РЕКОМЕНДАЦІЇ З ВИКОРИСТАННЯ СУЧАСНИХ МЕТОДІВ ВИЗНАЧЕННЯ СТАТІ МОЛОДНЯКУ СІЛЬСЬКОГОСПОДАРСЬКОЇ ПТИЦІ
ХАРКІВ 2007
1. Огляд методів визначення статі молодняку свійської птиці
Навчитися визначати стать добового молодняку птахівники намагалися вже давно. Відомі, наприклад, два стародавніх методи, що дозволяють розділяти добових курчат на курочок і півників з точністю 60–65%. В основі цих простих прийомів визначення статі лежать розходження в рефлекторних реакціях самців і самок на дискомфортне положення курчат (рис. 2).
Відповідно до першого способу, курчат піднімають за шию, при цьому ноги у півника в такому положенні будуть висіти рівно опущеними вниз, тоді як курочка буде намагатися підняти їх і корчити пальці.
Згідно до іншого способу, курчат потрібно тримати за лапки головою вниз, при цьому півник висить спокійно, а курочка намагається піднімати голову.
 |
 |
| Півник |
Курочка |
Півник |
Курочка |
Стародавні способи визначення статі курчат
Точність цих двох прийомів сексування курчат можна трохи підвищити, якщо врахувати, що у добових півників у порівнянні з курочками голова більш округла, помітніший гребінець, товщі ноги й сильніше загнути й дзьоб. Та й важать півники в середньому на один-два грамів більше, ніж курочки.
Визначення статі каченят за Н.В. Сидоровим. Простий спосіб визначення статі добових каченят запропоновано більше 50-ти років тому асистентом кафедри птахівництва Московської сільськогосподарської академії ім. К.А.Тімірязєва Н.Н. Сидоровим [13]. В основу цього способу визначення статі добових каченят свійської качки (але не мускусної) положено статеві розходження в будові нижньої гортані. У селезнів нижня частина гортані при вході в грудну клітку помітно розширена й виконує функцію резонатора. Це розширення має округлу форму діаметром 3–5 мм і добре прощупується пальцями оператора.

Місце розташування гортанного резонатора у селезня та положення каченяти в процесі визначення статі
Стать каченят визначається відразу ж після вибірки їх з інкубатора. Це пов’язане з тим, що нижня частина гортані поступово опускається в грудну клітку й чим старше каченя, тим важче прощупати резонатор у самців. Для визначення статі каченят цим способом його беруть у праву руку, при цьому пальці оператора торкаються грудки пташеняти.
Лівою рукою оператор відводить голівку каченят вперед від себе і піднімає догори дзьоб. Великий палець правої руки сортувальник прикладає до шийних хребців каченят, чим створює упор для його тіла в момент визначення статі. Одночасно з цим, вказівним пальцем правої руки оператор прощупує місце в нижній частині шиї. Місце для промацування рухливого резонатора у самців обмежено зверху двома нерухливими горбками (зрощення ключиць із лопатками), а знизу – одним горбком (зрощення ключиць із грудною кісткою). У центрі цього трикутника у селезнів і знаходиться четвертий рухливий горбок – резонатор, завбільшки з невелику горошину. У самочок же такого горбка не має, що й дає можливість визначати стать добових каченят.
Точність сексування каченят даним способом становить 94–98% при швидкості близько 300 гол./година. Однак слід зазначити, що ця процедура визначення статі достатньо трудомістка й нерідко призводить до травмування каченят, що, у свою чергу, викликає збільшення відходу молодняку в процесі його вирощування.
Японський метод (вентсексинг). На початку ХХ століття в Японії була розроблена процедура визначення статі добових курчат шляхом візуального обстеження клоаки й виявлення на її внутрішній стінці статевого горбка, що істотно розрізняється за формою й величиною в півників і курочок [14, 15]. У цей час японський метод визначення статі молодняку широко використовується у світовому птахівництві. У країнах СНД висококваліфіковані оператори з багаторічним стажем роботи сексують японським методом за 1 годину 600–800 добових курчат із середньою точністю 92–96%. Швидкість сексингу індичат, каченят і гусенят трохи нижче (500 гол./годину) при точності: для індичат – 88–92%, гусенят і каченят – 96–99%. У процесі визначення статі молодняку цим методом можливе травмування й перезараження пташенят патогенною мікрофлорою кишечника.
Генетичні методи (аутосексинг). Аутосексинг базується на розведенні спеціально виведеної міченої за статтю птиці, у якої статева приналежність добового молодняку визначається за забарвленням пуху (колорсексинг) або типами опереності крила (федерсексинг) [16,17]. У сучасному птахівництві особливо широко поширені аутосексні кроси яєчних і м’ясних курей, у яких як маркери статі використовуються системи альтернативних алелей – «сріблястість – золотистість» або «смугастість – однотонність» забарвлення оперення, а також «рання – пізня» опереність молодняку. Аутосексинг забезпечує високу точність (98–100%) і швидкість (1,5–7 тис. гол./годину) сортування молодняку за статтю. Він нешкідливий для пташенят, високотехнологічний і не вимагає тривалого навчання операторів-сортувальників.
Сексування молодняку за допомогою технічних засобів. Спроби спростити процедуру визначення статі молодняку за допомогою технічних засобів здійснювались неодноразово. При цьому, розробка різних по своїй суті методів проходила за приблизно однаковою схемою. Спочатку, дослідники намагалися знайти різницю між пташенятами чоловічої та жіночої статей за їх морфологічними, анатомічними, акустичними або фізіологічними ознаками. Якщо вдавалося виявити чіткий статевий диморфізм, то після цього розроблялася чутлива техніка сексування, яку можна було б автоматизувати й широко застосувати у виробничих масштабах. Нижче наведено методи сортування добового молодняку за статтю із застосуванням різних приладів або технічних пристроїв.
Зондовий метод. У середині XX століття в Японії був розроблений спосіб визначення статі молодняку за допомогою телескопічного пристрою, названого «чик-тестер» [13, 18]. Цей оптичний пристрій складається із трубки-окуляра зі скляним порожнім стрижнем (світловодом), що вводять через клоаку в кишечник курчат для огляду черевної порожнини. По осі скляного стрижня направляється потужний пучок світла, який висвітлює через тонку стінку кишечника статеві залози – сім’яники у півника, яєчник – у курочки, які розташовані поблизу хребта в області хрестця. У полі зору оператора у півників видно один із двох молочно-білих сім’яників, що мають форму рисових зерен приблизно однакових за розміром (рис. 4). У курочок же спостерігається плоский блідо-рожевий яєчник трикутної форми, розташований, як правило, з лівої сторони. Правий яєчник у самочок слабо розвинутий.
Спосіб введення стрижня «чик-тестера» через клоаку показаний на схемі. Оптична система приладу дозволяє операторові бачити сім’яники або яєчник пташенят, збільшені в 3–5 раз. Швидкість сексування молодняку зондовим методом – 400–500 гол./годину при точності визначення статі 95 – 98%.
Зондовий метод має ряд недоліків, властивих японському. Праця операторів монотонна й стомлююча, продуктивність праці при цьому невисока, а сам метод складний у здійсненні й вимагає тривалого навчання. До того ж у процесі визначення статі «чик-тестером» не виключається стресування, травмування й перезараження молодняку збудниками кишкових інфекційних захворювань. Все це знижує привабливість даного методу для промислового птахівництва.
Акустичний метод. Статевий диморфізм у будові нижньої гортані у добового молодняку приводить до розходжень у спектральній структурі звукових сигналів дискомфорту у самців і самок вже в момент вилуплення. Це явище положено в основу одного зі способів визначення статі добового молодняку птиці, пріоритет у розробці якого належить російським дослідникам [19–23, 176].
Застосування способу вимагає спеціальної електронно-акустичної апаратури, яка дозволяє визначати стать курчат яєчного напрямку продуктивності з точністю до 90%, м’ясного напрямку – з точністю до 80%, гусенят, каченят, індичат, цесарят і перепелят – до 85 – 95%, а пташенят деяких диких видів – до 90%.
Спосіб реалізується наступним чином. Для одержання сигналів дискомфорту добове пташеня опускають долілиць головою перед мікрофоном, з’єднаним з електронним аналізатором. У такому положенні пташеня, як правило, видає звуковий сигнал, що негайно обробляється по заданій програмі електронно-акустичним пристроєм. За результатами аналізу на приладі загоряється одна із трьох кольорових лампочок: червона – тестоване пташеня – самець, синя лампочка – самочка, зелена – стать пташеняти не визначена.
До недоліків даного способу можна віднести невисоку швидкість визначення статі молодняку (не більше 500 гол./година), потребу в дорогій електронно-акустичній апаратурі, знижену точність сексування, а також значне стресування пташенят.
Цитогенетичний метод. Описано методику сексування добових курчат за каріотипом клітин пульпи пера [24]. У курей (як і в інших видів птиці) дивергенція статевих хромосом зайшла далі, ніж у ссавців. «Чоловіча» статева Z-хромосома – самий довгий метацентрик каріотипу, тоді як «жіноча» статева W-хромосома є субметацентриком, що приблизно в 10 разів менше Z-хромосоми. Досліджуючи мітози кліток пульпи пера звичайним цитогенетичним методом, можна за кількістю Z-хромосом визначити стать пташенят. Якщо в каріотипі аналізованого пташеняти дві Z-хромосоми, значить це півник, а якщо тільки одна – курочка. Цитогенетичний метод визначення статі абсолютно точний, але трудомісткий і нетехнологічний. Найбільш виправдане його використання з метою генетичної експертизи.
Молекулярно-генетичний метод. Після того, як японські дослідники [25, 26] визначили й клонували специфічну нуклеотидную послідовність в W-хромосомі, була показана можливість визначення статі у курчат за допомогою нового методу блотт-гібридизації ДНК крові аналізованої особини зі специфічним праймером [27]. Стать молодняку чітко визначається не тільки з використанням для аналізу проб очищеної ДНК еритроцитів курчат, але й при дослідженні цільної крові. Варто підкреслити, що дана молекулярно-генетична процедура безпомилкового визначення статі молодняку поки ще трудомістка й дорого коштує.
Ваговий метод. Вивчення живої маси добових самців і самок домашньої птиці показало, що у більшості видів і порід добові самці в середньому важчі за самок [28–30]. За ступенем зменшення статевого диморфізму одноденних пташенят за масою тіла (розходження даються у відсотках) обстежені види птиці розташувалися в такій послідовності: мускусна качка (5,12%), фазан мисливський (5,08%), індичка (4,6%), гусак домашній (2,3%), курка домашня (1,8–2,5%), домашня качка (0,4%), японська перепілка (-0,1%), цесарка (-0,2%).
Оскільки статеві розходження в живій масі у добових мускусних каченят, фазанят, індичат і курчат м’ясних порід статистично вірогідні, то їх можна покласти в основу простого методу сексування молодняку шляхом його індивідуального зважування. Хоча ваговий метод сортування пташеняти дає низьку точність визначення статі (60–70%), він все-таки дозволяє створити рівновагові групи молодняку на перших етапах його вирощування.
Для практичної реалізації вагового методу необхідно визначити з точністю до 0,1 г середню живу масу репрезентативної вибірки несортованих за статтю пташенят (100 гол.) і шляхом наступного індивідуального зважування всього масиву молодняку розділити його на дві групи («важкі» і «легкі») за наступному принципом: якщо жива маса пташеняти дорівнює або вище середнього значення («важкі»), то така особина відноситься до самців, якщо нижче середнього («легкі») – до самок. Таке калібрування пташенят за масою тіла дві на дві групи й наступне їхнє роздільне вирощування в різних секціях дозволяють із перших же днів диференціювати годівлю й щільність посадки птиці, що позитивно зазначається на її подальшому розвитку.
Метод індивідуального зважування простий в експлуатації й нешкідливий для пташенят. При використанні електричних ваг типу ВЛТК-500, швидкість сортування молодняку досягає 600 гол./година.
Інші способи. Відомий спосіб сексування курчат, в основу якого положено розходження самців і самок за фототропною реакцією на фіолетове і червоне світло [31]. Також, у літературі описана можливість сексування курчат [32] за допомогою впливу на них інфрачервоного випромінюванням, до джерела якого спрямовуються курочки; півники ж при цьому залишаються на місці.
Пропонуються також методи ранньої діагностики статі курчат, засновані на статевих розходженнях в електропровідності їх тканин [33] та різні реакції самців і самок на низькочастотні звукові подразники [34].
Всі ці способи не знайшли застосування у виробничих умовах із-за низької точності (57–67%) і невисокої швидкості (120–140 гол./година) сексування курчат.
Таким чином, можна відзначити, що на сьогоднішній день відомо більше десяти різних методів визначення статі добового молодняку свійської птиці. Однак більшість із них не забезпечують високу точність і швидкість сексування пташенят. Більшість з цих методів трудомісткі й складні у використанні, роблять стресуючий вплив на пташенят, нерідко призводять до їхнього травмування. Тому в сучасному світовому птахівництві широко використовуються тільки три з вищеописаних методів визначення статі добового молодняку – японський і два генетичних (федер- і колорсексинг).
Через важливість цих методів для сучасного промислового птахівництва зупинимось на них докладніше в наступних главах.
Історія вентсексингу. Деякі найбільш досвідчені японські й китайські птахівники вже в XІХ столітті вміли визначати стать добових курчат шляхом огляду у них клоаки й виявлення в ній статевого горбка, характерного для півника. У порівняно завершеному виді клоакальний метод був розроблений у Японії в 20-х роках минулого століття видатними вченими Києші Масуі й Джюро Хашімото, коли в країні стала інтенсивно розвиватися птахівницька промисловість [35].
В 1927 р. К. Масуі вперше виклав суть свого методу на ІІІ Всесвітньому конгресі з птахівництва в Оттаві (Канада). А вже починаючи з 1933 р. метод став широко використовуватися в Канаді й США, де йому була привласнена робоча назва – вентсексинг (від англ. vent – отвір клоаки) [36].
Зараз у Японії існує добре продумана система підготовки сортувальників високої кваліфікації, здатних після навчання визначати стать добових курчат з точністю не нижче 98% при швидкості сортування 800–1000 голів за годину. Японські спеціалісти-сексатори користуються великим попитом у США і Європі, де їхня праця оцінюється не менш 5 000 євро на місяць. Останнім часом з японськими сексаторами успішно конкурують сортувальники курчат за статтю з Китаю, Лаосу й Південної Кореї, у зв’язку із чим ціни на послуги представників цієї професії знизилися.
У країнах СНД японський метод сортування добових курчат за статтю став використовуватися починаючи з 50-х років минулого сторіччя. Однак, дотепер, цей метод сортування не одержав належного розвитку, оскільки точність сексування складала в середньому 88–93% при швидкості 600–800 гол./година.
Зараз вентсексинг широко використовується в багатьох країнах світу, де добре розвинене птахівництво. Постійно відбувається вдосконалення як самої техніки маніпулювання з добовим молодняком, так і спеціального устаткування, використовуваного в даній технологічній операції.
В останні десятиліття японський метод визначення статі курчат успішно адаптований для сексування молодняку інших видів сільськогосподарської птиці – індичат, гусенят, каченят й цесарят. У зв’язку з розширенням видового складу домашньої птиці перспективна розробка клоакального методу для визначення статі у добових перепелят, страусят, фазанят і молодняку деяких інших видів сухопутної і водоплавної птиці, що перебувають на різних етапах одомашнювання.
Принцип методу. В основі японського методу визначення статі добового молодняку птиці лежить огляд розкритої клоаки пташеняти й виявлення в ній морфологічних розходжень у будові зародкового статевого горбка у самців та самок [14,15].
Первісна детермінація статі у ембріонів птиці обумовлена статевими хромосомами Z і W. У процесі ембріогенезу у самців (ZZ) формуються парні сім’яники й зародковий статевий горбок у клоаці, який згодом перетвориться в репродуктивний орган (пеніс). У ембріонів жіночої статі (ZW) формується, як правило, тільки ліва гонада (яєчник). До моменту вилуплення самок з яйця статевий горбок у них під дією жіночих статевих гормонів – естрогенів, регресує або здобуває іншу, чим у самців, форму.
Оскільки статевий диморфізм у будові нижнього відділу клоаки добре виражений у добового молодняку багатьох видів домашньої і дикої птиці, то японський метод можна вважати універсальним, придатним для сексування пташенят різних систематичних груп. Однак, він найкраще розроблений і найчастіше використовується для визначення статі у курчат, тому спочатку докладніше зупинимося на застосуванні вентсексингу в курівництві.
Вік курчат, оптимальний для вентсексингу. Точність визначення статі курчат японським методом у значній мірі залежить від їхнього віку. Вентсексинг молодняку яєчних порід курей рекомендується проводити відразу ж після виїмки курчат з інкубатора у віці 6–16 годин після вилуплення. До цього часу пташенята вже добре обсохли й у них зарубцювалася пуповина. У молодняку старше 16-годинного віку відмінна ознака статі поступово згладжується, що помітно знижує точність його сексування [37].
У курчат м’ясного й м’ясо-яєчного напрямків продуктивності оптимальним для сексування вважається вік 8–18 годин після виводу. Курчата-бройлери молодше 8-годинного віку звичайно ще вологі й пухкі, тому при визначенні у них статі можливі травми клоаки й стінок жовточного міхура [38].
Організація робочого місця. Продуктивність праці оператора багато в чому залежить від правильної організації його робочого місця. Для сортування добових курчат за статтю необхідно окреме приміщення з оптимальною температурою повітря (23–26 о
С), гарною вентиляцією й щільними шторами на вікнах.
Існує два варіанти роботи сортувальників молодняку за статтю японським методом – сидячи й стоячий. У країнах СНД більшість операторів сексують курчат сидячи за спеціально призначеним для цієї мети столом ССЦП-2. Крім спеціального стола на робочому місці сортувальника знаходиться настільна лампа, зручний стілець і ящики для розсортованих за статтю курчат.
Для освітлення розкритої клоаки курчат використовується електрична лампа розжарювання потужністю 75–100 Вт, яка жорстко закріплена на поверхні стола. Лампа має металевий абажур і рухливу станину, що дозволяє встановити джерело світла нижче рівня очей оператора (світло не повинно попадати йому в очі).
У поверхню стола вмонтований знімний ящик з нержавіючої сталі для утримання курчат. Ящик має сітчасте дно, під яким знаходиться кювета для посліду. Деякі оператори збирають посліду у скляну банку, яку після завершення роботи миють під проточною водою.
З обох сторін від оператора розташовані порожні пластмасові ящики для добових курочок (ліворуч) і півників (праворуч). У міру їхнього завантаження курочки й півники передаються іншим працівникам інкубаторію для наступної обробки, що може бути специфічна для одноденних самців і самок (вакцинація, маркірування, обрізання дзьоба або гребеня, припікання шпор і пазурів). В окремий ящик оператор-сортувальник кладе нетипових і непридатних для сексування особин (слабкі, каліки).
Перед початком роботи оператор заповнює центральний ящик несортованим молодняком, включає електричну лампу й, надягши марлеву пов’язку на обличчя, приступає до сортування курчат за статтю. По закінченню роботи він повинен виключити світло, витерти пил з поверхні стола, очистити й вимити кювету для збору посліду.
У країнах Західної Європи і Японії визначники статі курчат японським методом часто працюють стоячі. Прикладом тому служить нова система організації праці при визначенні статі добових бройлерів, розроблена й впроваджена в практику британським інженером Дж. Хейкоком. Модульна установка дозволяє одночасно працювати 14-ти операторам, хоча при бажанні її розміри можна легко збільшити.
Кожне робоче місце сексатора має три похилих воронкоподібних рукава-приймача, які виготовлені з нержавіючої сталі. Несортовані за статтю добові бройлери надходять до оператора через нахилений до нього центральний рукав. Два інших приймачі розташовані по обидві сторони від кожного сортувальника й через них курочки й півники плавно зсковзують у порожні ящики. Ця система збільшує на 35% продуктивність праці операторів і створює кращі санітарно-гігієнічні умови для курчат у процесі вентсексингу.
Незважаючи на те, що саму процедуру вентсексингу (на відміну від колор- і федерсексингу) поки не можна механізувати, у багатьох країнах світу постійно відбувається удосконалення столів, за якими працюють сортувальники молодняку за статтю. Так, наприклад, фахівці американської компанії «Ембрекс» розробили й впровадили в практику птахівничої фірми» Хай-лайн» виробничу лінію, важливим елементом якої є «карусель» сексування. Ця виробнича система дозволяє автоматизувати наступні технологічні операції, здійснювані в інкубаційному цеху: вибірку курчат з інкубатора, відділення їх від шкаралупи, конвеєрну подачу курчат до сексаторів, диференційований підрахунок самців, самок і некондиційних курчат, їхню вакцинацію проти хвороби Марека, розміщення курочок і півників у різні ящики.
Аналогічні виробничі лінії створені й в інших країнах світу з високорозвиненим птахівництвом. При їхньому використанні в значній мірі скорочується персонал інкубаторію, знижується рівень стресування добових курчат, прискорюється їхня ветеринарна обробка й відправлення на вирощування, підвищується збереженість молодняку в перший тиждень життя.
Незалежно від умов роботи, тривалість робочого дня сексатора молодняку японським методом становить 6 годин при обов’язковому десятихвилинній перерві щогодини. Негативно позначаються на результативності роботи операторів недостатнє освітлення, незручні столи й стільці, сторонній шум, щоденні перевантаження, тривалі перерви в роботі й відсутність оперативного індивідуального контролю за якістю визначення статі молодняку.
Техніка сексування курчат. Визначення статі курчат містить в собі ряд послідовних операцій, які безупинно переходять одна в іншу. Один повний цикл рухів оператора містить в собі такі дії:
– узяття курчати з ящика;
– оцінка його кондиційності;
– звільнення прямої кишки курчати від посліду;
– фіксація курчати в лівій руці;
– розкриття його клоаки;
– огляд клоаки й визначення статі курчати;
– розміщення півника і курочки у різні ящики.
Всі ці рухи рук досвідчений оператор виконує із вправністю, швидко й акуратно, витрачаючи на процес визначення статі одного курчати не більше 3-х секунд. Головна вимога до оператора при визначенні статі – не травмувати курчат! При цьому, з молодняком варто поводитись обережно, щоб уникнути вивихів ножних суглобів, травмування слизуватої оболонки клоаки й розривів неміцної оболонки жовточного міхура внаслідок надмірного здавлювання черевця курчат.
Зупинимося докладніше на техніці визначення статі курчат японським методом. Ключові моменти цієї процедури відображені на рис. 8–11. Сортувальник витягує курча з ящика лівою рукою і захоплює його пальцями так, щоб спинка пташеняти не торкалася долоні, а його голівка була спрямована до мізинця. Одночасно із цим оператор оцінює кондиційність курчат й, якщо потрібно – робить бракування й розміщення слабких і нетипових особин в окремий ящик.
Для видалення вмісту прямої кишки необхідно піднести курча до кюветі для збору посліду й трьома пальцями (великим, вказівним й середнім) тієї ж лівої руки обережно здавити пташеня з боків в області черевця (рис. 8). Струмок рідкого калу при цьому направляється в кювету. Якщо клоака залишається при цьому вологою та забрудненою, її варто осушити ватяним або марлевим тампоном.
Далі, фіксують курча в лівій руці при участю всіх п’яти пальців. Із цієї метою курча перевертають долілиць головою, а потім, захопивши пальцями правої руки його ніжки, затискають їх між вказівним і середнім пальцями лівої руки. При цьому шия пташеняти також злегка затиснута між мізинцем і підмізинним пальцями тієї ж лівої руки, а великий палець може підтримувати спинку курчати. Під час фіксації не слід сильно здавлювати пташеня, оскільки больовий стрес зробить його неспокійним, а це, у свою чергу утруднить огляд клоаки.
Уміле розкриття клоаки курчати – це завжди вирішальний момент при визначенні його статі. Відпрацьовування цієї операції потребує від оператора зосередженості й обережності. Як правило, досвідчені сортувальники починають розкривати клоаку пташеняти великим і вказівним пальцями правої руки, які розташовуються по краях (із протилежних сторін) клоаки. Для того, щоб нижня частина клоаки, на якій розташований статевий горбок самців, випнулася назовні, треба легким рухом позначених вище пальців зробити розтягання клоаки з одночасним натисненням даної ділянки убік живота курчат. Зафіксувавши клоаку курчат у випнутому положенні, треба краєм нігтьової пластинки великого пальця вже лівої руки обережно вивернути назовні внутрішню стінку клоаки з боку черевця курчати. На слизуватій поверхні цього відділу клоаки (вентральний відділ) у півників буде розташований статевий горбок, якого, як правило, немає у курочок.
Після огляду клоаки й розпізнавання образу самця або самки курчати надають статеву приналежність (півник або курочка) і поміщають його у відповідну тару. Як правило, на огляд клоаки курчат професіонали й початківці сортувальники витрачають приблизно однаковий час, а от підготовчі й завершальні рухи у майстрів проходять швидше.
Описані вище прийоми вентсексингу не є строгим стандартом і можуть бути змінені в деталях з урахуванням психофізичних особливостей і майстерності людини, що визначає стать курчат. Так, наприклад, деякі оператори-правши фіксують курчат в правій, а не в лівій руці. Досвідчені сортувальники, які розпізнають статевий горбок у півників з першого погляду, можуть не фіксувати ніжки пташеня, беручи для дослідження відразу двох курчат. Деякі оператори розкривають клоаку курчат тільки двома великими пальцями або одного великим і двома вказівними пальцями одночасно.
Останнім часом у Японії застосовується оновлений варіант клоакального методу, при якому оператор тримає курча в лівій руці спинкою догори. Зафіксувавши в такому положенні курча, вказівним пальцем правої руки він проводить вертикально зверху долілиць по клоаковому отвору, в результаті чого клоака розкривається й у її нижній частині стає видимим статевий горбок. Цей прийом дозволяє сортувальникові з багаторічним досвідом домагатися найкращих результатів точності сортування курчат за статтю (98–100%) при найвищій продуктивності праці (більше 1000 голів у годину).
Таким чином, існує кілька варіантів фіксації курчат й розкриття його клоаки. Кожний птахівник, що навчається вентсексингу, повинен сам вибрати для себе найбільш прийнятну техніку визначення статі. Не так важливо, який варіант сексування молодняку буде обраний. Головне, щоб було виконане основне завдання – правильно визначена його стать. При цьому важливо, щоб молодняк був мінімально стресований і не травмований. Потрібно постійно пам’ятати, що після вилуплення тканини курчат дуже м’які, а пупочний отвір ще повністю не зарубцювався. Тому при сексуванні курчат поводитись з ними треба дуже обережно.
Оскільки японський метод може сприяти перезараженню молодняку збудниками кишкових інфекційних захворювань (пуллороз, колібактеріоз, нейссеріоз, сальмонельоз та ін.), то операторові доцільно періодично, через кожні 10–20 маніпуляцій (визначень статі) дезінфікувати пальці рук у дезрозчині. Із цією метою пальці можна опускати у ванночку з 75%-ним розчином етилового спирту або ж обтирати їх марлевою серветкою, змоченої в цьому ж розчині.
Різновиди статевих горбків у курчат. Форма статевих горбків у добових півників і курочок сильно варіює, тому операторам необхідно придбати великий досвід, щоб безпомилково визначати стать курчат. Приблизно у 70% добових самців зустрічається типовий статевий горбок, що схожий на півсферу діаметром близько 1 мм. Іноді зустрічаються півники з дуже маленьким статевим горбком, або з горбком, роздвоєним по середині або маючим серцеподібну форму. У більшості півників статевий горбок пружний й гладкий, блискучий, червонуватого кольору. Від нього відходять уліво й вправо по одній рожевій шкірній складці, які відділені від горбка помітним поглибленням у його основі. Оскільки горбок є щільним утворенням, то при натисненні на нього пальцем він не зникає.
Типовий вид зовнішніх статевих ознак добової курочки – наявність на стінці клоаки рожевих складок і відсутність між ними горбка (55–60% курочок). Однак, не рідко зустрічаються курочки з невеликим гострим або плоским горбком рожевого кольору. При розтягуванні клоаки або натисканні на горбок жіночого типу він зникає, що є додатковою діагностичною ознакою жіночої статі.
Варіації статевого горбка у півників:
а
– збільшений куполоподібний статевий горбок (4–6%);
b
– типовий куполоподібний горбок (65–75%);
c
– маленький куполоподібний статевий горбок (8–15%), що може бути не помічений при швидкому огляді й прийнятий за жіночій тип;
d
і d1
– серцеподібний горбок одного типу, що міняє свою форму при різному натисканні пальців в області клоаки (0,5–1%). При більше сильному натисканні пальців (d) цей горбок важко диференціювати відносно статі. Ослаблення тиску на клоаку (d1
) дозволяє слизуватій оболонці повернутися в природне положення, що робить характерну для чоловічої статі півсферу статевого горбка добре помітною у нижній частині цього тривершинного утворення;
e
– глибоко розташований горбок, що при слабкому розкритті клоаки може бути не помічений (2–10%);
f
– перехідна форма горбка між варіантами с и в (1–6%);
g
– вертикально розщеплений половою горбок (0,5–4,0%);
h
– великий цебулеподібний горбок (0,5–3%).
Мінливість будови клоаки у курочок:
а
– найпоширеніша форма жіночого типу – повна відсутність якої-небудь структури, хоча б що віддалено нагадує горбок (55–60%);
b
– на місці горбка є V-образна складка (5–10%);
с
и с1
– горбок жіночого типу, що під різним тиском пальців опера – тора змінює свою форму (2–5%). При більше слабкому тиску на клоаку (з1) цей горбок легко можна поплутати із чоловічим;
d
– горбок подібний з типом с
, по винятком того, що він має плоский язикоподібний зовнішній вигляд (10–17%);
e
– піднесення у вигляді горизонтального гребеня (2–10%). Варіації цього типу важко диференціюються відносно статі;
f
– плоский горбок розщеплений вертикально по серединній лінії (1–8%). Цей тип горбка може бути легко переплутаний із чоловічим типом g
. Однак, більш плоска поверхня піднесення (на відміну від округлої чоловічої) дозволяє класифікувати його носія – як «жіночу особину».
g
– маленький гострий жіночий горбок, розташований трохи вліво від середньої лінії. При розкритті клоаки курчат горбок значно витикається й звішується долілиць, що чітко відрізняє його від чоловічого типу (2–8%);
h
– горбок жіночого типу, що відрізняється від чоловічого горбка занадто малими розмірами (1–5%).
На частоту зустрічальності типів статевих горбків впливає породна приналежність курчат й ступінь тиску пальців оператора на стінки клоаки. Досвідчені визначники статі вважають, що молодняк яєчного напрямку продуктивності легше сексувати, ніж курчат м’ясних порід і кросів. Це пов’язане з тим, що останніх слабкіше виражений статевий диморфізм у будові клоаки (статевий горбок у півників має менші розміри й часто розташований глибоко в клоаці). Крім того, курчата м’ясного напрямку продуктивності більші й пухкіші, що змушує операторів поводитися з ними дуже обережно.
Особливості сексування індичат. У індичат, на відміну від курчат, зі збільшенням часу після вилуплення (у годинах) розпізнавання статі за будовою клоаки замітно полегшується. Нижче наводимо опис статевого диморфізму в будові клоаки у добових індичат, узятий нами з навчального посібника Н.П. Третьякова й Г.С. Крок [39]. Зовнішній статевий орган добового індичати самця являє собою два однакові кулясті горбка, розташовані на краю клоаки з боку живота. Кулясті статеві горбки самця пружні, блискучі й у більшості особин червоного кольору.
У самочок зовнішні статеві органи виражені двома шкірними складками, з’єднаними між собою у вентральній частині клоаки. Форма складок трохи варіює. У більшості самок з нижньої сторони клоаки вони стовщені й витягнуті. Зустрічаються особини, у яких стовщенні складки більш округлі, що часто є причиною помилкової діагностики статі (самок приймають за самців). На відміну від статевих горбків самця, шкірні складки самки мають блідо-рожевий колір. При легкому масажуванні вони не блищать і стають плоскими, що також допомагає відрізнити самок від самців.
Техніка вентсексингу у добових індичат майже така ж, як у курчат. Відмінними рисами є те, що немає необхідності звільняти кишечник індичати від посліду, а також сильно розкривати у нього клоаку. Часто буває досить злегка відтягнути пальцями правої руки хвостик пташеняти до спини, щоб виявити горбки або складки, що диференціюють стать. Якщо ж статеві горбки у самців і шкірні складки у самок розташовані на 2–3 мм глибше краю клоаки, то тоді їх варто розкрити сильніше. Для цього великий і вказівний пальці правої руки розміщають праворуч і ліворуч від клоаки й, злегка розтягуючи її, нахиляють хвостик індичати у бік спинки.
Визначення статі гусенят, каченят, фазанят і перепелят. У добового молодняку водоплавної птиці (гусаки, качки, лебеді) статевий диморфізм у морфології клоаки виражений більш чітко, ніж у сухопутної (кури, індички, цесарки, фазани), що, у свою чергу, забезпечує більш високу точність їх сексування японським методом. У добових гусенят і каченят статевий горбок має форму злегка загнутого буравчика, розміром 1,5 – 2,0 мм [40]. Він має серединне положення в нижній частині клоаки й, у звичайному положенні пташеняти, схований у складках слизуватої оболонки. У самочок горбок завжди відсутній, а замість нього видні дві округлі шкірні складки, що зрослись між собою.
У добових пташенят водоплавної птиці, так само як і у індичат, немає необхідності звільняти кишечник від його вмісту. У них також відсутні жорсткі вимоги до віку молодняку, у якого визначається стать. Однак гусенята й каченята молодче 8-ми годинного віку ще дуже вологі, пухкі й слабкі. І тому раннє визначення їхньої статі може викликати у такого молодняку травми клоаки й стінок жовточного міхура.
Техніка сексування добових цесарят, фазанят і перепелят така ж, як і у курчат, однак у зв’язку з дуже маленькими розмірами статевих горбків у самців застосування японського методу в цьому випадку значно спрощується при використанні операторами бінокулярної лупи або збільшувального скла зі збільшенням в 3–5 разів.
Контроль точності вентсексингу. Оперативний контроль за точністю визначення статі молодняку здійснюють за допомогою анатомічного методу. Цей метод абсолютно точний, однак досить трудомісткий і сполучений зі значними матеріальними витратами, викликаними забоєм пташенят. В основу цього методу покладені анатомічні розходження самців і самок у будові гонад (статевих залоз), локалізованих у черевній порожнині [39]. Перед визначенням статі анатомічним методом курчат необхідно приспати концентрованими парами діетилового ефіру. Із цією метою курчат розміщають у герметично закриту тару, куди також кладуть тампон, змочений ефіром. Потім кожне приспане курча беруть у ліву руку й за допомогою кривих ножиців роблять розтин черевної порожнини. Далі, кінцями ножиців або пінцетом відсувають убік жовточний міхур і печінку, які прикривають статеві залози, прикріплені до хребетного стовпа курчат в області хрестця.
Ідентифікацію статі добового молодняку анатомічним методом здійснюють шляхом огляду їх гонад (рис. 13). У добових самців всіх видів птиці в області хрестця (у глибині черевної порожнини) розташовані парні сім’яники білуватого кольору, які нагадують рисові зерна. Лівий сім’яник у деяких видів птиці трохи більше правого. Потрібно пам’ятати, що сім’яники не міцно закріплені в черевній порожнині (підвішені на коротких брижах) і при недбалому розкритті вони можуть бути відірвані. У добових півників довжина гонад приблизно однакова й становить 4,0-4,5 мм при їхній товщині близько 1,5 мм.
Яєчник самки – непарний орган. Він крупніше сім’яників, рожевого кольору, плоский, має форму трикутника з нечіткими границями. Яєчник розташований, як правило, з лівої сторони, а права його частина майже повністю скорочена.
За нашими спостереженнями [30] швидкість визначення статі молодняку при використанні анатомічного методу коливається для різних видів свійської птиці від 120 (гусенята) до 200 (курчата) голів за 1 годину.
Контроль за точністю сексування курчат японським методом проводиться звичайно в господарствах, де утримуються кури яєчного напрямку продуктивності. При визначенні точності сексування у кожного оператора беруть для розтину по 50 голів добових півників і 20 голів курочок з різних ящиків. Після ідентифікації статі у вибірці забитих курчат підраховують кількість помилок і точність сексування молодняку окремо в групі самців (50 гол.) і самок (20 гол.), а також у цілому по аналізованій групі (70 гол.). Для цього використовують наведену нижче формулу:
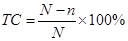 ,
,
де ТС – точність визначення статі японським методом; N – кількість сексованих пташенят; n – кількість помилкових визначень статі.
У племінних господарствах, що спеціалізуються на розведенні птиці м’ясного напрямку продуктивності (кури, індички, гуси, качки) контроль за точністю визначення статі добового молодняку можна проводити також ретроспективним методом. Для цього розсортованих за статтю японським методом самців і самок розміщують в окремі секції по 200–500 голів і вирощують роздільно до двомісячного віку. У цьому віці статеві ознаки у самців і самок уже помітно проявляються, що й дозволяє встановити справжню стать особин.
На відміну від інших видів свійської птиці визначити стать молодих і дорослих гусей за вторинними статевими ознаками важко, тому що за зовнішнім виглядом гуски й гусаки схожі. Тому, стать гусей у будь-якому віці визначають за наявністю в клоаці репродуктивного органа. Із цією метою кожну особину беруть за ноги й крила, перевертають її і кладуть спиною на стіл або коліна оператора так, щоб хвостова частина тулуба звисала. Зафіксувавши в такому положенні птицю, пальцями обох рук розкривають її клоаку і за наявністю або відсутністю спіралеподібного статевого органа визначають стать особини. У молодих гусаків довжина репродуктивного органа становить 4–5 мм, а у статевозрілих самців – 3–5 см. Більш докладно процедура вентсексингу молодих і дорослих гусаків описано нами в другому розділі.
За даними ряду авторів [34, 40, 41], використання японського методу дозволяє визначити стать добових курчат, цесарят і перепелят з точністю 86–99%, індичат – 90–95%, каченят і гусенят – 96–100%. Помилки у визначенні статі при використанні вентсексингу виникають за наступними причинами: велика кількість некондиційних курчат у партії, глибоке розташування в клоаці статевого горбка самця або занадто малі його розміри, перетримування курчат в інкубаторі, некваліфікована робота операторів (у т.ч. неповне розкриття клоаки пташеняти).
Продуктивність і оплата праці. Поряд з точністю сексування швидкість визначення статі молодняку й вихід кондиційних курчат, індичат, каченят і гусенят після сортування також є найважливішими показниками рівня професійної майстерності оператора. Звичайно, продуктивність праці сортувальника виміряється шляхом прямого хронометрування процесу визначення статі у 100 пташенят за допомогою секундоміра на початку й кінці роботи.
При гарній організації робочого місця досвідчений сортувальник визначає стать курчат яєчних порід зі швидкістю 700–900 голів/год, м’ясних порід – 500–600 голів/год, індичат, гусенят і каченят – до 500 голів/год. Якість сортування молодняку за статтю залежить від рівня професійної підготовки операторів, впровадження прогресивних методів організації праці, біологічної повноцінності добового молодняку й матеріальної зацікавленості сортувальників.
Відповідно до тарифікації робіт, виконуваних у тваринництві, робота оператора за визначенням статі добового молодняку сільськогосподарської птиці ставиться до шостого розряду. Норми виробітку й розцінки за 1000 розсортованих за статтю курчат, індичат, гусенят і каченят встановлюються у кожному птахівничому господарстві з урахуванням конкретних умов. Оплата праці операторів, як правило, залежить напряму від якості їхньої роботи, у першу чергу – від точності визначення статі молодняку. Наприклад, у дослідному господарстві Інституту птахівництва УААН «Борки» у 2007 році були прийняті наступні диференційовані розцінки, відносно 1000 розсортованих за статтю курчат:
– при точності 90, 0–95,0% – 40,00 грн;
– при точності 85, 0–89,9% – 35,00 грн;
– при точності 80, 0–84,9% – 25,00 грн.
При сортуванні м’ясних курчат, індичат, каченят і гусенят розцінки за 1000 голів розсортованого за статтю молодняку збільшуються на 40–50%. Крім основного заробітку, операторам виплачується щомісячна премія (до 20% від середнього заробітку) за високу точність визначення статі молодняку (96,0–100,0%).
Недоліки й слабкі сторони вентсексингу. Повноцінність добового молодняку, реалізованого на вирощування, залежить, насамперед, від генотипу й здоров’я батьків, якості інкубаційних яєць і режиму інкубації [34]. У той же час, недбале відношення до молодняку в інкубаторії може помітно погіршити його кондиційність і біологічну повноцінність. Зокрема, травми тонкої стінки жовточного міхура, виникаючі іноді в момент сексування пташенят японським методом, призводять до жовточного перитоніту й загибелі молодняку в початковий період вирощування. Наші спостереження за роботою досвідчених операторів показали, що в процесі сексування спочатку здорового молодняку відхід у вигляді слабких і калік склав у яєчних і м’ясних порід, відповідально, 0,28 і 1,57% [42].
Процедура сортування пташенят за статтю японським методом значно збільшує час між вибіркою молодняку з інкубатора й посадкою його в пташник, що додатково стресує птицю й знижує її збереженість при наступному вирощуванні на 0,4–2,0% [43, 44]. У деяких випадках (слабкий молодняк, некваліфіковані оператори) застосування вентсексингу збільшує відхід курчат у процесі вирощування на 8–15% [45]. Крім того, цей метод може сприяти горизонтальному перезараженню птиці збудниками кишкових інфекційних захворювань, що також збільшує смертність молодняку в перші тижні життя.
Таким чином, японський метод, незважаючи на широке застосування у світовому птахівництві, має ряд істотних недоліків. Вентсексинг трудомісткий і складний у використанні для сортувальників; не завжди забезпечує високу точність і швидкість сексування молодняку; вимагає тривалого періоду навчання операторів і спеціально обладнаних робочих місць. Цей метод також стресує добовий молодняк, нерідко травмує й перезаражає його збудниками інфекційних захворювань. Все це призводить до вповільнення темпу росту й підвищенню падежу птиці в період вирощування.
Відзначені недоліки японського методу сексування молодняку викликали необхідність розробки більше прогресивних способів діагностики статі добових пташенят – федер- і колорсексингу.
Історія федерсексингу. Про наявність у генотипі ряду порід курей зчепленого зі статтю домінантного гена, що одержав спочатку назву «азіатського гальма» оперення, уперше повідомив А.С. Серебровський [46] у 1922 році. Цей ген викликає затримку у розвитку оперення пташенят таким чином, що у 10-денних курчат ще не відзначається розвиток хвостового оперення, а махові пір’я у них сильно вкорочені. Пташенята, які рано оперюються того ж віку вже мають хвостові пір’я, а махові у них досягають основи хвоста.
В 1930 р. Д. Уоррен [47] показав, що вже в добовому віці існують чіткі фенотипові розходження між курчатами, які рано і пізно оперюються й запропонував використовувати зчеплене зі статтю успадкування цієї ознаки для сексування добового молодняку.
Пізніше П. Хертвігг і Т.Ріттерсхауз [48] запропонували позначити домінантний фактор, що сповільнює опереність молодняку, символом K
(«коротке крило»), а його рецесивний алель – k
(«довге крило»).
Р. Соумз у 1969 р. виявив нову мутацію в K
-локусі, і назвав її надповільною опереністю й позначив символом Kn
[49]. Фенотипічний ефект цього алеля виражений значно сильніше, ніж гена K
.
У 1977 році Мак Гібон [50] повідомив про існування четвертого алеля Ks
, і назвав контрольовану їм ознаку «повільної опереністю» і запропонував наступний порядок домінування у цьому локусі: Kn
надповільна > Ks
(повільна) > K
(пізня) > k
(рання опереність).
В. Асмундсон і К. Аббот [51] першими повідомили про зчеплений зі статтю домінантний гені пізньої опереності індичок, що за аналогією з курми можна використовувати для ідентифікації статі добових індичат. За своїм фенотипічним проявом алель K
індичок нагадує алель Kn
домашніх курей. І той і інший сильно сповільнюють швидкість розвитку оперення у молодняку й погіршують зовнішній вигляд дорослих особин. При цьому обоє ці алеля не використовуються як маркери статі добових індичат і курчат.
У цей час зчеплені зі статтю алелі K
-k
виявлені й у цесарки [177].
П. Хорн і Ф. Біскуп [52] описали зчеплений зі статтю ген пізньої опереності у домашніх голубів, що дозволяє проводити маркуючі стать схрещування, особливо кращі для м’ясних білоперих порід. На відміну від курей домінантний алель локусу K
у голубів контролює швидкий темп росту ювенального оперення, а рецесивний алель – уповільнений. Схрещування самців, які рано оперюються з самками, які пізно оперюються дає типове успадкування «хрест-навхрест» і дозволяє з великою надійністю сортувати гібридних голуб’ят за статтю.
Одна з головних вимог до фенотипічних маркерів статі добового молодняку – відсутність їхнього негативного впливу на господарсько-корисні ознаки лінійної й гібридної птиці. Переважна більшість дослідників, що вивчали вплив алелей К
и k
на продуктивність і життєздатність курей, не виявили розходжень у швидкості росту, конверсії корму, яйценосності, масі яєць і збереженості у особин які рано та пізно оперюються [53–69]. Тому, на цей час система алелей «пізня-рання» опереність молодняку широко використовується для діагностики статі курчат у м’ясному і яєчному курівництві (кроси «Гібро», «Кобб-500», «Ломанн браун», «Бованс», «Домінант», «Заславский», «Зміна», «Родоніт», «Ломанн м’ясний», «Росс білий», «Хаббард», «Конкурент», «КК Кубань 123» і ін.).
Генетичні основи федерсексингу. В основі генетичних методів визначення статі молодняку лежать закономірності спадкування зчеплених зі статтю ознак, добре помітних у добових пташенят. У якості фенотипічних маркерів статевої приналежності особини найчастіше використовуються такі ознаки екстер’єру, як швидкість росту зародкового пір’я на крилі, забарвлення й малюнок пуху молодняку й, у меншому ступені, колір плюсни.
Для того, щоб краще зрозуміти спадкоємну природу аутосексингу взагалі й федерсексингу зокрема, розглянемо генетичні механізми зчепленого зі статтю спадкування у птиці.
Стать добового молодняку, як і інші його ознаки, спадково визначена. Хромосомна теорія спадковості показала, що провідна роль у детермінації статі й у підтримці рівного співвідношення статей при виводі молодняку належить хромосомному апарату. У цей час достовірно встановлене, що диплоїдна кількість хромосом (2n) у курки дорівнює 78, в індички – 82, у цесарки – 74, у качки й голубки – 80.
Хромосоми однієї пари, за якими особини різної статі відрізняються один від одного, носять назви статевих, а інші хромосоми, однакові в обох статях, називаються аутосомами. У гомогаметної статі дві статеві хромосоми ідентичні, а у гетерогаметної – ні.
У птиці чоловіча стать гомогаметна (AA+ZZ), а жіноча – гетерогаметний (AA+ZW), тому самки продукують яйцеклітини двох типів: 50% із хромосомою Z і 50% із хромосомою W. Після запліднення яйцеклітин сперміями, що несуть тільки Z-хромосоми, виникає розщеплення потомства за статтю: 50% ♂♂ (AA+ZZ) і 50% ♀♀ (AA+ZW). У всіх синів одна Z-хромосома батьківська, інша – материнська. Дочки Z-хромосому одержують тільки від батька, а W-хромосому – тільки від матері. На відміну від Z-хромосоми, W-хромосома значно менше за розмірами й не містить більшості структурних генів, які локалізовані в чоловічій хромосомі. У зв’язку із цим, характер спадкування зчеплених зі статтю генів (ознак) відрізняється від менделівського спадкування аутосомних генів і обумовлений поводженням статевих хромосом у мейозі, а також у процесі запліднення.
Відповідно до теорії успадкування ознак, зчеплених зі статтю, для одержання аутосексних (мічених за статтю) курчат домінантний алель завжди повинен привноситися в схрещування гемізиготними самками, а рецесивний – гомозиготними самцями [16,17,70]. При такому напрямку схрещування буде спостерігатися успадкування ознак за типом «хрест-навхрест», тобто, домінантну ознаку матері завжди успадкують тільки сини, а рецесивну ознака батька – тільки дочки.
Оскільки алельні гени К
и k
локалізовані в Z-хромосомі й контролюють різну швидкість росту махового пір’я у добових курчат, то існує пряма залежність між статтю гібридного курчати й типом опереності його крила.
Механізм перехресного успадкування зчеплених зі статтю ознак молодняку птиці послужив відправною крапкою для розробки федерсексингу – безпомилкового методу визначення статі добових курчат.
Для одержання мічених за статтю курчат, яких можна сексувати за типами опереності крила, необхідно схрещувати гомозиготних півнів, які рано оперюються (k+
/k+
) батьківської лінії з гемізиготними самками, які пізно оперюються (К
/–) материнського лінії. У добовому віці всі гібридні півники успадкують фенотип матері й будуть з пізнім типом опереності крила (К
/k+
). Навпроти, усі курочки будуть характеризуватися ранньої опереністю (k+
/–) (фенотип батька).
Фенотипові розходження між одноденними самцями й самками чітко виражені, що дозволяє сексувати курчат з точністю до 100%.
Племінна робота з федерсексним кросом. Для забезпечення високої точності аутосексування курчат в процесі багаторічної експлуатації федерсексного кросу необхідно постійно підтримувати абсолютну гомозиготність батьківських і материнських ліній за генами ранньої і пізньої опереності молодняку (k
і К
, відповідно). Із цією метою при відтворенні чистих ліній треба фенотипувати весь добовий молодняк з наступним бракуванням нетипових для кожної конкретної лінії особин. Всі передані на вирощування півники й курочки батьківських ліній повинні бути тільки з раннім типом опереності (k
/k
, k
/–), а материнських – тільки з пізнім.
«Забруднення» вихідних ліній федерсексного кросу невластивими їм маркерними генами відбувається найчастіше в племінний сезон при виконанні наступних технологічних операцій:
– формування батьківського стада (використання півнів і курей без обліку їхнього походження у зв’язку із втратою індивідуальної криломітки);
– штучне запліднення птиці (переплутана сперма півнів-плідників);
– маркування племінних яєць (помилки в зазначеної лінійної приналежності);
– вивід молодняку (змішування курчат батьківських і материнських ліній у вивідних шафах інкубатора).
Оскільки закони генетики виконуються з неухильною точністю тільки при бездоганному дотриманні всіх правил і технологічних норм племінної роботи, то необхідно виключити всілякі помилки, що порушують чистоту схрещування, і, як наслідок цього, знижують точність федерсексування добових гібридних курчат.
Організація робочого місця. Сортування федерсексних курчат за статтю можна починати відразу ж після їхньої виїмки з інкубатора, але не пізніше, ніж через 18 годин після виводу. Більш пізнє визначення статі курчат цим методом небажано, оскільки структура оперення крила починає змінюватися й фенотипові розходження півників і курочок стають менш чіткими.
Перед початком роботи оператор повинен витратити кілька хвилин на психофізіологічну адаптацію, що містить у собі наступні моменти: а) підготовку робочого місця; б) визначення категорійності й віку (у годинах) курчат; в) регулювання освітленості робочого поля; г) перегляд схеми типів опереності й оперативне відновлення в пам’яті образу опереності крил у курочок і півників; д) попередній вибір серед курчат сортувального еталона курочки й півника. Всі ці завдання взаємопов’язані між собою і їх потрібно вирішувати в комплексі й паралельно. Головна мета підготовчого періоду – оператор якнайкраще повинен підготуватися до ефективного багатогодинного сексування добового молодняку.
При федерсексингу вимоги до робочого місця оператора менш жорсткі, чим при вентсексингу. Сортувальник може працювати як у положенні стоячи, так і у положенні сидячи за спеціальним столом. Головна умова ефективної роботи – комфортна позиція оператора й достатнє освітлення об’єкта в полі зору (не менш 500 люкс). Звичайні умови сортування дозволяють федерсексувати курчат зі швидкістю близько 1,5 тис. голів/година. Робота операторів за «каруселлю» сексування збільшує продуктивність праці до 2 тис. голів/година.
Техніка сексування курчат. При федерсексуванні курчат, на відміну від вентсексування, немає необхідності у видаленні посліду з кишечника пташеняти й повної його фіксації в руці. Для визначення типу опереності молодняку необхідно при гарному освітленні оглянути розгорнуте віялом крило обсохлого гібридного курчати. Із цією метою оператор бере курча в праву руку й тримає його ногами долілиць, а головою до себе. Крило пташеняти при цьому потрібно затиснути між великим і вказівним пальцями лівої руки. У цьому положенні на крилі курчати добре видно дві групи зародкового пір’я – першорядні махові й парні до них покрівні, а також другорядні махові й відповідні їм покрівні пір’я. Махові пір’я ростуть від нижнього краю крила, а покрівні – від верхнього. Стать федерсексних курчат визначається шляхом порівняння відносної довжини махових і покрівних пір’їн. Більш зручно це робити оглядаючи першорядні махові й покрівні, оскільки вони розташовані на кінчику крила.
Фенотипові розходження курочок і півників. У курочок, які рано оперюються сім первинних махових пір’їн мають вигляд трубочок довжиною 12–13 мм і приблизно на 1/3 довші й трохи товщі парних до них покрівних пір’їн. У півників з пізнім типом крила зачатки 7-ми первинних махових пір’їн за довжиною й товщиною однакові із покрівними або трохи коротші за них. Ступінь укорочення махових пір’їн у гібридних півників сильно варіює, що варто мати на увазі при сексуванні курчат.
Для зручності ідентифікації півників фенотип «пізня опереність» нами [40] умовно розділений на три підтипи (П1, П2, П3) залежно від відносних розмірів покрівних і махових пір’їн [71]. До підтипу П1 відносяться особини, у яких першорядні махові пір’я за довжиною рівні з покрівними. У півників підтипу П2 середня довжина покривного пір’я (8,0 мм) така ж, як і в півників підтипу П1 (7,9 мм), але першорядні махові в них коротші і їхня довжина становить від 3/4 до 1/2 довжини зачатків відповідних покрівних пір’їн (рис. 15). Добові півники підтипу П3 характеризуються сильно вкороченими маховими пір’їнами (2,5–2,7 мм) і незначно вкороченими покрівними (6,2–7,0 мм). Початки першорядних махових у них приблизно в 2,5–3,0 рази коротші, покрівних пір’їн.
Поряд із алелем пізньої опереності К, у материнських лініях федерсексних кросів з невеликою частотою (1–2%) зустрічається ген повільної опереності Кs
. Експресія гена повільної опереності у добових півників полягає у повній відсутності у них першорядного махового пір’я й значному вкороченні зачатків покрівних пір’їн.
Розходження між курочками й півниками насамперед пов’язані з довжиною першорядного махового пір’я, тоді як величина покрівних пір’їн у особинах обох статей приблизно однакова (близько 8 мм). Тому, диференціальна діагностика статі добових федерсексних курчат може базуватися на двох критеріях – абсолютній і відносній довжині першорядного махового пір’я. Абсолютна довжина першорядного махового пір’я у добових курочок з однієї партії варіює, зазвичай, від 11 до 13 мм, тоді як у півників – від 0 до 8 мм. Якщо прийняти за стандарт статі особини абсолютну довжину махового пір’я, то виникає необхідність у вимірі середнього махового пера за допомогою мірної лінійки, що гальмує процес сексування молодняку й значно знижує продуктивність праці оператора. Тому, більш надійною і простою є друга ознака статі курчат – відносна довжина махових пір’їн у порівнянні зі спареними з ними покрівними.
Проведений нами [30] міжвидовий аналіз типів опереності добових індичат, перепеляти й фазанят різних порід показав, що всім їм характерний винятково ранній тип опереності добового молодняку.
Ця обставина виключає можливість одержання у цих видів федерсексного гібридного потомства. Незважаючи на генетичну однотипність у швидкості опереності пташенят із сімейства фазанових, федерсексинг для них у майбутньому принципово можливий, і, насамперед, за рахунок переносу алеля пізньої опереності К від курей за допомогою генно-інженерних маніпуляцій або ж завдяки виявленню в популяціях цих видів спонтанних мутантів з пізнім типом опереності, на базі яких і будуть створені материнські лінії федерсексних кросів м’ясних видів птиці.
Причини помилок при використанні федерсексингу. Якщо батьківська й материнська лінії федерсексного кросу курей повністю консолідовані (гомозиготні) за альтернативними алелями, відповідно, ранньої й пізньої опереності курчат, то «генетичних» помилок у визначенні статі гібридного молодняку в принципі не повинно бути. У всіх федерсексних курчат буде спостерігатися чіткий зв’язок між їхньою статтю й фенотипом крила.
Однак, одиничні помилки все-таки можуть виникати через неуважність оператора. У зв’язку з тим, що у деяких півників з пізнім типом опереності (підтип П2) першорядні махові пір’я на 1/3 коротше покрівних, то при швидкому огляді їхніх крил пізній тип опереності добових самців іноді може бути помилково прийнятий за ранній тип опереності у самок. Таке не розпізнавання образа самця або самки за типом опереності призводить до помилок у визначенні статі федерсексного молодняку – частина півників буде помилково віднесена до групи курочок.
Автоматичний федерсексинг курчат. Щоб уникнути суб’єктивних помилок при фенотипуванні федерсексного молодняку, а також інтенсифікувати процес сортування курчат за статтю, американськими вченими університету штату Арканзас була нещодавно здійснена успішна спроба перекласти процедуру розпізнавання образа крила самців і самок на «плечі» електронного робота [72]. Оптична система автоматичної установки значної збільшує зображення пір’я крила добового курчати, показуючи приховані в первинному пусі ясні для подальшої ідентифікації образи типів опереності добових курчат. Установка містить у собі транспортер для почергової подачі курчат до об’єктива цифрової відеокамери, джерело ультрафіолетового освітлення, комп’ютер для аналізу отриманого зображення й механічний пристрій-сексатор.
Цифрова відеокамера сприймає ясне зображення крила зафіксованого на мить курчати. При цьому використовується ультрафіолетове освітлення з довжиною хвилі від 250 до 450 нм. Спеціальна світлова архітектура пристрою забезпечує витяг характерних рис і особливостей типів опереності крила курчат. Комп’ютерна система аналізує отримане зображення, розпізнає стать курчат й пускає в хід виконавчий пристрій-сексатор.
Про швидкість і точність сортування курчат за статтю за допомогою цього пристрою в описі патенту ніяких даних не наводиться.
Визначення статі курчат в 10-денному віці. Гібридний молодняк федерсексних кросів можна розділити за статтю й в 10-денному віці [71]. У цей період у курочок з раннім типом крила махові пір’я досягають основи хвоста, а хвостові пір’я мають довжину 1–1,5 см. У півників з пізнім та повільним типами опереності у цьому віці махові пір’їни слабо розвинені, а хвостові (кермові) повністю відсутні.

Десятиденні півник (ліворуч) і курочка (праворуч)
До 30-денного віку півники з пізнім типом опереності «наздоганяють» курочок з раннім типом опереності за загальною опереністю тіла й статевий диморфізм у них за цією ознакою зникає. Півники з повільним Ks типом опереності (алель Ks) тіла вирівнюються з іншими фенотипами тільки до 11-ти тижнів життя. Оскільки така істотна затримка у формуванні пір’яного покриву може призвести до виникнення канібалізму серед молодняку, то материнські лінії курей федерсексних кросів у процесі їхнього виведення повинні бути ретельно «очищені» від небажаного алеля Ks (повільна опереність курчат).
молодняк свійський птиця стать
У цей час усі три мануальних методи визначення статі курчат (вентсексинг, федерсексинг, колорсексинг) широко використовуються в промисловому птахівництві. Тому досить актуальним є перевірка порівняльної ефективності застосування цих методів як для промислового птахівництва, так і для пошукових науково-дослідних робіт.
Точність і швидкість сексування молодняку є головними характеристиками будь-якого методу визначення статі. Анатомічний метод абсолютно точний, однак він пов’язаний із забоєм молодняку й тому його використовують, як правило, тільки як контроль при перевірці ефективності японського й іншого способів визначення статі курчат. Крім того, анатомічний метод досить складний і трудомісткий. За нашими спостереженнями швидкість визначення статі молодняку при його використанні варіює у різних видів домашньої птиці від 118 (гусенята) до 180 (курчата) голів./година.
Усереднені результати численних виробничих перевірок [30, 95, 96] показали, що абсолютна точність сексування гібридних курчат (100%) характерна для більшості федерсексних комбінацій курей (локус K
) і для всіх колорсексних гібридів, у яких експресія зчеплених зі статтю алельних генів сріблястості (S
) і золотистості (s
) у пусі курчат здійснюється на тлі колумбійського обмежника еумеланіну (Co
). Висока точність діагностики статі колорсексних курчат (98,5–100%) характерна також для схрещуваннь, де матері чорно-смугасті (B/ – E/E
), а батьки – чорні (b/b E/E
) або золотаво-коричневі (b/b ey
/ey
) забарвлення оперення. Використання ж у аутосексних схрещуваннях системи алелей S-s
у сполученні з геном І
(домінантне біле забарвлення) помітно знижує точність сортування молодняку за статтю (95,5–98,0%).
Продуктивність праці при сексуванні курчат за типами опереності крила складає 1,5–2,0 тис. гол./годину, а при колорсексингу – 3,5–7,0 тис. гол./годину. Отже, у першому випадку оператор потрачає на визначення статі однієї особини 2,0 секунди, а в другому – 0,5–1,0 секунду.
Японський метод забезпечує меншу ефективність сексування курчат. При його використанні точність ідентифікації статі молодняку в різних партіях курчат варіює від 86,0 до 95,0% при швидкості сортування 700–850 гол./годину. Незважаючи на те, що стать курчат визначали досвідчені сортувальниці, все-таки у всіх групах після визначення статі японським методом спостерігався відхід молодняку у вигляді слабких і травмованих особин (від 1,0 до 6,2%). Середня за різними вибірками збереженість курчат, після застосування японського (мануального) методу склала 97,4%.
Оскільки клоакальний метод (вентсексинг) найбільш складний і вимагає добре підготовленого висококваліфікованого персоналу (в основному його практикують японські й корейські сексатори), оплата праці при його застосуванні найвища. Колорсексинг простий у застосуванні й не вимагає тривалого навчання. Його може виконувати будь-який працівник інкубаторію, у зв’язку із чим оплата праці по виконання цього методу сексинга найнижча. Визначення статі за типами опереності крила вимагає незначної спеціальної підготовки, тому оплата праці федерсексингу займає проміжне положення.
В XX столітті одержала бурхливий розвиток одна із провідних галузей тваринництва – промислове птахівництво. Основна риса цієї галузі – спеціалізація птахівничих підприємств і концентрація в одному місці не тільки особин одного виду або породи птиці, але й однієї статі. У зв’язку із цим птахівники-практики поставили перед ученими конкретне завдання: розробити прості й надійні методи визначення статі добового молодняку.
Для рішення цього важливого практичного завдання в ХХ столітті вченими було розроблено більше десятка експрес-методів сексингу пташенят, однак у практичному птахівництві широке поширення одержали тільки три з них: вентсексинг, федерсексинг і колорсексинг.
Між собою ці три методи істотно розрізняються за багатьма параметрами. Вентсексинг, наприклад, більш універсальний, але менш технологічний. У процесі його реалізації можливе травмування й перезараження молодняку. Навпроти, колорсексинг нешкідливий для пташенят, високотехнологічений і не вимагає тривалого навчання операторів. При його використанні вибірка молодняку з інкубатора, сортування його за статтю й вибракування слабких особин зводяться до однієї дії оператора. При цьому на одне пташеня він витрачає не більше 0,5 секунди.
Федерсексинг на даному етапі свого розвитку використовується тільки в курівництві, і за багатьма технічними характеристиках (точність і швидкість сексування курчат), а також за простотою освоєння наближається до колорсексингу.
Безумовно, описані у цих рекомендаціях методи визначення статі добового молодняку птиці будуть удосконалюватися й далі. Насамперед це відноситься до конструкції столів й іншого допоміжного устаткування, що використовується при сексуванні пташенят. Уже зараз американські фірми «Ембрекс» і «Систенг» пропонують споживачам виробничі лінії, що дозволяють повністю автоматизувати такі технологічні операції як вибірка молодняку з інкубатора, подача його до сексаторів, підрахунок самців, самок і некондиційних особин, вакцинація молодняку, розміщення самців і самок в окрему тару.
Завдяки розвитку сучасних дослідницьких технологій різко збільшилася можливість вивчення незначних статевих розходжень ембріонів птиці починаючи з перших діб їхнього розвитку. Це у свою чергу дозволило вловити чіткий статевий диморфізм ембріонів за рядом ознак, що стимулювало виникнення нових ідей і методичних підходів до надраннього сексування сільськогосподарської птиці. У зв’язку із цим можна прогнозувати інтенсивний розвиток методів сексування ембріонів птиці (сексування іn ovo). Ці методи будуть перш за все базуватись на ідентифікації генетичних маркерів статі птиці – морфологічних (забарвлення очей 7-денних ембріонів), цитогенетичних (хромосомні ZZ- і ZW-комплекси) і молекулярних (ДНК маркери W-хромосоми). Великі надії також покладають на розробку способів сексування ембріонів птиці на основі сканування статевих залоз методом ядерного магнітного резонансу й виявлення специфічних для кожної статі хімічних речовин в алантоісній і амніотичній рідинах зародка.
Переходом на новий рівень знань з генетики сільськогосподарської птиці стане розробка методів спрямованого одержання з інкубаційних яєць особин бажаної статі. До теперішнього часу проблема штучної регуляції пола була вирішена тільки у шовковичного шовкопряда, у якого за допомогою партеногенетичного розвитку й зчеплених зі статтю летальних генів одержують за бажанням тільки самців або самок.
Оскільки у птиці, як і в шовковичного шовкопряда, жіноча стать гетерогаметний (ZW), методичні підходи, застосовувані для регуляції статі в шовковичного хробака, можуть виявитися перспективними й для сільськогосподарської птиці.
Таким чином, у минулому столітті були розроблені й широко впроваджені в племінне й промислове птахівництво три ефективних методи визначення статі добового молодняку – вентсексинг, колорсексинг і федерсексинг. На початку XXІ століття сучасна біологічна наука підійшла до рубежу, за яким стане можлива надійна автоматизація діагностики статі не тільки в добового молодняку, але й у ранніх ембріонів свійської птиці.
1. Зелятров А.В. Прогнози развития производства бройлеров к 2000 году по рубежом // Птицеводство. – 1981. – №9. – С. 39–40.
2. Старчиков Н., Догадаев А. Влияние раздельного виращивания курочек и півників на их рост, развитие и продуктивние качества // Передовой науч.-произв. опит в птицеводстве: Экспресс-информация. – 1983. – №4. – С. 14–16.
3. Marks H.L. Sexual dimorphism in early feed and water intake of broilers // Poultry Sc. – 1985. – Vol. 64. – №3. – P. 425–428.
4. Marks H.L. The role of water intake on sexual dimorphism for early growth of broilers // Poultry Sc. – 1986. – Vol. 65. – №3. – P. 433–435.
5. Marks H.L. Sexual dimorphism in broilers following periods of equal water and feed intake // Poultry Sc. – 1987. – Vol. 66. – №3. – P. 481–389.
6. Дуюнов Э.А., Гадючко О.Т., Рябоконь Ю.А., Пордаш Джамиль Мери. Половой диморфизм и его связь с хозяйственно полезними признаками индеек // Научно-технический бюллетень / УНИИП. Харьков, 1988. – №25. – С. 10–14.
7. Ковацкий М., Лисенко Ф. Технология виращивания и содержания мускусних уток. – М.: Агропромиздат, 1986. – 6 с.
8. Рябоконь М.Г. Роздільностатеве вирощування аутосексних гусенят на м’ясо // Птахівництво. – 1983. – Вип. 36. – С. 56–57.
9. Jones E. Sexed benefits could be worth 55 million a year // Poultry World. – 1990. – Vol. 177. – №5. – P. 20–21.
10. Seemann J. The influence of age, sex and cutting of broilers // Quality of Poultry Meet. – 1981. – P. 28–30.
11. Андреев Н.Ф. Половой диморфизм индеек и его селекционное значение // Птицеводство: Респ. межвед. темат. науч. сб. / Укр. НИИ птицеводства. – 1979. – №28. – С. 13–15.
12. Jones E. Sexed benefits could be worth 55 million a year // broilers // Quality of Poultry Meet. – 1981. – P. 22–27.
13. Биховец А.У. Определение пола молодняка // Орлов М.В., Биховец А.У., Злочевская К.В. Инкубация. – М.: Колос, 1970. – Гл. 5. – С. 132–137.
14. Canfield T.H. Sex determination of day old chicks. – Poultry Sci. – Vol.19. – 1940. – Р. 235–238.
15. Canfield T.H. Sex determination of day old chicks. Poultry Sci. – Vol.20. – 1941. – №4. – Р. 327–330.
16. Hann C.M. Sex-linkage in poultry breeding // Bull. / Min. Agriculture Fisheries Food. – Lond.: H.M.S.O., 1966. – №38. -23 pp.
17. Silverudd M. Genetic basis of sexing automation in the fowl // Acta agr. scand. – 1978. – Vol. 28. – №4. – P. 169–195.
18. Бессарабов Б.Ф. Практикум по инкубации яиц и эмбриологии сельскохозяйственной птици. – М.: Агропромиздат. 1985. – 175 с.
19. А.С. 1044250 СССР, МКИ З А 01 К 45/00. Способ определения пола циплят / В.Д. Бутенко (СССР). – №2764325/30–15; Появл. 04.05.79; Опубл. 30.09.83; Бюл. №36. – С. 8.
20. А.С. 1503719 СССР, МКИ 4 А 01 К 45/00. Устройство для определения пола циплят (А.Г. Соловьев (СССР). – №4239503/30–15; Появл. 04.05.89; Опубл. 30.08.89; Бюл. №32. – С. 11.
21. Бутенко В.Д. Исследование аутосексного метода сортировки суточних циплят по полу // Сб. науч. тр. – Волгоградский СХИ, 1975. – Т. 18. – С. 122–128.
22. Тихонов А.В., Мусаев А.М., Гуцев В.М. Полуавтоматический радиоэлектронний определитель пола у суточних циплят мясних направлений // Новие прибори, устройства и технологические процесси, разрабонанние ученими МГУ. – М., 1982. – С. 3–8.
23. Тихонов А.В., Акустическая сигналипоция и экология поведения птиц. – М.: Изд-во Моск. ун – та, 1986. – 240 с.
24. Krishan A. A cytological method for sexing young chicks // Experientia. – Basel, 1962. – Vol. 18. – P. 101–102.
25. Kodama H., Saitoh H., Tone M., Kuhara S., Sasaki Y., Mizumo S. Nucleotide sequences and unusual electrophoretic behavior of the W chromosome-specific repeating DNA units of the domestic fowl, Gallus gallus domesticus // Chromosoma. – Berl., 1987. – Vol. 96. – P. 18–25.
26. Tone M., Nakano N., Takao E., Narisawa S., Mizuno S. Demonstration of W chromosome-specific repetitive DNA sequences in the domestic fowl, Gallus g. domesticus // Chromosoma. – Berl., 1982. – Vol. 86. – P. 551–569.
27. Uryu N., Nagata Y., Ito K., Saiton H., Mizuno S. Determination of the sex of chickens by a biotin-labeled deoxyribonucleic acid probe // Poultry Sc. – 1989. – Vol. 68. – №6. – P. 850–853.
28. Romanoff A.L. Chemical and physiological sex differencies in newly hatched chicks // Poultry Sci. – 1948. – Vol. 27, №5. – P. 643–645.
29. Бондаренко Ю.В. Особенности статевого диморфизма живой масси у суточних птенцов домашних птиц // Науч.-техн. бюл. / УНИИП. – Харьков, 1988. – №24. – С. 3–7.
|