Введение
Взаимодействие человека с природой - одна из наиболее сложных и трудно разрешимых проблем современности. Сегодня стало очевидным, что задачи сохранения окружающей среды и экономического развития взаимосвязаны: разрушая и истощая природную среду невозможно обеспечить устойчивое экономическое развитие.
Формирование комплексной и гармоничной системы природопользования - важная проблема, стоящая перед человеком.
Целью
данной работы является изучение экологических систем, сукцессии и ее видов.
Рабочей гипотезой
выдвинуто предположение что, изучение данной темы будет эффективным при условиях:
· изучения экологической пирамиды;
· рассмотрение видов сукцессии;
· изучение стабильности и развития экосистем.
Основными задачами
работы являются:
· объяснить понятие и сущность экосистемы;
· определить виды экологической сукцессии;
· изучить пути решения проблемы взаимодействия человека с природой.
1.
Экологические системы
1.1
. Состав и структура экологической системы. Экологическая пирамида
Экологическая система
- основная функциональная единица экологии, включающая в себя живые организмы (биоценоз) и среду обитания (экотоп), причем каждая из этих частей влияет на другую и обе необходимы для поддержания жизни.
Экосистемы представляют собой основные природные единицы на поверхности Земли. Это не только комплекс живых организмов, но и все сочетания физических факторов. Всюду, где можно наблюдать отчетливое единство растений и животных, объединенных отдельным участком окружающей среды, говорят об экологической системе.
Понятие экосистемы не ограничивается какими-то признаками ранга, размера, сложности и происхождения. Поэтому оно применимо как к относительно простым искусственным (аквариум, теплица, пшеничное поле), так и к сложным естественным комплексам организмов и среды их обитания (озеро, лес, океан).
В состав экосистемы
входят неживые и живые компоненты (таблица)
| Неживые (абиотические) компоненты
|
Живые (биотические) компоненты
|
| название |
описание |
название |
описание |
| 1) неорганические вещества |
(N2
, C02
, Н2
О и др.), включающиеся в природные круговороты |
1) продуценты |
автотрофные (самостоятельно питающиеся) организмы, главным образом, зеленые растения, которые создают органические вещества из простых неорганических веществ. Автотрофы составляют основную массу всех |
продолжение таблицы
| Неживые (абиотические) компоненты
|
Живые (биотические) компоненты
|
| название |
описание |
название |
описание |
| живых существ и полностью отвечают за образование всего нового органического вещества в любой экосистеме, т.е. являются производителями продукции |
| 2) органические соединения |
углеводы, белки, аминокислоты, гумусовые вещества и др., связывающие биотическую и абиотическую части экосистем |
2) макроконсументы
(консументы 1, 2 и т.д. порядка) |
гетеротрофные (питающиеся другими) организмы, главным образом, животные, которые поедают растения и другие организмы. В отличие от автотрофов продуцентов, гетеротрофы выступают как потребители и разрушители органических веществ. |
| 3) климатический режим |
освещенность, температура, влажность и другие физические факторы. |
3) микроконсументы (редуценты) |
гетеротрофные организмы, преимущественно бактерии и грибы, которые разрушают сложные соединения мертвой протоплазмы, поглощают некоторые продукты разложения и высвобождают неорганические питательные вещества, пригодные для использования продуцентами |
Таблица
Компоненты входящие в состав экосистемы.
Структура экосистемы
. В зависимости от характера питания в экосистеме строится экологическая пирамида
(пирамида питания), состоящая из нескольких трофических уровней:
1) (низший) занимают автотрофные организмы;
2) гетеротрофные организмы 1 порядка, использующие в пищу биомассу растений;
3) гетеротрофы 2 порядка, питающиеся гетеротрофами 1 порядка, и т.д.
В наземных экосистемах масса продуцентов больше, чем масса консументов, масса консументов 1-ого порядка больше, чем консументов 2-ого порядка и т.д. Это обусловлено тем, что пища используется не только на рост организмов, но и на удовлетворение энергетических затрат: дыхание, движение, размножение, поддержание температуры. Поэтому графически модель экосистемы имеет вид пирамиды (Рис. 1).
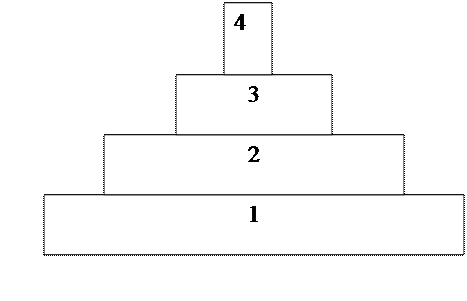
Рис. 1. Экологическая пирамида
1. Продуценты (растения); 2. Консументы 1порядка (травоядные)
3. Консументы 2 порядка (плотоядные, хищники); 4. Конечные консументы
Изучение пpиpодных экосистем в общем случае производится в стpуктуpном и функциональном аспектах. В стpуктуpном отношении исследуется видовой состав экосистемы: выясняется пеpечень видов микpооpганизмов, pастений и животных, населяющих экосистему, их количественное соотношение.
Информация, в экологических системах может пониматься как энергетически слабый сигнал, управляющий системой. Например, он может восприниматься ее организмами в форме закодированного сообщения о возможности многократно более мощных влияний со стороны других организмов, либо факторов среды, вызывающих их ответную реакцию. Так, слабые и совершенно нечувствительные для человека подземные толчки - предвестники более мощного разрушительного землетрясения, воспринимаются многими животными, своевременно покидающими свои норки.
Таким образом, информационная сеть экосистемы состоит из потоков сигналов физико-химической природы и определяет ее кибернетические возможности (кибернетика - искусство управления, гр.). Управление в экосистемах основывается на обратной связи, изображаемой обратной петлей, по которой часть сигналов с выхода системы поступает обратно на ее вход (рис.2). При этом их влияние на управление системой может резко усилится. В природе часто низкоэнергетические сигналы вызывают высокоэнергетические реакции.

Рис. 2. Механизм обратной связи
Понятие экологической системы иеpаpхично. Это означает, что всякая экологическая система опpеделенного уровня включает в себя pяд экосистем предыдущего уровня, меньших по площади и сама она, в свою очеpедь, является составной частью более кpупной экосистемы. Hапpимеp, пpавомеpно pассматpивать в качестве экосистемы аласную впадину, огpаниченную склонами межаласной возвышенности (рис.4). В свою очеpедь, эта система обычно включает в себя остаточное озеpо, болотные и луговые растительные сообщества со всеми населяющими его живыми существами. В качестве элементаpной экосистемы можно пpедставить себе кочку или мочежину на болоте, а более общей экосистемой, охватывающей множество аласов и межаласные пpостpанства, явиться соответствующая залесенная повеpхность теppасы или пенеплена.
Пpодолжая этот pяд ввеpх можно подойти к экологической системе Земли - биосфере, а двигаясь вниз - к биогеоценозу, как элементарной биохорологической (хора - пространство, гр.) единице биосферы. Учитывая pешающее значение на pазвитие живого вещества Земли зональных фактоpов, пpавомеpно пpедставить себе такой теppитоpиальный pяд соподчиненных экосистем:
· элементаpные
· локальные
· зональные
· глобальные.
И снова видно, что гpаницы экологических систем всегда откpыты. Однако, пpи этом подpазумевается некоторое теppитоpиальное огpаничение, необходимое и достаточное для получения нужных pезультатов исследования.
1.2. Биотический круговорот веществ и энергия в экологической системе
Круговорот веществ в экосистеме называется биотическим
. Перенос вещества и энергии в нем осуществляется, в основном, посредством трофических (пищевых) цепей.
Трофической (пищевой) цепью
называется перенос энергии пищи от ее источника - растений через ряд организмов путем поедания одних организмов другими. В основе этого процесса лежит следующая химическая формула:
C02
+ H2
0 + Q Cn
Hm
Ok
+ 02
. Cn
Hm
Ok
+ 02
.
Схема переноса веществ и энергии в природных экосистемах представлена на рисунке 2.

Рис.2. Схема переноса веществ и энергии в природных экосистемах.
Экологическую систему можно представить в виде диаграммы потока энергии (рис. 3).

Рис.3. Потоки энергии в простой трофической цепи.
Отдельные трофические уровни в ней изображены как резервуары, размер которых соответствует количеству энергии заключенной в них биомассы, а поперечник соединяющих их каналов - величине потоков энергии.
Энергия в экологическую систему попадает в виде потока солнечной энергии L. Большая часть ее (Lu
) рассеивается в виде теплоты. Часть энергии, эффективно поглощенная растениями (La), преобразуется фотосинтезом в энергию химических связей углеводов и других органических веществ (Pg
). Часть образовавшегося вещества окисляется в процессе дыхания растений, освобождая энергию R, а также используется в других биохимических процессах растений и в конечном счете рассеивается в виде тепла (Na
). Оставшаяся часть новообразованных органических веществ обусловливает прирост биомассы растений РП
|.
Прирост биомассы растений рано или поздно используется: часть потребляется первичными консументами, остальное перерабатывается редуцентами. Консументы питаются, размножаются, растут и также дают продукцию Р «2, которая поступает на следующий трофический уровень вторичным консументам.
Таким образом, при переходе от одного трофического уровня к другому часть доступной энергии не воспринимается (Nu
), часть отдается в виде тепла, экскрементов (Na
), а часть расходуется на дыхание (R). В среднем при переходе с одного трофического уровня на другой общая энергия уменьшается приблизительно в 10 раз (правило 10% Р. Линдемана). Чем длиннее пищевая цепь, тем меньше остается к ее концу доступной энергии. Поэтому число трофических уровней никогда не бывает слишком большим и чаще всего не превышает 4-5 уровней.
Поскольку в обратный поток поступает ничтожное количество изначально вовлеченной энергии (не более 0,25 - 0,35%). говорить о круговорот энергии нельзя. Существует лишь круговорот веществ, поддерживаемый потоком энергии.
1.3. Стабильность
и развитие экосистем
В нормальном состоянии любой экологической системе присуще устойчивое состояние, называемое гомеостазом,
характеризующееся динамическим (подвижным) равновесием между рождаемостью и смертностью, потреблением и освобождением вещества и энергии.
В тоже время любая экосистема входит в иерархию систем и поэтому подвергается внешним воздействиям, стремящимся вывести ее из равновесия. Если это атияние не слишком грубо, то нарушенные связи заменяются другими и процесс передачи вещества и энергии продолжается. Такое явление называется экологическим дублированием.
Экологическое дублирование
- процесс замены исчезнувшего по каким-либо причинам вида другим видом, который занимает его экологическую нишу. Так экосистемы сопротивляются воздействиям, нарушающим их стабильность.
Система тем надежнее и стабильнее, чем большее видовое разнообразие она имеет. Это обеспечивает широкие возможности для экологического дублирования.
В тоже время под влияние внешних и внутренних факторов в экологических системах происходят постоянные изменения. Некоторые виды экосистем, испытывая негативные воздействия, снижают свою численность, а иногда вовсе исчезают. Другие виды могут от этого выиграть, и их численность возрастает. Происходит вытеснение одних видов другими.
Процессы последовательных изменений состояния экосистем в пространстве или во времени, сопровождающиеся сменой состояний и свойств всех ее компонентов, называются сукцессиями.
Способность экосистемы относительно полно самовосстанавливаться и саморегулироваться в течение сукцессионного или эволюционного отрезка ее существования называется экологической надежностью
. Простейшим механизмом поддержания экологической надежности экосистемы является замена выбывшего по каким-либо причинам вида другим, экологически близким. При более глубоком нарушении замена происходит на уровне сообществ различного уровня вплоть до биогеоценозов.
2.
Сукцессия и ее виды
2.1. Понятие сукцессии
Восстановление экосистемой нарушенного равновесия проходит через четко определенные стадии.
Экосистему можно вывести из состояния равновесия многими способами. Обычно это бывает пожар, наводнение или засуха. После такого нарушения равновесия новая экосистема сама себя восстанавливает, и этот процесс носит регулярный характер и повторяется в самых разных ситуациях. Что же происходит в нарушенной экосистеме? На месте нарушения определенные виды и вся экосистема развиваются таким образом, что порядок появления этих видов одинаков для схожих нарушений и схожих ареалах. В этой последовательной смене одних видов другими и заключается суть экологической сукцессии.
Сукцессия
(от лат. successio — преемственность, наследование), последовательная смена одних фитоценозов (биоценозов, биогеоценозов) другими на определённом участке среды.
Сукцессии
- это постепенные необратимые направленные изменения в экосистемах, протекающие в результате внешних и внутренних причин на одной и той же территории под влиянием природных факторов или воздействий человека.
Различают множество форм сукцессии: пирогенную, катастрофическую, антропогенную и т.д. Причиной пирогенных сукцессий являются пожары; катастрофических - извержения вулканов, ураганы, необычный паводок, массовое размножение вредителей и т.п.; антропогенных - хозяйственная деятельность человека.
2.2. Причина сукцессий (частных)
Сукцессионные смены обычно связывают с тем, что существующая экосистема (сообщество) создает неблагоприятные условия для наполняющих ее организмов (почвоутомление, неполный круговорот веществ, самоотравление продуктами выделения или разложения и т.п.). Такие явления реальны, но не объясняют всех случаев смен экосистем. Например, в северных лесах внедрение под полог лиственных древесных сообществ ели связано прежде всего с тем, что последняя использует биологические свойства первых по слабому притенению почвы. Сами же почвенные условия остаются не только благоприятными для лиственных древостоев, но и постепенно улучшаются для них ( идет накопление питательных веществ, уменьшается кислотность и т.п.). Следовательно, здесь нет оснований говорить о самоотравлении или других подобных причинах смен.
Не подтверждается безоговорочно и точка зрения о том, что появление ели под пологом лиственных лесов и древостоев связано с тем, что в молодом возрасте этот вид требует затенения. Известно, например, что ель и в молодом возрасте прекрасно растет при полном освещении (значительно лучше, чем под пологом других древесных видов). Об этом, в частности, свидетельствуют многочисленные примеры создания культурных фитоценозов ели (посадкой молодых растений или посевом семян) на открытых площадях.
2.3. Человеческий фактор
Наряду с природными факторами причинами динамики экосистем все чаще выступает человек. К настоящему времени им разрушено большинство коренных (климаксных) экосистем. Например, степи почти полностью распаханы (сохранились только на заповедных участках). Преобладающие площади лесов представлены переходными (временными) экосистемами из лиственных древесных пород (береза, осина, реже ива, ольха и другие). Эти леса обычно называют производными, или вторичными. Они, как отмечалось выше, являются промежуточными стадиями сукцессий. К сменам экосистем ведут также такие виды деятельности человека, как осушение болот, чрезмерные нагрузки на леса. Например, в результате отдыха населения (рекреации), химических загрязнений среды, усиленного выпаса скота, пожаров и т.п. Антропогенные воздействия часто ведут к упрощению экосистем. Такие явления обычно называют дегрессиями. Различают, например, пастбищные, рекреационные и другие дегрессии. Смены такого типа обычно завершаются не климаксными экосистемами, для которых характерно усложнение структуры, а стадиями катоценоза, которые нередко заканчиваются полным распадом экосистем. Климаксные экосистемы обычно чувствительны к различным вмешательствам в их жизнь. К подобным воздействиям, кроме хвойных лесов, чувствительны и другие коренные сообщества, например, дубовые леса. Это одна из причин катастрофической гибели дубрав в современный период и замена их, как и хвойных лесов, менее ценными, но более устойчивыми временными экосистемами из березы, осины, кустарников или трав. Последнее особенно типично при разрушении степных и лесостепных дубрав.
2.4. Виды сукцессий
Сукцессии, с которыми мы познакомились на примере лесной зоны, называют первичными по той причине, что они начинаются с исходно безжизненного пространства (субстрата). Кроме отвалов горных пород, такие сукцессии могут начинаться на песчаных обнажениях, продуктах извержения вулканов (застывшая лава, отложения пепла) и т.п.
Наряду с первичными выделяют вторичные сукцессии. Последние отличаются от первичных тем, что они начинаются обычно не с нулевых значений,а возникают на месте нарушенных или разрушенных экосистем. Например, после вырубок лесов, лесных пожаров, при израстании площадей, находившихся под сельскохозяйственными угодьями. Основное отличие этих сукцессий заключается в том, что они протекают несравненно быстрее первичных, так как начинаются с промежуточной стадии (трав, кустарников или древесных растений-пионеров) и на фоне более богатых почв. Конечно, вторичная сукцессия возможна только в тех случаях, если человек не будет оказывать сильное и постоянное влияние на развивающиеся экосистемы. В последнем случае, как отмечалось выше, процесс пойдет по схеме дегрессий и завершится стадией катоценоза и опустынивания территорий.
Различают также автотрофные и гетеротрофные сукцессии. Рассмотренные выше примеры сукцессий относятся к автотрофным, поскольку все они протекают в экосистемах, где центральным звеном является растительный покров. С его развитием связаны смены гетеротрофных компонентов. Такие сукцессии потенциально бессмертны, поскольку все время пополняются энергией и веществом, образующимися или фиксирующимися в организмах в процессе фотосинтеза либо хемосинтеза. Завершаются они, как отмечалось, климаксной стадией развития экосистем.
К гетеротрофным относятся те сукцессии, которые протекают в субстратах, где отстутствуют живые растения (продуценты), а участвуют только животные (гетеротрофы) или мертвые растения. Этот вид сукцессий имеет место только до тех пор, пока присутствует запас готового органического вещества, в котором сменяются различные виды организмов-разрушителей. По мере разрушения органического вещества и высвобождения из него энергии сукцессионный ряд заканчивается, система распадается. Таким образом, эта сукцессия по природе своей деструктивна. Примерами гетеротрофных являются сукцессии, имеющие место, например, при разложении мертвого дерева или трупа животного. Так, при разложении мертвого дерева можно выделить несколько стадий смен гетеротрофов. Первыми на мертвом, чаще ослабленном дереве, поселяются насекомые-короеды. Далее их сменяют насекомые, питающиеся древесиной (ксилофаги). К ним относятся личинки усачей, златок и других. Одновременно идут смены грибного населения. Они имеют примерно следующую последовательность: грибы-пионеры (обычно окрашивают древесину в разные цвета), грибы-деструкторы, способствующие появлению мягкой гнили, и грибы-гумификаторы, превращающие часть гнилой древесины в гумус. На всех стадиях сукцессий присутствуют также бактерии. В конечном счете органическое вещество в основной массе разлагается до конечных продуктов: минеральных веществ и углекислого газа. Гетеротрофные сукцессии широко осуществляются при разложении детрита (в лесах он представлен лесной подстилкой). Они протекают также в экскрементах животных, в загрязненных водах, в частности, интенсивно идут при биологической очистке вод с использованием активного ила, насыщенного большим количеством организмов.
2.5. Общие закономерности сукцессионного процесса
Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса:
1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы. но по мере развития сукцессии эти показатели возрастают.
2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания.
3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. В связи с этим по мере развития сукцессий уменьшается вероятность вспышек численности отдельных видов.
4. Интенсифицируются процессы круговорота веществ, поток энергии и дыхание экосистем.
5. Скорость суцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительные сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах.
6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна.
Динамические процессы при этом не приостанавливаются, а лишь замедляются.
Продолжаются динамические процессы, обуславливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического (флуктуационного) плана.
7. В зрелой стадии климаксного сообщества биомасса обычно достигает максимальных или близких к максимальным значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса.
Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а, во- вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов.
Эти положения нельзя распространять на все климаксные сообщества. Например, нет реальных предпосылок для увеличения численности гетеротрофов в хвойных лесах по сравнению с лиственными. Скорее, в последних больше потребителей зеленой продукции и, вероятнее, вспышки численности отдельных видов- фитофагов, например, насекомых.
Нет также ни теоретических предпосылок, ни фактических данных, которые бы свидетельствовали, что в зрелой климаксной системе, например в еловых лесах, масса хвои достигает чрезмерно высоких значений. Это противоречит принципам адаптации к увеличению биогенной геохимической энергии организмами как условию их выживания (второй биогеохимический принцип В.И. Вернадского). Весь опыт лесоводства также свидетельствует о наиболее высокой продуктивности климаксных лесных сообществ (применительно к лесной зоне хвойных или смешанных хвойно- лиственных лесов). В противном случае, с точки зрения получения продукции (древесины), неизбежен вывод о нецелесообразности ориентации на выращивание и сохранение климаксных стадий лесов.
Применительно к другим экосистемам, например луговым, можно согласиться с тем, что возможности получения продукции на климаксной стадии уменьшаются, однако не потому, что сокращается ее нарастание (прирост, продуктивность), а по той причине, что более значительная часть ее отчуждается гетеротрофами в результате образования устойчивых цепей выедания.
Другими словами, продуктивность экосистем на климаксных стадиях сукцессий высока. Как правило, максимальна вследствие более полного освоения пространства. Однако возможности снятия человеком первичной продукции лимитируются (иногда до нулевых значений) вследствие включения ее в цепи питания.
Заключение
Экологическая система представляет собой любую совокупность живых оpганизмов и сpеды их обитания, взаимосвязанных обменом веществ, энеpгии, и инфоpмации, котоpую можно огpаничить в пpостpанстве и во вpемени по значимым для конкpетного исследования пpинципам.
В реферате была рассмотрена иерархия экологических систем и на примерах показано, что любая экологическя система состоит из подсистем. Их количество и качественное различие не могут быть строго фиксированы, но определяются физико-географическими и иными условиями жизнеобитания. Или, исходя из правила полноты составляющих: число функциональных составляющих экосистемы и связей между ними в условиях квазистационарного ее состояния - всегда оптимально.
Нарушение этого правила, вызванное внутренним саморазвитием системы, или внешним на нее воздействием, выводит систему из состояния равновесия и стимулирует ее переход в иное качество.
В данной работе также была рассмотрена экологическая сукцессия.
Сукцессия любого масштаба заканчивается формированием зрелого сообщества, и в экосистеме все популяции приходят в состояние динамического равновесия.
В ходе сукцессии постепенно нарастает видовое разнообразие.
С энергетических позиций сукцессия - это такое неустойчивое состояние сообщества, которое характеризуется несоответствием двух показателей: валовой продуктивности и энергетических затрат всей системы на поддержание жизнедеятельности – дыхания.
Изымая избыток чистой продукции из сообществ, находящихся в начале развития сукцессии, мы задерживаем ее, но не подрываем основу существования сообщества.
Вмешательство же в стабильные, климаксовые системы неминуемо вызывает нарушение сложившегося равновесия. Пока нарушение не превышает самовосстановительной способности системы, вторичные сукцессии могут вернуть ее к исходному состоянию - этим пользуются, например, при планировании рубок леса.
Таким образом цель данной работы была достигнута, все поставленные задачи решены.
Список использованных источников
1. Акимова, Т.А. Экология: Учебник для вузов / Т.А. Акимова, ВЛЗ. Хаскин. - М: ЮНИТИ, 1998, - 445 с.
2. Ильина Т.А. Экология и охрана природы [Электронный ресурс] – Режим доступа: http://revolutionecology/00074999_0.html.
3. Кормилицин, В.И. Основы экологии: Учеб, пособие / В.Ц. Кормилидин. - М.: Интерстиль. 1997. - 368 с.
4. Маврищев, В.В. Основы общей экологии: Учеб. пособие / В.В, Маврищев. - Мн.: Выш. шк., 2000, - 317 с.
5. Охрана окружающей среды: Учеб, для техн. спец, вузов / Под ред. С.З.
Белова. - М.: Высшая школа, 1991. - 319 с.
6. Реймерс, Н.Ф. Охрана природы и окружающей человека среды: Словарь-справочник / Н.Ф. Реймерс. - М: Просвещение, 1992. - 320 с.
7. Шимова, О.С. Основы экологии и экономика природопользования: Учебник / О.С. Шимова, Н.К. Соколовский. - Мн.: БГЭУ, 2001 -367 с.
8. Экология и безопасность жизнедеятельности: Учеб. пособие для вузов / Под ред. Л.А. Муравья. - М. ЮНИТИ-ДАНА, 2000. - 447 с.
9. Экология: Учебное пособие / Общая ред. С.А. Боголюбова. - М: Знание, 1997. - 288 с.
|