ФЕДЕРАЛЬНОЕ АГЕНСТВО ПО ОБРАЗОВАНИЮ
Пензенский государственный педагогический университет
им. В. Г. Белинского
Факультет Естественно-географический
Кафедра биохимии
Дипломная работа
Активность карбоксипептидазы N и ангиотензинпревращающего фермента в сыворотке крови у онкологических больных при химиотерапевтическом воздействии
Студент
_____________________ Лобзина Е. С.
Руководитель
______________________ Сметанин В. А.
К защите допустить.
Протокол № от « _ » 200 г
Зав. кафедрой
____________________________ Генгин М.Т.
Пенза, 2009 г.
Содержание
Список сокращений
Введение
Глава 1. Обзор литературы
1.1. Противоопухолевая химиотерапия
1.1.1. Механизм действия противоопухолевых препаратов
1.1.2. Осложнения химиотерапии
1.2. Пептидергическая система
1.2.1. Механизм образования активных форм регуляторных пептидов
1.2.2. Роль биологически активных пептидов
1.2.3. Ферменты обмена регуляторныхпептидов
1.2.3.1. Ангиотензинпревращающий фермент
1.2.3.2. Карбоксипептидаза N
1.3. Функционирование пептидергической системы при онкологических заболеваниях
1.4. Роль оксида азота (II) в онкогенезе
Глава 2. Материалы и методы
2.1. Материал исследования
2.2. Методы исследования
2.2.1. Метод определения активности карбоксипептидазы N
2.2.2. Метод определения активности ангиотензинпревращающего фермента
2.2.3. Метод определения содержания белка
2.2.4. Метод количественного определения оксида азота (II) в сыворотке крови
2.3. Статистическая обработка результатов исследования
Глава 3. Результаты и обсуждение
3.1. Исследование активности карбоксипептидазы N в сыворотке крови онкологических больных при химиотерапевтическом воздействии
3.2. Исследование ангиотензинпревращающего фермента в сыворотке крови онкологических больных при химиотерапевтическом воздействии
3.3. Исследование содержания нитрит–иона в сыворотке крови у онкологических больных при химиотерапевтическом воздействии
Выводы
Список литературы
Список сокращений
АПФ – ангиотензинпревращающий фермент
ККС – калликреин-кининовая система
КПN – карбоксипептидаза N
ЛАП – лейцин-аминопептидаза
ММП – матриксные металлопротеазы
РААС – ренин-ангиотензин-альдостероновая система
РЩЖ – рак щитовидной железы
iNOS – индуцибельная NO-синтаза
NO – оксид азота II
TNFa – фактор некроза опухолей
Введение
В структуре заболеваемости и смертности онкологическая патология занимает одно из ведущих мест. С каждым годом число людей, страдающих этим заболеванием, растет [2,34,49].
Развитие опухоли в организме приводит посредством различных механизмов к глубоким физико-химическим, нейрогуморальным и ферментативным сдвигам. Опухоли паразитируют в организме, извлекая необходимые для роста вещества из здоровых клеток и секретируя токсины локально или в системный кровоток[5,18,30,38,49].
Существует три направления лечения раковых заболеваний: хирургическое, радиологическое и химиотерапевтическое. Химиотерапия злокачественных опухолей – это использование лекарственных веществ, тормозящих пролиферацию или необратимо повреждающих опухолевые клетки [10,30,43]. Главной причиной, послужившей стимулом к развитию лекарственной терапии злокачественных опухолей, явилась неудовлетворенность результатами хирургического и лучевого лечения [7].
Особенность применения химиопрепаратов заключается в близкой сопряженности лечебного и токсического эффекта. К побочным действиям относят проявления нейро-, гепато-, кардиотоксичности, нарушения системы свертывания крови и эндокринных органов [15,25,32,37]. Итогом побочных действий являются, в том числе, нарушения в функционировании калликреин-кининовой и ренин-ангиотензиновой систем, важными компонентами которых являются ангиотензинпревращающий фермент и карбоксипептидаза N [52]. Кроме того, эти ферменты влияют на уровень брадикинина, стимулирующего продукцию оксида азота (II), который, в свою очередь, являясь мультифункциональной молекулой, может регулировать пролиферативную активность и апоптоз опухолевых клеток [28].
Целью
настоящей работы было изучение роли карбоксипептидазы N и ангиотензинпревращающего фермента в период химиотерапии у онкологических больных.
При выполнении работы были поставлены следующие задачи:
1. Изучить активность ангиотензинпревращающего фермента, карбоксипептидазы N и концентрацию оксида азота (II) у здоровых людей.
2. Изучить активность ангиотензинпревращающего фермента, карбоксипептидазы N и концентрацию оксида азота (II) у онкологических больных до химиотерапии.
3. Изучить активность ангиотензинпревращающего фермента, карбоксипептидазы N и концентрацию оксида азота (II) у онкологических больных после химиотерапии.
4. Установить корреляционные взаимосвязи между активностью ферментов и концентрацией оксида азота (II) в сыворотке крови у онкологических больных до и после химиотерапии.
Научная новизна и практическая ценность работы.
Изучена активность карбоксипептидазы N, ангиотензинпревращающего фермента и концентрация оксида азота (II) в сыворотке крови у онкологических больных в период химиотерапии. Установлены корреляционные взаимосвязи между активностью исследуемых ферментов и уровнем оксида азота (II) при химиотерапевтическом воздействии.
Полученные результаты позволяют расширить наши представления о биологической роли карбоксипептидазы N и ангиотензинпревращающего фермента, а также получить более полную картину о влиянии химиопрепаратов на организм при онкологических заболеваниях и могут быть использованы для коррекции терапии, оценки его эффективности и контроля больных после окончания курса лечения.
Апробация работы
Материалы данной работы представлены на 58 научной конференции студентов ПГПУ, на XVI Международной научно-практической конференции «Экология и жизнь» (24-25 апреля 2009 г.).
Глава 1. Обзор литературы
1.1
Противоопухолевая химиотерапия
Химиотерапия злокачественных опухолей предполагает применение лекарственных веществ избирательно действующих на рост опухоли и опухолевую ткань. Противоопухолевый эффект определяется соотношением действия вещества или продуктов его превращения в организме непосредственно на опухолевые клетки и организм в целом [9,10,40,43].
Основные принципы химиотерапии опухолей:
1. Подбор препарата соответственно спектру его противоопухолевого действия.
2. Выбор оптимальной дозы и режима введения препарата, обеспечивающих лечебный эффект без побочных явлений.
3. Учет факторов, требующих коррекции доз и режимов во избежание тяжелых осложнений химиотерапии [17,25].
Существует прямая зависимость между разовой и суммарной дозой и терапевтическим эффектом. Доза препаратов должна быть максимально переносимой и вызывать максимальный противоопухолевый эффект [9,37,43,44].
Химиопрепараты обладают определенной специфичностью на различные виды злокачественных новообразований. Быстро растущие опухоли более чувствительны к химиотерапии, чем медленно растущие. Метастазы опухолей более чувствительны к химиопрепаратам, чем первичные новообразования. Эффективность химиотерапии обратно пропорциональна массе опухоли, при которой начинают лечение. Чем больше опухоль, тем меньше эффект, и наоборот [25,46,55].
Современная химиотерапия – это комбинированная химиотерапия, когда используются одновременно от 2 до 6 цитостатиков с разным механизмом действия. При этом исходят из представления о том, что нарушения различных биохимических реакций в опухолевой клетке уменьшают шансы на то, что уцелеют резистентные клоны опухоли. Обычно в эти комбинации включают препараты с разной токсичностью, что может увеличить противоопухолевый эффект без увеличения общей токсичности [10,37].
Основной проблемой химиотерапии является неспособность цитостатиков полностью уничтожить опухоль в большинстве случаев. Клеточная популяция опухоли весьма гетерогенна, что делает вероятным существование клеточных линий, резистентных к химиотерапии. По мере гибели чувствительных к цитостатикам клеток химиорезистентные штаммы получают избирательное преимущество в росте [7,25,45].
1.1.1 Механизм действия противоопухолевых препаратов
По своему происхождению и механизму действия существующие противоопухолевые препараты делятся на следующие группы:
1. Алкилирующие агенты: хлорбутин, эмбихин, допан, ломустин.
2. Антиметаболиты: метотрексат, меркаптопурин, фторурацил, фторафур.
3. Антибиотики: дактиномицин, блеомицин, рубомицин, адриамицин.
4. Алкалоиды растительного происхождения: винбластин, винкристин, таксол [10,25,30].
По происхождению химиопрепараты можно разделить на синтетические и природные. К синтетическим препаратам относятся алкилирующие агенты и антиметаболиты. К продуктам природного происхождения относят антибиотики и алкалоиды. Противоопухолевые средства способны уничтожать опухолевую клетку (цитотоксический эффект) или угнетать их пролиферативную активность (цитостатический эффект) [43,51]. Механизм действия цитостатических веществ основан на их реагировании с молекулами жизненно важных химических соединений, составляющих субстрат клеток (белками, нуклеиновыми кислотами, ферментами), вследствие чего наступает изменение структуры и физико-химических свойств внутриклеточных компонентов, подавление биокаталитических процессов, лежащих в основе обмена веществ, нарушение митотического процесса и гибель клеток [51].
Вещества алкилирующего действия взаимодействуют с нуклеиновыми основаниями двойной спирали ДНК. Они присоединяются к ДНК путем реакции алкилирования – замещения атома водорода нуклеинового основания на метильную группу цитостатика. В результате образуются аномальные пары оснований. Это приводит к прямому подавлению транскрипции либо к образованию дефектной РНК и синтезу аномальных белков.
Антиметаболиты. Структурная или функциональная схожесть с молекулами- метаболитами позволяет им блокировать синтез нуклеотидов и тем самым угнетать синтез ДНК или РНК либо напрямую встраиваться в структуры ДНК и РНК, блокируя процессы репликации и синтеза белков.
Антибиотики. Непосредственно воздействуют на ДНК путем интеркаляции, запускают механизм свободно-радикального окисления с повреждением мембран клеток и внутриклеточных структур, а также ДНК. Нарушение структуры ДНК ведет к нарушению процессов репликации и транскрипции.
Алкалоиды растительного происхождения. Цитостатический эффект винка-алколоидов обусловлен деполимеризацией тубулина. Процесс клеточного деления останавливается в фазе митоза. Другие препараты способствуют полимеризацию тубулина, вызывая образование дефектных микротрубочек и необратимую остановку клеточного деления. По-видимому, противоопухолевый эффект проявляемый некоторыми алкалоидами зависит также от подавления ими некоторых других биохимических процессов [10,22,25,30,40,43].
1.1.2 Осложнения химиотерапии
Одна из проблем химиотерапии – токсичность цитостатических препаратов [25]. Применяемые даже в терапевтических дозах химиопрепараты вызывают целый ряд побочных явлений, что связано с их повреждающим действием на ряд органов и систем организма. Невысокая избирательность действия противоопухолевых препаратов объясняется отсутствием качественных отличий в биохимии, темпе роста, способности к репарации после повреждения между опухолевыми и нормальными клетками. Наряду с подавлением ими различных этапов обмена нуклеиновых кислот в опухолевых клетках, они оказывают влияние и на обмен нуклеиновых кислот быстро размножающихся популяций нормальных клеток – иммунокомпетентных, костного мозга, желудочно-кишечного тракта, репродуктивных органов [15,17,25,29,41].
Ряд побочных эффектов сравнительно специфичен для отдельных цитостатиков. К таким побочным действиям относят проявления нейро-, гепато-, кардиотоксичности, нарушения системы свертывания крови, эндокринных органов. Подобные специфические осложнения химиотерапии опухолевых заболеваний зависят от особенностей фармакологических свойств цитостатиков и их метаболизма [15,37].
Нейротоксичность обнаруживается лишь у некоторых противоопухолевых препаратов независимо от их способности проникать через ГЭБ. Проявляется в слабости скелетной мускулатуры, судорожных мышечных сокращений, развитии глаукомы. Имеются данные о нарушениях обмена аланина, лейцина и серина в синапсах [15,17].
Гепатотоксическое действие цитостатиков выражается в разной степени. Наибольшую часть осложнений составляют гепатопатии, не выходящие за пределы отклонений в показателях лабораторных тестов. При длительной цитостатической терапии наблюдаются гипербилирубинемия и гиперхолестеринемия, снижение уровня протромбина и коагуляционных факторов крови. При лечении производными платины гепатотоксичность проявляется в обратимом повышении аминотрансфераз. Антиметаболиты вызывают изменение отдельных функций печени вплоть до желтухи [15,44].
Кардиотоксическое побочное действие в основном присуще противоопухолевым антибиотикам. Отмечают боли в области сердца и нарушение ритма [15,44,49].
Нефротоксичность (характерно для производных платины) зависит от дозы введенного препарата и проявляется повышением содержания мочевины, мочевой кислоты и креатинина в плазме, снижением креатинового клиренса [44].
Алкалоиды наряду с общетоксическим действием влияют на утилизацию глутаминовой кислоты и аргинина, определенным образом действуют на обмен пролина, глутамина, триптофана. Усиливают отдачу гипоксантина, но не тормозят утилизацию клетками аденина. Не вызывают существенных нарушений со стороны печени за исключением уменьшения содержания альбуминов крови. Со стороны почек отмечают уменьшение содержания натрия [44].
Антиметаболиты угнетают гемопоэз, обладают антикоагулянтными свойствами, что проявляется в нарушении свертываемости крови.
Проявление токсичности лимитирует использование противоопухолевых агентов. Развитие осложнений приводит к необходимости снижения дозы цитостатиков или увеличению интервалов между курсами [25,40,43].
1.2 Пептидергическая система организма
1.2.1 Механизм образования активных форм регуляторных пептидов
Активные формы пептидов представляют собой полифункциональную группу веществ, которым отводится важная роль природных биорегуляторов. Это природные или синтетические соединения, молекулы которых построены из остатков -аминокислот, соединенных между собой пептидными (амидными) связями C(O)–NH. Большинство регуляторных пептидов образуется из физиологически неактивных белков-предшественников, путем посттрансляционного процессинга [32]. Секретируемые белково-пептидные продукты синтезируются на мембраносвязанных рибосомах ЭПР. Благодаря наличию на N-конце сигнальной последовательности, состоящей из остатков гидрофобных аминокислот, обеспечивается транслокация пептида через мембраны ЭПР. В полости ЭПР отщепление этой последовательности осуществляется при участии сигнальной пептидазы. Далее процессинг осуществляется в ходе передвижения молекул пропептидов по гранулярному ЭПР, комплексу Гольджи и в секреторных везикулах [14,61].
Сначала под действием эндопептидаз образуются неактивные пептиды, содержащие со стороны С- или N-конца “лишние” остатки аминокислот, которые затем удаляются экзопептидазами с карбоксипептидазо-B- и аминопептидазо-B-подобной активностью [4].
Уровень биологически активных пептидов в организме в значительной степени определяется активностью ферментов их обмена, к которым в частности принадлежат АПФ и КПN [13,27,32].
В связи с этим, большой интерес представляет изучение активности данных ферментов у онкологических больных при химиотерапевтическом воздействии, которое прямо или косвенно влияет на какую-либо систему организма.
1.2.2 Роль биологически активных пептидов
Область биологической активности пептидов чрезвычайно широка. Они влияют на состояние сердечно-сосудистой, иммунной, половой, эндокринной, пищеварительной и других систем, изменяют энергетический обмен в организме, участвуют в регуляции работы центральной нервной системы. КПN и АПФ играют важную роль в обмене ангиотензина и брадикинина [31,54].
Ангиотензины – пептиды, образующиеся в организме из белка плазмы ангиотензиногена. Почечный фермент ренин отщепляет от молекулы ангиотензиногена неактивный декапептид ангиотензин I. Другой фермент крови – АПФ – преимущественно в ткани легких отщепляет с карбоксильного конца молекулы ангиотензина I дипептид с образованием ангиотензина II. Ангиотезин IIявляется физиологическим фактором роста клеток, обладает митогенными (учащающими деление) свойствами и, тем самым, стимулирует гиперплазию и пролиферацию клеток. Пептид повышает активность симпатоадреналовой системы, увеличивая синтез адреналина и обусловливая высвобождение норадреналина из окончаний симпатических нервов, что стимулирует гипертрофию сердца и сосудов. Ангиотензин II оказывает сильное сосудосуживающее действие, вызывает быстрое и длительное повышение артериального давления. Кроме того, он увеличивает синтез альдостерона, что сопровождается реабсорбцией натрия и воды. В надпочечниках из ангиотензина II образуется ангиотензин III, обладающий положительной инотропной активностью. Далее при участии аминопептидазы Nобразуется ангиотензин IV, предположительно, участвующий в регуляции гемостаза [31,36,41,54].
Брадикинин – полипептид, состоящий из 9 аминокислот. Брадикинин способен расширять просвет периферических и коронарных сосудов, снижать артериальное давление, способствует синтезу NО в эндотелии. Пептид повышает проницаемость капилляров, сокращает гладкую мускулатуру бронхов и других органов, вызывает болевой эффект. Он стимулирует синтез и освобождение простагландинов и фактора некроза опухолей ( TNFa ) в различных тканях, освобождение ряда интерлейкинов, способствует процессам репарации и обладает инсулиноподобным действием, стимулируя захват глюкозы периферическими тканями, модулирует передачу нервных импульсов в ЦНС и периферической нервной системе, изменяет состояние гематоэнцефалического барьера [13,52,63]. Брадикинин участвует в широком спектре физиологических и патофизиологических эффектов, и особенно в развитии воспаления [52].
Разрушение брадикинина обусловлено наличием в крови и тканях высокоактивных ферментов – кининаз, осуществляющих физиологический контроль уровня кининов. Наиболее важную роль в метаболизме брадикинина играют два фермента – кининаза I (Карбоксипептидаза N), и кининаза II (ангиотензинпревращающий фермент).
1.2.3 Ферменты обмена вазоактивных пептидов
1.2.3.1 Ангиотензинпревращающий фермент. Ангиотензинпревращающий фермент (КФ3.4.15.1, АПФ, ангиотензин - конвертирующий фермент, кининаза II, дипептидилкарбоксипептидаза I,) является ключевым ферментом, связывающим между собой ренин – ангиотензиновую и калликреин – кининовую системы. Фермент присутствует в плазме крови, нервных клетках, клетках почечных канальцев, сердечной мышце, матке, слюнных железах. Основная локализация в организме человека – эндотелий сосудистой стенки [3]. По структуре АПФ представляет собой гликопротеин, существующий в виде мембрано- связанной формы и являющийся интегральным белком. Молекула фермента, представляющая собой одну полипептидную цепь, локализована экстрацеллюлярно, гидрофобный трансмембранный участок включает 17 аминокислотных остатков и находится в положении 1230-1247, а внутриклеточный гидрофильный участок состоит из 30 остатков. Имеется и растворимая форма АПФ отличающаяся от мембраносвязанной отсутствием трансмембранного и внутриклеточного участков. Мембраносвязанная форма имеет молекулярную массу 170кДа и включает С- и- N-гомологичные домены, обладающие энзиматической активностью. Предполагается, что домены АПФ могут иметь разные функции в организме. Возможно, между N- и C-доменами имеется участок, доступный для ферментативного расщепления. Таким образом, N-домен может освобождаться или из находящегося в растворе полноразмерного фермента, или из мембраносвязанной формы, составляя С-домен на мембране [21]. В принципе такой процесс может происходить где угодно в организме. Вопрос о функциональной роли доменов до сих пор остается неясным. Однако полученные к настоящему времени данные об обнаружении эндогенных субстратов, специфичных для N-домена, и о разном взаимодействии ингибиторов АПФ с доменами, а также присутствие в организме однодоменных форм фермента свидетельствуют в пользу физиологической значимости доменов. Каждый из доменов содержит активный центр, которые отличаются по скорости гидролиза пептидов, по степени торможения специфичными ингибиторами АПФ [54,56].
АПФ – металлопротеиназа, которая содержит в активном центре ион цинка и активируется ионами Сl-, NO3- ,SO42-, ингибируется соединениями, содержащими SH-группу, хелаторами (ЭДТА, о-фенантролин), брадикининпотенциирующим фактором (Ki
= 40 нм),2-меркаптоэтанолом. Кроме того, существуют специфические ингибиторы АПФ – каптоприл (К = 20 нм), лизиноприл (К= 3-10 нм), и эналаприл (К =25-35 нм) [4,20]. = 20 нм), лизиноприл (К= 3-10 нм), и эналаприл (К =25-35 нм) [4,20].
рН-Оптимум действия АПФ составляет 7,2-7,6. Препараты АПФ, выделенные из различных органов человека (легких, сердца, печени, мозга, плазмы крови) существенно не различались по следующим физико-химическим параметрам: молекулярной массе, изоэлектрической точке, рН-оптимуму, константе ингибирования известными ингибиторами АПФ. При этом их иммунологические и каталитические свойства могут быть различными [21].
При действии на физиологические субстраты АПФ может вызывать либо превращение неактивной формы в активную, инактивацию биологически активного пептида, либо трансформацию его активности. Так, участвуя в отщеплении С-концевого гистидиллейцина от ангиотензина I, он превращает его в физиологически активный ангиотензин II, инактивирует брадикинин путем последовательного удаления двух С-концевых дипептидов, расщепляет такие функционально активные пептиды, как мет-энкефалин, нейротензин, эндорфин, вещество Р, (действуя в этих превращениях как эндопептидаза), играя роль одного из регулирующих факторов в обмене этих биологически активных веществ [20,36,41]. АПФ принимает участие в процессинге энкефалинов, гидролизуя энкефалинсодержащие пептиды - Met-энкефалин-Arg6-Phe7 в мет-энкефалин и Met-энкефалин- Arg6-Glu7-Leu8 в Met-энкефалин-Arg6 [4].
АПФ является физиологическим регулятором концентрации в плазме пептида AcSDKP (N-AcSer-Asp-Lys-Pro), влияющего на пролиферацию гемопоэтических и других клеток.
Фермент участвует в регуляции артериального давления. Кроме того, он вовлечен в реализацию таких функций как репродуктивные процессы, защитные и иммунные реакции организма. Участие фермента в том или ином процессе определяется как его локализацией, так и особенностью действия на регуляторные пептиды [20,56]. Являясь фактором, связывающим ККС и РААС – систем, вовлеченных в регуляцию большинства функций организма, реагирует на изменения, возникающие при патологических процессах. В связи с этим представляет интерес изучение активности фермента у онкологических больных при химиотерапевтическом воздействии.
1.2.3.2 Карбоксипептидаза N. Карбоксипептидаза N(КФ 3.4.12.7. КП N, аргинин-карбоксипептидаза, кининаза I) обнаружена Erdosи соавт. в плазме крови человека в 1962 году. Названа так потому, что по свойствам и специфичности он близок к панкреатической карбоксипептидазе B, но отличается от нее тем, что не имеет неактивного предшественника. Относится к ферментам вневизикулярной локализации – внеклеточной жидкости и внешней поверхности цитоплазматических мембран. Фермент локализован в плазме крови, обнаружен в стенках кровеносных сосудов, слизистой оболочке носа, моче [13].
Фермент имеет Mr 280 кДа и состоит из четырёх субъединиц трёх типов: двух с Mr 88 кДа и по одной с Mr 55 кДа и 48 кДа. Субъединицы с Mr 88 кДа гликозилированы (на долю углеводов приходится 29% массы), не обладают ферментативной активностью и, по-видимому, стабилизируют фермент в плазме крови [61]. Субъединицы с Mr 48 кДа и 55 кДа обладают ферментативной активностью и не содержат в своём составе углеводных остатков [13,58].
КП Nотносится к металлокарбоксипептидазам. Активный центр фермента имеет форму кармана, в полости которого находится ион Zn2+
. В цельной сыворотке крови человека фермент активируется ионами Co2+
и в меньшей степени Ni2+
, инактивируется ионами тяжелых металлов, бензоилом–L-аргинином, и хелатными соединениями. Тормозящее действие ингибиторов КП N проявляется не только в опытах invitro, но и invivo. При внутривенном введении ингибитора (2-меркаптоэтанол, ЭДТА) усиливалось гипотензивное действие брадикинина. Каталитическая активность фермента оптимальна при pH 7,0 – 7,8, и зависит от природы буферной смеси. В фосфатном буфере максимум активности находится при pH 7,5; в зоне pH 7,0 – 6,0 наблюдается резкое падение активности; в трис-буфере pH – оптимум составляет 7,0. Фермент чувствителен к кислой среде. При pH среды 2,0 – 3,0 он инактивируется необратимо, а при pH 6,5 – 5,0 – обратимо. Трапезникова С.С. и Пасхина Т.С. провели работу по изучению свойств карбоксипептидазы Nиз сыворотки крови человека, очищенного в 273 раза. Полученные препараты, по данным ультрацентрифугирования, состояли из двух компонентов с константой седиментации 5,3S и 6,5S. Отмечена четко выраженная зависимость фермента от температуры: максимальная активность проявляется при 37°С, при 30°С она снижается наполовину. Лиофилизированные препараты сохраняют свою активность в течение 6 месяцев [4,13].
Помимо способности отщеплять С-концевой аргинин в брадикинине (истинная кининазная активность) и Met5
-энкефалин-Arg6
, лизин в Met5
-энкефалина-Lys6
, карбоксипептидаза N гидролизует более простые синтетические субстраты: гиппурил-L-аргинин, гиппурил–орнитин (пептидазная активность), а также расщепляет эфиры гиппурил-L-аргининовой кислоты [27]. При этом скорость расщепления субстратов, содержащих C-концевые остатки лизина в 5-6 раз выше, чем соответствующих пептидов с C-концевым аргинином. Скорость расщепления гиппурил-аргининовой кислоты в несколько раз выше, чем гиппурил-аргинина. Эстеразная активность фермента угнетается ионами тяжелых металлов, в особенности Cd2+
, а также аргининовой кислотой и ЭДТА [13].
Биологическая роль КП N во многом остаётся неясной. Инактивируя брадикинин, она способна вовлекаться в регуляцию артериального давления и тонуса кровеносных сосудов. Однако реальный вклад фермента в инактивацию брадикинина in vivo не превышает 10-12%. Некоторые авторы предполагают, что КПN может играть роль модулятора действия брадикинина [57,62].
Активность КПN изменяется при воспалительных и аллергических реакциях, так как подкисление реакции среды в тканях будет способствовать накоплению кининов из-за торможения кининазной активности фермента. Поскольку фермент расщепляет пептиды, участвующие в развитии воспалительных реакций (брадикинин и анафилотоксины), и его активность в крови снижается при введении рекомбинантного интерлейкина-1b, вероятно, что КПN вовлекается в развитие воспалительных реакций [52,63].
Фермент участвует не только в процессинге энкефалинов, но и в их инактивации. Являясь основным ферментом ККС, он участвует в морфогенезе клеток, увеличении проницаемости сосудистой стенки, регуляции активности каскадных протеолитических систем плазмы крови: гемокоагуляции, фибринолиза, комплемента, развитии злокачественных новообразований [13,52].
Таким образом, КП N вносит вклад в регуляцию многих физиологических процессов и развитие патологических состояний. Поэтому, представляет интерес исследование активности фермента при онкологических заболеваниях в период химиотерапии.
1.3 Функционирование пептидергической системы при онкологических заболеваниях
Злокачественный рост характеризуется изменением активности и спектра ферментов. Ферментный спектр обусловлен локализацией, гистоцитогенезом и степенью дифференцировки опухоли. Если в дифференцированной опухоли ферменты соответствуют таковым данного органа или ткани, то при низкой степени дифференцировки активность и спектр ферментов значительно изменены. Большое значение придается ферментам в инвазии опухолевых клеток. В трансформированных клетках протеиназ синтезируется заметно больше, чем в нормальных клетках. При ряде злокачественных новообразований наблюдается выход пептидаз в межклеточное пространство и увеличение их активности. В сыворотке больных со злокачественными опухолями можно часто обнаружить увеличение активности одних ферментов и уменьшение других. Источником увеличения активности ферментов в сыворотке может являться усиленное выделение его опухолью в кровяное русло. При изучении локализации пептидазной активности в опухолях человека высокая пептидазная активность была выявлена в периферических отделах инвазивно растущих опухолей [19,26,27,41,59].
В процессе онкогенеза участвуют матриксные металлопротеазы (ММП). Транскрипция этих ферментов зависит от целого ряда факторов: цитокинов, факторов роста и некроза опухолей, химических агентов. Участие ММП в опухолевой трансформации, а также в процессах инвазии и метастазирования хорошо доказано in vitro и in vivo. Установлено, что экспрессия ММП коррелирует с деструктивными изменениями в матриксе и с туморогенным фенотипом клеток, а также зависит от вида опухоли и ткани. ММП могут участвовать в процессе канцерогенеза, воздействуя на различные пути передачи сигнала в клетке, основные компоненты соединительнотканного матрикса, на межклеточные взаимодействия, а также продуцируя различные биологически активные молекулы [35]. В опухолевых клетках может образовываться большое количество коллагеназы (представитель семейства ММП), мишенью действия которой является коллаген, представляющий собой барьер для распространения опухолевого процесса. Ингибитор её активности способен ограничивать инвазивность опухолевых клеток. Таким образом, коллагеназы ассоциированы с возникновением метастазирующего фенотипа клеток и играют ключевую роль в процессах инвазии и метастазирования.В процесс неопластической трансформации вовлечены лизосомные цистеиновые протеиназы. Ферменты деградируют многие белки и компоненты внеклеточного матрикса, осуществляют деструкцию ткани. Эти протеиназы осуществляют протеолиз короткоживущих белков, которые регулируют злокачественный рост [29,33,35,55,60].
При раке молочной железы, раке легкого и раке толстой кишки увеличенный уровень коллагеназы типа IV (матриксной металлопротеиназы типа 2, желатиназы A) - один из признаков высокого риска метастазов.
При раке мочевого пузыря повышение уровня коллагеназы типа IV в крови соответствует увеличению массы опухоли [33].
Многие опухоли характеризуются повышенной продукцией коллагеназы I типа, гидролизующей основные компоненты экстрацеллюлярного матрикса и соединительной ткани [35].
Отмечено изменение изоферментного спектра аминопептидазы. При раке легких исчезает изофермент I растворимой аминопептидазы. При злокачественных опухолях печени и желудка появляется новый пик активности фермента [18]. В лейкозных клетках, полученных от больных разными формами лимфопролиферативных заболеваний, обнаружены аминопептидазы по крайней мере двух видов: металло- и SH-зависимые ферменты. В кроветворных клетках была обнаружена аминопептидаза N, главным образом, в популяциях миелоидно-моноцитарного ряда, находящихся на разных стадиях дифференцировки, и рассматривалась как маркер этих клеток. Активность лейцил-аминопептидазы (ЛАП) усиливается в ранние сроки после наступления холестаза (механическая желтуха, вызванная злокачественными новообразованиями). Наибольшее усиление активности фермента при закупорке общего желчного протока опухолью, раке поджелудочной железы [1,8,12]. При новообразованиях печени, наблюдается увеличение активности лейцил–аминопептидазы сыворотки. В меньшей степени активность ЛАП увеличивается при метастатических поражениях печени. У 46 % больных с опухолями описано увеличение активности фермента в моче. Но его исследование не имеет диагностического значения при новообразованиях ротовой полости и области шеи [1,26].
На поздних стадиях рака легких может быть выявлено снижение активности АПФ[47,48].
При раке щитовидной железы (РЩЖ) в ткани (операционный материал) наблюдался значительный рост активности сериновых и цистеиновых протеолитических ферментов. Активность сериновых протеиназ при РЩЖ в 6 раз превышала таковую в контрольной ткани. Активность цистеиновых протеиназ при РЩЖ в 6 раз была выше, чем в контрольной ткани. Это послужило основанием для использования данных показателей как дополнительных критериев РЩЖ [35,39].
У женщин со II стадией злокачественного поражения эндометрия исследовалась пептидгидролазная активность сыворотки крови. Исследования, проведенные в разновозрастных группах доноров, не выявили достоверных изменений в активности изучаемых протеиназ. Активность трипсиноподобных протеиназ в сыворотке крови достоверно увеличивалась у онкобольных женщин и имела тенденцию к увеличению с возрастом. Активность лизосомных катепсиноподобных протеиназ в сыворотке крови онкологических больных в 2,5-2,7 раза выше по сравнению с аналогичными показателями здоровых женщин, что может свидетельствовать об инвазивной стадии данного процесса, который характеризуется нарушением целостности здоровой ткани и стабильности клеточных мембран. С другой стороны данный процесс может сопровождаться усилением биосинтеза изучаемых протеиназ или снижением биосинтеза их эндогенных ингибиторов [19,35].
В облученной опухолевой ткани при раке почки было обнаружено, повышение активности ингибиторов сериновых и цистеиновых протеиназ в 4 и 5,5 раз, и снижение ферментативной активности на 74% и 45% для сериновых и цистеиновых протеиназ, соответственно. При сопоставлении активности протеолиза в опухолевой ткани после лучевой терапии и в необлученной опухолевой ткани обнаружено снижение ферментативной активности сериновых и цистеиновых протеиназ. Их активность под влиянием лучевой терапии уменьшалась, составляя всего 7% и 20% от активности до облучения, и после облучения не изменялась, а активность ингибитора цистеиновых протеиназ – увеличивалась в 2 раза [35,59,60].
При злокачественных образованиях толстой кишки активность нейтральных и кислых протеиназ в опухолевой ткани в 1,76 – 2 раза выше, чем в неизмененной ткани кишки. После предоперационного облучения прямой кишки активность протеиназ в облученной опухоли оказывается в 2,8 – 3,58 раза ниже, чем в опухоли не подвергнутой облучению [1,11].
В сыворотке крови онкологических больных (рак легкого, рак молочной железы) исследовали некоторые компоненты калликреин-кининовой системы, а также активность "нейтральных" и "кислых" протеаз и их ингибиторов до и после терапевтического нейтронного облучения. Исследования показали, что предоперационное облучение быстрыми нейтронами на область первичного очага тормозит процессы кининообразования в крови онкологических больных: на 20% снижается уровень калликреина на фоне увеличения содержания неактивного профермента. Общая активность сериновых протеиназ остается неизменной, однако отмечается уменьшение концентрации основного ингибитора калликреина – a2
-макроглобулина. На этом фоне повышается активность кининоразрушающего фермента карбоксипептидазы N. Вместе с тем, активность "нейтральных" и "кислых" протеаз после облучения имеет четко выраженную тенденцию к уменьшению, а уровень общей антипротеолитической активности остается практически неизменным. При этом отмечается стабилизация состояния клеточных мембран. Между степенью торможения образования протеолитических ферментов и клинически определяемой регрессией размеров опухоли существует достоверная корреляционная зависимость [35,62].
1.4 Роль оксида азота (
II
) в онкогенезе
Оксид азота II обладает мультипотентными свойствами, определяемые как цитотоксичностью радикала, так и его коммуникативной активностью. При онкологических заболеваниях эта молекула может проявлять и противоопухолевые свойства, и участвовать в патогенезе неоплазий. NO необходим для обеспечения цитотоксического действия макрофагов на опухолевые клетки [23,42]. Выделяемый макрофагами, NO подавляет опухолевые клетки либо блокируя их железосодержащие ферменты, либо повреждая их клеточные структуры. Вызывая повреждение ДНК, NO включается в модуляцию апоптоза: активирует экспрессию p53, который вызывает задержку деления клеток в фазе G1. В то же время p53 по принципу обратной отрицательной связи подавляет синтез iNOS и таким образом предохраняет трансформированные клетки от гибели. Установлено, что синтез iNOS индуцируется TNFa. Уже через 6 – 12 ч. после действия индуктора NO достигает уровня, при котором начинает сказываться его влияние на опухолевые клетки [24,28]. NO участвует в образовании новых сосудов, что необходимо для удовлетворения потребностей раковых клеток в питании. С другой стороны, вследствие этого улучшается доставка оксида азота в опухолевые клетки. NO, выделяемая эндотелиальными или опухолевыми клетками, может стимулировать процесс инвазии в стенку сосуда, поскольку прикрепление к эндотелию сосудов необходимое звено метастазирования. Радикал обладает дезагрергирующим действием. Поэтому следует ожидать, что усиленный синтез NO подавляет межклеточное сцепление клеток в опухоли и облегчает их распространение по организму. Клеткам с минимальной продукцией NO будет труднее отделяться от первичной опухоли и образовывать метастазы. [42].
Таким образом, существуют механизмы поддерживающие баланс между участием NO как в прометастатических, так и в антиопухолевых реакциях [42].
ГЛАВА 2. Материалы и методы исследования
2.1 Материал исследования
Активность ферментов определяли в сыворотке крови онкологических больных. Кровь брали из локтевой вены, далее ее инкубировали 30 минут при комнатной температуре, центрифугировали 20 минут при 4000 g и получали сыворотку, в которой определяли активность карбоксипептидазы N и ангиотензинпревращающего фермента.
Изучение активности ферментов было проведено у 24 мужчин в возрасте от 49 до 70 лет с опухолями легких, желудочно-кишечного тракта, мочеполовой системы. Больные подвергались химиотерапевтическому воздействию. Они составили две экспериментальные группы. Первая группа – больные до начала проведения им химиотерапии (12 человек); вторая – те же больные, но после окончания курса лечения (12 человек). В качестве контроля выступала группа из 13 здоровых мужчин такого же возраста.
2.2 Методы исследования
2.2.1 Метод определения активности карбоксипептидазы
N
Активность КПN определяли в сыворотке крови нингидриновым методом [61].
Опытные пробы содержали 20 мкл 3,5 мМ раствора CoSO4
, приготовленного на 100 мМ Трис-HCl буфере, pH 7,6 и 40 мкл препарата фермента. Контрольные пробы содержали 20 мкл 100 мМ Трис-HCl, pH 7,6 и 40 мкл препарата фермента. Пробы преинкубировали 8 мин при 37o
C, реакцию начинали прибавлением в опытные пробы 10 мкл гиппурил-арг, приготовленного на 100 мМ Трис-HCl буфере, pH 7,6 (конечная концентрация в пробе 5 мкМ). Реакцию проводили 120 мин при 37o
C и останавливали прибавлением 30 мкл 10 % трихлоруксусной кислоты.
Пробы центрифугировали 20 мин при 4000 об/мин, отбирали 50 мкл надосадочной жидкости, приливали 1 мл нингидринового реактива. Далее пробы встряхивали, выдерживали 12 мин на кипящей водяной бане и измеряли оптическую плотность на КФК-2 при 595 нм против H2
O.
Активность КПN определяли как разность оптической плотности опытных и контрольных проб и выражали в нмоль аргинина, образовавшегося за 1 мин инкубации в пересчете на 1 мг белка.
Концентрацию белка в пробах определяли биуретовым методом.
2.2.2 Метод определения активности ангиотензинпревращающего фермента
Активность АПФ определяли в сыворотке крови нингидриновым методом по образованию гли-арг из кбз-гли-гли-арг при рН 8,2 как активность, ингибируемую каптоприлом. Препарат фермента (40 мкл) смешивали с 20 мкл 35 мкМ каптоприла в 100 мМ Трис-НСl буфере, рН 8,2, или 20 мкл буфера и преинкубировали 8 мин при 37˚С. Реакцию начинали прибавлением 10 мкл раствора кбз-гли-гли-арг в вышеуказанном буфере (конечная концентрация в пробе 5 мкМ). Через 120 мин реакцию останавливали прибавлением 30 мкл 10% раствора трихлоруксусной кислоты [59]. Пробы центрифугировали 30 мин при 4000 об/ мин. Отбирали 50 мкл надосадочной жидкости и определяли количество образовавшегося гли-арг нингидриновым методом [61]. Пробы колориметрировали на КФК-2 при l=590 нм. Активность АПФ определяли как разность в оптической плотности проб не содержащих и содержащих каптоприл. Активность фермента выражали в нмоль гли-арг, образовавшегося за 1 мин инкубации в пересчете на 1 мг белка. Концентрацию белка определяли биуретовым методом.
2.2.3 Метод определения содержания белка
Содержание общего белка определяли биуретовым методом. Принцип метода основан на том, чтоионы меди в щелочной среде взаимодействуют с пептидными связями белков сыворотки крови с образованием комплекса красного цвета, интенсивность окраски которого пропорциональна концентрации общего белка.
Опытные пробы содержали 50 мкл сыворотки крови и 2,5 мл рабочего раствора биуретового реагента (концентрат биуретового реагента развести дистиллированной водой в соотношении 1:19). Контрольные пробы содержали 50 мкл дистиллированной воды и 2,5 мл рабочего раствора биуретового реагента. В калибровочную пробу добавляли 50 мкл калибровочного раствора общего белка (раствор бычьего сывороточного альбумина с концентрацией 60 г/л с добавлением хлористого натрия 9 г/л и азида натрия 1 г/л).
Содержимое пробирок тщательно перемешивали, избегая образования пены, инкубировали при комнатной температуре (+ 18 - 25 С) в течение 30 минут, после чего измеряли величину оптической плотности калибровочной и опытных проб против контрольной пробы при длине волны 540 нм. С) в течение 30 минут, после чего измеряли величину оптической плотности калибровочной и опытных проб против контрольной пробы при длине волны 540 нм.
Концентрацию общего белка рассчитывали, как отношение оптической плотности опытной пробы к оптической плотности калибровочной пробы умноженная на концентрацию общего белка в калибровочном растворе (60 г/л).
2.2.4 Метод количественного определения
NO в сыворотке крови
Для определения концентрации NO0,5 мл сыворотки депротеинизировали добавлением 1 мл 0,5 н. раствора NaOH и 10 % раствора ZnSO4
для осаждения белковых компонентов. Пробы центрифугировали при 5000 об/мин. 30 мин. К 1мл надосадочной жидкости добавляли равное количество 1 % раствора реактива Грисса и инкубировали в течение 10 мин. при 25С. Затем колориметрировали при 540 нм против контрольной пробы. Содержание NO оценивали по количеству конченого стабильного метаболита оксида азота (II) – нитрит иона. Суммарную концентрацию нитрит-иона определяли колориметрически по развитию окраски в реакции диазотирования нитритом сульфаниламида, входящего в реактив Грисса. Количество нитрит-иона выражали в мкг на мл.
2.3 Статистическая обработка результатов исследования
Достоверность отличий между средними определяли с использованием t-критерия Стьюдента [16]. Корреляционный анализ проводили с помощью программы «Статистика» (версия 6.0).
ГЛАВА 3. Результаты и обсуждение
3.1 Исследование активности карбоксипептидазы
N в сыворотке крови онкологических больных при химиотерапевтическом воздействии
Результаты исследования показали увеличение активности КПN у онкологических больных в период до проведения химиотерапии по сравнению с контрольной группой в 2 раза, и уменьшение активности фермента после проведения химиотерапии, по отношению к периоду до начала лечения в 2,6 раза(рис.1).
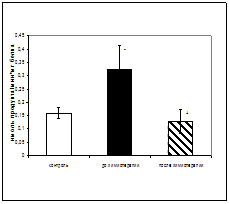
Рис.1. Активность КПN у онкологических больных при химиотерапевтическом воздействии (нмоль продукта, образовавшегося за 1 мин инкубации на 1 мг белка, M±m, n=12¸18; достоверность отличий: * - p< 0,05 относительно контроля, + - p<0,05 относительно до химиотерапии).
Калликреин-кининовая система, к которой относится и КПN играет не только центральную роль в процессах гемокоагуляции и фибринолиза, но и контролирует различные стадии развития злокачественных новообразований [52,63]. Возрастание активности КПN у онкологических больных, вероятно, опосредованно через уменьшение уровня брадикинина может препятствовать высвобождению фактора некроза опухолей (TNFa), способного лизировать большой набор опухолевых клеток invivo и invitro. Цитотоксическое действие TNFa на
опухолевую клетку связано с деградацией ДНК и нарушением функционирования митохондрий. Таким образом, увеличение активности КПN может способствовать прогрессии опухоли. Снижение активности КПNпосле химиотерапии до уровня контрольных значений может указывать на существенный противоопухолевый эффект данного терапевтического воздействия. Таким образом, динамическое исследование активности фермента в сыворотке крови больных до и после химиотерапии может быть использовано в качестве дополнительных биохимических критериев при наблюдении за эффективностью проводимого лечения [6,33].
3.2 Исследование активности ангиотензинпревращающего фермента
в сыворотке крови онкологических больных при химиотерапевтическом воздействии
Результаты исследования показали, что активность АПФ у онкологических больных до химиотерапии соответствовала контрольным значениям, однако после проведения лечения активность фермента возрастала по сравнению с двумя другими группами в 1,8 и 1,7 раза соответственно (рис.2).

Рис. 2. Активность АПФ у онкологических больных при химиотерапевтическом воздействии (нмоль продукта, образовавшегося за 1 мин инкубации на 1 мг белка, M±m, n=12¸18; достоверность отличий: * - p<
0,05 относительно контроля; ++ - p< 0,01 относительно до химиотерапии).
Известно, что наличие опухоли в организме приводит к нарушениям гемостаза. Также известно об отрицательном влиянии химиопрепаратов на свертывающую систему крови и гемостаз [30,37]. Таким образом, химиотерапия лишь усугубляет уже имеющиеся в организме нарушения системы свертывания крови. Возможно, именно с этим связано повышение активности АПФ после проведения лечения. Можно предположить, что свой эффект по усилению тромботических осложнений АПФ реализует двумя путями: через синтез ангиотензина II и через деградацию брадикинина. Ангиотензин II является вазопрессором, стимулятором образования свободных радикалов, в частности супероксидных анионов, которые инактивируют NO, промотируют образование пероксинитрита и снижают эффективность NO-опосредуемой сосудистой дилатации. Таким образом, ангиотензин II способствует развитию эндотелиальной дисфункции, в результате чего усиливается выработка вазоконстрикторов и прокоагулянтов, и как следствие активируется система свертывания. В то время как брадикинин стимулирует синтез NO в эндотелии, участвуя в дилатации сосудов и усиливая скорость местного кровотока [52,54].
Повышение активности фермента после лечения возможно также связано с проявлением нефротоксичности препаратов-производных платины (именно эти препараты применялись при химиотерапии) [44]. Вероятно, что поражение клеток почек, которые содержат значительное количество АПФ, приводит к выделению фермента в кровь и увеличению его активности.
Одновременное отсутствие достоверных отличий в активности АПФ у онкологических больных в периоде до химиотерапии и здоровых людей, и наличие нарушений в свертываемости крови у первых, может указывать на то, что в формировании патологий гемостаза участвуют и другие механизмы.
3.3 Исследование содержания нитрит-иона в сыворотке крови у онкологических больных при химиотерапевтическом воздействии
Результаты исследования показали, что содержание нитрит-иона в сыворотке крови онкологических больных до проведения им химиотерапии уменьшается по сравнению с контрольной группой на 19,4 %. После проведения химиотерапии концентрация нитрит повышается по отношению к периоду до лечения на 12 %, но остается ниже контрольных значений на 8,4 %(рис.3).

Рис.3 Содержание NO2
-
в сыворотке крови онкологических больных при химиотерапевтическом воздействии (мкг/мл, достоверность отличий: *- p<0,05 относительно контроля; *** - p<0,001 относительно контроля; ++ - p< 0,01 относительно до химиотерапии).
NO может выступать не только как индуктор неоплазий, но и как эффектор противоопухолевой защиты. Секретируемый клетками, инфильтрующими опухоли (макрофаги, лимфоциты, нейтрофилы), радикал выполняет роль противоопухолевого агента, т.е. способен уничтожать опухолевые клетки и остановить их рост [23,42,53]. Согласно Hibbs и соавт. цитостатический и цитотоксический эффекты макрофаги осуществляют посредством NO. При активации T-лимфоцитами макрофаги активируют синтез iNOS, которая превращает аргинин в NO. Оксид азота IIвыделяется из макрофагов и проникает в опухолевые клетки. Там молекула ингибирует три группы ферментов: митохондриальной дыхательной цепи, цикла Кребса и синтеза ДНК. В этих условиях энергопродукция и деление клеток становится невозможным и клетка погибает [24]. Таким образом, снижение содержания NO может быть вызвано ослаблением защиты (иммунной системы) организма и является одним из факторов, приводящих к развитию опухолей.
Вероятно, более высокий уровень NOв сыворотке крови после химиотерапии объясняется усилением его продукции под влиянием ряда цитостатиков [28]. Возможно, что частично, терапевтическая активность цитостатиков зависит от этой реакции, аувеличение концентрации оксида азота II в данной ситуации может расцениваться как благоприятный прогностический признак. Таким образом, управление синтезом NO может иметь значение для повышения эффективности противоопухолевой терапии.
Достоверная отрицательная взаимосвязь между увеличением активности КПN на фоне снижения концентрации оксида азота II в сыворотке крови до химиотерапии (табл.), возможно вызвано тем, что повышение активности КПN приводит к подавлению высвобождения фактора некроза опухоли ТNFa, который индуцирует синтез iNOS, это приводит к такому снижению продукции NO, при котором он не оказывает противоопухолевого воздействия [24]. Полученные данные свидетельствуют о генерализованном снижении естественных защитных сил организма.
Таблица.
Корреляционная взаимосвязь содержания NO и активности ферментов
| Активность ферментов |
Содержание NO |
Активность АПФ |
Активность КПN |
| До химио- терапии |
После химио-
терапии
|
До химио- терапии |
После химио-
терапии
|
До химио- терапии |
После химио-
терапии
|
| АПФ до химиотерапии |
—— |
—— |
—— |
—— |
| АПФ после химиотерапии |
—— |
-1,00*** |
—— |
—— |
| КПN до химиотерапии |
-1,00*** |
—— |
—— |
| КПN после химиотерапии |
—— |
—— |
—— |
Достоверность корреляционных взаимосвязей: —— достоверной корреляции не обнаружено, *** - p < 0,001.
Отрицательная корреляция между активностью АПФ и уровнем NO, по всей видимости, связана с тем, что фермент, опосредованным образом вызывает деградацию оксида азота II [42]. Вероятно, по этой причине после химиотерапии на фоне высокой активности АПФ концентрация NOу пациентов оказывается ниже, чем в контрольной группе.
Выводы
1. Обнаружено увеличение активности ангиотензинпревращающего фермента у онкологических больных прошедших химиотерапию по сравнению с периодом до начала лечения и контрольной группой.
2. Обнаружено увеличение активности карбоксипептидазы N у онкологических больных до проведения химиотерапии по сравнению с контрольной группой и больными, прошедших химиотерапию.
3. Обнаружено более низкое содержание оксида азота (II) у онкологических больных по сравнению с контрольной группой и повышение уровня оксида азота (II) после проведения химиотерапии по сравнению с периодом до лечения.
4. Полученные результаты представляют интерес для понимания биологической роли ангиотензинпревращающего фермента и карбоксипептидазы
N при онкологических заболеваниях с различной локализацией опухоли и могут быть использованы для разработки методов профилактики и коррекции различных нарушений, возникающих до и после химиотерапии.
Список литературы
1. Абдурасулов Д.М. Множественные опухолевые поражения. – Т.: Медицина, 1980. – 326с.
2. Абелев Г.И. Что такое опухоль // Соросовский образовательный журнал. – 1997. - №10. – С. 85- 90
3.Альтшулер Б.Ю., Ройтман А.П., Долгов В.В. Методические аспекты определения ангиотензинпревращающего фермента // Клиническая лабораторная диагностика. – 2000. – №12. – С. 10– 14.
4. Антонов В.К. Химия протеолиза. – М.: Наука, 1991. – 503с.
5. Балаж А. Биология опухолей. Сомнения и надежды. Пер с венг.: – М.: Мир, 1987. – 206с.
6. Бессмельцев С.С., Рыбакова Л.П., Грицкевич Н.Л. Диагностическое и прогностическое значение определения церулоплазмина, ацетилхолинэстеразы и общей протеолитической активности в сыворотке крови больных множественной миеломой // Вопросы онкологии. – 1999, том 45. - № 6. – С.398 – 403.
7. Блохин Д.Ю. Фенотип множественной лекарственной устойчивости опухолевых клеток, обусловленный нарушением программы клеточной гибели // Вестник Российской академии медицинских наук. – 2004. - № 12. – С.16 – 19.
8. Блохин Н.Н., Итин А.Б., Клименков А.Г. Рак поджелудочной железы и внепеченочных желчных путей. – М.: Медицина,1982. – 272с.
9. Блохин Н.Н., Переводчикова Н.И. Химиотерапия опухолевых заболеваний. – М.: Медицина, 1984.
10. Бычков М.Б. Химиотерапия злокачественных опухолей // Архив патологий. – 1996, том 58. - №4. – С. 15 – 17.
11. Варбанец В. Ф. Активность протеолитических ферментов и их ингибиторов в опухолях толстой кишки // Вопросы медицинской химии. -1990, том 36. -№2. – С. 33 – 35.
12. Василенко В.Х. Опухоли желудка. Клиника и диагностика. – М.:Медицина,1989. – 288с.
13. Верееменко К.Н. Кининовая система. – К.: Здоровье, 1977.
14. Вернигора А.Н., Генгин М.Т. Протеолитические ферменты: субклеточная локализация, свойства и участие в обмене нейропептидов // Биохимия. – 1996,т.6, №5. – С. 771– 785.
15. Гершанович М.Л. Осложнения при химио- и гормонотерапии злокачественных опухолей. – М.: Медицина, 1982. – 224с.
16. Гланц С. Медико-биологическая статистика: Пер. с англ. – М, Практика, 1998. – 459с.
17. Гнатышак А.И. Общая клиническая онкология. – Львов: Выща шк., 1988. – 238с.
18. Голубев А.М. Изоферменты новообразований. – М.: Медицина, 1981. – 144с.
19. Диагностика и лечение злокачественных новообразований. Сборник научных трудов. / Кавецкий Р.Е. – К.: Наукова думка,1979. – 180с.
20. Елисеева Ю.Е. Ангиотензинпревращающий фермент, его физиологическая роль. // Вопросы медицинской химии. – 2001. – № 1. – С. 53 – 60.
21. Елисеева Ю.Е. Структурно-функциональные особенности ангиотензинпревращающего фермента. // Биоорганическая химия. – 1998, том 24. – №4. – С. 262 – 270.
22. Зеленин К.Н. Возникновение и развитие химиотерапии. // Соросовский образовательный журнал. – 2001, том 7. – №5. – С. 23 – 28.
23. Зеленин К.Н. Оксид азота (II): новые возможности давно известной молекулы. // Соросовский образовательный журнал. – 1997. – №10. – С. 109 – 110.
24. Ивашкин В.Т., Драпкина О.М. Клиническое значение оксида азота и белков теплового шока. – М.: Геотар-мед, 2001. – 87с.
25. Клиническая онкология: справочное пособие. / З.С. Фрадкин, И.В. Залуцкий – Мн.: Беларусь,2003. – 784с.
26. Клиническая ферментология. / Э. Щеклик. – Варшава: Польское государственное медицинское изд-во, 1966. – 491с.
27. Колодзейская М.В., Пилявская А.С. Пептидазы. – К.: Наукова думка,1982. – 173с.
28. Кондакова И.В., Какурина Г.В., Чойнозов Е.Л. Влияние доноров оксида азота на противоопухолевый эффект доксорубицина. // Бюллютень СО РАМН. – 2005. – №2(116). – С.92 – 95.
29. Крутяков В.М. Биотерапия рака и ингибиторы протеаз. // Вопросы онкологии. – 2001, том 47. – №1. – С. 106 – 107.
30. Кулиев Ш.Б. Проблемы свертывания крови в онкологии. – Б.: Азернешр, 1979. – 273с.
31. Лебедев А.А. Система ренин-ангиотезин. // Соросовский образовательный журнал. – 1998. – №3. – С. 35 – 40.
32. Ленинджер А., Основы биохимии, пер. с англ., т. 1 3, M., 1985.
33. Локшина Л.А. Протеолитические ферменты в процессах онкогенеза. // Вопросы медицинской химии. – 1991, том 37. – №6. – С.15 – 21.
34. Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: В 2-х томах. Т.2. Пер. с англ.: – М.: Мир, 1993. – 384с.
35. Материалы конференции “Структура и функция протеолитических ферментов”. // Вопросы медицинской химии. – 2000. – №5.
36. Машковский М.Д. Лекарственные средства: В 2-хтомах Т.1. – М.: ООО “ Изд-во новая волна” , 2002. – 540с.
37. Онкология. / Н.Н. Трапезников. – М.: Медицина, 1981. – 255с.
38. Патологическая физиология и биохимия: Учебное пособие для вузов / - М.: Издательство «экзамен», 205. – 480с.
39. Пачес А.И., Пропп Р.М. Рак щитовидной железы. – М.: Медицина, 1984. – 320с.
40. Переводчикова Н.И. Клиническая химиотерапия опухолевых заболеваний. – М.: Медицина, 1976. – 200с.
41. Поляк С.М., Рожанская Т.И., Яковлева Е.П. Регуляторы активности ферментов и их применение в медицине. – М.: Медицина, 1989. – 128с.
42. Проскуряков С.Я., Конопляников А.Г., Иванников А.И. Оксид азота в неопластическом процессе. // Вопросы онкологии, 2001, том 47. – №3. – С. 257 – 266.
43. Противоопухолевая химиотерапия. Справочник. / Н.И. Переводчикова. – М.: Медицина, 1993. – 155с.
44. Противоопухолевые препараты: справочник. / З.П. Булкина. – К.: Наукова думка,1991. – 304с.
45. Сивашский М.С., Салганин Р.И. Гетерогенность раковых клеток в отношении их реакции на цисплатину. // Вопросы онкологии, 2001,том 47. – №1. – С. 66 – 68.
46. Слинчак С.М. Онкология. – К.: Выща шк.,1989.
47. Трахтенберг А.Х. Рак легкого. – М.: Медицина, 1987. – 304с.
48. Харченко В.Г., Кузьмин И.В. Рак легкого. Руководство для врачей. – М.: Медицина, 1994. – 480с.
49. Цыганенко А.Я., Жуков В.И., Мясоедов В.В., Завгородний И.В. Клиническая биохимия ( учебное пособие для студентов медицинских вузов). – М.: Триада – Х, 2002. – 504с.
50. Черенков В.Г. Клиническая онкология. – М. Медицина, 2005. – 447с.
51. Чернов В.А. Цитостатические вещества в химиотерапии злокачественных новообразований. – М.: Медицина, 1964. – 319с.
52. Яровая Г. А. Калликреин-кининовая система: новые факты и концепции (обзор) // Вопросы медицинской химии. – 2001, том 47. – № 1. – с. 20-42.
53. Ahmad H., Srivastova R.C., Agarwal R., Mukhtar H. Nitric oxide synthase and skin tumor promotion. // BBRC. – 1997. – Vol.232. – P. 328 – 331.
54. Blume A., Herdegen T., Unger T. Angiotensin peptides and inducible transcription factors. J. Mol. Med.77. – P. 339 – 357.
55. Boyd D. Invasion and metastasis. // Cancer Metastasis Rev. – 1996. – Vol.15. – P. 77 – 89.
56. Camdell DJ. Circulating and tissue angiotensin system. J. Clin. Invest., 1987. – Vol.79. – №1-6.
57. Erdos E.G., Sloane E.M., Wohler I.M. Carboxypeptidase in blood and other fluids // Biochem. Pharmacol. – 1964. – №13. – P. 893– 905.
58. Fulong Tan., Deepthi K., Skidgel R.A., Kaul R.K., Erdos E.G. The deduced protein sequence of the human carboxypeptidase N high molecular weight subunit reveals the presence of leucine-rich tandem repeats. J. of biological chemistry, 1990. – Vol.265. – №1. – P. 13 – 19.
59. Kennedy A.R. Cancer prevention by protease inhibitors. // Prev. Med/ - 1993. – Vol.22. – P. 796 – 811.
60. Kennedy A.R. Chemopreventive agents: protease inhibitor. // Pharmacol. Ther. – 1998. – Vol.78. – P. 167 – 209.
61. Lee P.V., Takahashi T.N. An improved colorimetric determination of amino acid with the use of ninhydrin // Analyt. Biochem. – 1966. – 14, № 1. – P. 71– 77.
62. Skidgel R.A., Erdos E.G. Cellular carboxypeptidases // Immunol. Rev. – 1998. – V. 161, № 2. – P. 129-141.
63. The kallikrein – kinin system in health and disease. – Munich: Limbach – verlag braunschweig. – 1989. – P. 374.
|