использование
радиоактивных
изотопов
Аффинные
иммуносорбенты
Высокая
степень избирательности
связи антиген
- антитело открывает
перспективу
создания аффинных
хроматографических
колонок для
очистки любых
белков, не имеющих
подобно ферментам
и прочим биологически
родственным
парам, своих
специфических
партнеров.
Общий план
действий здесь
таков:
1. Иммунизировать
кролика неким
хорошо очищенным
белком.
2. Тот
же белок ковалентно
закрепить на
матрице, например,
в колонке
BrCN-активированной
сефарозы.
3. Иммунную
антисыворотку
крови кролика
пропустить
через эту колонку.
Все прочие
антитела легко
смыть, а специфические
для нашего
белка антитела
на ней останутся.
4. Их можно
потом снять
изменением
рН буфера или
подачей высокой
концентрации
соли. (Антитела
прекрасно
хранятся)
5. Если
мы сумеем надежно
посадить эти
антитела на
другую колонку,
причем так, что
они не утратят
способности
связываться
со своим антигеном,
то с ее помощью
можно будет
извлечь из
любой сложной
смеси тот белок,
который нас
интересует,
а затем снять
и его в совершенно
очищенном виде"
Для
конкретизации
способа осуществления
этого плана
нам потребуются
дополнительные
сведения о
структуре и
свойствах
антител. Во-первых,
укажем точное
место нахождения
антител в составе
крови.
Если
кровь налить
в пробирку и
подождать пока
она свернется,
а затем центрифугированием
убрать осадок,
то с ним уйдут
и красные кровяные
клетки (эритроциты),
и белые (лейкоциты
и лимфоциты),
и белок фибриноген,
который осуществляет
свертывание.
Останется
прозрачная
жидкость, именуемая
сывороткой
крови. В ней
растворены
сахара, гормоны,
витамины, соли
и прочие низкомолекулярные
вещества, а
также разнообразные
и многочисленные
белки крови.
Основную их
массу составляют
два класса
белков: альбумины
и глобулины.
Их можно разделить
добавлением
сульфата аммония
до концентрации
в 2М.
Глобулины
выпадают в
осадок. Если
же всю сыворотку
подвергнуть
фракционированию
электрофорезом
при рН8,6, то вперед
быстро уйдут
альбумины, а
затем последуют
а-, (3-й у-глобулины.
Вот в
этой последней
фракции, так
называемых
иммуноглобулинов
и находятся
антитела. Существует
несколько типов
иммуноглобулинов.
Из них богаче
других представлен
в крови и используется
для исследовательских
целей один -
иммуноглобулин
типа G,
обозначаемый
IgG.
Сыворотку
крови животного,
успевшего
выработать
"иммунный
ответ", т.е. антитела,
специфичные
для введенного
извне антигена,
и называют
антисывороткой.
Иногда
в ее полном
названии указывают
и стимулировавший
эту антисыворотку
антиген и даже
его происхождение.
Например,
"антисыворотка
кролика против
эритроцитов
козы".
Название
длинное, зато
полностью
описывает
конкретный
случай: в кровь
кролика были
введены эритроциты
из крови козы.
Они - чужие и
потому вызвали
иммунный ответ.
Вообще-то
говоря, у человека
известно четыре
варианта: от
IgGI до IgG4, да
еще некоторое
количество
других иммуноглобулинов:
IgM, IgA, IgE,
играющих несколько
различные роли.
Но мы
в эти детали
вникать не
будем. Различия
их структур
незначительны
и более 60% приходится
на долю IgGI. Именно
его мы будем
далее подразумевать
под термином
"антитело".
Молекулярная
масса IgGI составляет
146 тысяч дальтон.
Вся
его пространственная
конфигурация
установлена
методом
рентгеноструктурного
анализа. Под
электронным
микроскопом
антитело выглядит
подобно латинской
букве "Y".
На ее
"стержне" и
обеих "ветвях"
можно различить
отдельные
участки ("домены"),
где полипептидная
цепь белка
скручена и
образует некое
подобие полусферы,
как это изображено
на рис.1.
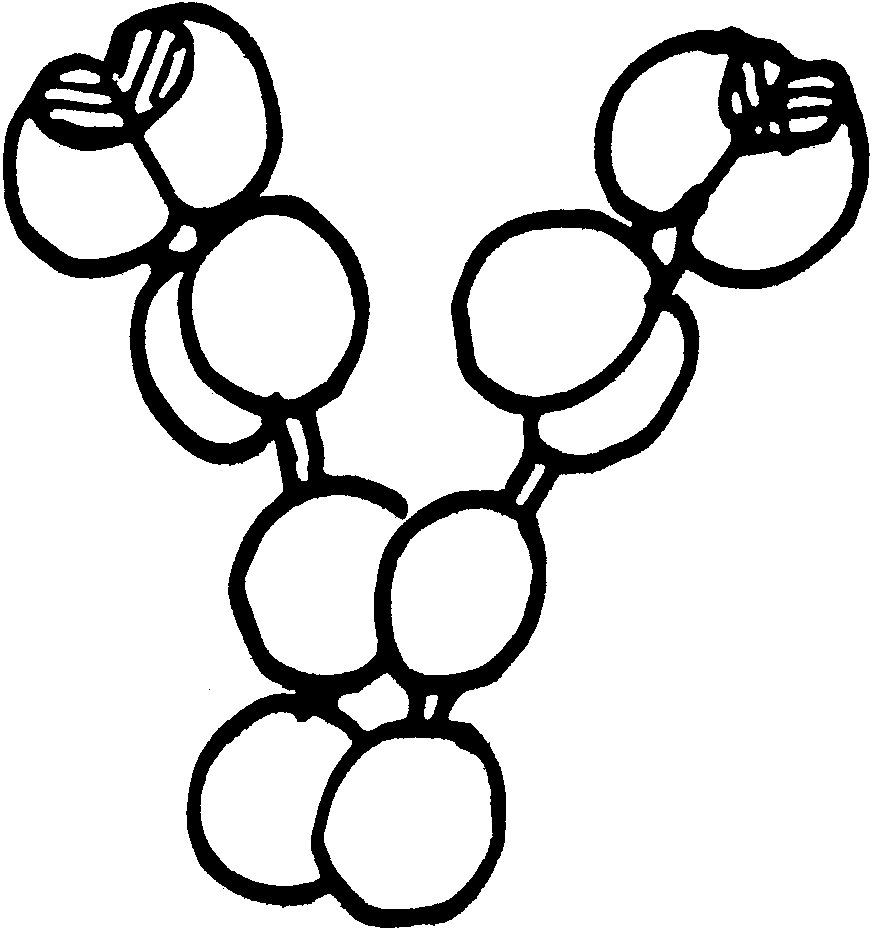
Рис.
1
Заметная
на рисунке
парность доменов
отражает
действительное
подобие строения
членов каждой
пары, но все
они водородными,
гидрофильными,
а главное -
дисульфидными
мостиками
связаны в одну
молекулу.
Самое
важное для нас
то обстоятельство,
что все четыре
домена "стержня"
и ближние к
нему пары доменов
"ветвей" неизменны
по своему
аминокислотному
составу и строению
у всех антител
("константная
часть").
И только
две пары концевых
доменов представляют
собой вариабильную
часть антитела,
определяющую
его антигенную
специфичность.
В этих доменах
есть участки,
которые именуются
"гипервариабильными
областями"
(на рис. - заштрихованы).
В них
различия антител,
специфичных
для различных
антигенов,
особенно ярко
выражены. В
каждой паре
концевых доменов
они образуют
некое подобие
"ущелья".
Можно
ожидать, что
именно здесь
связывается
антиген. Такое
предположение
нашло прямое
подтверждение
после того, как
удалось кристаллизовать
концевой фрагмент
одного из антител
вместе со связанным
с ним антигеном
(витамином К).
Анализ
рентгенограммы
показал, что
молекула витамина
лежит в "ущелье"
области узнавания
и различные
ее части сближены
и взаимодействуют
с определенными
аминокислотами
в гипервариабильной
области.
На рис.
изображена
пространственная
конфигурация
одного из
вариабильных
концевых доменов
IgGl. Полипептидная
цепочка из 104
аминокислот
свернута так,
что три ее
заштрихованных
(гипервариабильных)
участка выходят
на поверхность
домена примерно
в одной плоскости,
образуя один
из склонов
"ущелья" связи.
Точками
на этом же рисунке
обозначены
места соединения
с парным доменом
оконечности
той же ветви,
образующим
второй склон.
Аминокислотный
состав гипервариабильных
участков на
обоих склонах,
очевидно, разный.
Но "ущелье"
на конце второй
ветви молекулы
антитела является
точной копией
"ущелья" на
первой ветви.
Непосредственную
область узнавания
и связывания
антигена в
"ущелье" на
конце ветви
образуют около
20 аминокислот.
Но еще не менее
70 аминокислот
каждого из
концевых доменов
участвуют в
определении
пространственной
конфигурации
этой области.
Но вернемся
к нашей программе
создания
аффинно-иммунной
колонки. Понятна
возможность
реализации
четырех ее
первых пунктов.
Из сыворотки
иммунизированного
кролика, двухмолярным
раствором
сульфата аммония
осаждают суммарную
фракцию глобулинов.
Затем
ее растворяют,
диализом освобождают
от соли и наносят
в буфере рН6,5
на колонку
ДЕАЕ-целлюлозы.
При таком значении
рН IgG не
задерживается
на колонке и
выходит сразу
очищенным,
поскольку все
прочие глобулины
остаются на
колонке.
Одновременно
подготавливают
колонку на
основе BrCN-активированной
сефарозы и
фиксируют на
ней белок, которым
был иммунизирован
кролик. Раствор
суммарной
фракции IgG пропускают
через эту колонку
и промывают
ее буфером. В
результате
на колонке
задерживаются
только антитела,
специфичные
для этого
белка-антигена.
Их можно снять
0,1М раствором
уксусной кислоты
или 4,5М раствором
MgClg. Полученный
таким образом
раствор нужных
антител нейтрализуют,
диализуют от
солей и, если
надо, концентрируют
упариванием
воды.
Теперь
встает последний
вопрос. Как
связать эти
антитела с
матрицей еще
одной колонки,
причем так,
чтобы не пострадала
их антигенная
активность
и специфичность?
Нелегкий вопрос!
Но тут
природа сделала
исследователям
роскошный
подарок. Оказалось,
что некий белок
из оболочки
бактерии
Staphylococcus aureus,
названный
"белком А",
способен прочно
связываться
с неизменной
частью ("стержнем")
большинства
IgG.
За словом
"оказалось"
скрывается
многое. Вряд
ли белок с таким
свойством
специально
искали именно
у этой бактерии.
Скорее всего
его обнаружили
случайно. (Но,
как говорит
пословица,
ученый тот, кто
видит то же,
что видят все,
но замечает
то, что не заметил
никто!).
Так
или иначе, но
обнаружение
указанного
свойства белка
А открыло путь
к осуществлению
последнего
этапа нашей
программы. На
колонку активированной
сефарозы химически
прочно сажают
белок А (даже
есть в продаже
готовая "Protein
А - Sepharose CL-4В")
и через нее в
определенных
условиях пропускают
раствор специфических
антител. Они
закрепляются
на колонке
через свои
"стержни" и
белок А.
Оба
конца разветвления
остаются свободными.
Колонка готова
- создан аффинный
иммуносорбент
для немедленной
очистки из
любой белковой
смеси того
белка, который
был использован
для иммунизации
кролика. Снять
его с колонки
после очистки
не представляет
труда - кислотой
или солью...
Аффинные
иммуносорбенты
нашли широкое
применение
и, кстати сказать,
не только для
очистки белков.
В 1978 г. Stumphet сумели
этим методом
из смеси фрагментов
ядерной ДНК
отделить гибридные
ДНК-РНК куски.
По этим гибридам
определили
гены, ответственные
за синтез
рибосомальных
РНК. (Кролика
в этом случае
иммунизировали
двухнитевым
синтетическим
гибридом
полиадениловой
кислоты и
полидезоксириботимидиновой
кислоты) Оказалось,
что такой гибрид
отлично имитирует
в отношении
специализации
антител гибрид
ДНК-РНК.
Отыскание
антигена после
электрофореза
смеси белков
Это -
еще один весьма
полезный вариант
использования
иммуноспецифического
связывания
антител с их
антигенами.
Представим
себе, что после
разделения
смеси белков
электрофорезом
в ПААГ получается
большое число
белковых полос,
из которых одна
должна содержать
искомый белок.
Здесь
можно использовать
сочетание
избирательности
иммуноспецифического
связывания
с очень чувствительным
способом
использования
радиоактивности.
Интересующим
исследователя
белком иммунизируют
кролика и очищают
из его крови
антисыворотку.
Разработана
и имеется в
продаже специальная,
так называемая
"диазобумага".
Она
обладает способностью
по своим активным
диазогруппам
ковалентной
химической
связью присоединять
любые белки.
По
окончании
электрофореза
с верхней поверхности
геля фильтровальной
бумагой удаляют
избыток влаги
и кладут на
него диазобумагу,
зафиксировав
ее положение
на геле проколами
по двум углам
бумаги и геля.
Потом эту пару,
гелем вниз,
помещают в
систему "блоттинга",
для переноса
ДНК из агарозного
геля на нитроцеллюлозный
фильтр.
То есть
кладут гель
на лист толстой
фильтровальной
бумаги, постоянно
смачиваемой
буфером, а на
диазобумагу
накладывают
стопку фильтровальной
бумаги и через
пластинку
стекла прижимают
грузом. Током
жидкости, идущей
вверх по капиллярам,
часть белков
из каждой полосы
переносится
на диазобумагу
и там прочно
фиксируется.
Неиспользованные
диазогруппы
между белковыми
"репликами"
закрывают
вымачиванием
в 10% -ном растворе
этаноламина,
который в отношениях
с диазогруппой
имитирует
белок. Затем
диазобумагу
помещают в
раствор полученной
ранее антисыворотки
и выдерживают
12 часов при 37°С.
За это
время завершается
реакция связывания
специфических
антител, имеющихся
в антисыворотке
с фиксированным
на бумаге антигеном.
(Для всех остальных
белков в антисыворотке,
- даже суммарной,
- нет специфических
антител) Оставшиеся
свободными
антитела отмывают
буфером.
Наконец,
диазобумагу,
на которой
имеется реплика
искомого белка
вместе со
связавшимися
с ним антителами,
инкубируют
2 часа при 37°С в
растворе меченого
радиоактивным
йодом белка
А (он доступен
в виде готового
препарата).
Не
сорбированный
на "стержнях"
антител белок
А отмывают
крепким солевым
раствором.
Бумагу сушат
и методом
авторадиографии
(описанным в
следующей
главе) на рентгеновской
пленке локализуют
положение
полосы искомого
белка. Для
последующего
правильного
совмещения
пленки с диазобумагой
(а через нее и
с гелем), на бумаге
перед авторадиографией
делают две
пометки радиоактивными
чернилами.
Радиоактивные
изотопы, используемые
в биологии
Явление
радиоактивного
распада изучается
в школьном
курсе физики,
и потому физическую
природу радиоактивности
мы здесь рассматривать
не будем. Перечислю
только те
характеристики
этого явления,
которые существенны
для дальнейшего
изложения.
1. Известны
три вида радиоактивного
излучения: а,
(3 и у. ос-частицы
представляют
собой ядра
гелия; (З-частицы
- это обычные
электроны
(Р-электроны)
и у - очень коротковолновое
электромагнитное
изучение. В
биологии используются
только два
последних вида.
2. Р-электроны
испускаются
ядрами атомов
и потому химические
свойства
соответствующих
радиоактивных
изотопов (их
валентности
и химические
связи) при этом
не изменяются.
3. Интенсивность
излучения
любого радиоактивного
препарата
уменьшается
со временем,
поскольку
уменьшается
число атомов,
способных к
радиоактивному
распаду в смеси
с теми, в которых
он уже произошел.
Скорость этого
уменьшения
характеризуется
так называемым
временем
полураспада
(ТУ).
То есть
временем, за
которое любое
данное количество
изотопа (а с
ним и интенсивность
излучения)
уменьшается
вдвое. Очевидно,
что эта характеристика
остается неизменной
в течение всего
"периода жизни"
этого изотопа.
Время полураспада
для различных
изотопов колеблется
в очень широких
пределах.
Для
изотопов,
используемых
в биологии, -
от нескольких
суток до нескольких
тысяч лет (см.
таблицу ниже).
Знания величины
ТУ очень существенно:
в одних случаях
даже при длительном
хранении
радиоактивного
препарата
интенсивность
его излучения
практически
не изменяется,
в других - она
может упасть
почти до нуля
за пару недель.
4. Интенсивность
излучения
данного радиоактивного
препарата в
данный момент
времени характеризуется
числом радиоактивных
распадов (актов
излучения) в
минуту. Его
чаще всего
выражают в
единицах "Кюри"
(Ки). Это - очень
крупная единица.1Ки==2,2
1012 распадов
в минуту.
Другая
величина, принятая
в системе единиц
СИ, оказалась,
напротив, слишком
мала: 1 Беккерель
(Бк) = 1 распаду
в секунду. Поэтому
до сих пор
интенсивность
предпочитают
выражать числом
милликюри (мКи)
или микрокюри
(мкКи).
5. Практический
интерес представляет
удельная
радиоактивность
(УА), т.е. радиоактивность
1 миллимоля
радиоактивного
вещества, в
котором часть
атомов уже
претерпела
радиоактивный
распад. Кроме
того, в подавляющем
большинстве
случаев истинно
радиоактивные
изотопы составляют
лишь некоторую
долю от общего
числа атомов
(или молекул)
вещества, именуемого
радиоактивным.
УА выражают
в единицах
Ки/ммоль или
мКи/ммоль.
6. Очень
важной характеристикой
радиоактивного
излучения
является энергия
вылетающих
частиц (у-излучение
тоже происходит
отдельными
порциями энергии
- "у-квантами").
И Р-электроны,
и у-кванты,
испускаемые
различными
атомами одного
и того же радиоактивного
препарата,
могут иметь
разные величины
энергии.
В качестве
характеристики
изотопа принято
указывать
величину максимально
возможной
энергии соответствующих
частиц или
квантов. Ее
выражают в
"электронвольтах"
(эВ) или килоэлектронвольтах
(КэВ). Из курса
физики известно,
что 1эВ соответствует
энергии, приобретаемой
электроном
при пролете
в вакууме ускоряющего
электрического
поля с разностью
потенциалов
в 1 Вольт.
В нижеследующей
таблице собраны
все эти данные
для изотопов,
используемых
в биологии.
| Изотоп |
ТУз |
УА
(Ки/ммоль) |
Энергия
(КэВ) |
|
|
|
Р-электронов,
Emax |
у-излучения |
| "Н |
12,3 лет |
29 |
19 |
- |
| 14^ |
5730 лет |
62 (мКи/ммоль) |
156 |
- |
| 35g |
87,4 сут. |
1 500 |
167 |
- |
| 32р |
14,3 сут. |
9 000 |
1708 |
,- |
| 125J |
60 сут. |
2 000 |
6 |
35 |
| 131J |
8 сут. |
16 000 |
250-810 |
80-720 |
(Цифра,
стоящая слева
вверху у химического
символа изотопа,
указывает его
массу в единицах
массы атома
нормального
водорода. Она
может быть
больше или
меньше массы
соответствующего
нерадиоактивного
атома. Объяснение
этому - в курсе
физики)
Иногда
УА препарата
выражают не
в Кюри, а числом
импульсов в
минуту (имп/мин),
которые регистрирует
прибор, и относят
не к миллимолю,
а к миллиграмму
или микрограмму
вещества
(имп/мин/мг). Строго
говоря, это
некорректно,
так как включает
сюда эффективность
счета импульсов,
которая может
быть далеко
не стопроцентной.
Но
для сравнительных
измерений,
например, для
наблюдения
за ходом очистки
индивидуального,
радиоактивно
меченого белка
- вполне приемлемо.
Кроме того,
радиоактивность
чаще всего
используют
для "детектирования"
- отделения
меченых препаратов
определенной
природы от
таких же, но не
меченых. Здесь
эффективность
счета не существенна.
Следует
отдавать себе
отчет в том,
что указанные
в таблице значения
максимальной
энергии полезны
для сопоставления
энергетических
возможностей
(прохождения
через преграды)
каждого из
изотопов, но
отнюдь не
представляют
энергию большинства
реально испускаемых
частиц или
квантов.
Это
хорошо видно
из графиков
распределения
по энергиям
электронов,
представленных
на рис. По осям
абсцисс на этих
графиках отложены
величины энергии
в КэВ, естественно,
в разных масштабах
(см. цифры). По
осям ординат
относительное
содержание
электронов
указанной
энергии. (Кривые
не доходят до
оси ординат,
так как энергии
электронов,
близкие к нулю
не поддаются
измерению)
Указанные
значения Е
соответствуют
средней величине
энергии электронов
соответствующего
изотопа, на
которую следует
ориентироваться
при оценке
возможностей
их регистрации.
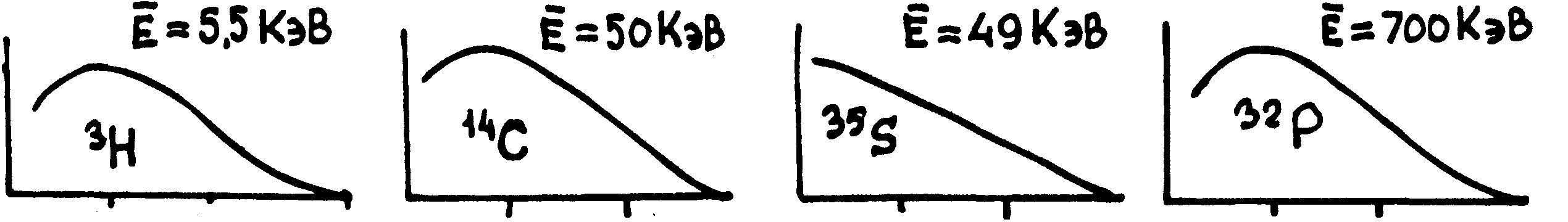
Рис.
2
Литература
Курашвили
Л.В., Николаев
П.Н. Диагностическая
значимость
исследования
холестерина
в ЛПВП у ожоговых
больных. III Всесоюзная
конференция
по проблеме:
Современные
средства первой
помощи и методы
лечения ожоговой
болезни /Тезисы/
- Москва, 1986. - С.186-187.
Курашвили
Л.В., Савченко
Р.П. Изменение
показателей
липидного
обмена у больных
с хронической
почечной
недостаточностью,
находящихся
на программированном
гемодиализе
/Лабораторное
дело. - N 12. - 1986. - С.717-719.
Курашвили
Л.В., Николаев
П.Н. Новое в
лабораторной
диагностике
ожоговых больных.
Научно-практическая
конференция,
посвященная
140-летию областной
больницы им.Н.
Н. Бурденко и
110-летию со дня
рождения академика
Н.Н. Бурденко.
Тез. докладов.
- Пенза, 1986. - С.103-105