
БИОЛОГИЯ
Под редакцией академика РАМН профессора В.Н. Ярыгина
В двух книгах
•
Книга 2 •
Издание пятое исправленное и дополненное
Рекомендовано Министерством образования Российской Федерации
в качестве учебника для студентов медицинских специальностей
высших учебных заведений
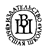
Москва «Высшая школа» 2003
УДК
574/578
ББК 28.0 Б63
Авторы:
В.Н. Ярыгин, В.И. Васильева, И.Н. Волков, В.В. Синельщикова
Рецензенты:
кафедра медицинской биологии и генетики
Тверской государственной медицинской академии
(зав. кафедрой — проф. Г. В. Хомулло
);
кафедра биологии Ижевской государственной медицинской академии (зав. кафедрой — проф. В.А. Глумова
)
ISBN
5-06-004589-7
(кн. 2)
© ФГУП «Издательство «Высшая школа», 2003
ISBN
5-06-004590-0
Оригинал-макет данного издания является собственностью издательства «Высшая школа», и его репродуцирование (воспроизведение) любым способом без согласия издательства запрещается.
Настоящая книга — продолжение учебника «Биология» для студентов медицинских специальностей. Она включает разделы, посвященные биологическим закономерностям, которые проявляются на популяционно-видовом и биогеоценотическом уровнях организации жизни на Земле.
Содержание учебника, как и в 1-й книге, включает изложение теоретических проблем, описание конкретных биологических объектов и оценку особенностей человека как биосоциального вида в процессах эволюции жизни на всех уровнях, вплоть до биосферного.
В данной книге есть некоторые подходы к изложению материала, не встречавшиеся ранее в подобных учебниках. К ним относятся более глубокое отображение и обоснование врожденных пороков развития человека, которые могут быть расценены как онто- и филогенетически обоснованные. Кроме того, дано описание экологических групп паразитов человека в соответствии с их адаптацией к специализированным условиям существования в разных органах.
Нумерация глав продолжает 1-ю книгу.
Авторы
Рассмотренные ранее биологические явления и механизмы, относящиеся к молекулярно-генетическому, клеточному и онтогенетическому уровням организации жизни, в пространственном отношении ограничивались отдельно взятым организмом (многоклеточным или, одноклеточным, прокариотическим или эукариотическим), а во временном — его онтогенезом, или жизненным циклом. Популяционно-видовой уровень организации относится к категории надорганизменных.
Жизнь представлена отдельными видами, являющимися совокупностями организмов, которые обладают свойствами наследственности и изменчивости.
Эти свойства становятся основой эволюционного процесса. Механизмами, обусловливающими такой результат, являются избирательная выживаемость и избирательное размножение особей, принадлежащих к одному виду. В природных условиях особенно интенсивно размножение происходит в популяциях, которые являются минимальными самовоспроизводящимися группами особей внутри вида.
Каждый 'из некогда существовавших или ныне живущих видов представляет собой итог определенного цикла эволюционных преобразований на популяционно-видовом уровне, закрепленный изначально в его генофонде.
Последний отличается двумя важными качествами. Во-первых, он содержит биологическую информацию о том, как данному виду выжить и оставить потомство в определенных условиях окружающей среды, а во-вторых, обладает способностью к частичному изменению содержания заключенной в нем биологической информации. Последнее является основой эволюционной и экологической пластичности вида, т.е. возможности приспособиться к существованию в иных условиях, меняющихся в историческом времени или от территории к территории. Популяционная структура вида, приводящая к распаду генофонда вида на генофонды популяций, способствует проявлению в исторической судьбе вида в зависимости от обстоятельств обоих отмеченных качеств генофонда — консервативности
и пластичности.
Таким образом, общебиологическое значение популяционно-видового уровня состоит в реализации элементарных механизмов эволюционного процесса, обусловливающих видообразование.
Значение происходящего на популяционно-видовом уровне для здравоохранения определяется наличием наследственных болезней, заболеваний с очевидной наследственной предрасположенностью, а также выраженными особенностями генофондов разных популяций людей. Процессы, происходящие на этом уровне, в сочетании с экологическими особенностями различных территорий составляют основу перспективного направления современной медицины — эпидемиологии неинфекционных болезней.
Видом
называют совокупность особей, сходных по основным морфологическим и функциональным признакам, кариотипу, поведенческим реакциям, имеющих общее происхождение, заселяющих определенную территорию (ареал), в природных условиях скрещивающихся исключительно между собой и при этом производящих плодовитое потомство.
Видовая принадлежность особи определяется по соответствию ее перечисленным критериям: морфологическому, физиолого-биохимическому, цитогенетическому, этологическому, экологическому и др. Наиболее важные признаки вида — его генетическая
(репродуктивная
) изоляция,
заключающаяся в нескрещиваемости особей данного вида с представителями других видов, а также генетическая устойчивость в природных условиях,
приводящая к независимости эволюционной судьбы.
Со времен К. Линнея вид является основной единицей систематики. Особое положение вида среди других систематических единиц (таксонов) обусловливается тем, что это та группировка, в которой отдельные особи существуют реально.
В составе вида в природных условиях особь рождается, достигает половой зрелости и выполняет свою главную биологическую функцию: участвуя в репродукции, обеспечивает продолжение рода. В отличие от вида таксоны надвидового ранга, такие, как род, отряд, семейство, класс, тип, не являются ареной реальной жизни организмов. Выделение их в естественной системе органического мира отражает результаты предшествующих этапов исторического развития живой природы. Распределение организмов по надвидовым таксонам указывает на степень их филогенетического родства.
Важнейшим фактором объединения организмов в виды служит половой процесс.
Представители одного вида, скрещиваясь друг с другом, обмениваются наследственным материалом. Это ведет к перекомбинации в каждом поколении генов (аллелей), составляющих генотипы отдельных особей. В результате достигаются нивелировка различий между организмами внутри вида
и длительное сохранение основных морфологических, физиологических и прочих признаков, отличающих один вид от другого. Благодаря половому процессу происходит также объединение генов (аллелей), распределенных по генотипам разных особей, в общий генофонд
(аллелофонд)1
вида. Этот генофонд заключает в себе весь объем наследственной информации, которым располагает вид на определенном этапе его существования.
Определение вида, приведенное выше, не может быть применено к размножающимся бесполым путем агамным (некоторые микроорганизмы, синезеленые водоросли), самооплодотворяющимся и строго партеногетическим организмам. Группировки таких организмов, эквивалентные виду, выделяют по сходству фенотипов, общности ареала, близости генотипов по происхождению. Практическое использование понятия «вид» даже у организмов с половым размножением нередко затруднено. Это обусловлено динамичностью видов,
проявляющейся во внутривидовой изменчивости, «размытости» границ ареала, образовании и распаде внутривидовых группировок различного объема и состава (популяций, рас, подвидов). Динамичность видов является следствием действия элементарных эволюционных факторов (см. гл. 11).
В природных условиях организмы одного вида расселены неравномерно. Имеет место чередование участков повышенной и пониженной концентрации особей (рис. 10.1). В результате вид распадается на группировки или популяции, соответствующие зонам более плотного заселения. «Радиусы индивидуальной активности» отдельных особей ограничены. Так, виноградная улитка способна преодолеть расстояние в несколько десятков метров, ондатра —в несколько сотен метров, песец — в несколько сотен километров. Благодаря этому размножение (репродуктивные ареалы) в основном приурочено к территориям с повышенной плотностью организмов.
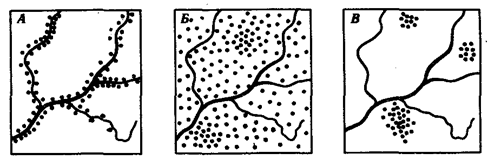
Рис. 10.1. Неравномерное расселение особей по ареалу вида.
А—
ленточный; Б—
пятнистый; В—
островной типы
Вероятность случайных скрещиваний (панмиксии), обусловливающих из поколения в поколение эффективную перекомбинацию генов, внутри «сгущений» оказывается выше, чем в зонах между ними и для вида в целом. Таким образом, в репродуктивном процессе генофонд вида представлен генофондами популяций.
Популяцией
называют минимальную самовоспроизводящуюся группу особей одного вида, населяющих определенную территорию (ареал) достаточно долго (в течение многих поколений). В популяции фактически осуществляется сравнительно высокий уровень панмиксии, и она в определенной степени отделена от других популяций той или иной формой изоляции1
.
10.2.1. Экологическая характеристика популяции
Экологически популяция характеризуется величиной, оцениваемой по занимаемой территории (ареалу), численности особей, возрастному и половому составу. Размеры ареала
зависят от радиусов индивидуальной активности организмов данного вида и особенностей природных условий на соответствующей территории. Численность особей
в популяциях организмов разных видов различается. Так, количество стрекоз Leucorrhinia
albifrons
в популяции на одном из подмосковных озер достигало 30 000, тогда как численность земляной улитки Cepaea
nemoralis
оценивалась в 1000 экземпляров. Существуют минимальные значения численности, при которых популяция способна поддерживать себя во времени. Сокращение численности ниже этого минимума приводит к вымиранию популяции.
Величина популяции постоянно колеблется, что зависит от изменений экологической ситуации. Так, осенью благоприятного по кормовым условиям года популяция диких кроликов на одном из островов у юго-западного побережья Англии состояла из 10 000 особей. После холодной малокормной зимы число особей снизилось до 100.
Возрастная структура
популяций организмов разных видов варьирует в зависимости от продолжительности жизни, интенсивности размножения, возраста достижения половой зрелости. В зависимости от вида организмов она может быть то более, то менее сложной. Так, у стадных млекопитающих, например дельфинов белух Delphinapterus
leucas
,
в популяции одновременно находятся детеныши текущего года рождения, подросший молодняк прошлого года рождения, половозрелые, но, как правило, не размножающиеся животные в возрасте 2—3 лет, взрослые размножающиеся особи в возрасте 4—20 лет. С другой стороны, у землероек Sorex
весной рождаются 1—2 приплода, вслед за чем взрослые особи вымирают, так что осенью вся популяция состоит из молодых неполовозрелых животных.
Половой состав
популяций обусловливается эволюционно закрепленными механизмами формирования первичного (на момент зачатия), вторичного (на момент рождения) и третичного (во взрослом состоянии) соотношения полов. В качестве примера рассмотрим изменение полового состава популяции людей. На момент рождения оно составляет 106 мальчиков на 100 девочек, в возрасте 16—18 лет выравнивается, в возрасте 50 лет насчитывает 85 мужчин на 100 женщин, а в возрасте 80 лет — 50 мужчин на 100 женщин.
10.2.2. Генетические характеристики популяции
Генетически популяция характеризуется ее генофондом (аллелофондом). Он представлен совокупностью аллелей, образующих генотипы организмов данной популяции. Генофонды природных популяций отличает наследственное разнообразие (генетическая гетерогенность, или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие
заключается в присутствии в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости.
Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Так, при скрещивании организмов, различающихся по 1000 локусов1
, каждый из которых представлен десятью аллелями, количество вариантов генотипов достигает 101000
, что превосходит число электронов во Вселенной.
Генетическое единство
популяции обусловливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяции. Генетическое единство проявляется также в общей генотипической изменчивости популяции при изменении условий существования, что обусловливает как выживание вида, так и образование новых видов.
10.2.3. Частоты аллелей. Закон Харди — Вайнберга
В пределах генофонда популяции доля генотипов, содержащих разные аллели одного гена; при соблюдении некоторых условий из поколения в поколение не изменяется. Эти условия описываются основным законом популяционной генетики, сформулированным в 1908 г. английским математиком Дж. Харди и немецким врачом-генетиком Г. Вайнбергом. «В популяции из бесконечно большого числа свободно скрещивающихся особей
в отсутствие мутаций, избирательной миграции
организмов с различными генотипами и давления естественного отбора
первоначальные частоты аллелей сохраняются из поколения в поколение».
Допустим, что в генофонде популяции, удовлетворяющей описанным условиям, некий ген представлен аллелями А1
и А2
, обнаруживаемыми с частотой р и q. Так как других аллелей в данном генофонде не встречается, то р+q = 1. При этом q = 1—р.
Соответственно особи данной популяции образуют р гамет с аллелем А1
и q гамет с аллелем А2
. Если скрещивания происходят случайным образом, то доля половых клеток, соединяющихся с гаметами А1
,равна р, а доля половых клеток, соединяющихся с гаметами A2
, — q. Возникающее в результате описанного цикла размножения поколение F1
образовано генотипами Al
A1
, A1
A2
, A2
A2
, количество которых соотносится как (р + q) (р + q) = р2
+ 2pq + q2
(рис. 10.2). По достижении половой зрелости особи AlAi и АгА2 образуют по одному типу гамет — A1
или A2
— с частотой, пропорциональной числу организмов указанных генотипов (р и q). Особи A1
A2
образуют оба типа гамет с равной частотой 2pq/2.
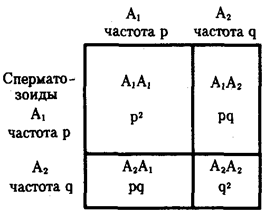
Рис. 10.2. Закономерное распределение генотипов в ряду поколений в зависимости от частоты образования гамет разных типов (закон Харди—Вайнберга)
Таким образом, доля гамет A1
в поколении F1
составит р2
+ 2pq/2 = р2
+ р(1—р) = p, а доля гамет А2
будет равна q2
+ 2pq/2 = q2
+ + q(l-q) = q.
Так как частоты гамет с разными аллелями в поколении fiв сравнении с родительским поколением не изменены, поколение F2 будет представлено организмами с генотипами Al
A1
, A1
A2
и А2
А2
в том же соотношении р2
+ 2pq + q2
. Благодаря этому очередной цикл размножения произойдет при наличии р гамет A1
и q гамет А2
. Аналогичные расчеты можно провести для локусов с любым числом аллелей. В основе сохранения частот аллелей лежат статистические закономерности случайных событий в больших выборках.
Уравнение Харди—Вайнберга в том виде, в котором оно рассмотрено выше, справедливо для аутосомных генов. Для генов, сцепленных с полом, равновесные частоты генотипов Al
A1
, A1
A2
и А2
А2
совпадают с таковыми для аутосомных генов: р2
+ 2pq + q2
. Для самцов (в случае гетерогаметного пола) в силу их гемизиготности возможны лишь два генотипа A1
—или А2
—, которые воспроизводятся с частотой, равной частоте соответствующих аллелей у самок в предшествующем поколении: р и q. Из этого следует, что фенотипы, определяемые рецессивными аллелями сцепленных с хромосомой Х генов, у самцов встречаются чаще, чем у самок.
Так, при частоте аллеля гемофилии, равной 0,0001, это заболевание у мужчин данной популяции наблюдается в 10 000 раз чаще, чем у женщин (1 на 10 тыс. у первых и 1 на 100 млн. у вторых).
Еще одно следствие общего порядка заключается в том, что в случае неравенства частоты аллеля у самцов и самок разность между частотами в следующем поколении уменьшается вдвое, причем меняется знак этой разницы. Обычно требуется несколько поколений для того, чтобы возникло равновесное состояние частот у обоих полов. Указанное состояние для аутосомных генов достигается за одно поколение.
Закон Харди — Вайнберга описывает условия генетической стабильности популяции.
Популяцию, генофонд которой не изменяется в ряду поколений, называют менделевской.
Генетическая стабильность менделевских популяций ставит их вне процесса эволюции, так как в таких условиях приостанавливается действие естественного отбора. Выделение менделевских популяций имеет чисто теоретическое значение. В природе эти популяции не встречаются. В законе Харди — Вайнберга перечислены условия, закономерно изменяющие генофонды популяций. К указанному результату приводят, например, факторы, ограничивающие свободное скрещивание (панмиксию), такие, как конечная численность организмов в популяции, изоляционные барьеры, препятствующие случайному подбору брачных пар. Генетическая инертность преодолевается также благодаря мутациям, притоку в популяцию или оттоку из нее особей с определенными генотипами, отбору.
10.2.4. Место видов и популяций
в эволюционном процессе
Вследствие общей адаптивной (приспособительной) направленности эволюции виды, возникающие в результате этого процесса, являются совокупностями организмов, так или иначе приспособленных к определенной среде. Эта приспособленность сохраняется на протяжении длительного ряда поколений благодаря наличию в генофондах и передаче потомству при размножении соответствующей биологической информации. Из этого следует, что при мало меняющихся условиях обитания сохранность вида во времени зависит от стабильности, консерватизма его генофонда. С другой стороны, стабильные генофонды не обеспечивают выживания в случае изменения условий жизни в историческом развитии планеты. Такие генофонды дают меньше возможностей для расширения ареала вида и освоения новых экологических ниш в текущий исторический период.
Популяционная структура вида позволяет совместить долговременность приспособлений, сформировавшихся на предшествующих этапах развития, с эволюционными и экологическими перспективами. Генофонд вида фактически распадается на генофонды популяций, каждый из которых отличается собственным направлением изменчивости. Популяции — это генетически открытые в рамках вида группировки организмов.
Межпопуляционные миграции особей, сколь бы незначительными они ни были, препятствуют углублению различий и объединяют популяции в единую систему вида. Однако в случае длительной изоляции некоторых популяций от остальной части вида первоначально минимальные различия нарастают. В конечном итоге это приводит к генетической (репродуктивной) изоляции, что и означает появление нового вида. В эволюционный процесс непосредственно включены отдельные популяции, а завершается он образованием вида.
Таким образом, популяция
является элементарной эволюционной единицей, тогда как вид —
качественным этапом эволюции, закрепляющим ее существенный результат.
Согласно синтетической теории эволюции, элементарное эволюционное явление, с
которого начинается видообразование, заключается в изменении генетического состава (генетической конституции, или генофонда) популяции. События и процессы, способствующие преодолению генетической инертности популяций и приводящие к изменению их генофондов, называют элементарными эволюционными факторами.
Важнейшими из них являются мутационный процесс, популяционные волны, изоляция, естественный отбор.
Изменения наследственного материала половых клеток в виде генных, хромосомных и геномных мутаций происходят постоянно. Особое место принадлежит генным мутациям.
Они приводят к возникновению серий аллелей и, таким образом, к разнообразию содержания биологической информации.
Вклад мутационного процесса в видообразование носит двоякий характер. Изменяя частоту одного аллеля по отношению к другому, он оказывает на генофонд популяции прямое действие. Еще большее значение имеет формирование за счет мутантных аллелей резерва наследственной изменчивости. Это создает условия для варьирования аллельного состава генотипов организмов в последовательных поколениях путем комбинативной изменчивости. Благодаря мутационному процессу поддерживается высокий уровень наследственного разнообразия природных популяций. Совокупность аллелей, возникающих в результате мутаций, составляет исходный элементарный эволюционный материал.
В процессе видообразования он используется как основа действия других элементарных эволюционных факторов.
Хотя отдельная мутация — событие редкое, общее число мутаций значительно. Допустим, что некая мутация возникает с частотой 1 на 100 000 гамет, количество локусов в геноме составляет 10 000, численность особей в одном поколении равна 10 000, а каждая особь производит 1000 гамет. При таких условиях по всем локусам за поколение в генофонде вида произойдет 106
мутаций. За среднее время существования вида, равное нескольким десяткам тысяч поколений, количество мутаций составит 1010
. Большинство мутаций первоначально оказывает на фенотип особей неблагоприятное действие. В силу рецессивности мутантные аллели обычно присутствуют в генофондах "популяций в гетерозиготных по соответствующему локусу генотипах.
Благодаря этому достигается тройственный положительный результат: 1) исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном; 2) сохраняются нейтральные мутации, не имеющие приспособительной ценности в настоящих условиях существования, но которые смогут приобрести такую ценность в будущем; 3) накапливаются некоторые неблагоприятные мутации, которые в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов (эффект гетерозиса). Таким образом создается резерв наследственной изменчивости популяции.
Доля полезных мутаций мала, однако их абсолютное количество в пересчете на поколение или период существования вида может быть большим. Допустим, что одна полезная мутация приходится на 1 млн. вредных. Тогда в рассматриваемом выше примере среди 106
мутаций за одно поколение 104
будет полезной. За время существования вида его генофонд обогатится 104
полезными мутациями.
Мутационный процесс, выполняя роль элементарного эволюционного фактора, происходит постоянно на протяжении всего периода существования жизни, а отдельные мутации возникают многократно у разных организмов. Генофонды популяций испытывают непрерывное давление мутационного процесса.
Это обеспечивает накопление мутаций, несмотря на высокую вероятность потери в ряду поколений единичной мутации.
Популяционными волнами
или волнами жизни
(С.С. Четвериков) называют периодические или апериодические колебания численности организмов в природных популяциях. Это явление распространяется на все виды животных и растений, а также на микроорганизмы. Причины колебаний часто имеют экологическую природу. Так, размеры популяций «жертвы» (зайца) растут при снижении давления на них со стороны популяций «хищника» (рыси, лисицы, волка). Отмечаемое в этом случае увеличение кормовых ресурсов способствует росту численности хищников, что, в свою очередь, интенсифицирует истребление жертвы (рис. 11.1).
Вспышки численности организмов некоторых видов, наблюдавшиеся в ряде регионов мира, были обусловлены деятельностью человека.В XIX—XX вв. это относится к популяциям кроликов в Австралии, домовых воробьев в Северной Америке, канадской элодеи в Евразии. В настоящее время существенно возросли размеры популяций домовой мухи, находящей прекрасную кормовую базу в виде разлагающихся пищевых отбросов вблизи поселений человека. Напротив, численность 12 популяций домовых воробьев в городах падает вследствие прекращения широкого использования лошадей. Масштабы колебаний численности организмов разных видов варьируют. Для одной из зауральских популяций майских жуков отмечены изменения количества особей в 106
раз.
Рис. 11.1 Колебания численности особей в популяциях жертвы (заяц-беляк, сплошная линия) и хищников (А -
рыси; B
- лисицы; Д
- волка)
Численность особей выражена в процентах к минимальному из зарегистрированных значений, принятому за 100%
Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. При росте численности организмов наблюдается слияние ранее разобщенных популяций и объединение их генофондов.
Так как популяции по своему генетическому составу уникальны, в результате такого слияния возникают новые генофонды с измененными по сравнению с исходными частотами аллелей. В условиях возросшей численности интенсифицируются межпопуляционные миграции особей,
что также способствует перераспределению аллелей. Рост количества организмов обычно сопровождается расширением занимаемой территории.
На гребне популяционной волны некоторые группы особей выселяются за пределы ареала вида и оказываются в необычных условиях существования.
В таком случае они испытывают действие новых факторов естественного отбора. Повышение концентрации особей в связи с ростом их численности усиливает внутривидовую борьбу за существование.
При спаде численности наблюдается распад крупных популяций.
Возникающие малочисленные популяции характеризуются измененными генофондами. В условиях массовой гибели организмов редкие мутантные
аллели могут быть генофондом потеряны. При сохранении редкого аллеля его концентрация в генофонде малочисленной популяции автоматически возрастает. На спаде волны жизни часть популяций, как правило, небольших по размерам, остается за пределами обычного ареала вида. Чаще они, испытывая действие необычных условий жизни, вымирают. Реже, при благоприятном генетическом составе, такие популяции переживают период спада численности. Будучи изолированными от основной массы вида, существуя в необычной среде, они нередко являются родоначальниками новых видов.
Популяционные волны — это эффективный фактор преодоления генетической инертности природных популяций. Вместе с тем их действие на генофонды не является направленным. В силу этого они, так же как и мутационный процесс, подготавливают эволюционный материал к действию других элементарных эволюционных факторов.
Ограничение свободы скрещиваний (панмиксии) организмов называют изоляцией.
Снижая уровень панмиксии, изоляция приводит к увеличению доли близкородственных скрещиваний. Сопутствующая этому гомозиготизация усиливает особенности генофондов популяций, которые создаются вследствие мутаций, комбинативной изменчивости, популяционных волн. Препятствуя снижению межпопуляционных генотипических различий, изоляция является необходимым условием сохранения, закрепления и распространения в популяциях генотипов повышенной жизнеспособности.
В зависимости от природы факторов ограничения панмиксии различают географическую, биологическую и генетическую изоляцию. Географическая изоляция
заключается в пространственном разобщении популяций благодаря особенностям ландшафта в пределах ареала вида — наличию водных преград для «сухопутных» организмов, участков суши для видов-гидробионтов, чередованию возвышенных участков и равнин. Ей способствует малоподвижный или неподвижный (у растений) образ жизни. Так, на Гавайских о-вах популяции наземных улиток занимают долины, разделенные невысокими гребнями. Сухость почвы и редколесье затрудняют преодоление этих гребней моллюсками. Выраженная, хотя и неполная, изоляция в течение многих поколений привела к ощутимым различиям фенотипов улиток из разных долин. В горах о-ва Оаху, например, один из видов улиток Achatinella
mustelina
представлен более чем сотней рас, выделяемых по морфологическим признакам.
Пространственная изоляция может происходить и в отсутствие видимых географических барьеров. Причины ее в таком случае кроются в ограниченных «радиусах индивидуальной активности». Так, у «береговой» рыбы бельдюги Zoarces
viviparus
от устья у концу фьорда уменьшается число позвонков и лучей некоторых плавников. Сохранение изменчивости объясняется оседлым образом жизни бельдюги. Такая изменчивость наблюдается и у подвижных видов животных, например перелетных птиц с гнездовым консерватизмом. Молодь ласточек, например, возвращается с зимовки на место своего рождения и гнездится в радиусе до 2 км от материнского гнезда. Скрещивания у ласточек ограничиваются группой близко селящихся особей. В отличие от разделения барьерами
эту разновидность географической изоляции обозначают как разделение расстоянием.
Биологическая изоляция
возникает вследствие внутривидовых различий организмов и имеет несколько форм. К экологической изоляции
приводят особенности окраски покровов или состава пищи, размножение в разные сезоны, у паразитов — использование в качестве хозяина организмов разных видов. Так, в Молдове есть две несмешивающиеся популяции мышей —желтогорлая лесная мышь и степная. Фактором разделения их является состав пищи. Разобщение популяций способствовало выявлению и усилению особенностей фенотипа степных мышей. Они мельче и имеют иную форму черепа. В описанном примере экологическая изоляция дополняется территориальной. Сезонные расы, выделяемые по месту и времени икрометания, описаны у лососевых, осетровых, карповых рыб.
Длительная экологическая изоляция способствует дивергенции популяций вплоть до образования новых видов. Так, предполагают, что человеческая и свиная аскариды, морфологически очень близкие, произошли от общего предка. Их расхождению, согласно одной из гипотез, способствовал запрет на употребление человеком в пищу свиного мяса, который по религиозным соображениям распространялся длительное время на значительные массы людей.
Этологическая
(поведенческая
) изоляция
существует благодаря особенностям ритуала ухаживания, окраски, запахов, «пения» самок и самцов из разных популяций. Так, подвиды щеглов — седоголовый Carduelis
carduelis
carduelis
и черноголовый С. с. brevirostis
—
имеют выраженные отметины на голове. Серые вороны Corvus
corone
cornix
из крымской и североукраинской популяций, внешне неразличимые, отличаются карканьем.
При физической
(механической
) изоляции
препятствием к скрещиванию являются различия в структуре органов размножения или просто разница в размерах тела. У растений такая форма изоляции возникает при приспособлении цветка к определенному виду опылителей.
Описанные формы изоляции, особенно в начальный период их действия, снижают, но не исключают полностью межпопуляционные скрещивания.
Генетическая
(репродуктивная
) изоляция
создает более жесткие, иногда непреодолимые барьеры скрещиваниям. Она заключается в несовместимости гамет, гибели зигот непосредственно после оплодотворения, стерильности или малой жизнеспособности гибридов.
Иногда разделение популяции сразу начинается с генетической изоляции. К этому приводят полиплоидия или массивные хромосомные перестройки, резко изменяющие хромосомные наборы гамет мутантов по сравнению с исходными формами. Полиплоидия распространена среди растений (рис. 11.2). Разные виды плодовой мухи нередко различаются хромосомными перестройками. Гибриды от скрещивания близкородственных форм со сниженной жизнеспособностью известны для серой и черной ворон. Указанный фактор изолирует популяции этих птиц в Евразии (рис. 11.3). Чаще генетическая изоляция развивается вторично вследствие углубления морфологических различий организмов из популяций, длительно разобщенных другими формами изоляции — географической, биологической. В первом случае генетическая изоляция предшествует дивергенции признаков и начинает процесс видообразования, во втором — она его завершает.


Рис.11.2. Растения и наборы хромосом в соматических клетках Solanumnigrum:
а—г:
36, 72, 108 и 144 хромосомы
Изоляция в процессе видообразования взаимодействует с другими элементарными эволюционными факторами. Она усиливает генотипические различия, создаваемые мутационным процессом и генетической комбинаторикой. Возникающие благодаря изоляции внутривидовые группировки отличаются по генетическому составу и испытывают неодинаковое давление отбора.
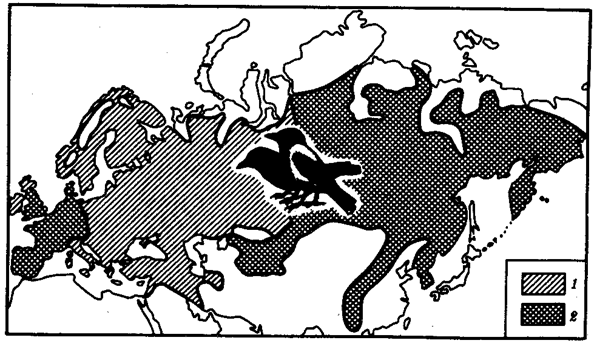
Рис. 11.3. Сниженная жизнеспособность гибридов как фактор разделения популяций серой и черной ворон:
1
— ареал серой вороны, 2 —
ареал черной вороны
В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Благодаря индивидуальной изменчивости в условиях конкретной среды обитания приспособленность разных генотипов (фенотипов) различна. В эволюционном контексте приспособленность
определяют как произведение жизнеспособности в данной среде, обусловливающей большую или меньшую вероятность достижения репродуктивного возраста, на репродуктивную способность особи. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора.
Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции.
Необходимой предпосылкой отбора является борьба за существование — конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная
(избирательная
) смертность.
В конечном итоге отбор обеспечивает дифференциальное
(избирательное
) воспроизведение
(размножение
) генотипов.
Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов.
Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия,
отдельные особи — объектами действия,
а конкретные признаки — точками приложения
отбора.
Эффективность отбора по качественному и количественному изменению генофонда популяции зависит от величины давления и направления его действия. Величину давления отбора
выражают коэффициентом отбора
S, который характеризует интенсивность устранения из репродуктивного процесса или сохранения в нем соответственно менее или более приспособленных форм по сравнению с формой, принятой за стандарт приспособленности. Так, если некий локус представлен аллелями A1
и А2
, то популяция по генотипам делится на три группы: A1
A1
; A1
A2
; A2
A2
. Обозначим приспособленность этих генотипов W0
, W1
, W2
. Выберем в качестве стандарта первый генотип, относительная приспособленность которого максимальна и равна 1. Тогда приспособленности других генотипов составят доли этого стандарта:
Al
A1
A1
A2
A2
A2
W0
W1
W2
или W0
/W0
= 1, W1
/W0
=l—S1
, W2
/W0
=l—S2
.
Величины S1
и S2
означают пропорциональное снижение в очередном поколении воспроизводства генотипов A1
A2
и А2
А2
сравнительно с генотипом Al
A1
.
Отбор особенно эффективен в отношении доминантных аллелей при условии их полного фенотипического проявления и менее эффективен в отношении рецессивных аллелей, а также в условиях неполной пенетрантности. На результат отбора влияет исходная концентрация аллеля в генофонде. При низких и высоких концентрациях отбор происходит медленно. Изменение доли доминантного аллеля в сравнении с рецессивным при коэффициенте отбора 0,01 приведено ниже.
| Возрастание, %
|
Число поколений
|
| 0,1-1 |
250 |
| 1-50 |
500 |
| 50-98 |
5000 |
| 98-99,9 |
90000 |
В теории, упрощая ситуацию, допускают, что отбор через фенотипы действует на генотипы благодаря различиям в приспособительной ценности отдельных аллелей. В реальной жизни приспособительная ценность генотипов зависит от влияния на фенотип и взаимодействия всей совокупности генов. Оценка величины давления отбора
по изменению концентрации отдельных аллелей технически часто невыполнима. Поэтому расчет проводят по изменению концентрации организмов определенного фенотипа.
Пусть в популяции присутствуют организмы двух фенотипических классов А и В в отношении СA
/СB
= U1
. Вследствие различий по приспособленности происходит естественный отбор (селекция), который изменяет соотношение особей с фенотипами А и В. В следующем поколении оно станет СA
/СB
= U2
=U1
(1 + S), где S — коэффициент отбора. Отсюда S = U2
/U1
— 1. При селективном преимуществе фенотипа А U2
> U1
, aS > 0. При селективном преимуществе фенотипа В U2
< U1
и S < 0. Если приспособленность фенотипов А и В сопоставима и U2
= U1
, aS=0. В рассмотренном примере при S > 0 отбор сохраняет в популяции в ряду поколений фенотипы А и устраняет фенотипы В, при S < 0 имеет место обратная тенденция. Отбор, сохраняющий определенные фенотипы, по своему направлению является положительным,
тогда как отбор, устраняющий фенотипы из популяции,— отрицательным.
В зависимости от результата различают стабилизирующую, движущую и дизруптивную формы естественного отбора (рис. 11.4). Стабилизирующий отбор
сохраняет в популяции средний вариант фенотипа или признака. Он устраняет из репродуктивного процесса фенотипы, уклоняющиеся от сложившейся адаптивной «нормы», приводит к преимущественному размножению типичных организмов. Так, сотрудник одного из университетов США подобрал после снегопада и сильного ветра 136 оглушенных воробьев Passer
domesticus
.
Из них 72 выживших воробья имели крылья средней длины, тогда как 64 погибшие птицы были либо длиннокрылыми, либо короткокрылыми. Стабилизирующая форма соответствует консервативной роли естественного отбора.
При относительном постоянстве условий среды благодаря этой форме сохраняются результаты предшествующих этапов эволюции.
Движущий
(направленный
) отбор
обусловливает последовательное изменение фенотипа в определенном направлении, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. При смене условий обитания благодаря этой форме отбора в популяции закрепляется фенотип, более соответствующий среде. После того как новое значение признака придет в оптимальное соответствие условиям среды, движущая форма отбора сменяется стабилизирующей. Примером такого отбора является замещение в популяции гавани Плимут (Англия) крабов Carcinus
maenas
с широким головогрудным щитком животными с узким щитком в связи с увеличением количества ила.
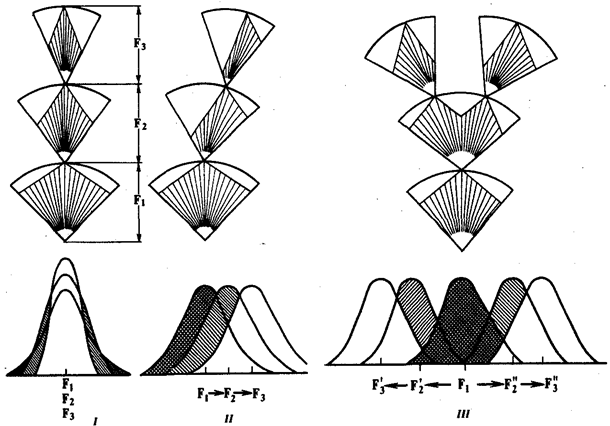
Рис. 11.4. Формы естественного отбора:
I
—стабилизирующая, II—
движущая, III—
дизруптивная;
F1
—
F3
—
последовательные поколения особей
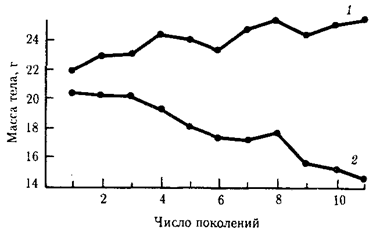
Рис. 11.5. Изменение массы тела мышей из двух популяций в последовательных поколениях вследствие отбора по этому признаку
1
— тяжелые животные, 2
— легкие животные
Направленный отбор составляет основу искусственного отбора. Так, в одном эксперименте на протяжении ряда поколений из популяции шестинедельных мышей отбирали для скрещивания наиболее тяжелых и наиболее легких животных. Избирательное воспроизведение по признаку массы тела привело к образованию двух самостоятельных популяций, соответственно с возрастающей и убывающей массой тела (рис. 11.5). По окончании опыта, занявшего 11 поколений, ни одна из этих популяций не вернулась к первоначальной массе.
Дизруптивный
(разрывающий
) отбор
сохраняет несколько разных фенотипов с равной приспособленностью. Он действует против особей со средним или промежуточным значением признаков. Так, в зависимости от преобладающего цвета почвы улитки Cepaea
nemoralis
имеют раковины коричневой, желтой, розовой окраски. Дизруптивная форма отбора «разрывает» популяцию по определенному признаку на несколько групп. Она поддерживает в популяции состояние генетического полиморфизма.
В зависимости от формы отбор сокращает масштабы изменчивости, создает новую или сохраняет прежнюю картину разнообразия. Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения соотношений аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направленны. Отбор приводит генофонды в соответствие с критерием приспособленности. Он осуществляет обратную связь между изменениями генофонда и условиями обитания, накладывает на эти изменения печать биологической целесообразности (полезности). Естественный отбор действует совместно с другими эволюционными факторами. Поддерживая генотипическое разнообразие особей в ряду поколений, мутационный процесс, а также популяционные волны, комбинативная изменчивость создают для него необходимый материал.
Естественный отбор нельзя рассматривать как «сито», сортирующее генотипы по приспособленности. В эволюции ему принадлежит творческая роль.
Исключая из размножения генотипы с малой приспособительной ценностью, сохраняя благоприятные генные комбинации разного масштаба, он преобразует картину генотипической изменчивости, складывающуюся первоначально под действием случайных факторов, в биологически целесообразном направлении. Результатом творческой роли отбора является процесс органической эволюции, идущей в целом по линии прогрессивного усложнения морфофизиологической организации (арогенез), а в отдельных ветвях — пути специализации (аллогенез).
Мутации и комбинативная изменчивость, периодические колебания численности организмов, изоляция изменяют генофонды популяций случайным образом. Их совместное действие с естественным отбором в процессе видообразования придает биологической изменчивости в целом приспособительный характер. Выполнению отбором упорядочивающей роли препятствуют изменения частот аллелей, зависящие от случайных причин. Таковыми в данном случае являются причины, обусловливающие преимущественное размножение генотипов вне связи с их приспособительной ценностью. Так как динамика частот аллелей в генофондах последовательных поколений носит статистический характер (см. закон Харди — Вайнберга), размах случайных колебаний этого показателя возрастает по мере снижения численности особей в популяции.
Расчеты показывают, что при воспроизведении 5000 потомков родительской популяции с частотой некоего аллеля р = 0,50 колебания концентрации этого аллеля в 99,994% вариантов дочерних популяций в силу случайных причин (в отсутствие отбора по этому аллелю) не выйдут за пределы 0,48—0,52. Если же родительская популяция мала и воспроизводит 50 потомков, то размах случайных колебаний концентрации наблюдаемого аллеля в том же проценте вариантов дочерних популяций составит 0,30—0,70. Случайные, но не обусловленные действием естественного отбора колебания частот аллелей называют генетико-автоматическими процессами
или дрейфом генов.
При значительном размахе колебаний в последовательных поколениях создаются условия для потери популяцией некоторых аллелей и закрепления других. В результате происходят гомозиготизация особей и затухание изменчивости.
Предположим, что популяция состоит из четырех особей и имеет аллель с частотой р = 0,125. Это означает, что указанный аллель присутствует в генофонде данной популяции в единственном экземпляре у одной из особей, гетерозиготной по соответствующему локусу. Любое случайное стечение обстоятельств, исключающее такую особь из размножения (лесной пожар, выстрел охотника и т.п.), приведет к утрате аллеля. Генофонд дочерней популяции будет его лишен. Вероятность утраты составит 1/2 в случае одного, 1/4 — двух, 1/8 — трех потомков у данной особи. В популяции из 4000 организмов при р = 0,125 минимум 500 особей имеют соответствующий аллель, причем в гомозиготном состоянии. Вероятность исключения всех этих особей из размножения в силу случайных обстоятельств ничтожно мала. Это гарантирует переход аллеля в генофонд следующего поколения и его сохранение.
Дрейф генов обусловливает утрату (р = 0) или закрепление аллелей в гомозиготном состоянии у всех членов популяции (р = 1) вне связи с их приспособительной ценностью. Он играет важную роль в формировании генофондов малочисленных групп организмов, изолированных от остальной части вида.
Процесс образования видов осуществляется в результате взаимодействия элементарных эволюционных факторов. Видообразование
в типичных случаях заключается в разделении первоначально единого вида на два или более новых. Это связано с возникновением межпопуляционных изоляционных барьеров и углублением различий между генофондами популяций под действием естественного отбора вплоть до генетической изоляции. Такой процесс, ведущий к увеличению количества видов, называют дивергентным
или истинным видообразованием.
Выделяют также филетическое видообразование.
Оно заключается в постепенном превращении во времени одного вида в другой. Этот способ наблюдается, если изменения условий захватывают весь ареал. Известны примеры видообразования путем гибридизации.
Перечисленные способы видообразования схематически изображены на рис. 11.6.
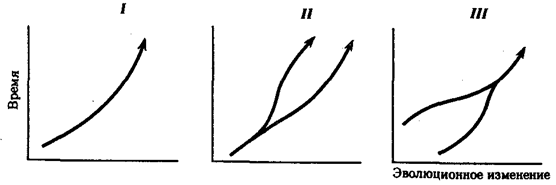
Рис. 11.6. Основные способы видообразования:
I
—филетическое, II
—дивергентное (истинное), III
—путем гибридизации
Существуют аллопатрический и симпатрический пути образования видов. При аллопатрическом видообразовании,
называемом также географическим, препятствия к скрещиванию первично обусловлены пространственным разобщением популяций. Генетическая изоляция развивается вторично. Так, некогда в Австралии существовал один вид попугайчиков рода Pachycephala
.
В условиях засушливого периода единый ареал разделился на западную и восточную зоны. Со временем особи двух популяций приобрели морфофизиологические различия, которые сделали невозможным скрещивание, когда ареал вновь стал общим. Произошло образование из одного предкового вида двух новых.
При симпатрическом видообразовании
новый вид образуется внутри ареала исходного вида. С самого начала изоляция является генетической. Такое положение создается в результате полиплоидии вследствие нарушений нормального хода мейоза, при крупных хромосомных перестройках или межвидовой гибридизации. Аллопатрическое видообразование происходит медленно и дает виды, как правило, отличающиеся по морфофизиологическому критерию от вида-родоначальника. Симпатрический путь относительно быстрый и дает виды, близкие к исходному по морфофизиологическим показателям.
Большинство видов, особенно животных, возникают аллопатрическим путем. Симпатрическое видообразование на основе полиплоидии характерно для растений. Так, разные виды пшениц составляют ряд с наборами 14, 28, 42 хромосомы. В клетках дикого хлопчатника 26 хромосом, культурного — 52. Культурная слива возникла путем гибридизации терна с алычой. Примером гибридогенного вида является рябинокизильник, распространенный в лесах центральной Сибири. Симпатрический путь видообразования у паразитов часто связан с освоением популяцией новых хозяев. Анализ генного состава и межхромосомных различий между человеком и человекообразными обезьянами дает повод предположить, что разделение этих двух ветвей могло идти симпатрическим путем.
Процесс видообразования с участием такого фактора, как естественный отбор, создает разнообразие живых форм, приспособленных к условиям обитания. Среди разных генотипов, возникающих в каждом поколении благодаря резерву наследственной изменчивости и перекомбинации аллелей, лишь ограниченное число обусловливает максимальную приспособленность к конкретной среде. Можно предположить, что дифференциальное воспроизведение этих генотипов в конце приведет к тому, что генофонды популяций будут представлены лишь «удачными» аллелями и их комбинациями. В итоге произойдет затухание наследственной изменчивости и повышение уровня гомозиготности генотипов.
В природных популяциях, однако, наблюдается противоположное состояние. Большинство организмов являются высокогетерозиготными. Отдельные особи гетерозиготны частично по разным локусам, что повышает суммарную гетерозиготность популяции. Так, методом электрофореза на 126 особях рачка Euphausia
superba
,
представляющего главную пищу китов в антарктических водах, изучали 36 локусов, кодирующих первичную структуру ряда ферментов. По 15 локусам изменчивость отсутствовала. По 21 локусу имелось по 3—4 аллеля. В целом в этой популяции рачков 58% локусов были гетерозиготными и имели по 2 аллеля и более. В среднем у каждой особи по 5,8% гетерозиготных локусов. Средний уровень гетерозиготности у растений составляет 17%, беспозвоночных — 13,4, позвоночных — 6,6%. У человека этот показатель равен 6,7%. Столь высокий уровень гетерозиготности нельзя объяснить только мутациями в силу относительной их редкости.
Наличие в популяции нескольких равновесно сосуществующих генотипов в концентрации, превышающей по наиболее редкой форме 1%1
, называют полиморфизмом.
Наследственный полиморфизм создается мутациями и комбинативной изменчивостью. Он поддерживается естественным отбором и бывает адаптационным (переходным) и гетерозиготным (балансированным).
Адаптационный полиморфизм
возникает, если в различных, но закономерно изменяющихся условиях жизни отбор благоприятствует разным генотипам. Так, в популяциях двухточечных божьих коровок Adalia
bipunctata
при уходе на зимовку преобладают черные жуки, а весной—красные (рис. 11.7). Это происходит потому, что красные формы лучше переносят холод, а черные интенсивнее размножаются в летний период.
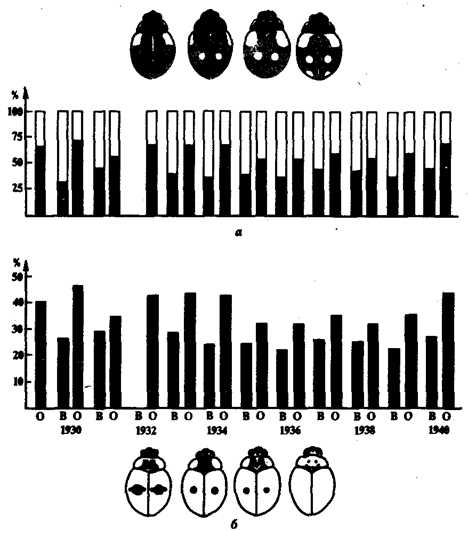
Рис. 11.7. Адаптационный полиморфизм у двухточечных божьих коровок:
а—
соотношение черной (зачернено) и красной форм при весеннем (В) и осеннем (О) сборе; б—
частота доминантного аллеля черной окраски в весенней и осенней популяциях
Балансированный полиморфизм
возникает, если отбор благоприятствует гетерозиготам в сравнении с рецессивными и доминантными гомозиготами. Так, в опытной численно равновесной популяции плодовых мух Drosophila
melanogaster
,
содержащей поначалу много мутантов с более темными телами (рецессивная мутация ebony), концентрация последних быстро падала, пока не стабилизировалась на уровне 10% (рис. 11.8). Анализ показал, что в созданных условиях гомозиготы по мутации ebony и гомозиготы по аллелю дикого типа менее жизнеспособны, чем гетерозиготные мухи. Это и создает состояние устойчивого полиморфизма по соответствующему локусу.

Рис. 11.8. Балансированный полиморфизм по локусу окраски тела в опытной популяции плодовых мух: I
—серая муха (дикий тип), II—
мутантная муха с черной окраской тела
Явление селективного преимущества гетерозигот называют сверхдоминантностью.
Механизм положительного отбора гетерозигот различен. Правилом является зависимость интенсивности отбора от частоты, с которой встречается соответствующий фенотип (генотип). Так, рыбы, птицы, млекопитающие предпочитают обычные фенотипические формы добычи, «не замечая» редких.
В качестве примера рассмотрим результаты наблюдений, выполненных на обыкновенной наземной улитке Cepaea
nemoralis
,
раковина у которой бывает желтая, различных оттенков коричневого цвета, розовая, оранжевая или красная. На раковине может быть до пяти темных полос. При этом коричневая окраска доминирует над розовой, а они обе — над желтой. Полосатость является рецессивным признаком. Улитки поедаются дроздами, использующими камень как наковальню, чтобы разбить раковину и добраться до тела моллюска. Подсчет числа раковин разной окраски вокруг таких «наковален» показал, что на траве или лесной подстилке, фон которых достаточно однороден, добычей птиц чаще оказывались улитки с розовой и полосатой раковиной. На пастбищах с грубыми травами или в живых изгородях с более пестрым фоном чаще поедались улитки, раковины которых окрашены в светлые тона и не имели полос.
Самцы относительно редких генотипов могут иметь повышенную конкурентоспособность за самок. Селективное преимущество гетерозигот обусловливается также явлением гетерозиса. Повышенная жизнеспособность межлинейных гибридов отражает, по-видимому, результат взаимодействия аллельных и неаллельных генов в системе генотипов в условиях гетерозиготности по многим локусам. Гетерозис наблюдается в отсутствие фенотипического проявления рецессивных аллелей. Это сохраняет скрытыми от естественного отбора неблагоприятные и даже летальные рецессивные мутации.
В силу разнообразия факторов среды обитания естественный отбор действует одновременно по многим направлениям. При этом конечный результат зависит от соотношения интенсивности разных векторов отбора.
Конечный результат естественного отбора в популяции зависит от наложения многих векторов отборов и контротборов. Благодаря этому достигается одновременно и стабилизация генофонда, и поддержание наследственного разноообразия.
Балансированный полиморфизм придает популяции ряд ценных свойств, что определяет его биологическое значение. Генетически разнородная популяция осваивает более широкий спектр условий жизни, используя среду обитания более полно. В ее генофонде накапливается больший объем резервной наследственной изменчивости. В результате она приобретает эволюционную гибкость и может, изменяясь в том или ином направлении, компенсировать колебания среды в ходе исторического развития.
В генетически полиморфной популяции из поколения в поколение рождаются организмы генотипов, приспособленность которых неодинакова. В каждый момент времени жизнеспособность такой популяции ниже уровня, который был бы достигнут при наличии в ней лишь наиболее «удачных» генотипов. Величину, на которую приспособленность реальной популяции отличается от приспособленности идеальной популяции из «лучших» генотипов, возможных при данном генофонде, называют генетическим грузом.
Он является своеобразной платой за экологическую и эволюционную гибкость. Генетический груз — неизбежное следствие генетического полиморфизма.
Наряду с общей приспособленностью, .зависящей от генотипа в целом и измеряемой выживаемостью и успехом в размножении, в процессе эволюции возникают приспособления,
или адаптации, для решения организмом экологических задач,
предъявляемых средой обитания. Отдельные адаптации — это постоянно возникающие в процессе развития жизни, изменяющиеся, самосовершенствующиеся, иногда исчезающие, эволюционно обусловленные приспособления к конкретным факторам среды. В результате выработки адаптации достигается состояние адаптированности,
или соответствия морфологии, физиологии, поведения организмов занимаемым ими экологическим нишам. Под экологической нишей
понимают всю совокупность условий среды и образа жизни данного организма.
Процесс выработки адаптации происходит постоянно. В него вовлечены многие признаки организма. Эволюция птиц от рептилий включала, например, последовательные изменения костей, мышц, покровов, конечностей. Увеличение грудины, перестройка гистологической структуры костей, придавшей им наряду с прочностью легкость, развитие оперения, обусловившего лучшие аэродинамические свойства и терморегуляцию, превращение пары конечностей в крылья, обеспечило решение проблемы полета. У некоторых представителей птиц впоследствии развились приспособления к наземному или водному образу жизни (страус, пингвин). Вторичные адаптации захватили также ряд признаков: пингвины, например, сменили крылья на плавники, а их покровы стали водонепроницаемы.
Одна экологическая задача может решаться путем выработки разных адаптации. Так, средством термоизоляции у медведей, песцов является густой мех, а у китообразных — жировой подкожный слой.
Адаптации возникают в ответ на конкретную экологическую задачу. В силу этого они всегда относительны. Относительность адаптации заключается в ограниченности их приспособительного значения определенными условиями обитания. Так, приспособительная ценность пигментированности бабочек березовых пядениц по сравнению со светлыми формами очевидна лишь на закопченных стволах деревьев. Приведенный пример показывает также, что о степени адаптивности признака можно судить лишь сравнив два его разных состояния.
Приспособление образуется только при наличии в генофонде вида наследственной информации, позволяющей изменить структуру и функции в требуемом направлении. Так, млекопитающие и насекомые используют для дыхания соответственно легкие и трахеи, которые развиваются из разных зачатков под контролем разных генов. Нередко основу нового приспособления составляет предсуществующая структура. Последняя выполняла другие функции, но изменилась в таком направлении и до такой степени, что смогла взять на себя новые функции.
Наличие структур, способных расширить или изменить круг функций, называют преадаптацией.
Например, когда-то у рыб, обитавших в мелководных водоемах со стоячей и бедной кислородом водой, появились полые выросты в передней части пищевода и мускулистые плавники. Первая структура способствовала решению задачи дыхания, а вторая — перемещения по грунту. Они позволили некоторым рыбам покидать на время водоемы. Первоначально такие выходы совершались, видимо, в дождливые дни или влажные ночи. Именно так делает в настоящее время американский сомик-кошка Ictalurus
nebulosus
.
Впоследствии эти структуры развились в легкие и конечности наземных животных. К адаптации иногда приводит новая мутация. Включившись в систему генотипа, она изменяет фенотип в направлении более эффективного решения экологических задач. Этот путь возникновения адаптации называют комбинативным.
Есть несколько классификаций адаптации. По механизму действия
выделяют приспособления пассивной защиты (высокая плодовитость;
покровительственная, отпугивающая окраска), активной защиты, перемещения и добывания пищи (совершенствование аппарата движения, нервной системы, органов чувств; развитие средств нападения у хищных), к общественному образу жизни (разделение «труда» у пчел), к сожительству с другими организмами (симбиоз, паразитизм).
В зависимости от характера изменения
различают адаптации с усложнением или упрощением морфофизиологической организации. Последнее типично для паразитов. По масштабу
приспособления делят на специализированные и общие. С помощью специализированных адаптации организм решает конкретные задачи в узкоограниченных условиях жизни вида. К примеру, особенности строения языка муравьеда таковы, что обеспечивают питание муравьями. Общие адаптации позволяют решать многие задачи в широком спектре условий среды. К ним относят внутренний скелет позвоночных и наружный членистоногих, гемоглобин как переносчик кислорода и др.
При наличии таких адаптации осваиваются разнообразные экологические ниши. Они обеспечивают значительную экологическую и эволюционную пластичность и обнаруживаются у представителей крупных таксонов организмов. Так, первичный роговой покров предковых форм рептилий в процессе исторического развития дал покровы современных рептилий, птиц, млекопитающих. Масштаб приспособления выявляется в ходе эволюции той группы организмов, у которой оно возникло впервые.
Биологическая целесообразность
организации живых существ проявляется в гармонии между морфологией, физиологией, поведением организмов разных видов и средой их обитания. Она заключается также в удивительной согласованности строения и функций отдельных частей и систем самого организма. Сторонники теологического объяснения происхождения жизни видели в биологической целесообразности проявление мудрости Творца природы, будь то Бог или абстрактный разум. Теологическое объяснение биологической целесообразности исходит из принципа «конечной цели», согласно которому жизнь развивается направленно в силу внутренне присущего стремления к известной цели.
Со времен Ж.-Б. Ламарка существуют гипотезы, связывающие целесообразность в мире жизни с принципом биологического соответствия ответа организмов на изменения во внешних условиях и наследованием таких благоприобретенных признаков.
Убедительным доказательством в пользу целесообразности изменений под влиянием среды долго признавался факт «привыкания» микроорганизмов к лекарственным препаратам —сульфаниламидам, антибиотикам. Опыт В. и Э. Ледербергов показал, что это не так. Схема опыта приведена на рис. 11.9.
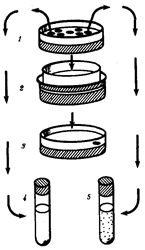
Рис. 11.9. Схема опыта, опровергающего факт непосредственного приспособления микроорганизмов к среде с антибиотиком (пояснение см. в тексте)
В чашке Петри на поверхности твердой питательной среды микроб образует колонии (1
). Специальным штампом отпечаток всех колоний переносили на среду (2
)со смертельной дозой антибиотика (J). Если в таких условиях вырастала хоть одна колония, то она происходила от колонии микробов, также устойчивых к данному препарату. В отличие от других колоний первой чашки Петри (4
) она давала рост в пробирке с антибиотиком (5
).
Если число исходных колоний было велико, то среди них, как правило, находилась и устойчивая.
Таким образом, речь идет не о направленном приспособлении микроба, а о состоянии преадаптации. Оно обусловлено наличием в геноме микроорганизма аллеля, блокирующего действие антибиотика. В одних случаях «устойчивые» микробы образуют фермент, разрушающий лекарственное вещество, в других — стенка клетки непроницаема для препарата. Появлению штаммов микроорганизмов, устойчивых к лекарственным препаратам, способствует неправильная тактика врачей, которые, желая избежать побочных эффектов, назначают низкие для микробов, сублетальные дозы.
Появление форм, резистентных к ядам, наблюдается у насекомых, млекопитающих. Это явление имеет сходную основу: среди мутантных организмов находится устойчивая форма, которая подвергается положительному отбору в условиях действия отравляющего вещества. Например, устойчивость крыс к варфарину, используемому для их уничтожения, зависит от присутствия в генотипе определенного доминантного аллеля. Возможность прямого, непосредственного приспособления организмов к среде обитания допускалась некоторыми биологами еще в 40—50 гг. прошлого столетия.
Биологическая целесообразность строения и функций организмов складывается в процессе развития жизни. Она представляет собой историческую категорию.
Об этом свидетельствует смена типов организации, занимающих господствующее положение в органическом мире планеты. Так, господство амфибий на протяжении почти 75 млн. лет сменилось господством рептилий, затянувшимся на 150 млн. лет. В периоды господства любой группы обычно случается несколько волн вымирания. Они изменяют относительный видовой состав соответствующего крупного таксона, например класса.
Проявление адаптации и биологической целесообразности в целом объясняется действием в природе естественного отбора. Из множества случайных мутаций он сохраняет и накапливает наследственные изменения, имеющие приспособительную ценность. Это объяснение позволяет понять, почему биологическая целесообразность, если ее рассматривать в пространстве и времени, является относительным свойством живых существ. Становится также ясным, почему в конкретных условиях обитания отдельные приспособления достигают лишь той степени развития, которая достаточна для выживания в сравнении с приспособлениями конкурентов.
Размножение человека осуществляется половым путем, а репродуктивные ареалы в той или иной степени ограничены определенной группой населения. Это позволяет выделить в человечестве сообщества, аналогичные популяциям в биологическом понимании этого термина. В антропогенетике популяцией
называют группу людей, занимающих общую территорию и свободно вступающих в брак. Изоляционные барьеры, препятствующие заключению брачных союзов, нередко носят выраженный социальный характер (например, различия в вероисповедании). Благодаря этому в формировании популяций людей главную роль играет не общность территории, а социальные факторы.
Размер, уровень рождаемости и смертности, возрастной состав, экономическое состояние, уклад жизни являются демографическими показателями
популяций людей. Генетически они характеризуются генофондами (аллелофондами). Демографические показатели оказывают серьезное воздействие на состояние генофондов человеческих популяций, главным образом через структуру браков. Большое значение в определении структуры браков имеет размер группы.
Популяции из 1500—4000 человек называют демами
, популяции численностью до 1500 человек — изолятами.
Для демов и изолятов типичен относительно низкий естественный прирост населения — соответственно порядка 20% и не более 25% за поколение. Частота внутригрупповых браков в них составляет 80—90% и свыше 90%, а приток лиц из других групп сохраняется на уровне 1—2% и менее 1%. В силу высокой частоты внутригрупповых браков члены изолятов, просуществовавших четыре поколения (примерно 100 лет) и более, являются не менее чем троюродными братьями и сестрами (сибсами).
В больших по размерам популяциях распределение аллелей отдельных генов в генотипах индивидуумов последовательных поколений подчиняется закону Харди — Вайнберга. Это используют в медико-генетической практике для расчета доли гетерозигот — носителей определенного рецессивного аллеля. Так, в Швеции в 1965—1974 гг. страдающие фенилкетонурией встречались с частотой примерно 1 : 40000. Исходя из закона Харди — Вайнберга, по локусу, представленному двумя аллелями, три возможных генотипа (Al
A1
, A1
A2
и А2
А2
) распределяются с частотой р2
, 2pq, q2
. Следовательно, q2
= 1/40000, aq
=
1/200. Частота доминантного аллеля нормального обмена фенилаланина р=1—q=l— 1/200 = 199/200. Тогда частота гетерозигот 2pq = 2 х (1/200) • (199/200) = 2 • (199/40000). При найденных частотах доминантного и рецессивного аллелей популяция численностью 40 000человек содержит одного больного фенилкетонурией (А2
А2
) и 400 носителей неблагоприятного аллеля в гетерозиготном состоянии (A1
A2
). Остальные члены популяции гомозиготны по благоприятному доминантному аллелю (Al
A1
). Ниже приведены данные о частоте гетерозиготного носительства и соответствующей ей частоте рецессивных гомозигот с фенотипическим проявлением определенного аллеля.
Встречаемость гомозигот
(в пересчете на число членов популяции
)
|
Встречаемость гетерозигот
(в пересчете на число членов популяции
)
|
| 1 : 10 |
1 : 2,3 |
| 1 : 100 |
1 : 5.6 |
| 1 : 1000 |
1 : 16 |
| 1 : 10 000 |
1 : 51 |
| 1 : 100 000 |
1 : 159 |
| 1 : 1 000 000 |
1 : 501 |
Даже по редким рецессивным аллелям количество гетерозигот оказывается достаточно высоким, чтобы это учитывалось при медико-генетическом консультировании вступающих в брак.
Клеточные и физико-химические механизмы наследственности и изменчивости универсальны для всех живых существ, включая человека. Установлена определенная зависимость жизнеспособности индивидуума от особенностей его генотипа. Большую часть своей истории человечество было совокупностью более или менее изолированных в репродуктивном отношении относительно малочисленных групп. Вплоть до настоящего времени сохраняются изоляты. В отдельные исторические периоды происходили миграции значительных масс людей. Они сопровождались объединением ранее разобщенных групп, освоением новых территорий со своими климато-географическими условиями. В настоящее время миграции населения усилились в связи с ростом численности людей, совершенствованием средств транспорта, неравномерньм развитием экономики. Благодаря отмеченному генофонды популяций людей испытывали ранее и продолжают испытывать действие элементарных эволюционных факторов. Социальность человека вносит в это действие определенную специфику.
12.2.1. Мутационный процесс
Мутационный процесс у человека
сходен с таковым у других организмов по всем основным показателям — средней частоте мутирования на локус или геном за поколение, генетико-физиологическим характеристикам мутаций, наличию антимутационных барьеров. Это совпадение неслучайно. Основные характеристики спонтанного мутагенеза формировались на начальных этапах эволюции жизни под действием таких постоянных факторов, как ультрафиолетовое и иные виды излучения, температура, определенная химическая среда.
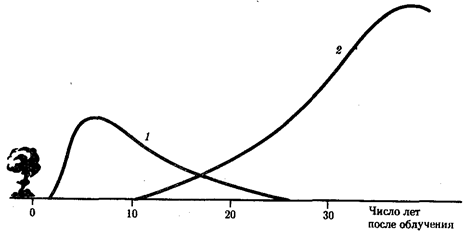
Рис. 12.1. Относительная среднестатистическая вероятность заболевания раком после однократного равномерного облучения тела дозой в 0,01 Гр в популяции людей, переживших атомную бомбардировку:
1
— лейкозы, 2 —
все другие виды раковых заболеваний
Хотя оценка частоты возникновения мутаций у людей встречает серьезные трудности, некоторые подходы к получению таких данных имеются. Согласно одному из них, максимальная определяемая вероятность новой мутации составляла 2,24 · 10-5
на один локус в поколении.
В настоящее время давление мутационного процесса на генофонд человечества, по-видимому, усиливается благодаря росту индуцированных мутаций. Их причиной нередко служат факторы, возникающие в связи с производственной деятельностью человека в условиях научно-технической революции, например ионизирующее излучение. Подсчет прироста количества мутаций сверх фоновых значений встречается с теми же трудностями, о которых шла речь выше. Согласно ориентировочным данным, доза в 1 Гр (грей), получаемая при низком уровне радиации мужчинами, индуцирует от 1000 до 2000 мутаций с серьезными фенотипическими последствиями на каждый миллион живых новорожденных. У женщин эта цифра ниже — 900.
Мутагенные факторы индуцируют мутации как в половых, так и в соматических клетках. В последнем случае результат может состоять в повышении частоты определенных заболеваний, прежде всего злокачественных опухолей. В отношении ионизирующих излучений, в частности, это лейкозы. Далее идут рак молочной железы и щитовидной железы (рис. 12.1).
12.2.2. Популяционные волны
Численность населения планеты за обозримый исторический период в целом возросла. В эпоху неолита (10—6 тыс. лет назад) число людей было равно примерно 5 млн., в период появления городов (4,5—3,5 тыс. лет назад) — 20—40 млн., во времена Римской империи к началу новой эры — 200 млн., к 1600 г. - 500 млн., в 1800 г. -1 млрд., в настоящее время — более 6 млрд. Представляя общую тенденцию в истории человечества, эти данные указывают также на изменение во времени темпов прироста народонаселения. Этот показатель, однако, изменялся в историческом развитии неравномерно.
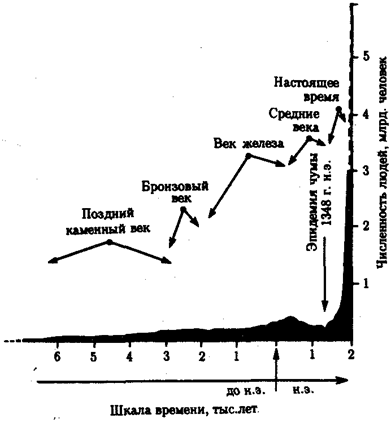
Рис. 12.2. Сокращение численности людей в средние века
в связи с эпидемией чумы
Моменты ускорения прироста численности людей совпадают с важнейшими достижениями человечества — развитием земледелия примерно 8000 лет назад, началом индустриализации, эрой научно-технической революции. Важным следствием увеличения темпа прироста является изменение плотности населения. Так, в эпоху охотников и собирателей (30—20 тыс. лет назад) она составляла менее 1 человека на 3 км2
, в век бронзы и железа (4—3 тыс. лет назад) — 10 человек, в настоящее время — от 4 до 900—1200 на 3 км2
. Даже сейчас 50% населения размещаются всего на 5% площади обитаемой суши. Крайне неравномерное распределение людей на Земном шаре имело место на любом этапе развития человечества. Ускорение роста численности при ограниченности заселяемой территории способствует усилению миграций.
На фоне общей тенденции к повышению численности людей имели место отдельные снижения этого показателя (рис. 12.2). Причиной снижения, отраженного на диаграмме динамики населения планеты, явилась эпидемия чумы с большой смертностью, которая в средние века распространилась на значительные территории. Предполагают, что сокращение численности людей на территории Европы происходило также в начале каменного века. Причина состояла в уничтожении племенами, освоившими технику коллективной охоты, основного источника пищи — крупных травоядных животных. Заметные колебания численности на ограниченных территориях происходили вследствие, например, особо опасных инфекций. Они были закономерными явлениями в жизни целых народов еще в начале текущего столетия (рис. 12.3).
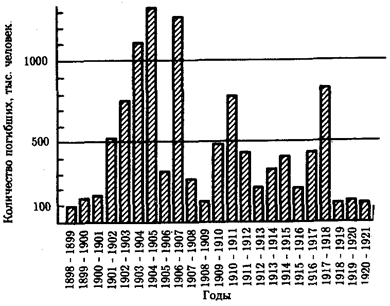
Рис. 12.3. Смертность от чумы в Индии по годам. (1898—1922)
Периодические колебания численности людей на обширных или ограниченных территориях, изменяя плотность населения и вызывая миграции, влияли на состояние генофондов человеческих популяций.
12.2.3. Изоляция
Человеческое общество длительно развивалось как совокупность изолированных производственных коллективов, внутри которых в основном и совершались браки. Природа изоляционных барьеров между популяциями людей разнообразна. В ранней истории человечества важное место принадлежало, по-видимому, географической изоляции. Специфическими для человеческого общества являются формы изоляции, зависящие от разнообразия культур, экономических укладов, религиозных и морально-этических установок.
Фактор изоляции оказывал влияние на генофонды популяций людей. Длительным проживанием в состоянии относительной культурной и географической изоляции объясняют, например, некоторые антропологические особенности представителей малых народностей: своеобразный рельеф ушной раковины бушменов, большая ширина нижнечелюстного диаметра коряков и ительменов, исключительное развитие бороды айнов. Среди горных таджиков, проживающих в одном районе, выделены группы с разным соотношением индивидуумов по антигенам эритроцитов системы АВО. Причиной различий является изолированность от главных перевальных путей сообщения.
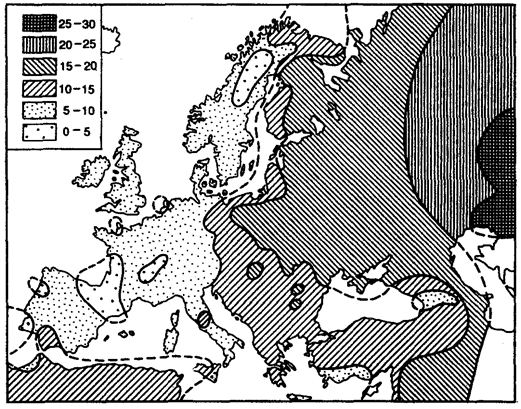
Рис. 12.4. Градиент распределения аллеля В системы группы АВО в Европе
Сохранению высокого уровня генетической изоляции двух популяций, существующих на одной территории, способствуют отличия по физическим признакам или образу жизни. Однако такие барьеры со временем ослабевают. Об этом свидетельствует судьба популяций белых и негров в США и Бразилии. К настоящему времени доля генов от белых составляет у американских негров 25%, а у бразильских — 40%. Между двумя генетически различающимися популяциями, разделенными географически, иногда вклиниваются другие популяции, через которые и происходит обмен генами. В таких случаях наблюдается градиент признака. Так, частота аллеля группы крови В в Европе постепенно повышается с запада на восток (рис. 12.4). У коренного населения Пиренейского п-ова этот ген практически отсутствует, тогда как в районе Астрахани его частота достигает 30%.
В настоящее время круг возможных браков неуклонно расширяется. Разрушение многовековых изоляционных барьеров — процесс, по-видимому, необратимый.
12.2.4. Генетико-автоматические процессы
Предположительно человеческие популяции в палеолите состояли из нескольких сотен индивидуумов. Всего одно-два столетия тому назад люди жили преимущественно поселениями в 25—35 домов. Вплоть до самого последнего времени количество индивидуумов в отдельных популяциях, непосредственно участвующих в размножении, редко превышало 400—3500 человек. Причины географического, экономического, расового, религиозного, культурного порядка ограничивали брачные связи масштабами определенного района, племени, поселения, секты. Высокая степень репродуктивной изоляции малочисленных человеческих популяций на протяжении многих поколений создавала благоприятные условия для дрейфа генов.
Генетико-автоматические процессы, или дрейф генов, приводят к сглаживанию изменчивости внутри группы и появлению случайных, не связанных с отбором различий между изолятами. Именно это выявили наблюдения за особенностями фенотипов малочисленных групп населения в условиях, например, географической изоляции. Так, среди жителей Памира резус-отрицательные индивидуумы встречаются в 2—3 раза реже, чем в Европе. В большинстве кишлаков такие люди составляют 3—5% популяции. В некоторых изолированных селениях, однако, их насчитывается до 15%, т.е. примерно как в европейской популяции.
В крови человека имеются гаптоглобины, которые связывают свободный гемоглобин после разрушения эритроцитов, чем предотвращают его выведение из организма. Синтез гаптоглобина Нр1-1 контролируется геном Нр1
. Частота этого гена у представителей двух соседних племен на Севере Южной Америки составляет 0,205 и 0,895, отличаясь более чем в 4 раза.
Примером действия дрейфа генов в человеческих популяциях служит эффект родоначальника.
Он возникает, когда несколько семей порывают с родительской популяцией и создают новую на другой территории. Такая популяция обычно поддерживает высокий уровень брачной изоляции. Это способствует случайному закреплению в ее генофонде одних аллелей и утрате других. В результате частота очень редкого аллеля может стать значительной.
Так, члены секты амишей в округе Ланкастер штата Пенсильвания, насчитывающей к середине девятнадцатого века примерно 8000 человек, почти все произошли от трех супружеских пар, иммигрировавших в Америку в 1770 г. В этом изоляте обнаружено 55 случаев особой формы карликовости с многопалостью, которая наследуется по аутосомно-рецессивному типу. Эта аномалия не зарегистирирована среди амишей штатов Огайо и Индиана. В мировой медицинской литературе описано едва ли 50 таких случаев. Очевидно, среди членов первых трех семей, основавших популяцию, находился носитель соответствующего рецессивного мутантного аллеля — «родоначальник» соответствующего фенотипа.
В XVIII в. из Германии в США иммигрировало 27 семей, основавших в штате Пенсильвания секту дункеров. За 200-летний период существования в условиях сильной брачной изоляции генофонд популяции дункеров изменился в сравнении с генофондом населения Рейнской области Германии, из которой они произошли. При этом степень различий во времени увеличивалась. У лиц в возрасте 55 лет и выше частоты аллелей системы групп крови MN ближе к цифрам, типичным для населения Рейнской области, чем у лиц в возрасте 28—55 лет. В возрастной группе 3—27 лет сдвиг достигает еще больших значений (табл. 12.1).
Таблица 12.1. Прогрессивное изменение концентрации аллелей системы групп крови MN в популяции дункеров
| Возраст членов изолята, лет
|
Концентрация аллеля
LM
|
LN
|
Более 55
От 28 до 55
От 3 до 27
|
0,55
0,66
0,735
|
0,45
0,34
0,265
|
Рост среди дункеров лиц с группой крови М и снижение — с группой крови N нельзя объяснить действием отбора, так как направление изменений не совпадает с таковым в целом для населения штата Пенсильвания. В пользу дрейфа генов говорит также то, что в генофонде американских дункеров увеличилась концентрация аллелей, контролирующих развитие заведомо биологически нейтральных признаков, например оволосения средней фаланги пальцев, способности отставлять большой палец кисти (рис. 12.5).

Рис. 12.5. Распространение нейтральных признаков
в изоляте дункеров штата Пенсильвания:
а—
рост волос на средней фаланге пальцев кисти, б—
способность отставлять большой палец кисти
На протяжении большей части истории человечества дрейф генов оказывал влияние на генофонды популяций людей. Так, многие особенности узкоместных типов в пределах арктической, байкальской, центрально-азиатской, уральской групп населения Сибири являются, по-видимому, результатом генетико-автоматических процессов в условиях изоляции малочисленных коллективов. Эти процессы, однако, не имели решающего значения в эволюции человека.
Последствия дрейфа генов, представляющие интерес для медицины, заключаются в неравномерном распределении по группам населения Земного шара некоторых наследственных заболеваний. Так, изоляцией и дрейфом генов объясняется, по-видимому, относительно высокая частота церебромакулярной дегенерации в Квебеке и Ньюфаундленде, детского цестиноза во Франции, алкаптонурии в Чехии, одного из типов порфирии среди европеоидного населения в Южной Америке, адреногенитального синдрома у эскимосов. Эти же факторы могли быть причиной низкой частоты фенилкетонурии у финнов и евреев-ашкенази.
Изменение генетического состава популяции вследствие генетико-автоматических процессов приводит к гомозиготизации индивидуумов. При этом чаще фенотипические последствия оказываются неблагоприятными. Вместе с тем следует помнить, что возможно образование и благоприятных комбинаций аллелей. В качестве примера рассмотрим родословные Тутанхамона (рис. 12.6) и Клеопатры VII (рис. 12.7), в которых близкородственные браки были правилом на протяжении многих поколений.
Тутанхамон умер в возрасте 18 лет. Анализ его изображения в детском возрасте и подписи к этому изображению позволяют предположить, что он страдал генетическим заболеванием — целиакией, которая проявляется в изменении слизистой оболочки кишечника, исключающем всасывание клейковины. Тутанхамон родился от брака Аменофиса III и Синтамоне, которая была дочерью Аменофиса III. Таким образом, мать фараона была его сводной сестрой. В могильном склепе Тутанхамона обнаружены мумии двух, по всей видимости мертворожденных, детей от брака с Анкесенамон, его племянницей. Первая жена фараона была или его сестрой, или дочерью. Брат Тутанхамона Аменофис IV предположительно страдал болезнью Фрелиха и умер в 25—26 лет. Его дети от браков с Нефертити и Анкесенамон (его дочерью) были бесплодны. С другой стороны, известная своим умом и красотой Клеопатра VII была рождена в браке сына Птоломея Х и его родной сестры, которому предшествовали кровнородственные браки на протяжении по крайней мере шести поколений.
Рис. 12.6. Родословная фараона XVIII династии Тутанхамона
Рис. 12.7. Родословная Клеопатры VII
12.2.5. Естественный отбор
В процессе видообразования естественный отбор
переводит случайную индивидуальную изменчивость в биологически полезную групповую — популяционную, видовую. Стабилизирующая его форма сохраняет «удачные» комбинации аллелей от предшествующих этапов эволюции. Отбор поддерживает также состояние генетического полиморфизма. Смена биологических факторов исторического развития социальными привела к тому, что в человеческих популяциях отбор утратил функцию видообразования. За ним сохранились функции стабилизации генофонда и поддержания наследственного разнообразия.
В пользу действия стабилизирующей формы естественного отбора говорит, например, большая смертность среди недоношенных и переношенных новорожденных по сравнению с доношенными. Направление отбора среди таких детей зависит, по-видимому, от снижения общей жизнеспособности. Отрицательный отбор по одному локусу можно проиллюстрировать на примере системы групп крови «резус» (Rh).
Около 85% населения Европы имеет в эритроцитах антиген Rh и образует группу Rh-положительных индивидуумов. Остальные люди из европейской популяции лишены этого антигена и являются Rh-отрицательными. Синтез антигена Rh контролируется доминантным аллелем в и происходит у лиц с генотипами DD и Dd. Резус-отрицательные люди являются рецессивными гомозиготами (dd). При беременности Rh-отрицательной женщины (dd) Rh-положительным плодом (мужчина DD или Dd, плод Dd) при нарушении целостности плаценты в родах Rh-положительные эритроциты плода проникают в организм матери и иммунизируют его (рис. 12.8). При последующей беременности Rh-положительным плодом (Dd) анти-Rh-антитела проникают через плаценту в организм плода и разрушают его эритроциты. Развивается гемолитическая болезнь новорожденного. Ведущим ее симптомом является тяжелая анемия.
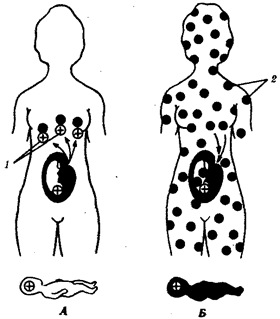
Рис. 12.8. Генетическая основа резус-конфликта. А —
первая беременность; Б —
вторая беременность:/—резус-антигены, 2—
антитела к резус-антигену
В настоящее время медицина располагает способами борьбы с этой формой патологии в виде быстрого переливания младенцу Rh-отрицательной крови или введения анти-Rh-антител для предотвращения иммунизации матери. В отсутствие медицинской помощи новорожденный с гемолитической болезнью нередко погибал. При Rh-отрица-тельном фенотипе матери Rh-положительный плод всегда гетерозиготен (Dd). Это означает, что со смертью такого индивидуума из генофонда популяции, к которой он принадлежит, удаляется равное количество доминантных и рецессивных аллелей локуса «резус». Отбор в данном случае направлен против гетерозигот. При неравенстве исходных частот удаляемых из генофонда аллелей такой отбор приводит к постепенному снижению доли более редкого из них. В европейской популяции таковым является рецессивный аллель d. Подсчитано, что снижение его доли с 15 до 1% путем отбора против гетерозигот потребует 600 поколений, или около 15 000 лет.
Под действием отбора находятся аллели, контролирующие синтез антигенов системы групп крови АВ0. Об этом свидетельствует снижение против ожидаемого количества детей в АВ0-несовместимых браках женщин 0-группы с мужчинами А, В или АВ-групп (табл. 12.2).
Таблица 12.2. Влияние АВ0-несовместимости
на плодовитость и эмбриогенез
| Показатель
|
АВ0-совместимые
|
АВ0-несовместимые
|
| Число обследованных семей |
812 |
617 |
| Выкидыши, % |
10,3 |
15,3 |
| Бездетные браки, % |
9,8 |
18,2 |
| Среднее число детей на одну мать |
2,6 |
2,2 |
Особенность отбора по локусам группоспецифичных антигенов эритроцитов, таких, как «резус» или АВО, заключается в том, что он действует на уровне зиготы или ранних стадий эмбриогенеза. Замечено, что в браках, где мать и отец несовместимы по фактору «резус», число детей с гемолитической болезнью меньше ожидаемого. Так, среди белого населения США Rh-отрицательные люди составляют примерно 16%. В такой популяции частота аллеля в равна 40%, а вероятность развития гемолитической болезни при случайном подборе брачных пар — 9%. На самом деле частота этой болезни составляет 1:150—1:200. Одним из факторов снижения является одновременная несовместимость родителей по локусу АВ0. В этом случае Rh-положительные эритроциты плода, попавшие в организм матери, разрушаются и иммунизации не происходит.
Отрицательный отбор действует в большинстве популяций людей по аллелям аномальных гемоглобинов. Его особая жесткость обусловливается тем, что он направлен против гомозигот. Ребенок, умирающий, например, от серповидно-клеточной анемии, является гомозиготным по аллелю S. Каждая такая смерть устраняет из генофонда популяции аллели одного вида. Это приводит к сравнительно быстрому снижению изменчивости по соответствующему локусу. Во многих популяциях людей частота аллелей аномальных гемоглобинов, в том числе и S, не превышает 1%.
Высокая частота аллелей таких аномальных гемоглобинов, как S, С, D, Е, в некоторых районах планеты иллюстрирует действие естественного отбора по поддержанию в человеческих популяциях состояния балансированного генетического полиморфизма. Отрицательный отбор в отношении аллеля S перекрывается мощным положительным отбором гетерозигот HbAHbS благодаря высокой жизнеспособности последних в очагах тропической малярии (рис. 12.9).
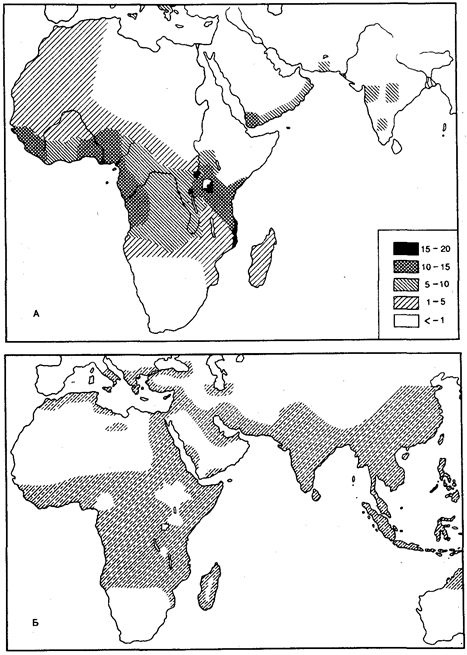
Рис. 12.9. Совпадение распространения в Старом Свете аллеля
серповидно-клеточности (А
) и тропической малярии (Б
)
Исследования в Уганде показали, что количество возбудителей в 1 мл крови зависит от генотипа ребенка и составляет до 10 000 у HbSHbS, до 160 000 у HbAHbS и до 800 000 у НbAHbА. В Северной Греции обследовали 48 семей, в которых наблюдались и серповидно-клеточность, и малярия. Среди братьев и сестер, больных серповидно-клеточной анемией, болело малярией 16 из 25 с генотипами HbAHbA и 1 из 23 с генотипами HbAHbS.
Таким образом, нормальные люди болели в 13 раз чаще, чем индивидуумы с аномальным генотипом. Устранение фактора контротбора приводит к снижению частоты аллеля серповидно-клеточности. Этой причиной, действующей на протяжении уже нескольких столетий наряду с метисацией, объясняют относительно низкую частоту гетерозигот HbAHbS среди североамериканских негров (8—9%) в сравнении с африканскими (около 20%).
В приведенных примерах действию отрицательного отбора, снижающего в генофондах некоторых популяций людей концентрацию определенных аллелей, противостоят контротборы, которые поддерживают частоту этих аллелей на достаточно высоком уровне. Результатом наложения многочисленных и разнонаправленных векторов отбора является формирование и поддерживание генофондов популяций в состоянии, обеспечивающем возникновение в каждом поколении генотипов достаточной приспособленности с учетом местных условий. Благодаря социально-экономическим преобразованиям, успехам лечебной и особенно профилактической медицины влияние отбора на генетический состав популяций людей прогрессивно снижается.
Человечеству свойствен высокий уровень наследственного разнообразия, что проявляется в многообразии фенотипов. Люди отличаются друг от друга цветом кожных покровов, глаз, волос, формой носа и ушной раковины, рисунком эпидермальных гребней на подушечках пальцев и другими сложными признаками. Выявлены многочисленные варианты отдельных белков, различающиеся по одному или нескольким аминокислотным остаткам и, следовательно, функционально. Белки являются простыми признаками и прямо отражают генетическую конституцию организма. У людей не совпадают группы крови по системам эритроцитарных антигенов «резус», АВ0, MN. Известно более 130 вариантов гемоглобина, более 70 вариантов фермента глюкозо-6-фосфатдегидрогеназы (Г6ФД), который участвует в бескислородном расщеплении глюкозы в эритроцитах. В целом не менее 30% генов, контролирующих у человека синтез ферментных и других белков, имеют несколько аллельных форм. Частота встречаемости разных аллелей одного гена варьирует.
Так, из многих вариантов гемоглобина лишь четыре обнаруживаются в некоторых популяциях в высокой концентрации: HbS (тропическая Африка, Средиземноморье), НЬС (Западная Африка), HbD(Индия), НЬЕ (Юго-Восточная Азия). Концентрация других аллелей гемоглобина повсеместно не превышает, видимо, 0,01—0,0001. Вариабельность распространенности аллелей в популяциях людей зависит от действия элементарных эволюционных факторов. Важная роль принадлежит мутационному процессу, естественному отбору, генетико-автоматическим процессам, миграциям.
Мутационный процесс создает новые аллели. И в человеческих популяциях он действует ненаправленно, случайным образом. В силу этого отбор не приводит к выраженному преобладанию концентрации одних аллелей над другими. В достаточно большой популяции, где каждая пара родителей из поколения в поколение дает двух потомков вероятность сохранения новой нейтральной мутации через 15 поколений составляет всего 1/9.
Все многообразие вариантов белков, отражающее разнообразие аллелей в генофонде человечества, можно разделить на две группы. К одной из них относятся редкие варианты, встречающиеся повсеместно с частотой менее 1%. Появление их объясняется исключительно мутационным процессом. Вторую группу составляют варианты, обнаруживаемые относительно часто в избранных популяциях. Так, в примере с гемоглобинами к первой группе относятся все варианты, кроме HbS, HbC, HbD и HbE. Длительные различия в концентрации отдельных аллелей между популяциями, сохранение в достаточно высокой концентрации нескольких аллелей в одной популяции зависят от действия естественного отбора или дрейфа генов.
К межпопуляционным различиям в концентрации определенных аллелей приводит стабилизирующая форма естественного отбора. Неслучайное распределение по планете аллелей эритроцитарных антигенов АВ0 может быть, например, обусловлено различной выживаемостью лиц, отличающихся по группе крови, в условиях частых эпидемий особо опасных инфекций. Области сравнительно низких частот аллеля I0
и относительно высоких частот аллеля IB
в Азии примерно совпадают с очагами чумы. Возбудитель этой инфекции имеет Н-подобный антиген. Это делает людей с группой крови О особенно восприимчивыми к чуме, так как они, имея антиген Н, не способны вырабатывать противочумные антитела в достаточном количестве. Указанному объяснению соответствует факт, что относительно высокие концентрации аллеля I0
обнаруживаются в популяциях аборигенов Австралии и Полинезии, индейцев Америки, которые практически не поражались чумой.
Частота заболеваемости «натуральной» оспой, тяжесть симптомов, смертность выше у лиц с группой крови А или АВ в сравнении с лицами, имеющими группу крови 0 или В (рис. 12.10). Объяснение состоит в том, что у людей первых двух групп отсутствуют антитела, частично нейтрализующие оспенный антиген А. Лица с группой крови 0 в среднем имеют возможность прожить дольше, однако для них выше вероятность заболеть язвенной болезнью. В таблице 12.3 указаны некоторые аллели и генотипы, имеющие приспособительное значение в отдельных географических и экологических регионах.
Рис. 12.10. Относительная частота и показатели тяжести заболевания «натуральной» оспой у лиц с разными группами крови АВО
Таблица 12.3. Примеры аллелей, имеющих адаптивное значение
| Аллели и генотипы
|
Географическая распространенность
|
Адаптивное значение
|
Группы крови системы АВ0, аллель В
Аллель А
Трансферрины — белки, связывающие железо,
аллель TfDI
Кислая фосфатаза эритроцитов, аллель Acpr
Аллель АсР и группа
крови АВ
Аллель АсР и группа
крови А или аллель АсР и
группа крови АВ
Сухая ушная сера, аллель d
|
Повсеместно, чаще в Азии
Повсеместно
Высокая частота в зоне тропического пояса
Высокая частота у бушменов и негроидов Центральной Африки
Высокая частота на о.Новая Гвинея
Высокая частота у жителей Чукотки и Аляски
Высокая частота в Дальневосточном регионе
|
Относительная устойчивость к чуме
Относительная устойчивость к язве желудка и двенадцатиперстной кишки
Резистентность ко многим инфекционным заболеваниям
Высокая активность
фермента при повышениях
температуры
Устойчивость к тропи
ческой малярии
Легкость адаптации в условиях холодного климата
При генотипе dd низки
уровень холестерина и
концентрация липидов в
крови, высокая концентрация лизоцима в ушной сере
|
Вместе с тем для популяций из одного географического района, но изолированных в репродуктивном отношении, причиной различий в концентрации аллелей АВО мог быть дрейф генов. Так, частота группы крови А достигает у индейцев племени черноногих 80%, а у индейцев из штата Юта — 2%.
В основе стойкого сохранения в популяции людей одновременно нескольких аллелей одного гена лежит, как правило, отбор в пользу гетерозигот, который ведет к состоянию балансированного полиморфизма. Классическим примером такой ситуации является распространение аллелей гемоглобинов S, С, и Е в очагах тропической малярии.
Выше приведены примеры полиморфизма по конкретным локусам, который объясняется действием известного фактора отбора. В естественных условиях в силу воздействия на фенотипы организмов комплекса факторов отбор осуществляется по многим направлениям. В результате формируются генофонды, сбалансированные по набору и частотам аллелей, обеспечивающие в данных условиях достаточную выживаемость популяций. Это справедливо и для популяций людей. Так, люди с группой крови 0 более восприимчивы к чуме, чем люди с группой В. Туберкулез легких у них лечится с большим трудом, чем у лиц с группой крови А. Вместе с тем лечение больных сифилисом людей с группой крови 0 быстрее вызывает переход болезни в неактивную стадию. Для лиц с группой крови 0 вероятность заболеть раком желудка, раком шейки матки, ревматизмом, ишемической болезнью сердца, холециститом, жёлчно-каменной болезнью примерно на 20% ниже, чем для лиц с группой А.
Генетический полиморфизм по многим локусам мог быть унаследован людьми от предков на досапиентной стадии развития. Полиморфизм по таким системам групп крови, как АВ0 и Rh, обнаружен у человекообразных обезьян. Факторы отбора, действие которых создавало современную картину распределения аллелей в популяции людей, для подавляющего большинства локусов точно не установлены. Примеры, рассмотренные выше, указывают на их экологическую природу.
Учитывая слабую техническую оснащенность, плохие экономические и гигиенические условия жизни основной массы населения планеты на протяжении значительной части истории человечества, можно представить, какую большую роль играли возбудители особо опасных инфекций, паразитарных заболеваний, туберкулеза. В этих условиях наследственный полиморфизм способствовал расселению людей, обусловливая удовлетворительную жизнеспособность в разных экологических ситуациях. Определенный вклад в наблюдаемое распределение аллелей внесли массовые миграции населения и сопутствующая им метизация. В период до Великих географических открытий и начала колониальной экспансии смешение больших контингентов людей разной расовой принадлежности имело место в Восточной Африке, Индии, Средней Азии, Западной Сибири, Алтае-Саянском нагорье, Индокитае. Впоследствии это наблюдалось в Южной и Центральной Америке.
Генетический полиморфизм является основой межпопуляционной и внутрипопуляционной изменчивости людей. Изменчивость проявляется в неравномерном распределении по планете некоторых заболеваний, тяжести их протекания в разных человеческих популяциях, разной степени предрасположенности людей к определенным болезням, индивидуальных особенностях развития патологических процессов, различиях в реакции на лечебное воздействие. Наследственное разнообразие долго было препятствием успешному переливанию крови. В настоящее время оно же создает большие трудности в решении проблемы пересадок тканей и органов.
Так же как и в популяциях других организмов, наследственное разнообразие снижает реальную приспособленность популяций людей. Бремя генетического груза человечества можно оценить, введя понятие летальных эквивалентов.
Считают, что число их в пересчете на гамету колеблется от 1,5 до 2,5 или от 3 до 5 на зиготу. Это означает, что то количество неблагоприятных аллелей, которое имеется в генотипе каждого человека, по своему суммарному вредному действию эквивалентно действию 3—5 рецессивных аллелей, приводящих в гомозиготном состоянии к смерти индивидуума до наступления репродуктивного возраста.
При наличии неблагоприятных аллелей и их сочетаний примерно половина зигот, образующихся в каждом поколении людей, в биологическом плане несостоятельна. Такие зиготы не участвуют в передаче генов следующему поколению. Около 15% зачатых организмов гибнет до рождения, 3 — при рождении, 2 — непосредственно после рождения, 3 — умирает, не достигнув половой зрелости, 20 — не вступают в брак, 10% браков бездетны.
Неблагоприятные последствия генетического груза в виде рецессивных аллелей, если они не приводят к гибели организма, проявляются в снижении ряда важных показателей состояния индивидуума, в частности его умственных способностей. Исследования, проведенные на популяции арабов в Израиле, для которой характерна высокая частота близкородственных браков (34% между двоюродными и 4% между дважды двоюродными сибсами), показали снижение умственных способностей у детей от таких браков.
Исторические перспективы человека в силу его социальной сущности не связаны с генетической информацией, накопленной видом Homo
sapiens
в ходе эволюции. Тем не менее человечество продолжает «оплачивать» эти перспективы, теряя в каждом поколении часть своих членов из-за их генетической несостоятельности.
Действие элементарных эволюционных факторов приводит к дивергенции популяций и образованию новых видов. Многообразные виды связаны друг с другом разной степенью родства. Виды, дивергировавшие от общих предков относительно недавно, сохраняют нередко значительное фено- и генотипическое сходство. Дивергенция, произошедшая в отдаленное время, в результате оставляет меньшее количество общих фенотипических признаков и особенностей геномов. Виды, практически не связанные родством, характеризуются серьезными различиями.
Благодаря тому что процесс микроэволюции, завершающийся видообразованием, протекает постепенно на протяжении всего существования жизни на Земле, в живой природе образовались естественные группировки видов разного ранга родства, называемые таксонами.
Тип — это наиболее крупный таксой, объединяющий виды, которые имеют сходный общий план строения благодаря отдаленному родству. Типы подразделяются на классы, далее следуют отряды, семейства и роды. Дивергенция видов, входящих в состав одного рода, произошла относительно недавно (рис. 13.1).
Рис. 13.1. Эволюция таксономических групп
Процесс формирования таксонов надвидового ранга называют макроэволюцией.
Процесс макроэволюции изучают методами палеонтологии, сравнительной анатомии и сравнительной эмбриологии. Современные методы изучения макроэволюции — иммунологический и молекулярно-биологический — подробнее описаны в § 15.2.
13.1.1. Уровень организации
Говоря о прогрессивной эволюции природы в целом и животного мира в частности, следует определить понятие уровень организации.
Это понятие прямо не связано со степенью сложности строения животного.
Действительно, степень сложности не только морфофизиологической организации, но и генома человека и современных человекообразных обезьян практически совпадает, но уровень организации человека несомненно выше.
Предложено несколько вариантов оценки уровня организации животных. Наиболее приемлемым из них является, вероятно, определение уровня организации через характер взаимоотношения организмов данной группы со средой обитания.
Примитивные организмы проявляют себя в среде пассивно. Выживание и дальнейшая эволюция их обеспечиваются в первую очередь интенсивным процессом размножения, благодаря которому компенсируются огромные потери, возникающие в процессе естественного отбора. В противоположность им высокоорганизованные организмы более активно проявляют себя в среде обитания и не только противостоят ей, но и способны преобразовывать ее, приспосабливая для своих нужд. Плодовитость их невелика.
Рассмотренный критерий уровня организации может применяться только по отношению к группам организмов, связанных более или менее отдаленным родством. Так, пресмыкающиеся более высоко организованы, чем земноводные, потому, что они активнее ведут себя •в среде обитания (размножение их может происходить в разнообразных условиях), при более низкой плодовитости выживаемость их потомства значительно выше. Уровень организации млекопитающих еще более высок благодаря постоянной температуре тела, сложному поведению, внутриутробному эмбриональному развитию и заботе о потомстве. Уровень организации человека чрезвычайно высок благодаря его социальному способу адаптации к среде и активному ее преобразованию (см. гл. 17).
13.1.2. Типы эволюции групп
В зависимости от того, изменяется ли уровень организации в эволюционирующих группах, выделяют два основных типа эволюции: аллогенез и арогенез.
При аллогенезе
у всех представителей данной группы сохраняются без изменения основные черты строения и функционирования систем органов, благодаря чему уровень организации их остается прежним. Аллогенная эволюция происходит в пределах одной адаптивной зоны —
совокупности экологических ниш, различающихся в деталях, но сходных по общему направлению действия основных средовых факторов на организм данного типа. Интенсивное заселение конкретной адаптивной зоны достигается благодаря возникновению у организмов идиоадаптаций —
локальных морфофизиологических приспособлений к определенным условиям существования. Пример аллогенеза с приобретением идиоадаптаций к разнообразным условиям обитания в отряде насекомоядных млекопитающих см. на рис. 13.2.

Рис. 13.2. Аллогенез в отряде насекомоядных млекопитающих
Наземные формы: а—прыгунчик, б—землеройка, в—еж; земноводные формы: г—кутора, д—
выдровая землеройка, е—
выхухоль; роющие формы: ж—
крот, з—златокрот
Арогенез
— такое направление эволюции, при котором у некоторых групп внутри более крупного таксона появляются новые морфофизиологические особенности, приводящие к повышению уровня их организации. Эти новые прогрессивные черты организации называют ароморфозами.
Ароморфозы позволяют организмам заселять принципиально новые, более сложные адаптивные зоны. Так, арогенез ранних земноводных был обеспечен появлением у них таких основных ароморфозов, как пятипалые конечности наземного типа, легкие и два круга кровообращения с трехкамерным сердцем. Завоевание адаптивной зоны с более сложными для жизни условиями (наземной по сравнению с водной, воздушной по сравнению с наземной) сопровождается активным расселением в ней организмов с появлением у них локальных идиоадаптаций к различным экологическим нишам.
Таким образом, периоды арогенной эволюции группы могут сменяться периодами аллогенеза, когда в результате возникающих идиоадаптаций новая адаптивная зона заселяется и используется наиболее эффективно. Если в ходе филогенеза организмы осваивают более 49
простую по сравнению с исходной адаптивную зону, например, переходя к прикрепленному неподвижному образу жизни или становись паразитами, то они претерпевают морфофизиологический регресс, или дегенерацию, утрачивая часть прогрессивных черт, которыми обладали их предки, и оказываясь на более низком уровне организации. Нередко и новая, более простая, адаптивная зона дает возможность широких идиоадаптаций к не менее разнообразным условиям обитания в ней, т.е. обеспечивает аллогенез (рис. 13.3).
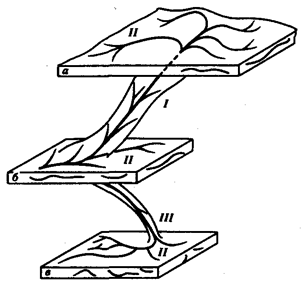
Рис. 13.3. Направления эволюции групп:
I
— арогенез,II
— аллогенез,III
— дегенерация; а, б, в —
адаптивные зоны
Современный органический мир характеризуется огромным многообразием форм, отличающихся как по уровню организации, так и по разнообразию локальных адаптации именно благодаря смене типов эволюции, протекающей своеобразно в каждой из эволюционирующих групп. Наряду с высокоорганизованными формами существуют примитивные, свободноживущие и паразитические организмы, возникшие эволюционно относительно недавно, и реликтовые группы, время бурного адаптационного процесса которых давно прошло.
13.1.3. Формы эволюции групп
Существуют две элементарные формы филогенеза: филетическая
и дивергентная
эволюция.
Филетическая эволюция
— это изменения, происходящие в одном филогенетическом стволе, эволюционирующем во времени как единое целое. Реконструированы филогенетические ряды слона, лошади и других организмов, в которых можно наблюдать постепенное нарастание количества и степени выраженности признаков, характерных для современных форм (рис. 13.4).
Рис. 13.4. Филетическая эволюция конечности в сем. Лошадиные.
I
—преобразование скелета конечности; II
—схема филетической эволюции:
предковая форма—фенакодус, 1
— эогиппус, 2—
миогиппус, 3—
парагиппус, 4—плиогиппус, 5—
лошадь; Ап—А5
— этапы эволюции
Дивергентная эволюция
заключается в образовании на основе одной предковой группы двух или нескольких производных (рис. 13.5). Она приводит к дифференциации более крупных таксонов на более мелкие, например классов на отряды, родов на виды.
Как филетическая, так и дивергентная эволюция протекают на общей генетической базе, поэтому между организмами сохраняется более или менее выраженное генотипическое и морфофункциональное сходство.
Сопоставление филогенеза в разных группах позволяет выделить и некоторые общие закономерности соотносительной эволюции. Так, при попадании в одну и ту же среду обитания двух или более филогенетических групп неродственных организмов у них обычно проявляется конвергенция
признаков. При этом сходные экологические задачи они решают сходным образом. Конвергентные адаптации возникают в этом случае на разной генетической основе, затрагивают в первую очередь поверхностные признаки, не распространяясь на общий план строения и наиболее существенные черты организации соответствующих групп. Примером конвергентной эволюции являются форма тела и особенности локомоции в воде у акуловых рыб, водных пресмыкающихся — ихтиозавров, костистых рыб, пингвинов, ластоногих и китообразных млекопитающих, внутреннее строение которых полностью соответствует особенностям, характерным для классов, к которым они относятся (рис. 13.6).
Другая форма соотносительной эволюции — параллелизм —
реализуется в двух или нескольких группах, связанных более или менее отдаленным родством, которое основано на дивергенции от общего предка. В связи с общностью части генофондов, унаследованных от предков, у них возникают сходные адаптации в условиях действия факторов отбора в одинаковом направлении. Параллельное филетическое развитие двух родственных групп обеспечивается реализацией закона гомологических рядов (см. разд. 13.3.5). Примером параллелизма является эволюция одногорбого и двугорбого верблюдов соответственно в Африке и Центральной Азии, имеющих сходные адаптации к жизни в пустынях
Параллелизм можно наблюдать и в родственных группах организмов, разобщенных временем. Так, во второй половине кайнозойской эры в семействе кошачьих такой признак, как саблезубость (гипертрофия клыков верхней челюсти), возникал по крайней мере четырежды у представителей четырех разных родов (рис. 13.7).
13.1.4. Биологический прогресс и биологический регресс
Эволюция любого типа — арогенная, аллогенная или идущая по типу мофофизиологического регресса в природе — в целом приводит к всесветному расселению живых организмов разного уровня организации.
Рассматривая эволюцию отдельных таксонов, можно убедиться в том, что некоторые из них находятся в состоянии расцвета, в то время как другие вымирают. Успех группы организмов в эволюционном процессе оценивают как состояние биологического прогресса,
критериями которого являются: 1) увеличение количества представителей соответствующей группы; 2) расширение ареала распространения;
3) активизация видоообразования в роде, увеличение количества родов в семействе, семейств в отряде и т.д.
Явление, противоположное биологическому прогрессу, — биологический регресс —
свидетельствует об угасании филогенетической группы, ведущем к ее вымиранию. Так же как и биологический прогресс, состояние регресса может длиться очень долго. Оставшиеся немногочисленные представители некогда биологически прогрессивной группы, найдя соответствующую их организации экологическую нишу, могут продолжать существовать на протяжении сотен миллионов лет. Они называются реликтами,
обладают консервативностью организации и крайне медленно эволюционируют. Примерами реликтов являются современная кистеперая рыба — латимерия, ближайшие родственники которой жили в девонском периоде более 200 млн. лет назад, а также дерево гинкго, непосредственные предки которого существовали на Земле уже в первой половине мезозойской эры.

Рис. 13.6. Конвергентная эволюция формы тела позвоночных. I
—водные животные разного происхождения; II
—схема конвергентной эволюции:
1—
акула, 2—ихтиозавр, 3—
дельфин, 4—
костистая рыба, 5—
пингвин, 6—
тюлень; A—A1
, B—B1
— этапы эволюции
На протяжении исторического развития одна и та же группа может претерпевать периоды бурного расцвета, стабильного состояния, или стагнации, и упадка. Так, в девонском периоде появились первые земноводные, вставшие на путь арогенной эволюции. Заселив наземную среду обитания, они господствовали на ней в течение 75 млн. лет. На границе палеозойской и мезозойской эр (230 млн. лет назад) бурный биологический прогресс претерпевают пресмыкающиеся, быстро вытесняющие земноводных, которые с тех времен и до современного состояния переживают биологический регресс.
Расцвет пресмыкающихся длился до последней трети мезозойской эры, когда (около 65 млн. лет назад) он сменился биологическим регрессом, после чего освобождающиеся вымирающими динозаврами экологические ниши стали занимать бурно эволюционирующие млекопитающие и птицы, биологический прогресс которых продолжается до настоящего времени. В то же время такой класс, как костистые рыбы, пройдя период бурного расцвета к началу мезозойской эры и прочно заняв разнообразные места обитания, до настоящего времени находится в состоянии стабильного господства среди позвоночных в водной среде обитания.
Состояние биологического прогресса может быть обусловлено как аро- и аллогенезом, так и морфофизиологическим регрессом. Действительно, занятие новых экологических ниш как в новых адаптивных зонах, так и в уже освоенных приводит и к активизиции размножения, и к расширению ареала, и к усилению видообразования.
Рис. 13.7. Параллельная эволюция саблезубости у кошачьих. I
—
черепа саблезубых кошек в разные периоды кайнозойской эры; II—
схемы параллельной эволюции:
а
—
махайрод (олигоцен - 37-25 млн. лет назад), б
—
смилодон (миоцен - 25-9 млн. лет назад) в
—
лжесаблезубая кошка (олигоцен - 37-25 млн. лет назад), г—
саблезубый тигр (плейстоцен —
1,8 млн.-10 тыс. лет назад); А-А2
—
этапы синхронного параллелизма, B-B2
- этапы асинхронного
13.1.5. Эмпирические правила эволюции групп
Из многочисленных обнаруженных эмпирическим путем правил и закономерностей эволюции групп остановимся на важнейших.
Согласно правилу необратимости эволюции,
эволюция является необратимым процессом, и организмы не могут вернуться к прежнему состоянию, уже пройденному их предками ранее. Действительно, каждое эволюционно значимое изменение представляет собой комбинацию многих мутаций и появление новых регуляторных взаимодействий, подхваченных естественным отбором, но возникших случайно и независимо друг от друга. Поэтому понятно, что возвращение генофонда данной группы организмов или даже одного генотипа к состоянию, характерному для предков, статистически практически невероятно. Однако в ряде случаев за счет отдельных обратных мутаций или за счет мутаций регуляторных генов возможно повторное возникновение отдельных признаков атавистической природы (разд. 13.3.4).
Правило прогрессивной специализации
утверждает, что филогенетическая группа, эволюционирующая по пути приспособления к данным конкретным условиям, и в дальнейшем будет продвигаться по пути углубления специализации. Генетические основы этого правила заключены в том, что в г«роцессе естественного отбора в условиях данной адаптивной зоны отсеиваются те гены генофондов популяций, которые не соответствуют ей. В результате возникает ограниченность способности генофондов к изменениям в разных направлениях.
Примером прогрессивной специализации служат морфологические преобразования конечностей в эволюционной ветви лошадей (см. рис. 13.4). При переходе к жизни на открытых пространствах с плотной почвой у предков лошади уменьшается количество пальцев до одного, что не позволяет современным лошадям населять другие биотопы. Другой пример — эволюция Юго-Восточно-азиатских человекообразных обезьян гиббонов. Специализация их к древесному образу жизни при отсутствии хватательного хвоста, имеющегося, например, у всех американских древесных обезьян, привела к возникновению своеобразного типа движения — брахиации, при котором осуществляются прыжки по веткам в подвешенном состоянии на передних конечностях. Они при этом резко удлиняются, большой палец значительно редуцируется, а рука становится почти неспособной к манипуляциям мелкими предметами. При передвижении по земле руки гиббонов уже не участвуют в локомоции.
Прогрессивная специализация резко уменьшает экологическую пластичность видов и часто является причиной их вымирания при изменившихся условиях. Уменьшение размеров и упрощение видового состава тропических лесов Индонезии являются причиной биологического регресса такого узкоспециализированного вида человекообразных обезьян, как орангутан.
Следствием правила прогрессивной специализации является правило происхождения новых групп организмов от малоспециализированных предков.
Примеров, иллюстрирующих это правило, много. Действительно, млекопитающие произошли от пресмыкающихся, сохранивших в своей организации некоторые черты земноводных. Прогрессивная линия эволюции приматов, ведущая к человеку, не связана с рамапитеком (специализированной древесной формой), как полагали раньше, а берет начало от неспециализированных африканских приматов, ведущих полуназемный образ жизни (см. § 15.2 и 15.3). Генетической основой этого правила является отсутствие жесткого одностороннего отбора генотипов и сохранение в связи с этим их большего разнообразия у неспециализированных форм и, следовательно, их высокой экологической пластичности.
13.2.1. Закон зародышевого сходства
Исследователи начала XIX в. впервые стали обращать внимание на сходство стадий развития эмбрионов высших животных со ступенями усложнения организации, ведущими от низкоорганизованных форм к прогрессивным. Сопоставляя стадии развития зародышей разных видов и классов хордовых, К. Бэр сделал следующие выводы.
1. Эмбрионы животных одного типа на ранних стадиях развития сходны.
2. Они последовательно переходят в своем развитии от более общих признаков типа ко все более частным. В последнюю очередь развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, индивидуальные черты.
3. Эмбрионы разных представителей одного типа постепенно обособляются друг от друга (рис. 13.8).
К. Бэр, не будучи эволюционистом, не мог связывать открытые им закономерности индивидуального развития с процессом филогенеза. Поэтому сделанные им обобщения имели значение не более чем эмпирических правил.
Развитие эволюционной идеи в последующем позволило объяснить сходство ранних зародышей их историческим родством, а приобретение ими все более частных черт с постепенным обособлением друг от друга — действительным обособлением соответствующих классов, отрядов, семейств, родов и видов в процессе эволюции.
Вскоре после открытия закона зародышевого сходства Ч. Дарвин показал, что этот закон свидетельствует об общности происхождения и единства начальных этапов эволюции в пределах типа.
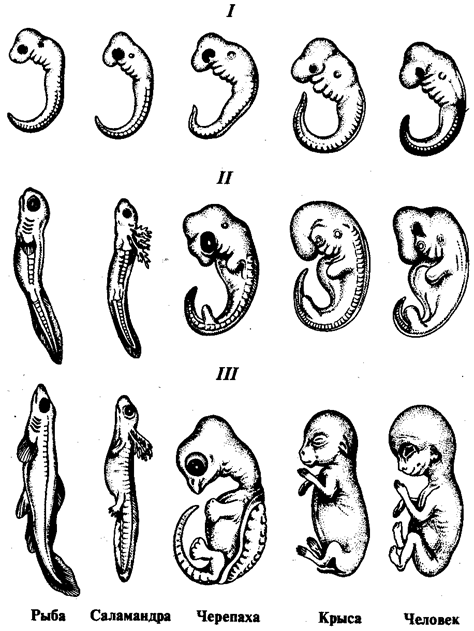
Рис. 13.8. Сходство зародышей разных классов позвоночных на разных этапах (I
—
Ill
)онтогенеза
13.2.2. Онтогенез — повторение филогенеза
Сопоставляя онтогенез ракообразных с морфологией их вымерших предков, Ф. Мюллер сделал вывод о том, что ныне живущие ракообразные в своем развитии повторяют путь, пройденный их предками. Преобразование онтогенеза в эволюции, по мнению Ф. Мюллера, осуществляется благодаря его удлинению за счет добавления к нему дополнительных стадий или надставок. На основе этих наблюдений, а также изучения развития хордовых Э. Геккель (1866) сформулировал основной биогенетический закон, в соответствии с которым онтогенез представляет собой краткое и быстрое повторение филогенеза.
Повторение структур, характерных для предков, в эмбриогенезе потомков названо рекапитуляциями.
Рекапитулируют не только морфологические признаки — хорда, закладки жаберных щелей и жаберных дуг у всех хордовых, но и особенности биохимической организации и физиологии. Так, в эволюции позвоночных происходит постепенная утрата ферментов, необходимых для распада мочевой кислоты — продукта метаболизма пуринов. У большинства беспозвоночных конечный продукт распада мочевой кислоты — аммиак, у земноводных и рыб — мочевина, у многих пресмыкающихся — аллантоин, а у некоторых млекопитающих мочевая кислота вообще не расщепляется и выделяется с мочой. В эмбриогенезе млекопитающих и человека отмечены биохимические и физиологические рекапитуляции: выделение ранними зародышами аммиака, позже мочевины, затем аллантоина, а на последних стадиях развития — мочевой кислоты.
Однако в онтогенезе высокоорганизованных организмов не всегда наблюдается строгое повторение стадий исторического развития, как это следует из биогенетического закона. Так, зародыш человека никогда не повторяет взрослых стадий рыб, земноводных, пресмыкающихся и млекопитающих, а сходен по ряду черт лишь с их зародышами. Ранние стадии развития сохраняют наибольшую консервативность, благодаря чему рекапитулируют более полно, чем поздние. Это связано с тем, что одним из наиболее важных механизмов интеграции ранних этапов эмбриогенеза является эмбриональная индукция, а структуры зародыша, формирующиеся в первую очередь, такие, как хорда, нервная трубка, глотка, кишка и сомиты, представляют собой организационные центры зародыша, от которых зависит весь ход развития.
Генетическая основа рекапитуляции заключена в единстве механизмов генетического контроля развития, сохраняющемся на базе общих генов регуляции онтогенеза, которые достаются родственным группам организмов от общих предков.
13.2.3. Онтогенез — основа филогенеза
Опираясь только на основной биогенетический закон, невозможно объяснить процесс эволюции: бесконечное повторение пройденного само по себе не рождает нового. Так как жизнь существует на Земле благодаря смене поколений конкретных организмов, эволюция ее протекает благодаря изменениям, происходящим в их онтогенезах. Эти изменения сводятся к тому, что конкретные онтогенезы отклоняются от пути, проложенного предковыми формами, и приобретают новые черты.
К таким отклонениям относятся, например, ценогенезы —
приспособления, возникающие у зародышей или личинок и адаптирующие их к особенностям среды обитания. У взрослых организмов ценогенезы не сохраняются. Примерами ценогенезов являются роговые образования во рту личинок бесхвостых земноводных, облегчающие им питание растительной пищей. В процессе метаморфоза у лягушонка они исчезают и пищеварительная система перестраивается для питания насекомыми и червями. К ценогенезам у амниот относят зародышевые оболочки, желточный мешок и аллантоис, а у плацентарных млекопитающих и человека — еще и плаценту с пуповиной.
Ценогенезы, проявляясь только на ранних стадиях онтогенеза, не изменяют типа организации взрослого организма, но обеспечивают более высокую вероятность выживания потомства. Они могут сопровождаться при этом уменьшением плодовитости и удлинением зародышевого или личиночного периода, благодаря чему организм в постэмбриональном или постличиночном периоде развития оказывается более зрелым и активным. Возникнув и оказавшись полезными, ценогенезы будут воспроизводиться в последующих поколениях. Так, амнион, появившийся впервые у предков пресмыкающихся в каменноугольном периоде палеозойской эры, воспроизводится у всех позвоночных, развивающихся на суше, как у яйцекладущих — пресмыкающихся и птиц, так и у плацентарных млекопитающих.
Другой тип филогенетически значимых преобразований филогенеза — филэмбриогенезы.
Они представляют собой отклонения от онтогенеза, характерного для предков, проявляющиеся в эмбриогенезе, но имеющие адаптивное значение у взрослых форм. Так, закладки волосяного покрова появляются у млекопитающих на очень ранних стадиях эмбрионального развития, но сам волосяной покров имеет значение только у взрослых организмов.
Такие изменения онтогенеза, будучи полезными, закрепляются естественным отбором и воспроизводятся в последующих поколениях. В основе этих изменений лежат те же механизмы, которые обусловливают врожденные пороки развития: нарушение пролиферации клеток, их перемещения, адгезии, гибели или дифференцировки (см. § 8.2 и 9.3). Однако от пороков их так же, как и ценогенезы, отличает адаптивная ценность, т.е. полезность и закрепленность естественным отбором в филогенезе.
В зависимости от того, на каких этапах эмбриогенеза и морфогенеза конкретных структур возникают изменения развития, имеющие значение филэмбриогенезов, различают три их типа.
1.Анаболии,
или надставки, возникают после того, как орган практически завершил свое развитие, и выражаются в добавлении дополнительных стадий, изменяющих конечный результат.
К анаболиям относят такие явления, как приобретение специфической формы тела камбалой лишь после того, как из икринки вылупляется малек, неотличимый от других рыб, а также появление изгибов позвоночника, сращение швов в мозговом черепе, окончательное перераспределение кровеносных сосудов в организме млекопитающих и человека.
2.Девиации —
уклонения, возникающие в процессе морфогенеза органа. Примером может являться развитие сердца в онтогенезе млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и трехкамерное строение, но стадия формирования неполной перегородки, характерной для пресмыкающихся, вытесняется развитием перегородки, построенной и расположенной иначе и характерной только для млекопитающих (см. § 14.4). В развитии легких у млекопитающих также обнаруживается рекапитуляция ранних стадий предков, позднее морфогенез идет по-новому (см. разд. 14.3.4).
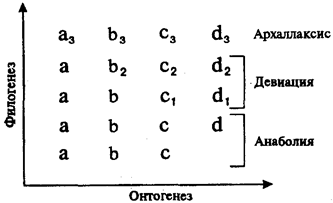
Рис. 13.9. Преобразования онто- и филогенеза в связи с возникающими филэмбриогенезами
Буквами обозначены этапы онтогенеза, цифрами — филэмбриогенетические преобразования
3.Архаллаксисы —
изменения, обнаруживающиеся на уровне зачатков и выражающиеся в нарушении их расчленения, ранних дифференцировок или в появлении принципиально новых закладок. Классическим примером архаллаксиса является
развитие волос у млекопитающих, закладка которых наступает на очень ранних стадиях развития и с самого начала отличается от закладок других придатков кожи позвоночных (см. § 14.1).
По типу архаллаксиса возникают хорда у примитивных бесчерепных, хрящевой позвоночник у хрящевых рыб (см. разд. 14.2.1.1), развиваются нефроны вторичной почки у пресмыкающихся (см. разд. 14.5.1).
Ясно, что при эволюции за счет анаболии в онтогенезах потомков полностью реализуется основной биогенетический закон, т.е. происходят рекапитуляции всех предковых стадий развития. При девиациях ранние предковые стадии рекапитулируют, а более поздние заменяются развитием в новом направлении. Архаллаксисы полностью не допускают рекапитуляции в развитии данных структур, изменяя сами их зачатки.
Если сопоставить схему филэмбриогенезов с таблицей К. Бэра (рис. 13.9), иллюстрирующей закон зародышевого сходства, то станет понятно, что Бэр уже был очень близок к открытию филэмбриогенезов, но отсутствие эволюционной идеи в его рассуждениях не позволило более чем на 100 лет опередить научную мысль.
В эволюции онтогенеза наиболее часто встречаются анаболии как филэмбриогенезы, лишь в малой степени изменяющие целостный процесс развития. Девиации как нарушения морфогенетического процесса в эмбриогенезе часто отметаются естественным отбором и встречаются поэтому значительно реже. Наиболее редко в эволюции проявляются архаллаксисы в связи с тем, что они изменяют весь ход эмбриогенеза, и если такие изменения затрагивают зачатки жизненно важных органов или органов, имеющих значение эмбриональных организационных центров (см. разд. 8.2.6), то часто они оказываются несовместимыми с жизнью.
В одной и той же филогенетической группе эволюция в разных системах органов может происходить за счет разных филэмбриогенезов.
Так, в онтогенезе млекопитающих прослеживаются все этапы развития осевого скелета в подтипе позвоночных (анаболии), в развитии сердца рекапитулируют лишь ранние стадии (девиация), а в развитии придатков кожи рекапитуляции вообще отсутствуют (архаллаксис). Знание типов филэмбриогенезов в эволюции систем органов хордовых необходимо врачу для прогнозирования возможности возникновения у плодов и новорожденных врожденных пороков развития атавистической природы (см. разд. 13.3.4). Действительно, если в системе органов, эволюционирующей путем анаболии и девиаций, возможны атавистические пороки развития за счет рекапитуляции предковых состояний, то в случае архаллаксисов это исключается полностью.
Кроме ценогенезов и филэмбриогенезов в эволюции онтогенеза могут обнаруживаться еще и отклонения времени закладки органов — гетерохронии —
и места их развития — гетеротопии.
Как первые, так и вторые приводят к изменению взаимосоответствия развивающихся структур и проходят жесткий контроль естественного отбора. Сохраняются лишь те гетерохронии и гетеротопии, которые оказываются полезными. Примерами таких адаптивных гетерохронии являются сдвиги во времени закладок наиболее жизненно важных органов в группах, эволюционирующих по типу арогенеза. Так, у млекопитающих, и в особенности у человека, дифференцировка переднего мозга существенно опережает развитие других его отделов.
Гетеротопии приводят к формированию новых пространственных и функциональных связей между органами, обеспечивая в дальнейшем их совместную эволюцию. Так, сердце, располагающееся у рыб под глоткой, обеспечивает эффективное поступление крови в жаберные артерии для газообмена. Перемещаясь в загрудинную область у наземных позвоночных, оно развивается и функционирует уже в едином комплексе с новыми органами дыхания — легкими, выполняя и здесь в первую очередь функцию доставки крови к дыхательной системе для газообмена.
Гетерохронии и гетеротопии в зависимости от того, на каких стадиях эмбриогенеза и морфогенеза органов они проявляются, могут быть расценены как филэмбриогенезы разных типов. Так, перемещение зачатков головного мозга, приводящее к его изгибу, характерному для амниот, и проявляющееся на начальных этапах его дифференцировки, является архаллаксисом, а гетеротопия семенника у человека из брюшной полости через паховый канал в мошонку, наблюдающаяся в конце эмбриогенеза после окончательного его формирования, — типичная анаболия.
Иногда процессы гетеротопии, одинаковые по результатам, могут являться филэмбриогенезами разных типов. Например, у различных классов позвоночных очень часто встречается перемещение поясов конечностей. У многих групп рыб, ведущих придонный образ жизни, брюшные плавники (задние конечности) располагаются кпереди от грудных, а у млекопитающих и человека плечевой пояс и передние конечности в дефинитивном состоянии находятся значительно каудальнее места их первоначальной закладки. В связи с этим иннервация плечевого пояса у них осуществляется нервами, связанными не с грудными, а с шейными сегментами спинного мозга. У упомянутых выше рыб брюшные плавники иннервируются нервами не задних туловищных, а передних сегментов, расположенных кпереди от центров иннервации грудных плавников. Это свидетельствует о гетеротопии закладки плавников уже на стадии самых ранних зачатков, в то время как перемещение переднего пояса конечностей у человека происходит на более поздних этапах, когда иннервация их уже полностью осуществлена. Очевидно, в первом случае гетеротопия представляет собой архаллаксис, в то время как во втором — анаболию.
Ценогенезы, филэмбриогенезы, а также гетеротопии и гетерохронии, оказавшись полезными, закрепляются в потомстве и воспроизводятся в последующих поколениях до тех пор, пока новые адаптивные изменения онтогенеза не вытеснят их, заменив собой. Благодаря этому онтогенез не только кратко повторяет эволюционный путь, пройденный предками, но и прокладывает новые направления филогенеза в будущем.
Организм, или особь, — отдельное живое существо, в процессе онтогенеза проявляющее все свойства живого. Постоянное взаимодействие особи с окружающей средой в виде организованных потоков энергии и вещества поддерживает ее целостность и развитие. В структурном отношении организм представляет собой интегрированную иерархическую систему, построенную из клеток, тканей, органов и систем, обеспечивающих его жизнедеятельность. Подробнее остановимся на органах и системах жизнеобеспечения.
Органом
называют исторически сложившуюся специализированную систему тканей, характеризующуюся отграниченностью, постоянством формы, локализации, внутренней конструкции путей кровообращения и иннервации, развитием в онтогенезе и специфическими функциями. Строение органов часто очень сложно. Большинство из них полифункционально, т.е. выполняет одновременно несколько функций. В то же время в реализации какой-либо сложной функции могут участвовать различные органы.
Группу сходных по происхождению органов, объединяющихся для выполнения сложной функции, называют системой
(кровеносная, выделительная и др.).
Если одну и ту же функцию выполняет группа органов разного происхождения, ее называют аппаратом.
Примером служит дыхательный аппарат, состоящий как из органов собственно дыхания, так и из элементов скелета и мышечной системы, обеспечивающих дыхательные движения.
В процессе онтогенеза происходит развитие, а часто и замена одних органов другими. Органы зрелого организма называют дефинитивными;
органы, развивающиеся и функционирующие только в зародышевом или личиночном развитии, — провизорными.
Примерами провизорных органов являются жабры личинок земноводных, первичная почка и зародышевые оболочки высших позвоночных животных (амниот).
В историческом развитии преобразования органов могут иметь прогрессивный или регрессивный характер. В первом случае органы увеличиваются в размерах и становятся более сложными по своему строению, во втором — уменьшаются в размерах, а их строение упрощается.
Если у двух организмов, находящихся на разных уровнях организации, обнаруживаются органы, которые построены по единому плану, расположены в одинаковом месте и развиваются сходным образом из одинаковых эмбриональных зачатков, то это свидетельствует о родстве данных организмов. Такие органы называют гомологичными.
Гомологичные органы часто выполняют одну и ту же функцию (например, сердце рыбы, земноводного, пресмыкающегося и млекопитающего), но в процессе эволюции функции могут и меняться (например, передних конечностей рыб и земноводных, пресмыкающихся и птиц).
При обитании неродственных организмов в одинаковых средах у них могут возникать сходные приспособления, которые проявляются в возникновении аналогичных
органов. Аналогичные органы выполняют одинаковые функции, строение же их, местоположение и развитие резко различны. Примерами таких органов являются крылья насекомых и птиц, конечности и челюстной аппарат членистоногих и позвоночных.
Строение органов строго соответствует выполняемым ими функциям. При этом в исторических преобразованиях органов изменение функций непременно сопровождается и изменением морфологических характеристик органа.
13.3.1. Дифференциация и интеграция
в эволюции органов
Основным принципом эволюции органических структур является принцип дифференциации.
Дифференциация представляет собой разделение однородной структуры на обособленные части, которые в силу различного положения, связей с другими органами и различных функций приобретают специфическое строение. Таким образом, усложнение структуры всегда связано с усложнением функций и специализацией отдельных частей. Дифференцированная структура выполняет несколько функций, и строение ее сложно.
Примером филогенетической дифференциации может являться эволюция кровеносной системы в типе хордовых. Так, у представителей подтипа бесчерепных она построена очень просто: один круг кровообращения, отсутствие сердца и капилляров в системе жаберных артерий.
В надклассе рыб имеются двухкамерное сердце и жаберные капилляры. У земноводных впервые появляется разделение кровеносной системы на два круга кровообращения, а сердце становится трехкамерным. Максимальная дифференциация характерна для кровеносной системы млекопитающих, сердце которых четырехкамерное, а в сосудах достигается полное разобщение венозного и артериального кровотоков.
Отдельные части дифференцирующейся, ранее однородной структуры, специализируясь на выполнении одной функции, становятся функционально все более зависимыми от других частей данной структуры и от организма в целом. Такое функциональное соподчинение отдельных компонентов системы в целостном организме называют интеграцией.
Четырехкамерное сердце млекопитающих представляет собой пример высокоинтегрированной структуры: каждый отдел выполняет лишь свою специальную функцию, не имеющую никакого смысла в отрыве от функций других отделов. Поэтому сердце снабжено автономной системой функциональной регуляции в виде парасимпатического атриовентрикулярного нервного узла и при этом строго подчинено нейрогуморальной системе регуляции организма в целом.
Таким образом, одновременно с дифференциацией наблюдается и подчинение частей целостной системе организма, т.е. процесс интергации.
13.3.2. Закономерности морфофункциональных преобразований органов
В основе филогенетических преобразований органов лежит их полифункциональность и способность к количественным изменениям функций. Практически все органы выполняют не одну, а несколько функций, причем среди них всегда выделяется главная, а остальные второстепенны. Строение такого полифункционального органа обязательно соответствует главной функции. Так, рука человека может использоваться для лазания по деревьям, плавания, даже хождения. Но основной ее функцией является трудовая деятельность. В связи с этим и строение руки в максимальной степени соответствует функции труда.
Один из основных принципов эволюции органов — принцип расширения и смены функций.
Расширение функций сопровождает обычно профессивное развитие органа, который по мере дифференциации выполняет все новые функции. Так, парные плавники рыб, возникшие как пассивные органы, поддерживающие тело в воде в горизонтальном положении, с приобретением собственной мускулатуры и прогрессивным расчленением становятся еще и активными рулями глубины и поступательного движения. У придонных рыб они обеспечивают также их передвижение по дну. С переходом позвоночных на сушу к перечисленным функциям конечностей добавились хождение по Земле, лазание, бегание и др.
Расширение функций сопровождается специализацией, благодаря которой главной функцией становится одна из бывших ранее второстепенными. Бывшая главной функция преобразуется во второстепенную и может впоследствии даже исчезнуть. Орган при этом меняется таким образом, что его строение становится максимально соответствующим выполнению главной функции. Так, переход предков ластоногих и китообразных к водному образу жизни привел к преобразованию их парных конечностей в ласты, практически утратившие способность обеспечивать передвижение по суше. Жизнь ленивцев, представителей отряда неполнозубых, на деревьях привела к формированию у них крючкообразных конечностей, с помощью которых возможно лишь медленное перемещение по веткам в подвешенном состоянии с почти полной утратой способности движения по земле.
Нередко функции, выполняемые органами, могут измениться кардинально. Так, плавательный пузырь рыб, будучи гидростатическим органом, у кистеперых рыб становится дополнительным органом дыхания, а у земноводных он преобразуется в легкое, и основной функцией его становится дыхательная. У пресмыкающихся и млекопитающих, ведущих наземный образ жизни, легкие выполняют только дыхательную функцию, но первичная функция плавательного пузыря сохраняется за легкими у крокодилов, ластоногих и китообразных, ведущих водный образ жизни, а также у наземных форм во время плавания.
В других случаях видоизменения органов в связи со сменой их функций столь велики, что выполнение ими функций, бывших ранее главными, становится невозможным. Так, передние жаберные дуги предков хрящевых рыб преобразовались в челюсти, а у наземных позвоночных они стали выполнять функции звукопроводящего аппарата, превратившись в слуховые косточки (см. разд. 14.2.1). Участие их в пищеварении и дыхании стало невозможным.
В прогрессивной эволюции органов очень важным является принцип активации функций.
Он наиболее часто реализуется на начальных этапах эволюции органов в том случае, когда малоактивный орган начинает активно выполнять функции, существенно при этом преобразуясь.
Так, крайне малоподвижные парные плавники хрящевых рыб становятся активными органами движения уже у костистых.
Более часто в филогенезе наблюдается интенсификация функций,
являющаяся следующим этапом эволюции органов после активации. Благодаря этому орган обычно увеличивается в размерах, претерпевает внутреннюю дифференцировку, гистологическое строение его усложняется, нередко наблюдается многократное повторение одноименных структурных элементов, или полимеризация
структуры. Примером является усложнение структуры легких в ряду наземных позвоночных за счет ветвления бронхов, появления ацинусов и альвеол на фоне постоянной интенсификации его функций.
Высокая степень дифференцировки может сопровождаться уменьшением количества одинаковых органов, выполняющих одну и ту же функцию, или их олигомеризацией.
Это явление наблюдается, к примеру, в эволюции артериальных жаберных дуг, которые закладываются у хрящевых рыб в количестве 6—7 пар, у костных рыб их становится 4 пары, а у млекопитающих и человека сохраняются в дефинитивном состоянии лишь части 3, 4 и 6-й пар (см. § 14.4).
Иногда в процессе интенсификации функций наблюдается тканевая субституция органа —
замещение одной ткани другой, более соответствующей выполнению данной функции. Так, хрящевой скелет хрящевых рыб сменяется на костный у более высокоорганизованных классов позвоночных.
В противоположность интенсификации и активации ослабление функций
ведет в филогенезе к упрощению строения органа и его редукции, вплоть до полного исчезновения.
13.3.3. Возникновение и исчезновение
биологических структур в филогенезе
В процессе эволюции закономерным является как возникновение
новых структур, так и их исчезновение.
В основе лежит принцип дифференциации, проявляющийся на фоне первичной полифункциональности и способности функций изменяться количественно. Любая структура при этом возникает на основе предшествующих структур вне зависимости от того, на каком уровне организации живого осуществляется процесс филогенеза. Так, известно, что около 1 млрд. лет назад исходный белок глобин вслед за дупликацией исходного гена дифференцировался на мио- и гемоглобин — белки, входящие в состав соответственно мышечных и кровяных клеток и дифференцировавшиеся в связи с этим по функциям. В филогенезе центральной нервной системы хордовых также можно видеть дифференцировку и смену функций структур: головной мозг формируется из переднего конца нервной трубки. Таким же образом новые биологические виды образуются в виде изолированных популяций исходных видов (см. § 11.6), а новые биогеоценозы — за счет дифференцировки предсуществующих (см. § 16.2).
В связи с тем что ниже будут рассмотрены филогенезы конкретных систем органов, подробнее остановимся на закономерностях возникновения и исчезновения органов. Примером возникновения
органов служит происхождение матки плацентарных млекопитающих от парных яйцеводов. При удлинении эмбрионального развития млекопитающих возникает необходимость более длительной задержки зародыша в организме матери. Это может осуществляться только в каудальных отделах яйцеводов, полость которых при этом увеличивается, а стенка дифференцируется таким образом, что к ней прикрепляется плацента, обеспечивающая взаимосвязь организма матери и плода. В процессе естественного отбора сохранялись и успешно размножались в первую очередь те млекопитающие, в организмах самок которых потомство развивалось наиболее долго. В итоге возник новый орган — матка, обеспечивающий зародышу оптимальные условия внутриутробного развития (см. разд. 14.5.3) и повышающий выживаемость соответствующих видов.
В возникновении такого более сложного и специализированного органа, как глаз, наблюдаются те же закономерности. В основе формирования органа зрения, как и всех органов чувств, лежат клетки кожного эпителия, среди которых дифференцируются и рецепторные, в частности светочувствительные. Объединение их в группы приводит к возникновению примитивных обособленных органов зрения, позволяющих животным лишь оценивать освещенность. Погружение такого светочувствительного органа под кожу обеспечивает сохранность нежных клеток, но при этом зрительная функция может осуществляться только благодаря возникновению прозрачности покровов. Чувствительность к свету примитивного органа зрения усиливается при утолщении прозрачных покровов и приобретении ими способностей преломлять свет и фокусировать его лучи на чувствительных клетках глаза. Сложный орган требует вспомогательного аппарата — защитных структур, мышц, приводящих его в движение, и т.д. Возросший уровень сложности организации глаза с необходимостью сопровождается усложнением регуляции его функций, что и выражается в усилении его интеграции как целостной системы.
Исчезновение,
или редукция, органа в филогенезе может быть связана с тремя разными причинами и имеет различные механизмы. Во-первых, орган, выполнявший ранее важные функции, может оказаться в новых условиях вредным. Против него срабатывает естественный отбор, и орган довольно быстро может полностью исчезнуть. Примеров такого прямого исчезновения органов немного. Так, многие насекомые малых океанических островов бескрылы вследствие постоянной элиминации из их популяций летающих особей ветром. Чаще наблюдается исчезновение органов благодаря их субституции новыми структурами, выполняющими прежние функции с большей интенсивностью. Так исчезают, например, у пресмыкающихся и млекопитающих предпочки и первичные почки, заменяясь функционально вторичными почками. Таким же образом у рыб и земноводных происходит вытеснение хорды позвоночником.
Самый частый путь к исчезновению органов — через постепенное ослабление их функций. Такие ситуации возникают обычно при изменении условий существования. Орган, почти не выполняющий функций, выходит из-под контроля естественного отбора и проявляет обычно повышенную изменчивость. Возникающие изменения вызывают нарушение коррелятивных связей с другими частями организма. Благодаря этому такой орган зачастую становится вредным и против него начинает действовать естественный отбор.
В медицинской практике широко известно, что рудиментарные органы и у человека характеризуются широкой изменчивостью. Третьи большие коренные зубы, или «зубы мудрости», например, характеризуются не только значительной вариабельностью строения и размеров, но и разными сроками прорезывания, а также особой подверженностью кариесу. Иногда они вообще не прорезываются, а нередко, прорезавшись, в течение ближайших лет полностью разрушаются. То же касается и червеобразного отростка слепой кишки (аппендикса), который в норме может иметь длину от 2 до 20 см и быть расположенным по-разному (за брюшиной, на длинной брыжейке, позади слепой кишки и т.д.). Кроме того, воспаление аппендикса (аппендицит) встречается значительно чаще, чем воспалительные процессы в других отделах кишечника.
Процесс редукции органа противоположен по отношению к его нормальному морфогенезу. Прежде всего выпадают закладки таких частей органа, которые в норме формируются последними. При недоразвитии конечностей у человека обычно в первую очередь недоразвиваются фаланги I и V пальцев, закладывающиеся последними. У китообразных, совершенно лишенных задних конечностей благодаря ослаблению их функций в филогенезе, все же остаются закладки элементов тазового пояса, формирующиеся в процессе морфогенеза наиболее рано.
Исследования генетических основ редукции органов показали, что структурные гены, регулирующие морфогенез, не исчезают, в то время как существенным изменениям подвергаются гены, регулирующие время закладки рудиментарных органов, либо гены, ответственные за феномен индукционных взаимодействий в развивающемся зародыше. Действительно, при пересадках мезодермального материала дна ротовой полости зародыша ящерицы в ротовую полость развивающегося цыпленка возможно формирование у последнего зубов типичного строения, а пересадка кожной мезодермы ящерицы под эпидермис спины цыпленка приводит к формированию в нем типичных роговых чешуи вместо перьев.
Недоразвившиеся органы носят название рудиментарных
или рудиментов.
К рудиментам у человека относят, во-первых, структуры, потерявшие свои функции в постнатальном онтогенезе, но сохраняющиеся и после рождения (волосяной покров, мышцы ушной раковины, копчик, аппендикс как пищеварительный орган), и, во-вторых, органы, сохраняющиеся только в эмбриональном периоде онтогенеза (хорда, хрящевые жаберные дуги, правая дуга аорты, шейные ребра и др.).
Крайне редко редукция органов оказывается настолько полной, что от них не остается даже эмбриональных закладок. Примером такого полного исчезновения органов является редукция парных конечностей у большинства змей. Обычно же эмбриональные закладки органов, выполнявших серьезные функции даже у весьма отдаленных предков и давно утративших свое значение в постнатальном онтогенезе, продолжают развиваться у потомков в эмбриогенезе неопределенно долгое время. В первую очередь это относится к органам, связанным тесными морфогенетическими корреляциями с другими зачатками в зародышевом развитии. Так, хорда у всех позвоночных выполняет роль организационного центра, под контролем которого развиваются нервная трубка, позвоночник и дифференцируются сомиты. Нарушение развития даже участка хорды влечет за собой грубые пороки развития соответствующих структур.
Не менее тесными взаимными связями характеризуются элементы мочеполовой системы позвоночных. Так, мочеточники, связывающие нефроны предпочки с внешней средой у круглоротых, преобразуются у рыб и более высокоорганизованных классов позвоночных в два мочеполовых протока, выполняющих разные функции (см. разд. 14.5.3), а рудиментирующиеся нефроны первичной почки у пресмыкающихся и млекопитающих стимулируют развитие вторичной почки.
13.3.4. Атавистические пороки развития
Различного рода нарушения эмбриогенеза могут привести к формированию у высокоорганизованных организмов и человека таких признаков, которые при нормальных условиях у них не встречаются, но присутствуют у более или менее отделенных предков. Такие признаки называют атавизмами.
Если они снижают жизнеспособность и проявляются как морфологические аномалии, то их называют атавистическими
или анцестральными
(от франц. ancetre — предок) пороками развития.
По механизмам формирования различают три варианта атавизмов. Наиболее часто встречаются атавизмы, связанные с недоразвитием органов на тех этапах морфогенеза, когда они рекапитулировали предковое состояние. К примерам аномалий такого рода относятся двух- и трехкамерное сердце, гипоплазия, или недоразвитие диафрагмы, срединная расщелина твердого нёба, или «волчья пасть», и др. Атавизмы другого рода — результат нарушения редукции — персистирование (сохранение) и дальнейшее развитие эмбриональных структур, также рекапитулирующих морфологию, характерную для предков. К ним относят персистирование боталлова протока и правой дуги аорты (см. § 14.4), наличие ребер, связанных с шейным отделом позвоночника, боковые свищи шеи (см. разд. 14.2.1, 14.3.4). Третий тип атавистических пороков развития возникает в связи с нарушением перемещения органов в онтогенезе, результатом чего является их расположение в тех частях тела, где при нормальных условиях они находятся у предковых форм. У человека широко известны тазовое расположение почек (см. разд. 14.5.1), крипторхизм (неопущение яичника), высокое стояние плечевого пояса и др.
Во всех случаях атавизмов ведущими механизмами их возникновения являются не обратные мутации, приводящие к формированию предкового фенотипа, а, вероятно, мутации регуляторных генов, которые контролируют скорость морфогенеза и запуск процессов, направленных на редукцию органов. Действительно, для формирования любой структуры в многоклеточном организме необходимо слаженное функционирование десятков и даже сотен структурных генов. Одновременное возникновение адекватных друг другу мутаций целого комплекса таких генов крайне маловероятно. Однако на базе генных комплексов, унаследованных от предков, многие предковые структуры закономерно рекапитулируют в ходе эмбриогенеза, а время этих рекапитуляции и редукций контролируется значительно проще и небольшим количеством генов. Мутации таких регуляторных генов гораздо более вероятны.
13.3.5. Аллогенные аномалии и пороки развития
Атавистические пороки развития, объясняющиеся ходом предшествующей эволюции, относят к разряду филогенетических пороков. К этой же группе аномалий принадлежат и так называемые аллогенные аномалии.
Это врожденные пороки, имеющие в своей основе генетические дефекты. Они встречаются одновременно у ряда родственных организмов и являются выражением закона гомологических рядов.
Н.И. Вавилова 1935 г. обратил внимание на то, что растения разных видов и родов, сходные по происхождению, обладают сходными рядами наследственной изменчивости, и объяснил эту закономерность сходством основных характеристик аллелофондов видов, еще недавно претерпевших дивергенцию. Эта закономерность была названа законом гомологических рядов.
Действие закона гомологических рядов распространяется на широкий круг биологических объектов. Попытки сопоставить гомологии генотипа с морфофункцйональными гомологиями не всегда успешны. Действительно, нет уверенности в том, что такое, с точки зрения сравнительной анатомии, бесспорно гомологическое образование, как хорда зародыша человека и личинки асцидии (животного, относящегося к подтипу Личиночно-хордовые Urochordata типа Хордовые), является абсолютно гомологичным и с позиций генетического контроля его морфогенеза. Однако изучение кариотипов цитогенетическим методом с использованием дифференциальной окраски хромосом и последовательности нуклеотидов в молекулах ДНК методом гибридизации (см. § 15.2) у близких и более отделенных видов привело к выводу о том, что закон гомологических рядов распространяется по крайней мере на виды в пределах рода, семейства, отряда, а по многим признакам — ив рамках класса. Так, известно, что ген, ответственный за синтез фактора VIII в системе свертывания крови, расположен в Х-хромосоме не только у человека, но и у приматов и даже у собак. Поэтому и у этих видов животных гемофилия встречается преимущественно у самцов, наследуясь так же, как и у человека.
В природе хорошо известен ген альбинизма, гомологичный у всех млекопитающих, а также и у других классов позвоночных. Среди млекопитающих распространен аутосомно-доминантный ген ахондроплазии, или хондродистрофической карликовости, нарушающий рост 70 длинных трубчатых костей. В морфогенезе мягких тканей морды млекопитающих и лица человека имеется срастание двух половин верхней губы за счет избирательной клеточной адгезии. У ряда травоядных животных, питающихся грубыми злаками или колючими ветками (грызуны, зайцы, верблюды), срастание верхней губы нерационально. В процессе естественного отбора у них закрепились адаптивные мутации, нарушающие клеточную адгезию верхней губы. У человека такое нарушение называют заячьей губой.
У людей аллогенные аномалии встречаются очень часто. Даже такой генетический дефект, как синдром Дауна, является аллогенной аномалией: известны случаи рождения детенышей гориллы с трисомией хромосомы, соответствующей 21-й паре хромосом человека. Симптоматика такой трисомии соответствует клинике синдрома Дауна у человека.
Точная диагностика и изучение механизмов возникновения аллогенных состояний представляют большой интерес и практически важны потому, что дают возможность использования животных с соответствующими признаками в качестве экспериментальных моделей для изучения клиники и лечения их у человека.
Рассмотренные выше филогенетические преобразования отдельных биологических структур протекают в эволюционирующих группах организмов как целостных системах. Устойчивые взаимозависимости органов и систем, проявляющиеся в филогенезе, называют координациями.
Механизмы соотносительных преобразований биологических структур различны, в связи с чем выделяют три их группы: биологические, динамические
и топографические.
Биологические координации
наблюдаются между структурами, непосредственно не связанными ни по функциям, ни по месту положения. Основным связующим звеном между ними являются адаптации к определенным условиям обитания. Так, у большинства эндопаразитов (см. § 18.6) сильно развиты половая система и органы прикрепления к телу хозяина, но при этом недоразвиты органы чувств и опорно-двигательный аппарат. Млекопитающие, обитающие на деревьях, обычно имеют стереоскопическое зрение и сильно развитый мозжечок. Позвоночные, которые дышат кислородом, растворенным в воде, имеют хвостовой плавник, жабры, слабо развитый передний мозг и содержат гемоглобин, способный связываться с кислородом при низком его парциальном давлении в среде. Все позвоночные, имеющие матку, обладают также совершенной системой терморегуляции.
Динамические координации
выражаются во взаимном соответствии структур, связанных функционально. Тесные динамические координации имеются, например, между органами кровеносной и дыхательной систем. Так, животные, дышащие легкими, имеют трех- или четырехкамерное сердце и два круга кровообращения. Степень развитости нервных центров всегда соответствует интенсивности функционирования иннервируемых органов. Например, строение органа обоняния и обонятельные доли мозга у млекопитающих высоко развиты, в то время как у птиц примитивное строение периферической части обонятельного анализатора соответствует малым размерам обонятельных долей мозга. Это связано с тем, что в ориентации млекопитающих обоняние играет первостепенную роль, а для птиц оно не имеет большого значения.
Топографические координации
проявляются между структурами, связанными друг с другом пространственно. Так, для каждого типа животного царства характерен своеобразный общий план строения, выражающийся в определенном взаимном расположении основных органов и систем. Например, у всех представителей типа Хордовые на спинной стороне тела расположена нервная трубка, под ней лежат хорда, пищеварительная трубка и брюшной кровеносный сосуд, а по бокам тела — производные мезодермы (см. разд. 13.5.4).
Все типы координации характеризуются высокой степенью устойчивости. Так, хордовые животные, известные с конца протерозойской эры, сохранили неизменными общий план строения до настоящего времени, на протяжении более 500 млн. лет. Феномен паразитизма возник значительно раньше появления хордовых, и поэтому комплекс адаптации к паразитическому образу жизни (см. § 18.6) является еще более древним. Длительно существуют и другие, более частные координации, возникающие вместе с появлением новых таксономических или экологических групп организмов.
Высокая устойчивость филогенетических координации обеспечивается целостностью онтогенеза каждой конкретной особи, развитие всех биологических структур которой протекает в строгом взаимном соответствии. Такое соответствие структур развивающегося организма в онтогенезе называют онтогенетическими корреляциями.
Различают геномные, эргонтические и морфогенетические корреляции.
Геномные корреляции
обеспечиваются целостностью генетической конституции развивающегося организма. Ведущими механизмами геномных корреляций являются генный баланс генотипа, сцепленное наследование генов, различные формы взаимодействия генов, а также плейотропность. Так, генные системы, регулирующие процессы пролиферации и избирательной гибели клеток на различных этапах органогенеза (см. разд, 8.2.1, 8.2.4), приводят к аллометрическому росту органов (см. разд. 8.3.3), благодаря чему появляются, например, удлиненный клюв, шея и задние конечности у большинства болотных птиц, длинная шея и ноги у жирафа, а также отличающиеся друг от друга пропорции тела у мужчин и женщин.
Морфогенетические корреляции
возникают между органами, пространственно связанными между собой. Они основаны либо на феномене эмбриональной индукции (см. разд. 8.2.6), либо на общности эмбриональных закладок органов. Эти корреляции проявляются уже на ранних стадиях онтогенеза, когда еще отсутствуют функциональные связи между формирующимися органами. Так, зачаток хорды обусловливает развитие нервной трубки на спинной стороне зародыша и дифференцировку скелетогенной ткани внутренних частей сомита — склеротома в хрящ или кость, а глазной бокал (вырост переднего мозга) — формирование хрусталика при морфогенезе глаза.
Примером развития ряда структур из одного общего зачатка является формирование у млекопитающих и человека из закладок 1-й и 2-й пар жаберных дуг первичных челюстей, подъязычной кости, части хрящей гортани, шиловидного отростка черепа и трех слуховых косточек. При нарушении развития закладок жаберных дуг обычно образуется комплекс аномалий указанных структур, называемый аномаладом первых жаберных дуг.
Эргонтические корреляции
обусловлены функциональной взаимозависимостью органов и частей организма. Они возникают на более поздних стадиях развития, когда органы начинают функционировать, и проявляются, например, в соответствии между степенью развитости мышцы, костных выступов, к которым она прикрепляется, и интенсивностью ее кровоснабжения. К такого рода корреляциям относят также соответствие вторичных половых признаков развитию гонад.
Ведущие корреляции в онтогенезе — геномные.
В конечном счете именно они лежат в основе других корреляций, значение которых на протяжении онтогенеза меняется. Это связано с первичностью изменений генотипа в процессе филогенеза.
Система корреляций и сама подвергается эволюционным преобразованиям. В процессе эволюции видоизменяются вначале более частные корреляции, в то время как наиболее общие могут воспроизводиться в конкретных онтогенезах очень длительное время. В результате в ходе исторического развития происходит как отбор наиболее общих корреляций, имеющих значение при любых перестройках организма и в различных условиях обитания (общий план строения, соответствие между степенью развитости нервного центра и иннервируемыми органами), так и накопление локальных корреляций частного значения, формирующихся у организмов разных видов и отражающих специфику их образа жизни.
Корреляции общего значения обусловливают преемственность формообразовательных процессов в череде поколений организмов данного типа организации, а частные корреляции — многообразие конкретных форм жизни.
Между филогенетическими координациями и онтогенетическими корреляциями имеется теснейшая связь. Очевидно, что корреляции существуют и воспроизводятся в поколениях благодаря тому, что на протяжении предшествующей эволюции органов они преобразовывались скоординированно. С другой стороны, филогенетические координации в последующей эволюции организмов будут реализовываться благодаря воспроизведению онтогенетических корреляций в ходе индивидуального развития конкретных особей. Таким образом, в виде соотношения корреляций и координации проявляется диалектическое единство онто- и филогенеза как целостного процесса исторического развития живого.
Сопоставление конкретных форм соотносительных преобразований органов приводит к выводу о том, что морфогенетические корреляции и топографические координации точно так же, как эргонтические корреляции и динамические координации, взаимно обусловливают друг друга. В паре геномные корреляции — биологические координации также обнаруживается соответствие.
Осознание целостности и взаимообусловленности индивидуального и исторического развития необходимо врачу в связи с тем, что соотносительное преобразование биологических структур лежит в основе нормального развития организма человека, а нарушение даже одного незначительного компонента развивающейся системы может повлечь за собой возникновение целого комплекса нарушений в других ее элементах. С этим связано то, что практически все врожденные пороки развития, а также многочисленные генные болезни характеризуются не отдельными патологическими признаками, а синдромами, представляющими собой комплексы симптомов, связанных между собой процессом формирования.
Сердце у всех позвоночных закладывается на ранних этапах развития кпереди от глотки под челюстной дугой. В его морфогенезе участвует глотка как эмбриональный индуктор. Если это свойство глотки нарушено, тсгсердце может задержаться на двух- и трехкамерном уровне развития, при этом может быть нарушено и его перемещение в загрудинную область — шейная эктопия сердца
(см. § 14.4). Эти явления — результат нарушений морфогенетических корреляций в развитии шейной области. Часто этот порок развития сопровождается нарушением отходящих от сердца сосудов (персистирование общего эмбрионального ствола, двух дуг аорты и т.д.) и недоразвитостью легких.
В возникновении данных аномалий ведущим механизмом выступает нарушение эргонтических корреляций сердце — сосуды — легкие. Первичным нарушением в описанном комплексе признаков является, вероятно, нарушение генетического контроля эмбриональной индукции, описанное ранее (см. разд. 8.2.6). Таким образом, приведенный пример иллюстрирует взаимоотношение разных форм соотносительных преобразований органов при формировании сложного комплекса патологических признаков, имеющих в целом атавистическую природу.
Примером нарушения чисто геномных корреляций является синдром Дауна. Увеличение доз генов 21-й хромосомы или ее части приводит к формированию тяжелой умственной отсталости, ослаблению тонуса мышц, аномалиям мозгового черепа и мягких частей лица, светлой пигментации волос и глаз.
Кроме филогенетических координации, подкрепляемых в каждом поколении онтогенетическими корреляциями, целостность развивающегося организма отражают и такие соотносительные преобразования органов, как субституция и гетеробатмия.
Субституция —
это такое эволюционное преобразование, при котором один орган замещается другим, выполняющим обычно ту же функцию с большей интенсивностью. При этом наблюдается развитие этих органов в разных направлениях. Один обычно подвергается редукции, другой — эволюционирует прогрессивно. Так, хорда замещается позвоночником и превращается в рудиментарное образование, а первичные хрящевые челюсти позвоночных заменяются вторичными костными (см. разд. 14.2.1). Это примеры гомотопной
субституции, когда новый орган возникает на месте старого. При гетеротопной
субституции заменяющий орган находится на новом месте. Так, функцию печени как органа кроветворения берет на себя красный костный мозг. Выделительная функция выполняется у рыб и земноводных туловищной почкой, а у пресмыкающихся и млекопитающих — тазовой.
Гетеробатмия —
это такое эволюционное преобразование, при котором в одной группе организмов обнаруживается разный уровень эволюционной продвинутое™ и специализации разных частей одного и того органа, разных органов одной и той же системы или разных частей организма. Примером может являться человек, головной мозг которого за короткое время антропогенеза претерпел колоссальные морфофизиологические изменения, в то время как пищеварительная система соответствует уровню развития других приматов.
Гетеробатмия, наблюдающаяся внутри одной и той же системы органов в разных филогенетических группах, обусловливает феномен компенсации функций,
благодаря которому одни и те же экологические задачи решаются разными способами. Так, грызуны и копытные млекопитающие питаются одинаковой растительной пищей, но у первых наиболее выраженные адаптации к растительноядности проявляются в строении зубов и морфофизиологии слюнных желез, в то время как вторые на фоне примитивной зубной системы имеют высокоспециализированные желудок и кишечник. Явления гетеробатмии и, следовательно, компенсации функций имеют огромное эволюционное значение в связи с тем, что в организме, даже вступившем на путь узкой специализации, всегда остаются органы и системы относительно мало специализированные, которые при меняющихся условиях могут еще прогрессивно развиваться, раскрывая перед такими филогенетическими группами новые адаптивные возможности.
Все живые организмы подразделяются на неклеточных (вирусы) и клеточных (все остальные). Несмотря на то что филогенетические взаимоотношения между ними неясны, вирусы как облигатно-парази-тические формы (см. § 18.3), возможно, возникли от более высокоорганизованных организмов за счет упрощения в процессе адаптации к паразитизму. В то же время не исключена возможность существования вирусов как фрагментов нуклеиновых кислот еще на предбиологическом этапе эволюции и приобретения ими основных свойств живых организмов позже, при попадании в клетки. Клеточные организмы подразделяются на про- и эукариот. Эукариоты, вероятно, произошли от прокариот (см. § 1.5).
13.5.1. Типы питания и основные группы
живых организмов в природе
Древние эукариоты, будучи одноклеточными, по характеру питания специализировались на группы организмов, активно добывающих пищу путем ее поиска и захвата, и формы, добывающие продукты питания за счет их всасывания из среды. Первый способ питания называют голозойным,
второй — голофитным.
Прогрессивная эволюция первого способа питания сопровождает возникновение царства Животные Animalia и весь их последующий филогенез. Поэтому главными чертами животного организма являются способность к перемещению, активному захвату и переработке пищи. Второй способ питания, наоборот, предусматривает приобретение клетками дополнительных защитных оболочек, усложняющих их захват и переваривание и, следовательно, неподвижность. Клетки части организмов, эволюционирующих по этому пути, приобрели клеточную стенку из полисахарида хитина, позволяющую всасывание высокомолекулярных органических веществ. Такие организмы перешли к сапрофитному питанию,
т. е. к потреблению разлагающихся органических веществ, и дали начало филогенезу царства Грибы Fungi.
Клеточная стенка других организмов построена из целлюлозы, позволяющей всасывать из среды лишь воду, углекислый газ и неорганические ионы. Приобретение ими способности к фотосинтезу явилось ключевым моментом в прогрессивной эволюции царства Растения Plantae.
Подробнее остановимся на филогенезе царства Животные. Важнейшим прогрессивным событием в эволюции животных является возникновение многоклеточности.
13.5.2. Происхождение многоклеточных животных
Прежде всего необходимо определить понятие многоклеточное животное.
Важнейшими чертами многоклеточности являются следующие:1) тело животного состоит из большого количества клеток; 2) клеткидифференцированы на половые и соматические, а последние различаются также по структуре и функциям; 3) клетки расположены в организме в несколько слоев; 4) клетки интегрированы в целостную систему благодаря существованию жидкостной внутренней среды и нервной системы.
В процессе эволюции первая характеристика многоклеточности достигается наиболее просто: среди простейших уже имеется огромное количество видов колониальных организмов. Остальные черты многоклеточности связаны с возникновением многослойности, обеспечивающей как различные условия существования клеткам, расположенным на поверхности и внутри тела, так и появление внутренней среды, объединяющей их воедино. На рис. 13.10 представлены различные варианты объединения клеток в надклеточные комплексы. Ясно, что только интеграция клеток в шаровидную структуру дает им возможность оказаться в разных условиях, дифференцироваться и взаимодействовать друг с другом.
Рис. 13.10. Варианты объединения клеток в надклеточные образования:
I
—нитевидная колония клеток, II
—колония в виде однослойного пласта клеток, III
—двуслойная колония клеток, IV—
сферический многоклеточный организм с клетками разных типов
Родоначальником многоклеточных в настоящее время считают шаровидную колонию жгутиковых, половые клетки которых перемещались в глубь колонии, а соматические первично выполняли как функцию перемещения всей колонии в пространстве, так и пищеварения за счет переваривания фагоцитированных пищевых частиц, захваченных из воды.
Осуществление одной и той же клеткой функций движения и пищеварения малоэффективно. С этим связана последующая специализация клеток в направлении преимущественно пищеварения или обеспечения движения. Результатом является возникновение фагоцитобласта
(внутреннего слоя амебовидных клеток, занимающихся пищеварением) и кинобласта
(наружного слоя клеток со жгутиками, обеспечивающими движение).
Стойкая дифференцировка соматических клеток по функциям и строению, возникшая первоначально на фоне выделения двух клеточных слоев, явилась ключевым моментом в происхождении многоклеточных. Именно с двуслойностью связано появление жидкой внутренней среды, через которую клетки обмениваются химическими сигналами, а также дальнейшее обособление и специализация части поверхностных клеток в направлении восприятия внешних раздражителей и передача возбуждения на другие клетки, располагающиеся в отдалении от них. Таким образом возникают предпосылки к формированию нервной системы.
Гипотетический предок многоклеточных животных назван фагоцителлой
(рис. 13.11). Он плавал в толще воды за счет биения ресничек кинобласта, а питался, захватывая взвешенные в среде частички пищи и переваривая их клетками фагоцитобласта. На более поздних этапах эволюции происходили многочисленные адаптации потомков фагоцителлы к многообразным условиям существования при оседании их на дно или при перемещении к поверхности, а также при изменении источников питания (захват мелких или крупных, живых или мертвых пищевых частиц).
Рис. 13.11. Этапы происхождения многоклеточности:I
,II
—сферические колонии жгутиковых, III
—V—
фагоцителлы разной степени сложности; 1
—кинобласт, 2
—рыхлый фагоцитобласт, 3—
скопление чувствительных клеток на переднем конце тела, 4—
ротовое отверстие, 5—
половые клетки, 6—эпителизованный фагоцитобласт
Большое значение в эволюции потомков фагоцителлы имели также изменения характера движения: пассивное движение или прикрепленный образ жизни обусловливают лучевой тип симметрии, в то время как активное перемещение в определенном направлении предусматривает формирование двубоковой, или билатеральной, симметрии. В результате возникло огромное многообразие форм многоклеточных животных.
Представления о происхождении многоклеточных, изложенные здесь, являются развитием гипотезы И.И. Мечникова (1877—1880) о том, что многоклеточность возникла на базе шаровидных колоний жгутиковых, часть клеток которых иммигрировала внутрь для переваривания пищевых частиц, в результате чего оформилась дифференцировка на экто-
и энтодерму.
13.5.3. Основные этапы прогрессивной эволюции многоклеточных животных
Важным шагом в эволюции многоклеточных животных явилось возникновение третьего зародышевого листка — мезодермы.
Мезодерма обеспечивает возможность дифференцировки мышечной, соединительной тканей и скелета, а также многоклеточных половых желез, в которых созревающие гаметы оказываются надежно защищены от неблагоприятных средовых воздействий. Практически все трехслойные животные ведут активно подвижный образ жизни, благодаря чему приобретают билатеральный тип симметрии. Вместе с тем у трехслойных животных с интенсивным обменом веществ, активно перемещающихся с помощью мышц, возникают проблемы с выведением большого количества продуктов диссимиляции из тканей — производных мезодермы, в то время как эктодермальные и энтодермальные клетки выделяют их за счет диффузии соответственно либо наружу, либо в просвет пищеварительной полости. Поэтому именно у трехслойных впервые появляется и прогрессивно эволюционирует выделительная система.
Следующий значительный этап эволюции животных — возникновение вторичной полости тела,
или целома,
первоначально функционирующего как гидростатический скелет, а также выполняющего половую и выделительную функции в связи с тем, что продукты диссимиляции и половые клетки попадают в целом и только потом выделяются наружу (см. § 14.5).
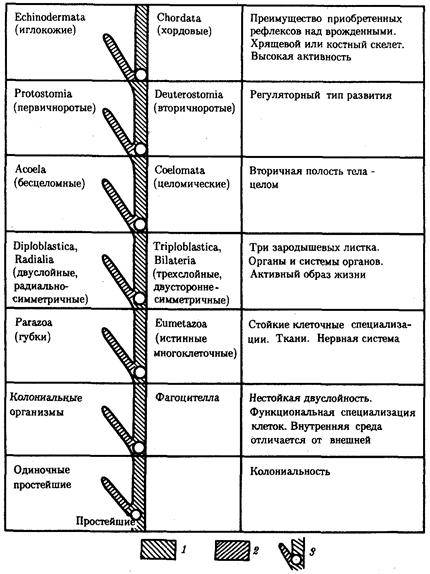
Рис. 13.12. Главные направления эволюции групп в животном царстве:
1
—прогрессивное направление, 2—
адаптивное направление, 3—
узловые моменты в прогрессивной эволюции; каждому узловому моменту соответствует его характеристика, обозначенная в правом столбце
Существенным этапом дальнейшей эволюции многоклеточных является возникновение регуляторного типа эмбрионального развития (см. разд. 8.3.1 и 8.3.2), в результате которого в развивающемся зародыше доминирует целостность морфогенетических процессов над их составляющими. Благодаря этому зародыш развивается относительно автономно в соответствии со своей генетической программой и способен компенсировать даже серьезные повреждения. Организмы, характеризующиеся такими особенностями, относят к группе вторичноротых,
в отличие от первичноротых,
у которых эмбриональное развитие протекает по мозаичному типу (см. разд. 8.3.1).
Наиболее крупные систематические группировки в царстве Животные называют типами.
За период существования жизни на Земле их было не менее 35. К настоящему времени некоторые из них вымерли; сейчас на Земле обитают животные 26 типов.
На рис. 13.12 приведена схема главных направлений эволюции в Животном царстве, а на рис. 13.13 изображены представители основных типов современных животных. Интересно, что к концу протерозойской эры (2,7 млрд. лет тому назад) на Земле уже существовали представители всех типов животного мира и основные узловые моменты в прогрессивной эволюции животных ими были пройдены.
Существование разнообразных живых организмов на Земле на протяжении около 3 млрд. лет, а также возникновение человека как биосоциального существа определяет в настоящее время картину современного органического мира.
Рис. 13.13. Основные типы животного царства и их филогенетические взаимоотношения
13.5.4. Характеристика типа Хордовые
Хордовые представляют собой наиболее высокоорганизованный тип в животном мире. Их общими чертами являются следующие.
1. Внутренний осевой скелет представлен хордой, которая присутствует в эмбриогенезе у всех представителей типа, а у высших дополняется, а затем и замещается позвоночником.
2. Над хордой располагается центральная нервная система в виде нервной трубки с полостью — невроцелем.
3. В боковых стенках глотки находятся жаберные щели, соединяющие ее полость с внешней средой. У рыб и некоторых земноводных они сохраняются в течение всей жизни, у высших хордовых — только в эмбриональном периоде.
4. Тело построено метамерно. У низших хордовых и у зародышей высших сегментация распростраянется на все системы органов, у высших ярко выражена только в эмбриональном периоде. Позже частично сохраняется только в опорно-двигательном аппарате, нервной и кровеносной системах.
5. Органами поддержания равновесия и движения являются конечности, причем у низших хордовых большее значение имеют непарные, а у высших — парные.
6. Общий план строения хордовых представлен на рис. 13.14. На спинной стороне расположена нервная трубка, под ней — хорда или заменяющий ее позвоночник. Глубже находится пищеварительная трубка с развивающейся из нее дыхательной системой, а под ней — вентральный пульсирующий кровеносный сосуд или сердце. По бокам от нервной трубки и хорды лежат сомиты, а по бокам от кишки — спланхнотомы, внутри которых расположен целом.
Рис. 13.14. Общий план строения хордового животного. А —
вид сбоку; Б —
поперечный срез:
1
—нервная трубка, 2—
хорда, 3—
плавники, 4—
сегменты тела, 5—пищеварительная трубка, 6—
пульсирующий сосуд или сердце. 7—жаберные щели в глотке, 8—
целом, 9—
сомит
13.5.5. Систематика типа Хордовые
Из четырех подтипов хордовых — Полухордовые Hemichordata, Личиночно-хордовые Urochordata, Бесчерепные Acrania и Позвоночные Vertebrata — остановимся на двух последних, имеющих отношение к прогрессивному направлению в эволюции этого типа животных.
Подтип Бесчерепные состоит лишь из одного класса — Головохордовые Cephalochordata, к которому относится ланцетник; подтип Позвоночные включает следующие классы: Круглоротые Cyclostomata, Хрящевые рыбы Chondrichthyes, Костные рыбы Osteichthyes, Земноводные Amphibia, Пресмыкающиеся Reptilia, Птицы Aves и Млекопитающие Mammalia.
13.5.6. Подтип Бесчерепные
Acrania
Характерной особенностью животных этого подтипа, представителем которого является ланцетникBranchiostoma lanceatum
(рис. 13.15), является малоподвижный придонный образ жизни и пассивное питание за счет фильтрации воды. Тело ланцетника полупрозрачно, покрыто однослойным цилиндрическим эпителием. Дерма развита слабо. Хорда тянется от головного до хвостового конца, а нервная трубка замкнута на спинной стороне неполно. В ней расположены светочувствительные глазки Гессе, обеспечивающие ланцетнику лишь восприятие света и темноты.
Около половины длины пищеварительной трубки составляет глотка, которая начинается позади ротовой полости и пронизана более чем 100 парами жаберных щелей, ведущих в околожаберную полость. Последняя открывается во внешнюю среду на брюшной стороне тела. Кишка в передней ее части снабжена печеночным выростом, гомологичным печени других хордовых, и заканчивается анальным отверстием. При поступлении воды через рот в глотку и далее в околожаберную полость осуществляется как отцеживание взвешенных в воде пищевых частиц, так и газообмен в кровеносных сосудах межжаберных перегородок.
Кровеносная система замкнута. Имеется один круг кровообращения, функцию сердца выполняет пульсирующая брюшная аорта.
Органы выделения — метамерно расположенные на межжаберных перегородках нефридии, состоящие из воронок, которые собирают продукты диссимиляции из целома и выводят их в околожаберную полость.
Метамерно организованы и половые железы, выделяющие гаметы по мере их созревания также в околожаберную полость и далее во внешнюю среду вне зависимости от наличия поблизости половозрелых организмов противоположного пола.
Данные сравнительной анатомии и эмбриологии позволяют сделать вывод о том, что предками бесчерепных были мелкие свободнопла-вающие двусторонне-симметричные существа с сегментированной мускулатурой и с небольшим количеством жаберных щелей, открывающихся наружу. Так же как и у ланцетника, питание их было пассивным и осуществлялось за счет фильтрации воды через жаберные щели, а половые продукты периодически выбрасывались в воду. Эти первичные бесчерепные дали начало двум ветвям в эволюции хордовых. Представители одной из них перешли к придонному образу жизни. От них произошли предки современных бесчерепных. Другая ветвь характеризовалась усилением двигательной активности. Арогенная эволюция в этой ветви хордовых привела к формированию подтипа Позвоночные.
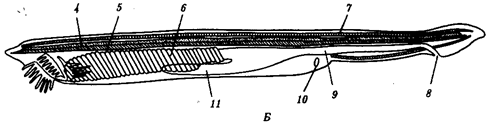
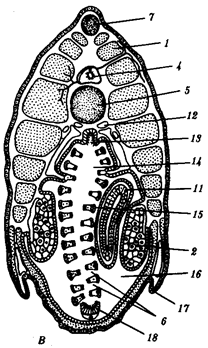
Рис. 13.15. Строение ланцетника. А—
общий вид; Б—
сагиттальный срез; В
—поперечный срез:
1
—миомер, 2—
гонада, 3—
щупальца предротовой воронки, 4—
спинной мозг, 5—
хорда, 6
—жаберные щели, 7—лучи (камеры) спинного плавника, 8—
анальное отверстие, 9—
кишечник,
10—
атриопор, 11
—печеночный вырост, 12—
корень спинной аорты, 13—
наджаберная бороздка, 14—
глотка, 15—
жаберная перегородка, 16—
атриальная полость, 17
—металлевральная складка, 18—
эндостиль
13.5.7. Подтип Позвоночные
Vertebrata
Основные особенности образа жизни позвоночных — их активные перемещения в пространстве, обеспечивающие им эффективное разыскивание пищи и представителей противоположного пола при размножении. Это достигается следующими ароморфными чертами их организации: усовершенствованный опорно-двигательный аппарат (сегментированный позвоночник взамен хорды); конечности с собственными мышцами и скелетом; дифференцировка головы с черепом, содержащим головной мозг (состоит из пяти отделов); развитые органы чувств. Интенсификации обмена веществ способствует выделение специализированных органов дыхания — жабр или легких, а также дифференцировка сердца из брюшного кровеносного сосуда. Выведение продуктов диссимиляции осуществляется специализированным компактным органом — почкой. Половые продукты выделяются гонадами позвоночных только в период размножения в момент встречи особей противоположных полов (рис. 13.16).
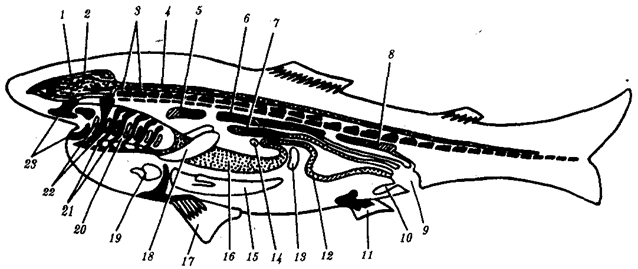
Рис. 13.16. Строение позвоночного челюстноротого животного:
1
—череп, 2—
головной мозг, 3—
позвоночник, 4—
спинной мозг, 5—
головная почка, 6—туловищная почка, 7—половая железа с протоком,8—
тазовая почка, 9—
клоака, 10—
мочевой пузырь, 11
—задняя парная конечность, 12—
кишка, 13 —
селезенка, 14—
поджелудочная железа, 15—
печень, 16
—желудок, 17—
передняя парная конечность, 18—
легкое, 19—
сердце, 20—
глотка, 21—
жаберные дуги, 22—
жаберные щели, 23—
челюсти
Семь перечисленных выше классов типа позвоночных фактически являются ступенями, соответствующими поэтапному повышению уровня организации в этом филогенетическом стволе эволюционного древа животного мира (рис. 13.17).
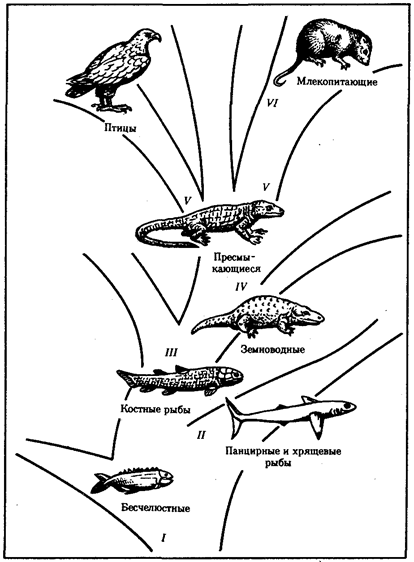
Рис. 13.17. Узловые моменты в прогрессивной эволюции хордовых:
I
—появление хрящевого скелета, дифференцировка центральной нервной системы на головной и спинной мозг, II—
появление челюстей, III
—появление парных конечностей наземного типа и легких, IV—
преодоление барьера влажности, V—
теплокровность, VI—
внутриутробное развитие
Класс Круглоротые Cyclostomata — наиболее архаичный среди позвоночных. Представители его смогли дожить до настоящего времени в основном благодаря переходу к паразитическому образу жизни. Представителями являются миноги. Главная их особенность — неподвижность рта, которая обусловлена особым строением жаберных дуг, выполняющих только функцию опоры для глотки. Имеются непарные спинные и хвостовые плавники.
Активизация питания — активный захват пищи —
важнейшее ароморфное приобретение надкласса Рыбы Pisces. Благодаря этому в отличие от бесчелюстных круглоротых рыбы и все более высокоорганизованные позвоночные относятся к группе Челюстноротые Gnathostomata. Челюсти возникли в результате преобразования передних жаберных дуг (см. разд. 14.2.1). Кроме того, у рыб прогрессивно развиваются парные конечности с собственным скелетом, а у наиболее высокоорганизованных из них — и плавательный пузырь, выполняющий гидростатические функции.
Следующий этап в прогрессивной эволюции позвоночных — выход на сушу,
обеспечивающийся дифференцировкой четырех парных конечностей и утратой значения хвостового плавника как органа движения у земноводных. В связи с этим земноводные вместе с более прогрессивными позвоночными объединяются в группу Четвероногие Tetrapoda. Выход на сушу сопровождается преобразованием плавательного пузыря в легкие и появлением в связи с этим двух кругов кровообращения и трехкамерного сердца.
Пресмыкающиеся Reptilia успешно преодолели барьер влажности. Они имеют сухую кожу, предохраняющую организм от высыхания, и стали независимыми от водной среды даже при размножении, приобретя способность к внутреннему оплодотворению и откладке яиц с плотными оболочками и запасами жидкости. Амниотическая оболочка их зародышей, формирующаяся в ходе эмбриогенеза, создает благоприятные условия для их развития вне зависимости от влажности окружающей среды. Поэтому пресмыкающиеся вместе с птицами и млекопитающими относятся к группе Амниоты Amniota, или Первичноназемные животные.
Ключевой адаптацией птиц и млекопитающих является их теплокровность,
или гомойотермность,
обеспечивающая резкий биологический прогресс этих классов благодаря независимости от климатических факторов среды. Эти классы объединяются в группу Гомойотермные.
Наконец, Плацентарные млекопитающие Placentalia характеризуются внутриутробным эмбриональным развитием, в еще меньшей степени, чем у пресмыкающихся, зависящим от окружающей среды. Выкармливание потомства молоком значительно повышает шансы на выживание.
Кроме того, преимущественное развитие в центральной нервной системе коры больших полушарий
переднего мозга обеспечивает кардинальное усложнение поведения на фоне превалирования условных рефлексов над безусловными и формирования сложных инстинктов. Биологическая организация наиболее прогрессивного отряда млекопитающих — Приматы Primates — явилась предпосылкой происхождения человека именно в рамках этого отряда.
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
Покровы
любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное — экто- и мезодермальное — происхождение. Из эктодермы развивается эпидермис, из мезодермы — дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях — дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной.
Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных — стегоцефалов — соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез,
секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см.разд. 23.1).
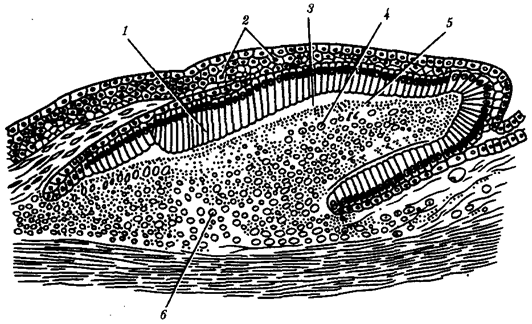
Рис. 14.1. Закладка плакоидной чешуи:
1
—клетки-эмалеобразователи, 2—
эпидермис, 3—
эмаль, 4—
склеробласты—дентинообразователи, 5—
дентин, 6—
сосочек дермы
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает.
Роговые чешуйки у некоторых рептилий тонки и эластичны, у других—сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие — насекомоядные, грызуны и некоторые другие — наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3—7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы
многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы
, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых — разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.

Рис. 14.2. Расположение волос у млекопитающих. А—на хвосте грызунов; Б—
на других участках тела:
1
—роговые чешуйки, 2—
группы волос, расположенные в шахматном порядке
|
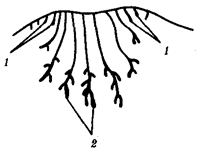
Рис. 14.3. Строение развивающегося соска млекопитающего
Виден постепенный переход от потовых (1
) к млечным (2
) железам
|
Потовые железы
млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы.
У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна (рис. 14.4).
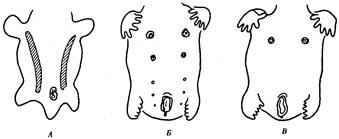
Рис. 14.4. Эмбриогенез передней брюшной стенки человека. А —
зародыш в возрасте 5 недель (видны млечные линии); Б —
дифференцировка пяти пар сосков; В —
зародыш в возрасте 7 недель
Рис. 14.5. Атавистические аномалии развития кожи.
А —
гипертрихоз; Б —
полимастия
Сальные железы
образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата,
претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
14.2.1. Скелет
У хордовых скелет внутренний.
По строению и функциям подразделяется на осевой, скелет конечностей и головы.
В подтипе Бесчерепные имеется только осевой скелет
в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей — хордом,
возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками,
развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом.
Это один из выраженных примеров гомотопной субституции органов (см. § 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).
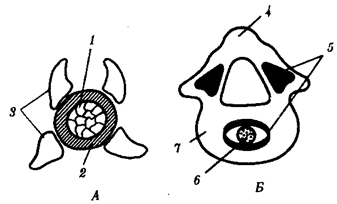
Рис. 14.6. Развитие позвонка. А—ранний этап; Б—
последующая стадия:
1
—хорда, 2—
оболочка хорды, 3—
верхние и нижние позвонковые дуги, 4—
остистый отросток, 5—
зоны окостенения, 6—рудимент хорды, 7—
хрящевое тело позвонка
Замещение хорды позвоночным столбом — более мощным органом опоры, имеющим сегментарное строение,— позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией — заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный
и хвостовой.
Это связано с перемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный
и крестцовый
отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй — опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный
отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5—10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,— у хвостатых земноводных и пресмыкающихся , поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12—13 пар ребер.
Рис. 14.7. Аномалии развития осевого скелета. А —
рудиментарные шейные ребра (показаны стрелками); Б —
несращение остистых отростков позвонков в грудной и поясничной областях. Спинномозговые грыжи
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida —дефект позвоночного канала.
При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа
(рис. 14.7).
В возрасте 1,5—3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8—11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
Продолжением осевого скелета спереди является осевой,
или мозговой, череп,
служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный,
или лицевой череп,
образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую
и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он
выполняет функцию первичной верхней челюсти. Нижний, или меккелев,
хрящ — первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного
хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным
(рис. 14.9).
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным.
Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка — столбик,—
характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко,
а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку
и молоточек.
Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).
Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных.
А—
хрящевая рыба; Б—
земноводное; В—
пресмыкающееся; Г—
млекопитающее:
1
—нёбно-квадратный хрящ, 2—меккелев хрящ, 3—
гиомандибулярный хрящ, 4—гиоид, 5—
столбик, 6—
накладные кости вторичных челюстей, 7—наковаленка, 8—
стремечко, 9—
молоточек; гомологичные образования обозначены соответствующей штриховкой
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки — столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой' и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности — грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей.
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными
(рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль — стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.
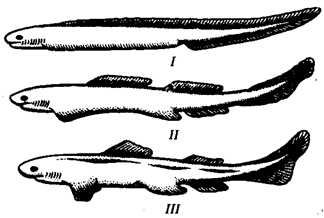
Рис. 14.10. Формирование передних и задних конечностей из метаплевральныхскладок: I
—III
—гипотетические этапы эволюции
В результате из передних отделов складок развились грудные, а из задних — брюшные плавники (рис. 14.10). Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками (рис. 14.11). За счет слияния оснований хрящевых лучей возникли плечевой
и тазовый пояса. Остальные
их участки дифференцировались в скелет свободных конечностей.

Рис. 14.11. Древняя акулообразная рыба с многочисленными парными конечностями
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7—12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными (рис. 14.12, А, Б
) и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания (см. разд. 14.3.4) стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители — стегоцефалы — обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб (рис. 14.12, B
)
Рис. 14.12. Скелет конечности кистеперой рыбы (А
),
его основание (Б
) и скелет передней лапы стегоцефала (В
): I—
плечевая кость, 2—локтевая кость, 3—
лучевая кость
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3—4 ряда, в пястье располагается 7—5 костей, а далее также радиально лежат фаланги 7—5 пальцев.
У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух — у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду — и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно (рис. 14.13). Однако в процессе онтогенеза у большинства наземных позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять (рис. 14.14).
Рис. 14.13. Скелет передней конечности наземных позвоночных. А
—лягушка- Б
—саламандра; В
—крокодил; Г
—летучая мышь; Д
—человек: 1
—плечевая кость, 2—лучевая кость, 3
—кости запястья, 4
—пясти, 5
—фаланги пальцев, 6
—локтевая кость
Рис. 14.14. Строение развивающейся конечности позвоночного: рр — prepollex, pin — postminimus — дополнительные рудиментарные I и VII пальцы
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3—4-го шейных позвонков, а нижние — на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих отделов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой — с конечностями, переместившимися на новое место (рис. 14.15; см. также разд. 14.2.2.2).
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Так, полидактилия,
или увеличение количества пальцев, наследующаяся как аутосомно-доминантный признак, является результатом развития закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Известен феномен полифалангии, характеризующийся увеличением числа фаланг обычно большого пальца кисти. В основе его возникновения — развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
Серьезным пороком развития является нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1—2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля
или врожденным высоким стоянием лопатки (рис. 14.16). Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем что такое нарушение часто сопровождается аномалиями рёбер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения — не только нарушение перемещения органов, но и обусловленное этим нарушение морфогенетических корреляций (см. § 13.4).
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопитающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его прямохождением и трудовой деятельностью. К ним относят: 1) изменения стопы переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению еесводов, служащих для амортизации при ходьбе; 2) изменения позвоночного столба — его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении; 3) изменения черепа — резкое уменьшение его лицевой части и увеличение мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка;4) специализация верхних (передних) конечностей как органа труда; 5) появление подбородочного выступа в связи с развитием членораздельной речи.
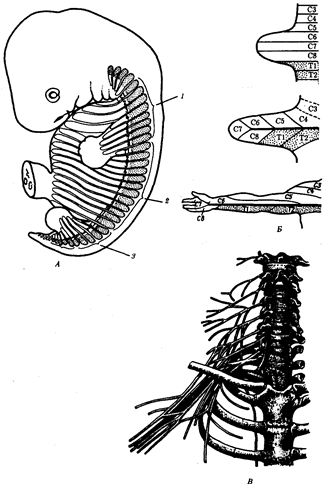
Рис. 14.15. Формирование передних конечностей, их гетеротопия и иннервация в онтогенезе человека. А
—врастание шейных миотомов в формирующуюся переднею конечность зародыша; Б
—развитие кожной иннервации руки; В
— расположение шейного и плечевого сплетений, участвующих в иннервации руки:
1
—шейные миотомы, 2—
грудные миотомы, 3
—поясничные миотомы; буквами С, Т, L обозначены шейные, грудные и поясничные сегменты
Рис. 14.16. Болезнь Шпренгеля (пояснение см.в тексте)
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии
скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
14.2.2. Мышечная система
У представителей типа Хордовые мускулатура подразделяется по характеру развития и иннервации на соматическую и висцеральную.
Соматическая мускулатура
развивается из миотомов и иннервируется нервами, волокна которых выходят из спинного мозга в составе брюшных корешков спинномозговых нервов. Висцеральная мускулатура
развивается из других участков мезодермы и иннервируется нервами вегетативной нервной системы. Вся соматическая мускулатура поперечно-полосата, а висцеральная может быть как поперечно-полосатой, так и гладкой (рис. 14.17).
Рис. 14.17. Соматическая и висцеральная мускулатура позвоночных:
1
—соматическая мускулатура, развивающаяся из миотомов, 2—
висцеральная мускулатура жаберной области
Наиболее существенные изменения претерпела висцеральная мускулатура, связанная с висцеральными дугами передней части пищеварительной трубки. У низших позвоночных большая часть этой мускулатуры представлена общим сжимателем висцерального аппарата — m. constrictor superficialis, покрывающим всю область жаберных дуг со всех сторон. В области челюстной дуги эта мышца иннервируется тройничным нервом
(V), в области подъязычной дуги — лицевым
(VII), в области первой жаберной дуги — языкоглоточным
(IX), наконец, ее часть, лежащая каудальнее,— блуждающим
нервом (X). В связи с этим все производные соответствующих висцеральных дуг и мышц, связанных с ними, иннервируются впоследствии у всех позвоночных перечисленными нервами.
В передней части сжимателя обособляется крупная мускулатурная масса, обслуживающая челюстной аппарат. Позади висцерального аппарата дифференцируется трапециевидная мышца m. trapezius, отдельными пучками прикрепляющаяся к последним жаберным щелям и переднему краю спинного отдела плечевого пояса. Часть поверхностного сжимателя в области подъязычной дуги у пресмыкающихся разрастается, охватывает шею снизу и с боков и образует сжиматель шеи m. sphincter colli. У млекопитающих эта мышца делится на два слоя: глубокий и поверхностный. Глубокий сохраняет прежнее название, а поверхностный называется platysma myoides и располагается подкожно. Эти две мышцы разрастаются на всю область головы и дают начало сложной системе лицевой подкожной мускулатуры, которую у приматов и человека называют мимической.
Поэтому вся мимическая мускулатура иннервируется так же, как и мышца, из которой она происходит,— лицевым нервом.
Мышцы собственно жаберного аппарата с утратой жаберного дыхания редуцируются, но отдельные их элементы сохраняются в виде мышц подъязычного аппарата, глотки и гортани. Трапециевидная мышца полностью теряет связь с жаберным аппаратом и становится исключительно мышцей плечевого пояса. Часть ее у млекопитающих отходит от сосцевидного отростка черепа и приклепляется к ключице и грудине, обособляется — грудино-ключично-сосцевидная мышца т. sternocleidomastoideus. Иннервирующая эту мышцу задняя часть блуждающего нерва становится самостоятельным черепно-мозговым нервом XI пары, добавочным —
п. accessorius.
Основные этапы филогенеза висцеральной мускулатуры жаберной области рекапитулируют в эмбриогенезе млекопитающих и человека. Знание этих рекапитуляции позволяет объяснить сложность иннервации мышц лица и шеи, объединенных с ними общностью происхождения.
Мускулатура головы.
У всех позвоночных животных в процессе эмбриогенеза нижние концы миотомов образуют выросты в вентральном направлений, охватывающие полость тела снаружи и срастающиеся по центральной линии на брюшной стороне. Таким образом, закладки соматической мускулатуры становятся сегментарными не только на дорсальной стороне в связи с сегментацией сомитов, но и на вентральной. В миотомах и в их вентральных отростках идет образование продольных мускульных волокон.
Миотомы, лежащие на головном конце тела, распадаются на мезенхиму и образуют зачатки отдельных мышц. Из первого миотома головы формируются верхняя внутренняя и нижняя прямые и нижняя косая мышцы глаза, иннервируемые глазодвигательнъш нервом
п. oculomotorius (III пара). Из второго миотома — верхняя косая мышца, иннервируемая блоковым нервом
п. trochlearis (IV пара); а из третьего — наружная прямая мышца, получающая иннервацию от отводящего нерва
п. abducens (VI пара).
Задние миотомы головы, образующие мощные вентральные отростки, распространяющиеся вокруг висцеральной мускулатуры глоточной области, образуют подъязычную мускулатуру, которая у рыб слабо дифференцирована, а начиная с земноводных распадается на m. sternohyoideus, т. omohyoideus и т. geniohyoideus. У наземных позвоночных за счет последней мышцы формируется собственная мускулатура языка — m. genioglossus и m. hyoglossus. Вся подъязычная мускулатура иннервируется подъязычным нервом
п. hypoglossus, который у амниот становится типичным черепно-мозговым нервом.
Мускулатура туловища и конечностей.
У бесчерепных, а также у рыб вся мускулатура туловища состоит из ряда мышечных сегментов, или миомеров, правой и левой сторон, которые вместе образуют так называемые боковые мышцы.
Каждый миомер развивается из миотома одного сомита и иннервируется первоначально двигательной ветвью одного спинномозгового нерва. Миомеры отделены друг от друга миосептами — соединительнотканными перегородками. Такой же перегородкой, идущей продольно, бокойая мышца разделена на спинную и брюшную мышцы.
Уже у рыб направление пучков мышечных волокон в миомерах начинает изменяться на разной глубине мышечного слоя. Эта дифференцировка значительно более выражена у наземных позвоночных и приводит у них к постепенному обособлению различных слоев брюшной и спинной мускулатуры. В результате этого возникают сложные группы мышц, от первоначальной четкой метамерии которых остаются только следы в виде глубоких мышц спины и шеи, связывающих друг с другом соседние позвонки. Значение спинной мускулатуры наземных позвоночных снижается в связи с передвижением большинства из них с помощью конечностей, а брюшные мышцы претерпевают смену функций: первоначально принимающие участие в перемещениях тела, они у пресмыкающихся и млекопитающих служат для изменения объема грудной и брюшной полостей в процессе дыхания.
Мускулатура парных плавников рыб закладывается в виде ряда мускульных почек, вырастающих от вентральных концов миотомов. Каждая из этих почек подразделяется на два мышечных зачатка, врастающих в основание закладки плавника с его спинной и брюшной сторон. Первые функционально становятся мышцами, отводящими плавник, вторые — приводящими. У наземных позвоночных из мышечного зачатка, гомологичного отводящей мышце плавника, развивается группа разгибателей пятипалой конечности, а из зачатка ее антагонистов — группа сгибателей. В пределах каждой группы идет дифференцировка на поверхностные и глубокие мышечные пучки, становящиеся самостоятельными мышцами. В целом мышцы наземных позвоночных, гомологичные мышцам плавников рыб, образуют первичную мускулатуру конечностей.
Она иннервируется нервами плечевого и пояснично-крестцового сплетений, образованных в процессе перемещения поясов конечностей в ходе эмбриогенеза (см. разд. 14.2.1).
При дальнейшей дифференцировке миотомов грудной клетки развивается группа мышц, приводящая в движение сам плечевой пояс, или вторичная мускулатура.
К ней относят широчайшую мышцу спины, большую и малую грудные, а также зубчатую мышцы. Они иннервируются непосредственно спинномозговыми нервами сегментов спинного мозга, расположенных каудальнее тех его участков, которые осуществляют иннервацию первичной мускулатуры. Задняя пара конечностей не имеет вторичной мускулатуры в связи с тем, что гетеротопия тазового пояса по отношению к позвоночнику в процессе эволюции менее выражена.
Изменение среды обитания и характера движений позвоночных привело к усилению и обособлению большого числа мышц, обслуживающих конечности, и к относительной редукции собственно мышц туловища. Такие мышцы, как грудные, широчайшая мышца спины и трапециевидная, в значительной мере покрывают туловищную мускулатуру и даже частично вытесняют ее функционально.
Уникальной особенностью организации хордовых является филогенетическая, эмбриональная, а также функциональная связь пищеварительной и дыхательной систем.
Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка — передняя часть пищеварительной трубки, пронизанная сквозными отверстиями — жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источником его питания является взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки.
У более высокоорганизованных хордовых начиная с рыб пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка.
Рис. 14.18. Кишечная трубка позвоночных (А
) и ее дифференцировка в эмбриогенезе человека (Б
):
а—
жаберные щели в глотке;1
—ротовая полость, 2—
1—5-й глоточные карманы, 3—
глотка, 4—
пищевод, 5—
желудок, 6—
двенадцатиперстная кишка, 7—дорсальная поджелудочная железа, 8—
вентральная поджелудочная железа, 9—
тонкая кишка. 10
—клоака, 11—
мочевой пузырь, 12—
желточный мешок, 13—
желчный пузырь, 14—
печень,15—
легкие,16
—закладка щитовидной железы
Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе — морфогенетическими и эргонтическими корреляциями. Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют stomodeum. Слизистая оболочка, выстилающая этот участок, эктодермального происхождения и в развитии связана с кожным эпидермисом и его производными (см. § 14.1). Средняя часть кишки начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum (рис. 14.18).
14.3.1. Ротовая полость
Рассмотрим вначале эволюцию ротовой полости
и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком.
Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом.
Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо,
полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания (рис. 14.19, А— В
).
До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть
(рис. 14.19, Г
). Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений.
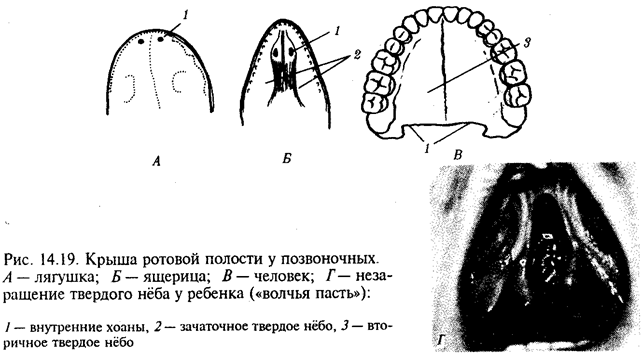
Зубы
позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб (см. § 14.1). У них наблюдается непрерывный переход от типичных чешуи к зубам в ротовой полости (рис. 14.20). Зубы первоначально расположены во много рядов и покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем, так же как у земноводных и рыб, дифференцировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной.
У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми их поколениями. Многократную смену зубов называют полифиодонтизмом.
Рис. 14.20. Переход от плакоидной чешуи к зубам по краю ротового отверстия акулы: 1
— плакоидная чешуя, 2—
закладка новых зубов, 3—
эмаль, 4—
дентин, 5—
зубы
Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной.
Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они вознили у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее количество зубов у млекопитающих уменьшается и достигает у высших приматов 32. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень.
Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и коренные. Это явление называют дифиодонтизмом.
У человека возможны атавистические аномалии зубной системы,
связанные с нарушениями как дифференцировки зубов, так и с их количеством. Редкой аномалией является гомодонтная зубная система, в которой все зубы имеют коническую форму. Более частой патологией является трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе (рис. 14. 21). Это свидетельствует о том, что у человека возможно образование большего количества зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельством тенденции к дальнейшему уменьшению количества зубов у человека является то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно — до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы.

Рис. 14.21. Сверхкомплектные зубы, прорезавшиеся у человека (указаны стрелками)
На дне ротовой полости позвоночных располагается непарный выступ — язык,
который у рыб представляет собой складку слизистой оболочки, лишенную мышц. Его движения осуществляются вместе с челюстями и подъязычной костью. У наземных позвоночных в языке располагаются мышцы, и они определяют его собственную подвижность. У пресмыкающихся и млекопитающих язык закладывается из трех зачатков: одного непарного и двух парных, лежащих кпереди от первого. Парные зачатки позже срастаются и дают начало телу языка. У большинства рептилий это срастание неполное, и язык раздвоен на конце. Весьма редким пороком развития языка у человека является раздвоенность его конца как результат несращения парных зачатков в эмбриогенезе.
Слюнные железы
у рыб, заглатывающих пищу вместе с водой, отсутствуют и появляются только при переходе на сушу. Земноводные, обитающие на земле, имеют несколько слюнных желез, выделяющих только слизь. Слюна пресмыкающихся содержит уже и пищеварительные ферменты, а у некоторых змей—и токсины (см. §23.1). У млекопитающих слюнные железы многочисленны: это и мелкие — зубные, щечные, нёбные, язычные, гомологичные железам земноводных и пресмыкающихся, и крупные — подъязычные, подчелюстные и околоушные. Подъязычная и подчелюстная железы — результат дифференцировки подъязычной железы пресмыкающихся, околоушная — новое приобретение млекопитающих.
14.3.2. Глотка
Глотка — орган, выполняющий у всех хордовых две функции: дыхательную и пищеварительную. У ланцетника она пронизана большим количеством жаберных щелей (более 150 пар). У рыб жаберные щели в количестве 5—7 закладываются как слепые парные выросты глотки — жаберные мешки. Навстречу им выпячиваются кожные покровы — жаберные карманы. В месте их соприкосновения происходит прорыв тканей эктодермы кожи и энтодермы глотки и возникают сквозные жаберные щели.
У личинок земноводных в глотке образуется четыре пары жаберных щелей (рис. 14.22), а у пресмыкающихся прорыв их происходит только в эмбриональном развитии; вскоре после этого они зарастают. У млекопитающих в эмбриогенезе начинается закладка глоточных мешков и жаберных карманов, которые в норме никогда не прорываются и не образуют жаберных щелей (рис. 14.23).
При нарушении эмбриогенеза во время закладки зачатков жаберных щелей они могут прорываться и даже сохраняться в постэмбриональном периоде у млекопитающих и человека. Эту аномалию называют латеральными свищами шеи.
Они открываются на коже боковой поверхности шеи, а другим концом впадают в глотку. Чаще встречается феномен под названием латеральные кисты шеи.
Эта аномалия связана с сохранением в постнатальном развитии эмбрионального материала глоточных жаберных мешков или эктодермальных жаберных карманов. Участки этих образований могут заполняться жидкостью, увеличиваться в размерах и склонны к малигнизации. Необходимы своевременная диагностика этих образований и их хирургическое удаление.
Первая жаберная щель у всех наземных позвоночных начиная с земноводных в процессе формирования превращается в евстахиеву трубу, барабанную полость и наружный слуховой проход. Располагаясь топографически в зоне редукции первичных челюстей и подъязычной висцеральной дуги, преобразующихся в слуховые косточки, она меняет свои функции, включаясь в систему слухового анализатора (см. разд. 14.2.1).
Рис. 14.22. Закладка жаберных щелей у позвоночных. А—
фронтальный срез личинки тритона; Б—
срез через глотку зародыша мыши (головной конец зародыша сильно изогнут, поэтому на фотографии видны два среза одной нервной трубки):
1—5—
жаберные карманы и жаберные щели, 6—
зачаток легкого, 7—полость глотки, 8—
нервная трубка
14.3.3. Средняя и задняя кишка
Кишечная трубка претерпевает в ходе филогенеза следующие прогрессивные преобразования: увеличение общей длины, дифференцировку на отделы и образование крупных многоклеточных желез.
Если длина этого отдела пищеварительной трубки ланцетника составляет примерно 1/3 длины тела, то у млекопитающих возможно превышение ее длины по отношению к телу в 10 раз и более. Этим достигается удлинение времени контакта пищевых частиц с ферментами пищеварения и увеличение всасывающей поверхности. На органном уровне это сопровождается появлением складчатости слизистой оболочки кишки, возникновением ворсинок и крипт. Кишечник ланцетника абсолютно не дифференцирован. В средней
его части расположен печеночный вырост — простое слепое выпячивание брюшной стенки кишки, выстланное железистыми клетками. Функционально этот вырост представляет собой зачаточную печень.
Он является единственной многоклеточной железой пищеварительной системы ланцетника.
Рис. 14.23. Развитие области глотки и ее аномалии у человека.
А—
зародыш в возрасте 5 недель; Б—
тот же зародыш (срез через область глотки по пунктирной линии, изображенной на рис. А
); В—
схема латеральных свищей шеи; Г—
проекции наиболее часто встречающихся латеральных шейных свищей:
I
—IV
—жаберные дуги; 1
—кожа, 2—
сонная артерия, 3—
глотка, 4—
нёбные миндалины, 5—
подъязычная кость, 6—
гортань, 7—сквозной шейный свищ,8—10—
несквозные свищи
У рыб за глоткой следует короткий пищевод, затем желудок, слабо от него отграниченный. В кишечнике выделяются тонкий
и толстый
отделы, последний открывается анусом во внешнюю среду. В отличие от ланцетника у рыб хорошо развита печень, снабженная желчным пузырем. Поджелудочная железа у различных рыб построена по-разному. В некоторых случаях она представлена отдельными мелкими дольками в стенке кишечника, в мезентерии, рассеяна в ткани печени. Нередко уже у рыб она представляет собой компактный орган, включающий в себя как экзокринную часть, которая ответственна за синтез пищеварительных ферментов, так и эндокринную, которая выделяет гормоны, регулирующие углеводный обмен. Таким образом, у рыб наблюдаются все основные стадии эволюции сложной многоклеточной железы, структура и функции которой впоследствии принципиально практически не изменяются.
Единственное отличие пищеварительной трубки земноводных от трубки рыб — ее удлинение и впадение толстой кишки в клоаку.
Существенной особенностью пищеварительной трубки пресмыкающихся является возникновение в ней слепой кишки.
У большинства из них она зачаточна, но у некоторых развита весьма существенно. Слепая кишка — важное эволюционное приобретение пресмыкающихся, позволяющее им расширить рацион питания и использовать растительную пищу, переваривание которой сложно и требует участия симбиотических простейших и бактерий. Слепая кишка особенно богата микрофлорой, под действием которой осуществляются процессы брожения, позволяющие наиболее полно использовать пластические вещества растительного происхождения.
У млекопитающих за счет дальнейшей дифференцировки пищеварительного тракта, и в частности больших размеров слепой кишки, резко увеличивается разнообразие используемых продуктов питания и возникает пищевая специализация на травоядность и плотоядность; всеядность встречается редко, в частности в отряде приматов.
Задняя кишка
плацентарных млекопитающих дифференцирована, клоака отсутствует и прямая кишка заканчивается анусом.
В онтогенезе человека рекапитулируют основные стадии филогенеза кишечной трубки и ее производных. Из пороков развития, связанных с задержкой эмбриогенеза, известны гипоплазии всей пищеварительной системы, укорочение кишки и недоразвитие любых ее отделов, а также печени и поджелудочной железы. В зависимости от степени недоразвития тяжесть аномалии может быть как незначительной, так и несовместимой с жизнью.
Клинический интерес представляет гетеротопия тканей поджелудочной железы
в стенке тонкого кишечника или желудка. Иногда гетеротопированные фрагменты могут симулировать опухолевую трансформацию слизистой оболочки. По данным патологоанатомов, частота этой аномалии — от 0,6 до 5%. Механизм гетеротопии — нарушение клеточной миграции зачатков железы из стенки кишечной трубки. С недоразвитием и задержкой дифференцировки связана и такая аномалия, как персистирование клоаки,
при которой мочеполовые пути и прямая кишка объединены. При нормальном развитии после 8-й недели эмбриогенеза клоака должна полностью дифференцироваться на прямую кишку, мочевыделительные и половые протоки.
14.3.4. Органы дыхания
Жабры —
наиболее ранние специализированные органы дыхания, появляющиеся среди хордовых впервые у рыб. Они представляют собой тонкие складки слизистой оболочки глотки, лежащие на жаберных дугах, снабжаемые венозной кровью через жаберные артерии и распадающиеся здесь на капилляры.
Позади последней жаберной дуги у кистеперых рыб за счет выпячивания вентральной стороны глотки формируется парное образование — плавательный пузырь,
который выполняет в первую очередь гидростатические функции, уравновешивая тело рыбы в толще воды. Связь между ним и глоткой, имеющаяся в эмбриогенезе, не прерывается и позже, поэтому воздух при попадании в глотку может беспрепятственно проходить в плавательный пузырь. Эта морфофизиологи-ческая особенность при изменении условий существования кистеперых рыб явилась предпосылкой преобразования плавательного пузыря в легкие (рис. 14.24).
Рис. 14.24. Плавательный пузырь рыб (А—
костной; Б
—кистеперой) и развитие легких у человека (В—
ранние стадии): 1
—средняя кишка, 2—
плавательный пузырь, 3—
глотка, 4—развивающиеся легкие
У земноводных, связанных по происхождению с кистеперыми рыбами, в личиночном состоянии функционируют жабры, а во взрослом — органами дыхания становятся легкие.
В связи с отсутствием грудной клетки и диафрагмы воздух в них попадает из ротовой полости за счет глотательных движений, осуществляющихся подбородочно-подъязычной мышцей. Материал жаберных дуг, следующих за подъязычной дугой, частично редуцируясь, входит в состав хрящей гортани,
которая, появляясь впервые у земноводных, является первым органом, относящимся к нижним дыхательным путям. Легкие начинаются непосредственно от гортани. Они крупноячеисты и имеют относительно малую дыхательную поверхность, в связи с чем газообмен в большей степени осуществляется через кожные покровы (см. § 14.1).
Пресмыкающиеся, вышедшие на сушу окончательно, имеют как верхние дыхательные пути (не полностью отграниченную от ротовой полости полость носа), так и нижние — гортань, трахею
и бронхи.
Их легкие мелкоячеисты, содержат многочисленные внутренние перегородки и обладают большой дыхательной поверхностью. Впервые появляется диафрагма, которая в дыхании принимает скорее пассивное участие, так как она либо лишена мышечных элементов, либо разделяет грудную и брюшную полости не полностью. Механизм дыхания основан на сокращении межреберных мышц, приводящих в движение грудную клетку.
У млекопитающих дыхательные пути, как и у предков, выстланы мерцательным эпителием. Они полностью отделены от пищеварительной системы и только перекрещиваются с ней в глотке. Бронхи многократно разветвляются, вплоть до бронхиол, ведущих в альвеолы — легочные пузырьки, имеющие в сумме огромную площадь поверхности (у человека до 90 м ). Основной мышцей, изменяющей объем грудной полости, становится диафрагма.
В эмбриогенезе человека отражается первоначальное единство пищеварительной и дыхательной систем. На этом основано формирование большой группы врожденных пороков развития пищевода и трахеи
типа эзофаготрахеальных свищей
(рис. 14.25). Нарушением альвеолярной дифференцировки легочной ткани могут быть объяснены дизонтогенетические бронхолегочные кисты —
округлые полости в легких, отграниченные от окружающих тканей примитивно построенной стенкой недифференцированного бронха, а также кистозная гипоплазия
(недоразвитие) легких (рис. 14.26). При этой патологии недоразвитыми оказываются целые доли легкого, которые представляют собой многочисленные полости, связанные с крупными бронхами и имеющие малую поверхность. Газообмен в таких участках легких резко ослаблен. Широко известны также гипоплазии диафрагмы от небольших дефектов в ее куполе до полной аплазии. Последняя аномалия несовместима с жизнью и встречается обычно вместе с другими множественными пороками развития.

Рис. 14.25. Различные формы эзофаготрахеальных свищей (А—Г
):
1—
пищевод, 2—
трахея
Необходимым условием существования высокоорганизованных крупных многоклеточных организмов является наличие жидкой подвижной внутренней среды,
которая обеспечивает интеграцию организма в целостную систему, выполняя транспортные функции. Эти функции являются основными для кровеносной системы. Конкретная функция кровеносной системы зависит от того, что она транспортирует: питательные вещества, кислород, углекислый газ, другие продукты диссимиляции или гормоны. Кровеносная система всех хордовых замкнутая и состоит из двух основных артериальных сосудов: брюшной
и спинной аорт.
По брюшной аорте венозная кровь продвигается кпереди, обогащается кислородом в органах дыхания, а по спинной — кзади. Из спинной аорты кровь через систему капилляров возвращается по венам в брюшную аорту. Брюшная аорта или ее часть, периодически сокращаясь, проталкивает кровь по сосудам (рис. 14.27).
Рис. 14.26. Кистозная гипоплазия легкого
Рис. 14.27. Строение кровеносной системы водных (А
) и наземных (Б
) позвоночных:
1
—жаберные артерии, 2—сонная артериям—передняя кардинальная вена, 4—
задняя кардинальная вена,5—
спинная аорта, 6—
кювьеров проток, 7—подкишечная вена, 8—
печеночная вена, 9—брюшная аорта, 10—
задняя (нижняя) полая вена, 11
—воротная вена печени, 12—
легочная вена, 13—
легочная артерия; стрелками указано направление движения крови
14.4.1. Эволюция общего плана строения
кровеносной системы хордовых
У ланцетника кровеносная система наиболее проста (рис. 14.28, А
).Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, которые по количеству соответствуют числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к головному мозгу, так и назад. Передние ветви этих двух сосудов являются сонными артериями.
На уровне заднего конца глотки задние ветви образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние
или задние кардинальные вены,
расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток.
Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени
в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд — печеночную вену,
по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта,
становящаяся позже собственно аортой, и сонные артерии.
Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами,
правый кювьеров проток преобразуется в верхнюю полую вену,
а левый, сильно редуцировавшись, — в коронарный синус сердца.
Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Более активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг в конечном счете до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце,
состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника (рис. 14.28, Б
).
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное,
но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд — артериальный конус,
разветвляющийся последовательно на три пары сосудов:
кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое— малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами,
а кювьеровы потоки, в которые они впадают вместе с подключичными венами, — передними полыми венами
(рис. 14.28, В
).
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а из правой — легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных (рис. 14.28, Г
).
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению
венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца
и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены — рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими (рис. 14.28, Д
).
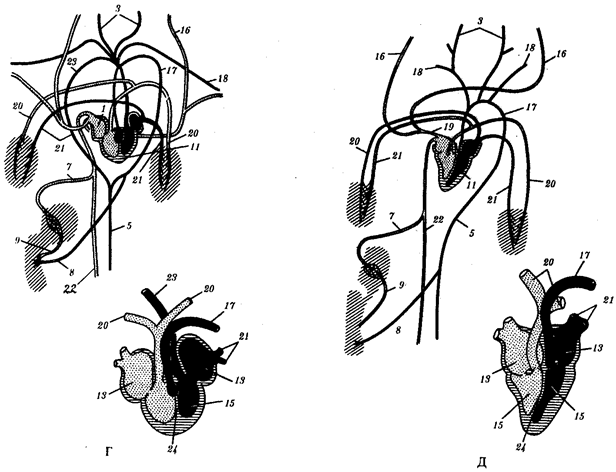
Рис 14.28. Строение кровеносной системы и сердца хордовых животных.
А—
ланцетника; Б—
рыбы; В—
земноводного; Г—
пресмыкающегося; Д—млекопитающего:
1
—брюшная аорта, 2—
жаберные артерии, 3—
сонная артерия, 4
—передняя кардинальная вена,5
—
спинная аорта, 6—
задняя кардинальная вена, 7— печеночная вена, 8—
чревная артерия, 9
—воротная вена печени, 10—
кювьеровы протоки, 11
—сердце, 12—
артериальный конус, 13—
предсердие, 14—
венозный синус, 15—
желудочек, 16
—яремная вена, 17—
левая дуга аорты, 18
—подключичная артерия, 19—
передняя полая вена, 20—
легочная артерия, 21—
легочная вена, 22—
задняя полая вена, 23
—правая дуга аорты, 24—
межжелудочковая перегородка
В эмбриональном развитии млекопитающих и человека рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце
закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным (рис. 14.29). Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
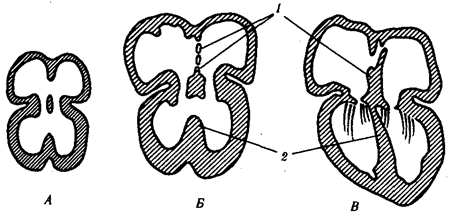
Рис. 14.29. Развитие перегородок сердца у зародыша человека в возрасте 30 сут (А
) 35 сут (Б
) и 2 мес (Д): 1
—межпредсердная, 2—межжелудочковая
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих и человека. Так, закладка сердца у человека осуществляется на 20-е сутки эмбриогенеза, как у всех позвоночных, позади головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение.
Нарушения развития сердца
могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5—5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком (см. рис. 14.31, В
).
Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфо-генезе сердца большое значение имеют онтогенетические корреляции (см. § 13.4). Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
14.4.2. Филогенез артериальных жаберных дуг
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных (рис. 14.30). В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг,
соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты — основными сосудами большого круга кровообращения.
Рис. 14.30. Эволюция артериальных жаберных дуг позвоночных. А—
закладка в эмбриогенезе; Б
—жаберные дуги рыб; В—
жаберные дуги хвостатых земноводных- Г
—жаберные дуги пресмыкающихся; Д—
жаберные дуги млекопитающих:
I
—VI -
жаберные дуги; 1
—сонные артерии, 2
—боталлов проток, 3
—артериальные дуги большого круга кровообращения, 4—
легочные артерии
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент— плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытии детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты
4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытии) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) — персистирование артериального,
или боталлова, протока
(рис. 14.31, А
)представляющего собой часть корня спинной аорты между 4-й и'6-и парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития — персистирование первичного эмбрионального ствола,
в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке (рис. 14.31, Б
). Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов —
отхождение аорты от правого желудочка, а легочного ствола — от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен.
Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия — неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
Рис. 14.31. Врожденные пороки развития сердца и сосудов у человека:
А —
персистирование артериального (боталлова) протока у человека:
1
—легочный ствол, 2—
левая дуга аорты, 3—
артериальный (боталлов) проток;
Б—
онтофилогенетические основы формирования пороков развития сердца и отходящих от него сосудов у человека: I
—сердце зародыша человека в возрасте 5 недель; II
—то же, в разрезе III—
разделение артериального конуса у зародыша человека в возрасте 6 недель, IV—
сердце рыбы, V—
то же, в разрезе, VI
—персистирование неразделенного артериального конуса у человека: 1—
артериальные жаберные дуги, 2—
артериальный конус, 3—
предсердие, 4—
кардинальные вены, 5—
желудочек, 6
—общая легочная артерия, 7—левая дуга аорты;
В —
аномалии межпредсердной (I
, II
) и межжелудочковой (III
)перегородок
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела — целбмом.
В эмбриогенезе закладка мочеполовой системы — нефрогонотом —
формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целоми-ческих животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции — через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых (рис. 14.32).
Рис. 14.32. Взаимоотношения целома (1
), половой железы (2
) и общего выделительного канала (3
)
14.5.1. Эволюция почки
Органами выделения позвоночных являются почки —
парные компактные органы, структурная единица которых представлена нефроном.
В наиболее примитивном виде это — воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток — мочеточник.
В филогенезе позвоночных почка прошла три этапа эволюции: предпочка — головная, или пронефрос;
первичная почка—туловищная, или мезонефрос, и вторичная почка
-
тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2—12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки (рис. 14.33, А
).
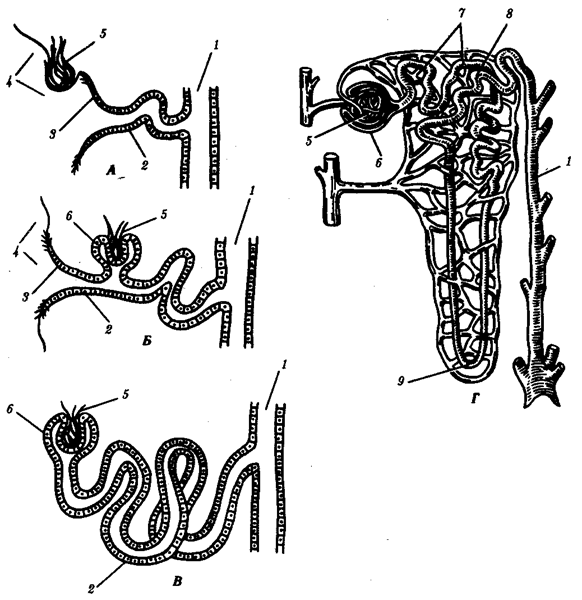
Рис. 14.33. Эволюция тефрона. А—
предпочка; Б, В—
первичная почка; Г—
вторичная почка:
1
—собирательная трубочка, 2—
выделительный канадец, 3—
нефростом, 4—
целом, 5—
капиллярный клубочек, 6—
капсула, 7, 8—
извитой канадец, 9—
петля нефрона
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки,
содержащие до нескольких сотен нефронов. В ходе онтогенеза нефроны увеличиваются в количестве за счет их почкования друг от друга с последующей дифференцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие — утрачивают её (рис. 14.33, Б, В
).
Выделительные канальцы удлиняются и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки.
Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Канадец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Генле
(рис. 14.33, Г
).
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество — мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые участки земли. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки — в тазовой области. В них прослеживаются черты первичного метамерного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки (рис. 14.34).
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития почек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка,
имеющая один или даже несколько мочеточников; возможно и полное ее удвоение.
Часто наблюдается тазовое расположение почки,
связанное с нарушением ее перемещения на 2—4-м месяцах зародышевого развития (рис. 14.35).
Рис. 14.34. Пятинедельный зародыш человека с тремя поколениями почек:
1
—предпочка, 2—
первичная почка, 3—
вторичная почка
Рис. 14.35. Онтофилогенетически обусловленные пороки развития почек:
1
—удвоение почки, 2—
удвоенный мочеточник, 3—
тазовая эктопия почки, 4—
надпочечники
14.5.2. Эволюция половых желез
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки обособляются у зародышей очень рано — уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола.
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник
всегда имеет фолликулярное строение, т.е. содержит пузырьки — фолликулы, в каждом из которых находится одна будущая яйцеклетка.
При созревании яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники
содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды
выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом.
Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных — круглоротых — таких половых желез, передняя часть которых является яичником, а задняя — семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов (см. разд. 6.1.2) становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма —
неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
14.5.3. Эволюция мочеполовых протоков
В эмбриогенезе всех позвоночных при развитии предпочки вдоль тела, от головного конца к клоаке, закладывается канал, по которому продукты диссимиляции из нефронов поступают во внешнюю среду. Это пронефрический канал.
При развитии первичной почки этот канал либо расщепляется на два канала, идущих параллельно, либо второй канал образуется в продольном утолщении стенки первого. Один из них — вольфов —
вступает в связь с нефронами первичной почки. Другой — мюллеров —
срастается передним концом с одним из нефронов предпочки и образует яйцевод, открывающийся передним концом в целом широкой воронкой, а задним — впадающий в клоаку.
Вне зависимости от пола у всех позвоночных обязательно формируются как вольфов, так и мюллеров каналы, однако судьба их различна как у разных полов, так и у представителей разных классов. У самок рыб и земноводных вольфов канал всегда выполняет функцию мочеточника, а мюллеров — яйцевода. У самцов мюллеров канал редуцируется и обе функции — половую и выделительную — выполняет вольфов канал. Семенные канальцы при этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду вместе с мочой.
У пресмыкающихся и млекопитающих большая часть вольфова канала не принимает участия в выведении мочи и только его наиболее каудальная часть в области впадения в клоаку образует выпячивание, становящееся мочеточником вторичной почки. Сам же вольфов канал у самцов выполняет функцию семяизвергательного канала. Мюллеров канал у них подвергается редукции. У самок вольфов канал редуцируется (за исключением его каудальной части, формирующей мочеточник), а мюллеров — становится яйцеводом (рис. 14.36). У плацентарных млекопитающих мюллеров канал дифференцируется на собственно яйцевод, матку и влагалище. Будучи парным образованием, как и все элементы половой системы, мюллеров канал сохраняет парность строения у яйцекладущих и частично у сумчатых млекопитающих, у которых имеется два влагалища, две матки и два яйцевода. В дальнейшей эволюции происходит срастание мюллеровых каналов с образованием одного влагалища и матки, которая может быть либо двойной, как у многих грызунов, либо двураздельной, как у хищных; либо двурогой, как у насекомоядных и китообразных, либо простой, как у приматов и человека.
Соответственно дифференцировкам мюллерова канала самок у самцов пресмыкающихся и млекопитающих развиваются копулятивные органы.
У большинства пресмыкающихся, а также у сумчатых млекопитающих они парные. У плацентарных с одним влагалищем копулятивный орган непарный, но в его развитии обнаруживается срастание парных зачатков.
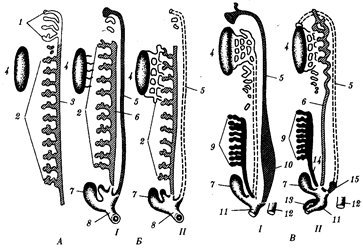
Рис. 14.36. Эволюция почки и мочеполовых каналов.
А —
нейтральное зародышевое состояние; Б —
анамнии; В —
амниоты:
I
—самки, II
—самцы; 1
—предпочка, 2—
первичная почка, 3—
канал предпочки, 4—
половая железа, 5—
мюллеров канал, 6—вольфов канал, 7—мочевой пузырь, 8—
клоака, 9—
вторичная почка, 10
—матка, 11
—мочеполовой синус, 12—
задняя кишка, 13—
половой член, 1
4
—мочеточник вторичной почки, 15—
мужская «маточка»
В эмбриогенезе человека закладываются парные вольфовы и мюллеровы каналы. Позже в зависимости от пола происходит их редукция. Рудимент мюллерова канала у мужчин располагается в предстательной железе и называется мужской маточкой —
utriculus masculinus. Канальцы передней части первичной почки у них вступают в связь с семенниками и преобразуются в придаток семенника — эпидидимис.
У плодов женского пола возможно нарушение редукции вольфовых каналов,
которые располагаются по бокам от влагалища. Эта аномалия опасна возможностью образования кист и злокачественного перерождения. Распространенными пороками развития являются также различные формы удвоения матки
(1 случай на 1000 перинатальных вскрытии). Они развиваются как результат нарушения срастания мюллеровых каналов. Нарушение срастания парных зачатков полового члена в эмбриогенезе человека может привести к формированию такого порока развития, как его удвоение
(рис. 14.37).
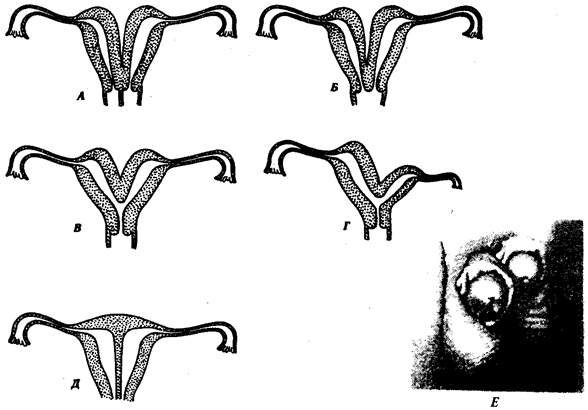
Рис. 14.37. Аномалии матки и полового члена у человека. А
—Д
—различные формы удвоения матки и влагалища; Е—
удвоение полового члена
У большинства многоклеточных животных, в том числе и у позвоночных, существует две системы регуляции функций, интегрирующие организм в целостную систему, — нервная и эндокринная. Нервная система осуществляет быстрое реагирование организма на изменения условий среды посредством рефлексов. Эндокринная система с помощью гормонов обеспечивает более медленные, но и более стойкие приспособительные реакции. В связи с тесным взаимодействием обеих регуляторных систем в их филогенезе проявляются тесные динамические координации, а в онтогенезе — эргонтические корреляции.
14.6.1. Центральная нервная система
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы.
Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых — ланцетника — центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования — глазки Гессе.
Кроме того, основные дистантные органы чувств — зрения, обоняния и слуха — образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри — невроцелем.
У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок (рис. 14.38). Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.

Рис 14.38. Нервная трубка ланцетника: 1
— невроцель, 2
— глазки Гессе
У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией.
Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных — обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными — и средним, слуховыми — и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей — переднего, среднего и заднего — и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозг
современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал,
а в головном — желудочки мозга.
Ткань мозга состоит из серого вещества
(скопления нервных клеток) и белого
(отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней —
archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный
тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой —
neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок (рис. 14.39).
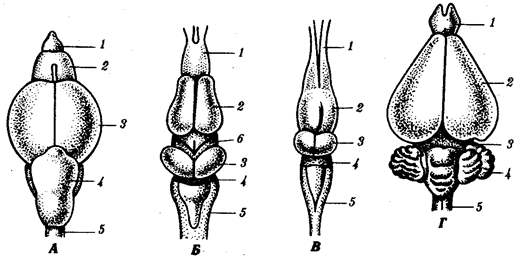
Рис. 14.39. Эволюция головного мозга позвоночных. А—рыба; Б—
земноводное; В—
пресмыкающееся; Г—
млекопитающее:
1
—обонятельные доли, 2—
передний мозг, 3—
средний мозг, 4—
мозжечок, 5—
продолговатый мозг, 6—
промежуточный мозг
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (I
),
средний (II
) и задний (III
).
Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2
) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4
) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
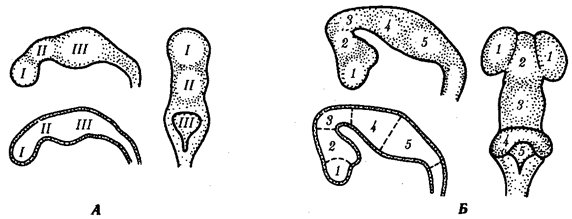
Рис. 14.40. Мозговые пузыри в эмбриогенезе мозга человека. А —
стадия трех мозговых пузырей; Б —
стадия пяти мозговых пузырей (остальные пояснения см. в тексте)
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека (см. § 15.3).
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли
платиневрию,— отсутствие замыкания нервной трубки (рис. 14.41). Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия —
выражается в нарушении морфо-генеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия
(отсутствие извилин) и олигогирия
с пахигирией
(малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
Рис. 14.41. Рахисхиз (платиневрия)
14.6.2. Эндокринная система
Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами —
биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции.
Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной потому, что она более проста и не требует развития таких сложных структур, как нервная система.
Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции.
Химическая структура гормонов
многообразна. Это могут быть белки (инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин). Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, — способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной.
Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательством этого является обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших и низших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей.
В эмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы и дни развития. В процессе дробления они регулируют течение клеточного цикла. Позже — перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных — соответственно на стадии первичного органогенеза.
При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих и человека гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых — образование оболочек икринок в яйцеводах, у третьих — некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения.
Функции других гормонов, например адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена.
При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных. Так, адреналин в организме млекопитающих и человека замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Из главной эта функция у высокоорганизованных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же как и на эволюцию органов, распространяются основные закономерности макроэволюции (см. гл. 13).
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции.
Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная
и паращитовидная железы. Эпифиз
развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа
имеют сложное происхождение.
Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус —
нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма в единое целое: нервной и эндокринной. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему.
Эволюционным предшественником гипоталамуса является так называемый инфундибулярный вырост ланцетника, состоящий из нейросекреторных клеток и находящийся на вентральной стороне переднего конца нервной трубки.
Начиная с рыб гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные
и моноаминовые.
Первые — гормоны, влияющие на функции внутренних органов — вазопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Под их действием стимулируется или подавляется секреция гормонов соответствующими гипофизарными клетками.
Подробнее остановимся на эволюции гипофиза.
Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом — воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие.
Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиальными. Промежуточная доля — производная от передней (рис. 14.42).
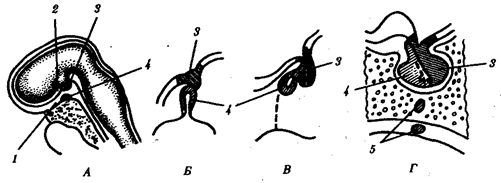
Рис. 14.42. Развитие гипофиза у человека. А—
сагиттальный срез 4-недельного зародыша; Б—
объединение зачатков гипофиза из стенки глотки и основания мозга; В—
формирование трех долей гипофиза на 8-й неделе развития; Г
—сформированный гипофиз:
1
—крыша ротовой полости, 2—
основание мозга,3
—выпячивание основания мозга (задняя доля гипофиза), 4—
карман Ратке (промежуточная и передняя доли гипофиза), 5—
аномалия развития (добавочные дольки передней доли в стенке глотки и внутри клиновидной кости, оставшиеся в области выпячивания кармана Ратке)
У хрящевых рыб во взрослом состоянии сохраняется первоначальная связь передней доли гипофиза с эпителием ротовой полости. За счет ее задней части формируется также средняя доля. Обе доли вырабатывают гонадотропные гормоны. У костных рыб и личинок земноводных имеются передняя и промежуточная доли, а у взрослых амфибий, переходящих к наземному существованию, появляется также задняя, регулирующая водный обмен. Средняя доля у них перестает выделять гонадотропный гормон, но вырабатывает пролактин.
В связи с наземным образом жизни пресмыкающихся и млекопитающих у них наиболее прогрессивно развивается задняя доля гипофиза, что связано с интенсификацией водного обмена.
Передняя доля вырабатывает соматотропный гормон (гормон роста) и ряд гормонов, регулирующих функции других желез внутренней секреции, а средняя — пролактин и некоторые другие. При этом усиливается дифференцировка и интенсифицируются функции ядер гипоталамуса, находящихся в функциональной связи со всеми долями гипофиза.
У человека в эмбриогенезе развитие гипофиза соответствует основным этапам его эволюции. Очень часто, в 30—40%, у нормальных людей под слизистой оболочкой крыши глотки, в основании клиновидной кости, обнаруживается группа клеток длиной 5—6 мм и шириной 0,5—1 мм, по структуре и функциям соответствующая передней доле гипофиза. Это результат нарушения перемещения клеток при закладке гипофиза в эктодерме ротовой полости в области турецкого седла. Эту 139 аномалию называют эктопией аденогипофиза,
она не сопровождается патологическими проявлениями. Более опасно сохранение полости в области кармана Ратке — киста кармана Ратке.
Она располагается между передней и промежуточной долями, содержит слизь и в ряде случаев имеет тенденцию к росту и даже к переходу в злокачественное новообразование. Растущие кисты сдавливают гипофиз и вызывают его гипофункцию. Больные нуждаются в оперативном вмешательстве.
Щитовидная железа, гормон которой — тироксин —
регулирует энергетический обмен, среди хордовых как компактный орган впервые появляется у рыб. Однако уже у ланцетника отдельные тироксин-синтезирующие клетки обнаруживаются в желобке на вентральной стороне глотки. Щитовидная железа рыб закладывается также в виде желобка на вентральной стороне глотки между 1-й и 2-й жаберными щелями в области зачатка основания языка. Позже этот клеточный материал погружается под слизистую оболочку и формирует фолликулы, характерные для щитовидной железы. У других позвоночных железа закладывается так же, как у рыб, но затем она перемещается в область подъязычной кости (у земноводных) или в шейную область (у пресмыкающихся и млекопитающих).
У человека в эмбриогенезе щитовидной железы происходит рекапитуляция предковых состояний. Гетеротопия ее осуществляется посредством миграции клеток в виде тяжа, полого внутри, называющегося щитоязычным протоком — canalis thyreoglossus. При нормальном развитии этот проток полностью редуцируется, рудиментом его является слепое отверстие foramen caecum в корне языка.
Свидетельством эмбрионального перемещения железы является и расположение верхней щитовидной артерии a. thyreoidea superior, которая, начинаясь от общей сонной артерии, резко поворачивает вниз и входит в ее ткань. На ранних этапах эмбрионального развития эта артерия направляется вверх к зачатку щитовидной железы, а затем меняет свое направление вместе с его перемещением книзу.
Персистирование участков подъязычного протока в постнатальном периоде сопровождается накоплением в них жидкости и образованием срединных кист шеи,
которые могут располагаться в любом месте от корня языка до верхней границы щитовидного хряща. Иногда, кисты загнаиваются и прорываются с образованием срединных свищей шеи.
Не менее известны такие пороки развития, как эктопия щитовидной железы
, которая может развиться в результате нарушения клеточной миграции (рис. 14.43).
Из эпителия глотки в области III-V жаберных карманов у позвоночных развиваются мелкие эндокринные образования, связанные по месту окончательного положения с щитовидной железой. Это паращитовидные железы
и ультимобранхиальные тельца.
Первые, выделяя гормон паратиреоидин,
повышающий содержание ионов кальция в крови и уменьшающий их количество в костях, развиваются как самостоятельные железы только у наземных позвоночных, а у земноводных — лишь после метаморфоза. Клетки вторых выделяют кальцитонин,
являющийся антагонистом паратиреоидина. Таким образом, паращитовидные железы и ультимобранхиальные тельца являются регуляторами кальциевого обмена. Значение их наиболее велико у наземных позвоночных. В филогенетическом ряду позвоночных они постепенно перемещаются из глоточной области в сторону щитовидной железы, а у млекопитающих даже срастаются с ней. Паращитовидные железы еще сохраняют самостоятельность, а клетки ультимобранхиальных телец мигрируют между фолликулами щитовидной железы и сохраняются под названием парафолликулярных клеток. Таким образом, на примере щитовидной, паращитовидных желез и ультимобранхиальных телец видна интеграция железистых структур в сложное надерганное образование, выполняющее целый комплекс взаимосвязанных функций.
Рис. 14.43. Аномалии щитовидной железы:
1—
нормальное расположение.
железы, 2—
место эмбриональной закладки железы, 3—
варианты аномального расположения железы; стрелками показано направление перемещения зачатка щитовидной железы в эмбриогенезе
У человека известны аномалии расположения паращитовидных желез, связанные с нарушением их гетеротопий.
Надпочечники позвоночных имеют двойственное происхождение. У рыб и земноводных ткани, соответствующие мозговому и корковому веществам этих желез, расположены отдельно друг от друга. Зачатки мозгового вещества связаны по происхождению с симпатическими нервными узлами и расположены метамерно. Зачатки коркового вещества развиваются из утолщений эпителия брюшины. У наземных позвоночных мозговое и корковое вещества объединяются в компактные эндокринные железы, имеющие сложное гистологическое строение. У млекопитающих они прилежат к переднему концу почек. Мозговое вещество выделяет в основном адреналин —
регулятор кровообращения и энергетического обмена, а корковое вещество — разнообразные стероидные гормоны,
влияющие на минеральный, углеводный обмены и функции почек.
Неограниченный прогресс в эволюции живой материи проявился в возникновении человека как биосоциального существа. Появление человека с его целенаправленной трудовой деятельностью качественно изменило облик планеты Земля и определило новое направление эволюции природы в целом. Будучи биологическим видом, человек — единственный на Земле организм, обладающий социальной сущностью. Социальная сущность человека определяет его современное состояние и прогнозирует будущее. Биологическое в нем при этом становится его наследством, поэтому эволюция человека на современном этапе идет не столько по биологическим законам, сколько по законам развития человеческого общества. Биологическая эволюция представляет собой биологическую форму движения материи, а эволюция общества — социальную. Естественно, что социальная форма движения материи возникла лишь с появлением человека. Поэтому изучение вопросов антропогенеза — это не только биологическая, но и философская проблема.
Человек как биологический вид относится к отряду приматов
(рис. 15.1).
Рис. 15.1. Место человека в отряде приматов:
1
—предок приматов, 2—
вымершие и современные полуобезьяны, 3—
настоящие обезьяны Нового, Старого света и человек
С биологических позиций важно ответить на вопрос: почему человек возник именно в отряде приматов, а не в другом эволюционно прогрессивном отряде млекопитающих. Ключевой адаптацией представителей отряда приматов являются древесный образ жизни и передвижение по ветвям с помощью хватательных конечностей, имеющих противопоставленный большой палец и уплощенные ногти вместо когтей. Пальцевые подушечки и внутренние поверхности кистей и стоп имеют кожные узоры в виде гребешков и бороздок, увеличивающих контакт пальцев с ветками и усиливающих их тактильную чувствительность. Локтевая кость свободно вращается вокруг лучевой. Эти особенности при освобождении рук от участия в движении являлись предпосылкой к использованию их в трудовой деятельности. Другими прогрессивными чертами приматов, связанными с энергичными перемещениями в трехмерном пространстве, являются бинокулярное зрение, сильно развитые мозжечок и кора больших полушарий головного мозга.
В отличие от большинства отрядов млекопитающих приматов характеризует также выраженная социальность: жизнь небольшими стадами со сложной системой иерархии и общественным воспитанием потомства. Плодовитость приматов чрезвычайно низка — обычно рождается один детеныш, беспомощный и нуждающийся в уходе на протяжении многих лет. Характерной особенностью поведения приматов является выраженный инстинкт подражания, обеспечивающий быстрое научение и возможность передачи социального опыта в ряду поколений. Перечисленные черты отряда приматов явились необходимыми преадаптациями к возникновению человека при появлении комплекса благоприятных условий.
Обычно выделяют следующие этапы эволюции
человека:
1. Древнейшие стадии гоминизации — происхождение родаHomo.
2. Эволюция родаHomo
до возникновения современного человека.
3. Эволюция современного человека.
Первый этап антропогенеза есть чисто биологическая эволюция. На втором этапе к элементарным факторам биологической эволюции подключается действие социального фактора, который на третьем этапе является доминирующим. Методологические подходы к изучению разных этапов антропогенеза различны.
Изучение 1-го этапа производят методами палеонтологии
и сравнительной анатомии.
В связи с появлением элементов материальной культуры 2-й этап изучают также методами археологии.
На 3-м этапе эволюционные события происходят в основном на молекулярно-генетическом уровне и проявляются на популяционном уровне. Основными способами изучения эволюции современного человека поэтому являются биохимический, цитогенетический
и популяционно-статистический методы.
Изучение первых двух этапов эволюции человека сталкивается с необходимостью датировки палеонтологического материала и элементов материальной культуры. Для определения абсолютного возраста ископаемых остатков человека и его предков широко используют физические методы,
в частности радиометрические. С помощью масс-спектрометров определяют изотопный состав изучаемого объекта и по соотношению элементов с учетом периода полураспада входящих в его состав радиоактивных изотопов выявляют возраст образца. Ископаемый костный материал содержит в своем составе минеральные компоненты и белок коллаген, разрушающийся чрезвычайно медленно. На этом основан коллагеновый метод
абсолютной датировки ископаемых остатков: чем меньше коллагена содержится в образце, тем более велика его древность.
В последнее время в антропологии все более активно применяют методы иммунологии, молекулярной биологии и цитогенетики. В связи с огромной важностью этих методов остановимся на них подробнее. Для определения прямого родства организмов друг с другом используют иммунологический метод,
основанный на изучении иммунологических реакций антиген — антитело. Его можно применять для изучения степени родства не только современного человека с человекообразными обезьянами, но и ныне живущих видов с ископаемыми. Для этого следовые количества белка, извлекаемые из костей ископаемых форм, используют для получения антител, которые и применяют в иммунных реакциях с белками современных видов. Из современных человекообразных обезьян к человеку иммунологически наиболее близок шимпанзе, наиболее далеко от человека отстоит орангутан.
Иммунологическим методом было обнаружено, что белки рамапитека, человекообразной обезьяны Южной Азии (абс. возраст 13 млн. лет), более сходны с белками орангутана, чем человека и шимпанзе. Эти данные вместе с результатами морфологических и палеонтологических сопоставлений заставили отказаться от представления о том, что рамапитек является прямым предком человека, и связать его с эволюционной линией орангутана. Из этого следует, что разделение человеческой линии эволюции с африканскими человекообразными обезьянами произошло значительно позже, чем 13 млн. лет назад.
В последние годы антропогенез эффективно изучают также биомолекулярными методами.
В основе этих методов изучения эволюции лежит допущение, что мера сходства двух таксонов соответствует мере их родства. Поэтому организмы, имевшие общего предка в недалеком прошлом, будут более'сходными друг с другом, чем имевшие его очень давно. Сущность биомолекулярных методов состоит в использовании данных, полученных при сопоставлении белков и нуклеиновых кислот организмов разных видов для определения их родства и древности соответствующих филогенетических ветвей. При этом считают, что степень различий в аминокислотном составе белков и в нуклеотидных последовательностях ДНК позволяет судить о времени расхождения сравниваемых видов от предковой формы. Естественно, что оценивать степень родства и давность расхождения филогенетических ветвей можно лишь по накоплению нейтральных мутаций в геноме, так как вредные мутации быстро элиминируются из генофондов. В действительности нельзя считать большинство мутаций селективно нейтральными.
Расчеты показывают, что при нейтральности мутаций скорость их накопления постоянна только при измерении времени в количестве поколений, а не в годах. Так как продолжительность жизни поколений у различных видов различна, то и скорости накопления различий нуклеотидных последовательностей будут более велики у короткоживущих видов по сравнению с долгоживущими. Кроме того, фактическая скорость эволюции может значительно варьировать в различные временные интервалы в различных группах и по разным признакам. В определении применимости методов молекулярной биологии имеет значение возможность возникновения конвергентного сходства молекул, причем вероятность его повышается с увеличением изучаемых временных интервалов. Эволюция генов и белков часто может опережать реальное расхождение популяций, в первую очередь за счет адаптивного генетического полиморфизма. Однако молекулярно-биологические методы применимы для оценки родства и времени дивергенции в качестве приблизительных «молекулярных часов» при сравнении средних скоростей замен нуклеотидов в ДНК в целом и аминокислот во многих белках за длительные интервалы времени.
Биомолекулярный подход — лишь один из путей определения эволюционных расстояний, который работает только наряду с классическими методами палеонтологии и антропологии, причем в результатах при этом возможны серьезные расхождения. Так, при изучении скелета человека, обнаруженного на территории Эквадора, по данным радиоуглеродного и аминокислотного анализа была установлена его древность в 28 тыс. лет. При использовании же коллагенового анализа возраст того же скелета оказался датированным не более чем в 2,5 тыс. лет.
В антропологии применяют несколько методов оценки степени дифференцированности таксонов, дополняющих друг друга: гибридизация ДНК, определение аминокислотных последовательностей белков, изучение генов гистосовместимости и т.д. Наиболее достоверные данные получены путем гибридизации ДНК,
так как скорость эволюции ДНК в целом более постоянна, чем скорость изменения белков. Гибридизация ДНК показала, что момент дивергенции эволюционных ветвей человека и шимпанзе наступил 6,5—6,7 млн. лет назад (рис. 15.2).
Однако между эволюцией структуры генома в виде накопления генных мутаций и морфофизиологической эволюцией часто нет прямой зависимости (рис. 15.3). Это может быть связано с тем, что в формировании практически всех сложных фенотипических признаков принимают участие различные генные системы. Таким образом, скорость эволюции белков у двух разных родственных видов может быть одинакова, а скорость эволюции в целом, оцененная по комплексу фенотипических признаков, при этом оказывается различной. Сравнение аминокислотных последовательностей белков шимпанзе и человека привело к выводу, что около 99% их белков абсолютно идентичны. Из этого следует, что и структурные гены человека и шимпанзе сходны в наибольшей степени. С чем же связаны столь значительные морфофизиологические отличия обоих видов? Можно предполагать, что это зависит от различного распределения белков в клетках организма в процессе развития, что, в свою очередь, определяется различиями программы считывания сходной наследственной информации во времени и пространстве. Иными словами, это может быть обусловлено эволюционными преобразованиями в большей степени не структурных, а регуляторных генов.
Рис. 15.2. Среднее время дивергенции высших приматов по часам ДНК
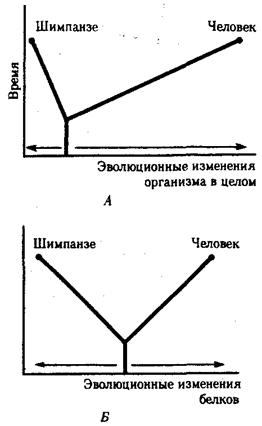
Рис. 15.3. Соотношение скорости морфофизиологической (А
) и молекулярной (Б
) эволюции
Поскольку изучение хромосомного материала возможно только у ныне живущих организмов, применение цитогенетического метода
ограничивается современным человеком и человекообразными обезьянами. Дифференциальная окраска хромосом позволяет не только сопоставлять хромосомы разных видов приматов и человека и изучать хромосомный полиморфизм современного человека, но и решать некоторые вопросы эволюции.
Выяснено, что кариотип человекообразных обезьян отличается по количеству хромосом от кариотипа человека на одну пару (23 пары хромосом человека и 24 пары шимпанзе). У человека и шимпанзе идентичны 13 пар хромосом. Хромосома 2-й пары человека точно соответствует двум соединенным хромосомам шимпанзе, а остальные хромосомы отличаются друг от друга незначительно. Так, хромосома 5-й пары шимпанзе соответствует такой же хромосоме человека, но небольшой ее перицентрический участок инвертирован на 180° по сравнению с человеческой хромосомой. Инверсии такого рода обнаружены в кариотипах человека и шимпанзе еще в восьми хромосомах. Эти данные вместе с указаниями на сходство белков человека и шимпанзе свидетельствуют об их значительной эволюционной близости.
Сопоставление кариотипов людей, происходящих из разных популяций, приводит к выводу о полиморфизме хромосом, в первую очередь по размерам гетерохроматиновых участков. Наследуемость индивидуальных вариаций хромосом и их неравномерное распределение в разных популяциях (в частности, расовые различия по размерам длинного плеча Y-хромосомы) делают возможным популяционно-цитогенетический подход
в изучении эволюции современного человека.
Долгое время многие палеонтологи и антропологи считали наиболее древними представителями семейства гоминид южно-африканских двуногих приматов
Australopithecus africanus.
Возраст большей части южно-африканских находок австралопитеков определяется в интервале 3—1 млн. лет. Строение скелетов этих организмов свидетельствует об их прямохождении. Масса мозга — 450—550 г при общей массе тела от 25 до 65 кг. Зубная система близка к человеческой: клыки малых
146
размеров, зубы расположены в виде широкой дуги, как у человека. Это свидетельствует о всеядности.
В местах обнаружения остатков австралопитеков имеется множество костей, расколотых тяжелыми предметами. Большое число черепов животных расколоты с левой стороны, и это свидетельствует о том, что австралопитеки были в основном правшами. Некоторые австралопитеки, видимо, начинали осваивать огонь. В 1960—1970-х гг. в результате раскопок в Эфиопии и позже в Танзании и Кении были обнаружены большие группы гоминид, древность которых определяется в 4—2,8 млн. лет. По этим материалам в 1978 г. был описан новый вид австралопитековA. afarensis,
более примитивный, чем A. africanus,
но тоже двуногий и с чертами «промежуточности» по отношению к современным человекообразным обезьянам и человеку (рис. 15.4). Объем мозговой полости черепа оказался в пределах 380—450 см3
, т.е. практически не отличался от объема мозга современного шимпанзе.

Рис. 15.4. Австралопитек афарский. Реконструкция
Действительно, австалопитеки имеют большее сходство с человеком не по объему и строению мозга, а по строению зубной системы и способу передвижения. Изучение австралопитековых показало, что именно двуногость, а не большой объем мозга, явилась ключевой адаптацией ранних гоминид. В 1964 г. по находкам, сделанным в Танзании, был выделен видHomo habilis,
или Человек умелый, имеющий абсолютный возраст 2—1,7 млн. лет. Отличительными чертами его являются двуногость, в целом прогрессивное строение кисти, зубной системы, объем мозговой коробки от 540 до 700 см3
, что примерно в полтора раза превышает объем мозга австралопитеков. На внутренней поверхности черепа обнаруживаются признаки прогрессивных нейроморфологических изменений, определяющиеся по отпечаткам головного мозга: выраженная асимметрия полушарий и развитие двух речевых центров как условие для возникновения членораздельной речи. Большой палец стопы не отведен в сторону. Это свидетельствует о том, что морфологические перестройки, связанные с прямохождением, у него полностью завершились. Вместе с остатками Н.
habilis
найдены орудия труда со следами целенаправленной обработки, свидетельствующие о ранних формах трудовой деятельности.
Перечисленные признаки, ведущим из которых является прогрессивное развитие мозга, характеризуют организм уже с иной морфофункциональной организацией по сравнению с австралопитеками. Эти признаки считаются специфичными для родаHomo.
Сопоставление морфологии африканского и афарского австралопитеков с Н.
habilis
и современным человеком позволяет предположить следующую схему филогенетических взаимоотношений ранних гоминид (рис. 15.5). Африканский австралопитек, считавшийся раньше общим предком как родаHomo,
так и других австралопитековых, является в этой схеме представителем боковой ветки эволюции, приведшей к узкой специализации и образованию мощных форм типаA. robustus,
которые вымерли около 1 млн. лет назад. Общим предком всех австралопитековых и родаHomo
является в соответствии с этой схемойA. afarensis.
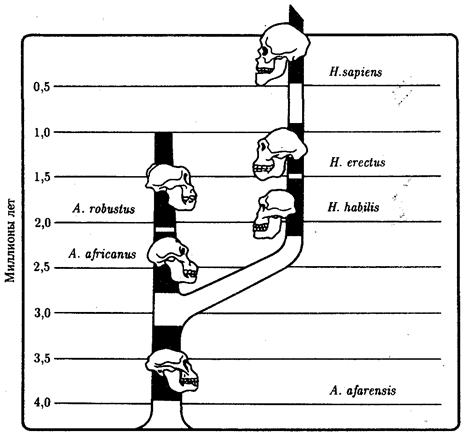
Рис. 15.5. Филогенетические взаимоотношения ранних гоминид
Таким образом, на протяжении 1—1,5 млн. лет представители двух близких родов и, возможно, нескольких видов семейства гоминид сосуществовали, причем не только во времени, но и на перекрывающихся территориях. В основе дивергенции различных линий ранних гоминид и австралопитековых могли лежать разного рода механизмы изоляции, в первую очередь генетические: мутации в виде хромосомных перестроек. Это означает, что эволюция австралопитековых шла постепенно, приводя благодаря дивергенции к морфологическому и экологическому разнообразию.
Ведущими факторами эволюции
на прегоминидной стадии антропогенеза являлись, несомненно, факторы биологической эволюции, главным из которых является естественный отбор.
Об этом свидетельствует, в частности, большое видовое разнообразие австралопитековых, обитавших в различных условиях практически на всей территории Южной, Центральной и Северо-Восточной Африки. В это же время в происхождении родаHomo
имело место скачкообразное изменение наследственного материала.
В разных органах и системах прегоминид обнаруживалась асинхронность филогенеза.
Есть предположение о том, что эволюция коры больших полушарий мозга состоит из двух компонентов, разобщенных по времени: соматического, обеспечивающего сенсорно-моторные функции, и несоматического, связанного с высшими психическими функциями. Если локомоторный комплекс подвергался длительным постоянным изменениям, то головной мозг эволюционировал скачкообразно. Элементы скачкообразности в эволюции некоторых структур ранних гоминид могли быть обусловлены «транспозиционными взрывами» (перестройками генома за счет подвижных генетических элементов, переносящих комплексы генов на новое место). Это могло сопровождаться постепенным развитием других морфофизиологических свойств в результате накопления малых мутаций под контролем естественного отбора. Но именно в период становления Н.
habilis
возникла, вероятно, часть хромосомных перестроек в геноме человека, о которых говорилось выше.
Следующей ступенью гоминизации после появления Н.
habilis
считается возникновение архантропов,
представителем которых является сборный вид Н.
erectus,
или Человек прямоходящий. Материальная культура и ярко выраженная социальность позволили ему быстро и эффективно расселиться по всей территории Африки и Евразии и освоить обширный ареал, разнообразный в климатическом отношении. Действительно, орудия труда Н.
erectus
более прогрессивны, чем у Н.
habilis,
а масса мозга (от 800 до 1000 г) явно превышает минимальную массу (750 г), при которой возможно существование речи. Наличие при этом речевых центров, возникших впервые у Н.
habilis,
предполагает и развитие второй сигнальной системы.
Выделяют три группы Н.
erectus:
обитавшие в Европе, Азии и Африке. Долгое время древнейшими архантропами считались азиатские представители из Индонезии и Восточного Китая — питекантроп и синантроп.
Однако находки последних лет на территории Израиля (1982) и Кении (1984), датирующиеся соответственно 2,0 и 1,6 млн. лет, сопровождающиеся элементами материальной культуры и признаками использования огня, показали, что арогенная эволюция гоминид происходила на африканском континенте и на Ближнем Востоке. Это позволило связать происхождение Н.
erectus
с
восточноафриканскими формами Н.
habilis.
Наличие большого количества находок архантропов древностью 1,5—0,1 млн. лет в отдаленных от Африки регионах — в Юго-Восточной и Восточной Азии, в Центральной Европе и даже на Британских о-вах — свидетельствуют об активных адаптациях их к разнообразным условиям существования (рис. 15.6). В связи с тем что небольшое различие ископаемых остатков Н.
erectus
не соответствует значительному разнообразию природно-климатических условий указанных территорий, можно заключить, что в этих адаптациях значительную роль играли наряду с факторами биологической эволюции также и социальные факторы: совместное изготовление укрытий, орудий труда и использование огня.

Рис. 15.6. Места обнаружения ископаемых остатков Человека прямоходящего
Роль Н.
erectus
как этапа антропогенеза никогда не подвергалась сомнению. Что же касается палеоантропа,
или неандертальского человека,
то его роль в происхождении человека в настоящее время оспаривается. Это связано в первую очередь с обнаружением большого количества ископаемых остатков человека с промежуточными чертами между Н.
erectus
и человеком современного физического типа. Кроме того, палеонтологические находки последних лет дают повод судить о недооценке интеллектуальных возможностей неандертальцев. На всех стоянках обнаружены следы костров и обгоревшие кости животных, что свидетельствует об использовании огня для приготовления пищи. Орудия труда их гораздо совершеннее, чем у предковых форм. Масса мозга неандертальцев около 1500 г, причем сильное развитие получили отделы, связанные с логическим мышлением. Костные остатки неандертальца из Сен-Сезер (Франция) были найдены вместе с орудиями труда, свойственными верхнепалеолитическому человеку, что свидетельствует об отсутствии резкой интеллектуальной грани между неандертальцем и современным человеком. Имеются данные о ритуальных захоронениях неандертальцев на территории Ближнего Востока.
Эти и ряд других находок позволили в конце 60-х годов выделить палеоантропов в отдельный подвидHomo sapiens neanderthalensis
в отличие от неоантропа
H.s. sapiens,
который, таким образом, тоже получил ранг подвида. Наиболее древние ископаемые остатки его возрастом 100 тыс. лет обнаружены также на территории Северо-Восточной Африки. Многочисленные находки палеоантропов и неоантропов на территории Европы, датирующиеся 37—25 тыс. лет, свидетельствуют о существовании обоих подвидов в течение нескольких тысячелетий.
В тот же период неоантропы обитали уже не только в Европе и Африке, но и в отдаленных районах Азии (о. Тайвань, о. Окинава) и даже в Америке. Эти данные указывают на необычайно быстрый процесс расселения современного человека, что может быть доказательством «взрывного», скачкообразного характера антропогенеза в этот период как в биологическом, так и в социальном смысле. H.s.
neanderthalensis
в виде ископаемых остатков не обнаруживается позже рубежа в 25 тыс. лет. Быстрое исчезновение палеоантропов может быть объяснено вытеснением их людьми с более совершенной техникой изготовления орудий труда и метисацией с ними.
С возникновением человека современного физического типа роль биологических факторов в его эволюции свелась к минимуму, уступив место социальной эволюции. Об этом отчетливо свидетельствует отсутствие существенных различий между ископаемым человеком, жившим 30—25 тыс. лет назад, и нашим современником.
С момента возникновения Н.
sapiens
социальное в человеке стало его сущностью и биологическая эволюция видоизменялась, проявляясь в возникновении широкого генетического полиморфизма (см. § 12.3).
Генетическое разнообразие на уровне генов и в меньшей степени хромосом обеспечивает разнообразие генотипов особей. Разнообразные генотипы по-разному проявляются в меняющихся условиях среды, давая огромное фенотипическое многообразие людей.
В основе морфофизиологического полиморфизма человечества лежат полиморфизм наследственного материала на уровне генома и модификационная изменчивость. Эти факторы обеспечивают не только индивидуальное морфофизиологическое многообразие, но и внутривидовую групповую дифференциацию человечества на расы и адаптивные экологические типы.
15.4.1. Расы и расогенез
На протяжении длительного времени в антропологии господствовали представления о значимости расовой дифференцировки человечества и о большой роли естественного отбора в формировании основных расовых признаков. Применение методов молекулярной антропологии в значительной степени изменило представление о расах и расогенезе.
Морфологические и в меньшей степени физиологические признаки дают возможность выделить внутри человечества три основные большие расы:
европеидную, австрало-негроидную и монголоидную.
Европеоиды
имеют светлую или смуглую кожу, прямые или волнистые волосы, узкий выступающий нос, тонкие губы и развитый волосяной покров на лице и теле. У монголоидов
кожа также может быть как светлой, так и темной, волосы обычно прямые, жесткие, темно пигментированные, косой разрез глаз и эпикант («третье веко»). Негроиды
характеризуются темной кожей, курчавыми или волнистыми волосами, толстыми губами и широким, слегка выступающим носом. Имеются отличия рас и по некоторым физиологическим и биохимическим показателям: интенсивность потоотделения с единицы площади кожи у негроидов выше, чем у европеоидов, средние показатели уровня холестерина в плазме крови наиболее велики у европеоидов.
В рамках каждой большой расы выделяются отдельные антропологические типы с устойчивыми комплексами признаков, называющиеся малыми расами.
Существует три основных подхода к классификации рас: без учета их происхождения, с учетом происхождения и родства и на основе популяционной концепции. В соответствии с первым подходом три большие расы включают в себя 22 малые, причем между большими расами располагаются по две переходные малые. Схема расовой классификации изображается при этом в виде круга (рис. 15.7). Несмотря на то что при такой классификации не учитывается происхождение рас, само существование малых переходных рас, сочетающих в себе одновременно признаки двух больших рас (эфиопская, южносибирская, уральская и т.д.), свидетельствует, с одной стороны, о динамизме расовых комплексов признаков, а с другой — об условности членения человечества даже на большие расы.
Гибридизация ДНК между большими выборками представителей малых рас в рамках одной большой показала высокую степень гомологии нуклеотидных последовательностей. Гибридизация ДНК представителей пар разных больших рас выявляет их значительную отдаленность друг от друга. Изучение гомологии нуклеотидных последовательностей западных европеоидов и представителей малой уральской расы и центрально-азиатских монголоидов с той же самой уральской расой дает среднее значение. Эти данные свидетельствуют о том, что переходные малые расы совмещают в себе не только морфологические признаки в соответствии с их промежуточным положением, но оказываются промежуточными и в отношении генетическом. Из этого следует, что они либо гибридогенны, либо сохранили в своей организации более древние черты, характерные для этапа существования человечества, предшествующего формированию больших рас.
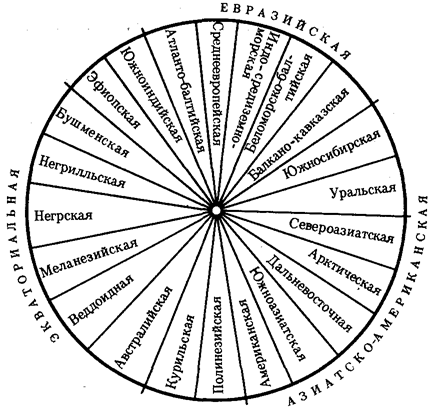
Рис. 15.7. Расовая классификация человечества без учета происхождения рас
Классификация с учетом происхождения рас изображается в виде эволюционного древа с коротким общим стволом и расходящимися от него ветвями (рис. 15.8). В основе таких классификаций лежит обнаружение черт архаизма и эволюционной продвинутости отдельных рас, в соответствии с чем разные большие и малые расы занимают разное положение на ветвях такого древа. Выявление архаичных и прогрессивных черт среди морфологических признаков носит субъективный характер, благодаря чему схемы расовых классификаций такого рода очень многообразны. Но самым большим недостатком подхода
к классификации рас исходя из их происхождения является попытка расположить расы на разных уровнях эволюционного древа, т.е. признание их биологической неравноценности.
Кроме того, данные палеоантропологических исследований показывают, что вплоть до верхнего палеолита на территориях, обитаемых людьми, практически нигде не сформировались расовые типы человека, с которыми были бы генетически связаны современные большие расы. Это подтверждает анализ верхнепалеолитических находок скелетов людей современного физического типа из сунгирских погребений (Россия), живших приблизительно 26 тыс. лет назад. Все черепа, принадлежащие им, характеризуются мозаичным сочетанием расовых признаков и не могут быть отнесены ни к одной из современных рас. Этим данным соответствует и описание ископаемого скелета из Южной Калифорнии, пролежавшего в земле 21,5 тыс. лет и характеризующегося отсутствием выраженных монголоидных черт, несмотря на то что аборигенным населением Америки являются монголоиды.
Только более поздние мезолитические находки свидетельствуют о формировании у человека расовых признаков. Так, известны мезолитические черепа с территории Северной Африки возрастом 10—8 тыс. лет с явными признаками не просто негроидной, а малой эфиопской расы. Сходные данные получены на территории Европы и в других регионах. Все это указывает на то, что процесс формирования расовых признаков — довольно поздний, идущий параллельно в разных регионах на рубеже верхнего палеолита — мезолита на фоне исходной разнородности расовых признаков у человека современного физического типа.
Первичное появление на протяжении эволюции признаков малых, а не больших рас позволяет сделать вывод о том, что европеоидная, монголоидная и негроидная расы имеют мозаичное происхождение
и представляют собой крупные популяции, объединенные не столько общностью происхождения, сколько климатогеографическими характеристиками условий существования и адаптивностью большинства основных признаков.
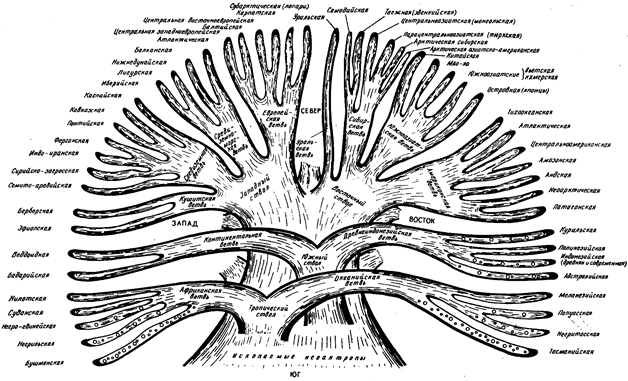
Рис. 15.8. Расовая классификация человечества с попыткой реконструкции происхождения рас
Это заключение хорошо согласуется с популяционной концепцией рас.
Суть ее заключается в следующем. Если принять, что большие расы человека представляют собой огромные популяции, то малые расы — субпопуляции больших, локальные естественные общности людей внутри которых — конкретные этнические образования (нации, народности) — являются более малыми популяциями. Если предположить при этом, намеренно упрощая ситуацию, что этносы не разделяются на элементарные популяции, и считать их просто состоящими из конкретных особей, то получится сложная структура, включающая в себя четыре уровня иерархии (рис. 15.9).
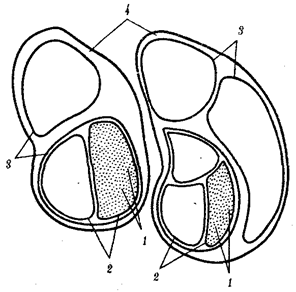
Рис. 15.9. Расы как выражение генетического полиморфизма человечества:
1
—отдельные индивидуумы, 2—
этносы, 3—
малые расы, 4—
большие расы
На основании исследований распределения различных групп крови и белков в популяциях человека произведено сравнение доли каждого из четырех уровней меж- и внутрипопуляционных различий в общем объеме генетического полиморфизма человека по этим признакам.
| Уровни различий
|
Доля генетического разнообразия, %
|
Индивидуумы
Этносы
Малые расы
Большие расы
Итого
|
84
5
3
8
100
|
Таким образом, от тотального генетического полиморфизма человечества расовые признаки составляют только 8%, в то время как основная доля генетического разнообразия определяется многообразием отдельных индивидуумов. Иными словами, немец может быть генетически гораздо ближе к полинезийцу, чем к другому немцу, живущему в соседней квартире. Изучение геногеографии популяций человека показало, что географическое распределение частот генов групп крови системы АВ0, MN, Lutheran, Duny, Diego и др., а также различных форм ферментов и иммуноглобулинов не соответствует ареалам расселения ни одной из рас. Так, по группам крови АВ0 и MN жители Европы оказываются ближе к африканцам, в то время как по системе иммуноглобулинов они ближе к монголоидам Азии. Сходные результаты получены и в отношении распределения в популяциях вариантов митохондриальной ДНК.
Эти данные свидетельствуют о том, что биохимический полиморфизм человека
эволюционно возник раньше и развивался дольше по сравнению с возникновением комплексов расовых признаков. Из этого следует, что расы не представляют собой особых изолированных групп людей, характеризующихся наборами специфических генов. Расовые же характеристики являются не более чем отдельными проявлениями общего генетического полиморфизма, выражающегося в первую очередь в сложных морфологических признаках. Некоторые из них адаптивны, другие сформировались на основе коррелятивной изменчивости, но все они касаются лишь ряда второстепенных особенностей (цвета кожи, волос, глаз и т.д.) и не затрагивают таких общечеловеческих признаков, как морфология головного мозга, а также строение и функции руки как органа труда.
На основании определения числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния
между ними. Эта величина для больших рас человека составляет 0,03. Она гораздо ниже цифр, характерных для истинных подвидов (0,17—0,22), и еще более мала по сравнению с межвидовым расстоянием (0,5—0,6 и более). В животном мире генетическое расстояние, равное 0,03, соответствует обычно генетическим отличиям местных популяций друг от друга. Все эти данные свидетельствуют о том, что понятие расы условно, второстепенно и не позволяет подводить под иерархическую классификацию рас глубокую биологическую, а значит, и социальную базу.
15.4.2. Адаптивные экологические типы человека
Человечество, заселившее уже около 15 тыс. лет назад все более или менее благоприятные для жизни природно-географические зоны, встретилось с необходимостью адаптироваться к самым разнообразным условиям существования. Адаптации человека к среде, как уже указывалось, проявляются в основном на социальном уровне, однако человечество на ранних этапах эволюции подвергалось непосредственному действию биотических и абиотических экологических факторов в значительно большей степени по сравнению с современной эрой научно-технического прогресса. Комплексы таких факторов имели разнонаправленное действие на человеческие популяции. В результате в разных климатогеографических зонах сформировались разнообразные адаптивные типы людей.
Адаптивный тип
представляет собой норму биологической реакции на комплекс условий окружающей среды и проявляется в развитии морфофункциональных, биохимических и иммунологических признаков, обеспечивающих оптимальную приспособленность к данным условиям обитания.
В комплексы признаков адаптивных типов из разных географических зон входят общие
и специфические
элементы. К первым относят, например, показатели костно-мускульной массы тела, количество иммунных белков сыворотки крови человека. Такие элементы повышают общую сопротивляемость организма к неблагоприятным условиям среды. Специфические элементы отличаются разнообразием и тесно связаны с преобладающими условиями в данном месте обитания — гипоксией, жарким или холодным климатом. Именно их сочетание служит основанием к выделению адаптивных типов: арктического, тропического, зоны умеренного климата, высокогорного, пустынь и др.
Разберем особенности условий жизни человеческих популяций в различных климатогеографических зонах и адаптивные типы людей, сформировавшиеся в них.
Условия обитания в Арктике характеризуются постоянно низкими температурами воздуха, скудной растительностью, богатством животного мира и сезонной периодичностью поступления продуктов питания: растительная пища доступна только в короткие летние месяцы, а животная — в периоды нереста рыбы, гнездования птиц, размножения оленей и морского зверя. Продукты питания богаты витаминами, белками, жирами и микроэлементами, но бедны углеводами растительного происхождения. Воздух и почва в Заполярье содержат очень мало микроорганизмов, низкие температуры препятствуют сохранению цист патогенных простейших, яиц и личинок гельминтов.
Из перечисленных факторов наибольшее влияние на формирование комплекса признаков арктического адаптивного типа
оказали, по-видимому, холодный климат и преимущественно животная пища. Арктическому комплексу признаков свойственны относительно сильное развитие костно-мускульного компонента тела, большие размеры грудной клетки, высокий уровень гемоглобина, относительно большое пространство, занимаемое костным мозгом, повышенное содержание минеральных веществ в костях, высокое содержание в крови белков, холестерина, повышенная способность окислять жиры. Среди аборигенов Арктики почти не встречаются лица с астеническим телосложением.
В целом арктический тип характеризуется усиленным энергетическим обменом, который отличается стабильностью показателей в условиях переохлаждения. Имеют свои особенности и механизмы терморегуляции. Так, при одинаковой степени охлаждения у канадских индейцев резко падает температура кожи, но уровень обмена веществ меняется незначительно, а у пришлого белого населения наблюдается меньшая степень снижения кожной температуры, но появляется сильная дрожь, т.е. интенсифицируется обмен.
В тропиках и субтропиках располагается чуть ли не большая часть Ойкумены. Этот регион отличается в целом большим количеством тепла и влаги и сглаженностью сезонных колебаний условий обитания. Вместе с тем благодаря особенностям рельефа наблюдается значительная контрастность распределения тепла и влаги — массивы влажных лесов нередко соседствуют с засушливыми плато, обширными равнинами и редколесьем. В экваториальной и субэкваториальной областях сосредоточены огромные количества растительной биомассы. Влажные тропические леса относительно бедны животными, тогда как в саваннах животный мир разнообразен и включает крупных стадных животных, издавна используемых человеком как объект охоты. Почва, воздух и вода содержат большое количество микроорганизмов, яиц гельминтов и цист патогенных простейших. Богатство и разнообразие животного мира обеспечивает существование огромного количества промежуточных и окончательных хозяев биогельминтов и переносчиков возбудителей трансмиссивных заболеваний. К преобладающим экологическим факторам, под влиянием которых формировался комплекс признаков тропического адаптивного типа,
относят жаркий влажный климат и рацион с относительно низким содержанием животного белка.
В тропической области наблюдается исключительно широкая вариабельность групп населения в расовом, этническом и экономическом отношениях. Это проявляется в поразительном размахе изменчивости, например по соматическим признакам. Тем не менее преобладающие экологические факторы, особенно климатический, способствовали образованию определенного комплекса морфофизиологических признаков обитателей тропиков и субтропиков. К характерным признакам тропического типа относят удлиненную форму тела, сниженную мышечную массу, относительное уменьшение массы тела при увеличении длины конечностей, уменьшение окружности грудной клетки, более интенсивное потоотделение за счет повышенного количества потовых желез на 1 см2
кожи, низкие показатели основного обмена и синтеза жиров, сниженную концентрацию холестерина в крови.
Антропологическое изучение современных обитателей зоны умеренного климата под углом зрения формирования биологических механизмов адаптации к природным условиям затруднено, так как значительная часть людей проживает в промышленно развитых странах с большой долей городского населения. Результаты наблюдения позволяют, однако, судить о том, что и в этом случае в процессе исторического развития человеческих популяций сформировался комплекс признаков, соответствующий особому адаптивному типу умеренного пояса.
По соматическим показателям, уровню основного обмена население умеренного пояса занимает промежуточное положение между коренными жителями арктического и тропического регионов. Это соответствует условиям биогеографической среды в зоне умеренного климата. Для нее характерны неравномерное распределение районов, отличающихся по количеству тепла и влаги, типу растительности (от сухих степей и полупустынь до тайги), богатству животного мира. Вместе с тем температура и влажность воздуха здесь не достигают экстремальных величин, хорошо выражен сезонный ритм биоклиматических условий.
Условия высокогорья для человека во многих отношениях экстремальны. Их характеризуют низкое атмосферное давление, сниженное парциальное давление кислорода, холод, относительное однообразие пищи. Основным экологическим фактором формирования горного адаптивного типа
явилась, по-видимому, гипоксия. У жителей высокогорья независимо от климатической зоны, расовой и этнической принадлежности наблюдаются повышенный уровень основного обмена, относительное удлинение длинных трубчатых костей скелета, расширение грудной клетки, повышение кислородной емкости крови за счет увеличения количества эритроцитов, содержания гемоглобина и относительной легкости его перехода в оксигемоглобин.
15.4.3. Происхождение адаптивных экологических типов
Человечество возникло в тропической зоне Африки, и следует полагать, что поэтому наиболее древним и исходным для остальных экологических типов человека является тропический тип.
Об этом свидетельствует и наибольший полиморфизм морфофункциональных признаков человека в рамках этого типа на территории Центральной и Северо-Восточной Африки. В связи с этим уместно обратиться к учению Н.И. Вавилова о центрах происхождения и многообразия культурных растений, в соответствии с которым именно в зонах первоначального вовлечения биологических видов в социальную среду наблюдается наиболее выраженный наследственный полиморфизм местных популяций по большому количеству признаков.
Об этом свидетельствует и то, что именно в тропической Африке в непосредственной близости друг от друга, т.е. под действием сходных экологических факторов, обитают наиболее низкорослые и наиболее высокорослые племена, например пигмеи, готтентоты и бушмены, с одной стороны, и массаи — с другой. Пределы изменчивости африканских популяций, например, по росту и массе тела таковы, что в них укладываются все известные на Земле человеческие популяции. Основные же черты тропического типа в Африке, отмеченные выше, остаются неизменными. Они же характеризуют монголоидные популяции Индокитая, Малайского архипелага и некоторые группы индейских племен зоны влажных тропических лесов Центральной и Южной Америки.
Это указывает на то, что тропический адаптивный тип развивается в результате асинхронного параллелизма в эволюции человека, причем вначале на африканском континенте, а потом и в других областях (см. раздел 3.13). Отсюда следует и еще один вывод: адаптивный тип формируется на фоне расогенеза и вне зависимости от него. В процессе адаптогенеза в популяциях человека можно проследить и проявление закона гомологических рядов (см. разд. 13.3.5). Это выражается, например, в том, что в бассейне Меконга в Индокитае и на острове Суматра имеются пигмеоидные популяции, по антропометрическим признакам соответствующие африканским пигмеям.
Экологический тип умеренного
пояса сформировался на базе исходного генетического и фенотипического полиморфизма тропического типа при расселении популяций человека в умеренных зонах Евразии и позже — Северной Америки. Он оформился в рамках двух больших рас: европеоидной и монголоидной.
При заселении человеком арктической
зоны в Евразии и Северной Америке произошло формирование арктического типа. Независимо от него среди индейцев Южной Патагонии и Огненной Земли в приантарктической зоне Южной Америки возникли популяции индейцев, по основному комплексу признаков соответствующие арктическому типу. Это еще один убедительный пример параллелизма эволюции человеческих популяций и реализации закона гомологических рядов, а также доказательство вторичности адаптивных типов по отношению к большим расам человечества.
Формирование горного
адаптивного типа иллюстрирует общие закономерности адаптогенеза, отмеченные выше. Этот тип также развился независимо от расовой и этнической принадлежности популяций — среди европеоидов Альп, Кавказа, Памира и Гималаев, а также в монголоидных популяциях Тибета, Тянь-Шаня и Анд. В связи с тем что высокогорья заселялись человеком в последнюю очередь, горный экологический тип является по происхождению самым молодым. Интересно, что, несмотря на особенно выраженную расовую и этническую разнородность этого типа, комплекс основных признаков его является монолитным. Однако в отличие от других типов, вероятно, именно горный проявляется в основном только на фенотипическом уровне и не имеет наследственной природы. Об этом свидетельствует то, что количество эритроцитов в крови и объем грудной клетки людей, переселяющихся в условия высокогорья и обратно, могут меняться на протяжении жизни одного поколения.
Следовательно, адаптивные типы человека не только отражают его прошлое, но формируются и в настоящем, а разные типы имеют и разную по длительности историю.
В настоящее время пока еще на фенотипическом уровне идет формирование адаптивного типа человека городской среды,
который характеризуется широкой лабильностью психических реакций, обеспечивающих способность переживать состояния постоянного стресса, и рядом морфофизиологических особенностей, оптимальных для жизни в специфических условиях города (см. разд. 17.3.2).
Формирование экологических типов человека в значительной степени обеспечило всесветное расселение людей. Меняющаяся среда обитания под действием антропогенных факторов ставит перед популяциями человека новые задачи, решаемые и сегодня за счет не только социальных, но и биологических адаптации. Схему эволюционных взаимоотношений больших рас и адаптивных типов человека см. на рис. 15.10.
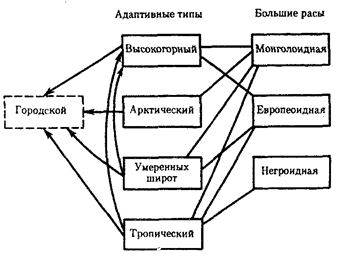
Рис. 15.10. Адаптивные типы человека и большие расы
Живые существа, населяющие территории с разнообразными условиями обитания, испытывают на себе влияние последних и сами оказывают действие на окружающую среду. Закономерности взаимоотношений организмов и среды их обитания, законы развития и существования биогеоценозов, представляющих собой комплексы взаимодействующих живых и неживых компонентов в определенных участках биосферы, изучает специальная биологическая наука — экология.
Экологические закономерности проявляются на уровне особи, популяции особей, биоценоза, биогеоценоза. Биоценозом
(сообществом организмов) называют пространственно ограниченную ассоциацию взаимодействующих растений и животных, в которой доминируют определенные виды или физический фактор. Предметом экологии, таким образом, являются физиология и поведение отдельных организмов в естественных условиях обитания (аутэкология
), рождаемость, смертность, миграции, внутривидовые отношения (динамика популяций
),
межвидовые отношения, потоки энергии и круговороты веществ (синэкология
).
К основным методам экологии относят полевые наблюдения, эксперименты в природных условиях, моделирование процессов и ситуаций, встречающихся в популяциях и биоценозах.
Среда —
это совокупность элементов, которые действуют на особь в месте ее обитания. Элемент среды, способный оказывать прямое влияние на живой организм хотя бы на одной из стадий индивидуального развития, называют экологическим фактором.
В соответствии с распространенной и удобной классификацией экологические факторы делят на биотические
и абиотические,
хотя указанное деление до некоторой степени условно. Абиотический фактор — температура — может, например, регулироваться изменением состояния популяции организмов. Так, при температуре воздуха ниже 13°С интенсифицируется двигательная активность пчел, что повышает температуру в улье до 25—30°С. Учитывая социальную сущность человека, проявляющуюся в его активном отношении к природе, целесообразно выделение также антропогенных экологических факторов.
По мере роста народонаселения и технической вооруженности человечества удельный вес антропогенных экологических факторов неуклонно возрастает.
Согласно другой классификации, различают первичные
и вторичные периодические экологические
факторы. С действием первичных факторов жизнь столкнулась на ранних стадиях эволюции. К ним относят температуру, изменение положения Земли по отношению к Солнцу. Благодаря им в эволюции возникла суточная, сезонная, годичная периодичность многих биологических процессов. Вторичные периодические факторы являются производными первичных. Например, уровень влажности зависит от температуры, поэтому в холодных областях планеты атмосфера содержит меньше водяных паров.
Непериодические факторы
действуют на организм или популяцию эпизодически, внезапно. К ним относят стихийные силы природы — извержение вулкана, ураган, удар молнии, наводнение, а также хищника, настигающего жертву, и охотника, поражающего цель- Благодаря многообразию экологических факторов наблюдается закономерное расселение видов по планете. Колебания интенсивности их действия проявляются в исчезновении некоторых видов с определенных территорий, изменении плотности популяций, показателей рождаемости, смертности. Под влиянием экологических факторов в эволюции сложились такие адаптивные модификации, как зимняя или летняя спячка, диапауза.
Любая особь, популяция, сообщество испытывают одновременное воздействие многих факторов, но лишь некоторые из них являются жизненно важными. Такие факторы называют лимитирующими,
их отсутствие или наличие в концентрации ниже и выше критических уровней делает невозможным освоение среды организмами определенного вида. Благодаря наличию лимитирующих экологических факторов для каждого биологического вида существуют оптимум
и пределы выносливости.
Так, устрицы наилучшим образом развиваются в воде с содержанием солей 1,5—1,8%. При снижении концентрации солей до 1,0% более 90 % личинок погибает в течение двух недель, а при концентрации 0,25% все поголовье их гибнет за одну неделю. Повышение концентрации соли по сравнению с оптимальной величиной также оказывает неблагоприятное действие на устриц. В общем виде зависимость выживаемости организмов определенного вида от интенсивности лимитирующего экологического фактора представлена графически на рис. 16.1. Взаимодействие нескольких экологических факторов усложняет картину. Так, некоторые виды тропических орхидей в природе при относительно высокой температуре воздуха растут только в тени. При искусственном понижении температуры окружающего воздуха они прекрасно развиваются в условиях прямой инсоляции.
Способность вида осваивать разные среды обитания выражается величиной экологической валентности.
Виды с малой экологической валентностью называют стенотопными, с
большой — эвритопными.
Эвритопные виды могут быть представлены несколькими экотипами —
разновидностями, приспособленными к выживанию в средах, различающихся по некоторым факторам. Так, сложноцветное растение тысячелистникAchillea millefolium
образует равнинные и горные экотипы. При выращивании горного экотипа в равнинных условиях растения сохраняют присущие им особенности на протяжении ряда поколений.
Рис.16.1. Интенсивность действия экологического фактора и выживаемость вида
Всю полноту взаимодействий и взаимозависимости живых существ и элементов неживой природы в области распространения жизни отражает концепция биогеоценоза.
Биогеоценоз —
это динамическое и устойчивое сообщество растений, животных и микроорганизмов, находящееся в постоянном взаимодействии и непосредственном контакте с компонентами атмосферы, гидросферы и литосферы. Биогеоценоз состоит из биотической (биоценоз
) и абиотической (экотоп
) частей, которые связаны непрерывным обменом веществом, и представляет собой энергетически и вещественно открытую систему (рис. 16.2). В него поступают энергия Солнца, минеральные вещества почвы, газы атмосферы, вода. Из него выделяются теплота, кислород, углекислый газ, биогенные вещества, переносимые водой, перегной.
Биогеоценоз содержит следующие обязательные компоненты (рис. 16.3): 1) абиотические неорганические и органические вещества среды; 2) автотрофные организмы — продуценты биотических органических веществ; 3) гетеротрофные организмы (консументы) — потребители готовых органических веществ первого (растительноядные животные) и следующих (плотоядные животные) порядков; 4) детритоядные организмы — редуценты-разрушители, разлагающие органическое вещество.
Как через любую диссипативную (т.е. рассеивающую энергию) систему, через биогеоценоз протекает регулируемый поток энергии. Эта энергия затрачивается на обеспечение постоянного круговорота веществ, поддержание целостности системы и обеспечение ее эволюции. Энергия проходит через серию трофических уровней, являющихся звеньями цепей питания.
Рис. 16.2. Биогеоценоз — открытая экологическая система
Первичным источником энергии служит солнечное излучение, энергия которого составляет 4,6 • 1026
Дж/с (1,1 • 1026
кал/с). 1/2000000 этого количества энергии достигает поверхности Земли, при этом 1,0—2,0% ассимилируются растениями, 30—70% поглощенной энергии используется ими для обеспечения собственной жизнедеятельности и синтеза органических веществ.
Энергия, накопленная в растительной биомассе, составляет чистую первичную продукцию
биогеоценоза. Фитобиомасса используется в качестве источника энергии и материала для создания биомассы потребителей первого порядка — растительноядных животных и далее по пищевой цепи. Количество энергии, расходуемой на поддержание собственной жизнедеятельности, в цепи трофических уровней растет, а продуктивность падает. Обычно продуктивность последующего трофического уровня составляет не более 5—20% предыдущего. Это находит отражение в соотношении на планете биомасс растительного и животного происхождения.
Так, суммарная биомасса организмов, обитающих на суше, составляет примерно 3 • 1012
т. Лишь 1—3% этого количества — зообиомасса. Масса животного вещества, приходящегося на людей, составляет около 0,0002% от суммарной массы живого вещества планеты. Объем энергии, необходимый для обеспечения жизнедеятельности организма, растет с повышением уровня морфофунк-циональной организации. Соответственно количество биомассы, создаваемой на более высоких трофических уровнях, снижается. Например, в разных биогеоценозах 95—99,5% зообиомаесы приходится на беспозвоночных животных.
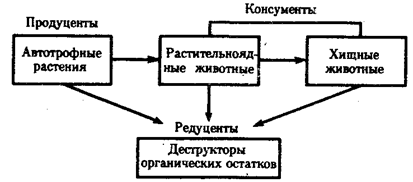
Рис. 16.3. Живые организмы — компоненты биогеоценоза
Прогрессивное снижение ассимилированной энергии в ряду трофических уровней находит отражение в структуре экологических пирамид.
Продукция живого вещества растительноядными животными составляет в данном случае 12,5%, а человеком — 0,6% продукции растений. Снижение количества доступной энергии на каждом последующем трофическом уровне сопровождается уменьшением биомассы и численности особей. Таким образом, пирамиды биомассы и численности организмов для данного биогеоценоза повторяют в общих чертах конфигурацию пирамиды продуктивности.
Размеры биогеоценозов, выделяемых экологами, различны. Совокупности определенных биогеоценозов образуют главные природные экосистемы,
имеющие глобальное значение в обмене энергии и вещества на планете. К ним относят: 1) тропические леса; 2) леса умеренной климатической зоны; 3) пастбищные земли (степь, саванна, тундра, травянистые ландшафты); 4) пустыни и полупустыни; 5) озера, болота, реки, дельты; 6) горы; 7) острова; 8) моря.
Главным компонентом биогеоценоза, от состояния которого зависят его существование и изменения во времени, служит биоценоз.
Биоценозы отличаются по видовому составу, и важнейшей их характеристикой является постоянное прямое или опосредованное взаимодействие популяций организмов друг с другом. Влияние любой популяции распространяется до экологически отдаленных элементов биоценоза через взаимодействие с конкурентами, хищниками, жертвами. Так, насекомоядные птицы не оказывают прямого действия на растения, но, снижая численность насекомых, питающихся листьями или опыляющих растения, они тем самым воздействуют на воспроизведение фитобиомассы. Последнее существенно для состояния популяций и продуктивности растительноядных животных, хищников, паразитов. Экологические влияния отдельной популяции распространяются в биоценозе во всех направлениях, но по мере прохождения последовательных звеньев в цепи взаимодействия интенсивность влияния ослабевает.
Показателями структуры и функционирования биоценозов служат их видовой состав, число трофических уровней, первичная продуктивность, интенсивность потока энергии и круговоротов веществ. Структура биоценозов складывается в процессе эволюции, причем каждый вид организмов эволюционирует таким образом, чтобы занять в биоценозе определенное место. Совместное историческое развитие многих видов на одной территории способствует их специализации к использованию лишь части наличных пищевых ресурсов и ограниченному местообитанию. В результате достигается состояние взаимоприспособленности видов друг к другу, или коадаптации,
которая служит обязательным условием стабильности биоценоза.
В качестве примера рассмотрим ситуацию, возникшую в искусственном оз. Гатун, которое образовалось в начале XX столетия в зоне Панамского канала. В течение нескольких десятилетий биоценоз озера отличался стабильностью благодаря коадаптации организмов основной пищевой цепи: фитопланктон — зоопланктон — планктоноядные рыбы. Последние, поедая зоопланктон, снижали его численность, что способствовало поддержанию количества фитопланктона на достаточно высоком уровне. В 1967 г. случайно в озеро была интродуцирована хищная, прожорливая рыба туканаре. Она быстро сократила численность планктоноядных рыб, что привело к размножению зоопланктона и сокращению количества фитопланктона. Одновременно снизилась численность обитающих на озере крачек и зимородков, питающихся рыбой, и повысилась численность комаров, личинки которых прежде поедались рыбой.
Таким образом, появление нового вида вызвало серьезные нарушения в экономике биоценоза озера и временно дестабилизировало его структуру. В дальнейшем, по мере развития коадаптации, при измененном видовом составе стабильность биоценоза может восстановиться. Состояние коадаптации достигается даже между видами-антагонистами: хищником и жертвой, хозяином и паразитом.
Наиболее устойчивыми являются биогеоценозы, характеризующиеся: 1) большим видовым разнообразием, 2) наличием неспециализированных видов, 3) слабой степенью отграниченности от соседних экологических систем и 4) большой биомассой. Действительно, разнообразие видового состава биоценозов обеспечивает реальное существование не столько цепей, сколько сетей питания, поскольку на каждом трофическом уровне находятся организмы разных видов, способные заместить друг друга в выполнении функций биотического круговорота веществ при изменении экологической ситуации (рис. 16.4).
Неспециализированные виды, способные обитать в меняющихся условиях и использовать разные источники питания, объединяют разные трофические уровни экологической пирамиды, упрочивая тем самым ее структуру. Обмен видами между соседними биоценозами может обеспечить восстановление даже существенно нарушенного экологического равновесия. Большое количество вещества, накопленного в виде биомассы, обладает свойствами буферности, обеспечивая систему веществом и энергией при длительном действии неблагоприятных экологических факторов, например, во время полярной ночи в высоких широтах или при длительных сезонных наводнениях в странах с муссонным климатом.
Рис. 16.4. Пищевая сеть в биогеоценозе смешанного леса
Тесные коадаптации популяций разных видов, входящих в состав биоценоза, проявляются, как и любые другие эволюционные события; на фенотипическом уровне, но по существу они — результат микро- и макроэволюционных процессов, затрагивающих в первую очередь их генофонды. Поэтому экологический гомеостаз базируется на коадаптациях популяционных генофондов и проявляется как выражение свойства наследственности
на биогеоценотическом уровне. Приобретение экологической системой новых видов или их утрата, изменение скорости и объема круговорота веществ, связанное с изменениями генофондов популяций биоценоза, а также приспособление его в целом как системы к меняющимся экологическим факторам есть проявление свойства изменчивости.
Другие характеристики живых систем — обмен веществ, выступающий в биогеоценозе в виде биогенного круговорота, и самовоспроизведение, в результате которого на базе исходного биогеоценоза возможно возникновение дочерних экосистем, — также проявляются на этом уровне организации жизни. Благодаря этому в биогеоценозах реализуется и такое фундаментальное свойство живого, как способность эволюционировать.
Любая территория, пригодная к жизни по набору абиотических факторов, заселяется. Этот процесс называют сукцессией.
В соответствии с трофической структурой биоценоза первостепенная роль в освоении новых местообитаний принадлежит растительным организмам. Развитие растительности в местообитаниях, где прежде растений не было, обозначают как первичную сукцессию,
а в местах с предсуществовавшим, но разрушенным растительньм покровом — как вторичную.
В процессе сукцессии изменяются видовой состав биоценоза и характеристики местообитания. Вслед за растениями в сукцессию вовлекаются представители животного мира, а развивающийся био-геоценоз становится все более богатым видами; цепи питания в нем усложняются, разветвляются и превращаются в сети питания. Среди животных растет количество всеядных видов, активизируется функция редуцентов, возвращающих органическое вещество из почвы в состав биомассы, благодаря чему ее объем неуклонно растет.
Сукцессия завершается климаксом —
образованием сообщества, видовой состав которого в дальнейшем изменяется незначительно. Скорость сукцессии по мере приближения к состоянию климакса снижается. Процесс практически прекращается, когда добавление или исключение видов не приводит к изменению среды развивающегося биогеоценоза, т.е. между элементами биоценоза и физической средой по достижении климакса устанавливается равновесие.
Из наблюдений за заселением песчаных дюн или вновь образованных потоков лавы в результате первичной сукцессии, а также вырубок или заброшенных пашен в процессе вторичной сукцессии следует, что для достижения состояния климакса требуются сотни и тысячи лет. Климаксные сообщества, возникающие в результате сукцессии разных местообитаний, различаются по производимой биомассе: тропический лес, леса умеренной зоны, болота. Максимальный объем биомассы ограничивается климатом соответствующего района.
Примером сукцессии служит зарастание некрупных пресноводных водоемов. Последовательное отмирание и придонное отложение мелких планктонных организмов, донных водорослей, водоплавающих растений, сопровождаемые сменой преобладающих видов животных и микроорганизмов, обусловливают трансформацию водных биогеоце-нозов в биогеоценозы болотного типа.
Климаксные сообщества на протяжении определенного времени характеризуются состоянием устойчивого равновесия, что проявляется в их способности возвращаться в исходное состояние после кратковременных внешних воздействий, изменяющих условия существования, и противостоять этим воздействиям. Так, в одном из климаксных биогеоценозов при временном понижении осадков на 50 % по сравнению с их обычным количеством продукция фитобиомассы снижалась на 25%, а численность популяций растительноядных — всего на 10%. Устойчивость подобных сообществ зависит как от гомеостатических реакций организмов и популяций, так и от условий физической среды. В приведенном примере она могла быть обусловлена запасом влаги в почве и реакцией растений на засуху. Несмотря на высокую степень устойчивости биогеоценозов, глобальное изменение условий среды, связанное с эпохальными сменами климата, приводит и к эволюции климаксных экологических систем.
В настоящее время под действием антропогенных факторов климаксные экологические системы сменяются менее устойчивыми либо в связи с прямым их разрушением, либо за счет загрязнения окружающей среды.
Так, в районе Москвы почвенно-климатические условия соответствуют развитию биогеоценозов дубовых лесов, господствовавших здесь до XV—XVIII вв. Вырубка дубрав и хозяйственное освоение территорий привели к появлению на их месте обедненных биогеоценозов березово-осиновых и еловых лесов. Участки древних дубрав сохранились местами в старинных парках и на границе Москвы в ее северо-восточной части, но поддержание состояния относительного равновесия в них требует уже значительных материальных затрат.
Взаимная адаптация популяций разных видов, включенных в состав эволюционирующего биогеоценоза, представляет собой процесс их соотносительной эволюции, сопровождающейся направленными изменениями аллелофондов этих популяций. В результате система аллелофондов биогеоценоза в целом как уровня организации жизни изменяется.
Таким образом, эволюция биогеоценоза базируется на эволюции отдельных популяций разнообразных организмов, а результатом ее является возникновение сообщества, включающего в себя новые виды, каждый из которых выполняет присущую только ему функцию в целостной системе.
В настоящее время термином «экология человека» обозначают комплекс полностью еще не очерченных вопросов, касающихся взаимодействия человека с окружающей средой. Главной особенностью экологии человека как самостоятельной области науки служит ее междисциплинарный характер, так как в ней сходятся социологические, философские, географические, естественнонаучные, медико-биологические проблемы. Экология человека
изучает закономерности возникновения, существования и развития антропоэкологических систем, которые представляют собой сообщество людей, находящееся в динамической взаимосвязи со средой и удовлетворяющее благодаря этому свои потребности.
Размеры таких систем различны в зависимости от численности и характера организации человеческих популяций. Это могут быть изоляты, демы, нации, наднациональные ассоциации, различающиеся по способу производства, укладу жизни, наконец, человечество в целом. Большое значение в определении размера антропоэкологической системы имеют природные условия. Наиболее многочисленные современные популяции, объединяющие более 80% человечества, обитают на 44% суши в области тропических лесов и саванн, а также в зоне умеренного пояса с кустарниковой растительностью или смешанными лесами.
Засушливые земли и зона пустынь, на которые приходится 18% суши, являются местом обитания 4% населения.
Главной отличительной чертой антропоэкологических систем по сравнению с природными экосистемами служит наличие в их составе человеческих сообществ,
которым в развитии всей системы принадлежит доминирующая роль. Сообщества людей различаются по способу производства материальных ценностей и структуре социально-экономических отношений, от чего зависят способ организации труда, объем и способ распределения производимой продукции между членами сообщества. Активностью сообществ людей на занимаемой территории определяется уровень воздействия их на окружающую среду. Развивающиеся сообщества (например, в период индустриализации) характеризуются наряду с ростом численности населения, увеличением потребностей его в продуктах питания, сырье, водных ресурсах, размещении отходов. Это повышает нагрузку на природную среду, интенсифицирует использование биотических и абиотических факторов.
В процессе существования антропоэкологических систем взаимодействие людей и природной среды осуществляется по двум главным направлениям. Во-первых, происходят изменения биологических и социальных показателей отдельных индивидуумов и сообщества в целом, направленные на удовлетворение требований, предъявляемых человеку средой. Во-вторых, осуществляется перестройка самой среды для удовлетворения требований человека. На протяжении истории человечества соотношение названных изменений сдвигалось в сторону преобладающей роли второго направления. Естественная среда, в которой зарождалось человечество, в результате перехода к культурному земледелию и скотоводству уступила место частично очеловеченной среде
сельских жителей. С возникновением городов современного типа произошел переход к существованию сообществ людей в полностью очеловеченной среде,
границы распространения которой неуклонно расширяются.
Общим результатом биологических и социальных процессов в антропоэкологических системах служит индивидуальная и групповая приспособленность человеческих сообществ к жизни в средах обитания, различающихся по природным условиям, формам хозяйствования и культуры. Особенность такой приспособленности в отличие от приспособленности к среде популяций любых других живых организмов состоит в том, что человек адаптируется к условиям жизни не только физиологически, но прежде всего экономически, технически, эмоционально. Различные стороны и направления индивидуальной и групповой адаптации человека, вся совокупность условий жизни и экологических связей людей являются предметом изучения экологии человека. Именно это делает ее междисциплинарной наукой. В курсе биологии допустимо ограничиться рассмотрением отдельных вопросов, имеющих непосредственное отношение к задачам охраны здоровья людей. Среди них важное значение принадлежит биологической изменчивости популяций людей в связи с биогеографическими особенностями среды, а также медико-биологической характеристике антропогенных экологических систем. Экологические вопросы паразитологии включены в соответствующий раздел учебника и излагаются ниже.
Благодаря наиболее высокому уровню организации человека, которого он достиг как биосоциальное существо, его взаимоотношения со средой обитания имеют существенные особенности (рис. 17.1).
Человек как экологический фактор в отличие от животных не только пользуется природными ресурсами, но, действуя на нее целенаправленно и осознанно, господствует над ней, адаптируя условия к
своим потребностям. Это достигается благодаря тому, что человек в отличие от растений и животных, использующих для своих потребностей энергию Солнца или органического вещества, накопленного в процессе фотосинтеза, применяет различные источники энергии, в том числе недоступные другим живым организмам: энергию ископаемого топлива, водных потоков, атомную и термоядерную. Энерговооруженность и техническая оснащенность человека неуклонно растут, и это позволяет ему заселять самые разнообразные условия обитания и снимает естественные барьеры ограничения численности человеческих популяций.
Рис. 17.1. Среда обитания людей
Человечество представляет собой единственный на Земле вид, всесветно обитающий, что превращает его в экологический фактор с глобальным распространением влияния. Благодаря воздействию на все главные компоненты биосферы влияние человечества достигает самых отдаленных экологических зон планеты. Печальным примером этому служит, в частности, обнаружение опасных пестицидов в печени пингвинов и тюленей, отловленных в Антарктиде, где никогда ни один из них не применялся. Еще одна особенность человека как экологического фактора заключается в активном, творческом характере его деятельности. Энергия, которой манипулируют люди, обращается ими на изменение среды обитания. Экологический оптимум существования человека на основе его биологических механизмов ограничен, и возможность широкого расселения достигается не путем изменения людьми их собственной биологии, а путем создания очеловеченной среды.
Созданием вокруг себя искусственной среды обусловливается также и специфика человека как объекта действия экологических факторов. Это действие всегда опосредовано результатами производственной деятельности людей. Естественные экосистемы вытесняются антропогенными экосистемами, абсолютно доминирующим экологическим фактором которых является человек. Среда обитания человека включает биоприродный и социально-культурный компоненты, или естественную
и искусственную среды.
В естественной и искусственной средах человек представлен как социальное существо.
Факторы естественной и искусственной среды оказывают на человека постоянное влияние. Результаты действия природных факторов, различающихся в разных районах обитаемой части планеты, на протяжении истории человечества проявляются в настоящее время в экологической дифференциации населения Земного шара, подразделении его на расы и адаптивные типы (см. § 15.4). Социальные факторы обусловливают образование и закономерную смену хозяйственно-культурных типов
сообществ людей. Они представляют собой комплекс хозяйства и культуры, характеризующий народы, которые различаются по происхождению, но обитают в сходных природно-ресурсных условиях и находятся на одинаковом социально-экономическом уровне.
В настоящее время на планете сосуществуют различные по времени возникновения, производительности труда, благосостоянию и демографическим показателям населения хозяйственно-культурные типы сообществ людей. В ограниченном числе сохраняется «присваивающий» тип с преобладанием экономической роли охоты, рыболовства, собирательства (пигмеи-охотники на территории Заира, племена аэта, кубу, обитающие в лесах Юго-Восточной Азии, отдельные группы индейцев в бассейне реки Амазонки). Достаточно широко представлены хозяйственно-культурные типы, экономическую основу которых составляют ручное (мотыжное) или плужное (пашенное) земледелие и скотоводство. В связи с научно-технической революцией в промышленно развитых странах сложились хозяйственно-культурные типы с высокоразвитым товарным земледелием и животноводством.
Формирование хозяйственно-культурных типов зависит от естественной среды обитания людей. Эта зависимость была наиболее сильна на ранних стадиях развития человеческого общества. Однако уже тогда и особенно в более поздние периоды развития человечества зависимость формирования хозяйственно-культурных типов от природных условий опосредовалась уровнем социально-экономического развития народа. На всех этапах истории общество активно приспосабливает природу к собственным нуждам. Инструментом такого приспособления, связующим звеном между естественной и очеловеченной средой служит трудовая деятельность людей, в процессе которой человек создает хозяйственную и культурную среду, от которой зависят образ жизни, показатели здоровья, структура заболеваемости,
Среда обитания человека представляет собой переплетение взаимодействующих естественных и антропогенных экологических факторов, набор которых различается в разных природно-географических и экономических регионах планеты. В таких условиях необходим единый интегральный критерий качества среды
с точки зрения ее пригодности для обитания человека. Согласно Уставу Всемирной, организации здравоохранения, принятому в 1968 г., этим критерием служит состояние здоровья населения.
В исследованиях по экологии человека термин «здоровье» используют в широком смысле как показатель полного физического и душевного благополучия.
Главная линия развития экологии человека в настоящее время нацелена на решение проблем управления средой, выработку путей рационального природопользования, оптимизации условий жизни людей в различных антропоэкологических системах.
Благодаря биосоциальной природе человека адаптации его к условиям обитания имеют отчасти биологическую, но главным образом социальную природу. В настоящее время преобладающее значение для освоения человеком новых сред обитания и создания лучших условий жизни в уже освоенных средах имеют социально-гигиенические мероприятия, результатом которых служит совершенствование средств и систем жизнеобеспечения, достижение состояния комфорта в местообитаниях людей. Адаптации создаются по отношению к факторам как природной, так и искусственной среды, поэтому они носят не только экологический, но и социально-экономический характер.
Каждый человек представляет собой индивидуальность, поэтому экологические и социально-экономические адаптации дополняются психологическими. Индивидуальные и групповые адаптации человека в отличие от биологических адаптации растений и животных обеспечивают наряду с выживанием и воспроизведением потомства выполнение им социальных функций, важнейшей из которых является труд.
Социально-гигиенические мероприятия, направленные на оптимизацию условий жизни и производственной деятельности, включают устройство жилищ и других помещений, конструкцию одежды, организацию питания и водоснабжения, рациональный режим труда и отдыха, сознательно направленную тренировку организма и многое другое. При этом исходят из принципа «разумного максимума» удобств, при котором поддерживается высокая работоспособность и сохраняется здоровье населения, но в то же время не допускаются изнеживание организма и чрезмерные экономические затраты.
В основе адаптации человека лежат социально-экономические механизмы, однако важная роль принадлежит также состоянию естественных приспособительных и защитных механизмов, составляющих биологическое наследство людей. Достаточно демонстративно эта роль выявляется при переходе в местообитания с экстремальными условиями, которые проявляются благодаря наличию на заселяемой территории экологического фактора или комбинации факторов, оказывающих на здоровье человека выраженное неблагоприятное действие.
Они могут складываться не только в естественных (Арктика, высокогорье), но и в антропогенных (крупные города) местообитаниях. Так, выходцев из зоны умеренного климата, прибывающих на работу в Арктику или Антарктиду, встречают суровый климат, необычные для средних широт атмосферные явления, резко пониженное количество микроорганизмов в почвах и воздухе, жизнь в относительно малочисленных, скученных коллективах. Как правило, такие люди по прибытии в Заполярье длительное время испытывают болезненные состояния и ощущения, усиливающиеся, например, при смене полярных дня и ночи. Они проявляются в повышении артериального давления и учащении пульса, которые сменяются затем понижением давления (иногда до уровня 70/30 мм рт. ст.) и урежением пульса. Эти явления, обозначаемые некоторыми исследователями как метеоневроз, сопровождаются падением работоспособности.
Описанная ситуация находит отражение в рекомендациях гигиенистов, ограничивающих продолжительность работы для вновь прибывших в Заполярье. Так, при температуре до —30°С и скорости ветра 4—8 м/с основной состав полярной станции может работать на открытом воздухе полный рабочий день, тогда как вновь прибывшие — не более 1 ч. У полярников количество лейкоцитов в крови обычно снижено до уровня 3000—3500 в 1 мм3
. В период смены состава зимовщиков при контакте с вновь прибывшими, как правило, наблюдается почти поголовная заболеваемость простудными и кишечными заболеваниями. Выявляются признаки утомления и даже истощения нервной системы — ухудшается оперативная память, снижается надежность работы человека, увеличивается продолжительность скрытого периода двигательных реакций.
По прошествии времени у части людей функциональные показатели возвращаются к нормальному уровню. У других они остаются измененными по сравнению с исходными значениями, наблюдавшимися до приезда в Арктику или Антарктику, однако восстанавливаются работоспособность и самочувствие. В таких случаях говорят об акклиматизации
людей к новым условиям обитания. Для растений и животных, перенесенных в необычную среду обитания, критерием акклиматизации служит их выживание, для людей — восстановление высокого уровня трудоспособности. Изменение физиологических механизмов при акклиматизации нередко имеет сложный характер.
Так, у адаптированных к жизни в холодном климате людей при охлаждении наблюдается изменение кровоснабжения кожных покровов. При этом степень изменения различается в разных частях тела. Например, у акклиматизированных в Заполярье людей на холоде тепловой поток с рук возрастает на 40%, тогда как с груди — на 19%. Таким образом, благодаря поддержанию достаточно высокой температуры сохраняется работоспособность рук.
В адаптациях человеческих популяций к новым экстремальным условиям, в которых они оказываются, огромную роль играет их исходный генетический полиморфизм
(см. § 12.3). В каждой популяции человека можно выделить разнородные конституциональные типы, отличающиеся друг от друга особенностями адаптации к новым условиям благодаря различиям их генотипических характеристик. Особенно отчетливо отличаются друг от друга типы «стайер» и «спринтер». Организм стайера довольно слабо приспособлен к выдерживанию мощных кратковременных нагрузок, однако после относительно короткой перестройки он способен переносить длительные равномерные воздействия экологических факторов в неадекватных условиях.
Тип «спринтер» может осуществлять мощные физиологические реакции в ответ на сильные, но непродолжительные воздействия экстремальными экологическими условиями. Длительное действие неблагоприятных факторов даже относительно небольшой интенсивности переносится спринтерами плохо. Наряду с этими крайними типами существует промежуточный вариант — «микст», характеризующийся средними адаптационными способностями.
У спринтеров и стайеров обнаруживаются различия по ряду антропометрических показателей, имеющих генетическую природу: масса тела, рост, объем грудной клетки, а также по функциональным признакам: величина артериального давления, емкость легких, соотношение форменных элементов крови, ее свертываемость и др. Отмеченные конституциональные типы отличаются друг от друга также различной заболеваемостью. Так, спринтеры более склонны иметь сердечно-сосудистые заболевания. Течение их в этой группе лиц более тяжелое. Тип «спринтер» оказывается более легко адаптирующимся в экстремальных экологических ситуациях на протяжении первых месяцев и лет после попадания в соответствующие условия. Особенности стайеров менее выигрышны в условиях акклиматизации, но по истечении указанного срока их состояние значительно улучшается.
Статистические обследования человеческих популяций в разных экологических условиях показали, что в г. Новосибирске среди людей со средним возрастом 30,5 лет 22% составляют спринтеры, 12% — стайеры, 66% — миксты. Среди людей, переселившихся в экстремальные условия БАМа, на протяжении 1-го года спринтеров было 32%, стайеров — 25%, микстов — 43%. В конце 2-го года жизни на БАМе соотношение типов сместилось резко в сторону стайеров и составило 17,6% спринтеров, 53% стайеров и 29,4% микстов. Это произошло в связи с тем, что большая часть людей, генетическая конституция которых не соответствует стайерам, покинула экстремальную зону, в результате чего среди оставшихся больше половины составили именно стайеры.
Знания о конституциональных особенностях и адаптивных возможностях стайеров и спринтеров имеют большое значение для осуществления мероприятий по регулированию и улучшению систем жизнеобеспечения человеческих популяций в регионах с экстремальными экологическими условиями.
Длительное существование групп людей на территориях, различающихся преобладающими климатическими, алиментарными и другими факторами, привело к образованию воспроизводящихся в ряду поколений комплексов признаков. Эти комплексы соответствуют экологическим типам людей и обусловливают более высокий уровень приспособленности к проживанию в определенной биогеографической среде за счет биологических механизмов. Такое положение свидетельствует об относительности понятия экстремальности природных условий в тех или иных регионах планеты. Вместе с тем перед человечеством стоит задача глубокого освоения и более плотного заселения малокомфортных территорий — пустынь и полупустынь, высокогорных, полярных, тропических районов, океанов и морей (на долю которых приходится 71% поверхности планеты). Космоса.
Решение этих задач связано с социально-экономическим прогрессом человечества.
17.3.1. Город
Город
во многом напоминает такие экологические системы, как пещерные, глубоководные и иные биогеоценозы, зависящие в основном от поступления в них энергии и вещества извне. Они полностью или частично лишены продуцентов и поэтому называются гетеротрофными.
Город от большинства природных экологических систем отличается следующими особенностями:
1) более интенсивным метаболизмом на единицу площади, для чего используется в первую очередь не солнечная энергия, а энергия горючих материалов и электричества;
2) более активной миграцией веществ, в которую вовлекается перемещение металлов, пластмасс и т.д., причем не столько в пределах системы, сколько на входе и на выходе из нее;
3) более мощным потоком отходов, многие из которых вообще не реутилизируются и являются более токсичными, чем естественное сырье, из которого они получены.
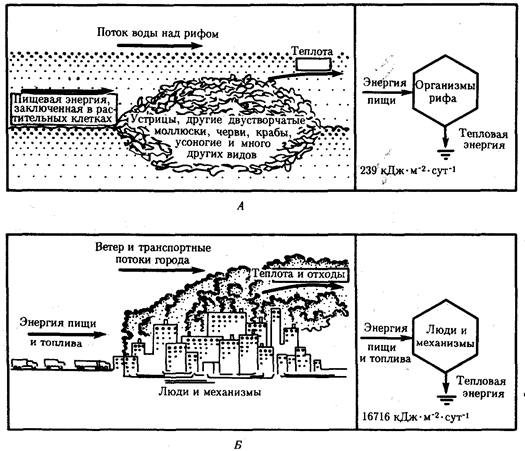
Рис. 17.2. Гетеротрофные-экологические системы. А —
устричная банка; Б—
современный город
Следовательно, для эффективного функционирования города как экологической системы необходима более тесная связь его с окружающей средой и большая зависимость его от нее. Действительно, хотя в большинстве городов имеются мощные зеленые насаждения, органическая продукция их не играет существенной роли в снабжении города. Кислород, выделенный ими, не покрывает его расходов на дыхание людей, животных, а главное — на технологические процессы промышленных предприятий.
Без постоянных поступлений пищи, строительных материалов, горючего, электричества и воды город вскоре прекратил бы существование. На рис. 17.2 представлены схемы двух экологических систем гетеротрофного типа: устричной банки и города. Обратите внимание на то, что 1 м2
городской системы потребляет в 70 раз больше энергии, чем соответствующая площадь естественного биогеоценоза; а также на более интенсивные потоки энергии и вещества на входе — выходе из системы.
Площадь суши, занятая в настоящее время городами, составляет 1—5% в разных районах мира. Однако воздействие их на окружающую среду огромно. Город может влиять на окружающие его биогеоценозы не только как потребитель органического вещества и кислорода, но и как мощнейший загрязнитель, действующий нередко на огромном расстоянии. Так, продукты питания и промышленное сырье могут завозиться в города с расстояний в несколько тысяч километров. Загрязнение воздуха вредными промышленными выбросами в Центральной Европе приводит к выпадению кислотных дождей в Скандинавии, что вызывает деградацию местных лесных биогеоценозов. В США подсчитано, что для обеспечения питанием города с населением в 1 млн. человек, занимающего 259 км2
территории, необходимо около 0,8 млн. га, или 8090 км2
пашни. В слаборазвитых странах города потребляют меньше вещества и энергии, однако отсутствие в них очистных сооружений на промышленных предприятиях и для бытовых отходов приводит обычно к более сильному воздействию на окружающую природу по сравнению с городами промышленно развитых стран.
Современный город сам не производит продуктов питания и других органических веществ, не обогащает воздух кислородом, почти не возвращает воду и неорганические материалы в круговорот веществ. Поэтому в широком экологическом смысле город не может считаться экологической системой. Для того чтобы рассмотреть город как биогеоценоз с характерными для него признаками устойчивости, саморегуляции и саморазвития, необходимо расширить его границы с учетом тех близких и отдаленных сред, которые определяют его жизнеобеспечение. Поэтому изучение экологических проблем городов необходимо вести не только в самих городах, но и далеко за их пределами.
17.3.2. Город как среда обитания людей
Для современного состояния развития человеческого общества характерна интенсивная урбанизация. Растет количество жителей больших городов. В странах с высокой плотностью населения происходит слияние соседних городов и образование обширных территорий с высоким уровнем урбанизации — мегаполисов.
Условия жизни в городах своеобразны: с одной стороны, в городе легче решаются проблемы трудоустройства, снабжения продуктами питания, медицинского обслуживания, с другой — в городах наиболее выражены преобразования человеком природной среды, что часто приводит к отрицательным последствиям.
Неблагоприятно действует на человека высокая плотность населения в городах, облегчающая циркуляцию возбудителей многих инфекционных и паразитарных заболеваний. Промышленные и бытовые отходы загрязняют почву, воду и воздушный бассейн. Аэрозольные загрязнения воздуха приводят к повышению облачности и образованию тумана, нарушают теплообмен таким образом, что города становятся своеобразными «тепловыми островами». Так, в центре Вашингтона последние весенние заморозки отмечаются в среднем 29 марта, а первые осенние — 10 ноября, в то время как на окраине города весенние заморозки кончаются на один месяц позже, а осенние начинаются на месяц раньше. Летний период в городах в целом оказывается гораздо более жарким, и температура в городах даже умеренных широт периодически поднимается до 40°С. При этом смертность среди населения, в особенности среди людей, страдающих хроническими сердечно-сосудистыми заболеваниями, может увеличиваться в 5 раз и более.
Высокая облачность и туманы в городах приводят к ослаблению освещенности, а также снижают интенсивность ультрафиолетового излучения, достигающего поверхности Земли. Недостаток света приводит к учащению случаев гиповитаминоза в и рахита у городских детей и снижает их сопротивляемость к простудным и детским инфекционным заболеваниям. Другими неблагоприятными факторами городской среды являются шум и вибрация, в результате именно здесь чаще встречаются поражения слухового аппарата и неврозы.
Высокий темп жизни, постоянные стрессы, гиподинамия горожан и высокая калорийность пищи способствуют нарушению функций нервной, сердечно-сосудистой систем, обмена веществ. Уровень заболеваемости в городах в 1,5%—2 раза выше, чем в сельской местности.
Города характеризуются также низким уровнем рождаемости, а рост их населения происходит в основном за счет притока людей из сельской местности.
17.3.3. Агроценозы
В отличие от городов агроценозы,
или сельскохозяйственные экосистемы, характеризуются основным компонентом — автотрофными организмами,
которые обеспечивают их органическим веществом и выделяют кислород. От естественных биогеоценозов они отличаются следующими особенностями.
1. Кроме солнечной энергии для поддержания агроценозов необходимы затраты дополнительной энергии: химической в виде удобрений, механической в виде работы мышц человека и животных, а также энергии горючих материалов и электричества.
2. Видовое разнообразие организмов резко снижено и представлено отдельными сельскохозяйственными культурами, иногда даже только одной, с сорняками и вредителями сельскохозяйственных растений, а также ограниченным количеством видов домашних животных.
3. Доминирующие виды растений и животных находятся под контролем искусственного отбора. Агроценозы организуются таким образом, чтобы получать максимальное количество продуктов питания. В настоящее время около 10% свободной от льда суши занято пахотными землями, еще 20% используются как пастбища.
Существует два основных типа сельскохозяйственных экосистем: экстенсивные
и интенсивные
агроценозы. Первые существуют с использованием в основной мышечной энергии человека и животных. Продукция этих систем используется для питания семей мелких фермеров и для продажи или обмена на местном рынке. Вторые связаны с крупными затратами химической энергии и машин. Продукты питания производятся здесь в количестве, превышающем местные потребности, и они вывозятся на продажу, играя важную роль в экономике.
Около 60% сельскохозяйственных угодий используются экстенсивно. Большая часть из них сосредоточена в странах Азии, Африки и Южной Америки. В ряде случаев они могут быть весьма сложными и гармонировать с природными экосистемами. Эффективность экстенсивных агроценозов может быть очень высокой, особенно в случаях культивирования на ограниченных территориях значительного количества видов растении и животных. Так, на о-вах Новая Гвинея и Филиппины население получает продукты питания, содержащие примерно 96 млн. кДж/га при затратах труда, эквивалентного б млн. кДж. Соотношение полученной и затраченной энергии соответствует 16 : 1.
Однако даже наиболее продуктивные экстенсивные агроценозы не могут производить достаточно много избыточных продуктов, чтобы снабжать ими большие города. Таким образом, неиндустриальное сельское хозяйство эффективно сберегает энергию, но оно малопродуктивно при учете количества продуктов, произведенных одним фермером.
Интенсивные агроэкосистемы занимают 40% обрабатываемых земель. Они сосредоточены в основном в Европе, Центральной и Северной Америке и в Австралии. Эффективность их столь высока, что 4% населения США, живущего в сельской местности, обеспечивают не только всю страну основными продуктами питания, но и экспорт.
Урожайность многих сельскохозяйственных культур, используемых в индустриальных агроценозах, в настоящее время достигает биологически возможного максимума. Характерно увеличение потерь сельскохозяйственных угодий в связи с эрозией почвы, а также ухудшением качества воды в результате стока поверхностных вод, насыщаемых пестицидами и минеральными удобрениями, в озера и реки.
В последние годы в индустриальном сельском хозяйстве наметились тенденции, направленные на совместное возделывание нескольких культур, уменьшение размеров посевных площадей с чередованием их с садами, водоемами, виноградниками, пастбищами и лесопосадками. На фоне применения технологий обработки почвы без глубокой вспашки, использования в основном органических удобрений и преимущественно биологических мер борьбы с вредителями и сорняками эти мероприятия способствуют созданию искусственных экосистем, по ряду показателей приближающихся к естественным биогеоценозам. Истощение почв и эрозия их, а также загрязнение окружающей среды при такой системе хозяйствования сводятся к минимуму.
Наиболее наглядными примерами влияния человека на процесс эволюции видов являются одомашнивание животных
и выведение сортов культурных растений.
Этот процесс продолжается целенаправленно не менее 70 тыс. лет. Существующая генотипическая изменчивость была использована для выведения тысяч сортов растений и пород животных. Таким образом, человек создал огромное разнообразие организмов, которые не могли бы быть получены естественным образом и существовать в естественной среде.
Предковые формы многих культур исчезли с лица Земли. Многие из сегодняшних видов так сильно отличаются от исходных, что их можно уже считать новыми видами антропогенного происхождения. Такова, например, кукуруза, которая в процессе эволюции под контролем человека в естественных условиях утратила способность к самостоятельному размножению: перед прорастанием ее семена обязательно должны быть освобождены от початка.
Параллельно с эволюцией собственно культурных растений человек стимулирует адаптации огромного количества видов сорных растений и животных-вредителей сельскохозяйственных культур, часто строго приуроченных к определенным культурам. Это вынуждает селекционеров выводить сорта растений, устойчивых к вредителям и болезням. Так, сорта садовой земляники отличаются по чувствительности к грибковому заболеванию — серой гнили, а маниока (распространенная тропическая культура) — даже по чувствительности к поеданию местной саранчой.
Интродукция (введение в культуру) растений и животных из отдаленных мест их естественных обитании часто оканчивается неудачей в связи с тем, что в новых биогеоценозах они не могут включиться в эволюционно отработанную систему биотического круговорота веществ, поэтому их успешное культивирование обычно возможно лишь в условиях садов и зоопарков с гарантированным постоянным уходом. Однако для некоторых видов новые условия обитания оказываются даже более благоприятными, чем на их родине, в результате чего возможно их эффективное включение в состав местной фауны и флоры и даже вытеснение некоторых конкурирующих аборигенных видов.
Так, совершенно новые водные экологические системы создало растение элодея канадская, которое попало в Европу через ботанические сады около 100 лет тому назад и встречается сейчас практически во всех пресных водоемах с медленным течением. Ее быстрое вегетативное размножение привело'в ряде мест даже к нарушению судоходства.
Столь же недолга история другого растения — высокого кустарника буддлейи Давида, завезенного в Европу из субтропических зон Центрального Китая. Это растение оказалось чрезвычайно устойчивым к низким зимним температурам, к загрязнению воздуха и способно к очень быстрому росту. В Европе оно занимает пустыри, свалки и заброшенные строительные площадки, создавая основу новых биогеоценозов в условиях, мало пригодных для роста большинства аборигенных растений. Цветы буддлейи посещают 20 местных видов бабочек, много видов пчел, мух и жуков. Листьями стали кормиться гусеницы более 12 видов европейских чешуекрылых, а некоторые виды других насекомых перешли к питанию лепестками ее цветков и семенами. 182
Не менее впечатляющие примеры преобразующей деятельности человека на экологические системы дает изучение флоры субтропических районов Западной Грузии, абсолютное большинство кустарников и особенно травянистых растений которой происходит из стран Восточной и Юго-Восточной Азии. Сельскохозяйственное и промышленное производство в Грузии создает новые комплексы условий, в которых растения, адаптированные к жизни в девственных колхидских лесах, не выживают. Широкие международные связи Грузии через портовые города и работа ботанических садов по культивированию иноземных растений привели к быстрому освоению ими новых территорий, причем в основном в садах, на полях, вдоль дорог, на железнодорожных насыпях и в зонах прямого действия промышленных предприятий. В устойчивые экологические системы колхидских лесов они проникнуть не смогли.
Интересно, что консументами в таких антропогенных биогеоценозах могут являться не только некоторые наиболее экологически пластичные насекомые, синантропные грызуны и воробьи. На территории Южного Закавказья описано несколько видов ящериц, популяции которых представлены исключительно самками, размножающимися партеногенетически. Происходят они от местных видов ящериц, обитающих в условиях среды, не измененной человеком. Переход к партеногенезу снижает возможности адаптации высокоорганизованных животных в естественных биогеоценозах, но облегчает возможности обитания в обедненных экологических системах, продуценты которых представлены практически исключительно иноземными видами растений.
Таким образом, рассмотренные примеры свидетельствуют о том, что как целенаправленная, так и непланируемая преобразовательная деятельность человека не только приводит к исчезновению отдельных видов животных и растений, но и является фактором эволюции популяций, видов и целых экологических систем практически во всех регионах, затронутых его хозяйственной деятельностью.
Болезни животных и человека можно классифицировать по этиологическому принципу как эндогенные
и экзогенные.
В основе эндогенных заболеваний лежат аномалии структуры или функционирования наследственного аппарата. Экзогенные заболевания имеют разную природу: это травмы, нарушения питания, авитаминозы и т.д. Кроме того, это болезни, вызываемые живыми организмами: вирусами, прокариотами и животными. Болезни, вызываемые вирусами и прокариотическими организмами, называют инфекционными. Болезни, вызываемые животными, называют инвазионными или паразитарными. .
Медицинская паразитология изучает особенности строения и жизненных циклов паразитов, взаимоотношения в системе паразит — хозяин, а также методы диагностики, лечения и профилактики инвазионных болезней.
В связи с тем что большинство паразитов человека относится к типу Простейшие Protozoa, а также к группе Черви (гельминты) — плоские Plathelminthes и круглые Nemathelminthes, — в рамках паразитологии выделяют разделы: медицинскую протозоологию
и медицинскую гельминтологию.
Немало животных, имеющих медицинское значение и в типе Членистоногие Arthropoda. Некоторые из них сами являются возбудителями заболеваний, другие — переносчиками возбудителей паразитарных и инфекционных болезней. Биологию членистоногих — возбудителей и переносчиков (клещей и насекомых) — изучает медицинская арахноэнтомология.
Паразиты могут обитать в любых органах человека, поэтому врач любой специальности может встречаться с паразитарными заболеваниями и обязан уметь распознавать их, лечить больных и проводить профилактику заражения паразитами (рис. 18-1).
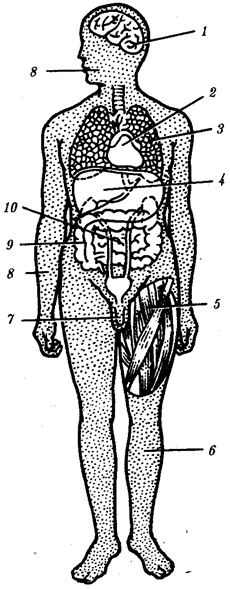
Рис. 18.1. Локализация паразитов в организме человека:
1
— головной мозг (эхинококк, цистицерки свиного цепня, трипаносома, токсоплазма и др.), 2 —
кровь (малярийный плазмодий, трипаносома и др;), 3 —
легкие (эхинококк, легочный сосальщик и др.), 4
—.печень (многие сосальщики, эхинококк, токсоплазма и др.), 5
— скелетные мышцы (трихинелла, цистицерки свиного цепня и др.), 6
— подкожная клетчатка (ришта и др.), 7
— мочеполовая система (шистосомы, трихомонада и др.), 8 —
кожа открытых участков тела (лейщмания и др.), 9
— толстая кишка (дизентерийная амеба, балантидий, власоглав и др.), 10
— тонкая кишка (лямблия, все цепни, лентецы, аскарида, острица и др.)
Организмы разных видов в биоценозах находятся в постоянном взаимодействии друг с другом. Существуют две основные формы межвидовых взаимодействий: антибиоз
и симбиоз.
Антибиоз —
невозможность сосуществования двух видов организмов, основанная на конкуренции прежде всего за источники питания. Примером служат взаимоотношения сапрофитных бактерий и ряда плесневых грибов. Первые способны быстро заселять среды, богатые органическими веществами, за счет интенсивного размножения, а вторые, значительно уступая им в этом, приобрели способность делать субстрат неблагоприятным для жизнедеятельности бактерий, выделяя в него продукты своего метаболизма — антибиотики.
В результате среда используется либо грибами, либо бактериями, успевшими попасть в нее и размножиться раньше.
Симбиоз
в переводе с греческого означает «сожительство». Формы симбиоза разнообразны. В некоторых случаях отношения между организмами разных видов являются взаимополезными настолько, что раздельное их существование вообще невозможно. Такой симбиоз называют мутуализмом.
Примером мутуалистических взаимоотношений является сожительство человека с микрофлорой его кишечника, основным компонентом которой являются разнообразные штаммы бактерий кишечной палочкиEscherichia coli.
Бактерии в таком сожительстве находят благоприятную среду обитания и неисчерпаемый источник питания. Нормальное же пищеварение в кишечнике человека и всасывание рядов витаминов возможно только при участии бактерий. После длительного лечения больных различными инфекционными заболеваниями с помощью антибиотиков у них нередко наряду с подавлением жизнедеятельности болезнетворных бактерий наблюдается состояние дисбактериоза —
гибель нормальных бактерий кишечника и усиленное размножение бактерий, нечувствительных к антибиотику, и микроскопических грибов, которые в свою очередь сами могут явиться причиной заболевания. Для восстановления нормальной кишечной микрофлоры часто необходимым является искусственное заселение пищеварительной системы человека симбионтными штаммами кишечной палочки.
Комменсализм —
форма симбиоза, при которой один вид использует остатки или излишки пищи другого, не причиняя ему видимого вреда. Часто комменсалы даже поселяются в теле хозяина, не снижая его жизнеспособности. Примером комменсалов являются непатогенные ротовая и кишечная амебы, живущие в пищеварительной системе человека и питающиеся бактериями.
При хищничестве
между организмами разных видов существуют только пищевые взаимоотношения, а пространственные отсутствуют. Хищники используют представителей другого вида для питания однократно, убивая их.

Рис. 18.2. Терракотовая статуэтка «паразита» – персонажа римской комедии «Паллиата» (184 г. до н.э.)
Наибольшее значение для медицины имеет паразитизм —
форма межвидовых взаимоотношений, при которой один вид использует другой как источник питания и среду обитания. В тех случаях, когда паразит не живет в организме хозяина, он посещает его для питания многократно.
Слово паразит
стало применяться в Древней Греции для обозначения пассивных участников жертвоприношений во время религиозных обрядов. Позже паразитами стали называть непрошеных гостей, а также персонажей драматических произведений, не выполняющих в действии серьезных функций (рис. 18.2). Понятие «паразит» в современном биологическом смысле стало применяться в Европе с XV-XVI вв.
Разные формы симбиотических взаимоотношений организмов не являются абсолютно стойкими и могут переходить друг в друга. Так, комменсалы могут становиться паразитами при ослаблении иммунитета хозяина. При нормальном питании и физическом здоровье хозяина некоторые паразиты могут долгое время не оказывать на него патогенного действия. Некоторые хищники, питающиеся мелкими животными, могут становиться паразитами крупных, и наоборот. Об этом подробнее см. разд. 18.5.
Условность классификации биотических связей и нечеткость их отграниченности друг от друга являются отражением эволюции не только взаимодействующих видов, но и самих межвидовых взаимоотношений.
Формы паразитизма чрезвычайно многообразны, и классификация их возможна по разным основаниям. С точки зрения обязательности паразитического образа жизни для данного вида различают истинный и ложный, а также облигатный и факультативный паразитизм.
При истинном
паразитизме взаимоотношения между паразитом и хозяином являются закономерными и имеют эволюционную основу. Паразитология изучает в основном феномен истинного паразитизма.
Ложный
паразитизм — явление для данного вида случайное. В нормальных условиях данный вид ведет свободный образ жизни. При попадании в организм хозяина ложный паразит может некоторое время сохранять жизнеспособность и нарушать жизнедеятельность хозяина. Примерами ложного паразитизма являются случаи обнаружения пиявок в носовой полости и носоглотке человека. Ложный паразитизм пиявок может привести хозяина к смерти в связи с закупоркой дыхательных путей или из-за носовых кровотечений, которые они могут вызвать.
Облигатный
паразитизм — паразитизм, являющийся обязательным для данного вида организмов. Абсолютное большинство видов паразитов относятся к этой группе.
Факультативные
паразиты способны вести свободный образ жизни, но, попадая в организм хозяина, проходят в нем часть цикла своего развития и нарушают его жизнедеятельность. Таковы многие виды синантропных мух, личинки которых могут нормально развиваться либо в пищевых продуктах человека, либо в его кишечнике, вызывая кишечный миаз
(см. разд. 21.2.4).
По времени контакта хозяина и паразита паразитизм бывает временным и постоянным. Временные
паразиты обычно посещают хозяина только для питания. Это в основном кровососущие членистоногие. Постоянные
паразиты подразделяются на стационарных и периодических.
Стационарные
паразиты всю жизнь проводят на хозяине или внутри него. Примерами являются вши, чесоточный клещ, трихинелла спиральная и многие другие. Периодические
паразиты часть своего жизненного цикла проводят в паразитическом состоянии, остальное время обитают свободно. Типичным паразитом такого рода является угрица кишечная.
Нередко паразитический образ жизни ведут только личинки, в то время как половозрелые формы являются свободноживущими. Паразитизм такого рода называют ларвальным
(личиночным). Примерами служат вольфартова муха, оводы и др. (см. разд. 21.2.4). Противоположное явление, когда паразитом является половозрелая форма, а личинка обитает в открытой природе, называют шлагинальным
паразитизмом. К паразитам этого типа относят, например, анкилостомид, личинки которых живут в почве, а взрослые стадии — в двенадцатиперстной кишке человека.
Особенно большое медицинское значение имеет классификация паразитов по их локализации в организме хозяина. Эктопаразиты
находятся на покровах хозяина. К ним относят кровососущих насекомых и клещей. Эндопаразиты
обитают внутри хозяина. Их подразделяют на паразитов, обитающих в полостных органах,
связанных с внешней средой (пищеварительная, дыхательная и мочеполовая системы), и паразитов тканей внутренней среды
(опорно-двигательный аппарат, система крови, соединительная ткань). Примерами первых являются аскарида, легочный сосальщик, урогенитальная трихомонада, вторых — ришта, малярийный плазмодий, лейшмании.
Любой подход к классификации паразитизма не дает возможность строго разграничить формы этого сложного экологического явления. Многие виды на протяжении жизненного цикла могут быть по отношению к разным хозяевам и ларвальными, и имагинальными паразитами. Так, сосальщики на начальных этапах развития ведут свободный образ жизни. Позже их личинки обитают в промежуточном хозяине, затем вновь образуются свободноживущие личинки, которые, обнаружив второго промежуточного или окончательного хозяина, паразитируют у него на половозрелой стадии.
В процессе жизнедеятельности нередко паразиты осуществляют миграцию в организме хозяина и способны таким образом вначале обитать в полостных органах, а затем перемещаться в ткани внутренней среды. Таковы трихинелла и свиной цепень. Возможен переход от эктопаразитизма к паразитированию в тканях внутренней среды. К таким видам относятся, например, личинки вольфартовой мухи.
Отсутствие четких границ между разными формами паразитизма отражает объективную ситуацию — эволюцию этого экологического феномена.
Своеобразной экологической группой паразитов являются сверхпаразиты.
В качестве среды обитания и источника питания ими используются другие паразитические организмы. Обычно сверхпаразиты еще более мелкие и низко организованы, чем паразиты (рис. 18.3). Они могут поражать как простейших, так и многоклеточных паразитов. Сверхпаразитизм — очень широко распространенное явление. Так, подсчитано, что только один вид свободноживущих бабочек — луговой мотылекLoxostege sticticalis —
является хозяином 40 видов паразитов, за счет которых существуют еще 12 видов сверхпаразитов. Среди сверхпаразитов, обитающих в паразитах человека, известны несколько видов микроспоридий, относящихся к классу споровиков и встречающихся в цитоплазме балантидия (см. разд. 19.2.3), в клетках паренхимы цепней (см. разд. 20.1.2) и в гонадах аскарид (см. разд. 20.2.1).
Сверхпаразиты имеют огромное экологическое значение, выполняя функции стабилизаторов численности популяций паразитов. Медицинское значение сверхпаразитов еще не изучено, но не исключено, что и в популяциях человека они могут выполнять роль факторов, сдерживающих численность паразитов.
Рис. 18.3. Сверхпаразиты:
А —
рачокErythrops
(I
) с паразитическим ракообразнымAspidophryxus
(2
),
на котором находится сверхпаразитAspidoecia normani
(3
);
Б —
наездникIxodiphagus sp.
откладывает яйцо в нимфу собачьего клеща
Паразитический образ жизни могут вести самые разнообразные организмы, не имеющие между собой ничего общего. Паразитизм известен в царстве Вирусы, среди прокариотических организмов, в царствах Грибы, Растения, Животные. Все вирусы являются паразитами. Их изучает отдельная наука — вирусология.
Паразитические прокариоты изучаются микробиологией,
паразитические грибы — микологией,
паразитизм в растительном мире — фитопатологией,
паразиты-животные, или зоопаразиты, — паразитологией.
От общего числа известных видов животных зоопаразиты составляют 6—7%.
Однако среди первично примитивных форм паразитизм встречается более часто, нежели среди высокоорганизованных организмов. Действительно, большая часть паразитов относится к типам Простейшие, Плоские черви. Круглые черви и Членистоногие. В целом в 17 классах вторичноротых животных паразитизм встречается только у отдельных видов, в то время как из 37 классов первичноротых в 20 классах паразитизм — обычное явление, а 6 классов представлены исключительно паразитами.
В подтипе Позвоночные паразитизм встречается в классе Круглоротые, к которому относятся миноги и миксины — водные организмы, присасывающиеся к покровам рыб и питающиеся кровью, и в классе Млекопитающие в отряде рукокрылых. Это группа южноамериканских летучих мышей-вампиров. В обоих случаях паразитизм позвоночных представлен в виде временного и больше напоминает хищничество, с которым связан по происхождению.
Таким образом, одной из предпосылок к паразитическому образу жизни является исходно низкий уровень организации. Кроме того, большинство паразитов имеют малые размеры, по крайней мере по сравнению с хозяевами. Поэтому наибольшее число специализированных паразитов встречается в систематических группах, представленных мелкими животными.
Явление паразитизма, как и любой другой экологический феномен, возникло разными путями. С одной стороны, по-разному развиваются взаимные адаптации паразитов и хозяев в разных систематических группах организмов — классах и типах, с другой — различны направления эволюции, ведущие к возникновению разнообразных форм паразитизма.
Первый подход к исследованию происхождения паразитизма конкретен. Он рассматривается при изложении материала по частной паразитологии в разделах, посвященных описанию характеристик типов и классов паразитических организмов и их экологических групп. Второй подход вскрывает общие закономерности перехода к паразитическому существованию вне зависимости от систематического положения организмов, занимающих новые экологические ниши.
Наиболее просто объясняется происхождение эктопаразитизма.
Один из путей к этому — через увеличение количества источников питания
с последующей их сменой. Так, многие насекомые имеют колюще-сосущий ротовой аппарат, питаясь соками растений. Но питание за счет прокалывания ткани и всасывания жидкости и есть способ поглощения пищи всеми кровососущими членистоногими, ряд которых, потребляя кровь человека и теплокровных животных, продолжает пользоваться также и соками растений, s
Другой путь, ведущий к эктопаразитизму, —
хищничество.
Активные хищники, осваивающие для питания все более крупные жертвы, становятся вначале временными, а затем и постоянными эктопаразитами за счет удлинения контактов с организмом хозяина. Так, многие пиявки, ведущие себя как хищники по отношению к мелким организмам, становятся паразитами более крупных животных, питаясь их кровью. Увеличение продолжительности питания — основное направление перехода от временного к постоянному эктопаразитизму. Действительно, из большого количества кровососущих форм членистоногих наиболее длительное питание на хозяине характерно именно для постоянных паразитов, степень контакта которых с хозяевами наиболее высока.
Иной путь возникновения эктопаразитизма — через усиление контакта так называемых гнездовых паразитов
с поверхностью тела хозяина. Животные, обитающие в убежище другого вида, могут питаться его перьями, волосами и отпадающими чешуйками кожного эпидермиса. Переход к постоянному обитанию на поверхности тела хозяина дает паразиту большие преимущества. Возможно, так возник паразитизм пухоедов, власоедов птиц и млекопитающих и группы клещей — обитателей эпидермиса животных и человека.
Основная масса случаев эндопаразитизма в полостных органах, имеющих связь с внешней средой, представляет собой явление, развившееся в результате случайного заноса
в организм цист, яиц или личинок свободноживущих видов, предварительно имеющих адаптации к обитанию в почве или в воде, содержащей избыток органического вещества. Примером является угрица кишечная, которая в своем развитии сохранила возможность обитать и размножаться как в почве, так и в организме человека (см. разд. 20.2.1.1).
Возможен переход к паразитированию в одном хозяине после предварительной адаптации к обитанию в другом,
служащем источником питания первого. Так, известен целый ряд гельминтов, которые, обитая в кишечнике рыбы, не перевариваются в пищеварительной системе хищников, съевших паразитов вместе с хозяином и продолжающих паразитировать в кишечнике или тканях уже нового вида (см. разд. 20.1.2.1).
Не исключается и вариант перехода к полостному паразитизму видов, предварительно адаптированных к эктопаразитизму.
Этим путем,вероятно, эволюционируют некоторые насекомые, большую часть цикла развития проводящие в ротовой полости птиц, но выходящие для размножения на перьевой покров их головы.
Наиболее сложно и многообразно происхождение паразитов тканей внутренней среды.
Один из путей — через изменение инстинкта откладки яиц и предварительных адаптации к эктопаразитизму. Таким путем, вероятно, произошел тканевой паразитизм личинок мух и оводов (см. разд. 21.2.4), откладывающих яйца на поверхности кожи и слизистых оболочек животных и человека. Личинки при этом вскоре погружаются под покровы и ведут типичный эндопаразитический образ жизни.
Многие паразиты приспособились к обитанию в тканях после освоения полостных органов, связанных с внешней средой. Так, по-видимому, шла эволюция паразитизма у ленточных червей, у трихинеллы спиральной (см. разд. 20.2.1.2). В цикле развития этих паразитов имеются формы, обитающие как в кишечнике, так и в тканях.
Некоторые паразиты внутренней среды возникли, вероятно, предварительно адаптировавшись к обитанию в пищеварительной системе членистоногих, а с переходом последних к гематофагии заселили новую и труднодоступную экологическую нишу — кровь и другие ткани мезодермального происхождения.
Таким образом, путей перехода к паразитизму у разных видов животных много, но несомненным остается одно: паразитизм — явление вторичное.
Об этом свидетельствует наличие в жизненных циклах многих, даже наиболее специализированных паразитов, свободноживущих стадий, рекапитулирующих свободный образ жизни предков (см., например, разд. 20.1.1).
Переход к паразитическому образу жизни сопровождается появлением у паразитов ряда адаптации,
облегчающих их существование, развитие и размножение в специфических условиях организма хозяина Разнообразие форм паразитизма, различное систематическое положение паразитов (их принадлежность к разным отрядам, классам и типам), а также обитание их в разных органах и системах хозяина обусловливают многообразие этих адаптации.
Однако некоторые приспособления являются абсолютно универсальными. К ним в первую очередь относятся высокая плодовитость
и особенности половой системы.
Действительно, возможность оставления потомства и попадания его в благоприятную среду — организм хозяина — у паразитов часто ничтожна. В связи с этим интенсивность размножения паразитов по сравнению со свободноживущими формами гораздо более велика. Достигается это разными способами. У многоклеточных это сильная степень развития половой системы и образование огромного количества половых продуктов. Этому способствуют первичный гермафродитизм плоских червей, изначально высокая плодовитость круглых червей и основной массы членистоногих. Нередко высокая интенсивность полового размножения дополняется размножением личиночных стадий жизненного цикла. Особенно это характерно для сосальщиков, личинки которых размножаются партеногенетически, а у некоторых ленточных червей — внутренним или наружным почкованием.

Рис. 18.4. Органы прикрепления паразитов
Присасывательные диски у инфузории (Л) и лямблии (Б
) (указаны стрелками), сложная присоска (В
)и кутикулярные шипики на коже (Г) у сосальщиков, крючья и сложные присоски на головках ленточных червей (Д, Е, Ж
),
конечности, служащие для прикрепления, у ракообразного (3
) и насекомого (И
)
Некоторые паразиты из типа простейших приобретают способность к множественному делению — шизогонии,
когда из одного паразита может образоваться более 1000 дочерних особей, или к спорогонии,
в результате которой из одной особи могут образоваться десятки тысяч организмов следующего поколения. Практически у всех эктопаразитов и паразитов, обитающих в полостных органах, имеются адаптации для прикрепления
к телу хозяина. Они встречаются у простейших (присасывательные диски лямблии), у гельминтов (присоски, шипики, крючья плоских червей, хитинизированный ротовой аппарат ряда круглых червей) и паразитических членистоногих (своеобразные конечности) (рис. 18.4).
Эндопаразиты, обитающие в полостных органах, имеют покровы,
обладающие антиферментными свойствами, быстро регенерирующие либо вообще непроницаемые для ферментов хозяина. Паразиты, живущие в тканях, часто там инкапсулируются (рис. 18.5).
Рис. 18.5. Личинка трихинеллы, инкапсулированная в мышце
Рис. 18.6. Пищеварительная система пиявки:
1 – слепые выросты кишечника
Паразиты, питающиеся кровью (представлены в основном членистоногими), имеют колюще-сосущий ротовой аппарат,
а также сильно растяжимый хитиновый покров, часто разветвленную пищеварительную трубку (рис. 18.6), антикоагулянтные свойства слюны и консервантные свойства ферментов пищеварительной системы.
Эндопаразиты, активно отыскивающие хозяина, обладают органами ориентации
в среде, используемыми для поисков хозяина (светочувствительные глазки, термо- и хеморецепторы), и органами передвижения
(рис. 18.7).
Передний конец тела паразитов, внедряющихся в организм хозяина, снабжен органами проникновения —
специализированными железами, колющими стилетами и т.д. Это касается даже некоторых простейших, способных проникать в ткани хозяина через неповрежденные покровы.
Все паразиты, развивающиеся со сменой хозяев, используют в качестве таковых виды, связанные между собой непосредственными пищевыми взаимоотношениями или обитающие с ними в одной среде. Большинство промежуточных хозяев являются источником питания для основных. Другой распространенный путь попадания паразита в организм хозяина — это использование многочисленных переносчиков,
которые обеспечивают не только постоянную циркуляцию паразитов в экологических системах, но и их широкое расселение.
Высшей степенью адаптации паразитов к хозяевам является наблюдаемая часто полная зависимость
паразита от жизнедеятельности хозяев. При этом паразит нередко вызывает такие реакции хозяина, которые обеспечивают максимальную вероятность заражения последнего. Так, самки остриц, откладывая яйца в области анального отверстия, вызывают зуд. Расчесывание зудящих мест способствует распространению яиц этого паразита руками по окружающим предметам. Таким же образом обеспечивается расселение чесоточного клеща. Зуд в пораженной конечности, прекращающийся от соприкосновения с водой, способствует циркуляции в природе такого паразита, как ришта. Высокая температура больных паразитарными заболеваниями, распространяющимися с помощью кровососущих членистоногих, привлекает переносчиков нередко с больших расстояний и также оказывается полезной для паразитов.
Нередко особенности жизнедеятельности паразитов оказываются синхронизированными
с образом жизни хозяев. Так, откладка яиц шистосомами происходит обычно в самое жаркое время суток, когда наиболее вероятным оказывается контакт хозяев с водой, куда для развития должны попасть яйца этих паразитов. В это же время в поверхностных слоях воды скапливаются в поисках хозяев церкарии этих шистосом. Таким образом облегчается циркуляция паразита сразу на двух стадиях его жизненного цикла (см. разд. 20.1.1.2). Если в циркуляцию паразита включены несколько хозяев, то наблюдаются их взаимные адаптации, оказывающиеся выгодными паразитам и обеспечивающие его эффективное развитие. Так, выход микрофилярий в кровеносные сосуды человека происходит в часы суток, соответствующие периоду максимальной активности кровососущих насекомых, являющихся их переносчиками (см. разд. 20.2.1.2).
Рис. 18.7. Приспособления, служащие для проникновения в организм хозяина. А—
простейшее токсоплазма; Б —
церкарии — личинка кровяного сосальщика:
1
— коноид, 2 — колющие стилеты
Нередко паразиты даже модифицируют поведение
одних хозяев таким образом, что в результате облегчается их попадание к другим. Так, рыбы, пораженные личинками ленточных червей, плавают в основном у поверхности воды и чаще вылавливаются рыбаками и хищными животными. Ленточные черви, использующие в качестве промежуточных хозяев копытных животных, снижают их жизнеспособность, и, таким образом, хищники поедают их в первую очередь. Сосальщики, заражающие травоядных животных и человека через случайное проглатывание насекомых, вызывают обездвиживание последних и плотное прикрепление к растениям, облегчающее им попадание в пищеварительный тракт хозяина (см. раздел 20.1.1.3. стр. 245).
Одновременно с перечисленными признаками свойства паразитов переживать неблагоприятные условия внешней среды
являются также несомненными адаптациями к паразитизму. Большинство простейших, заражение которыми происходит без участия переносчиков, во внешней среде способны инцистироваться. Яйца большинства гельминтов обладают феноменальной устойчивостью к неблагоприятным воздействиям. Капсулы с личинками трихинелл переносят не только промораживание, кипячение, но и многократное прохождение через пищеварительную систему рыб, земноводных, птиц, насекомых и ракообразных, не теряя жизнеспособности.
Для большинства паразитов, переживших нахождение во внешней среде или в промежуточном хозяине в покоящихся стадиях и попавших в организм окончательного хозяина, существует комплекс условий, являющийся сигналом к началу активной жизнедеятельности. У млекопитающих он часто неспецифичен: это температура тела около 37°С, водная среда и высокая ее кислотность, а также наличие ферментов желудочного сока. Такие условия характерны для желудка любого млекопитающего, поэтому цисты, яйца и другие инвазионные стадии паразитов, обитающие у разных млекопитающих могут, попадая в желудок человека, начинать развитие. Некоторые из них проходят лишь часть цикла и, не находя специфических условий, гибнут (см. разд. 20.2.1.3), успевая, однако, привести к тяжелым последствиям.
Указанные особенности паразитов, общие для многих из них, не состоящих в родстве, возникли в разных их группах независимо друг от друга, иллюстрируя конвергентный характер эволюции организмов разных видов, классов и типов, адаптирующихся к сходным условиям.
Онтогенез паразитов обычно бывает сложнее, чем развитие свобод-ноживущих видов. Действительно, свободноживущие организмы довольно легко преодолевают проблемы размножения и расселения, что значительно усложнено у паразитов. Поэтому большинство паразитов нередко развиваются со сложным метаморфозом,
включающим много личиночных стадий, обитающих в разных средах и выполняющих разные функции: расселения, активного роста, пассивного ожидания попадания в другую среду обитания и иногда даже размножения.
Совокупность всех стадий онтогенеза паразита и путей передачи его от одного хозяина к другому называют его жизненным циклом.
Личинки могут вести как свободный, так и паразитический образ жизни. Хозяин, в котором обитают личинки паразита, носит название промежуточного.
Значение промежуточных хозяев в циклах развития паразитов очень велико: они являются источниками заражения окончательных хозяев, часто выполняют расселительные функции, а иногда обеспечивают выживание популяций паразита в случае временного исчезновения окончательных хозяев.
Иногда в цикле развития паразита последовательно сменяются два-три промежуточных хозяина и даже больше. Хозяина, в котором развивается и размножается половым путем половозрелая стадия паразита, называют окончательным
или дефинитивным.
Заражение его осуществляется либо при поедании промежуточного хозяина, либо при контакте с последним в одной среде обитания.
Выделяют также понятие «резервуар паразита»,
или «резервуарный хозяин».
Это такой хозяин, в организме которого возбудитель заболевания может жить долго, накапливаясь, размножаясь и расселяясь по окружающей территории.
Наиболее часто резервуарами паразитов служат их дефинитивные хозяева. В том случае, когда продолжительность жизни промежуточного хозяина велика, а личинка в нем долго сохраняет жизнеспособность и иногда даже размножается, он также может выполнять роль резервуара. Продолжительность жизненного цикла разных паразитов очень сильно колеблется в зависимости от их систематического положения, видовой принадлежности и условий. Так, жизнь аргасовых клещей может продолжаться до 20 лет, кровяных сосальщиков — до 40, а детская острица и карликовый цепень живут не более 2 мес. Знание длительности онтогенеза паразитов необходимо для разработки мер профилактики паразитарных заболеваний.
Расселение паразитов
может происходить на разных стадиях их жизненного цикла. Расселение во времени обычно осуществляется покоящимися стадиями: развитие на этих стадиях приостанавливается до тех пор, пока не возникают новые условия, благоприятные для дальнейшего развития. Такими стадиями у простейших являются цисты,
а у гельминтов — обычно яйца
и иногда инкапсулированные личинки.
Обычно покоящиеся стадии очень устойчивы к изменениям внешней среды. Так, яйца аскариды могут сохраняться жизнеспособными до 7 лет, а цисты дизентерийной амебы — до 7 мес. При попадании покоящейся стадии в благоприятного хозяина перемещение последнего способствует расселению паразита (часто далеко за пределы ареала его первоначального существования). Цисты, яйца и инкапсулированные личинки могут также разноситься ветром, водными потоками и животными — механическими переносчиками.
Таким образом объясняется расширение ареалов распространения паразитов, не имеющих активных расселительных стадий в цикле развития. Многие паразиты, однако, имеют также свободноживущие подвижные стадии, служащие специально для расселения. Помимо расселения подвижные стадии часто выполняют функции поиска новых хозяев. Подвижный образ жизни промежуточных хозяев повышает вероятность контактов с окончательным хозяином. Перемещение окончательных хозяев, в которых обитают половозрелые паразиты, обеспечивает эффективное рассеивание цист, яиц и личинок паразитов по территории ареала.
Паразиты попадают к хозяевам разными путями. Нередко хозяев заражают переносчики — обычно кровососущие членистоногие. Такой способ передачи возбудителя называют трансмиссивным.
Существует два его варианта: инокулятивный
и контаминативный.
При первом возбудитель проникает в кровь хозяина через ротовой аппарат переносчика, при втором — выделяется переносчиком с фекалиями либо иным способом на кожу или слизистые оболочки и оттуда попадает в организм хозяина через рану от укуса, царапины, расчесы и т.п. (см. разд. 21.2.2; 21.2.3).
Другой способ заражения — через промежуточных хозяев.
В этом случае сам паразит не участвует в поисках хозяина, а промежуточный хозяин поедается окончательным. Столь же пассивно ведет себя паразит в случае заражения окончательного хозяина покоящимися стадиями — цистами, яйцами и инкапсулированными дичинками.
Ряд паразитов внедряются в организм хозяина на стадии свободноживущих личинок
через неповрежденную кожу и слизистые оболочки.
При любом способе заражения не исключена возможность попадания паразита к неподходящему хозяину. При этом развитие паразита либо вообще невозможно, либо прерывается на начальных стадиях.
Знание путей и способов проникновения паразитов в организм хозяина необходимо для разработки мер общественной и личной профилактики соответствующих заболеваний.
Существует много путей выведения паразитов
из организма хозяина. Так, паразиты, обитающие в пищеварительной системе, выделяют яйца, цисты или личинки с фекалиями. Живущие в мочеполовой системе — с мочой или содержимым влагалища, в легких — с мокротой. Паразиты внутренней среды обычно не покидают организм хозяина сами, а используют для расселения либо переносчиков, либо пассивно ожидают поедания хозяина другим хозяином.
Знание путей выведения паразитов или их покоящихся стадий из организма хозяина необходимо для правильной диагностики заболеваний. Действительно, если при постановке диагноза в одних случаях достаточно исследовать фекалии, мочу или мокроту больного с помощью микроскопа, то в других — приходится применять сложные иммунологические реакции или даже биопсию тканей больного1
.
Различные факторы генетической и негенетической природы обусловливают различную чувствительность организма хозяина к паразиту.
Среди негенетических факторов
выделяют возраст, питание, гормональный статус, сопутствующие заболевания и особенности их лечения. У детей с нарушенным белковым питанием более тяжело протекают амебиаз, стронгилоидоз и пневмоцистоз, в то время как тропическая малярия — легче, причем такие дети почти никогда не умирают от церебральных ее форм.
Люди, страдающие тяжелыми формами злокачественных опухолей, как правило, не заражаются висцеральным лейшманиозом. Железодефицитные анемии практически обеспечивают защиту человека от малярии, в то время как лечение препаратами железа сопровождается утяжелением течения этого заболевания.
Злокачественные опухоли толстой кишки и женской половой системы утяжеляют течение амебиаза и трихомоноза.
Запоры и все формы нарушения перистальтики кишечника способствуют утяжелению стронгилоидоза и аутоинвазии паразитом, его вызывающим. Поражение периферической нервной системы способствует протеканию чесотки в наиболее тяжелой форме.
Все формы иммунодефицитных состояний (СПИД, лечение кортикостероидными гормонами и иммунодепрессантами) приводят к утяжелению течения большинства инвазионных заболеваний.
Огромное значение в восприимчивости человека к паразитарным заболеваниям имеет его генетическая конституция.
Люди с группой крови II (А) оказываются наиболее восприимчивыми к лямблиозу. В Западной Африке в человеческих популяциях отсутствуют люди с группой крови Duffy, причем в этом регионе крайне редко встречается возбудитель трехдневной малярииPlasmodium vivax.
Показано, что восприимчивость человека к этому возбудителю зависит от наличия в крови антигена Duffy.
Разные формы гемоглобинопатий (талассемии, серповидно-клеточная анемия и др.), а также дефицит эритроцитарного фермента глюкозо-6-фосфатдегидрогеназы обеспечивают устойчивость к малярии. Так, известно, что в эритроцитах людей, гетерозиготных по гену серповидно-клеточной анемии, развитие малярийных плазмодиев идет быстрее и это приводит к преждевременному образованию гаметоцитов, что способствует ускоренному выведению паразитов из крови (см. разд. 19.3.2).
Не все рассмотренные примеры в настоящее время объяснены. Некоторые известны как эмпирические факты. Однако они убеждают в необходимости индивидуальной работы врача с больными паразитарными заболеваниями и делают необходимым дальнейшее изучение факторов восприимчивости человека к паразитам.
Защитные действия хозяина против паразитарной инвазии обеспечиваются главным образом иммунными механизмами.
Иммунные реакции хозяина возникают в ответ на действие антигенов двух разных типов: входящих в состав организма паразита и выделяющихся паразитами в окружающую среду.
Антигены первого типа,
кроме входящих в состав покровов, высвобождаются только после гибели паразитов. Они очень многообразны, но у многих, особенно родственных форм, часто бывают сходными. Поэтому антитела на эти антигены обладают слабой специфичностью. Антигены покровов разнообразны и специфичны. Часто они имеют гликопротеиновую природу и на разных этапах жизненного цикла паразитов могут меняться, поэтому выработка иммунитета к ним затруднена. Антигены второго типа
специфичны. Это компоненты слюны кровососущих паразитов, ферменты, выделяющиеся различными железами гельминтов.
Простейшие, обитающие вне клеток, покрываются антителами и в таком виде теряют свою подвижность. При этом облегчается их захват макрофагами. В некоторых случаях антитела обеспечивают агглютинацию (склеивание) паразитов, которые после этого гибнут. Внутриклеточные паразиты, обитающие в макрофагах, — лейшмании, токсоплазма — в случае активации макрофагов антителами могут перевариваться на месте пребывания. Против многоклеточных паразитов эти механизмы иммунной защиты не действенны. К неповрежденным покровам гельминтов антитела не прикрепляются. Иммунитет при гельминтозных заболеваниях поэтому частичный и действен в основном против личинок: мигрирующие личинки червей в присутствии антител замедляют или прекращают свое развитие. Некоторые типы лейкоцитов, в частности эозинофилы, способны прикрепляться к мигрирующим личинкам. Поверхность тела личинок при этом повреждается лизосомальными ферментами, что облегчает контакт тканей с антителами и часто приводит к гибели. Гельминты, прикрепляющиеся к стенке кишки, могут подвергаться воздействию клеточного иммунитета в слизистой оболочке. При этом перистальтика кишечника выбрасывает гельминтов во внешнюю среду.
При многих паразитарных заболеваниях между хозяином и паразитом устанавливаются компромиссные взаимоотношения: хозяин адаптируется к обитанию в его организме небольшого количества паразитов, а их существование в организме хозяина создает состояние иммунитета, препятствующего выживанию личинок, вновь попадающих в организм больного. Такое состояние называют нестерильным иммунитетом.
В сохранении нестерильного иммунитета хозяин заинтересован не только потому, что он предотвращает усиление степени инвазии: нередко в случае гибели паразита возникают серьезные тканевые реакции, способные привести хозяина к гибели. Примером таких реакций являются местные и общие осложнения после гибели личинок филярий в лимфатических узлах и в глазах, а также цистицерков свиного цепня в головном мозге. Пока паразиты живы, такие реакции вообще не проявляются. Поэтому во многих случаях система паразит — хозяин долгое время остается равновесной.
Многие паразиты на протяжении эволюции выработали механизмы, позволяющие им ослаблять неблагоприятное влияние иммунитета хозяина. Поэтому длительные и повторные инвазии возможны только в случае снижения иммунитета хозяина.
Многие простейшие обитают внутри клеток,
что делает их малодоступными для антител хозяина. Кроме того, такие паразиты способны размножаться и расселяться по организму хозяина, не выходя даже за пределы клеток, потому что клетки хозяина, в которых они находятся, нередко сохраняют способность к делению и перемещению.
Преимущества некоторым паразитам дает и локализация в тканевой жидкости, где концентрация антител обычно в 5 раз ниже, чем в плазме крови.
На паразитов, обитающих в просвете кишечника,
не действуют ни антитела плазмы крови, ни механизмы клеточного иммунитета. Некоторые взрослые гельминты покрыты толстой кутикулой, которая в неповрежденном виде почти не вызывает защитных реакций со стороны хозяина.
Ряд паразитов находятся в тканях внутренней среды в инкапсулированном состоянии.
В состав стенки капсулы входят как компоненты хозяина, так и паразита. В неповрежденном виде стенка такой капсулы почти непроницаема. Поэтому через нее не выходят антигены паразита и не проникают антитела хозяина.
Некоторые паразиты прибегают к очень своеобразному средству защиты — антигенной маскировке.
Они синтезируют поверхностные антигены, настолько сходные с белками хозяина, что организм не распознает их как чужеродные. Другие паразиты заимствуют антигены хозяина, включая и антигены групп крови, и, таким образом, также не воспринимаются хозяином как чужеродный материал. Иммунная защита против многоклеточных паразитов многократно усложняется многоступенчатостью их циклов развития, в которых каждая стадия может вырабатывать свой антигенный комплекс. К тому моменту, когда хозяин приобретает к нему иммунитет, паразит вступает в следующую стадию развития и меняет свой антигенный состав.
Паразитические простейшие способны менять антигенную структуру
своей оболочки, причем разными путями. Это может быть либо естественный отбор, формирующий возникновение популяций паразитов с новыми антигенными свойствами, как в случаях с малярийным плазмодием, либо, как у трипаносом, поочередная активация разных, но функционально сходных и родственных по происхождению генов, постоянно меняющих антигенную конституцию организма (см. разд. 19.3.2).
Популяции как хозяев, так и паразитов являются обязательными членами биогеоценозов, устойчивость которых зависит, в частности, от видового разнообразия живых организмов, входящих в их состав (см. § 16.1). Паразиты в экосистемах являются консументами второго и третьего порядков и играют существенную роль в биотическом круговороте веществ. Даже самые патогенные из них, вызывающие гибель большого числа особей хозяев, выступают, с одной стороны, как стабилизаторы численности хозяев, периодически изымая из популяций избыток организмов, который мог бы привести к нарушению экологического баланса. С другой стороны, наиболее тяжелое течение паразитарных заболеваний обычно наблюдается у особей с ослабленным иммунитетом, страдающих наследственными дефектами или с врожденной предрасположенностью к аллергическим реакциям. Гибель именно этих организмов оказывает на генетическую структуру популяций хозяина благотворную роль, элиминируя из его аллелофон-да аллели, снижающие жизнеспособность. Таким образом, взаимоотношения между популяциями хозяев и паразитов в условиях конкретных биогеоценозов способствуют их устойчивости и одновременно выступают как фактор естественного отбора,
снижая неспецифический генетический груз популяции хозяина (см. § 11.7 и 12.4).
Кроме того, утрата биогеоценозами наиболее восприимчивых к заражению паразитами особей хозяина сопровождается и уничтожением части паразитарной популяции, гибнущей вместе с ним. Это обеспечивает активизацию микроэволюционных процессов в оставшейся части популяции паразитов, способствуя в конечном счете появлению у них новых адаптации.
В связи с социальностью человека в настоящее время паразитизм как фактор естественного отбора в человеческих популяциях значения практически не имеет. Однако целенаправленная борьба человека с паразитами, осуществляющаяся разными способами, несомненно является важным фактором эволюции самих паразитов (см. гл. 22).
Изучение распределения паразитов в популяциях хозяина показало, что оно зависит от многих факторов. Одним из них является возраст хозяина.
Ряд паразитов чаще встречаются у взрослых хозяев. Действительно, у взрослых организмов больше шансов прийти в соприкосновение со многими паразитами и быть инвазированными. Например, влагалищная трихомонада поражает только половозрелых людей потому, что передается только половым путем.
Дифиллоботриозом
и описторхозом
человек заражается, поедая недостаточно термически обработанную рыбу. Такой путь заражения маловероятен для ребенка. Восточно-африканский трипаносомоз
встречается чаще у людей среднего возраста — охотников, путешественников, участников геологоразведочных партий в необжитых саваннах Африки. Эта закономерность часто проявляется и у промежуточных хозяев: взрослые крупные рыбы имеют больше возможностей стать носителями метацеркарий сосальщиков или плероцеркоидов ленточных червей, чем мелкие молодые.
Другие паразиты чаще встречаются у детей. Причины этого разнообразны. Во-первых, это связано с незрелостью иммунной системы детей, а во-вторых, с не отработанными еще навыками личной гигиены. Поэтому для детей характерен особый спектр паразитарных заболеваний, в заражении которых большое значение имеет состояние иммунитета, а инвазия осуществляется просто при проглатывании цист, яиц или личинок, без участия промежуточных хозяев. Это лямблиоз, энтеробиоз, гименолепидоз, аскаридоз.
Более частому заражению детей способствует их тесный и продолжительный контакт друг с другом в детских учреждениях.
На вероятность заражения также часто накладывает отпечаток профессия. Так, тениозом
и тениаринхозом
обычно заражаются работники мясокомбинатов, анкилостомидозами
в умеренных широтах — шахтеры, а в тропиках — работники сельского хозяйства. Дифиллоботриозом
чаще заражаются рыбаки, а альвеококкозом —
охотники и лица, обрабатывающие меховое сырье.
Некоторые паразиты, в первую очередь простейшие, а также острица, угрица кишечная, карликовый цепень и трихинелла, попав в организм человека, способны размножаться в нем. То же касается личинок сосальщиков и некоторых ленточных червей в промежуточных хозяевах. Все это способствует тому, что в популяциях хозяина паразиты распределяются не равномерно, а перерассеянно.
Это означает, что в меньшей части популяции хозяина сосредоточена большая часть популяции паразита.
Данная особенность имеет принципиальное значение для выживания хозяев, так как сводит к минимуму результаты массированных инвазий их паразитами, сохраняя основное ядро популяции интакт-ным. Перерассеянное распределение паразита в популяции хозяина необходимо учитывать при проведении профилактических мероприятий. Действительно, для выживания популяции паразита и передачи ее следующему поколению хозяев нередко важными оказываются лишь несколько особей или даже один организм, несущий в себе очень большое число паразитов.
Характерной особенностью паразитизма является соответствие определенного вида паразита определенному хозяину. Это соответствие называют специфичностью паразита.
Степень специфичности паразитов может быть различна: от строгой к определенному подвиду до форм, встречающихся в десятках различных видов хозяев. Примерами специфических паразитов человека являются малярийные плазмодии, острица детская и некоторые другие. Источником инвазии этими паразитами всегда является человек. Такие специфические паразиты человека вызывают заболевания, называемые антропонозными.
Ряд других паразитов, встречающихся у человека, может поражать также и человекообразных обезьян. Таковы, например, вши, вухерерия Бликрофта и некоторые другие. Источником инвазии ими в абсолютном большинстве случаев также является человек.
Многие паразиты обладают меньшей специфичностью, чаще встречаясь у домашних и диких животных, но могут поражать также и человека. К таким паразитам относятся печеночный сосальщик, широкий лентец, вольфартова муха и многие другие. Источником заражения человека в этом случае являются обычно животные. Заболевания, вызываемые этими паразитами, называют зоонозньши.
Деление инфекционных и паразитарных заболеваний на антропо-и зоонозные имеет огромное эпидемиологическое значение в связи с разными подходами к их профилактике. Для профилактики антропо-нозных болезней в первую очередь необходимо выявление и лечение больных. Полное излечение всех больных людей этими заболеваниями должно привести к полному исчезновению этих паразитов как биологических видов. В то же время профилактика зоонозных заболеваний значительно сложнее. Для ее успешного проведения необходимы мероприятия не только по оздоровлению человека» но и других хозяев, что значительно труднее, в особенности в отношении диких животных.
У одного и того же вида паразитов специфичность может быть выражена в разной степени на разных стадиях жизненного цикла. У некоторых видов специфичность более выражена на личиночных стадиях, а половозрелые организмы обитают в большом количестве окончательных хозяев. Таковы, например, сосальщики, личинки которых адаптированы к определенным видам моллюсков, а взрослые формы могут паразитировать у разных видов млекопитающих.
Многие ленточные черви, наоборот, строго специфичны по отношению к окончательным хозяевам и проявляют меньшую разборчивость к промежуточным хозяевам.
Изменение специфичности паразитов к хозяевам на протяжении жизненного цикла обеспечивает им широкую экологическую пластичность,
позволяющую выживать в меняющихся условиях, и открывает дальнейшие эволюционные перспективы.
Учет этой особенности паразитов необходим для проведения личной профилактики заражения соответствующими заболеваниями. Действительно, для предотвращения заражения лейшманиозами, трипано-сомозами или малярией (см. разд. 19.3.2) оказывается достаточным предохранение от укусов определенным видом кровососущих насекомых, в то время как для личной профилактики, например, токсоплаз-моза (см. разд. 19.3.1) необходим сложный комплекс мероприятий.
Большая группа паразитарных и инфекционных заболеваний характеризуется природной очаговостью.
Для них характерны следующие признаки: 1) возбудители циркулируют в природе от одного животного к другому независимо от человека; 2) резервуаром возбудителя служат дикие животные; 3) болезни распространены не повсеместно, а на ограниченной территории с определенным ландшафтом, климатическими факторами и биогеоценозами.
Компонентами природного очага
являются: 1) возбудитель; 2) восприимчивые к возбудителю животные — резервуары: 3) соответствующий комплекс природно-климатических условий, в котором существует данный биогеоценоз. Особую группу природно-очаговых заболеваний составляют трансмиссивные болезни,
такие, как лейшманиоз, трипаносомоз, клещевой энцефалит и т.д. Поэтому обязательным компонентом природного очага трансмиссивного заболевания является также наличие переносчика.
Структура такого очага приведена на рис. 18.8.


Рис. 18.8. Структура природного очага паразитарного заболевания. А —
лейшманиоза (трансмиссивного заболевания); Б —
дифиллоботриоза (нетрансмиссивного заболевания):
1
— возбудитель заболевания — лейшмания, 2 —
природный резервуар — монгольские песчанки, 3 —
переносчик возбудителя — москит, 4 —
норы грызунов в полупустынях Средней Азии, 5 —
возбудитель заболевания — широкий лентец, 6 —
природный резервуар — рыбоядные млекопитающие, 7 — промежуточные хозяева — циклопы и рыбы, 8
— крупные пресноводные водоемы Северной Евразии
Категорию заболеваний с природной очаговостью выделил акад. Е.Н. Павловский в 1939 г. на основании экспедиционных, лабораторных и экспериментальных работ. В настоящее время природно-очаговые заболевания активно изучают в большинстве стран мира. Освоение новых, незаселенных или малообжитых территорий приводит к открытию и новых, неизвестных ранее природно-очаговых заболеваний.

Рис.
18.9. Клещ Amblyomma sp.
Некоторые природно-очаговые заболевания характеризуются эндемизмом,
т.е. встречаемостью на строго ограниченных территориях. Это связано с тем, что возбудители соответствующих заболеваний, их промежуточные хозяева, животные-резервуары или переносчики встречаются только в определенных биогеоценозах. Так, только в отдельных районах Японии расселены четыре вида легочных сосальщиков из р.Paragonimus
(см. разд. 20.1.1.3). Расселению их препятствует узкая специфичность в отношении промежуточных хозяев, которые обитают только в некоторых водоемах Японии, а природным резервуаром являются такие эндемичные виды животных, как японская луговая мышь или японская куница.
Вирусы некоторых форм геморрагической лихорадки
встречаются только в определенных зонах Восточной Африки, потому что здесь расположен ареал их специфических переносчиков — клещей из р. АтЫуотта
(рис. 18.9).
Хорошо известными являются природно-очаговые заболевания, вызываемые лентецами р.Diphyllobothrium,
распространенными в некоторых районах Сибири. Так, на оз. Байкал известно несколько видов ленточных червей, которые обычно паразитируют у чаек, заражающихся при проглатывании инвазированной рыбы, например байкальского омуля. Рыбы заражаются при поедании рачков, а те в свою очередь становятся промежуточными хозяевами гельминтов от фекалий птиц, попадающих в воду. Съев инвазированную рыбу, заболеть дифиллоботриозом
может и человек (см. разд. 20.1.2.1).
Существуют природно-очаговые заболевания, имеющие более широкий ареал. Так, в бассейне р. Оби и Иртыша, а также в некоторых других зонах Сибири и Восточной Европы распространено заболевание описторхоз,
встречающееся у медведей, выдр, кошек, волков, лис, а также у человека. Возбудителем этого заболевания является кошачий сосальщик, расселение которого ограничивается, вероятно, комплексом природно-климатических факторов. Об этом свидетельствует то, что ближайший родственник этого паразита, сосальщик виверры,
встречается только в Юго-Восточной Азии и не выходит за пределы этого региона, являясь, таким образом, компонентом восточно-азиатских тропических биогеоценозов.
Небольшое количество природно-очаговых заболеваний встречается практически повсеместно. Это такие заболевания, возбудители которых, как правило, не связаны в цикле своего развития с внешней средой и поражают самых разнообразных хозяев. К заболеваниям такого рода относятся, например, токсоплазмоз
и трихинеллез.
Этими природно-очаговыми болезнями человек может заразиться в любой природно-климатической зоне и в любой экологической системе.
Абсолютное же большинство природно-очаговых болезней поражает человека только в случае попадания его в соответствующий очаг (на охоте, рыбной ловле, в туристических походах, в геологических партиях и т.д.) при условиях его восприимчивости к ним. Так, таежным энцефалитом
человек заражается при укусе инфицированным клещом, а описторхозом —
съев недостаточно термически обработанную рыбу с личинками кошачьего сосальщика.
Профилактика природно-очаговых заболеваний
представляет особые сложности. В связи с тем, что в циркуляцию возбудителя бывает включено большое количество хозяев, а часто и переносчиков, разрушение целых биогеоценотических комплексов, возникших в результате эволюционного процесса, экологически неразумно, вредно и даже технически невозможно. Лишь в тех случаях, если очаги являются небольшими и хорошо изученными, возможно комплексное преобразование таких биогеоценозов в направлении, исключающем циркуляцию возбудителя. Так, рекультивация опустыненных ландшафтов с созданием на их месте орошаемых садоводческих хозяйств, проводящаяся на фоне борьбы с пустынными грызунами и москитами, может резко снизить заболеваемость населения лейшманиозами. В большинстве же случаев природно-очаговых болезней профилактика их должна быть направлена в первую очередь на индивидуальную защиту (предотвращение от укусов кровососущими членистоногими, термическая обработка пищевых продуктов и т.д.) в соответствии с путями циркуляции в природе конкретных возбудителей.
К типу Простейшие
относят организмы, тело которых состоит из одной клетки, функционирующей, однако, как целый организм. Клетки простейших способны к самостоятельному питанию, передвижению, защите от врагов и к переживанию неблагоприятных условий. В строении простейших обнаруживаются как все особенности эукарио-тических клеток, так и специфические органеллы, обеспечивающие выполнение организменных функций.
Питание простейших происходит с помощью пищеварительных вакуолей,
содержащих пищеварительные ферменты и связанных по происхождению с лизосомами. Оно осуществляется за счет фаго-
или пиноцитоза.
Остатки непереваренной пищи выбрасываются наружу. Некоторые простейшие содержат хлоропласты и способны питаться за счет фотосинтеза.
Большинство простейших имеют органеллы передвижения:
жгутики, реснички и псевдоподии (временные подвижные выросты цитоплазмы). Формы органелл движения лежат в основе систематики простейших.
Пресноводные свободноживущие простейшие имеют органеллы, регулирующие водно-солевой баланс, — сократительные вакуоли.
Периодически они сокращаются и выделяют во внешнюю среду избытки воды и жидкие продукты диссимиляции. Морские и паразитические простейшие, живущие в среде с высокой концентрацией солей, могут не иметь сократительных вакуолей.
Размножение простейших осуществляется обычно разными формами деления —
разновидностями митоза. Характерен также половой процесс: в виде слияния клеток — копуляция —
ил» обмен наследственным материалом — конъюгация.
Большинство простейших имеют одно ядро,
но встречаются и многоядерные
формы. Ядра некоторых простейших характеризуются полиплоидностью.
В жизненном цикле большинства простейших выделяют стадию трофозоита —
активно питающуюся и перемещающуюся форму — и стадию цисты. Циста —
неподвижная форма жизненного цикла простейших, покрытая плотной оболочкой и характеризующаяся резко замедленным обменом веществ. Паразитические простейшие инцисти-руются, попадая во внешнюю среду. В таком состоянии они способны переноситься ветром, водой и животными на огромные расстояния и таким образом расселяться. При попадании цисты в благоприятные условия происходит эксцистирование и простейшее начинает активно функционировать в состоянии трофозоита.
В настоящее время известно около 10 000 видов простейших. Основными средами их обитания являются вода и почва. Многие простейшие перешли к паразитическому или к комменсальному образу жизни.
Болезни, вызываемые простейшими, называют протозойными.
Большинство простейших имеют время генерации от 6 до 24 ч. В связи с этим их размножение в организме хозяина обычно сопровождается экспоненциальным увеличением размеров их популяций до тех пор, пока этот процесс не замедлится или не остановится защитными механизмами хозяина или другими внешними факторами. Это означает, что один паразитический организм в принципе способен, размножившись, привести к гибели своего хозяина. В этом плане простейшие — возбудители заболеваний — сходны с возбудителями инфекционных болезней, например с патогенными бактериями и вирусами.
Медицинское значение имеют простейшие, относящиеся к классам Саркодовые, Жгутиковые, Инфузории и Споровики.
19.1.1. Класс Саркодовые
Sarcodina
Представители этого класса — самые примитивные простейшие. Форма их тела непостоянна. Передвигаются они с помощью ложноножек. Обитают в пресных водах, в почве, морях. В биогеоценозах выполняют функции консументов и редуцентов. Некоторые саркодовые адаптировались к комменсальному и паразитическому образу жизни. Медицинское значение имеют представители отряда амеб Amoebina. Паразитические амебы обитают у человека в основном в пищеварительной системе. Некоторые саркодовые, ведущие свободный образ жизни и обитающие в почве и загрязненной воде, при попадании в организм человека могут вызывать тяжелые заболевания, нередко заканчивающиеся смертью.
19.1.2. Класс Жгутиковые
Flagellata
Тело жгутиковых кроме цитоплазматической мембраны покрыто еще и пелликулой —
специальной оболочкой, обеспечивающей постоянство его формы. Имеется один или несколько жгутиков, органелл движения, представляющих собой нитевидные выросты эктоплазмы. Внутри жгутиков проходят фибриллы из сократительных белков. Некоторые жгутиковые имеют также ундулирующую мембрану —
своеобразную органеллу передвижения, в основе которой лежит тот же жгутик, не выступающий свободно за пределы клетки, а проходящий по наружному краю длинного уплощенного выроста цитоплазмы. Жгутик приводит ундулирующую мембрану в волнообразное движение. Основание жгутика всегда связано с кинетосомой,
органеллой, выполняющей энергетические функции. Ряд жгутиковых имеет также и опорную органеллу — аксостиль —
в виде плотного тяжа, проходящего внутри клетки.
Разные виды паразитических жгутиковых у человека обитают в различных органах. Циклы их развития очень разнообразны.
19.1.3. Класс Инфузории
Infusoria
Для инфузорий, как и для жгутиковых, характерно наличие пелликулы, им свойственна постоянная форма тела. Органеллы передвижения — многочисленные реснички, покрывающие все тело и представляющие собой полимеризованные жгутики. У инфузорий обычно два ядра: крупное — макронуклеус,
регулирующее обмен веществ, и малое — микронуклеус,
служащее для обмена наследственной информацией при конъюгации. Макронуклеусы инфузорий полиплоидны, микронуклеусы — гаплоидны или диплоидны. Сложно организован аппарат пищеварения. Имеется постоянное образование: клеточный рот — цито-стом,
клеточная глотка — цитофаринкс.
Пищеварительные вакуоли перемещаются по эндоплазме, при этом литиче''кие ферменты выделяются поэтапно. Это обеспечивает полноценное переваривание пищевых частиц. Непереваренные остатки пищи выбрасываются через порошицу —
специализированный участок клеточной поверхности.
Инфузории — наиболее высоко организованные простейшие. Паразитов среди них относительно немного. У человека паразитирует единственная инфузория — балантидий,
которая обитает в пищеварительной системе.
19.1.4. Класс Споровики
Sporozoa
Все споровики — паразиты и комменсалы животных и человека. Органеллы движения у них отсутствуют. Питание споровиков осуществляется за счет поглощения пищи всей поверхностью тела. Многие споровики — внутриклеточные паразиты. Они претерпели наиболее глубокую дегенерацию. Цикл развития включает стадии бесполого размножения, нолового процесса в виде копуляции
и спорогонии.
Бесполое размножение осуществляется путем простого или множественного деления — шизогонии.
Половому процессу предшествует образование половых клеток — мужских и женских гамет.
Гаметы сливаются, а образовавшаяся зигота
покрывается оболочкой, под которой происходит спорогония — множественное деление с образованием спорозоитов
(рис. 19.1).
Ниже описаны паразитические и комменсальные простейшие, обитающие в разных органах человека. От специфики органа, являющегося средой обитания паразита, зависят пути проникновения и патогенное действие паразита, методы диагностики соответствующих заболеваний и меры их профилактики. Поэтому простейшие, с медицинской точки зрения, могут быть разделены на виды, обитающие в полостных органах, которые имеют связь с внешней средой, и живущие в тканях внутренней среды человека. Кроме того, выделяют группу свободноживущих простейших, случайное попадание которых в организм человека может приводить к острейшим патологическим процессам и даже к смерти. Соответствующие три экологические группы простейших описаны отдельно.
Рис. 19.1. Цикл развития споровиков:
1
— спорозоиты под оболочкой цисты, 2 —
спорозоит, 3 —
паразит в клетке хозяина, 4 —
дочерние паразиты в клетке хозяина, 5
и 6 —
мужская и женская гаметы, 7 — зигота
Простейшие, относящиеся к этой экологической группе, став паразитами, не претерпели глубокой дегенерации в связи с тем, что условия их обитания относительно мало отличаются от внешней среды. Большинство этих паразитов анаэробны. Циклы их развития просты:
у многих имеются стадии трофозоита и цисты. Некоторые не образуют и цист. Заражение большинством этих паразитов осуществляется путем проглатывания цист или трофозоитов. В их распространении большое значение имеют насекомые — механические переносчики возбудителей. Диагностика соответствующих заболеваний основана на обнаружении цист или трофозоитов в выделениях из пораженных органов. В профилактике заражения основное значение имеет соблюдение правил личной гигиены.
Многие виды, живя в просвете пищеварительной трубки и других органов и находя там благоприятную среду, являются комменсалами. Однако в будущем возможно усиление степени их контакта с хозяином и переход к паразитированию.
19.2.1. Простейшие, обитающие в полости рта
Ротовая амеба
Entamoeba gingivalis
(кл. Саркодовые) — комменсал, обитающий на деснах, зубном налете и в криптах нёбных миндалин более чем у 25% здоровых людей (рис. 19.2, А
).
У лиц с заболеванием полости рта встречается чаще. Размеры клетки 6—30 мкм, псевдоподии широкие. Питается бактериями и лейкоцитами, при кровотечении из десен может захватывать и эритроциты. Цист не образует.
Рис. 19.2. Комменсальные простейшие, обитающие в полости рта. А — ротовая амеба; Б
— Trichomonas tenax:
1 —
ядро, 2 —
пищеварительная вакуоль
Trichomonas tenax
(кл.
Жгутиковые) — такой же комменсал, как и предыдущий вид. Форма тела грушевидная, длина 6—13 мкм (рис. 19.2, Б
).
На переднем конце расположены четыре жгутика, сбоку находится ундулирующая мембрана длиной около 3
/4
длины тела. Встречается у 30% здоровых людей, причем у взрослых чаще, чем у детей. Обитает в складках слизистой оболочки рта, кариозных полостях зубов, криптах миндалин при хроническом тонзиллите, а при низкой кислотности желудочного сока встречается и в желудке. Цист не образует, как и предыдущий вид. Передача от человека к человеку обоих видов осуществляется при поцелуях, пользовании общей посудой и зубными щетками, а также с капельками слюны и мокроты при чихании и кашле.
Оба вида самостоятельного медицинского значения не имеют, однако считается, что при патологических процессах в полости рта могут утяжелять их течение.
19.2.2. Простейшие, обитающие в тонкой кишке
В тонкой кишке человека паразитирует единственный вид простейших — лямблия
Lamblia intestinalis
(кл. Жгутиковые) — возбудитель лямблиоза,
которым чаще болеют дети. Форма паразита напоминает грушу, разрезанную вдоль. Длина тела 10—18 мкм. В расширенной части, на уплощенной стороне расположен присасывательный диск, с помощью которого лямблии присасываются к ворсинкам кишечника. Вдоль тела проходят две тонкие опорные органеллы — аксостили. Симметрично в клетке располагаются два ядра и четыре пары жгутиков (рис. 19.3, А).
Рис. 19.3. Простейшие, обитающие в тонкой и толстой кишках. А —
лямблия; Б —
дизентерийная амеба; В —
кишечная амеба; Г —
кишечная трихоманада; Д— балантидий кишечный: а —
трофозонты, б —
цисты
Трофозоиты используют питательные вещества с поверхности клеток кишечного эпителия. Захват пищи осуществляется пиноцитозом.
Большие количества лямблии, которые покрывают обширные поверхности кишечной стенки, нарушают процессы всасывания и присте-ночного пищеварения. Попадая в нижние отделы тонкой кишки, лямблии инцистируются. Зрелые цисты имеют овальную форму, четыре ядра и несколько аксостилей. Во внешней среде цисты сохраняют жизнеспособность в течение нескольких недель. Заражение человека происходит при проглатывании цист.
Лабораторная диагностика — обнаружение цист в фекалиях и тро-фозоитов в содержимом двенадцатиперстной кишки, полученном при дуоденальном зондировании.
Личная профилактика — соблюдение правил гигиены питания. Общественная профилактика —
санитарное благоустройство туалетов, предприятий общественного питания.
19.2.3. Простейшие, обитающие в толстой кишке
Толстая кишка очень благоприятна для обитания простейших. Паразиты располагаются вблизи слизистой оболочки, в слизи, покрывающей эпителий крипт. Большинство этих простейших являются комменсалами, питаясь бактериями и слущенными клетками кишечника. Два вида простейших — дизентерийная амеба и балантидий — патогенны, но в организме здорового человека они могут долгое время вести комменсальный образ жизни.
Дизентерийная амеба
Entamoeba histolylica
(кл. Саркодовые) — возбудитель амебиаза
(рис. 19.3, Б
).
Амебиаз встречается повсеместно, но чаще в зонах с влажным жарким климатом. В цикле развития амебы имеется несколько стадий, морфологически и физиологически отличающихся друг от друга. Мелкая вегетативная форма
обитает в просвете кишки. Размеры ее 8—20 мкм. В цитоплазме можно обнаружить бактерии и грибки — элементы микрофлоры кишечника.
Крупная вегетативная форма также обитает в просвете кишки в гнойном содержимом язв кишечной стенки. Ее размеры — до 45 мкм. Цитоплазма четко разделена на прозрачную, стекловидную эктоплазму и зернистую эндоплазму. В ней расположены ядро с характерной темно окрашенной кариосомой и эритроциты, которыми она питается. Крупная форма энергично передвигается с помощью широких псевдоподий. В глубине пораженных тканей располагается тканевая форма.
Она мельче крупной вегетативной формы и не имеет в цитоплазме эритроцитов. Цисты обнаруживаются в фекалиях хронически больных и паразитоносителей, у которых заболевание проходит бессимптомно. Цисты имеют округлую форму диаметром 8—15 мкм и от одного до четырех ядер в виде колечек.
Жизненный цикл паразита сложен (рис. 19.4). Человек заражается амебиазом, проглатывая цисты паразита с водой или пищевыми продуктами, загрязненными землей. В просвете толстой кишки из цисты образуется, за счет следующих друг за другом делений, восемь мелких клеток, превращающихся в мелкие вегетативные формы. Вреда человеку они не приносят. Они могут вновь инцистироваться и выходить наружу. При ухудшении условий существования хозяина мелкие вегетативные формы способны превращаться в крупные, которые вызывают образование язв. Погружаясь глубже, они превращаются в тканевые формы, которые в особо тяжелых случаях могут попадать в кровь и разноситься по всему организму. При этом возможно образование абсцессов в печени, легких и других органах. В остром периоде заболевания у больного в фекалиях обнаруживаются не только цисты, но и трофозоиты.
Диагноз ставится на основе обнаружения в фекалиях трофозоитов с заглоченньми эритроцитами. Четырехъядерные цисты могут свидетельствовать скорее о хроническом течении заболевания или о пара-зитоносительстве.
Профилактика — как при лямблиозе.
Балантидий
Balantidium coli
(кл. Инфузории) — возбудитель балантидиаза.
Это крупное простейшее, длиной до 200мкм.
Сохранены многие признаки свободноживущих инфузорий: все тело покрыто ресничками, имеются цитостом и цитофаринкс. Под пелликулой расположен слой прозрачной эктоплазмы, глубже находится эндоплазма с органеллами и двумя ядрами. Макронуклеус имеет гантелевидную или бобовидную форму, рядом с ним находится маленький микронуклеус. Циста балантидия овальна, до 50—60 мкм в диаметре, покрыта двуслойной оболочкой, ресничек не имеет. Микронуклеус обычно не виден, а в цитоплазме отчетливо выделяется сократительная вакуоль (см. рис. 19.3, Д
).
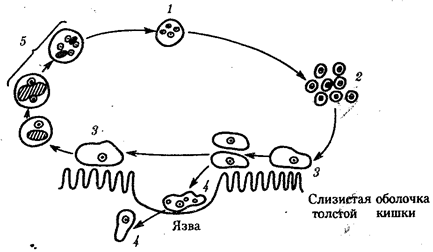
Рис. 19.4. Цикл развития дизентерийной амебы:
1
— циста, 2 —
мелкая вегетативная форма, 3 —
крупная вегетативная форма, 4 —
тканевая форма, 5
— инцистирование
Балантидий может жить в кишечнике человека, питаясь бактериями и не принося ему вреда, но иногда внедряется в стенку кишки, вызывая образование язв с гнойным и кровянистым отделением. В этом случае в его цитоплазме часто обнаруживаются форменные элементы крови хозяина. Для заболевания характерны длительные поносы с кровью и гноем, а иногда и перфорация кишечной стенки с перитонитом. Как и при амебной дизентерии, В. coli
может попадать в кровеносное русло и оседать в печени, легких и других органах, вызывая там образование абсцессов.
Особенностью этих инфузорий является их способность вырабатывать фермент гиалуронидазу, благодаря которой они внедряются и в неповрежденную стенку кишки, где на гистологических препаратах обнаруживаются целые скопления тканевых трофозоитов, морфологически не отличимых от живущих в просвете кишки, но не способных к образованию цист. Кроме человека, балантидий встречается также у крыс и свиней, которые и являются его основным резервуаром.
Лабораторная диагностика — обнаружение цист и трофозоитов в мазках фекалий больного.
Профилактика — как при лямблиозе, однако в связи с зоонозной природой балантидиаза следует также вести борьбу с грызунами и обеспечивать гигиеническое содержание свиней.
Кроме описанных паразитов в пищеварительной трубке человека, особенно в ее нижних отделах, обитает ряд условно патогенных видов простейших. Среди них особенно интереснаDientamoebafragilis
чертами адаптации к обитанию в кишечнике человека: она сосуществует в своеобразном симбиозе с острицами (см. разд. 20.2.1.1). Цист эта амеба не образует, а трофозоиты прикрепляются к яйцам остриц, через которые и происходит заражение новых хозяев. Вероятно, заражение этой амебой возможно и без яиц острицы, при прямом контакте. Размножаясь в большом количестве в кишке человека, эта амеба может вызвать недолгие поносы. Такое же медицинское значение имеет и известный жгутиконосец — кишечная трихомонадаTrichomonas hominis
(см. рис. 19.3, Г
).
В плане дифференциальной диагностики следует упомянуть также кишечную амебу
Entamoeba coli —
нормального симбионта толстой кишки человека. Она очень похожа на дизентерийную амебу, но является типичным комменсалом (см. рис. 19.3, В
).
Трофозоиты имеют размеры 20—40 мкм и передвигаются медленно. Питаются бактериями, грибами, а если у хозяина имеется кишечное кровотечение, то и форменными элементами крови.
В окружающую среду выделяется в виде цист, содержащих восемь ядер и имеющих более крупные размеры, чем у Е.
histolytica
(около 18 мкм).
19.2.4. Простейшие, обитающие в половых органах
В половых органах человека обитает влагалищная трихомонада
Trichomonas vaginalis
(кл. Жгутиковые) — возбудитель трихомоноза.
Длина этого паразита 14—30 мкм. Форма тела грушевидная. На переднем конце находятся четыре жгутика. До середины клетки доходит также небольшая ундулирующая мембрана. По середине тела тянется аксостиль, выступающий из клетки на ее заднем конце. Характерна форма ядра, овального, заостренного с двух концов, и напоминающего косточку сливы (рис. 19.5).
В пищеварительных вакуолях располагаются лейкоциты, эритроциты и бактерии, которыми этот паразит питается. Цист не образует. Эта трихомонада обитает у женщин во влагалище и в шейке матки, а у мужчин — в мочеиспускательном канале, мочевом пузыре и в предстательной железе. Зараженность женщин достигает 20—40%, мужчин — 15%. Серьезных повреждений хозяину эта трихомонада не наносит, но, тесно контактируя с эпителием мочеполовой системы, она вызывает врзникновение мелких воспалительных очагов под эпителиальным слоем и слущивание поверхностных клеток слизистой оболочки. Через нарушенную эпителиальную выстилку в просвет органа поступают лейкоциты. У мужчин заболевание обычно завершается спонтанным выздоровлением примерно через 1 мес. У женщин трихомоноз может протекать несколько лет.
Лабораторная диагностика — обнаружение живых подвижных трихомонад в мазке из выделений мочеполовых путей.
Профилактика — соблюдение правил личной гигиены при половых контактах.

Рис. 19.5 Трихомонада влагалищная
19.2.5. Одноклеточные паразиты, обитающие в легких
Pneumocystis carinii
относится к своеобразной группе паразитов не имеющих полового процесса. Является возбудителем пневмоцистоза.
Современные исследования особенностей биохимической и генетической организации позволили отнести этого паразита не к типу простейших, а к особой группе царства Грибов (Furgi). Однако в связи со сходством жизненного цикла пневмоцистиса со многими паразитическими простейшими и с его серьезным медицинским значением авторы сочли уместным поместить его описание в данном разделе книги.
Паразиты имеют неправильную форму, размеры их от 1 до 5 мкм. В их цитоплазме содержатся митохондрии, а диссимиляция происходит аэробно.
Обитает в альвеолах легких человека и многих млекопитающих.
Человек заражается воздушно-капельным путем, вдыхая клетки пневмоцистиса.
В легких паразиты располагаются на альвеолярном эпителии, прикрепляясь к нему псевдоподиями. Здесь они размножаются простым делением, они увеличиваются в размерах, образуют толстую оболочку и делятся обычно несколько раз образуя споры. При разрыве оболочки споры паразитов выходят из нее и начинается следующий цикл размножения в данном организме или в другом хозяине, в случае попадания к нему при вдыхании (рис. 19.6).
У 1—10% здоровых людей отмечены Р.
carinii.
Инвазия протекает обычно бессимптомно, однако, в случае ослабления иммунитета хозяина размножение паразитов усиливается, они закупоривают бронхиолы, что приводит в тяжелых случаях к смерти от асфиксии.
Анализ смертности больных СПИДом в США, Европе и Японии показал, что в 70% случаев основной причиной смерти явился пневмоцистоз. Имеютсясведения о том, что если больных СПИДом удается спасти от пневмоцистоза, то жизнь их значительно продлевается.

Рис. 19.6. Пневмоцистис. А —
паразиты (о) на клетках эпителия альвеол (б); Б —
цикл развития: 1—4 —
паразиты в процессе деления, 5—7 —
образование спор
Обнаружить Р.
carinii
в мокроте больных оказывается крайне сложно. Поэтому для диагностики используют иммунологические методы, которые не обладают высокой специфичностью. В настоящее время разработан метод диагностики пневмоцистоза с использованием генетических зондов. У погибших больных диагноз ставят на основании обнаружения паразитов на разных стадиях их жизненного цикла в гистологических срезах легких.
Профилактика пневмоцистоза сложна в связи с широкой распространенностью паразита у здоровых людей и домашних животных. Поэтому эффективной мерой профилактики является только вакцинация людей с иммунодефицитом специальными вакцинами, получаемыми методами генетической инженерии.
Простейшие, обитающие в тканях, являются более специализированными паразитами по сравнению с предыдущей группой организмов. В связи с этим у большинства из них, особенно живущих внутрикле-точно, обнаруживаются признаки глубокой дегенерации:
исчезают орга-неллы передвижения и питания, форма тела становится непостоянной, поглощение питательных веществ осуществляется всей поверхностью тела за счет пиноцитоза и активного транспорта через мембраны.
На протяжении эволюции у них выработались адаптации к проникновению в организм хозяина разными способами, к перемещению в тканях. Многие из них инвазируют хозяина трансмиссивным путем, другие используют промежуточных хозяев, которыми питаются основные. Циклы развития большинства простейших этой группы сильно усложняются и часто являются лабильными: в ряде случаев возможным становится заражение одного промежуточного хозяина от другого, минуя основного. Часть паразитов этой группы адаптировалась к человеку не только как к биологическому, но и как к социальному существу, используя такие необычные факторы заражения, как хирургический инструмент и медицинские манипуляции.
Длительный контакт этих паразитов с иммунной системой хозяина выработал своеобразные адаптации к. избеганию действия иммунитета хозяина:
наиболее специализированные паразиты этой группы обитают не просто в тканях, а внутри клеток, в том числе даже в клетках иммунной системы. Другие, находящиеся в тканевой жидкости, в межклеточных пространствах и плазме крови, вынуждены на протяжении всего онтогенеза постоянно подвергаться действию гуморальных факторов иммунитета. В результате у них возникли своеобразные особенности клеточной поверхности, выражающиеся в том, что постоянно меняется ее антигенный состав.
Простейших, обитающих в тканях, следует подразделить на передающихся нетрансмиссивно и передающихся трансмиссивно.
19.3.1. Простейшие, обитающие в тканях
и передающиеся нетрансмиссивно
К этой группе простейших относятся близкие друг другу виды кокцидий (кл. Споровики), ведущих внутриклеточный образ жизни. Они паразитируют не только у человека, но и у большого количества видов домашних и диких животных. Таким образом, вызываемые ими заболевания являются зоонозными и природно-очаговыми, профилактика которых осложнена. Не менее сложна и диагностика, что связано, с одной стороны, со слабой выраженностью симптомов заболеваний, а с другой — с тем, что паразиты могут обитать в разных клетках и тканях хозяина и вероятность обнаружения их невелика.
Токсоплазма
Toxoplasma gondii —
возбудитель токсоплазмоза.
Имеет форму полумесяца, один конец которого заострен более другого. В центре располагается крупное ядро. Длина паразита 4—7 мкм (рис. 19.7).
Рис. 19.7. Токсоплазма (Л) и саркоциста (Б
):
1
— единичные паразиты, 2
— спорозоиты под общей оболочкой, 3 —
изолированная спора, 4—
саркоциста в мышечном волокне
Токсоплазма поражает огромное количество видов животных и человека. Иммунологические исследования показали, что на Земле токсоплазмами заражено более 500 млн. человек.
Жизненный цикл токсоплазмы типичен для споровиков: в нем чередуются стадии шизогонии, гаметогонии и спорогонии.
Основные хозяева паразита — домашние кошки и дикие виды сем Кошачьи. Они заражаются, поедая больных грызунов, птиц или инвазированное мясо крупных животных. Паразиты у них сосредоточиваются в клетках кишечника, размножаются шизогонией, а затем образуют гаметы. После копуляции гамет формируются ооцисты, которые выделяются во внешнюю среду. В них происходит спорогония, т. е. деление зиготы под оболочкой.
Такие спороцисты со спорозоитами рассеиваются кошками и попадают к промежуточным хозяевам, которыми могут быть человек, почти все млекопитающие, птицы и даже пресмыкающиеся. В клетках большинства их органов происходит бесполое размножение токсоплазм в форме множественного деления. В результате образуются группы, состоящие из многих сотен отдельных паразитов. Эти группы могут распадаться, и тогда отдельные токсоплазмы внедряются с помощью специфической органеллы проникновения — коноида —
в непораженные клетки, в которых вновь происходит шизогония.
Другие такие группы покрываются плотной оболочкой и формируют цисты. Цисты очень устойчивы и могут длительное время находиться в состоянии покоя в органах хозяев. В окружающую среду они не выделяются. Цикл развития замыкается при поедании кошками органов промежуточных хозяев с цистами.
Своеобразной особенностью цикла развития токсоплазм является то, что промежуточные хозяева могут заражаться ими не только от основного хозяина, но и при поедани» друг друга. Так, возможно заражение свиней при поедании ими трупов грызунов, погибших от токсоплазмоза, грызуны же заражаются друг от друга при каннибализме. Возможно и внутриутробное заражение плода от больной беременной самки, когда паразиты проникают через плаценту. Этот способ заражения обеспечивает устойчивое существование природных очагов токсоплазмоза и среди мелких грызунов, не склонных к каннибализму.
В соответствии с этим и человек как промежуточный хозяин может заразиться токсоплазмозом разными путями: 1) при поедании мяса инвазированных животных; 2) с молоком и молочными продуктами; 3) через кожу и слизистые оболочки при уходе за больными животными, при обработке шкур и разделке животного сырья; 4) внутриутробно через плаценту; 5) при медицинских манипуляцих переливания крови и лейкоцитарной массы, при пересадках органов, сопровождающихся приемом иммунодепрессивных препаратов. Последнее свидетельствует о том, что общее снижение иммунитета повышает вероятность заражения токсоплазмозом.
Обычно паразиты обладают весьма низкой патогенностью, но в некоторых условиях они могут вызвать очень тяжелые нарушения, что зависит как от индивидуальной чувствительности хозяев, так и от путей проникновения токсоплазм в организм человека.
Наиболее опасным является трансплацентарное заражение.
При этом возможно рождение детей с множественными врожденными пороками развития, в первую очередь головного мозга. При постановке диагноза используют методы иммунологических реакций, обнаружение токсоплазм при прямом микроскопировании материала, взятого от больного человека или трупа. Для исследования используют плаценту, печень, кровь, лимфатические узлы, головной мозг. Применяют также метод биологических проб. В этом случае лабораторным животным вводят кровь или спинномозговую жидкость больного. Мыши заболевают токсоплазмозом при таком способе заражения в острой форме, и обнаружение возбудителя у них не представляет сложности.
Профилактика — термическая обработка животных продуктов питания, санитарный контроль на бойнях и мясокомбинатах, предотвращение тесных контактов детей и беременных женщин с домашними животными.
Несколько близких к токсоплазме паразитов — саркоцисты
(Sarcocystis hominis, S. suihominis, S. lindemanni
) —
являются возбудителями саркоцистозов
и имеют сходный с ней цикл развития. Человек для этих паразитов является основным хозяином, а животные — промежуточными. Поэтому у человека, как и у кошки при токсоплазмозе, поражается кишечник. Но степень поражения его очень незначительна. По некоторым данным, частота инвазии людей саркоцистами достигает 7—60%, особенно там, где в соответствии с традициями население употребляет в пищу сырое или полусырое мясо. Врачи обычно не ставят правильного диагноза, а заболевания заканчиваются быстрым самоизлечением.
19.3.2. Простейшие, обитающие в тканях
и передающиеся трансмиссивно
В эту экологическую группу паразитических простейших входит большое количество видов из классов Жгутиковые и Споровики, имеющих сложные циклы развития. Все переносчики этих паразитов являются специфическими. Распространенность заболеваний приурочена к зонам встречаемости переносчиков. Некоторые паразиты этой группы поселяются только в организме человека, поэтому соответствующие заболевания являются антропонозными.
Другие формируют устойчивые природные очаги, поражая разные виды диких животных. Цист эти паразиты не образуют и непосредственно во внешнюю среду не выделяются. Диагностика заболеваний, вызываемых этими паразитами, основана на обнаружении их трофозоитов в крови и биоптатах тканей или на иммунологических реакциях.
Профилактика включает общие меры профилактики трансмиссивных заболеваний (борьба с переносчиками) и конкретные, зависящие от видовой специфики паразитов.
Лейшмании
Leischmania
(кл. Жгутиковые) — возбудители лейшманиозов.
Заболевания человека вызываются несколькими видами и подвидами паразитов, которые объединяются в четыре комплекса:L. donovani
—
возбудитель висцерального лейшманиоза,L. tropica —
возбудитель кожного лейшманиоза,L. mexicana —
возбудитель лейшманиоза Центральной Америки,L. brasiliensis —
возбудитель бразильского лейшманиоза. Все виды сходны морфологически и имеют одинаковые циклы развития. Они существуют в двух формах: в безжгутиковой, или лейшманиальной, и жгутиковой; или промастиготной (рис. 19.8).
Лейшманиальная форма очень мелка — 3—5 мкм в диаметре. Характерной чертой ее является круглое ядро, занимающее около '/4 цитоплазмы; жгутика нет, но перпендикулярно клеточной поверхности располагается палочковидный кинетопласт. Эти формы обитают в клетках ретикулоэндотелиальной системы человека и ряда млекопитающих (грызунов, собак, лис). Промастиготная форма удлинена — до 25 мкм, спереди находится жгутик, у основания которого хорошо виден такой же кинетопласт, что и в безжгутиковой стадии паразита. Обитает в пищеварительной системе москитов. Безжгутиковая форма, посеянная на культуральную среду, превращается в жгутиковую.
Рис. 19.8. Лейшмания. Безжгутиковая (А
) и жгутиковая (Б
) формы
Лейшманиозы широко распространены в странах с тропическим и субтропическим климатом на всех континентах там, где обитают москиты. Они—типичные природно-очаговые заболевания (см. § 18.13). Природными резервуарами являются грызуны, дикие и домашние хищники. Заражение человека происходит при укусе инвазированными москитами.
По патогенному действию лейшманий заболевания, которые они вызывают, делят на три основные формы: кожный, слизисто-кожный и висцеральный лейшманиозы.
При кожном лейшманиозе
очаги поражения находятся в коже. Это самый распространенный тип лейшманиоза, протекающий относительно доброкачественно. Возбудителями кожного лейшманиоза в Африке и Азии являютсяL. tropica,
а в Западном полушарии — L. mexicana
и ряд штаммовL. brasiliensis.
ЛейшманийL. tropica
и L.
mexicana
вызывают на коже длительно не заживающие язвы на месте укусов москитами. Язвы заживают через несколько месяцев после образования, а на их месте на коже остаются глубокие рубцы. Некоторые формыL. brasiliensis
способны распространяться по лимфатическим сосудам кожи с образованием многочисленных кожных язв в отдалении от мест укусов.
Слизисто-кожный лейшманиоз
вызывается подвидомL. brasiliensis brasiliensis.
При этой форме заболевания паразиты проникают из кожи по кровеносным сосудам в носоглотку, гортань, мягкое нёбо, половые органы, поселяются в макрофагах соединительных тканей этих органов и вызывают здесь деструктивные воспаления.
Висцеральный лейшманиоз
вызываетL. donovani.
Заболевание начинается через несколько месяцев или даже лет после заражения как системная инфекция. Паразиты размножаются в макрофагах и в моноцитах крови. Нарушаются функции печени, кроветворение. Очень велика интоксикация. При отсутствии лечения заболевание заканчивается смертью.
Лабораторная диагностика
основана на микроскопировании мазков из кожных язв при кожном и слизисто-кожном лейшманиозах, пунктатов лимфатических узлов и костного мозга при висцеральном лейшманиозе. В окрашенных препаратах обнаруживается лейшманиальная форма паразитов как внутри клеток, так и внеклеточно. В сомнительных случаях производят посев материала, взятого от больного, на специальную культуральную среду, на которой лейшманий приобретают промастиготную форму, активно передвигаются и легко обнаруживаются при микроскопировании. Используют также и биологические пробы — заражение лабораторных грызунов.
Профилактика —
в первую очередь, это борьба с переносчиками и уничтожение природных резервуаров (грызунов и бродячих собак), а также профилактические прививки.
Trypanosoma brucei gambiense
иT.b. rhodesiense
(кл. Жгутиковые) — возбудители африканского трипаносомоза,
или сонной болезни.
Паразит имеет извилистую заостренную с обеих сторон форму. Длина его 17—28 мкм. Стадии, паразитирующие у человека, имеют один жгутик, ундулирующую мембрану сбоку и хорошо заметный кинетопласт у основания жгутика.
Трипаносомы поселяются у человека в крови, лимфе, спинномозговой жидкости, в тканях головного и спинного мозга и в серозных полостях.T.b. gambiense
встречается в Западной Африке, а T.b. rhodesiense —
в Восточной и Юго-Восточной Африке.
Жизненный цикл
этих паразитов протекает в организме человека, домашних и диких млекопитающих, в первую очередь копытных. T.b. gambiense
чаще поражает человека, свиней и собак,T.b. rhodesiense —
диких животных — антилоп и носорогов. Переносчиком первого подвида является муха це-це,Glossina palpalis,
живущая поблизости от жилища человека, второго—G. morsitans,
обитающая в открытых саваннах и саванновых лесах. В связи с этим сонная болезнь, возбудителем которой являетсяT.b. gambiense,
встречается в антропогенных очагах культурных ландшафтов. Ежегодно регистрируется около 10 000 новых случаев заражения. Восточноафриканский трипаносомоз распространен значительно реже в естественной природе. В основном заболевают охотники, туристы, сезонные рабочие, каждый год — около 1500 человек.
Сонная болезнь без лечения протекает около 5 лет и выражается в нарастающей мышечной слабости, депрессии, истощении и сонливости. Возможны случаи самоизлечения, но обычно заболевание заканчивается смертью больного.
Восточноафриканский трипаносомоз протекает более злокачественно, длится не более 6 мес. и также заканчивается смертью.
Для паразитирования трипаносом у млекопитающих и человека характерны циклические подъемы интенсивности инвазии за счет их размножения, сопровождающиеся изменениями строения и антигенных свойств паразитов. Во время увеличения количества паразитов в крови преобладают трипаносомы удлиненной формы. Антигены, которые они образуют, вызывают формирование антител в организме хозяина. Под действием антител многие паразиты гибнут и интенсивность инвазии снижается. Выжившие трипаносомы укорачиваются и начинают вырабатывать другие антигены. Укороченные формы паразита, инвазионные для мухи це-це, в ее организме вновь приобретают удлиненную форму, инвазионную для человека. Изменение формы тела и смена антигенных свойств оболочки повторяются многократно. Таким образом, популяция паразита в хозяине выживает и избегает его иммунной реакции.
Антигенные свойства поверхности трипаносомы зависят только от одного белка — гликопротеина, полностью покрывающего всю клетку. Гликопротеин построен из 470 остатков аминокислот. Каждая новая волна размножения паразитов представляет собой новую популяцию трипаносом, обладающих новым поверхностным антигеном. Эти вариации антигенных свойств помогают паразиту преодолевать иммунный ответ хозяина и делают невозможной вакцинацию населения, обитающего в природных очагах трипаносомозов.
Смена антигенных свойств обеспечивается заменой поверхностных гликопротеинов, кодируемых разными генами, относящимися к одному мультигенному семейству. Один клон трипаносом может образовывать попеременно до 100 разных варьирующих гликопротеинов. Гены этих протеинов возникли в процессе эволюции, вероятно, путем дупликаций и последующей дифференцировки, как и другие семейства генов. В геноме трипаносомы имеется сайт экспрессии, в который поочередно перемещаются гены поверхностных гликопротеинов, приближаясь к промотору, обеспечивающему их специфическую активацию. Там они транслируются. Не исключено, однако, что сайт экспрессии в геноме трипаносомы не единственный, и даже возможно, что разные гены гликопротеинов активируются несколькими механизмами. В любом случае речь идет о своеобразной адаптации паразита к специфическим условиям существования, повышающей его выживаемость и открывающей ему широкие эволюционные перспективы.
Лабораторная диагностика —
исследование мазков крови и спинномозговой жидкости больного для выявления в них возбудителя. Используются также иммунологические реакции и заражение лабораторных животных.
Профилактика —
кроме борьбы с переносчиками применяют профилактическое лечение здоровых людей, живущих в очагах трипаносомоза, делающее организм невосприимчивым к инвазии. Практиковавшийся ранее отстрел диких животных, являющихся природным резервуаром паразита, вряд ли рационален в связи с возможным нарушением экологического баланса, который складывался в биогеоценозах на протяжении тысячелетий.
Trypanosoma cruy —
возбудитель американского трипаносомоза,
или болезни Чагаса.
Длина этой трипаносомы в крови человека достигает 20 мкм. Кинетопласт очень крупный, округлой формы. Характерной особенностью возбудителя является способность к внутриклеточному паразитизму. При этом трипаносомы проникают вначале в макрофаги кожи и слизистых оболочек, а затем и в клетки миокарда, нейроглии и мышц, теряя жгутики, ундулирующие мембраны, и превращаясь в безжгутиковые, или амастиготные, формы. Здесь и происходит размножение паразитов. В крови эти трипаносомы никогда не делятся. В конечном счете, пораженная клетка вся заполняется амастиготными формами трипаносом и разрывается, а паразиты инвазируют новые клетки. При этом часть их, превращаясь вновь в жгутиковую форму, поступает в кровь, откуда в дальнейшем они могут попасть в организм переносчика (рис. 19.9).
Рис. 19.9. Трипаносома. Безжгутиковая (А
) и жгутиковые {Б—Г
) формы
Переносчиками являются триатомовые клопы
pp. Triatoma, Rhodnius
иPanstrongylus
(см. разд. 21.2.2). В них трипаносомы размножаются и достигают состояния инвазионности, поступая в заднюю кишку. Вскоре после кровососания клопы испражняются на покровы человека или животного и трипаносомы проникают в кровь через раневое отверстие от хоботка или через неповрежденные слизистые оболочки губ, носа и глаз. Окончательными хозяевами кроме человека являются броненосцы, опоссумы, крысы, обезьяны и домашние животные — собаки, кошки и свиньи.
Болезнь поражает в основном детей младшего возраста, у которых протекает остро. В старшем возрасте заболевание переходит в хроническую форму.
Патогенное действие выражается в поражении органов, в клетках которых развиваются паразиты: характерны миокардиты, кровоизлияния в мозговые оболочки и менингоэнцефалит. Иногда заболевание протекает легко и заканчивается самопроизвольным излечением.
Диагностика —
в острой форме заболевания возможно обнаружение трипаносом в крови. При хроническом течении рационально введение крови больного морским свинкам, у которых они обнаруживаются в большом количестве на 14-е сутки. Существует еще и своеобразный метод диагностики — кормление на больном неинвазированных переносчиков-клопов, в кишечнике которых трипаносомы быстро размножаются и легко обнаруживаются. Применяют также и методы иммунодиагностики.
Трипаносомы представляют большой интерес не только потому, что способны вызывать у человека серьезные, смертельно опасные заболевания. Колоссальная экологическая пластичность, обеспечивающая им эффективное паразитирование в организме хозяина на протяжении нескольких лет, в условиях постоянно действующих механизмов иммунитета, позволяет этим паразитам осваивать и новых хозяев. Так, описанная в последние годы вспышка трипаносомоза в Эфиопии была вызвана адаптацией к человеку Т.b.
brucei —
подвида, паразитирующего обычно только у крупного рогатого скота и антилоп. Та же причина, вероятно, лежит в основе существования вспышек трипаносомозов в Индии и Малайзии, где у больных были выделены паразиты, обитающие обычно в крови грызунов и низших обезьян. В Центральной и Южной Америке кроме Т.
cruzy
известна и еще одна трипаносома — Т.
rangeli,
которая чаще паразитирует у кошек и собак, но способна инвазировать и человека, вызывая у него нетяжелую, быстро проходящую лихорадочную реакцию.
Малярийные плазмодии
Plasmodium
(кл. Споровики) — возбудители малярии.
Известны следующие виды малярийных плазмодиев, паразитирующие у человека: Р.
vivax —
возбудитель трехдневной малярии, Р.
falciparum
—
возбудитель тропической малярии, Р.
malariae —
возбудитель четырехдневной малярии, Р.
ovale—
возбудитель овале-малярии, близкой к трехдневной. Три первых вида широко распространены в тропических и субтропических климатических поясах, последний — только в тропической Африке. Все виды сходны морфологически и жизненными циклами, отличаясь друг от друга деталями строения и некоторыми особенностями цикла развития, проявляющимися в основном продолжительностью его отдельных периодов.
Жизненный цикл малярийных плазмодиев типичен для споровиков, включая стадии бесполого размножения в виде шизогонии, полового процесса и спорогонии. Окончательным хозяином паразитов является комар р.
Anopheles
(см. разд. 21.22), а промежуточным — только человек. Комар является одновременно и переносчиком. Поэтому малярия — типичное антропонозное трансмиссивное заболевание.
Со слюной зараженного комара при укусе плазмодии попадают в кровь человека (рис. 19.10). Развитие паразитов в организме человека происходит синхронно. С током крови они разносятся по организму и поселяются в клетках печени. Здесь они растут и размножаются шизогонией таким образом, что один паразит делится на тысячи дочерних особей. Клетки печени при этом разрушаются и паразиты, называющиеся на этой стадии мерозоитами,
поступают в кровь и внедряются в эритроциты. С этого момента начинается эритроцитарная часть цикла развития плазмодия. Паразит питается гемоглобином, растет и размножается шизогонией. При этом каждый плазмодий делится на 8—24 мерозоита. После разрушения эритроцита мерозоиты попадают в плазму крови и оттуда в новые эритроциты, после чего весь цикл эритроцитарной шизогонии повторяется.
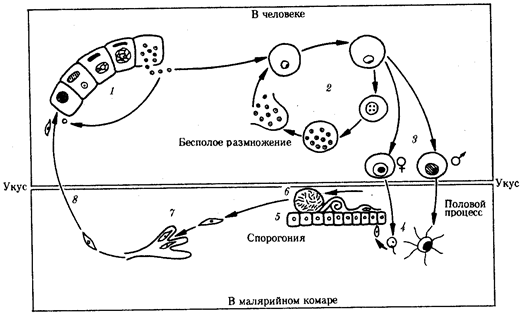
Рис. 19.10. Жизненный цикл малярийного плазмодия:
1
— преэритроцитарная шизогония в клетках печени, 2 —
эритроцитарная шизогония, 3 —
образование гаметоцитов, 4 —
оплодотворение, 5
— спорогония в стенке желудка комара, 6
— овоциста со споромитами, 7
— проникновение спорозоита в слюнные железы комара, 8—
заражение человека
Из части мерозоитов в эритроцитах образуются незрелые половые клетки — мужские и женские гаметоциты.
Они являются инвазионной стадией для комара. Дальнейшее их развитие возможно только в его пищеварительной системе. При укусе больного человека комаром гаметоциты попадают в желудок последнего, где из них образуются зрелые гаметы. В результате оплодотворения в желудке комара образуется подвижная зигота, которая перемещается на наружную поверхность стенки желудка и покрывается оболочкой, формируя ооцисту.
С этого момента начинается период спорогонии, когда содержимое ооцисты многократно делится, образуя около 10000 спорозоитов — тонких серповидных клеток, которые после разрыва оболочки поступают в слюнные железы комара. При кровососании спорозоиты поступают в кровяное русло человека.
Таким образом, в организме человека плазмодий размножается только бесполым путем — шизогонией, человек является его промежуточным хозяином. В организме комара проходят две другие стадии цикла развития паразита: половой процесс — гаметогония и образование спорозоитов за счет деления под оболочкой ооцисты — спорогония. Поэтому малярийный комар является окончательным хозяином этого паразита.
Выход большого количества мерозоитов из эритроцитов сопровождается выбросом в плазму крови значительной массы токсических продуктов жизнедеятельности. Их воздействие на организм приводит к резкому повышению температуры, ознобу, слабости и головным болям. Такое состояние возникает внезапно и длится в среднем 1,5—2 ч. Вслед за этим наступает чувство жара, сухость во рту, жажда. Температура тела достигает 40—41°С. Через несколько часов все перечисленные симптомы исчезают, и больные обычно засыпают. Весь приступ может продолжаться от 6 до 12 ч. При трехдневной и овале-малярии промежутки между приступами составляют 48 ч, число таких приступов может достигать 10—15, после чего они прекращаются за счет повышения уровня специфического иммунитета, но паразиты в крови еще могут обнаруживаться. В таком случае человек становится паразитоносителем и продолжает представлять опасность для окружающих как возможный источник заражения.
Естественный отбор приводит к возникновению новых антигенных вариантов возбудителя, которые обеспечивают возможность наступления рецидивов заболевания. Рецидивы могут повторяться несколько раз, но постепенно популяция эритроцитарных паразитов полностью погибает. Однако в течение 3—5 лет инвазия может вновь активизироваться за счет находящихся в латентном состоянии в печени экзоэритроцитарных шизонтов, которые могут выходить из печеночных клеток и внедряться в эритроциты. Таким образом, весь процесс болезни может начаться снова.
При малярии, вызываемой Р.
malariae,
приступы повторяются через 72 ч. Часто встречается и бессимптомное носительство. Экзоэритро-цитарной стадии в цикле развития этого паразита нет, поэтому поздние рецидивы невозможны, хотя инвазия характеризуется упорным течением и длится до 40 лет.
При тропической малярии вначале приступы развиваются через разные промежутки времени, а позже — через 24 ч. От осложнений со стороны центральной нервной системы или почек возможна смерть больного. Шизонты в клетках печени не сохраняются, а заболевание может продолжаться до 18 мес.
Все виды малярийных плазмодиев могут инвазировать человека и при гемотрансфузии (переливание крови). В этом случае ни у одного из паразитов не формируется экзоэритроцитарной стадии. Поэтому поздних рецидивов в этом случае не бывает. Гемотрансфузионный способ заражения наиболее часто встречается при четырехдневной малярии в связи с тем, что при этой форме болезни шизонты в эритроцитах находятся в очень малом количестве и могут не обнаруживаться при исследовании крови доноров.
Иногда человек может быть инвазирован одновременно двумя или тремя видами плазмодиев. В таком случае малярийные приступы не имеют четкой периодичности и клинический диагноз затруднен.
Лабораторный диагноз малярии можно поставить только в период, соответствующий стадии эритроцитарной шизогонии, когда в крови удается обнаружить паразитов.
Плазмодий, недавно проникший в эритроцит, имеет кольцевидную форму.
Его цитоплазма выглядит как ободок, окружающий крупную вакуоль с продуктами диссимиляции. Ядро паразита смещено к краю клетки. Следующая стадия называется амебовидным шизонтом.
У паразита появляются ложноножки, а вакуоль увеличивается. Наконец плазмодий занимает почти весь эритроцит. Следующая стадия развития паразита — фрагментация шизонта.
На фоне деформированного эритроцита обнаруживаются множественные мерозоиты, в каждом из которых лежит ядро. Кроме бесполых клеток в эритроцитах можно увидеть и гаметоциты. Они отличаются крупными размерами, не имеют псевдоподий и вакуолей (рис. 19.11).
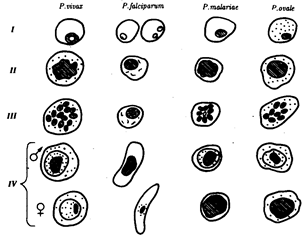
Рис. 19.11. Малярийные плазмодии. Стадии развития в эритроцитах:
I
—стадия кольца, II
—стадия амебовидного шизонта, III
—стадия фрагментации, IV—
гаметоциты
Профилактика малярии — раннее выявление и лечение больных, профилактическое лечение в зонах широкого распространения малярии. Как и при любых трансмиссивных заболеваниях, необходима прицельная борьба с переносчиками.
В 1965 г. в Австралии впервые были выявлены случаи заболеваний, вызываемых свободноживущими почвенными амебами, и с тех пор они регистрируются во многих странах. В большинстве случаев эти заболевания диагностируются только после смерти больных на основе гистологического исследования тканей, в которых обнаруживаются эти простейшие.
Крайняя тяжесть заболеваний, вызываемых этими амебами, объясняется тем, что паразитический образ жизни для них не является обязательным. Поэтому как у амеб, так и у человека на протяжении эволюции не возникло взаимных адаптации, в связи с чем система паразит — хозяин не является устойчивой.
Среди амеб этой группы наиболее известны представители родовNaegieria
и Acanthamoeba
(рис. 19.12).
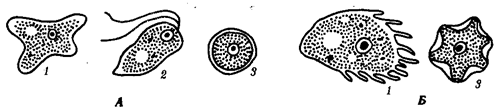
Рис. 19.12. Факультативные паразиты человека. А —
Naegieria sp.;
Б —
Acanthamoeba sp.:
1
, 2 —
вегетативные стадии, 3 —
цисты
Амебы р. Naegieria
в организм человека попадают при купании в грязной воде через носовую полость и проникают в мозговые оболочки. Здесь амебы размножаются и вызывают острый менингоэнцефалит, который почти всегда заканчивается смертью. Наиболее часто поражаются дети.
Амебы р.Acanthamoeba
образуют устойчивые цисты, которые попадают в организм человека не только через носоглотку и пищеварительную систему, но и при вдыхании, а также через травмированную кожу и роговицу. Заболевание проявляется по-разному в зависимости от путей попадания возбудителя в организм. Характерным является образование гранулем, содержащих амеб. У ослабленных больных и детей заболевание заканчивается менингоэнцефалитом и смертью.
Медицинская гельминтология изучает паразитов человека из группы червей. Черви—
это многоклеточные, трехслойные, первичноротые, двусторонне-симметричные животные. Их тело имеет удлиненную форму, а кожно-мускульный мешок состоит из гладких или поперечно-полосатых мышц и покровных тканей.
Черви — паразиты человека, относятся к типам Плоские
и Круглые
черви. Заболевания, вызываемые гельминтами, называют гельминтозами.
В большинстве случаев при одноразовой инвазии нарастания численности гельминтов в организме хозяина не. происходит: для успешного протекания циклов их развития необходима смена сред обитания. Из этого следует, что продолжительность заболевания
часто определяется продолжительностью жизни паразита и колеблется от нескольких недель при энтеробиозе до нескольких десятков лет при шистосоматозах. Тяжесть заболевания
зависит от количества паразитов, попавших в организм хозяина, и его индивидуальной чувствительности.
Гельминты могут обитать у человека практически во всех органах. В соответствии с этим различны пути проникновения их в организм человека, симптоматика заболеваний и методы диагностики.
На протяжении длительной эволюции при переходе к паразитизму у гельминтов возникли не только признаки общей дегенерации и адаптации к паразитическому образу жизни общего значения (особые покровы, мощная половая система), но и конкретные приспособления к обитанию в определенных органах. Хозяева, в свою очередь, приобрели соответствующие адаптации, обеспечивающие стабильное существование системы хозяин — паразит. Однако во многих случаях для попадания в определенный орган гельминты осуществляют миграцию по кровеносным сосудам или непосредственно через ткани и могут попасть в другие органы. Тогда говорят об атипичной,
или эктопической,
локализации. Заболевание при этом протекает тяжелее, диагностируется с трудом и часто может заканчиваться гибелью одного из компонентов системы: паразита или хозяина. Сказанное в полной мере относится и к видовой специфичности системы паразит — хозяин: гельминты, адаптированные только к человеческому организму, обусловливают более легкое течение заболевания у него, чем специфические паразиты животных, попадающие к человеку случайно.
Плоские черви имеют тело, уплощенное в дорсовентральном направлении. Полость тела отсутствует,
внутренние органы погружены в рыхлую соединительную ткань — паренхиму.
Кожно-мускульный мешок состоит из покровной ткани — тегумента, который представляет собой многоядерную неклеточную структуру, и трех слоев гладких мышц — продольных, поперечных и дорсовентральных. Движения, осуществляемые ими, медленны и несовершенны. Нервная система
состоит из нервных узлов на переднем конце тела, от которых кзади отходят продольные нервные тяжи. Пищеварительная система,
если она имеется, построена из глотки и кишечника, который слепо замкнут. Непереваренные остатки пищи выделяются через рот. Половая система
гермафродитна и построена очень сложно.
Выделение осуществляется с помощью протонефридиальной системы,
состоящей из отдельных выделительных клеток — протонефридиев. Они способны захватывать продукты диссимиляции и транспортировать их по внутриклеточным каналам, проходящим в их длинных отростках. Продукты экскреции поступают в собирательные трубочки, а оттуда либо непосредственно, либо через мочевой пузырь — во внешнюю среду.
Виды, имеющие медицинское значение, представлены в двух классах: Сосальщики и Ленточные черви.
20.1.1. Класс Сосальщики
Trematoda
Класс Сосальщики включает около 4000 видов. Все они паразиты. Форма тела большинства сосальщиков листовидная. В процессе адаптации к паразитизму выработались мощные присоски — органы прикрепления. Многие сосальщики, кроме того, имеют мелкие шипики, покрывающие все тело и облегчающие им прикрепление к хозяину. Мелкие виды имеют пищеварительную систему в форме мешка или двух слепо замкнутых каналов. У крупных видов пищеварительная система сильно разветвлена и наряду с собственно пищеварением выполняет также транспортную функцию, перераспределяя по организму продукты пищеварения. В остальном сосальщики повторяют организацию плоских червей, описанных выше (рис. 20.1).
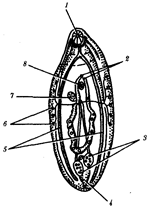
Рис. 20.1. Организация сосальщиков: 1
—
ротовая присоска, 2—
мужские и женские половые отверстия; 3—
семенники; 4—
мочевой пузырь; 5—
ветви кишечника, 6—
желточники; 6—
яичник; 8—
брюшная присоска
Сосальщики резко обособлены от других плоских червей своеобразием жизненного цикла,
в котором имеет место закономерное чередование поколений, способов размножения и хозяев (рис. 20.2). Половозрелая стадия всегда паразитирует в организме позвоночных животных. Выделяемое яйцо для успешного развития обычно должно попасть в воду. Из него выходит личинка — мирацидий, —
снабженная светочувствительными глазками и ресничками, с помощью которых она свободно перемещается. Личинка обычно способна активно отыскивать промежуточных хозяев, используя фото-, гео- и хемотаксис.
Мирадиции попадают в организм брюхоногого моллюска
определенного вида, строго специфичного для данного сосальщика. Здесь личинка превращается в материнскую спороцисту —
стадию, претерпевшую в связи с паразитизмом наиболее глубокую дегенерацию. В ней развиты почти исключительно органы женской половой системы, благодаря которым она размножается партеногенетически.
В результате этого образуются многоклеточные редии,
которые также способны к партеногенезу. Таким образом, возможно формирование нескольких поколений редий. Последнее из них генерирует церкариев,
покидающих организм моллюска и свободно плавающих в поисках основного или второго промежуточного хозяина.
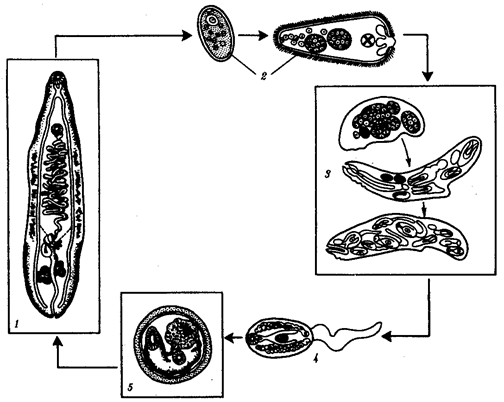
Рис. 20.2. Цикл развития сосальщиков:
1
—половозрелая форма в окончательном хозяине, 2—
яйцо и мирацидий в воде, 3—
личинки, размножающиеся партеногенетически в моллюске, 4—
церкарий в воде, 5—
метацеркарий во втором промежуточном хозяине
В первом случае церкарий либо самостоятельно внедряются в кожу хозяина, либо, инцистируясь на растениях, оказываются проглоченными травоядными животными или человеком. Во втором случае церкарий отыскивают животных, использующихся основными хозяевами для питания, и образуют в них покоящиеся стадии — инцистированные метацеркарий.
Церкарий в отличие от мирацидиев не обладают хемотаксисом, а использование только гео- и фототаксиса, а также инцистирование на траве не позволяют им находить специфичных хозяев. Поэтому основная масса церкариев погибает либо не найдя хозяев вообще, либо попав в организмы таких видов, развитие в которых невозможно. После проникновения инвазионных стадий сосальщиков в основного хозяина они мигрируют у него в организме и находят тот орган, где достигнут половой зрелости и будут обитать всю последующую жизнь.
Отсутствие свободноживущих видов в классе Сосальщики на фоне общего примитивизма их организации, сложный цикл развития, в котором участвуют специфические промежуточные и основные хозяева, огромная плодовитость, достигающаяся разными способами размножения, и сохранившаяся связь с водной средой обитания свидетельствуют о древности паразитического образа жизни этих организмов. Среди хозяев сосальщиков наиболее специфичными являются именно моллюски: для каждого вида паразита, обитающего на стадии мариты
нередко у нескольких видов основных хозяев, первым промежуточным хозяином практически всегда является только один вид улиток.
Это означает, что на протяжении эволюции между сосальщиками и моллюсками возникли наиболее совершенные взаимные адаптации. Моллюски были, вероятно, первыми и единственными хозяевами сосальщиков еще в период, предшествующий возникновению позвоночных. Об этом свидетельствует также сохранившийся в цикле развития сосальщиков партеногенез —
рудиментарная форма полового размножения, — проходящий именно в организме моллюсков. Наряду с этими чертами в цикле развития имеются такие признаки, которые указывают на рекапитуляции свободного образа жизни: обязательный выход яиц во внешнюю среду и наличие активно плавающих расселительных стадий с органами чувств — мирацидиев и церкарий.
Возникновение и дальнейший биологический и морфофизиологи-ческий прогресс позвоночных, освоивших все благоприятные для жизни среды, открыли и сосальщикам широкие возможности адаптивной эволюции за счет приспособления к новым хозяевам. Адаптации сосальщиков к позвоночным как к новым средам обитания не ограничились только использованием их широкого видового разнообразия. Сосальщики адаптировались к обитанию в самых разнообразных органах, тканях и системах — от кожных покровов и органов чувств до кровеносных сосудов внутренних органов.
Однако большинство сосальщиков-паразитов человека обитает в пищеварительной системе; некоторые виды живут в легких, другие — в кровеносных сосудах брюшной полости и малого таза. Человек заражается сосальщиками в зависимости от вида разными способами: при контакте с водой и проникновении церкарий через кожу, при поедании продуктов животного происхождения с метацеркариями и при употреблении растений в пищу, если на их листьях инцистированы церкарии паразитов.
После попадания в организм человека большинство сосальщиков осуществляют сложные миграции по пути к органам своей окончательной локализации. Миграция происходит по кровеносным сосудам, непосредственно по пространствам между органами и по полости тела. Во время миграции сосальщики вызывают у хозяина тяжелые интоксикации и аллергические состояния, но диагностировать заболевание в этот момент крайне сложно. Заболевания, вызываемые сосальщиками, называют трематодозами.
Для диагностики
трематодозов используют методы обнаружения яиц в фекалиях, моче или мокроте в зависимости от локализации паразитов, а также аллергические пробы.
В связи с тем что сосальщики, обитающие у человека, поражают также и ряд других видов млекопитающих, соответствующие трематодозы относят к природно-очаговым зоонозным заболеваниям, поэтому их полная ликвидация практически невозможна.
В зависимости от особенностей цикла развития сосальщиков, паразитирующих у человека, можно разделить на следующие группы: 1) развивающиеся с одним промежуточным хозяином и обитающие в пищеварительной системе; 2) развивающиеся с одним промежуточным хозяином и обитающие в кровеносных сосудах; 3) развивающиеся с двумя промежуточными хозяевами.
К сосальщикам этой экологической группы относится незначительное количество видов. Они характеризуются большими размерами, ротовая и брюшная присоски их расположены на переднем конце тела близко друг от друга. Паразиты обитают в печени или тонком кишечнике крупных травоядных млекопитающих и человека. Заражение происходит при поедании зелени и овощей, поливаемых прудовой водой, с которой могут заноситься церкарии, инцистирующиеся на листьях, превращаясь в адолескарш.
Травоядные животные, в том числе и домашние, заражаются гораздо чаще, чем человек. Они и являются наиболее частым источником заражения человека. Поэтому заражаются трематодозами этой группы обычно люди в сельской местности.
Лабораторная диагностика —
обнаружение яиц этих сосальщиков в фекалиях.
Профилактика заболеваний —
тщательное мытье и термическая обработка овощей и зелени в районах, где огороды поливают водой из стоячих водоемов, а также выявление, лечение больных животных и санитарная охрана пастбищ.
Печеночный сосальщик
Fasciola hepatica
(рис. 20.3) — возбудитель фасциолеза.
Тело паразита листовидное, передний конец клювообразно оттянут. Матка невелика и розеткой располагается позади брюшной присоски. Кзади от матки находятся сильно разветвленные семенники, яичники, желточники и ветви кишечника. Яйца крупные, желтовато-коричневого цвета. Фасциолез встречается чаще в странах с теплым влажным климатом.
Жизненный цикл
печеночного сосальщика типичен для этой группы паразитов. Окончательные хозяева паразита — крупные травоядные млекопитающие и человек. Промежуточный хозяин — малый прудовик Lymnea truncatula.
Интересно, что на территории Австралии нет моллюсков этого вида, а фасциолез распространен широко. Этот паразит, будучи занесенным сюда вместе со скотом, адаптировался к другому промежуточному хозяину —L. tomentosa.
Заражение основного хозяина происходит при поедании травы с заливных лугов. Случаи заражения человека связаны с употреблением щавеля и особенно часто — водяного кресса — полуводного растения, широко употребляющегося в пищу в Западной Европе. В кишечнике основного хозяина личинки освобождаются от оболочек, пробуравливают его стенку и попадают в полость брюшины, откуда мигрируют в печень, проникают через печеночную ткань в жёлчные ходы, где через 3—4 мес. достигают половой зрелости и начинают откладывать яйца.
Рис. 20.3. Сосальщики, имеющие одного промежуточного хозяина.
А —
печеночный; Б —
фасциолопсис
Печеночный сосальщик гигантский
Fasciola gigantica
отличается от предыдущего вида большими размерами (до 75 мм) и слабым выступанием вперед передней части тела. Цикл развития, диагностика и профилактика не отличаются от предыдущего вида. В связи с крупными размерами этого вида фасциолез, вызываемый им, протекает более тяжело. У человека фасциолез, вызываемый гигантским печеночным сосальщиком, встречается в Юго-Восточной Азии, на Гавайских о-вах, в Узбекистане.
Фасциолопсис
Fasciolopsis buski
(рис. 20.3, Б
) —
возбудитель фасциолопсидоза.
Этот паразит по размерам соответствует гигантскому печеночному сосальщику. Характерны для него очень толстое тело и две неразветвленные кишечные трубки.
Жизненный цикл
фасциолопсиса отличается тем, что его взрослые формы обитают в тонком кишечнике человека, домашних и диких свиней. Промежуточные хозяева — водные моллюски р.Segmentina.
Церкарии инцистируются наиболее часто на плодах водяного ореха и водяного каштана. При их поедании происходит заражение основного хозяина. Миграция паразита здесь не осуществляется. Паразит вызывает хронические желудочно-кишечные расстройства, сильно истощающие больных.
Диагностика
и профилактика
фасциолопсидоза соответствуют описанным выше. Заболевание широко распространено в Южной и Юго-Восточной Азии.
К этой группе относятся так называемые кровяные сосальщики — шистосомы.
Это раздельнополые организмы. Самцы имеют широкое тело, а самки — шнуровидное и в половозрелом состоянии находятся в гинекофорном канале на брюшной стороне самцов. Присоски невелики и располагаются на переднем конце тела. Все кровяные сосальщики обитают в тропических широтах Азии, Африки и Америки.
В связи с тем что половозрелые шистосомы живут в кровеносных сосудах, яйца, которые откладывают самки, имеют специальные приспособления для выведения в полостные органы и далее во внешнюю среду: они снабжены шипами, через которые выделяются ферменты, растворяющие ткани хозяина. За счет этих ферментов яйца пробуравливают стенки сосудов, проникают в ткани, могут попадать в кишечник или мочевой пузырь в зависимости от вида паразита. Опасным и характерным для этих паразитов является гематогенный занос яиц в различные органы, где вокруг них возникают локальные воспалительные процессы.
Как и у всех сосальщиков, личиночные стадии, размножающиеся партеногенезом, развиваются в водных моллюсках. Характерными являются церкарии —
они имеют раздвоенный хвост, а на переднем конце — железы проникновения, с помощью которых проникают через кожу в кровеносную систему окончательного хозяина при нахождении его в воде (см. рис. 18.7). При этом они вызывают кожные поражения — церкариозы,
выражающиеся в появлении сыпи, зуде и других аллергических проявлениях. При массовом попадании церкарии в легкие возникает пневмония.
Церкариозы могут вызываться не только шистосомами, паразитирующими у человека, но и несколькими видами кровяных сосальщиков, обитающих у водоплавающих птиц. Эти виды церкарии у человека обычно погибают либо в коже, либо в легких.
Личинки шистосом, патогенных для человека, мигрируют по организму и оседают в венах брюшной полости и малого таза, где и достигают половой зрелости. Патогенное действие половозрелых шистосом выражается в токсико-аллергических реакциях хозяина и местных проявлениях: характерны кровотечения из пораженных органов, образование изъязвлений и полипов, склонных к злокачественному перерождению.
Диагностика
заключается в обнаружении яиц шистосом в фекалиях или моче. Проводят также аллергические внутрикожные пробы и иммунобиологические реакции в пробирке.
Профилактика шистосоматозов:
необходимо остерегаться длительных контактов с водой в зонах распространения этих паразитов. В связи с тем что церкарии могут проникать и через слизистые оболочки, для питья следует использовать только обеззараженную воду. В целях общественной профилактики необходима охрана водоемов от загрязнения необеззараженными сточными водами. В ряде случаев возможна также борьба с моллюсками — промежуточными хозяевами паразитов — разными способами. В Бразилии эффективным оказался метод биологической борьбы с шистосомами — использование рыбки гуппи — Lebistes reticulatus,
которая активно поедает церкарий, выходящих из зараженных моллюсков.
Для профилактики церкариозов необходимо воздержаться от купания в пресноводных водоемах, в которых обитают водоплавающие птицы.
У человека часто паразитируют три вида шистосом, распространенных в тропиках Азии, Африки и Америки. В Центральной и Южной Африке описаны случаи инвазии человека еще четырьмя видами, обычно паразитирующими у животных. Кожные поражения в виде церкариозов могут наблюдаться в любой климатической зоне, где обитают водоплавающие птицы — окончательные хозяева ряда видов шистосом.
Schistosoma haematobium
(рис. 20.4, А
) —
возбудитель мочеполового шистосоматоза.
Самец длиной до 1,5, а самка — до 2,0 см. Поверхность тела мелкобугристая. Яйца очень крупные, размером до 0,16 мм; на конце находится длинный шип.
Промежуточными хозяевами являются моллюски из р.Bullinus, Planorbis
илиPlanorbarius;
окончательными хозяевами — человек и обезьяны, у которых паразит после миграции поселяется в венах мочевого пузыря и органах половой системы.
Встречается от Африки до Юго-Западной Индии.
Характерным для мочеполового шистосоматоза является гематурия (кровь в моче), боли в надлобковой области, нередко образование камней в мочевыводящих путях. В зонах распространения этого заболевания рак мочевого пузыря встречается в 10 раз чаще, чем в областях, свободных от шистосоматоза.
Рис. 20.4. Кровяные сосальщики.
А —
Schistosoma haematobium;
Б —
Sch..mansoni;
В —
Sch.
japonicum
При диагностике
обнаруживают яйца в моче, а также характерные изменения мочевого пузыря и влагалища: изъязвления, полипозные разрастания и местные воспалительные процессы.
Schistosoma mansoni
(рис. 20.4, Б
) —
возбудитель кишечного шистосоматоза.
В отличие от предыдущего вида имеет несколько меньшую длину (до 1,6 мм) и крупнобугристую поверхность тела. Яйца по размерам соответствуют яйцамSch. haematobium,
но шип находится на боковой поверхности. Промежуточными хозяевами являются моллюски р.Biomphalaria,
а окончательными — человек, обезьяны, собаки и грызуны. Ареал распространения шире, чем у предыдущего вида: он охватывает Северную, Экваториальную и Юго-Восточную Африку, Юго-Западную Азию; паразит занесен и в Западное полушарие — в Бразилию, Венесуэлу, Гайану и на Антильские о-ва.
У человека заселяет брыжеечные вены толстого кишечника и систему воротной вены печени. В связи с этим поражения возникают в первую очередь в толстом кишечнике (явления колита, понос с примесью крови, возможен полипоз толстой кишки) и в печени (венозный застой и цирроз).
При диагностике
обнаруживают яйца в фекалиях.
Schistosoma japonicum
(рис. 20.4, В
) —
возбудитель японского шистосоматоза.
Этот паразит по размерам не отличается отSch. haematobium,
но поверхность его тела совершенно гладкая. Яйца более округлые, чем у описанных видов, а шип, расположенный на боковой поверхности, имеет очень малые размеры. Промежуточные хозяева — моллюски р.Oncomelania,
а окончательные — человек и большое количество видов диких и домашних млекопитающих — грызуны, собаки, копытные. Ареал охватывает Восточную и Юго-Восточную Азию.
Паразит локализуется в венах кишечника, поэтому проявления болезни и диагностика соответствуют описанному кишечному шистосоматозу. Распространение яиц паразита по кровеносным сосудам в различные органы, в том числе в головной мозг, встречается чаще, чем при других формах шистосоматоза.
Эти сосальщики обитают у человека в разных органах, чаще в пищеварительной системе. Характерной чертой их жизненного цикла является наличие второго промежуточного хозяина,
которым могут быть самые разнообразные животные, иногда даже не связанные с водной средой обитания. Эти вторые хозяева, являясь источником питания для окончательных хозяев, используются паразитами только как транспортные средства, облегчающие замыкание жизненного цикла. Поэтому, попав в их организм, церкарии сосальщиков превращаются в покоящиеся стадии — метацеркарии,
не мигрируют и не развиваются до тех пор, пока вместе с ними не будут съедены окончательными хозяевами.
Способность церкарии инцистироваться во внешней среде хорошо известна. Вероятно, в ряде групп сосальщиков в процессе эволюции возникли адаптации к инцистированию во втором промежуточном хозяине, что повышает вероятность как выживания, так и попадания к окончательному хозяину. Использование представителей не только разных видов, но даже разных классов и типов в качестве таких вторых промежуточных хозяев (рыбы, ракообразные, насекомые) свидетельствует о независимости эволюции разных групп сосальщиков в этом направлении и о том, что эта особенность их жизненного цикла возникла относительно недавно. Различные направления адаптивной эволюции сосальщиков этой группы привели к тому, что они заселили не только разные органы окончательных хозяев, но и разные среды, в том числе выйдя на сушу и утратив связь с первоначальной водной средой обитания.
Сосальщиков, имеющих двух промежуточных хозяев, можно подразделить на связанных в цикле развития с водной средой и не связанных, цикл развития которых происходит на суше.
Сосальщики, цикл развития которых связан с водной средой
Паразиты этой экологической группы распространены очень широко и представлены большим количеством видов. Их расселение зависит от наличия пресноводных водоемов и степени подвижности вторых промежуточных хозяев, которыми могут быть рыбы или ракообразные. Заболеваемость среди людей определяется в первую очередь этническими традициями питания: употребление сырой рыбы и ракообразных, экзотические способы консервации продуктов питания(строганина, слабое просаливание, поверхностная термическая обработка и т. д.), а также профессиональной принадлежностью (рыбаки и члены их семей, геологи и охотники, проводящие много времени в естественной природе).
Рис. 20.5. Сосальщики, имеющие двух промежуточных хозяев.
А—
Metagonimusjokogawai,
Б—
Nanophyetes salmincola;
В—
кошачий сосальщик; Г—
клонорхис китайский; Д—
легочный сосальщик; Е—
ланцетовидный сосальщик; Ж—
сосальщик поджелудочной железы
Указаны средние размеры паразитов
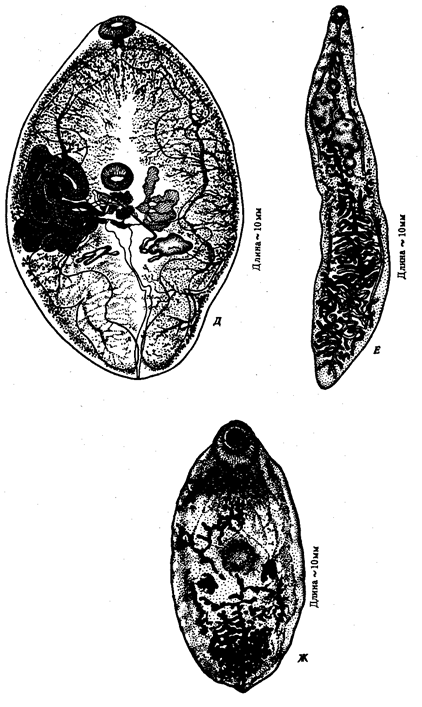
Продолжение рис. 20.5
Паразиты этой группы обитают у человека в тонкой кишке, в желчных ходах печени и в легких. Паразиты, живущие в тонкой кишке, при попадании в пищеварительную систему человека сразу задерживаются в кишечнике. Фаза миграции у них отсутствует. В связи с этим заболевания, вызываемые ими, протекают наиболее доброкачественно, часто бессимптомно, но иногда проявляются чередованием поносов и запоров.
Диагностика
основана на обнаружении яиц в фекалиях.
Вторым промежуточным хозяином являются разнообразные рыбы, у которых метацеркарии находятся на чешуе, плавниках, жабрах, реже в мышцах. Круг окончательных хозяев очень широк. Это рыбоядные птицы — пеликаны, бакланы, цапли и млекопитающие — норки, выдры, медведи, а также человек.
Паразиты этой группы распространены очень широко, но человека поражают лишь там, где этнические традиции питания способствуют этому.
Сосальщики, обитающие в кишечнике
Metagonimus yokogawai
(рис. 20.5, А
) —
возбудитель метагонимоза.
Это мелкий сосальщик длиной до 1,5 мм, тело его густо покрыто шипиками. Брюшная присоска расположена асимметрично, с правой стороны от средней линии. Яйца длиной до 0,028 мм.
Первыми промежуточными хозяевами являются моллюски из р.Melania,
вторыми — рыбы более 40 видов из сем. Карповые и Лососевые. У человека паразитирует в странах Дальневосточного региона, хотя у животных встречается также в Южной Европе.
Nanophyetes salmincola
(рис. 20.5, Б
) — возбудитель нанофиетоза.
Этот сосальщик имеет очень малые размеры — до 1,1 мм. Форма тела почти круглая. Яйца относительно крупные, до 0,056 мм длиной. Первый промежуточный хозяин — моллюски из р. Semisuicospira,
второй — рыбы сем. Лососевые, Хариусовые, Карповые и Подкаменщиковые. Окончательные хозяева — рыбоядные млекопитающие и человек. Встречается в бассейне р. Амур в Приморском крае, на Сахалине и на западном побережье Северной Америки. Заболевания человека описаны только у местного населения среднего и нижнего течения реки Амур.
Сосальщики, обитающие в желчных ходах печени
Особенностью паразитов этой группы является миграция личиной в организме человека, поэтому на первом этапе течения заболевания проявляются аллергические реакции. Длительное паразитирование большого количества этих сосальщиков в желчных ходах нарушает отток желчи, кровоснабжение и дает осложнения в 15—20% случаев цирротическими изменениями печени, а иногда и злокачественным перерождением печеночной ткани — возникновением первичного рака печени.
При диагностике
заболеваний следует исследовать фекалии для обнаружения яиц, а также провести дуоденальное зондирование, при котором возможно выделение через зонд не только яиц, но и половозрелых паразитов.
Кошачий сосальщик
Opisthorchisfelineus
(рис. 20.5, В
) —
возбудитель описторхоза.
Длина тела до 13 мм. Характерная особенность—два хорошо окрашивающихся лопастевидных семенника на заднем конце тела. Яйца длиной 26—30 мкм, с крышечкой. Описторхоз — эндемичное для России заболевание. Встречается у человека наиболее часто в Западной Сибири, но изредка проявляется и в европейской части СНГ — в Волжско-Камском бассейне, в бассейне рек Дона, Днепра, Днестра и Северского Донца. Обнаружен и в бассейне Немана. Известны природные очаги без участия человека также в Казахстане.
Первый промежуточный хозяин кошачьего сосальщика — моллюск Bithynia leachi,
второй — карповые рыбы, в мышцах которых локализуются метацеркарии паразита. Окончательные хозяева — различные дикие и домашние рыбоядные млекопитающие и человек.
Opisthorchis viveirini —
возбудитель описторхоза виверры.
Отличается от предыдущего вида крупнодольчатостью семенников, мелкими размерами тела (до 10 мм) и ареалом расселения. Это типичный тропический гельминт, распространенный в Таиланде, Лаосе и Малайзии, где в некоторых зонах зараженность населения достигает 90%. Как и у предыдущего вида, хозяевами являются моллюски из р.Bithynia
и карповые рыбы. Окончательные хозяева в первую очередь хищные млекопитающие из сем. Виверровые, реже кошки, собаки и человек.
Clonorchis sinensis
(рис. 20.5,I
) —
возбудитель клонорхоза
Этот сосальщик крупнее двух предыдущих — до 25 мм длиной. Характерна форма семенников: они ветвисты и располагаются друг за другом в задней части тела. Яйца длиной до 30 мкм. Распространен в Юго-Восточной Азии, в странах Дальнего Востока. В России — на юге Приморского и Хабаровского краев. Первые промежуточные хозяева—моллюски родовBithynia
и Parafossutaris,
вторые — более 70 видов карповых рыб, реже бычковые и сельдевые. Окончательные хозяева — человек и рыбоядные млекопитающие.
Сосальщики, обитающие
в легких
К этой группе сосальщиков относятся несколько близких видов из р.Paragonimus.
Заболевание, которое они вызывают, называетсяnapa
гонимоз.
В связи с тем что эти паразиты поселяются в легких, в организме человека они осуществляют сложную миграцию из кишечника через брюшную полость, диафрагму и плевру. Это вызывает со стороны хозяина особенно тяжелую токсико-аллергическую реакцию. Сама локализация паразитов в легких приводит к очаговой пневмонии. Осложнения — пневмосклероз и легочные абсцессы. В связи со сложностью путей миграции этих паразитов часто встречается атипичная локализация. Особенно опасно попадание паразита в головной мозг. Это сопровождается симптомами менингита, энцефалита, эпилепсией и атрофией зрительного нерва. В типичных случаях паразиты попарно находятся в ткани легкого в капсуле, сообщающейся через бронх с окружающей средой. Это дает им возможность беспрепятственно выводить яйца с мокротой.
Другой характерной особенностью биологии сосальщиков этой группы является использование ими в качестве вторых промежуточных хозяев различных пресноводных ракообразных, в мышцах которых располагаются метацеркарии. Поэтому заражение человека паразитом возможно лишь в таких этнокультурных зонах, где население традиционно употребляет в пищу сырых ракообразных. Заражение человека за пределами этих регионов носит случайный, спорадический характер. Стойкость национальных традиций в питании, а также подчеркнуто природно-очаговый характер парагонимоза осложняют профилактику этого заболевания.
Легочный сосальщик
Paragonimus westermani
(рис. 20.5, Д
) —
наиболее часто встречающийся возбудитель парагонимоза.
Кроме него известно еще пять видов сосальщиков, чаще паразитирующих у животных, но поражающих также и человека. Основной вид распространен по всему тропическому поясу Старого и Нового Света, исключая Австралию, а за пределами тропиков — в Дальневосточном регионе, включая южные районы Приморского края и Приамурья. Остальные виды описаны на ограниченных ареалах — в Японии, Южном Китае, Западной Африке.
Легочные сосальщики имеют необычную для сосальщиков форму тела: они напоминают семя апельсина и имеют размеры до 12 мм. Яйца до 0,118 мм длиной.
Первый промежуточный хозяин — моллюски из родовSemisulcospira, Oncomelania
и некоторых других. Второй промежуточный хозяин — крабы из р.Eriocheir, Potamon,
раки родовCambarus, Procambarus,
а также креветки р.Macrobrachium.
Окончательные хозяева — человек и животные, питающиеся ракообразными, — выдры, норки, свиньи, кошки, собаки и некоторые грызуны.
Для диагностики
заболевания необходимо исследовать мокроту больных, в которой обнаруживаются яйца, а также фекалии, куда яйца могут попадать при проглатывании мокроты. В пищеварительном тракте они не изменяются.
Личная профилактика
парагонимоза заключается в отказе от поедания сырых ракообразных. Общественная профилактика
соответствует мерам, применяемым против сосальщиков, развивающихся в водной среде с двумя промежуточными хозяевами.
Сосальщики, цикл развития которых не связан с водной средой
Эта группа сосальщиков интересна своеобразием адаптации к среде обитания. Яйца паразитов должны попасть на почву или растения. Они содержат зрелых мирацидиев, которые из яиц не выходят до тех пор, пока они не будут проглочены наземными моллюсками — их первыми промежуточными хозяевами. Типичные подвижные церкарии этими сосальщиками не образуются. Они выделяются моллюсками в виде слизистых комочков на растения, где и поедаются вторыми промежуточными хозяевами — насекомыми. Насекомые, пораженные метацеркариями этих паразитов, становятся малоподвижными и могут оказаться съеденными окончательными хозяевами — травоядными животными и человеком. Как и большинство сосальщиков, эти паразиты в организме окончательного хозяина осуществляют миграцию.
Особенности цикла развития позволяют этим сосальщикам расселиться очень широко и заселить безводные зоны с сухим климатом. В связи с тем что насекомые весьма редко попадают в пищу к человеку, заболевания человека нечасты. Однако в Юго-Восточной Азии культурные традиции допускают питание насекомыми. Поэтому на территории Лаоса, Таиланда, Южного Китая и Японии заболевания, вызванные этими паразитами, встречаются чаше, чем в других зонах.
Ланцетовидный сосальщик
Dicrocoelium lanceatum
(рис. 20.5, Е
) —
возбудитель дикролоцелиоза —
паразит, внешне напоминающий кошачьего сосальщика размерами и формой, но семенники у него имеют более округлую форму и расположены на передней стороне тела. Яйца его несколько крупнее — до 45мкм.
Распространен повсеместно.
Первый промежуточный хозяин — моллюски р.Helicella
илиZebrina,
второй — муравей р.Formica.
Человек заражается случайно, проглатывая инвазированного муравья. Поселяется этот сосальщик в жёлчных ходах печени.
Диагностика —
как при всех трематодозах с поражением печени.
Личная профилактика —
необходимо следить, чтобы в пищу не попадали муравьи. Общественная профилактика —
дегельминтизация скота и санитарная охрана пастбищ.
Сосальщик поджелудочной железы
Eurytrema pancreaticum
(рис. 20.5, Ж
) —
возбудитель эуритрематоза.
Тело сильно расширено, длиной до 15мм. Характерны очень крупные присоски и матка, лежащая на заднем конце тела. Яйца до 50 мкм длиной. Паразит у животных встречается в Южной и Юго-Восточной Азии и в южных районах СНГ — в Казахстане, Киргизии, на Дальнем Востоке. .
Первый промежуточный хозяин паразита — моллюск из р.Sradybaena,
второй — луговой кузнечик р.Conocephalus
или сверчок р.Oecanthus.
Человек заражается, употребляя в пищу кузнечиков, не прошедших термической обработки, поэтому заражение человека встречается только в странах Юго-Восточной Азии. Сосальщик поселяется в протоках поджелудочной железы и вызывает симптомы хронического панкреатита.
Диагностика —
обнаружение яиц в фекалиях и дуоденальном содержимом при зондировании.
Профилактика
заражения человека — отказ от поедания насекомых или их предварительная термическая обработка. В остальном — см. предыдущий вид.
20.1.2. Класс Ленточные черви
Cestoidea
Как и сосальщики, все ленточные черви — паразиты, главным образом позвоночных животных. Класс насчитывает около 3500 видов. Форма тела этих червей лентовидная. У большинства видов тело, или стробила,
разделено на многочисленные членики — проглоттиды.
На переднем конце находится головка, или сколекс,
несущая органы прикрепления — присоски, крючья
или присасывательные щели — ботрии.
За головкой следует несегментированная шейка, от которой сзади постепенно отпочковываются молодые проглоттиды. В них системы органов не дифференцированы. В средней части стробилы лежат членики с развитой мужской и женской половыми системами. Они называются гермафродитными.
Последние проглоттиды стробилы содержат почти исключительно матку, заполненную яйцами, и рудименты остальных органов. Эти членики называются зрелыми.
В процессе роста червя задние, зрелые, членики постепенно отрываются, а от шейки образуются все новые, молодые проглоттиды.
Пищеварительная система у ленточных червей отсутствует в связи с длительной эволюцией в условиях паразитизма. Питание осуществляется всей поверхностью тела за счет пиноцитоза тегументом. Нервная система и органы выделения построены по плану, характерному для всего типа плоских червей. Половая система состоит из тех же органов, что и у сосальщиков, но представлена в каждом ленточном черве в огромном количестве копий, соответствующих количеству проглоттид (рис. 20.6).
Цикл развития ленточных червей отражает их более глубокие адаптации
к паразитизму по сравнению с сосальщиками. Об этом свидетельствует то, что свободноживущие расселительные стадии имеются только в одной группе этих червей — лентецов, считающихся наиболее древними по происхождению. Большинство ленточных червей попадают во внешнюю среду только в виде яйца, но размножаются лишь в организме хозяина. Некоторые наиболее специализированные паразиты способны обеспечивать аутоинвазию
хозяина с помощью яиц, даже не выходящих во внешнюю среду.
Рис. 20.6. Организация ленточных червей:
а—
головка; б—
гермафродитный членик; в—
зрелый членик; г—
стробила;
1—
мужские половые органы; 2—
женские половые органы; 3—
каналы выделительной системы; 4—
матка со зрелыми яйцами
Цикл развития наиболее древних ленточных червей связан с водной средой, самые примитивные ленточные черви и близкие им формы являются эктопаразитами наиболее древних позвоночных — хрящевых рыб, обитая у них на кожных покровах, жабрах, а также в глотке и переходя, таким червей: образом, к эндопаразитизму. Появление и быстрый расцвет более прогрессивных позвоночных – костных рыб, земноводных и т.д. — привел и ленточных червей к освоению новых сред обитания вместе с новыми хозяевами, в том числе с наземными позвоночньми. Современные ленточные черви в цикле развития имеют две стадии: половозрелую и личиночную. Половозрелая стадия паразитирует обычно в тонком кишечнике позвоночных. Личиночная стадия, или финна, —
тканевый паразит в организме промежуточных хозяев, в основном позвоночных, но иногда также членистоногих (рис. 20.7).
С фекалиями окончательного хозяина яйца паразитов попадают во внешнюю среду. Они содержат личинку — онкосферу,
которая будет развиваться уже в промежуточном хозяине при попадании яиц в его пищеварительную систему. Здесь онкосфера с помощью крючьев проникает через кишечную систему в кровеносное русло или лимфатические сосуды и мигрирует по организму, оседая в печени, легких, мышцах, центральной нервной системе и т. д. Разрастаясь, она превращается в финну.
Финна —
промежуточная форма паразита, обычно с полостью внутри и со сформировавшейся головкой. Финны некоторых ленточных червей способны размножаться бесполым путем за счет внутреннего или наружного почкования. Это увеличивает вероятность заражения основного хозяина и выживания потомства паразита. Окончательный хозяин, обычно хищное животное или человек, проглатывает финну вместе с тканями промежуточного хозяина. При этом головка паразита прикрепляется к стенке кишки хозяина и начинается рост шейки, образование члеников и развитие гельминта.
Рис. 20.7. Цикл развития ленточных червей:
1
— половозрелая стадия в кишечнике окончательного хозяина, 2 —
яйцо по внешней среде, 3 —
финнозная стадия в тканях промежуточного хозяина
В связи с тем что окончательный хозяин заражается этими гельминтами, поедая промежуточных хозяев, взаимоотношения хозяина и паразита на разных стадиях развития червей различны.
Если основной хозяин, в кишечнике которого находится гельминт, страдает от него относительно несильно, то жизнеспособность промежуточного хозяина с финнами этих червей в легких, мозгу или печени оказывается резко снижена. Это увеличивает вероятность именно инвазированных организмов быть съеденными окончательным хозяином.
По особенностям биологии ленточных червей, имеющих медицинское значение, можно разделить на группы, жизненный цикл которых связан и не связан с водной средой. Вторая группа подразделяется на гельминтов: а) использующих человека как окончательного хозяина, б) обитающих в человеке как в промежуточном хозяине, в) проходящих в человеке весь жизненный цикл.
Соответственно этому пути заражения человека патогенное действие паразитов, диагностика, лечение и профилактика соответствующих заболеваний различны.
Болезни, вызываемые ленточными червями, называют цестодозами.
Многие виды ленточных червей поражают только человека, другие встречаются также и в природной обстановке, для них характерно существование классических природных очагов. Цестод, обитающих в человеке как основном хозяине, сближает то, что они живут в кишечнике и всегда в небольшом количестве. Это объясняется выраженной внутривидовой конкуренцией, в которой выживают лишь единичные особи. На интенсивности размножения это не отражается, так как плодовитость их огромна.
К ним относится несколько видов паразитов из р.Diphyllobothrium,
являющихся возбудителями дифиллоботриозов.
Гельминты этой экологической группы имеют наиболее архаичный вариант цикла развития:
у них сохранились активно плавающая личиночная стадия — корацидий —
и два промежуточных хозяина, обитающие в водной среде, — мелкие планктонные ракообразные из родовCyclops
иDiaptomus,
a также рыбы, питающиеся ими. В рачках обитает личинка, называемая процеркоид,
в рыбе — плероцеркоид.
Примитивной чертой плероцеркоида является его способность к активным перемещениям, причем в случае поедания крупной рыбой более мелких рыб, инвазированных плероцеркоидами, последние пробуравливают стенку кишки и выходят в брюшную полость и мышцы. Таким образом, крупные рыбы на протяжении всей жизни могут накопить в себе сотни плероцеркоидов. Хозяин заражается, поедая инвазированную рыбу. Поэтому основа личной профилактики — термическая обработка рыбных продуктов.
Широкий лентец
Diphyllobothrium latum
(рис. 20.8, А
) —
наиболее частый возбудитель дифиллоботриоза у человека. Стробила имеет длину около 10 м. Головка снабжена ботриями —
присасывательными щелями, зрелые членики характеризуются маткой розетковидной формы небольших размеров. Матка имеет связь с внешней средой, поэтому созревающие яйца свободно выводятся из нее. Яйца желтоватого цвета, длиной до 71 мкм, имеют на одном полюсе крышечку, а на другом — маленький бугорок. Паразит распространен широко в зоне с умеренным климатом. Заболевание встречается по берегам крупных рек и озер.

Рис. 20.8. Крупные ленточные черви. А —
широкий лентец; Б —
бычий цепень; В —
свиной цепень: а —
головки, б —
зрелые членики
Жизненный цикл
широкого лентеца типичен для этой группы паразитов. Яйца попадают в воду, и из них выходит корацидий, покрытый ресничками. Его проглатывают первые промежуточные хозяева — циклопы. Циклопов с процеркоидами съедают рыбы, в мышцах и икре которых накапливаются плероцеркоиды. Из рыб чаще всего поражаются окуни, ерши, налимы и щуки. В крупных хищных происходит дальнейшее накопление плероцеркоидов. Окончательными хозяевами являются крупные рыбоядные млекопитающие (медведи, собаки и т. д.) и человек. Таким образом, дифиллоботриоз — природно-очаговое заболевание. У человека это заболевание встречается чаще у рыбаков, туристов и людей, употребляющих в пищу слабопросоленную рыбу и икру домашнего производства.
Дифиллоботриоз — это опасное заболевание не только потому, что паразит имеет большие размеры тела и может вызвать кишечную непроходимость, но и в связи с тем, что он находится в антагонистических отношениях с нормальной микрофлорой кишечника, вызывая дисбактериоз. При этом нарушается всасывание витаминов группы В и развивается тяжелая анемия с нарушением кроветворения в красном костном мозге.
Диагностика
заболевания основана на обнаружении яиц и обрывков зрелых члеников паразита в фекалиях.
Личная профилактика
заражения — см. выше. Общественная профилактика —
охрана водоемов от фекального загрязнения.
Кроме этого паразита известны несколько других видов р.Diphyllobothrium,
имеющих меньшее значение в связи с малыми ареалами распространения (в Прибайкалье, на территории части Эвенкии, на Сахалине и т. д.) и с тем, что человек ими поражается редко, так как их окончательные хозяева обычно рыбоядные птицы и мелкие млекопитающие. Цикл развития их, методы профилактики и диагностики заболеваний не отличаются от данных, приведенных для широкого лентеца.
Эти гельминты — более специализированные паразиты по сравнению с предыдущей группой. Личинки их ни на какой из стадий развития не способны к перемещению. Матка в зрелых члениках не имеет связи с окружающей средой, поэтому яйца выводятся во внешнюю среду не поодиночке, а внутри зрелых члеников.
Ленточные черви, использующие человека
в качестве окончательного хозяина
Эта группа паразитов обитает у человека на половозрелой стадии только в тонком кишечнике. Большинство из них имеют крупные размеры и мощные органы прикрепления. Патогенное действие выражается в «эффекте отнятия пищи», в интоксикации продуктами жизнедеятельности, в подавлении размножения кишечных бактерий — дисбактериозе — и в нарушении всасывания витаминов в кишечнике. Кроме того, постоянное механическое раздражение кишечной стенки может приводить к антиперистальтическим движениям кишки, к завороту кишок и к другим формам кишечной непроходимости. У больных отмечается похудание, ухудшение или извращение аппетита и нарушение деятельности кишечника.
Диагноз
кишечных цестодозов ставят при обнаружении в фекалиях зрелых члеников паразитов. Яйца видов этой группы очень похожи, и постановка точного диагноза при обнаружении яиц невозможна.
Личная профилактика —
термическая обработка мяса. Общественная профилактика —
санитарный контроль мясопродуктов и санитарно-просветительная работа с населением.
Бычий цепень
Taeniarrhynchus saginatus
(рис. 20.8, Б
) —
возбудитель тениаринхоза,
достигает в длину 4—10 м. На головке имеет только четыре присоски. Гермафродитные членики квадратной формы, матка в них не разветвляется, а яичник состоит из двух долей. Зрелые членики сильно вытянуты. Матка очень разветвлена, число ее боковых ветвей достигает 17—34 пар. Яйца содержат онкосферы, расположенные под тонкой прозрачной оболочкой, которая быстро разрушается. Онкосферы имеют три пары крючьев и толстую, радиально исчерченную оболочку. Диаметр онкосфер около 10 мкм. Тениаринхоз распространен повсеместно, где население употребляет в пищу сырое или недостаточно обработанное говяжье мясо.
Жизненный цикл
бычьего цепня типичен. Основной хозяин только человек, промежуточный — крупный рогатый скот. Особенностью этого вида является то, что с фекалиями человека членики выделяются группами по 5—6. Корова, проглотив такие членики, становится промежуточным хозяином паразита. В ее мышцах формируются финны, называемые цистицерками.
Финна представляет собой пузырек, заполненный жидкостью, в котором находится сколекс. В мышцах финны могут сохранять жизнеспособность долгие годы. При поедании мяса такой коровы в желудке под действием кислой среды желудочного сока головка вывертывается, прикрепляется к стенке кишки и развивается новый цепень.
Диагностика
проводится несложно — при обнаружении зрелых члеников в фекалиях, так как членики имеют характерное строение.
Профилактика
тениаринхоза заключается в охране пастбищ от заражения фекалиями человека.
Свиной цепень
Taenia solium
(рис. 20.8, В
) —
возбудитель тениоза
и цистицеркоза.
Этот паразит меньше предыдущего, он достигает в длину 3 м. На головке кроме присосок у него находится венчик из 22—32 крючьев. В гермафродитных члениках не две, а три дольки яичника; матка в зрелых члениках имеет не более 12 пар боковых ответвлений. Яйца не отличаются от яиц предыдущего вида.
Жизненный цикл
свиного цепня типичен. Окончательный хозяин паразита — человек. Характерной особенностью является способность члеников активно выползать из заднепроходного отверстия поодиночке. При подсыхании оболочка их лопается и яйца могут свободно рассеиваться во внешней среде. Этому процессу могут способствовать птицы и мухи. Из яиц, проглоченных промежуточным хозяином — свиньей, развивается онкосфера и позже цистицерки, как и у предыдущего вида.
Промежуточными хозяевами этого гельминта кроме домашних и диких свиней могут быть кошки, собаки и человек: В этом случае у них, так же как и у свиней, развивается цистицеркоз. Человек может проглотить яйца свиного цепня случайно, но более часто цистицеркоз возникает как осложнение тениоза. При этом заболевании особенно часто возникает обратная перистальтика кишечника и рвота. Зрелые членики могут таким образом попасть в желудок, перевариться там, а освободившиеся онкосферы проникают в сосуды кишечника, разносятся кровью и лимфой по организму, где в печени, мышцах, легких, мозге и других органах формируются цистицерки. Это может привести к быстрому смертельному исходу.
Лабораторная диагностика
тениоза основана на обнаружении характерных зрелых члеников в фекалиях; диагностика цистицеркоза сложнее — путем рентгенологического обследования и постановки иммунологических реакций.
Для личной профилактики
тениоза необходимо термически обрабатывать свинину, а цистицеркоза — соблюдать правила личной гигиены. Общественная профилактика —
закрытое содержание свиней.
К этой экологической группе относится еще несколько видов ленточных червей, поражающих человека обычно случайно, для которых окончательным хозяином являются синантропные животные — крысы, мыши, собаки, а промежуточными — насекомые и клещи, обитающие в жилище человека, — тараканы, блохи и вредители продуктов питания. Заражение человека происходит при случайном проглатывании этих насекомых. Чаще заболеваниями, вызываемыми этими паразитами, заражаются дети, более активно, чем взрослые, контактирующие с домашними животными и не имеющие стабильных навыков личной профилактики и гигиены. Сведения об этих паразитах приведены в табл. 20.1.
Таблица 20.1. Ленточные черви, случайно использующие человека
как окончательного хозяина
| Вид паразита, название заболевания |
Промежуточный хозяин |
Окончательный хозяин |
Пути заражения человека |
Диагностика |
Профилактика |
Hymenolepis diminuta
Гименолепи-
доз диминут-ный
Dipilidium
саптит
Дипилидиоз
Inermicap-sifera sp.
Инермика-псифероз
Bertiella sp.
Бертиеллез
|
Блохи, тараканы, мучной хрущ, мучная огневка
Блохи и вла-соеды собак и
кошек
Клещи-паразиты кроликов и крыс
Клещи-паразиты обезьян
|
Грызуны, обезьяны, собаки
Собаки и кошки
Грызуны, кролики
Обезьяны
|
Проглаты-вание насекомых
Проглаты-вание блох и власо-едов
Проглаты-вание клещей
То же
|
Обнаруже-ние яиц в фекалиях
Обнаруже-ние под-
вижных чле-
ников в
фекалиях
Обнаруже-ние яиц в фекалиях
То же
|
Борьба с грызунами и бытовыми
насекомыми
Дегельминти-зация домаш-них животных
Не изучена
То же
|
Кроме перечисленных в таблице видов гельминтов использовать человека как окончательного хозяина могут еще несколько реже встречающихся видов. Все они представляют интерес как формы, расширяющие в процессе эволюции спектр окончательных хозяев, и как потенциальные паразиты человека в будущем могут иметь большее медицинское значение.
Ленточные черви,
использующие человека в качестве промежуточного хозяина
Гельминты этой группы обитают у человека и травоядных млекопитающих на стадии финны, поражая ткани внутренней среды: мышцы, кости, печень, почки, головной и спинной мозг и т. д. Часто они обнаруживаются также в легких. Финны этих паразитов могут жить в организме хозяина очень долго. При этом они постоянно растут и даже способны к бесполому размножению путем почкования. Финны резко снижают жизнеспособность инвазированных ими организмов, поэтому ларвольные
цестодозы, вызываемые паразитами этой группы, гораздо более тяжелы и опасны, чем все остальные. В патогенном действии паразита на хозяина имеют значение его локализация, скорость роста и выделения токсических продуктов диссимиляции.
Лабораторная диагностика
ларвальных цестодозов осложнена тем, что финны не имеют связи с окружающей средой и кроме продуктов диссимиляции ничего не выделяют. Диагноз ставят на основании рентгенологических, биохимических и иммунологических исследований.
Основные меры профилактики
направлены на предотвращение попадания в организм человека инвазионных стадий паразитов. Однако даже самая тщательная профилактика заболеваний этой группы не может гарантировать полного уничтожения паразитов, так как все ларвальные цестодозы — зоонозные природно-очаговые болезни.
Эхинококк
Echinococcus granulosus
(рис. 20.9, А
) —
возбудитель эхинококкоза.
Половозрелая форма имеет головку с крючьями и 3—4 членика разной степени зрелости. Последний из них зрелый, он содержит около 800 яиц. Общая длина тела до 5 мм. Яйца по форме и размерам сходны с яйцами свиного и бычьего цепней. Эхинококкоз у человека распространен во всех географических и климатических зонах, преимущественно в регионах с развитым отгонным животноводством.
Жизненный цикл
эхинококка связан с хищными животными семейства Псовые (волками, шакалами, собаками), которые являются его окончательными хозяевами. Взрослые членики способны активно ползать, распространяя яйца по шерсти хозяина и в окружающей среде. Их могут проглотить травоядные животные — коровы, овцы, олени или человек, становясь промежуточными хозяевами. Финна эхинококка — пузырь, нередко достигающий 20 см в диаметре. Он заполнен жидкостью с огромным количеством молодых сколексов, постоянно почкующихся от внутренней поверхности стенки финны. Окончательный хозяин заражается, поедая пораженные органы промежуточного.
Растущая финна сдавливает органы, вызывает их атрофию. Постоянное поступление продуктов диссимиляции в организм хозяина вызывает его истощение. Очень опасен разрыв эхинококкового пузыря: жидкость, заключенная в нем, может вызвать токсический шок. При этом мелкие зародышевые сколексы могут распространяться по организму, поражая другие органы. Множественный эхинококкоз обычно заканчивается смертью хозяина.
Рис. 20.9. Ленточные черви, использующие человека как промежуточного хозяина. А —
эхинококк; Б —
альвеококк: а
— половозрелые стадии,
б —
финны
Личная профилактика
заражения — мытье рук после контактов с пастушьими собаками. Общественная профилактика —
обследование и дегельминтизация собак, недопущение скармливания им органов больных животных.
Альвеококк
Alveococcus multilocularis
(рис. 20.9, Б
) —
возбудитель альвеококкоза.
Половозрелая форма отличается от эхинококка меньшими размерами тела (до 2 мм) и деталями строения крючьев и матки. Финнозная стадия состоит из множества мелких пузырьков, постоянно почкующихся друг от друга наружу, благодаря чему финна представляет собой постоянно растущий узел. В каждом пузырьке находится зародышевая головка паразита. Рост финны разрушает окружающие ткани наподобие злокачественной опухоли. Возможен и отрыв отдельных пузырьков с распространением патологического процесса по организму. Природные очаги альвеококкоза имеются в Сибири, Средней Азии, на Урале и Дальнем Востоке, а также в Северной Америке, в Центральной и Южной Европе.
Жизненный цикл
принципиально не отличается от эхинококка, но окончательными хозяевами этого паразита являются дикие хищники — лисы, песцы, волки, реже домашние — собаки, промежуточными — мышевидные грызуны, иногда человек.
Альвеококкоз — более тяжелое заболевание, чем эхинококкоз, в связи с инвазивным характером роста финны.
Личная профилактика —
как при эхинококкозе, общественная —
соблюдение правил гигиены при обработке шкур промысловых животных, а также запрещение скармливания собакам тушек грызунов.
Spiromeira erinacei
(рис. 20.10, А
) —
возбудитель спарганоза. Половозрелый паразит обитает в кишечнике кошачьих и псовых. Первыми промежуточными хозяевами являются циклопы, вторыми — земноводные, пресмыкающиеся, некоторые птицы и даже человек. При проматывании воды с циклопами происходит заражение. Плероцеркоид поселяется в соединительной ткани, под кожей, под конъюнктивой глаза и даже в мозге. Размеры его от 0,5 до 30 см. Заражение людей возможно при поедании мяса лягушек и змей. В таком случае человек также остается вторым промежуточным хозяином, так как в его теле продолжает паразитировать плероцеркоид.
Описанный вид гельминта расселен в Восточной и Юго-Восточной Азии, в Австралии, Америке и Африке.
Диагностика
осложнена. Обычно диагноз ставится во время хирургического вмешательства.
Профилактика —
фильтрование воды, употребляемой для питья, и термическая обработка экзотических продуктов питания — мяса лягушеки змей.
Рис. 20.10. Возбудители спарганоза.
А —
Spiromeira erinacei;
Б — Sparganum
proliferum
Sparganum protiferum
(рис. 20.10, Б
) —
плероцеркоид неизвестного вида ленточного червя, вероятно, также из р.Spiromeira.
Он характеризуется оригинальной особенностью — способностью почковаться наподобие альвеококка, но образует не более нескольких дочерних особей, морфологически связанных с материнской. В связи с этим и получил названиеproliferum —
разрастающийся. Размеры его совпадают с размерами предыдущего вида. Жизненный цикл этого вида изучен слабо, вероятно, он соответствует развитию предыдущего вида; гельминт чаще всего встречается в Корее, Вьетнаме и Японии. Кроме способов заражения, характерных для предыдущего вида, Sparganum
может проникать в организм человека через изъязвленные кожные покровы и слизистые оболочки при прикладывании к ним в качестве народного восточного лечебного средства мяса змей и лягушек, содержащего плероцеркоиды.
Два последних описанных вида встречаются редко и не представляют серьезной медицинской проблемы, но иллюстрируют необычность данной формы паразитизма и экзотичность способов заражения. Это отражает сложность и разнонаправленность эволюции паразитизма как экологического феномена.
Циклы развития, проходящие полностью в одном хозяине, встречаются не часто среди многоклеточных паразитов. Способность гельминта размножаться в хозяине длительное время, не покидая его на стадии яйца, — явление крайне редкое. Таким необычным паразитом человека стал карликовый цепеньHymenolepis nana
—
возбудитель гименолепидоза
(рис. 20.11). Это мелкий гельминт длиной до 5 см с головкой, снабженной крючьями и присосками. Зрелые членики столь нежны, что, развиваясь, полностью разрушаются еще в кишечнике, поэтому яйца быстро выходят из них и попадают в фекалии. Размер яиц до 40 мкм. Они имеют округлую форму и бесцветны.

Рис. 20.11. Карликовый цепень
Гименолепидоз встречается повсеместно, особенно в странах с сухим и жарким климатом. Поражаются преимущественно дети.
Жизненный цикл
этого паразита за время длительной адаптации к организму человека претерпел существенные изменения. Если его яйца, выделившись из организма с фекалиями, попадают в пищеварительную систему мучного хруща из р.Tenebrio,
в нем разовьется финнозная стадия червя — цистицеркоид.
При проглатывании инвазированного жука с непропеченным тестом в кишечнике человека из цистицеркоидов разовьются взрослые паразиты. В современных условиях такое явление происходит крайне редко. Если человек проглотит яйца карликового цепня при несоблюдении правил личной гигиены, то в ворсинках тонкого кишечника из них развиваются цистицеркоиды, которые позже, разрушая ворсинки, выпадают в просвет кишечника и превращаются в половозрелых гельминтов. Яйца этого паразита могут не выделяться во внешнюю среду вовсе, быстро достигая зрелости еще в кишечнике. В таком случае из них развиваются цистицеркоиды, как и в предыдущем случае, а затем и взрослые цепни.
Таким образом, у одного и того же вида сосуществуют три варианта развития: типичный для ленточных червей со сменой двух хозяев; с утратой промежуточного хозяина; вообще без выхода во внешнюю среду даже на стадии яйца. Эти три варианта соответствуют трем этапам эволюции биологии карликового цепня в процессе адаптации его к паразитированию в таком специфическом хозяине, как человек. Способность этого паразита к аутоинвазии хозяина даже без выхода во внешнюю среду обусловливает длительность заболевания на фоне смены поколений паразитов и сложность полного излечения в связи с тем, что в организме человека постоянно сосуществуют как личиночные формы в ворсинках, так и половозрелые — в просвете кишки. Однако в связи с тесным контактом цистицеркоидов карликового цепня с тканями хозяина у человека возникает иммунитет против них. Он затрудняет развитие следующих поколений паразитов, обеспечивая отторжение и гибель все большего количества цистицеркоидов, формирующихся в процессе аутоинвазии.
Таким образом, несмотря на аутоинвазию, гименолепидоз не может продолжаться бесконечно. После смены нескольких поколений паразитов наступает самоизлечение. Гименолепидоз сопровождается чередующимися поносами, болями в области живота и истощением. Часто при этом наблюдаются бессонница и эпилептоидные припадки. Для диагностики
важно обнаружение яиц в фекалиях. Профилактика
сводится к выявлению и лечению больных и соблюдению правил личной гигиены, в первую очередь в детских учреждениях. Имеются сведения, что у мышей и крыс также способны паразитировать некоторые штаммы этого паразита. В связи с этим профилактика гименолепидоза должна включать также борьбу с грызунами. Необходима и постоянная борьба с насекомыми — механическими переносчиками яиц гельминтов.
Форма тела этих организмов удлиненно-веретенообразная или нитевидная. Кожно-мускульный мешок состоит из многослойной, плотной, эластичной и нерастяжимой кутикулы, гиподермы, представляющей собой единую цитоплазматическую массу, не разделенную на отдельные клетки и содержащую большое количество ядер, и одного слоя продольных гладких мышц. Кутикула выполняет в основном защитную функцию. Мышцы располагаются в виде двух продольных тяжей — на спинной и брюшной сторонах тела. Их поочередное сокращение обеспечивает энергичные сгибательные и разгибательные движения и быстрое перемещение тела в пространстве.
Пищеварительная система — в виде сквозной трубки с ротовым и анальным отверстиями. Нервная система представлена продольными тяжами, соединенными кольцевидными перемычками. Выделительная система в основе имеет протонефридиальное строение, но количество выделительных клеток исчисляется единицами. Круглые черви раздельнополы. Половая система построена в виде дифференцированных по длине трубок, часть которых функционирует как яичники или семенники, часть — как семяпроводы или яйцеводы, а часть — как органы, в которых накапливаются и сохраняются зрелые половые продукты. Все внутренние органы расположены в первичной полости тела, заполненной жидкостью, которая придает всему организму упругость и обеспечивает обмен веществ между органами (рис. 20.12).
Рис. 20.12. Организация круглых червей. А—
самка; Б—
самец:
1
— нервная система, 2
— матка, 3—
кишечник, 4 —
яичники, 5 —
семенник
Своеобразной особенностью круглых червей является то, что в состав их тела входит всегда определенное количество клеток. Это ограничивает их способность к росту и регенерации. Медицинское значение имеют представители только класса Собственно круглые черви.
20.2.1. Класс Собственно круглые черви
Nematoda
Строение представителей этого класса в основном соответствует характеристике типа в целом. В связи с тем что свободноживущие круглые черви имеют обычно небольшие размеры и обитают в почве иногда на довольно большой глубине или в донном слое ила водоемов, где условия близки к анаэробным, многие из них способны жить без кислорода, обходясь гликолизом в процессе энергетического обмена. Кутикула, выполняющая защитные функции, также сформировалась еще у свободноживущих почвенных и донных видов нематод. Поэтому серьезной дегенерации при переходе к паразитизму они не претерпели. Адаптациями к обитанию в хозяине являются развитая половая система и у некоторых форм — органы фиксации, а также усложнение циклов развития с участием в ряде случаев промежуточных хозяев.
Круглых,червей, сохранивших связь с внешней средой, яйца или личинки которых развиваются в почве, называют геогельминтами.
Более специализированных паразитов, развивающихся с участием промежуточных хозяев, называют биогельминтами.
С медицинской точки зрения, следует выделить еще одну группу нематод, неспецифических паразитов человека, которые осуществляют лишь миграцию в его организме, не достигая половой зрелости. Болезни, вызываемые круглыми червями, называют нематодозами.
У этих червей яйца или личинки обязательно развиваются в поверхностных слоях почвы при доступе кислорода и достаточной влажности. За исключением острицы, все гельминты встречаются чаще в регионах с жарким и влажным климатом, который предоставляет личинкам и яйцам больше возможностей развиваться в почве. В Арктике и южных приполярных областях геогельминты не встречаются. Самцы и самки гельминтов легко различимы: самцы большинства видов имеют загнутый на брюшную сторону или спирально закрученный задний конец тела, в то время как у самок он прямой.
Геогельминты обитают в просвете кишки и размножаются яйцами, которые выводятся с фекалиями и развиваются далее в почве. Они либо сами через определенное время становятся инвазионными, либо из них развиваются личинки, ведущие некоторое время свободный образ жизни и позже становящиеся инвазионными. Геогельминты, поражающие человека, не могут паразитировать у животных. Соответственно этому нематодозы, вызываемые этими паразитами, являются антропонозными болезнями.
Заражение большей частью геогельминтов осуществляется при проглатывании яиц или личинок с продуктами, загрязненными почвой.
Для диагностики
всех нематодозов этой группы важно обнаружение яиц в фекалиях больного.
Профилактические меры
направлены на предотвращение попадания инвазионных яиц в пищеварительную систему—личная гигиена и гигиена питания, реже — другие меры. Часть геогельминтов, попадая в пищеварительную систему человека, быстро достигают половой зрелости и начинают размножаться в кишечнике, не мигрируя по организму хозяина. Личинки других перед достижением половой зрелости обязательно передвигаются по кровеносным сосудам и дыхательной системе и только после этого развиваются в кишечнике.
Геогельминты, развивающиеся без миграции
Эта группа червей представляет собой наиболее специализированных геогельминтов, яйца которых еще требуют присутствия кислорода, а развивающиеся личинки — уже нет. Отсутствие миграции по сосудам откладывает отпечаток на клинику соответствующих заболеваний. Во-первых, они протекают обычно достаточно доброкачественно, во-вторых, первые проявления их возникают только по достижении паразитами зрелости и начала откладки яиц.
Власоглав
Trichocephalus trichiurus
(рис. 20.13, А
) —
возбудитель трихоцефалеза.
Передний конец тела этого червя вытянут. В нем находится только пищевод. Задняя часть тела утолщена, здесь располагаются кишка и половая система. Общая длина паразита 3—5 см. Яйца светлые овальные, прозрачные, с двумя характерными пробочками, длиной до 50 мм.

Рис. 20.13. Геогельминты, развивающиеся без миграции.
А —
власоглав; Б —
острица
Власоглав поселяется в слепой и восходящей части толстой кишки, прикрепляясь к стенке передним концом тела и питаясь кровью и тканевой жидкостью из глубоких слоев слизистой оболочки. Яйца, выводимые с фекалиями, в почве достигают инвазионности за 3—4 недели. Заражение человека осуществляется при проглатывании инвазионных яиц. Половой зрелости паразиты достигают в кишке уже через несколько недель. Патогенное действие власоглава связано с интоксикацией хозяина продуктами жизнедеятельности и с нарушением функции кишечника. При массивной инвазии, возможны местные воспалительные явления, а также головные боли, головокружение и даже судороги.
В связи с тем что власоглав не питается содержимым кишечника и плотно прикреплен к его стенке, выведение этого паразита из организма человека более сложно по сравнению с многими другими геогельминтами.
Острица детская
Enterobiusvermicularis
(рис.
20.13, Б
) —
возбудитель энтеробиоза —
самого распространенного гельминтоза, который встречается в основном у детей. Мелкий червь с выраженным половым диморфизмом: самка достигает в длину 12 мм, тело ее прямое, сзади заостренное. Самцы мельче (до 5 мм) и свернуты спирально. Яйца овальны, несколько асимметричны, прозрачны и бесцветны, длиной до 50 мкм.
Жизненный цикл в целом соответствует циклу развития власоглава, но яйца достигают инвазионности после откладывания не за несколько недель, а всего за несколько часов. Поэтому при энтеробиозе часто происходит аутоинвазия.
Обитают острицы в нижнем отделе тонкой кишки, питаясь ее содержимым. Самки со зрелыми яйцами выползают из анального отверстия и откладывают яйца на коже промежности, вызывая при этом сильный зуд. При расчесывании зудящих мест яйца попадают на руки, затем на белье и игрушки. При проглатывании яиц из них быстро развиваются взрослые паразиты, продолжительность жизни которых около 1 мес. Патогенное действие остриц связано с тем, что зуд промежности приводит к нарушению сна, к нервному истощению. Возможно также нарушение стенки кишечника.
Лабораторная диагностика
имеет особенности: в фекалиях яйца обнаруживаются редко, поэтому их исследуют в мазке с перианальных складок промежности.
Личная профилактика —
соблюдение правил личной гигиены, особенно тщательное мытье рук после сна, короткая стрижка ногтей. Общественная —
регулярно обследовать детей в детских учреждениях, лечение больных, регулярные обследования также работников детских учреждений и предприятий общественного питания.
Геогельминты, развивающиеся с миграцией
Особенностью этой группы геогельминтов является то, что, попав в организм человека любым путем, они на стадии личинки проникают сначала в вены большого круга кровообращения, потом попадают в сердце и оттуда в малый круг. В капиллярах легких они выходят из кровеносного русла в альвеолы, активно поднимаются по бронхам, трахее, гортани. Эти перемещения вызывают кашлевый рефлекс, что способствует их выходу в глотку и вторичному проглатыванию. Только после этого, попадая в кишечник, личинки достигают половой зрелости.
Фазу миграции можно расценить как рекапитуляцию свободного образа жизни предковых форм. Она имеет большое медицинское значение в связи с тем, что в этот момент наступает особенно сильная интоксикация и аллергизация организма хозяина. Поэтому на начальных этапах инвазии паразитами этой группы у человека возможны аллергические реакции, кровоизлияния в легких и пневмонии. Позже возникают симптомы, связанные с локализацией паразитов в тонком кишечнике.
Лабораторная диагностика —
обнаружение яиц в фекалиях. Профилактика
зависит от вида паразита.

Рис. 20.14. Аскарида человеческая
Аскарида человеческая
Ascaris lumbricoides
(рис. 20.14) — возбудитель аскаридоза —
крупный гельминт, самки которого достигают 40 см длины, а самцы — 20 см. Зрелые яйца овальны и бугристы, оболочка их толстая и многослойная. Цвет желтовато-коричневый, длина до 60 мкм. Этот вид очень близок к свиной аскариде, которая в Юго-Восточной Азии может легко заражать человека и, наоборот, человеческая — свиней. Это свидетельствует об их относительно недавнем происхождении от одного предка и о времени начала дивергенции — не ранее начала одомашнивания свиней, которое происходило в Юго-Восточной Азии.
Жизненный цикл
типичен для гельминтов этой группы. Яйца созревают при высокой влажности почвы и температуре 18—25°С за 2—3 недели. Инвазия большим количеством аскарид может привести к закупорке кишечника или общего жёлчного протока. Известны случаи атипичной локализации аскарид: в гортани, среднем ухе, печени и даже сердце. При этом необходимо срочное хирургическое вмешательство.
Кривоголовка двенадцатиперстной кишки
Ankylostoma duodenale
и некатор
Necator americanus
(рис. 20.15)—оба вида относятся к сем. Анкилостомиды. Отличаются друг от друга формами ротовой капсулы и особенностями географического распространения. Кривоголовка имеет в ротовой полости четыре хитиновых зуба, а некатор — две широкие режущие пластинки полулунной формы. Оба паразита широко расселены по всему тропическому и субтропическому поясу, однако некатор чаще встречается в зонах с более жарким климатом. В остальном оба вида сходны настолько, что даже заболевания, вызываемые ими, не дифференцируются и называются анкилостомидозами.
Размеры анкилостомид около 10 мм. Головной конец загнут на брюшную сторону; яйца овальные, прозрачные, длиной до 60 мкм.
Жизненный цикл
анкилостомид своеобразен и свидетельствует о тесной связи их со свободноживущими предковыми формами. Яйца, попадающие в почву, быстро развиваются, и из них вскоре выходят личинки, которые, дважды линяя, через несколько месяцев становятся инвазионными и могут попадать в организм человека либо с загрязненными почвой овощами и фруктами, либо за счет активного внедрения через кожу.
Рис. 20.15. Анкилостомиды. А —
общий вид; Б—
ротовой аппарат:
а —
Кривоголовка, 6 —
некатор
Эта особенность обеспечивает возможность существования подземных очагов анкилостомидозов в шахтах, находящихся в зонах умеренного пояса. Основным условием возникновения таких очагов является высокая влажность грунта и загрязненность его фекалиями. Попав в кровь, личинки проделывают по сосудам путь, характерный для всех видов гельминтов этой группы. Окончательная локализация — двенадцатиперстная кишка, к ворсинкам которой они прикрепляются ротовыми капсулами, повреждая их и питаясь кровью и клетками слизистой оболочки. Оба паразита выделяют в ранки антикоагулянтные вещества и могут вызвать кишечные кровотечения и аллергизацию больных.
Лабораторная диагностика —
как при всех геогельминтозах.
Профилактика —
кроме общих мер, описанных выше, обязательное ношение обуви в районах, где распространены эти заболевания.
Угрица кишечная
Strongyloides stercoralis
—
возбудитель стронгилоидоза.
Это очень мелкий гельминт, до 2,2 мм длиной. Из всех известных паразитов человека — наименее специализированный.
Жизненный цикл
этого червя характеризуется значительной вариабельностью. Он может проходить целиком во внешней среде либо частично или полностью в организме хозяина. Чаще встречается чередование свободноживущего и паразитического поколений.
Паразитические половозрелые угрицы живут в кишечнике человека. Из яиц уже в кишечнике выходят личинки, судьба которых может быть двоякой. В первом случае после линьки в почве они могут стать инвазионными и проникают в организм человека либо через кожу, либо перорально с фруктами и овощами. Дальнейшее развитие в этом случае проходит так же, как у анкилостомид. Во втором случае личинки могут достичь половозрелости в почве.
Взрослые самки откладывают яйца и дают начало свободноживущим поколениям. При изменении условий жизни личинки могут стать инвазионными для человека и вновь перейти к паразитическому существованию. Присутствие паразитов в кишечнике вызывает чередующиеся поносы и запоры. Часто при этом паразиты заселяют и жёлчные ходы печени, вызывая желтуху.
Аллергизация организма при стронгилоидозе очень сильна, особенно на миграционной стадии развития.
Описаны случаи течения стронгилоидоза в течение 25 лет. Это объясняется не только длительностью жизни этого паразита,но ивозможностью аутоинвазии.
Как и анкилостомиды, угрица кишечная встречается в основном в тропической и субтропической зонах. Особенно широко расселена в Юго-Восточной Азии, Южной и Центральной Америке. Возможно существование подземных очагов в зонах с умеренным климатом, как и при анкилостомидозах.
При диагностике
надежным признаком заболевания является обнаружение живых личинок в фекалиях, рвотных массах и при дуоденальном зондировании.
Профилактика
стронгилоидоза соответствует профилактике анкилостомидозов.
Все нематоды этой группы, поражающие человека, живородящи и
большую часть цикла развития проводят у человека в тканях внутренней среды. Промежуточные хозяева их очень разнообразны — от циклопов и насекомых до медведей и человека. Окончательными хозяевами могут быть различные дикие и домашние животные, поэтому заболевания, которые вызывают эти паразиты, относятся к разряду природно-очаговых. Для попадания в места окончательной локализации биогельминты осуществляют миграцию по лимфатическим и кровеносным сосудам. Кроме того, они особенно активно взаимодействуют с иммунной системой хозяина. Поэтому в клинической картине нематодозов-биогельминтозов ведущими симптомами являются ток-сико-аллергические реакции.
Важно также механическое и местное токсическое действие. Диагноз биогельминтозов часто затруднен. Приходится прибегать к методам биопсии и иммунологическим реакциям. Профилактика зависит от путей заражения, которые бывают различны.
Биогельминты, заражение которыми происходит
при проглатывании личинок с тканями промежуточного хозяина
Для этих гельминтов характерна первичная локализация в кишечнике человека, а затем проникновение в кровь через его стенку. Профилактика
в связи с этим — создание препятствий для проникновения личинкой через рот.
Ришта
Dracunculus medinensis
(рис. 20.16) — возбудитель дракункулеза.
Длина самки до 120 см, самца — только 2 см. Заболевание распространено в зонах с тропическим и субтропическим климатом, раньше встречалось в Средней Азии.

Рис. 20.16. Ришта
Жизненный цикл
ришты связан с водной средой. Окончательные хозяева ришты — человек, обезьяны, домашние и дикие млекопитающие, у которых черви локализуются под кожей конечностей. У человека наиболее частая локализация — под кожей ног в области суставов. Описаны случаи обнаружения гельминта под серозной оболочкой желудка, под мозговыми оболочками, в стенке пищевода. Над передним концом зрелой самки образуется кожный пузырь, заполненный серозной жидкостью. Человек при этом ощущает сильный зуд, проходящий при соприкосновении с водой. Опускание ног в воду сопровождается разрывом пузыря и рождением живых микроскопических личинок, которые сразу проглатываются промежуточными хозяевами — циклопами. В полости тела циклопов они через несколько дней достигают инвазионности и при проглатывании с водой таких рачков попадают в кишечник, а затем мигрируют под кожу.
Весь жизненный цикл ришты длится 1 год. Интересно, что развитие паразитов у инвазированных людей происходит синхронно,
таким образом, что самки становятся способными рождать личинок одновременно почти у всех носителей паразита. Этим достигается резкое повышение вероятности заражения огромного количества циклопов, а затем и основных хозяев в течение небольшого интервала времени. Эта особенность жизненного цикла ришты имеет огромное адаптивное значение в зонах с засушливым климатом и редкими, повторяющимися из года в год дождевыми периодами. Она имеет и существенное медицинское значение в связи с тем, что в очагах распространения дракункулеза выявляется большое количество пораженных этим паразитом людей в течение небольшого временного интервала. Это облегчает врачу постановку диагноза, лечение и проведение профилактических мероприятий.
Кроме общих аллергических реакций обязательно проявляется и местное воздействие паразита: локальные воспалительные реакции и нарушение функций суставов, прилежащих к зоне поражения.
Диагностика при типичной локализации проста: паразит виден под кожей. Атипичное расположение гельминта требует применения иммунологических реакций.
Личная профилактика
также проста — кипячение или фильтрация питьевой воды, взятой из открытых водоемов. Общественная профилактика —
современное водоснабжение обеззараженной водой; выявление и лечение больных гарантирует успех в борьбе с этим заболеванием.
Рис. 20.17. Трихинелла.
А —
половозрелая стадия из кишечника; Б
— личинка, вышедшая из капсулы
Трихинелла
Trichinella spiralis
(рис. 20.17) — возбудитель трихинеллеза—
мелкий гельминт длиной до 4 мм. Распространен очень широко, на всех континентах и во всех природно-климатических зонах. Этому благоприятствуют такие особенности биологии паразита, как способность личинок переживать неблагоприятные условия на протяжении десятков лет. Хозяевами трихинеллы могут быть различные хищные и всеядные млекопитающие, а также человек. Распространение трихинелл происходит обычно при поедании животными друг друга.
Человек заражается, поедая мясо зараженных животных, чаще всего свиней. Проглоченные личинки в кишечнике быстро достигают половой зрелости. Оплодотворенные самки рождают живых личинок, которые пробуравливаются через стенку кишечника и, транспортируясь кровью, оседают в поперечно-полосатых мышцах: чаще всего в диафрагме, межреберных, дельтовидных. Здесь они после разрушения части мышечных волокон спирально скручиваются и инкапсулируются.
При попадании трихинелл в ослабленный организм возможно развитие первого поколения личинок не в мышцах хозяина, а в ворсинках его кишечника, после разрушения которых личинки вновь возвращаются в просвет кишки, достигают там половой зрелости и размножаются. Следующее поколение личинок уже оседает в мышцах хозяина. Этим достигается резкое увеличение размеров популяции паразита, которые в обычных случаях размножаются у одного хозяина лишь один раз, и соответственно резкое утяжеление заболевания, часто приводящее к смерти.
Личинки трихинелл после гибели хозяина сохраняют жизнеспособность даже после разложения его трупа. В пищеварительной системе трупоядных наземных и даже водных животных — жуков-мертвоедов, почвенных червей, мелких ракообразных, рыб, хищных птиц и чаек — личинки сохраняют жизнеспособность в течение нескольких дней. Перечисленные животные выступают в цикле развития паразитов как транспортные хозяева и способны передавать возбудителя по цепи питания до тех пор, пока личинки не попадут в организм хозяина, в котором возможно нормальное развитие паразита. Сложными цепями питания с передачей личинок транспортными хозяевами обеспечивается фактически всесветное распространение трихинеллы и существование таких природных очагов трихинеллеза, как очаги даже в зоне арктических пустынь.
Известно, что 67% ездовых собак в Гренландии и 50% белых медведей о. Шпицберген, многие тюлени и даже некоторые китообразные поражены трихинеллезом. Основной источник питания этих животных — рыба, морские беспозвоночные, яйца и птенцы морских птиц и т. д. Эти животные становятся транспортными хозяевами трихинеллы после того, как разлагающиеся трупы и помет птиц оказываются в воде и личинки паразита попадают в организм водных ракообразных, которыми питаются более крупные беспозвоночные и рыбы. Поедая их, млекопитающие постепенно накапливают в организме огромное количество трихинелл, становясь опасным источником заражения человека. Свиньи как наиболее частый источник инвазий человека заражаются, поедая не только крыс и мышей, но и насекомых, червей, рыбу, трупы и помет птиц, в которых сохраняются жизнеспособные личинки.
Патогенное действие трихинеллы включает как общеаллергические явления, так и нарушение функций мышц.
Диагноз
основывается на данных анамнеза — употребление мяса диких животных и не проверенной ветеринарной службой свинины, а также на результатах биопсии мышц.
Личная профилактика —
тщательная термическая обработка свинины и особенно мяса диких животных. Общественная профилактика —
санитарный надзор в свиноводстве, проверка свинины на торговых точках и на предприятиях общественного питания.
Биогельминты, передающиеся грансмиссивно
Трансмиссивный способ передачи вообще очень редок среди гельминтов. Он характерен только для этой небольшой группы нематод. Все паразиты этой группы распространены в тропиках. Они относятся к сем. Filarioidea и вызывают заболевания — филяриатозы.
Половозрелые особи, или филярии,
обитают в разных тканях внутренней среды основного хозяина, а рождаемые ими личинки, или микрофилярии,
периодически поступают в кровь и лимфу и некоторое время находятся там (рис. 20.18). Если в этот момент на хозяина нападают кровососущие насекомые, микрофилярии с кровью поступают в желудок, оттуда в мышцы переносчика и за одну или несколько недель достигают там инвазионности, переходят в хоботок переносчика и затем, при укусе, — в кровь основного хозяина. У некоторых видов филярии может быть несколько основных хозяев (человек, обезьяна, собака и т. д.), а переносчики всегда специфичны.
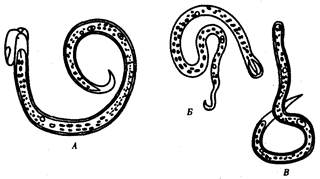
Рис. 20.18. Микрофилярии из крови человека.
А
—
Wuchereria bancrofti;
Б
—
Onchocerca
volvulus
В —
Brughia malayi
В связи с тем что в переносчиках происходит развитие личинок паразита, они являются одновременно и промежуточными хозяевами. Длительные взаимные адаптации филярий, их хозяев и переносчиков привели к своеобразному эффекту — у большинства филярий происходит выход личинок в кровяное русло в строго определенные часы суток, соответствующие периодам максимальной активности переносчиков. Так, у филярий, переносчиками которых являются комары, личинки поступают в кровь в вечерние и ночные часы суток. Если переносчиками являются слепни, то появление микрофилярий в крови приурочено к утренним и дневным часам. В том случае, когда переносчиками служат мокрецы, активность которых больше зависит от влажности воздуха, выход микрофилярий в кровь лишен периодичности. Эти особенности биологии филярий необходимо учитывать при постановке диагноза и брать кровь у больных для обнаружения в ней микрофилярий в то время, когда наличие их там наиболее вероятно. Профилактика
филяриатозов — выявление и лечение больных, борьба с переносчиками.
Особенности биологии наиболее распространенных филярий — паразитов человека — приведены в табл. 20.2.
Таблица 20.2.
Биология наиболее распространенных филярий, паразитов человека
| Название паразита и болезни |
Географическое распространение |
Промежу-точный хозяин и переносчик |
Основной хозяин |
Локализация половозрелых форм, симптомы |
Время появления микрофилярий в крови |
Wuchereria banctofti
Вухерериоз
Brughia
malayi
Бругиоз
|
Тропическая Азия,Африка,
ЮжнаяАмерика
Южная и
Восточная
Азия
|
Комары
»
|
Человек,
высшие
обезьяны
Человек,
обезьяна,
кошачьи
|
Лимфатические узлы и сосуды
Застой лимфы, отеки и развитие слоновости,аллергия
То же
|
Вечер и ночь
Ночь
|
Onchocerca volvulus
Онхоцеркоз
Loa loo
Лоаоз
Mansonella sp.
Мансонел-лез
Acanthohei-
lonema sp.
Акантохей-
лонематоз
|
Эваториальная Африка,
Центральная,
Северная и
Южная
Америка
Центральная и Западная экваториальная
Африка
Центральная и Южная
Америка
Экваториальная Африка, Южная Америка
|
Мошки
Слепни,
кусающие
днем; слепни, кусающие ночью
Мокрецы
»
|
Человек
Человек,
обезьяна
Человек
»
|
Под кожей груди, конечностей, головы
Образование узлов. Слепота при поражении глаз, аллергизация
Под кожей и
слизистой оболочкой
Аллергизация
В брыжейке, жировой ткани, под серозными оболочками
Аллергизация
В брыжейке,
жировой ткани, брюшной полости, в полостях перикарда и плевральной
Аллергизация
|
Нет периодичности
Утро и
день
Ночь
Нет периодичности
»
|
Эта группа нематод содержит гельминтов -— специфических паразитов собак, кошек и других млекопитающих. Человек для них является лишь случайным хозяином, в организме которого невозможно полноценное развитие. Поэтому личинки этих гельминтов, попадая в пищеварительную систему, способны осуществлять лишь начальные этапы миграционной стадии жизненного цикла: они проходят через стенку кишечника, попадают в сосуды, оттуда могут проникать в различные органы, включая головной и спинной мозг, и там гибнут. Патогенное действие их включает аллергизацию и интоксикацию хозяина, а затем воспалительные явления вокруг погибших паразитов.
Диагностика
заболеваний, вызванных этими паразитами, очень сложна. В подозрительных случаях применяют иммунологические реакции.
Toxocara canis
иToxascaris ieonina —
возбудители токсокароза
собак и кошек. Паразиты морфофизиологией и жизненным циклом напоминают аскарид. Миграция личинок в печени вызывает гепатит, а в головном мозге — менингитоподобные явления и даже смерть. Часто заражаются дети, активно контактирующие с домашними животными.
Профилактика —
гигиеническое содержание домашних животных.
Рис. 20.19. ЛичинкаAnisakis sp. A
—
в печени рыбы; Б —
из желудка человека
Гельминты из р.
Anisakis—
возбудители анизакиаза.
Паразитируют в желудке китообразных. Биогельминты. Промежуточными хозяевами их являются морские планктонные рачки, кальмары и рыбы, в мышцах и особенно печени которых инкапсулируются спирально скрученные личинки (рис. 20.19, А
).
Человек заражается, проглатывая личинок с недостаточно термически обработанными или сырыми морскими рыбными продуктами. Личинки у человека гибнут вскоре после проникновения их через стенку желудка.
Патогенное действие связано с нарушением целостности стенки желудка, что сопровождается сильными болями и явлениями гастрита. Возможны перфорации желудка и перитонит, заканчивающийся смертью больного.
Заболевание распространено в странах Дальневосточного региона, особенно широко известно в Японии, где морские продукты обычно употребляют в пищу в сыром виде.
Диагноз
ставится при гастроскопии, когда обнаруживают личинок, внедрившихся в слизистую оболочку желудка (рис. 20.19, Б
).
Профилактика
анизакиаза — санитарный контроль морских продуктов, их термическая обработка при приготовлении пищи.
Медицинская арахноэнтомология
изучает представителей типа Членистоногие Arthropoda, имеющих медицинское значение. Тип Членистоногие содержит более 1,5 млн. видов. Он характеризуется наличием хитинового покрова — скелетного и защитного образования — и членистых конечностей. Тело состоит из сегментов, сливающихся в три отдела: голову, грудь и брюшко. В некоторых группах членистоногих голова и грудь представляют собой единое образование — головогрудь,
иногда тело вообще не расчленено. На голове расположены органы чувств и ротовой аппарат — видоизмененные конечности. В пищеварительной системе имеются сложные железы. Органы дыхания в зависимости от систематического положения и образа жизни жабры, мешковидные легкие или трахеи. Кровеносная система незамкнутая, сердце находится на спинной стороне. На брюшной стороне — нервная цепочка из частично слившихся ганглиев, среди которых самые крупные — подглоточный и надглоточный — расположены на переднем конце тела.
Наибольшее медицинское значение имеют классы Паукообразные и Насекомые. В этих классах встречаются временные и постоянные паразиты, переносчики и возбудители инфекционных и паразитарных заболеваний. В классе Ракообразные встречаются только отдельные виды — промежуточные хозяева некоторых гельминтов. Информация о них изложена в соответствующих параграфах гл. 20.
Представители этого класса имеют два отдела тела: головогрудь и брюшко и шесть пар конечностей: хелицеры, педипальпы и четыре пары ходильных ног. Органами дыхания паукообразных являются мешковидные легкие и трахеи.
Медицинское значение имеют представители отрядов скорпионов Scorpiones, пауков Aranei, среди которых существует немало видов, ядовитых для человека, и клещей Acari. О ядовитых животных см. гл. 23.
21.1.1. Отряд Клещи Acari
Представители этого отряда имеют несегментированное тело. Ротовой аппарат в большинстве случаев представлен сложно .устроенным хоботком, в состав которого входят хелицеры и дополнительное образование — гипостом. Педипальпы
выполняют обонятельную и осязательную функции.
Развитие клещей происходит с метаморфозом. Из яйца вылупляется личинка,
имеющая три пары ног. За ней следует стадия нимфы.
У нее по четыре пары конечностей, но половая система не развита. Нимфа превращается во взрослую стадию — имаго,
отличающуюся более крупными размерами и развитой половой системой. Некоторые виды имеют несколько стадий нимф.
Экология клещей разнообразна. Среди них существуют как свободноживущие хищники, так и паразиты растений, животных и человека. Некоторые из видов клещей обитают в жилище человека. Не' будучи паразитами, они сохранили черты организации, характерные для свободноживущих видов. Другая группа клещей приобрела адаптации к временному эктопаразитизму, но большую часть времени проводит в естественной природе. В связи с этим они не претерпели глубокой дегенерации. Небольшое количество видов клещей паразитирует на человеке постоянно. Адаптация к паразитизму и общая дегенерация их наиболее выражены.
Три перечисленные группы клещей эволюционируют независимо друг от друга. Медицинское их значение различно.
Все клещи этой группы, нападающие на человека, характеризуются выраженными адаптациями к эктопаразитизму: ротовой аппарат их предназначен для прокалывания кожи и всасывания крови, пищеварительная система и покровы тела сильно растяжимы, что позволяет им питаться редко, иногда даже только один раз в жизни. Строгой специфичности хозяев у этой группы нет. Они легко могут переходить к питанию кровью разных видов животных и человека. Выделяемая в ранку слюна клещей обладает местным раздражающим и общетоксическим действием.
Массовые нападения клещей могут быть причиной не только поражений кожи — клещевых дерматитов, но и тяжелых лихорадочных состояний и нервных расстройств. Особенную опасность представляет, однако, не столько собственно питание кровью, сколько способность быть переносчиками возбудителей природно-очаговых протозойных, бактериальных, вирусных болезней, а также заболеваний, вызываемых риккетсиями.
Рассмотрим семейства Иксодовые Ixodidae и Аргазовые Argasidae.
Иксодовые клещи
довольно крупные — от нескольких миллиметров до 2 см в зависимости от степени насыщения. На переднем конце тела ротовой аппарат сильно выступает вперед. Основным компонентом хоботка является гипостом — длинный уплощенный вырост, несущий направленные кзади острые зубцы. Хелицеры имеют вид колющих стилетов, зазубренных с латеральных сторон. С их помощью в коже хозяина образуется резаная ранка, и в нее вводится гипостом.
Первая порция слюны, вводимая в ранку при укусе, обладает способностью застывать вокруг хоботка. В результате разведенные в стороны хелицеры и гипостом прочно заякориваются. Так достигается надежное прикрепление клеща к хозяину, позволяющее ему питаться на протяжении от нескольких часов до 15—20 дней. Средняя кишка имеет многочисленные выросты, заполняющиеся кровью при питании. Здесь кровь может сохраняться до нескольких лет (рис. 21.1). Вся спинная сторона самца покрыта нерастяжимым хитиновым щитком, у самки такой щиток занимает не более половины поверхности тела, поэтому покровы самки значительно более растяжимы и она может выпить гораздо больше крови. После питания самки откладывают от 1500 до 20 000 яиц в лесную подстилку, трещины почвы, в норы грызунов. Личинки
имеют три пары ходильных ног. Они питаются кровью ящериц и мелких грызунов.
Следующая стадия жизненного цикла — нимфа.
Она значительно крупнее личинок, имеет три пары ног и питается на зайцах, белках, крысах. После линьки нимфа превращается в половозрелую стадию.
Взрослый клещ сосет кровь крупных домашних и диких копытных, лис, собак и человека. В связи с тем что для каждого периода при переходе к следующей стадии цикла развития клещам необходимо питаться один раз, большинство иксодовых клещей меняет трех хозяев. Такой цикл развития называют треххозяинным.
Многие иксодовые клещи пассивно подстерегают своих хозяев, однако они локализуются в таких местах, где встреча с прокормителем наиболее вероятна: обычно они располагаются на высоте до 1 м на концах веточек и листьев кустарников вблизи троп, по которым передвигаются животные. Некоторые виды совершают активные поисковые движения.
Иксодовые клещи переносят возбудителей опасных заболеваний. Среди этих заболеваний наиболее известен клещевой весенне-летний энцефалит.
Вирусы размножаются в организме клеща и накапливаются в слюнных железах и яичниках. При кровососании происходит транс-миссивная передача
вирусов хозяину-прокормителю, а при откладке яиц — трансовариалъная передача
следующему поколению клещей. Наибольшее значение как переносчики и резервуары возбудителей имеют следующие виды иксодовых клещей.
Рис. 21.1. Адаптации иксодовых клещей к кровососанию:
1
— хелицеры, 2 —
педипальпы, 3 —
слюнные железы, 4 —
слепые выросты кишечника
Таежный клеш,
Ixodes persulcatus.
Размеры самца 2,5 мм, самки — до 4 мм. Распространен в таежной зоне Евразии от Дальнего Востока до горных районов Центральной Европы. Живет до 3 лет. Этот вид клещей особенно опасен, так как они наиболее часто нападают на человека.
Собачий клещ
Ixodes ricinus
(рис. 21.2). Встречается часто в смешанных, лиственных лесах и кустарниковых зарослях большей части Евразии. От таежного клеща отличается некоторыми деталями строения и более длительным циклом развития — до 7 лет.
Пастбищный клещ
Dermacentor sp.
(рис. 21.2). Чаще встречается на лугах, пастбищах, в лесостепной зоне и в горных лесах. От клещей рода Ixodes
отличается более коротким ротовым аппаратом, беловатым эмалевым рисунком на поверхности спинного щитка и фестончатым, а не гладким задним краем тела.
Клещи рода
Hyalomma
(рис. 21.2) — крупные иксодовые клещи (более 5 мм). Обитают в степной зоне и в горах субтропических районов Южной Европы. Характерной чертой строения являются очень толстые и длинные ноги.
Широкая распространенность иксодовых клещей в различных климатических зонах и биотопах от лесотундры и тайги до сухих субтропиков привела к расселению вируса таежного энцефалита
по огромному ареалу. Расселение вируса и его адаптация к разным видам переносчиков и огромному количеству разных видов природных резервуарных хозяев от землероек и рябчиков до оленей и коз способствовали возникновению его разнообразных штаммов, отличающихся по степени вирулентности. Наиболее тяжелое течение энцефалита со смертельным исходом более чем в 30% случаев и с серьезными необратимыми поражениями центральной нервной системы в виде параличей характерно при заражении дальневосточными штаммами возбудителя. Южно-европейские и восточноевропейские штаммы обладают значительно меньшей вирулентностью, и заболевание протекает либо вообще бессимптомно, либо в виде гриппа. При этом оно часто не диагностируется.

Рис. 21.2. Иксодовые клещи.
А —
собачий; Б —
таежный; В —
пастбищный; Г —
хиаломма
Из домашних животных таежным энцефалитом чаще всего заражаются козы в связи с особенностями их поведения на пастбищах: они предпочитают питаться листьями и ветвями кустарников, при этом чаще, чем коровы, продираются через заросли и используют в лесах малозаметные тропы диких животных. В таких условиях резко повышается возможность их укуса клещами. Кроме того, вместе с ветвями козы проглатывают сидящих на них клещей, поедают они также и клещей, присосавшихся к коже. Сами козы болеют энцефалитом в легкой форме, но вирус энцефалита выделяется ими с молоком. Употребляя такое молоко, способен заражаться и человек.
Способ заражения через козье молоко был впервые описан в Юго-Восточной Словакии в 1951 г., когда всего за несколько дней в небольшом городке Рожнява заболело энцефалитом 1660 человек. Все они покупали в магазине коровье молоко, в которое была подмешана порция молока от четырех коз из пригородной деревни. Вирусологические исследования показали, что и коровье молоко может быть заражено вирусом, но козье молоко содержит вирусы в более высокой концентрации и эпидемиологически гораздо опаснее.

Рис. 21.3. Поселковый клещ
Аргазовые клещи
отличаются от иксодовых тем, что их ротовой аппарат располагается на вентральной стороне тела и совершенно не выступает вперед. Щитка на спинной стороне тела нет. Вместо него покровы снабжены многочисленными хитиновыми бугорками и бляшками и очень сильно растяжимы. По краю тела со всех сторон проходит широкий рант. Продолжительность жизни более 20 лет. Распространены в странах с теплым или жарким климатом. Часто встречаются в Закавказье и Средней Азии, причем не только в естественных биотопах, но и в антропогенных ландшафтах и даже в хозяйственных постройках и жилищах человека.
В течение жизни питаются многократно и каждый раз на новом хозяине. В соответствии с этим самки в отличие от иксодовых клещей могут откладывать яйца несколько раз в жизни.
Эволюция аргазовых клещей как кровососущих членистоногих связана с адаптацией к ним спирохет р.Borrelia,
вызывающих у хозяев-прокормителей клещевые спирохетозы.
Наибольшее значение из них имеет клещевой возвратный тиф.
Спирохеты размножаются в кишечнике клеща и затем проникают во все внутренние органы, в том числе и в яичники, и передаются следующим поколениям клещей трансовариально. Циркуляция спирохет в очагах осуществляется с участием грызунов, собак и кошек. Домашний скот (коровы, лошади, овцы, верблюды) не участвует в передаче возбудителя, но, привлекая к себе клещей для питания, способствует возникновению популяций с высокой плотностью переносчиков, что и повышает вероятность заражения человека. Попадание спирохет в организм человека происходит не только через хоботок при укусе, но и непосредственно через кожу из экскрементов и продуктов выделения клещей.
Имеется несколько клинических вариантов клещевого возвратного тифа. Это зависит от вида возбудителя, причем разные виды спирохет передаются специфическими переносчиками — разными видами аргазовых клещей.
Среднеазиатский возвратный тиф
вызывает спирохета
Borrelia sogdiana,
ее переносчиком является поселковый клещOmithodorus papillipes
(рис. 21.3). Болезнь характеризуется периодическими лихорадочными приступами и воспалительными явлениями в дыхательной системе, а также поражениями центральной нервной системы. Обычно заболевание заканчивается выздоровлением через 2,5—4 мес. с момента заражения.
Более опасен восточноафриканский клещевой возвратный тиф,
сопровождающийся менингитами и параличами.
Надсемейство Гамазовые клещи
Gamasoidea — очень мелкие клещи, размером от 0,2 до 2,5 мм. Ведут хищнический или эктопаразитический образ жизни. Некоторые виды даже приспособились к эндопаразитиз-му. Все тело покрыто длинными щетинками. Ноги длинные и очень хорошо развиты. Хелицеры у паразитических видов превращены в колющие стилеты. Множественные укусы паразитических гамазовых клещей вызывают у человека местные поражения кожи — дерматиты.
Ряд клещей этой группы имеют значение как переносчики возбудителей некоторых вирусных, риккетсиозных и бактериальных заболеваний. Наиболее известны среди них крысиный сыпной тиф,
вирус которого передается клещомOmithonyssus bacoti,
риккетсиоз
Q
—
лихорадка, передающаяся куриным клещомDermanyssus gallinae
(рис. 21.4 А, Б
), а
также туляремия,
в переносе которой участвуют гамазовые клещи, паразитирующие на водяных крысах.
Семейство Краснотелковые клещи
Trombiculidae включает ряд видов, имеющих медицинское значение. Характерным для этих видов является личиночный паразитизм: половозрелые стадии цикла развития протекают в почве, личинки же — временные кровососущие эктопаразиты человека и животных. Большинство из них имеет ярко-красные бархатистые покровы (рис. 21. 4, В
).
Укусы личинок вызывают образование волдырей и сильный зуд. Массовое нападение личинок на человека наблюдается во второй половине лета, во время полевых работ. При этом проявления дерматита могут быть очень сильными. Это состояние называют осенней эритемой.
В результате расчесывания волдырей возможно внесение в кожу бактерий, осложняющих течение болезни.
Некоторые краснотелки Дальневосточного региона распространяют тяжелый трансмиссивный риккетсиоз — японскую речную лихорадку,
или цуцугамуши.
Природным резервуаром этого заболевания являются дикие грызуны.

Рис. 21.4. Гамазовые и Краснотелковые клещи.
А —
Dermanyssus gallinae;
Б —
Omithonyssus bacoti;
В —
Leptotrombidium sp.:
a
- личинка, б —
половозрелая особь
Борьба с кровососущими клещами —
одно из ведущих звеньев в системе противоэпидемиологических мероприятий. Уничтожение паразитических клещей в природе затруднено по экологическим соображениям. Наиболее рациональными являются методы биологической борьбы, не вредящие в целом биогеоценозам, но позволяющие избирательно уничтожать определенный вид паразита, заменяя его другим, не имеющим медицинского значения. Такие методы в отношении клещей находятся в состоянии разработки. Один из путей борьбы — использование животных, паразитирующих на клещах и их яйцах, вызывающих их гибель или снижающих жизнеспособность. При работе в местностях, эпидемиологически неблагоприятных по клещевому энцефалиту и геморрагическим лихорадкам, рационально использовать защитные костюмы с пропиткой их репеллентами. В отношении аргазовых и гамазовых клещей, обитающих в хозяйственных постройках, на скотных дворах или в птичниках, возможно применение химических акарицидных препаратов.
Клещи этой экологической группы — очень мелкие животные, обычно значительно меньше 1 мм. Ротовой аппарат грызущего типа. Способны к активным передвижениям в поисках пищи. Питаются пищевыми запасами — зерном, мукой, копченым мясом и рыбой, сушеными овощами и фруктами, а также эпидермальными чешуйками, слущивающимися с поверхности кожи человека, и спорами плесневых грибов.
Для здоровья человека эти клещи представляют опасность, во-первых, тем, что портят пищевые продукты, делая их несъедобными. Во-вторых, эти клещи могут кусать человека, вызывая тем самым зерновую чесотку, чесотку продавцов бакалейных товаров
и другие варианты дерматитов. Кроме того, с продуктами питания эти клещи могут попадать в пищеварительную систему человека, вызывая тошноту, рвоту, понос. Некоторые виды клещей способны переходить к существованию в анаэробных условиях кишечника и даже размножаться там. При попадании с пылью в дыхательные пути и легкие эти клещи вызывают акаридоз дыхательной системы.
Наиболее известные виды клещей этой группы —Tyroglyphus farinae
[мучной клещ), Т.
casei
(сырный клещ), а также Glyciphagus destructor.

Рис. 21.5. Домашний клещ
Особый интерес представляют в настоящее время так называемые домашние клещи,
обитающие в матрацах, коврах, мягкой мебели и в постельном белье. Наиболее известный домашний клещ —Dermatophagoides pteronyssinus
(рис. 21.5). Размеры его около 0,1 мм, а в 1 г домашней пыли обнаруживается от 100 до 500 экземпляров клещей этого вида. Подсчитано, что в матрасе одной двуспальной кровати может одновременно обитать популяция, состоящая из 2 млн. особей этого вида. Исследования аллергологов последних лет показали, что аллергические реакции к антигенам этого клеща обнаруживаются у 45—85% страдающих бронхиальной астмой, в то время как у неастматиков аллергия к клещам встречается в 5—30% случаев.
В течение нескольких последних лет в Папуа — Новая Гвинея вместе с мягкой мебелью и постельными принадлежностями из Европы были занесены также и домовые клещи. Заболеваемость бронхиальной астмой среди местного населения при этом возросла в 50 раз. Исходя из того что в настоящее время в развитых странах до 10% детей страдают бронхиальной астмой, причем для большинства из них аллергенами являются компоненты домашней пыли, становится понятным медицинское значение клещей этой экологической группы.
Основные меры борьбы с клещами, обитающими в пищевых продуктах,— понижение влажности и температуры в помещениях, где они хранятся.
Борьба с домовыми клещами
состоит в первую очередь в частых влажных уборках помещений, использовании мебели, подушек и матрацев из синтетических материалов, в которых эти животные не могут поселяться.
Эта экологическая группа клещей включает небольшое количество видов. Общими их признаками являются крайне мелкие размеры тела — 0,1—0,4 мм, сильная редукция конечностей, малая подвижность и цикл развития, полностью проходящий на хозяине. Переносчиками возбудителей заболеваний клещи этой группы не являются. Обитая постоянно на человеке и не будучи связанными с окружающей средой, они расселились вслед за человеком повсеместно.
К ним относится чесоточный зудень
Sarcoptes scabiei —
возбудитель чесотки человека
(рис. 21.6, А
).
Близкие виды вызывают чесотку у домашних и диких животных, но строгой специфичностью по отношению к хозяину не обладают. Человека могут поражать чесоточные зудни лошадей, овец, коз, верблюдов, собак и других животных. Они вызывают характерные поражения кожи, но на человеке живут недолго и почти не размножаются.
Рис. 21.6. Чесоточный зудень (А
) и железница угревая (Б
) ,
Самка чесоточного зудня длиной до 0,4 мм. Самец значительно меньше. Все тело покрыто щетинками разной длины, и на конечностях имеются присоски. Ротовой аппарат адаптирован к прогрызанию ходов в толще эпидермиса, где обитает этот паразит. Самка откладывает до 50 яиц за всю жизнь, которая длится около 15 сут. Чесоточные зудни могут поселяться в любом месте кожи, но наиболее часто — на нежных ее частях: в межпальцевых промежутках, на сгибах рук, на половых органах. Перемещаясь в толще кожи, клещи раздражают нервные окончания и вызывают нестерпимый зуд, усиливающийся ночью. При расчесывании ходов клещей они переносятся на другие участки кожи или на другого человека. Так происходит расселение клещей по телу хозяина и заражение чесоткой здоровых людей. Заразиться чесоткой можно также и при пользовании одеждой, постельными принадлежностями и личными вещами больного.
Диагностика
чесотки проста, так как поражения кожи клещами очень характерны. Они представляют собой прямые или извилистые полоски грязно-белого цвета. На одном из концов хода располагается пузырек, в котором находится клещ. Его можно перенести на предметное стекло в каплю 50 %-ного раствора глицерина и микроскопировать.
Для профилактики
чесотки необходимо соблюдение правил личной гигиены, выявление и лечение больных и дезинфекция их одежды, белья и полотенец, а также осторожность при общении с животными.
Железница угревая
Demodex folliculorum
(рис. 21.6, Б
) —
возбудитель демодикоза.
Эти клещи имеют червеобразную форму, длиной не более 0,4 мм. Они обитают в сальных железах и волосяных фолликулах кожи лица, шеи и плеч, располагаясь головным концом вниз, обычно группами по четыре особи. Часто встречаются у совершенно здоровых лиц, не вызывая никаких симптомов. Однако у ослабленных людей, в особенности со склонностью к аллергическим реакциям, железницы могут активно размножаться, вызывая закупорку протоков сальных желез, возникновение угрей розового цвета с гнойным содержимым.
Для диагностики
демодикоза выдавленное содержимое сальной железы или выдернутую ресницу помещают в каплю бензина на предметное стекло и микроскопируют. Таким образом могут быть обнаружены взрослые формы, нимфы, личинки и яйца паразита.
Расселение угрицы по человеческим популяциям происходит при личном контакте и пользовании общими полотенцами и бельем, причем у 40—60% населения можно обнаружить угрицу, живущую как комменсал. Поэтому профилактика
демодикоза сводится большей частью к лечению основных заболеваний, ослабляющих организм, а также выявлению и лечению больных с выраженными аллергическими реакциями.
Это самый многочисленный по числу видов класс животных. Общее их количество достигает 1 млн. Тело подразделяют на голову, грудь и брюшко. На голове находятся органы чувств — усики и глаза, сложный ротовой аппарат, строение которого связано со способом питания:
грызущий, лижущий, сосущий, колюще-сосущий и т. п. Грудь насекомых состоит из трех сегментов, каждый из которых несет по паре ходильных ног, построенных по-разному, в зависимости от способа передвижения и двигательной активности. Большинство свободноживу-щих насекомых имеют на груди также две пары крыльев, однако некоторые группы, перешедшие к паразитическому образу жизни, их утратили. Брюшко конечностей не имеет. Органы дыхания насекомых.—трахеи
(рис. 21.7). Остальные системы органов насекомых соответствуют организации членистоногих. Развитие насекомых происходит с метаморфозом — неполным,
когда из яйца вылупляется личинка, превращающаяся во взрослую форму или имаго постепенно, после нескольких линек, и полным,
при котором в ходе онтогенеза сменяются стадии яйца, личинки, куколки и имаго.
Среди насекомых, имеющих медицинское значение, выделяют следующие группы:
а) синантропные виды, не являющиеся паразитами;
б) временные кровососущие эктопаразиты;
в) постоянные кровососущие паразиты;
г) тканевые и полостные ларвальные (личиночные) паразиты.
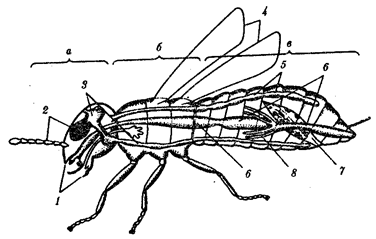
Рис. 21.7. Организация насекомого:
а —
голова, б —
грудь, в —
брюшко; 1
— ротовой аппарат, 2 —
органы чувств, 3 —
нервная система, 4 —
крылья, 5 —
кровеносная система, 6 —
пищеварительная трубка, 7 — половые органы, 8
— мальпигиевы сосуды
21.2.1. Синатропные насекомые, не являющиеся паразитами
К этой группе насекомых относят виды, которых привлекает своеобразие экологических условий человеческого жилища: постоянство действия микроклиматических факторов и независимость от сезонных изменений условий в природе, наличие постоянных источников питания и многочисленных убежищ. Поэтому адаптации у этих животных затрагивают в первую очередь поведенческие реакции — изменение инстинкта откладки яиц, предпочтение закрытых помещений и т. д. Эти насекомые связаны с человеком и его предками менее тесно по сравнению с другими группами и относительно недолго — с момента начала использования гоминидами естественных убежищ и строительства примитивных жилищ.
В связи с этим регрессивной морфофизиологической эволюции они не претерпели. Большинство таких насекомых — теплолюбивые виды тропического и субтропического происхождения, относящиеся к разным отрядам и семействам. Медицинское значение из них имеют лишь те, которые используют продукты питания, пищевые отходы или фекалии человека. Большинство из видов насекомых этой экологической группы являются механическими переносчиками возбудителей инфекционных и паразитарных заболеваний. Вирусы, бактерии, цисты простейших и яйца гельминтов переносятся ими на лапках, поверхности тела или в пищеварительной системе, не развиваясь и не размножаясь. В организме некоторых видов насекомых развиваются личиночные стадии ряда гельминтов (см. разд. 20.1.2.3).
К насекомым этой группы относятся тараканы, а также синантропные мухи, муравьи и жуки. Основными мерами борьбы с
синатропными насекомыми являются благоустройство жилища и поддержание в нем постоянной чистоты, хранение продуктов питания в недоступных для насекомых местах, в закрытой таре.
Тараканы
— всеядные насекомые довольно крупных размеров. В тропиках и субтропиках встречаются как в естественной природе, так и в жилище человека. Здесь же наиболее велико их видовое разнообразие. В умеренных широтах распространены только два вида — черный таракан
Blatta orientalis
и рыжий таракан
Blattella germanica
(рис. 21.8). Размеры черного таракана 19—26 мм, цвет черно-бурый. Рыжий таракан значительно мельче—до 11—12 мм, цвет его рыжеватый. Тело тараканов уплощено, на лапках имеются коготки и присоски, благодаря которым эти насекомые проникают в узкие щели и ползают в любом положении. Крылья их недоразвиты, и поэтому летать они почти не могут.
Развиваются тараканы, откладывая коконы, содержащие до 20 яиц. За 20—50 сут завершается эмбриональное развитие, и из оболочки кокона выходят мелкие светлые личинки. До достижения половой зрелости они линяют несколько раз. Питаются тараканы любыми пищевыми продуктами, а также кожей, бумагой, ватой и шерстью.

Рис. 21.8. Тараканы. А —
рыжий; Б —
черный
Кроме общих мер борьбы с тараканами используют отравленные приманки с добавлением борной кислоты, патогенных для них бактерий и т. д. В целом интенсивная дезинсекция обычно только снижает численность этих животных, не уничтожая их полностью. Это обеспечивается их слабой чувствительностью к ядохимикатам, наличием в трахеях специальных клапанов, закрывающихся при наличии в воздухе посторонних примесей, большой подвижностью, способностью к длительному голоданию, а также широким генетическим полиморфизмом, сформировавшимся в популяциях этих животных за длительное время контактов с человеком.
Мухи
— известны как наиболее активные механические переносчики возбудителей заболеваний. Как и у всех двукрылых, у мух одна пара передних крыльев; развитие происходит с полным метаморфозом. Важное медицинское значение имеет комнатная муха
Musca domestica
(рис. 21.9, А
).
Встречается в жилище человека во всех природных зонах. Размеры тела 6—8 мм, цвет серо-бурый. На груди выделяются четыре темные продольные полосы. Ротовой аппарат сосущего типа. Муха способна питаться не только жидкой, но и твердой пищей, предварительно смачивая ее слюной. Самка откладывает яйца в местах скопления гниющих органических веществ. За 5—10 сут развивается личинка, за 4—7 сут — куколка. Вышедшие из оболочек куколки мух становятся половозрелыми на 5—6-е сутки. За всю жизнь одна самка откладывает около 600 яиц. По сравнению с тараканами муха более опасна как механический переносчик возбудителей заболеваний, так как она более активно меняет источники питания и места пребывания, а местами массового их выплода являются выгребные ямы, помойки и нечистоты. На поверхности тела мухи и в ее пищеварительном тракте может находиться одновременно до 35 млн. разных микроорганизмов.
Кроме комнатной мухи такое же значение имеют синяя и серая мясные, зеленая падальная (рис. 21.9, Б — Г
) и ряд других.
Основная мера борьбы
с мухами — благоустройство мусоропроводов и мусоросборников, гигиена жилища.
Синантропные муравьи
представлены только одним видом. Это домовый муравей
Monomorium pharaonis,
типичный тропический вид, занесенный в последние десятилетия с продуктами питания в страны умеренных широт. Встречается только в хорошо отапливаемых жилищах человека. Благодаря крошечным размерам (1—1,5 мм) проникает в любые щели и легко переходит по мельчайшим трещинам в кирпичах из квартиры в квартиру, где чаще обнаруживается в кухнях, туалетах и ванных комнатах. Популяции муравьев редко образуют большие скопления и постоянно перемещаются, что осложняет борьбу с ними. Хорошим средством против этих насекомых являются пищевые приманки с борной кислотой. Поедание приманок муравьями снижает их жизнеспособность и плодовитость. Для достижения стойкого эффекта необходимо применять приманки в течение нескольких месяцев.

Рис.
21.9. Синантропные мухи. А —
комнатная; Б —
синяя мясная; В —
зеленая падальная; Г —
серая мясная
Жуки.
В отличие от тараканов, мух и домовых муравьев жуки из рода
Tenebrio
(рис. 21.10) не являются, за редким исключением, переносчиками возбудителей заболеваний. Они обитают в муке и крупе, подвижность их невысока. Представляют интерес в связи с тем, что в них развивается личиночная стадия карликового цепня — цистицеркоид (см. разд. 20.1.2.3). Заражение человека гименолепидозом может произойти при пропитывании инвазированного жука или его личинки с непропеченным хлебом или кондитерскими изделиями.

Рис. 21.10. Мучной жук
21.2.2. Насекомые — временные кровососущие паразиты
Насекомые этой экологической группы отличаются высокой подвижностью. Они посещают прокормителя только для питания, а остальное время проводят в естественной природной среде или в жилище и хозяйственных постройках человека. Временные кровососущие паразиты питаются многократно, часто на разных хозяевах, хотя и отдают предпочтение определенным видам теплокровных животных. Это облегчает циркуляцию возбудителей трансмиссивных заболеваний между животными разных видов и человеком, поэтому большинство заболеваний этой группы являются природно-очаговыми зоонозными болезнями. Исключение составляет только малярия.
Большинство переносчиков этой группы по отношению к переносимым возбудителям строго специфичны. Это объясняется особенностями их физиологии и морфологии, к которым на протяжении длительной эволюции у паразитов возникают специфические адаптации. Так, например, возбудители онхоцеркоза, которые могут переноситься комарами родовCulex
иAnopheles,
не инвазируют комаров р. Aedes.
Это связано с особенностями пищеварения последнего вида, у которого кровь сразу после питания свертывается и микрофилярии не могут мигрировать из кишечника в полость тела. В полости тела комаров родов Culex
иAnopheles
микрофилярии развиваются после свободной миграции из кишечника, кровь в полости которого долгое время не свертывается. Интересно, что при добавлении антикоагулянтов к крови, которой питается комар р. Aedes,
личинки филярий у него нормально развиваются.
Слюна кровососущих насекомых обладает антикоагулянтными свойствами, вызывает зуд и местное раздражение кожи. У некоторых людей возможны тяжелые аллергические реакции на их укусы.
К временным кровососущим паразитам относят представителей отрядов Блохи Siphonaptera, Полужесткокрылые Hemiptera и двукрылые Diptera.
Из перечисленных отрядов самыми специализированными паразитами является отряд Блохи, все представители которого ведут паразитический образ жизни. Среди полужесткокрылых и двукрылых абсолютное большинство видов — свободноживущие формы.
Отряд Блохи.
Это мелкие насекомые длиной от 1 до 5 мм. Паразитирование блох облегчается сплющенностью тела с боков, наличием на поверхности его большого количества щетинок, направленных остриями назад, и колюще-сосущим ротовым аппаратом. Задние конечности удлинены и служат для передвижения прыжками. Признаками дегенерации являются рудиментарные глаза и отсутствие крыльев. Развитие блох идет с полным метаморфозом.
Рис. 21.11. Блохи. А
—человеческая; Б
—крысиная; В—
желудок блохи суслика, блокированный бактериями чумы
Наиболее известны человеческая блоха
Pulex imtans
и крысиная блоха
Xenopsylla cheopis
(рис. 21.11, А, Б
).
Оба вида предпочитают питаться кровью соответственно человека и крыс, но легко переходят также на другие виды животных. Крысиная блоха живет в норах крыс, а человеческая — в трещинах пола, за плинтусами и обоями. Здесь самки откладывают яйца, из которых развиваются червеобразные личинки, питающиеся разлагающимися органическими веществами, в том числе фекалиями взрослых блох. Через 3—4 недели они окукливаются и превращаются в половозрелых насекомых.
Человека блохи посещают ночью. Укусы их болезненны и вызывают сильный зуд. Но основное значение блох в том, что они являются переносчиками бактерий — возбудителей чумы.
Бактерии чумы, попав в желудок блохи, размножаются там настолько интенсивно, что полностью закрывают его просвет. Это состояние называют чумным блоком
(рис. 21.11, В
).
Если блоха начинает питаться на здоровом животном или человеке, она, проколов кожу, в первую очередь отрыгивает в ранку бактериальный комочек, благодаря чему в кровь поступает сразу огромное количество возбудителей.
Природным резервуаром чумы являются грызуны — крысы, суслики, сурки и др. Эти животные болеют целым рядом других инфекционных заболеваний: туляремией, крысиным сыпным тифом
и т. д. Поэтому блохи известны как переносчики возбудителей и этих природно-очаговых заболеваний. Интересно, что кроме трансмиссивного способа заражения указанными болезнями существуют и другие пути: при контакте с зараженными животными, при питье воды из открытых водоемов и т. п., но при укусе блохой заражение является наиболее вероятным, а клиническая картина — наиболее тяжелой.
Борьба
с блохами — содержание жилых помещений и хозяйственных построек в чистоте, применение инсектицидов и различных средств борьбы с грызунами. Дают эффект и меры индивидуальной защиты, например репелленты, которыми пропитывают одежду и постельное белье.
Отряд Полужесткокрылые, или Клопы.
Характерной особенностью клопов является строение крыльев и ротового аппарата. Передние крылья в проксимальной части сильно хитинизированы, а в дисталь-ной — прозрачны. Колюще-сосущий ротовой аппарат образует два канала. Один из них служит для всасывания жидкой пищи, второй — для выведения секрета слюнных желез. Развитие с неполным метаморфозом. Медицинское значение имеют клопы из родовCimex, Triatoma
и некоторых близких им.
Постельный клоп
Cimex lectularius ~
наиболее адаптированный к паразитическому образу жизни вид. Тело его сплюснуто в дорсовент-ральном направлении и покрыто сильно растяжимым хитиновым покровом. Крылья полностью редуцированы. На человека клопы нападают ночью, а день проводят в укрытиях — в мебели, за обоями. Здесь же и размножаются. Способны голодать по нескольку месяцев. Известно, что в организме клопов могут длительно сохранять жизнеспособность возбудители многих трансмиссивных заболеваний: рик-кетсии сыпного тифа и спирохеты возвратного, висцерального лейшманиоза и чумы. Тем не менее доказательств роли клопов в переносе этих инфекционных болезней нет. Это связано, вероятно, с тем, что клопы р. Cimex
никогда не испражняются во время кровосо-сания.
На человека кроме постельного клопа часто нападают и другие клопы этого рода, паразитирующие на летучих мышах и птицах.

Рис. 21.12. Паразитические клопы. А —
постельный; Б -
поцелуйный
Несомненно больший интерес представляют южноамериканские поцелуйные клопы из родовTriatoma, Panstrongylus
и др. (рис. 21.12). Это крупные, до 4 см длиной, яркоокрашенные насекомые, ведущие ночной образ жизни. Днем они скрываются в различных убежищах. Являются специфическими переносчиками возбудителя южноамериканского трипаносомоза,
или болезни Чагаса
—Trypanosoma
с
r
и
z
i.
Заражению человека способствует легкость перехода клопов этой группы от одного хозяина к другому и такая особенность биологии, как обязательная дефекация сразу после укуса, при которой обеспечивается попадание инвазионной формы трипаносом в кровь через кожу, поврежденную хоботком клопа.
Борьба
с клопами сводится к улучшению жилищно-бытовых условий, к поддержанию чистоты и обработке стен, мебели и других поверхностей контактными ядами длительного действия (хлорофос, метафос и др.).
Отрад Двукрылые.
У представителей этого отряда пара крыльев, ротовые аппараты их имеют форму хоботка, предназначенного для высасывания жидкой пищи из растительных или животных тканей либо для слизывания ее с поверхности. Развитие с полным метаморфозом. Наибольшее значение представители этого отряда имеют как временные кровососущие паразиты и переносчики возбудителей трансмиссивных заболеваний. Массовые виды этого отряда, размножаясь в летний период в тундре и тайге в огромном количестве, делают жизнь человека почти невыносимой. Разные виды кровососущих двукрылых объединяются общим понятием гнус.
Самцы большинства кровососущих двукрылых питаются соками растений, а самки — преимущественно или исключительно кровью животных и человека. В большинстве случаев откладка яиц самками наступает только после кровососания.
Комары
(сем. Culicidae). Имеют тонкое стройное тело, длинные ноги и небольшую головку с ротовым аппаратом в виде длинного хоботка. Распространены повсеместно. Они сохраняют в организме и обладают способностью передавать животным и человеку возбудителей более 50 вирусных, бактериальных и паразитарных заболеваний. После оплодотворения самки активно ищут хозяина-прокормителя и способны обнаруживать его на расстоянии до 3 км с помощью обоняния, а затем и зрения. Больных с высокой температурой комары кусают более охотно.
Слабое локальное освещение в вечерние и предутренние часы стимулирует двигательную активность комаров и поиск ими хозяев. В связи с этим комары в темное время суток часто влетают в освещенные окна и двери жилых и хозяйственных построек. В дневные часы суток при высокой температуре и низкой влажности воздуха комары обычно находятся в укрытиях и не питаются. Указанные особенности поведения комаров отражаются на адаптациях к паразитизму возбудителей заболеваний, передающихся комарами: при большинстве заболеваний этой группы у больных повышается температура в вечерние и ночные часы суток, в это же время в крови обнаруживается максимальная концентрация инвазионных для комаров стадий возбудителей.
Комары откладывают яйца в воду или на влажную почву около воды. Личинки и куколки ведут водный образ жизни, а дышат атмосферным воздухом с помощью трахей. Личинки питаются взвешенными в воде мельчайшими органическими частичками. Наиболее известны комары из родовCulex
иAedes
(немалярийные комары) — переносчики возбудителей японского энцефалита, сибирской язвы, желтой лихорадки,
а такжеAnopheles
(малярийные комары) — специфические переносчики малярийного плазмодия.
Доказано, что восприимчивость комаров к заражению возбудителями малярии определяется генотипически и наследуется моногенно. Малярийные и немалярийные комары легко отличаются друг от друга на всех стадиях их жизненного цикла (рис. 21.13).
Яйца малярийных комаров р. Anopheles располагаются на поверхности воды поодиночке, и каждое снабжено двумя воздушными поплавками. Личинки их плавают в горизонтальном положении под поверхностью воды, а на предпоследнем членике имеют пару дыхательных отверстий. Куколки по форме напоминают запятые, находятся, как и личинки, под водной поверхностью и дышат кислородом воздуха через дыхательные рожки, имеющие форму широких воронок. Взрослые малярийные комары, сидя на предметах, располагаются под углом к их поверхности головкой книзу. Находящиеся по обе стороны от хоботка нижнечелюстные щупики равны ему по длине либо немного короче.
Рис. 21.13. Комары на разных стадиях жизненного цикла.
А —
Anopheles sp.;
Б—
Culex sp.:
I —
яйца, II
—
личинки.III
—
куколки, IV —
имаго; 1
—
воздушные попалвки, 2—
плотикиз яиц,3—
дыхательные отверстия, 4—
дыхательный сифон, 5—воронкообразные дыхательныерожки,
6—
цилиндрические дыхательные рожки, 7 — нижнечелюстные щупики
Немалярийные комары pp.Culex и Aedes откладывают яйца, слипающиеся группами в небольшие плотики серо-стального цвета. Личинки располагаются под поверхностью воды под углом к ней и имеют на предпоследнем членике длинный дыхательный сифон. Дыхательные рожки куколок имеют форму тонких цилиндрических трубочек, а нижнечелюстные щупики взрослых комаров коротки и достигают не более трети длины хоботка. Тело немалярийные комары держат параллельно поверхности, на которой сидят.
Борьба с
комарами наиболее результативна в отношении водных стадий жизненного цикла — личинок и куколок. Применяются мелиоративные методы — засыпка канав и карьеров со стоячей водой. Возможна обработка ядохимикатами отдельных водоемов с большой концентрацией личинок и куколок, а также мест массовых скоплений половозрелых стадий комаров в дневное время суток (сараи, скотные дворы). Наиболее эффективными являются биологические меры борьбы
в сочетании с гидромелиоративными, проводящимися в соответствии с государственными антималярийными программами. Так, в Западном Закавказье удалось быстро снизить численность комаров и заболеваемость населения малярией за счет мелиорации и разведения рыб — гамбузий, питающихся преимущественно личинками двукрылых. Для индивидуальной защиты
применяют репелленты и механические средства: марлевые пологи, сетки и т. д.
Москиты
(сем. Phlebotomidae). Более мелкие насекомые длиной 1,5—3,5 мм, имеющие короткий хоботок, сильно выступающий в виде горбика грудной отдел тела и обильное опущение тела и крыльев мелкими щетинками (рис. 21.14, А
).
Москиты встречаются в тропических и субтропических зонах на всех континентах. Они известны как переносчики разных видов лейшманий, вирусов лихорадки паппатачи
и возбудителей ряда других трансмиссивных заболеваний. Вирус лихорадки паппатачи передается в поколениях инвазированных москитов трансовариально. Москиты не способны к больших перелетам, поэтому природные очаги заболеваний, вызываемых перечисленными паразитами, обычно имеют небольшие размеры.
Яйца москиты откладывают в норы грызунов и другие затененные места с большим количеством органического вещества и высокой влажностью. Личинки развиваются около 2 мес, а затем окукливаются. Половозрелые стадии появляются через 10—12 сут.
Рис. 21.14. Мелкие кровососущие двукрылые.
А —
москит; Б —
мошка; В —
мокрец
Борьба
с москитами должна вестись комплексно и быть направленной на уничтожение природных очагов лейшманиозов и других трансмиссивных заболеваний: это уничтожение грызунов и мест выплода москитов, обработка инсектицидами поверхностей в хозяйственных постройках и жилищах. Эффективны также индивидуальные средства защиты от укусов.
Мошки
(сем. Simuliidae). Мелкие кровососущие насекомые длиной 2—6 мм. Ротовой аппарат короткий и очень мощный, предназначен для прокалывания кожи и слизывания крови. Крылья прозрачны, без пятен, конечности короткие и толстые (рис. 21.14, Б
).
Распространены повсеместно, но особую опасность представляют в Африке и тропической Америке, где являются переносчиками возбудителей онхоцеркоза.
Интересно, что мошки кусают человека и животных преимущественно в бедра и боковые поверхности туловища. Именно в этих областях скапливаются микрофилярии онхоцерков, что значительно увеличивает вероятность их передачи.
В большом количестве мошки встречаются поблизости от ручьев и речек с быстрым течением, чистой и прохладной водой. На дне таких водоемов самки откладывают яйца, из которых выходят личинки, ведущие прикрепленный и малоподвижный образ жизни. Через 2—3 недели формируется куколка, и еще через 1 неделю из ее оболочки выходят взрослые мошки. Развитие их идет синхронно, и огромное количество мошек на больших территориях появляются одновременно.
Одним из наиболее экологически чистых методов борьбы
с мошками является механическая очистка зон особенно быстрого течения рек и ручьев, где на дне и лежащих предметах скапливаются личинки мошек. При этом они отрываются от места прикрепления и уносятся течением. Многие из них при этом гибнут или поедаются различными хищниками. В редких случаях в очагах онхоцеркоза при высокой концентрации личинок мошек на небольших площадях возможно применение инсектицидов.
Мокрецы
(сем. Ceratopogonidae). Самые мелкие из кровососущих двукрылых. Их размеры от 1 до 4 мм. Большинство видов имеют пятнистые крылья, длинные многочленистые усики и колюще-сосущий хоботок (рис. 21.14, В
).
Обитают повсеместно, но так же, как мошки, в тропиках являются переносчиками возбудителей некоторых филяриатозов.
Размножаются мокрецы в небольших стоячих водоемах или во влажной почве. Развитие от яйца до имагинальной стадии продолжается около 1 мес. На человека нападают преимущественно при высокой влажности воздуха или во время дождя. Против мокрецов наиболее эффективны индивидуальные средства защиты.
Слепни
(сем. Tabanidae) — наиболее крупные кровососущие двукрылые, до 30 мм длиной. Общим видом напоминают крупных мух. Ротовой аппарат сочетает в себе черты колюще-сосущего и лижущего.
Рис. 21.15. Крупные кровососущие двукрылые. А — слепеньTabanus sp.;
Б —
слепень Chrysophthalmus sp.;
В —
муха осенняя жигалка
Все тело слепней покрыто тонкими щетинками (рис. 21.15, А, Б
).
Распространены повсеместно. В северных широтах являются переносчиками бактерий — возбудителей сибирской язвы
и туляремии,
а в тропиках и филярий — возбудителей лоаоза.
Американские слепни кусают человека и животных преимущественно в голову, где располагаются микрофилярии. Слепни — теплолюбивые и светолюбивые насекомые. Они активны в жаркие дневные часы на севере только летом, а в тропиках — в любом сезоне. На человека нападают обычно около воды.
Яйца самки слепней откладывают на околоводных растениях. Личинки развиваются во влажном иле около уреза воды и ведут хищнический образ жизни. Цикл развития в среднем продолжается около 1 года.
Против укусов слепней эффективны средства индивидуальной защиты.
Настоящие мухи
(сем. Muscidae). Насчитывают несколько кровососущих видов. Интерес представляют осенняя жигалка и мухи це-це.
Осенняя жигалка
Stomoxys calcitrans —
муха средних размеров серого цвета со сверлящим хоботком (рис. 21.15, В
).
Кровью питаются как самцы, так и самки. Личинки развиваются в навозе. В умеренных широтах массовая активность мух наблюдается в конце лета и начале осени, а в тропиках — круглый год. Держится обычно около животноводческих комплексов, активно нападает и на животных, и на человека. Является механическим переносчиком возбудителей туляремии
и сибирской язвы.
Для борьбы с
массовым выплодом жигалок необходимо гигиеническое содержание домашних животных, а предохранение от укусов достигается индивидуальными средствами защиты.
Мухи це-це р.
Glossina
широко распространены в экваториальной Африке. Довольно крупные мухи длиной до 13,5 мм. Ротовой аппарат сходен с хоботком осенней жигалки (рис. 21.15, В
).
Самки живородящи, рождают периодически по одной личинке, которая сразу окукливается, углубляясь в почву. Через 3 недели появляется имагинальная форма. Размножение происходит в тени деревьев и кустов на берегах водоемов. Несколько близких видов мух це-це отличаются друг от друга особенностями окраски, а главное — биологии: одни виды поселяются преимущественно около жилищ человека и питаются в основном его кровью и кровью домашних животных. Другие — обитают в естественной природе (в саваннах и лесах), предпочитая питаться кровью крупных диких копытных, а человека кусают случайно.Все виды являются специфическими переносчиками возбудителя африканского трипаносомоза
(см. разд. 19.3.2). Основной мерой борьбы
является индивидуальная защита от укусов.
21.2.3. Насекомые — постоянные кровососущие паразиты
К постоянным кровососущим паразитам человека из класса насекомых относятся только вши.
Человек для них является единственным хозяином, поэтому и трансмиссивные заболевания, возбудителей которых переносят вши, являются типичными антропонозами.
Вши характеризуются выраженными адаптациями к эктопаразитизму: размеры их невелики, конечности снабжены аппаратом фиксации к коже, волосам и одежде, ротовой аппарат колюще-сосущего типа, цикл развития упрощен (развитие с неполным метаморфозом), все стадии жизненного цикла обитают и питаются на хозяине. Постоянство паразитизма этих организмов сопровождается признаками их общей дегенерации: вши в отличие от большинства насекомых имеют не фасеточные глаза, а простые, конечности не обеспечивают быстрого передвижения, полностью редуцированы крылья.
У человека паразитируют два вида вшей: человеческая
Pediculus humanus
и лобковая
Phthirus pubis.
Вид человеческая вошь представлен двумя подвидами: Р.
h. Capitis —
головная, Р.
h. humanus —
платяная вошь (рис. 21.16).
Кроме человека головная вошь может поселяться на коже головы африканских человекообразных обезьян — шимпанзе и гориллы, что указывает на древность экологических связей с высшими узконосыми обезьянами. У других животных человеческие вши либо не пьют кровь, либо, будучи голодными, пьют, но потом погибают.
В то же время центрально-американские популяции головных вшей способны переходить от человека к капуцинам — низшим широконосым обезьянам, которых местное население традиционно держит в неволе. У диких капуцинов вши не встречаются. Это свидетельствует о широкой экологической пластичности и эволюционных перспективах данного подвида паразитов.
С возникновением человека современного физического типа и его широким расселением по территории с умеренным и холодным климатом вслед за ним стали распространяться и вши. Ношение одежды из шкур и позже из тканей в новых зонах обитания человека создало новые возможности расселения этих паразитов по телу хозяина. В результате появления адаптации к обитанию на одежде возник новый подвид — вошь платяная,
которая отличается от головной рядом морфофизиологических признаков, но свободно скрещивается с ней и дает плодовитое потомство. В соответствии с климатическими особенностями исходного ареала обитания платяная вошь встречается только в странах с холодным и умеренным климатом, а в тропиках лишь в условиях высокогорья. Лобковая вошь встречается реже, чем человеческая, но распространена повсеместно. Обитает на лобке, в подмышечных впадинах, иногда на бровях и ресницах. Паразитирование на человеке головной и платяной вшей называется педикулезом.
Зараженность лобковыми вшами носит название фтириаза.
Рис. 21.16. Вши. А —
головная; Б —
платяная; В —
лобковая; Г —
гниды вшей
Кроме разных мест обитания на человеке вши отличаются друг от друга морфологическими и физиологическими признаками и особенностями жизненного цикла. Самая крупная вошь — платяная. Ее размеры до 4,7 мм. Головная достигает длины 3 мм, а лобковая — не более 1,5 мм. Платяная и головная вши имеют четко отграниченные друг от друга головку, грудь и брюшко, а у лобковой грудь и брюшко слиты. Платяная вошь живет около 50, головная — около 40, а лобковая — до 30 сут. Головная и платяная вши питаются кровью по 2—3 раза в сутки, а лобковая — почти постоянно малыми порциями. Самка платяной и головной вшей откладывает до 300 яиц за всю жизнь, лобковой — до 50. Яйца приклеиваются к волосам или нитям одежды и называются гнидами.
Они очень устойчивы.
Слюна вшей обладает токсическими свойствами. Она вызывает ощущение жжения и зуда. У некоторых людей на укусы этих насекомых могут развиваться аллергические реакции. На месте укусов остаются поверхностные кровоизлияния. Ссадины на местах расчесывания инфицируются и загнаиваются. Волосы на голове при этом склеиваются, и образуется трудноизлечиваемое болезненное состояние — колтун.
Лобковая вошь является только паразитом, а головная и платяная — еще и специфическими переносчиками спирохетBorrelia reccurentis —
возбудителей возвратного тифа,
риккетсийRicketsia prowaceki
—
возбудителей эпидемического сыпного тифа,
R. wolhynica —
возбудителей волынской лихорадки.
Спирохеты размножаются и развиваются в полости тела вшей, поэтому заражение происходит при раздавливании паразитов и попадании их гемолимфы в ранку от укуса или в расчесы.
Риккетсий размножаются в стенке кишечника вшей и выделяются оттуда с фекалиями. Заражение человека происходит при попадании фекалий вшей с риккетсиями в кожные ранки или на слизистые оболочки глаз и дыхательных путей. В связи с тем что риккетсий сохраняют жизнеспособность в сухом виде в течение нескольких месяцев, возможно заражение человека ими без укуса вшами, например при контакте с одеждой больных, в складках которой сохранились сухие фекалии или мертвые насекомые.
Профилактика
описанных инфекционных заболеваний — это в первую очередь борьба с педикулезом. Основная мера профилактики педикулеза — соблюдение правил личной гигиены, особенно в местах массового скопления людей. Из химических средств уничтожения вшей используют мази и шампуни, содержащие инсектициды, а также лекарственные препараты, применяемые внутрь и действующие через кровь, которой питаются эти паразиты. Наиболее эффективны короткая стрижка для уничтожения гнид и обработка белья в дезинфекционных камерах.
21.2.4. Насекомые — тканевые и полостные эндопаразиты
В связи с высоким уровнем организации насекомых эндопаразитический образ жизни для них не характерен. Однако личинки ряда видов из отряда Двукрылые развиваются в трупах, гниющих остатках органического вещества, в почве. Некоторые из них приобрели адаптации к паразитическому образу жизни, поселяясь как в полостных органах, так и в тканях внутренней среды.
Личинки этой группы двукрылых способны к анаэробной диссимиляции и к передвижению внутри организма хозяина. Окукливание происходит во внешней среде, а взрослые формы ведут свободный образ жизни и мало отличаются от большей части других двукрылых.
Заболевания, вызываемые личинками двукрылых, называют миазами.
Вольфартова муха
Wohlfahrtia magnified
(рис. 21.17, А
) обитает в южных районах Европы, на Ближнем Востоке, в Средней Азии. Взрослые формы питаются нектаром цветов. Они рождают одновременно 150—190 живых личинок в кожные ранки и на неповрежденные слизистые оболочки крупных млекопитающих и человека. Личинки активно проникают в ткани хозяина, питаясь ими и производя серьезные повреждения. Через несколько суток личинки покидают хозяина и окукливаются в почве.
Более совершенные приспособления к заражению хозяина имеются у другой мухи — Dermatobia hominis,
обитающей в тропической зоне Америки. Самки этого вида откладывают яйца на поверхность тела самок комаров и мух-жигалок. При нападении этих кровососущих насекомых на человека или домашних животных под действием повышенной температуры тела хозяина из яиц мух вылупляются личинки и проникают в ткани через неповрежденную кожу. Дальнейшее развитие происходит так же, как и у вольфартовой мухи.
Личинки некоторых других мух — комнатной, падальной
и серой мясной —
иногда также могут попадать в ткани, окружающие раны и царапины, вызывая тканевые миазы. Они же, попадая в пищеварительную систему с испорченной пищей и загрязненной водой, могут вызывать доброкачественные кишечные миазы, заканчивающиеся самоизлечением.
Своеобразна группа оводов
Oestridae
(рис. 21.17, Б
).
Половозрелые оводы вообще не питаются и даже лишены ротового аппарата. Яйца откладывают на кожу или слизистые оболочки лошадей, оленей или овец. Личинки различных оводов развиваются в подкожной клетчатке, в стенках желудка или в носовой полости и придаточных пазухах носа, в глазнице. С фекалиями или через кожные покровы личинки выходят во внешнюю среду и окукливаются в почве. Человек поражается оводами редко, и полного развития их личинок в его тканях не происходит, но иногда оводовые миазы приводят к серьезным поражениям лица, глаз и внутренних органов.
Основная профилактика
тканевых миазов — применение репеллентов при контакте со скотом, около которого обычно концентрируется большое количество двукрылых.
Рис. 21.17. Возбудители миазов. А —
вольфартова муха;Б —
овод подкожный:
а —
взрослое насекомое, б—
личинка
Несмотря на то что в настоящее время для медицины существенно большее значение имеют онкологические, сердечно-сосудистые и респираторные заболевания, паразитарные и зоонозные болезни продолжают оставаться серьезной медицинской проблемой. Причины этого кроются в ускоренной эволюции возбудителей, переносчиков, резер-вуарных хозяев и биогеоценозов в целом, в первую очередь под действием комплекса антропогенных экологических факторов.
Хозяйственная деятельность человека в ряде случаев приводит к созданию новых комплексов условий, являющихся более благоприятными для существования очагов зоонозных заболеваний по сравнению с естественной природой даже в условиях урбанизации. Так, экологические обследования, проведенные в крупных городах тропического пояса, таких, как Гавана, Манила, Рио-де-Жанейро и др., показали, что многочисленные внутренние дворики, заросшие разнообразной декоративной растительностью, крытые галереи, балконы и затеняющие навесы, многочисленные мелкие водоемы и фонтаны, увлажняющие воздух, создают особо благоприятные условия для существования и размножения комараAedes egypti —
основного переносчика вируса желтой лихорадки (рис. 22.1). При этом возбудитель начинает циркулировать между людьми, минуя основной природный резервуар — диких приматов.
Рис. 22.1. КомарAëdes aegypti
Таким образом возникают очаги заболевания городского типа. Анализ эпидемиологической обстановки городов Западной и Центральной Европы, а также СНГ показал, что в городских парках и пригородных зонах массового отдыха населения нередко создаются благоприятные условия для существования больших групп переносчиков и циркуляции различных возбудителей природно-очаговых и трансмиссивных заболеваний. Этому способствуют разнообразие ландшафтов, часто создающихся искусственно, богатство растительного покрова, наличие бродячих кошек и одичавших собак, искусственное привлечение в зоны отдыха диких животных — белок, оленей, лосей, лесных и водоплавающих птиц, являющихся кормовой базой кровососущих членистоногих.
Так, в большинстве городских и пригородных парков Праги обнаружены устойчивые популяции собачьего клещаIxodes ricinus.
В городских и пригородных стоячих водоемах таких городов, как Москва, выплаживаются комары, а в речках с быстрым течением (р. Уводь в г. Иванове, р. Салгир в г. Симферополе, р. Учан-Су в г. Ялта) — мошки сем. Simuliidae. Даже не будучи инвазированными патогенными для человека возбудителями, переносчики из этих популяций представляют для человека постоянную потенциальную опасность.
Большое значение для обогащения биогеоценозов паразитофауной, переносчиками трансмиссивных заболеваний, патогенными вирусами, бактериями и другими микроорганизмами имеет изменение хозяйственной деятельности человека в сельской местности. Террасирование горных склонов в тропических и субтропических странах с целью задержки воды и выращивания риса привело к расширению не только ареалов расселения комаров р.Anopheles,
но и к распространению территорий малярийных районов в высокогорья.. Такая ситуация произошла в Непале, Индонезии, в странах Индокитая.
Горное и степное отгонное животноводство резко увеличивает кормовую базу аборигенных кровососущих членистоногих и способствует появлению новых природных очагов трансмиссивных заболеваний. Широко известны природные очаги клещевого энцефалита в Болгарских Родопах и в Словенских Альпах, возникшие и поддерживающиеся благодаря сезонному выпасу коз и овец на горных пастбищах.
Нередко к появлению новых очагов приводит и изменение жизненного уклада населения. Переход жителей зоны влажных тропических лесов Африки от присваивающего к производящему типу хозяйствования: возникновение плантаций бананов, ананасов, кофе и пряностей на месте девственных лесов — часто сопровождается устойчивым повышением заболеваемости населения желтой лихорадкой. Это связано с улучшением условий обитания и размножения комараAëdes egypti
на сельскохозяйственных плантациях по сравнению с лесными биогеоценозами.
Ухудшению паразитологической обстановки может способствовать и деятельность человека по преобразованию ландшафтов. Так, широко известно возникновение новых очагов мочеполового шистосоматоза на берегах Асуанского водохранилища в результате того, что население пустыни, занимавшееся ранее овцеводством и верблюдоводством, стало активно контактировать с водой, выращивая овощи, рис и ловя рыбу. В прибрежных мелководных зонах водохранилища создались благоприятные условия для развития промежуточных хозяев шистосом — моллюсков родовPlanorbis, Bullinus
и др. Этих факторов оказалось достаточно для интенсивного заражения населения. Сходная ситуация возникла в бассейне р. Вольта в Западной Африке, где в результате ирригационных работ возникли слабопроточные водохранилища, вследствие чего участилась заболеваемость населения дракункулезом.
Расширению ареала распространения паразитических видов может способствовать создание человеком и некоторых промышленных объектов. Так, анкилостомиды, будучи представителями тропической и субтропической фауны, встречаются также за пределами зон теплого влажного климата в шахтах с высокой температурой и влажностью при условии загрязнения горных пород органическими веществами.
Широкие перемещения человека по территории планеты неоднократно приводили к заселению паразитами новых территорий и созданию новых очагов трансмиссивных заболеваний. Один из самых интересных примеров этого — обнаружение 23 марта 1930 г. в водоеме вблизи бразильского города Натала около 2000 личинок комараAnopheles gambiae,
не встречавшегося ранее в Америке. В Западной Африке этот вид является одним из основных переносчиков возбудителей малярии. На протяжении ближайших лет этот африканский комар размножился в Южной Америке, и уже в 1937 г. начались эпидемии малярии, когда заболело около 100 000 человек, причем пятая часть из них умерла.
Анализ ситуации показал, что в 1930 г. из Африки в Бразилию прибыл быстроходный миноносец французского военно-морского флота. Место стоянки этого корабля в Бразилии находилось в 1 км от места выплода первых комаров. Несомненно, несколько экземпляров A. gambiae
были занесены в Америку именно этим кораблем.
Современный способ перевозки грузов на любые расстояния в контейнерах предоставляет комарам и другим членистоногим необычайно благоприятные условия для расселения. Известна даже небольшая эпидемия малярии среди работников перегрузочного пункта парижского аэропорта Орли после открытия контейнеров, в которых находились зараженные комары. Выяснилось также, что имагинальные стадии насекомых легко переносят многочасовые перелеты на любые расстояния в салонах современных самолетов и, попав в благоприятные условия, могут образовывать новые популяции на большом отдалении от исходного ареала.
Еще большие возможности усложнения эпидемической ситуации связаны с теми, что паразиты и переносчики, как и любые объекты живой природы, способны эволюционировать, адаптируясь к меняющимся условиям. Наиболее простым примером является изменение биологии размножения некоторых видов комаров. Так, упоминавшийся уже несколько раз переносчик вирусов желтой лихорадки и лихорадки денге комарAëdes egypti,
размножающийся в природе в небольших стоячих водоемах и даже в дуплах деревьев, заполненных водой, в антропогенных условиях способен откладывать яйца и развиваться в старых консервных банках, в брошенных автопокрышках, в небольших лужах на полу, в подвальных и полуподвальных помещениях. Благодаря этому формируются особые синантропные популяции этого комара.
На строительстве Панамского канала в 1880—1888 гг. от желтой лихорадки погибло около 20 000 человек. Больных помещали в больницу, где ножки всех коек стояли в сосудах с водой, чтобы их не беспокоили ползающие насекомые. Эти сосуды оказались благоприятной средой для размножения комаровAëdes egypti,
которые получали возможность инфицироваться вирусом желтой лихорадки уже при первом кровососании. Таким образом, помещение больницы послужило дополнительным очагом распространения заболевания.
Другие виды комаров р. A
ë
des,
а также р.Culex
в настоящее время создают устойчивые городские популяции в странах с умеренным климатом. В отапливаемых помещениях эти комары могут размножаться не только летом, но и в холодные месяцы года, не покидая при этом подвалов, квартир и учреждений. Естественно, что переход к синантропному образу жизни этих видов мог произойти лишь на базе широкого генетического полиморфизма в популяциях, способных исходно размножаться в очень мелких затененных водоемах с сильно загрязненной водой и с малой двигательной активностью имагинальных стадий.
Вырубка девственных тропических зарослей, где в кронах деревьев обитают определенные виды птиц, летучих мышей, обезьян и кровососущих членистоногих, слабо связанных с наземными участками биогеоценозов, приводит либо к быстрой гибели всего комплекса животных верхнего яруса леса, либо к адаптациям отдельных видов к меняющимся условиям. При этом у паразитических насекомых и клещей часто оказывается больше шансов выжить, чем у специализированных птиц и млекопитающих, питающихся плодами, семенами и листвой, — они могут перейти к питанию кровью человека и домашних животных. Адаптивная эволюция возбудителей трансмиссивных заболеваний диких животных может при этом привести к возникновению у человека новых, ранее не известных болезней. Так, вероятно, вирус желтой лихорадки был распространен до заселения Америки человеком среди примитивных обезьян, ведущих древесный образ жизни, а переносчиком его были в основном комары из р.Haemagogus.
Освоение Америки человеком вызвало адаптации комаров р. Aedes
к питанию человеческой кровью, а затем способствовало приспособлению вируса к обитанию у новых видов комаров и у человека.
Заселение Австралии европейцами, сопровождавшееся бурным развитием овцеводства, привело к заселению этого континента и комплексом паразитов, связанных в цикле развития с овцами. Некоторые из них, в частности печеночный сосальщик, адаптировались и к новым промежуточным хозяевам (см. разд. 20.1.1.1).
Применение человеком инсектицидов и акарицидов является не менее эффективным фактором, обеспечивающим естественный отбор переносчиков, а следовательно, и возбудителей трансмиссивных заболеваний, чем создание благоприятных условий для их размножения и развития. Известно множество примеров возникновения популяций переносчиков, генетически устойчивых к действию ядохимикатов.
Увеличение контактов человека с природой и ее преобразование, в особенности в последнее время (освоение Сибири, девственных лесов бассейна Амазонки, высокогорий Центральной и пустынь Средней 297
Азии и Африки), процессы одомашнивания новых видов диких животных (пушные звери, декоративные птицы и рыбы, лабораторные млекопитающие и т. д.), а также обогащение естественных биогеоценозов за счет введения в них новых видов животных и растений могут способствовать активизации процессов адаптивной эволюции возбудителей паразитарных и переносчиков трансмиссивных заболеваний. Это может привести к увеличению количества паразитарных и инфекционных трансмиссивных и зоонозных заболеваний у человека.
Большую опасность для популяций человека представляют также паразиты, в разных зонах Земного шара использующие разных основных хозяев. Так, сосальщикMetagonimus yokogawai
хорошо известен как паразит человека в бассейне среднего течения р. Амур. Этот же вид описан на территории Румынии, где он паразитирует в кишечнике кошек, а человека не поражает. То же касается другого дальневосточного вида сосальщиковNanophyetes salmincola,
встречающегося также на территории Аляски и Северной Канады, где он известен как паразит рыбоядных животных. Еще один сосальщик,Eurytrema pancreaticum
(сосальщик поджелудочной железы), известен как возбудитель эуритрематоза человека в Юго-Восточной Азии, но широко распространен также в Средней Азии и Казахстане как паразит крупного рогатого скота (см. с. 242, 245).
Несомненно, что изменение экологической обстановки и связанная с этим эволюция соответствующих биогеоценозов, с одной стороны, могут привести к расширению круга основных хозяев этих паразитов и вовлечению в него человека, тем более что адаптации к обитанию в человеческом организме у этих паразитов в других климатических зонах уже имеются. С другой стороны, освоению этими паразитами человека в качестве хозяина может способствовать изменение характера питания с использованием экзотических продуктов или миграция населения. При этом в зоне нового обитания могут оказаться не только новые паразиты, но и жизненный уклад с традициями питания и гигиены, способствующими их циркуляции в новых условиях.
Ядовитость широко распространена в природе. Ядовитым
называют организм, в котором вырабатываются или накапливаются вещества, способные при попадании в другой организм вызывать нарушения его жизнедеятельности или смерть.
Среди животных ядовитость встречается у представителей почти всех систематических групп — от простейших до млекопитающих.
Ядовитые вещества животного происхождения называют зоотоксинами.
Они служат либо для защиты от нападений хищников, отпугивая или вызывая их гибель, либо для нападения на жертву. Ядовитые животные подразделяются на группы первично- и вторичноядовитых. Первичноядовитые
животные вырабатывают токсины в специализированных железах либо накапливают в тканях ядовитые метаболиты.
Первичная ядовитость является видовым признаком. Вторичноядовитые
аккумулируют экзогенные яды из окружающей среды. Токсичность этих животных проявляется только при поедании их другими животными. К ним относятся насекомые, личинки которых питаются на ядовитых растениях, а также двустворчатые моллюски и рыбы, накапливающие в своем теле токсины синезеленых водорослей (рис. 23.1).
Первичноядовитые животные по способам применяемого яда делятся на активно- и пассивноядовитых.
Активноядовитые
имеют специальные органы, вырабатывающие токсины. Если у таких животных есть приспособления для введения яда в тело жертвы минуя пищеварительный тракт, то их называют вооруженными.
Это ядовитые змеи, жалящие насекомые и т. п. (рис. 23.2).
У невооруженных
ядовитых животных ядовитые органы лишены ранящих приспособлений. Таковы большинство ядовитых земноводных, некоторые насекомые, рыбы. Ядовитые секреты этих животных обладают как местным контактным действием, раздражая кожные покровы и слизистые оболочки, так и общим эффектом, наступающим после их всасывания в кровь.
Ядовитые метаболиты в организме пассивноядовитых
видов накапливаются в различных органах (рис. 23.3).

Рис. 23.1. Вторичноядовитые животные. А —
мидия съедобная; Б —
устрица

Рис. 23.2. Активноядовитые животные. А
—медуза корнерот; Б—
паук каракурт; В—
оса восточная; Г—
скат хвостокол; Д—
морской ерш
  
Продолжение рис. 23.2.
Акгивноядовитые животные. Е—
гадюка обыкновенная; Ж—
эфапесчаная; З—
кобра среднеазиатская

Рис. 23.3. Пассивноядовитые животные.
А—
жук стафилин береговой; Б—
колорадский жук; В —
бахчевая коровка;
Г—
саламандра огненная; Д—
маринка балхашская
На ранних этапах эволюции в качестве ядов могли выступать нормальные метаболиты, выделяющиеся в окружающую среду или накапливающиеся в организме. Эволюционно наиболее примитивная форма ядовитости — накопление токсичных метаболитов в организме, . т. е. первичная пассивная ядовитость. У животных, не имеющих иных способов защиты или нападения, в соответствии с правилом прогрессивной специализации могут появляться специализированные структуры, продуцирующие яд: за счет усиления защитной функции покровных клеток (у кольчатых червей, кишечнополостных, иглокожих) или за счет активизации работы и расширения функций желез внешней или внутренней секреции. Так, ядовитые железы змей являются преобразованными слюнными железами. Ядовитый аппарат перепончатокрылых насекомых — видоизмененная половая система самок. Пассивная, а также вторичная ядовитость почти не обеспечивают индивидуальной защиты животных, но ценой гибели одной особи повышают эффективность выживания популяции в целом. Использование ядовитости в сочетании с наличием аппарата введения яда является несомненно прогрессивным признаком с морфофизиологической точки зрения.
Большинство активноядовитых животных вырабатывают яды, представляющие собой смеси токсических полипептидов и литических ферментов. Таковы яды змей и пауков. Их токсины по происхождению связаны с пищеварительными ферментами слюнных желез, обладающих повышенной литической активностью. Они действуют только при попадании их в кровь. При введении их в пищеварительную систему они расщепляются и теряют токсический эффект.
Невооруженные активноядовитые животные в большинстве случаев имеют яды небелковой природы. Поэтому они наиболее эффективно действуют при попадании в пищеварительную систему.
Хищнический образ жизни ядовитых животных сопровождается обычно возникновением вооруженной формы ядовитого аппарата, а токсины обладают нейротропным действием, направленным в первую очередь на обездвиживание жертвы (змеи, скорпионы, осы).
Нередко нейротоксины обладают строгой специфичностью действия, соответствующей пищевой специализации животного.
Так, яд наездников парализует практически лишь гусениц, которыми питаются личинки этих хищников. В яде скорпионов возникли видоспецифичные нейротоксины, избирательно действующие на млекопитающих, насекомых и ракообразных.
У невооруженных ядовитых животных яды в основном обладают отпугивающим действием. Химическая природа их разнообразна, что свидетельствует о разных путях их происхождения. Это стероиды, органические кислоты и т. п. Такие яды встречаются у амфибий, жуков, некоторых многоножек и др.
В процессе эволюции ядовитости параллельно с видоизменением токсинов, органов, их вырабатывающих, и механизмов введения в тело жертвы эволюционируют также и механизмы резистентности
животных — производителей токсинов к собственным ядам. Наиболее известный из них — особое строение ядопродуцирующих желез, стенки которых препятствуют распространению токсинов по организму.
Нередко яды вырабатываются вместе с их ингибиторами и поэтому в таком виде не проявляют специфической активности.
При попадании в организм другого вида концентрация ингибиторов токсинов резко падает и ферменты активируются. Существуют и гуморальные антитоксические механизмы. Так, в крови некоторых змей циркулируют пептиды, инактивирующие токсическое действие ядов, а у ряда ядовитых амбифий и рыб мембраны клеток не имеют рецепторов к собственным токсинам, при этом клетки становятся нечувствительными к ним.
В результате обширной хозяйственной деятельности человека происходит исчезновение большого количества видов животных, причем не только хорошо известных, но и практически не изученных. В первую очередь это касается ядовитых форм, представляющих прямую или косвенную опасность для человека. Они подвергаются прямым преследованиям и уничтожаются, но, несомненно, большее значение имеют разрушение биотопов, в которых они обитают, и обеднение биогеоценозов, членами которых они являются.
Конкретной причиной вымирания отдельных видов ядовитых животных может быть действие разных факторов. Так, сокращение численности змей объясняется в первую очередь их прямым уничтожением, отловом и мелиоративными работами в зонах их обитания. Ядовитые насекомые исчезают большей частью за счет широкого применения пестицидов в сельском хозяйстве и в результате замены естественных экологических систем монокультурными сельскохозяйственными предприятиями, на полях и пастбищах которых они не находят благоприятных условий существования.
Ядовитые морские животные обычно не уничтожаются специально, но вместе с другими организмами в массовом количестве гибнут в результате загрязнения прибрежных зон моря токсичными отходами промышленности, а также при попадании в сети при промысловой ловле рыбы.
Все ядовитые животные, несомненно, нуждаются в охране не только как компоненты соответствующих биогеоценозов, устойчивость и эволюция которых зависят, в частности, и от них, но и как организмы, имеющие огромное медицинское и хозяйственное значение, несмотря на то что большинство их видов до сих пор не изучено.
Зоотоксины ряда видов животных используют как ценное сырье для фармацевтической промышленности и применяют для изготовления многих лекарственных препаратов. Некоторые токсины — источники химических реактивов. Важной областью применения зоотоксинов является производство сывороток, служащих для лечения отравлений.
Многие зоотоксины применяют в медико-биологических научных исследованиях, в результате которых выявляются их не известные ранее свойства. Так, в последние годы стало известно, что некоторые компоненты ядовитых секретов жаб обладают противоопухолевым свойством, яды иглокожих подавляют синтез нуклеиновых кислот и регулируют активность фермента АТФазы, а также транспорт Са2+
через мембраны клеток.
Некоторые виды ядовитых змей, скорпионов, пауков и насекомых содержатся в специальных заведениях — серпентариях, скорпионариях и инсектариях, где их используют для регулярного получения токсинов. В питомниках такого рода проводят планомерные токсикологические исследования и изучение биологии животных, разрабатывают способы их длительного содержания и размножения в неволе. Успехи в этой области должны способствовать восстановлению и сохранению природных биогеоценозов, компонентами которых являются ядовитые животные. Сохранению видов животных — поставщиков токсинов — должно служить также тщательное изучение химического состава их ядов и налаживание химического синтеза наиболее ценных из них, а также производство их искусственных аналогов.
Термин «биосфера» введен австрийским геологом Э. Зюссом в 1875 г. для обозначения особой оболочки Земли, образованной совокупностью живых организмов, что соответствует биологической концепции биосферы.
В указанном смысле названный термин используют ряд исследователей и в настоящее время.
Представление о широком влиянии живых существ на протекающие в природе процессы было сформулировано В.В. Докучаевым, который показал зависимость процесса почвообразования не только от климата, но и от совокупного влияния растительных и животных организмов.
В. И Вернадский развил это направление и разработал учение о биосфере как глобальной системе нашей планеты,
в которой основной ход геохимических и энергетических превращений определяется живым веществом. Он распространил понятие биосферы не только на сами организмы, но и на среду их обитания, чем придал концепции биосферы биогеохимический
смысл. Большинство явлений, меняющих в масштабе геологического времени облик Земли, рассматривали ранее как чисто физические, химические или физико-химические (размыв, растворение, осаждение, выветривание пород и т. д.). В.И. Вернадский создал учение о геологической роли живых организмов
и показал, что деятельность последних представляет собой важнейший фактор преобразования минеральных оболочек планеты.
С именем В.И. Вернадского связано также формирование социально-экономической концепции биосферы,
отражающей ее превращение на определенном этапе эволюции в ноосферу
(см. гл. 25) вследствие деятельности человека, которая приобретает роль самостоятельной геологической силы. Учитывая системный принцип организации биосферы, а также то, что в основе ее функционирования лежат круговороты веществ и потоки энергии, современной наукой сформулированы биохимическая, термодинамическая, биогеоценотическая, кибернетическая концепции биосферы.
Биосферой называют оболочку Земли, которая населена и активно преобразуется живыми существами. Согласно В.И. Вернадскому, биосфера — это такая оболочка, в которой существует или существовала в прошлом жизнь и которая подвергалась или подвергается воздействию живых организмов. Она включает: 1) живое вещество, образованное совокупностью организмов; 2) биогенное вещество, которое создается и перерабатывается в процессе жизнедеятельности организмов (газы атмосферы, каменный уголь, нефть, сланцы, известняки и др.); 3) косное вещество, которое образуется без участия живых организмов (продукты тектонической деятельности, метеориты); 4) биокосное вещество, представляющее собой совместный результат жизнедеятельности организмов и абиогенных процессов (почвы).
Биосфера представляет собой многоуровневую систему, включающую подсистемы различной степени сложности. Границы биосферы определяются областью распространения организмов в атмосфере, гидросфере и литосфере (рис. 24.1). Верхняя граница биосферы проходит примерно на высоте 20 км. Таким образом, живые организмы расселены в тропосфере и в нижних слоях стратосферы. Лимитирующим фактором расселения в этой среде является нарастающая с высотой интенсивность ультрафиолетовой радиации. Практически все живое, проникающее выше озонового слоя атмосферы, погибает. В гидросферу биосфера проникает на всю глубину Мирового океана, что подтверждает обнаружение живых организмов и органических отложений до глубины 10—11 км. В литосфере область распространения жизни во многом определяет уровень проникновения воды в жидком состоянии — живые организмы обнаружены до глубины примерно 7,5 км.
Атмосфера.
Эта оболочка состоит в основном из азота и кислорода. В меньших концентрациях она содержит углекислый газ и озон. Состояние атмосферы оказывает большое влияние на физические, химические и особенно биологические процессы на земной поверхности и в водной среде. Наибольшее значение для биологических процессов имеют кислород атмосферы, используемый для дыхания организмов и минерализации омертвевшего органического вещества, углекислый газ, расходуемый при фотосинтезе, а также озон, экранирующий земную поверхность от жесткого ультрафиолетового излучения. Вне атмосферы существование живых организмов невозможно. Это видно на примере лишенной жизни Луны, у которой нет атмосферы. Исторически развитие атмосферы связано с геохимическими процессами, а также жизнедеятельностью организмов. Так, азот, углекислый газ, пары воды образовались в процессе эволюции планеты благодаря (в значительной мере) вулканической активности, а кислород — в результате фотосинтеза.
Гидросфера.
Вода является важной составной частью всех компонентов биосферы и одним из необходимых факторов существования живых организмов. Основная ее часть (95%) заключена в Мировом океане, который занимает примерно 70% поверхности Земного шара. Общая масса океанических вод составляет свыше 1300 млн. км3
. Около 24 млн. км3
воды содержится в ледниках, причем 90% этого объема приходится на ледяной покров Антарктиды. Столько же воды содержится под землей. Поверхностные воды озер составляют приблизительно 0,18 млн. км3
(из них половина соленые), а рек—0,002 млн. км3
.
Количество воды в телах живых организмов достигает примерно 0,001 млн. км3
. Из газов, растворенных в воде, наибольшее значение имеют кислород и углекислый газ. Количество кислорода в океанических водах изменяется в широких пределах в зависимости от температуры и присутствия живых организмов. Концентрация углекислого газа также варьирует, а общее количество его в океане в 60 раз превышает его содержание в атмосфере. Гидросфера формировалась в связи с развитием литосферы, выделившей за геологическую историю Земли значительный объем водяного пара и так называемых ювенильных (подземных магматических) вод.
Рис. 24.1. Область распространения организмов в биосфере:
1
—уровень озонового слоя, задерживающего жесткое ультрафиолетовое излучение, 2—
граница снегов, 3—
почва, 4—
животные, обитающие в пещерах, 5—
бактерии в нефтяных скважинах
Литосфера.
Основная масса организмов, обитающих в пределах литосферы, сосредоточена в почвенном слое, глубина которого обычно не превышает нескольких метров. Почвы, будучи, по терминологии В.И. Вернадского, биокосным веществом, представлены минеральными веществами, образующимися при разрушении горных пород, и органическими веществами — продуктами жизнедеятельности организмов.
Живые организмы (живое вещество).
В настоящее время описано около 300 тыс. видов растений и более 1,5 млн. видов животных. Из этого количества 93% представлено сухопутными, а 7% — водными видами животных. Суммарная биомасса организмов сухопутных видов образована на 99,2% зелеными растениями (2,4 • 1012
т) и на 0,8% животными и микроорганизмами (0,2 • 1011
т). В океане, напротив, на долю растений приходится 6,3% (0,2 • 109
т), а на долю животных и микроорганизмов — 93,7% (0,3 • 1010
т) совокупной биомассы. Несмотря на то что океан покрывает немногим более 70% поверхности планеты, в нем содержится лишь 0,13% биомассы всех живых существ, обитающих на Земле.
Расчеты показывают, что растения составляют около 21% всех учтенных видов. Однако на их долю приходится более 99% биомассы, тогда как вклад животных в биомассу планеты (79% видов) составляет менее 1%. Среди животных 96% видов приходится на долю беспозвоночных и только 4% на долю позвоночных, среди которых млекопитающие составляют примерно 10%.
Приведенные соотношения иллюстрируют фундаментальную закономерность организации биосферы: в количественном отношении преобладают формы, достигшие в процессе эволюции относительно низких степеней морфофизиологического прогресса.
Живое вещество по массе составляет 0,01—0,02% от косного вещества биосферы, однако играет ведущую роль в биогеохимических процессах благодаря совершающемуся в живых организмах обмену веществ. Так как субстраты и энергию, используемые в обмене веществ, организмы черпают из окружающей среды, они преобразуют ее уже тем, что в процессе своего существования используют ее компоненты.
Ежегодная продукция живого вещества в биосфере составляет 232,5 млрд. т сухого органического вещества. За это же время в масштабе планеты в процессе фотосинтеза синтезируется 46 млрд. тонн органических углеродсодержащих веществ. Для этого требуется, чтобы 170 • 109
т С02
прореагировало с 68 • 109
т Н2
0.
Таким образом, в результате фотосинтеза ежегодно образуется 115х х 109
т сухого органического вещества и 123 • 109
т 02. В течение года в процесс фотосинтеза вовлекаются также 6 • 109
т азота, 2 • 109
т фосфора и другие элементы, например калий, кальций, сера, железо. Приведенные цифры показывают, что живое вещество является наиболее активным компонентом биосферы. Оно производит гигантскую геохимическую работу, способствуя преобразованию других оболочек Земли в геологическом масштабе времени.
Биотический круговорот.
Главная функция биосферы заключается в обеспечении круговоротов химических элементов. Глобальный биотический круговорот осуществляется при участии всех населяющих планету организмов. Он заключается в циркуляции веществ между почвой, атмосферой, гидросферой и живыми организмами. Благодаря биотическому круговороту возможно длительное существование и развитие жизни при ограниченном запасе доступных химических элементов. Используя неорганические вещества, зеленые растения за счет энергии Солнца создают органическое вещество, которое другими живыми существами (гетеротрофами — потребителями и деструкторами) разрушается, с тем чтобы продукты этого разрушения могли быть использованы растениями для новых органических синтезов.
Важная роль в глобальном круговороте веществ принадлежит циркуляции воды между океаном, атмосферой и верхними слоями литосферы. Вода испаряется и воздушными течениями переносится на многие километры. Выпадая на поверхность суши в виде осадков, она способствует разрушению горных пород, делая их доступными для растений и микроорганизмов, размывает верхний почвенный слой и уходит вместе с растворенными в ней химическими соединениями и взвешенными органическими частицами в океаны и моря. Подсчитано, что с поверхности Земли за 1 мин испаряется около 1 млрд. т Н2
0 (на образование 1 г водяного пара необходимо 2,248 кДж). Энергия, затрачиваемая на испарение воды, возвращается в атмосферу (рис. 24.2). Циркуляция воды между Мировым океаном и сушей представляет собой важнейшее звено в поддержании жизни на Земле и основное условие взаимодействия растений и животных с неживой природой.
Рис. 24.2. Круговорот воды в биосфере
Под влиянием этого процесса происходит постепенное разрушение литосферы, перенос ее компонентов в глубины морей и океанов.
На создание органического вещества расходуется всего 0,1—0,2% солнечной энергии, достигающей поверхности планеты. Благодаря этой энергии осуществляется значительный объем работы по перемещению химических элементов.
В качестве примеров биотического круговорота рассмотрим круговороты углерода и азота в биосфере (рис. 24.3; 24.4). Круговорот углерода
начинается с фиксации атмосферного диоксида углерода в процессе фотосинтеза. Часть образовавшихся при фотосинтезе углеводов используют сами растения для получения энергии, часть потребляется животными. Углекислый газ выделяется в процессе дыхания растений и животных. Мертвые растения и животные разлагаются, углерод их тканей окисляется и возвращается в атмосферу. Аналогичный процесс происходит и в океане.
Круговорот азота
также охватывает все области биосферы (рис. 24.4). Хотя его запасы в атмосфере практически неисчерпаемы, высшие растения могут использовать азот только после соединения его с водородом или кислородом. Исключительно важную роль в этом процессе играют азотфиксирующие бактерии. При распаде белков этих микроорганизмов азот снова возвращается в атмосферу.
Показателем масштаба биотического круговорота служат темпы оборота углекислого газа, кислорода и воды. Весь кислород атмосферы проходит через организмы примерно за 2 тыс. лет, углекислый газ — за 300 лет, а вода полностью разлагается и восстанавливается в биотическом круговороте за 2 млн. лет (рис. 24.5).
Рис. 24.3. Круговорот углерода в биосфере
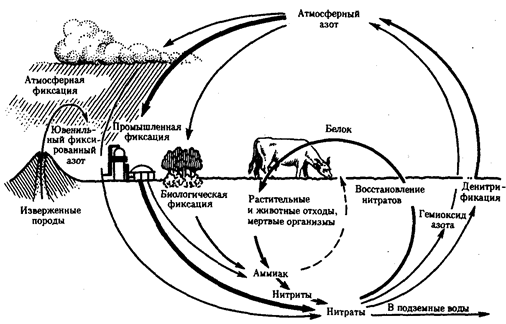
Рис. 24.4. Круговорот азота в биосфере
Благодаря биотическому круговороту биосфере присущи определенные геохимические функции: газовая —
биогенная миграция газов в результате фотосинтеза и азотфиксации; концентрационная —
аккумуляция в своих телах живыми организмами химических элементов, рассеянных во внешней среде; окислительно-восстановительная —
превращение веществ, содержащих атомы с переменной валентностью (например, Fе, Mn); биохимическая —
процессы протекающие в живых организмах.
Стабильность биосферы.
Биосфера представляет собой сложную экологическую систему, работающую в стационарном режиме. Стабильность биосферы обусловлена тем, что результаты активности трех групп организмов, выполняющих разные функции в биотическом круговороте,— продуцентов
(автотрофы), потребителей
(гетеротрофы) и деструкторов
(минерализующие органические остатки) — взаимоуравновешиваются. То, что в биосфере поддерживается постоянство ее главных характеристик (гомеостаз), не исключает способности ее к эволюции.
Рис. 24.5. Темпы циркуляции веществ в биосфере
Эволюция биосферы на Протяжении большей части ее истории осуществлялась под влиянием двух главных факторов:
естественных геологических и климатических изменений на планете и изменений видового состава и количества живых существ в процессе биологической эволюции. На современном этапе в третичном периоде к ним присоединился третий фактор —
развивающееся человеческое общество.
Этапы возникновения жизни, пути и механизмы ее эволюционного развития рассмотрены выше (см. гл. 1). Жизнь зародилась на Земле свыше 3,5 млрд. лет назад. Первыми живыми существами были анаэробы, которые получали энергию путем брожения. Так как брожение представляет собой относительно малопродуктивный способ энергообеспечения, примитивная жизнь не могла эволюционировать далее одноклеточной формы организации. Питание таких примитивных организмов зависело от опускавшихся на дно водоемов органических веществ, синтезируемых в поверхностных слоях воды абиогенным способом.
Недостаток органических веществ создал давление отбора, приведшее к возникновению фотосинтеза. Прогрессивное увеличение кислорода в воде за счет жизнедеятельности фотосинтезирующих организмов и его диффузии в атмосферу вызвало изменения в химическом составе оболочек Земли, прежде всего атмосферы, что в свою очередь сделало возможным и развитие более сложно организованных живых форм и быстрое распространение Жизни по планете. По мере увеличения содержания кислорода в атмосфере формируется достаточно мощный слой озона, защищающий поверхность Земли от проникновения жесткого ультрафиолетового излучения. В таких условиях жизнь смогла продвинуться к поверхности моря. Развитие механизма аэробного дыхания сделало возможным появление многоклеточных организмов. Примечательно, что первые такие организмы появились после того, как концентрация кислорода в атмосфере планеты достигла примерно 3%, что произошло около 600 млн. лет назад (начало кембрия).
Благодаря способности фотосинтезирующих морских организмов продуцировать такое количество кислорода, которое превышало потребности в нем обитателей планеты, стало возможным возникновение в процессе эволюции организмов более высокого уровня структурно-физиологической организации, их широкое расселение и проникновение Жизни в различные сферы обитания. В течение палеозойской эры живые существа не только заселили все моря, но и вышли на сушу. Развитие зеленых растений обеспечило образование больших количеств кислорода и органических веществ, что создавало благоприятные условия для последующей прогрессивной эволюции.
В середине палеозоя темпы потребления кислорода живыми организмами и расход его в абиотических процессах, а также темпы его образования сравнялись. Содержание кислорода в атмосфере начиная с этого периода истории Земли стабилизировалось на уровне примерно 20%.
С появлением человеческого общества в развитии биосферы намечается переход от биогенеза, обусловленного факторами биологической эволюции, к ноогенезу — развитию под влиянием разумной созидательной деятельности человечества.
Эволюция органического мира прошла несколько этапов. Первый из них — возникновение первичной биосферы с биотическим круговоротом, второй — усложнение структуры биотического компонента биосферы в результате появления многоклеточных организмов. Эти два этапа, осуществлявшиеся в связи с чисто биологическими закономерностями жизнедеятельности и развития, могут быть объединены в период биогенеза.
Третий этап связан с возникновением человеческого общества. Разумная по своим намерениям деятельность людей в масштабе биосферы способствует превращению последней в ноосферу. На рассматриваемом этапе эволюция происходит под определяющим воздействием человеческого сознания в процессе производственной (трудовой) деятельности людей, что свойственно периоду ноогенеза.
Понятие «ноосфера» было введено в науку французским философом Э. Леруа (1927). Ноосферой Леруа назвал оболочку Земли, включающую человеческое общество с его языком, индустрией, культурой и прочими атрибутами разумной деятельности.
Ноосфера, по мнению Э. Леруа, представляет собой «мыслящий пласт», который, зародившись в конце третичного периода, разворачивается с тех пор над миром растений и животных вне биосферы и над ней.
В противоположность приведенной трактовке В.И. Вернадский представляет ноосферу не как нечто внешнее по отношению к биосфере, а как новый этап в развитии биосферы, заключающийся в разумном регулировании отношений человека и природы.
Науку управления взаимоотношениями между человеческим обществом и природой можно назвать ноогеникой.
Основная цель ноогеники — планирование настоящего во имя будущего, а ее главная задача — исправление нарушений в отношениях человека и природы, вызванных прогрессом техники.
Помимо охранных функций ноогеника должна способствовать увеличению многообразия форм жизни путем создания новых видов растений, животных и микроорганизмов. Эти новые виды призваны не только служить источником пищи, кислорода, сырья для промышленности, но и помогать человеку, осуществляя буферные функции, бороться с вредными побочными результатами технического прогресса, способствовать еще более активному освоению неживой природы, сопровождать человека в космических полетах.
Таким образом, ноогеника не ставит целью достижение какого-то постоянного равновесия между человеком и природой, которое в принципе неосуществимо.
На начальных этапах существования человеческого общества интенсивность воздействия людей на среду обитания не отличалось от воздействия других организмов. Получая от окружающей среды средства к существованию в таком количестве, которое полностью восстанавливалось за счет естественных процессов биотического круговорота, люди возвращали в биосферу то, что использовали другие организмы для своей жизнедеятельности. Универсальная способность микроорганизмов разрушать органическое вещество, а растений — превращать минеральные вещества в органические обеспечивала включение продуктов хозяйственной деятельности людей в биотический круговорот.
В настоящее время человек извлекает из биосферы сырье в значительном и все возрастающем количестве, а современные промышленность и сельское хозяйство производят или применяют вещества, не только не используемые другими видами организмов, но нередко и ядовитые. В результате этого биотический круговорот становится незамкнутым. Вода, атмосфера, почвы загрязняются отходами производства, вырубаются леса, истребляются дикие животные, разрушаются природные биогеоценозы.
Нежелательные последствия неконтролируемой человеческой деятельности осознавали естествоиспытатели уже в конце XVIII — начале XIX в. (Ж.-Л.-Л. Бюффон; Ж.-Б. Ламарк). В настоящее время человечество стоит перед возможностью экологического кризиса,
т. е. такого состояния среды обитания, которое вследствие произошедших в ней изменений оказывается непригодным для жизни людей. Ожидаемый кризис по своему происхождению является антропогенным, так как. к нему ведут изменения в природе Земли, развивающиеся в связи с воздействием на нее человека.
По своим последствиям воздействия человеческого общества на среду обитания могут быть положительными и отрицательными. Последние особо привлекают к себе внимание. Основные пути воздействия людей на природу заключаются в расходовании естественных богатств в виде минерального сырья, почв, водных ресурсов; загрязнении среды, истреблении видов, разрушении биогеоценозов.
Естественные богатства планеты делятся на невосполняемые и восполняемые. К первым, например, относят полезные ископаемые, запасы которых ограничены. Тенденцию в изменениях восполняемых природных ресурсов можно проследить на примере леса. В настоящее время лесом покрыта примерно треть суши (без Антарктиды), тогда как в доисторические времена им было занято не менее 70%. Особенно пострадал лес в районах древних цивилизаций. Оголение горных склонов Ливана началось 5000 лет назад, когда по приказу царя Соломона 80 000 дровосеков вырубили для строительства дворца и храмов рощи ливанских кедров на значительной территории. Густые леса Далмации начали интенсивно уничтожать при создании римского флота, а затем при строительстве Венеции. На большей части территории Китая и Индии леса были почти полностью вырублены уже в позапрошлом тысячелетии.
Уничтожение лесов прежде всего резко нарушает водный режим планеты. Мелеют реки, их дно покрывается илом, что приводит в свою очередь к уничтожению нерестилищ и сокращению численности рыб. Уменьшаются запасы грунтовых вод, создается недостаток влаги в почве. Талая вода и дождевые потоки смывают, а ветры, не сдерживаемые лесной преградой, выветривают почвенный слой. В результате возникает эрозия почвы. Древесина, ветви, кора, подстилка аккумулируют минеральные элементы питания растений. Уничтожение лесов ведет к вымыванию этих элементов из почв и, следовательно, падению ее плодородия. С вырубкой лесов гибнут населяющие их птицы, звери, насекомые-энтомофаги. Вследствие этого беспрепятственно размножаются вредители сельскохозяйственных культур. Лес очищает воздух от ядовитых загрязнений, в частности он задерживает радиоактивные осадки и препятствует их дальнейшему распространению, т. е. вырубка лесов устраняет важный компонент самоочищения воздуха. Наконец, уничтожение лесов на склонах гор является существенной причиной образования оврагов и селевых потоков.
Таким образом, из-за нерационального землепользования человечество потеряло вследствие эрозии почв обширные территории, ставшие практически непригодными для земледелия. Так, за период, равный примерно 150 годам, в США эрозия привела к резкому снижению плодородия почв на территории, равной 120 млн. га.
Промышленные отходы, пестициды, применяемые для борьбы с вредителями сельскохозяйственных культур, радиоактивные вещества, образуемые, в частности, при испытании ядерного и термоядерного оружия, загрязняют природную среду. Так, только автомобили в крупных городах за год выбрасывают в атмосферу около 50 млн. м3
угарного газа, кроме того, каждый автомобиль ежегодно выделяет около 1 кг свинца. Обнаружено, что в организме людей, проживающих вблизи крупных магистралей, содержание свинца повышено. Из-за бесконтрольного использования количество инсектицида ДДТ (по данным 50-х годов), находящегося в распыленном состоянии на обширных территориях планеты и вредного для живых организмов, достигло 1 млн. т. Троекратная обработка в 60-е годы инсектицидом, сходным с ДДТ, района живописного оз. Клири в США привела к почти полному уничтожению популяции гагар, достигавшей 1000 пар. В среднем в организме жителя США в 1961 г. обнаружено около 925 мг хлорорганических соединений.
Считая с 1600 г., человеком было истреблено более 160 видов и подвидов птиц и около 100 видов млекопитающих. В настоящее время около 600 видов позвоночных животных находятся на грани полного истребления. К ним относятся киты, австралийские сумчатые (кенгуру), крокодилы, бегемоты, носороги, ряд крупных хищников.
Отдельные виды животных исчезают не только в результате их непосредственного истребления человеком. Между естественными и искусственными биоценозами все время идет борьба за территорию. Но человеческий труд оказывается фактором настолько мощным, что искусственные биоценозы, сами по себе малоустойчивые, тем не менее теснят естественные биоценозы.
Деятельность человека изменяет структуру земной поверхности, отчуждая под сельскохозяйственные угодья, строительство населенных пунктов, коммуникаций, водохранилищ территорию, занимаемую природными биогеоценозами. К настоящему времени указанным образом преобразовано около 20% суши.
К числу отрицательных влияний относится нерегулируемый промысел рыбы, млекопитающих, беспозвоночных, водорослей, изменение химического состава вод, воздуха, почвы в результате сбросов отходов промышленности, транспорта и сельскохозяйственного производства.
Положительное влияние человека выражается в выведении новых пород домашних животных и сортов сельскохозяйственных растений, создании культурных биогеоценозов, а также в разработке новых штаммов полезных микроорганизмов как основы микробиологической промышленности, развитии прудового рыбного хозяйства, интродукции полезных видов в новых условиях обитания.
Прогнозы будущего человечества с учетом экологических проблем, стоящих перед ним, представляют непосредственный интерес для всего населения планеты. По мнению экспертов, экологическая ситуация, складывающаяся на Земле, таит в себе опасность серьезных и, возможно, необратимых нарушений биосферы в том случае, если деятельность человечества не приобретет планомерный, согласующийся с законами существования и развития биосферы характер. Вместе с тем расчеты показывают, что человеческое общество не использует значительные резервы биосферы.
Одной из наиболее острых проблем современности является проблема быстрого роста населения Земли. Ежегодный прирост населения в абсолютном исчислении достигает 60—70 млн. человек, или примерно 2%. К 2000 г. численность населения достигла 6 млрд. человек. Площадь поверхности суши на планете равна 1,5 • 1014
м2
, что достаточно для размещения 15—20 млрд. человек со средней плотностью 300—400 человек на 1 км2
, имеющей место в настоящее время в Бельгии, Нидерландах, Японии.
Растущее население Земли должно быть обеспечено пищей. Известно, что производство продовольствия на душу населения растет медленнее, чем производство энергии, одежды, различных материалов. Многие миллионы людей в слаборазвитых странах испытывают нехватку продуктов. Вместе с тем из всей территории суши, пригодной для земледелия, в среднем по Земному шару под сельскохозяйственные угодья занято лишь 41%. При этом на используемой территории, по мнению разных экспертов, получают от 3—4 до 30% от возможного при современном уровне развития агротехники количества продуктов. Причины этого отчасти заключаются в недостаточной энерговооруженности сельского хозяйства. Так, в Японии, получая урожай, в 5 раз больший, чем в Индии (с 1 га сельскохозяйственных угодий), затрачивают в 20 раз больше электроэнергии и в 20—30 раз больше удобрений и пестицидов.
Уже сейчас 30% металлоизделий изготовляют из вторичного сырья. При существующей технологии из месторождений нефти извлекается лишь 30—50% запасов. Выход полезных ископаемых, таким образом, может быть увеличен путем разработки прогрессивных способов добычи. Около 95% энергии получают в настоящее время за счет сжигания ископаемого топлива, 3—4% за счет энергии речного стока и только 1—2% за счет атомного горючего. Использование атомной энергии в мирных целях решает проблему энергетического кризиса.
Преобразующая деятельность людей неизбежна, так как с ней связано благосостояние населения. Современное человечество располагает исключительно мощными факторами воздействия на природу планеты, которое по своим последствиям может быть и отрицательным, и положительным. Следование принципу научно обоснованного рационального природопользования
позволяет получить в целом позитивный итог.
Алексеева Т. И.
Адаптивные процессы в популяциях человека. М., 1986.
Бигон М., Харпер Дж., Таунсенд К.
Экология. Особи, популяции, сообщества. М., 1989. Т. 1, 2.
Гинецинская Т.А., Добровольский А.А.
Частная паразитология. М., 1978. Т. 1,2.
Дозье Т.
Опасные морские создания. М., 1985.
Заварзин А.А.
Основы сравнительной гистологии. Л., 1985.
Казначеев В. П.
Учение В. И. Вернадского о биосфере и ноосфере. Новосибирск,1989.
Крумбигель Г., Вольтер X.
Ископаемые. М., 1988.
Курс зоологии / Под ред. Б. С. Матвеева.
М., 1961. Т. 1, 2.
Левонтин Р.
Генетические основы эволюции. М., 1978.
Левушкин С.И., Шилов И.А.
Общая зоология. М., 1994.
Майр Э.
Популяции, виды и эволюция. М., 1974.
Орлов Б.Н., Гелашвили Д.Б., Ибрагимов А.К.
Ядовитые животные и растения СССР. М., 1990.
Павловский Е.Н.
Руководство по паразитологии человека. М., 1948. Т. 1, 2.
Паразитология человека / Под ред. В.Я. Подоляна.
Л., 1974. Патологическая анатомия болезней плода и ребенка / Под ред. Т.Е.
Ивановой
и Л.В. Леоновой.
М., 1989. Т. 1, 2.
Рогинский Я.Я., Левин М.Г.
Антропология. М., 1978.
Ромер А., Пирсоне Т.
Анатомия позвоночных. М., 1992. Т. 1, 2.
Рэфф Р., Кофмен Т.
Эмбрионы, гены и эволюция. М., 1986.
Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В.
Краткий очерк теории эволюции. М., 1987.
Фоули Р.
Еще один неповторимый вид. Экологические аспекты эволюции человека. М., 1990.
Шмальгаузен И.И.
Основы сравнительной анатомии. М., 19Э5.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ............................................................................................... 2
РАЗДЕЛ IV . ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ.............................................................................................................. 3
ГЛАВА 10. БИОЛОГИЧЕСКИЙ ВИД. ПОПУЛЯЦИОННАЯ СТРУКТУРА ВИДА.................................................................................................................. 4
10.1. ПОНЯТИЕ О ВИДЕ.................................................................................. 4
10.2. ПОНЯТИЕ О ПОПУЛЯЦИИ................................................................... 5
10.2.1. Экологическая характеристика популяции................................... 6
10.2.2. Генетические характеристики популяции...................................... 7
10.2.3. Частоты аллелей. Закон Харди — Вайнберга............................... 7
10.2.4. Место видов и популяций в эволюционном процессе................... 9
ГЛАВА 11....... . ВИДООБРАЗОВАНИЕ В ПРИРОДЕ. ЭЛЕМЕНТАРНЫЕ ЭВОЛЮЦИОННЫЕ ФАКТОРЫ................................................................. 11
11.1. МУТАЦИОННЫЙ ПРОЦЕСС............................................................... 11
11.2. ПОПУЛЯЦИОННЫЕ ВОЛНЫ.............................................................. 12
11.3. ИЗОЛЯЦИЯ............................................................................................ 14
11.4. ЕСТЕСТВЕННЫЙ ОТБОР.................................................................... 17
11.5. ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ (ДРЕЙФ ГЕНОВ).. 21
11.6. ВИДООБРАЗОВАНИЕ.......................................................................... 22
11.7. НАСЛЕДСТВЕННЫЙ ПОЛИМОРФИЗМ ПРИРОДНЫХ
ПОПУЛЯЦИЙ. ГЕНЕТИЧЕСКИЙ ГРУЗ....................................................... 24
11.8. АДАПТАЦИИ ОРГАНИЗМОВ К СРЕДЕ ОБИТАНИЯ...................... 27
11.9. ПРОИСХОЖДЕНИЕ БИОЛОГИЧЕСКОЙ ЦЕЛЕСООБРАЗНОСТИ. 29
ГЛАВА 12 . ДЕЙСТВИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ В ПОПУЛЯЦИЯХ ЛЮДЕЙ.......................................................................... 32
12.1. ПОПУЛЯЦИЯ ЛЮДЕЙ. ДЕМ, ИЗОЛЯТ.............................................. 32
12.2. ВЛИЯНИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ
НА ГЕНОФОНДЫ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ.................................. 33
12.2.1. Мутационный процесс.................................................................. 34
12.2.2. Популяционные волны................................................................. 35
12.2.3. Изоляция........................................................................................ 36
12.2.4. Генетико-автоматические процессы............................................. 38
12.2.5. Естественный отбор...................................................................... 41
12.3. ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ В ПОПУЛЯЦИЯХ ЛЮДЕЙ..... 45
12.4. ГЕНЕТИЧЕСКИЙ ГРУЗ В ПОПУЛЯЦИЯХ ЛЮДЕЙ.......................... 50
ГЛАВА 13. ЗАКОНОМЕРНОСТИ МАКРОЭВОЛЮЦИИ........................ 51
13.1. ЭВОЛЮЦИЯ ГРУПП ОРГАНИЗМОВ.................................................. 52
13.1.1. Уровень организации.................................................................... 52
13.1.2. Типы эволюции групп................................................................... 52
13.1.3. Формы эволюции групп............................................................... 55
13.1.4. Биологический прогресс и биологический регресс..................... 56
13.1.5. Эмпирические правила эволюции групп..................................... 60
13.2. СООТНОШЕНИЕ ОНТО- И ФИЛОГЕНЕЗА....................................... 61
13.2.1. Закон зародышевого сходства..................................................... 61
13.2.2. Онтогенез — повторение филогенеза........................................... 62
13.2.3. Онтогенез — основа филогенеза.................................................. 63
13.3. ОБЩИЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ОРГАНОВ.................. 67
13.3.1. Дифференциация и интеграция в эволюции органов.................. 68
13.3.2. Закономерности морфофункциональных преобразований органов 69
13.3.3. Возникновение и исчезновение биологических структур
в филогенезе.............................................................................................. 71
13.3.4. Атавистические пороки развития................................................. 74
13.3.5. Аллогенные аномалии и пороки развития................................... 75
13.4. Организм как целое в историческом и индивидуальном развитии. Соотносительные преобразования органов................................................... 76
13.5. СОВРЕМЕННАЯ СИСТЕМА ОРГАНИЧЕСКОГО МИРА................. 80
13.5.1. Типы питания и основные группы живых организмов в природе 81
13.5.2. Происхождение многоклеточных животных............................... 81
13.5.3. Основные этапы прогрессивной эволюции многоклеточных животных.................................................................................................................... 83
13.5.4. Характеристика типа Хордовые................................................... 86
13.5.5. Систематика типа Хордовые........................................................ 87
13.5.6. Подтип Бесчерепные Acrania........................................................ 87
13.5.7. Подтип Позвоночные Vertebrata.................................................. 89
ГЛАВА 14. ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ................ 92
14.1. Наружные покровы................................................................................ 92
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ............................................. 96
14.2.1. Скелет............................................................................................ 96
14.2.1.1. Осевой скелет........................................................................ 96
14.2.1.2. Скелет головы...................................................................... 99
14.2.1.3. Скелет конечностей............................................................ 102
14.2.2. Мышечная система...................................................................... 109
14.2.2.1. Висцеральная мускулатура............................................... 110
14.2.2.2. Соматическая мускулатура............................................... 111
14.3. ПИЩЕВАРИТЕЛЬНАЯ И ДЫХАТЕЛЬНАЯ СИСТЕМЫ................. 112
14.3.1. Ротовая полость.......................................................................... 114
14.3.2. Глотка.......................................................................................... 117
14.3.3. Средняя и задняя кишка............................................................. 119
14.3.4. Органы дыхания......................................................................... 121
14.4. КРОВЕНОСНАЯ СИСТЕМА.............................................................. 123
14.4.1. Эволюция общего плана строения кровеносной системы
хордовых................................................................................................. 124
14.4.2. Филогенез артериальных жаберных дуг................................... 129
14.5. МОЧЕПОЛОВАЯ СИСТЕМА............................................................. 132
14.5.1. Эволюция почки.......................................................................... 132
14.5.2. Эволюция половых желез........................................................... 135
14.5.3. Эволюция мочеполовых протоков............................................. 136
14.6. ИНТЕГРИРУЮЩИЕ СИСТЕМЫ....................................................... 138
14.6.1. Центральная нервная система.................................................... 139
14.6.2. Эндокринная система.................................................................. 143
14.6.2.1. Гормоны............................................................................. 144
14.6.2.2. Железы внутренней секреции............................................ 145
ГЛАВА 15. АНТРОПОГЕНЕЗ И ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА......................................................................................................................... 149
15.1. МЕСТО ЧЕЛОВЕКА В СИСТЕМЕ ЖИВОТНОГО МИРА............... 149
15.2. МЕТОДЫ ИЗУЧЕНИЯ ЭВОЛЮЦИИ ЧЕЛОВЕКА........................... 150
15.3. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗА 154
15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА....... 159
15.4.1. Расы и расогенез......................................................................... 160
15.4.2. Адаптивные экологические типы человека................................ 164
15.4.3. Происхождение адаптивных экологических типов................... 167
РАЗДЕЛ V . БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ......................................................................................................................... 170
ГЛАВА 16. ВОПРОСЫ ОБЩЕЙ ЭКОЛОГИИ........................................ 170
16.1. БИОГЕОЦЕНОЗ - ЭЛЕМЕНТАРНАЯ ЕДИНИЦА БИОГЕОЦЕНОТИЧЕСКОГО УРОВНЯ ОРГАНИЗАЦИИ ЖИЗНИ.......... 172
16.2. ЭВОЛЮЦИЯ БИОГЕОЦЕНОЗОВ...................................................... 177
ГЛАВА 17. ВВЕДЕНИЕ В ЭКОЛОГИЮ ЧЕЛОВЕКА............................ 179
17.1. СРЕДА ОБИТАНИЯ ЧЕЛОВЕКА....................................................... 180
17.2. ЧЕЛОВЕК КАК ОБЪЕКТ ДЕЙСТВИЯ ЭКОЛОГИЧЕСКИХ
ФАКТОРОВ. АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ............ 182
17.3. АНТРОПОГЕННЫЕ ЭКОЛОГИЧЕСКИЕ СИСТЕМЫ...................... 186
17.3.1. Город........................................................................................... 186
17.3.2. Город как среда обитания людей............................................... 188
17.3.3. Агроценозы................................................................................. 189
17.4. РОЛЬ АНТРОПОГЕННЫХ ФАКТОРОВ В ЭВОЛЮЦИИ ВИДОВ
И БИОГЕОЦЕНОЗОВ................................................................................... 190
ГЛАВА 18. МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ.
ОБЩИЕ ВОПРОСЫ.................................................................................... 192
18.1. ПРЕДМЕТ И ЗАДАЧИ МЕДИЦИНСКОЙ ПАРАЗИТОЛОГИИ....... 192
18.2. ФОРМЫ МЕЖВИДОВЫХ БИОТИЧЕСКИХ СВЯЗЕЙ
В БИОЦЕНОЗАХ........................................................................................... 194
18.3. КЛАССИФИКАЦИЯ ПАРАЗИТИЗМА И ПАРАЗИТОВ.................. 195
18.4. РАСПРОСТРАНЕННОСТЬ ПАРАЗИТИЗМА В ПРИРОДЕ............. 198
18.5. ПРОИСХОЖДЕНИЕ ПАРАЗИТИЗМА.............................................. 198
18.6. АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ.
ОСНОВНЫЕ ТЕНДЕНЦИИ......................................................................... 200
18.7. ЦИКЛ РАЗВИТИЯ ПАРАЗИТОВ И ОРГАНИЗМ ХОЗЯИНА.......... 205
18.8. ФАКТОРЫ ВОСПРИИМЧИВОСТИ ХОЗЯИНА К ПАРАЗИТУ...... 207
18.9. ДЕЙСТВИЕ ХОЗЯИНА НА ПАРАЗИТА........................................... 208
18.10. СОПРОТИВЛЕНИЕ ПАРАЗИТОВ РЕАКЦИЯМ ИММУНИТЕТА ХОЗЯИНА........................................................................................................................ 209
18.11. ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ ПАРАЗИТ - ХОЗЯИН НА УРОВНЕ ПОПУЛЯЦИЙ................................................................................................ 210
18.12. СПЕЦИФИЧНОСТЬ ПАРАЗИТОВ ПО ОТНОШЕНИЮ К ХОЗЯИНУ 212
18.13. ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ.................................... 213
ГЛАВА 19. МЕДИЦИНСКАЯ ПРОТОЗООЛОГИЯ................................ 217
19.1. ТИП ПРОСТЕЙШИЕ PROTOZOA...................................................... 217
19.1.1. Класс Саркодовые Sarcodina...................................................... 218
19.1.2. Класс Жгутиковые Flagellata...................................................... 218
19.1.3. Класс Инфузории Infusoria......................................................... 219
19.1.4. Класс Споровики Sporozoa........................................................ 219
19.2. Простейшие, обитающие в полостных органах, сообщающихся
с внешней средой........................................................................................... 220
19.2.1. Простейшие, обитающие в полости рта..................................... 220
19.2.2. Простейшие, обитающие в тонкой кишке.................................. 221
19.2.3. Простейшие, обитающие в толстой кишке................................. 223
19.2.4. Простейшие, обитающие в половых органах............................ 225
19.2.5. Одноклеточные паразиты, обитающие в легких....................... 226
19.3. Простейшие, обитающие в тканях....................................................... 227
19.3.1. Простейшие, обитающие в тканях и передающиеся нетрансмиссивно.................................................................................................................. 228
19.3.2. Простейшие, обитающие в тканях и передающиеся
трансмиссивно......................................................................................... 230
19.4. ПРОСТЕЙШИЕ — ФАКУЛЬТАТИВНЫЕ ПАРАЗИТЫ ЧЕЛОВЕКА 239
ГЛАВА 20. МЕДИЦИНСКАЯ ГЕЛЬМИНТОЛОГИЯ............................ 240
20.1. ТИП ПЛОСКИЕ ЧЕРВИ PLATHELMINTHES................................... 240
20.1.1. Класс Сосальщики Trematoda.................................................... 241
20.1.1.1. Сосальщики с одним промежуточным хозяином,
обитающие в пищеварительной системе.......................................... 244
20.1.1.2. Сосальщики с одним промежуточным хозяином,
обитающие в кровеносных сосудах................................................. 246
20.1.1.3. Сосальщики с двумя промежуточными хозяевами.......... 249
20.1.2. Класс Ленточные черви Cestoidea.............................................. 255
20.1.2.1. Ленточные черви,.............. жизненный цикл которых связан
с водной средой................................................................................. 258
20.1.2.2. Ленточные черви,......... жизненный цикл которых не связан
с водной средой................................................................................. 260
20.1.2.3. Ленточные черви,........... проходящие в организме человека
весь жизненный цикл......................................................................... 266
20.2. ТИП КРУГЛЫЕ ЧЕРВИ NEMATHELMINTHES................................ 267
20.2.1. Класс Собственно круглые черви Nematoda............................. 268
20.2.1.1. Круглые черви — геогельминты....................................... 269
20.2.1.2. Круглые черви — биогельминты...................................... 274
20.2.1.3. Круглые черви,...... осуществляющие в организме человека
только миграцию............................................................................... 280
ГЛАВА 21. МЕДИЦИНСКАЯ АРАХНОЭНТОМОЛОГИЯ................... 281
21.1. КЛАСС ПАУКООБРАЗНЫЕ ARACHNOIDEA.................................. 281
21.1.1. Отряд Клещи Acari..................................................................... 282
21.1.1.1. Клещи — временные кровососущие эктопаразиты.......... 282
21.1.1.2. Клещи — обитатели человеческого жилья....................... 288
21.1.1.3. Клещи — постоянные паразиты человека........................ 290
21.2. КЛАСС НАСЕКОМЫЕ INSECTA....................................................... 291
21.2.1. Синатропные насекомые, не являющиеся паразитами.............. 292
21.2.2. Насекомые — временные кровососущие паразиты................... 296
21.2.3. Насекомые — постоянные кровососущие паразиты................. 304
21.2.4. Насекомые — тканевые и полостные эндопаразиты................. 306
ГЛАВА 22. ЭВОЛЮЦИЯ ПАРАЗИТОВ И ПАРАЗИТИЗМА
ПОД ДЕЙСТВИЕМ АНТРОПОГЕННЫХ ФАКТОРОВ......................... 308
ГЛАВА 23. ЯДОВИТОСТЬ ЖИВОТНЫХ КАК ЭКОЛОГИЧЕСКИЙ ФЕНОМЕН.................................................................................................... 313
23.1. ПРОИСХОЖДЕНИЕ ЯДОВИТОСТИ В ЖИВОТНОМ МИРЕ......... 315
23.2. ЧЕЛОВЕК И ЯДОВИТЫЕ ЖИВОТНЫЕ............................................ 316
РАЗДЕЛ VI. ЧЕЛОВЕК И БИОСФЕРА..................................................... 318
ГЛАВА 24. ВВЕДЕНИЕ В УЧЕНИЕ О БИОСФЕРЕ............................... 318
24.1. СОВРЕМЕННЫЕ КОНЦЕПЦИИ БИОСФЕРЫ.................................. 318
24.2. СТРУКТУРА И ФУНКЦИИ БИОСФЕРЫ.......................................... 319
24.3. ЭВОЛЮЦИЯ БИОСФЕРЫ.................................................................. 325
ГЛАВА 25. УЧЕНИЕ О НООСФЕРЕ......................................................... 326
25.1. БИОГЕНЕЗ И НООГЕНЕЗ................................................................... 326
25.2. ПУТИ ВОЗДЕЙСТВИЯ ЧЕЛОВЕЧЕСТВА........................................ 327
НА ПРИРОДУ. ЭКОЛОГИЧЕСКИЙ КРИЗИС............................................ 327
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА......................................................... 331
Биология.
В 2 кн. Кн. 2: Учеб. для медиц. спец. вузов/ Б63 В.Н. Ярыгин, В.И. Васильева И.Н. Волков, В.В. Синелыцикова;
Под ред. В.Н. Ярыгина. — 5-е изд., испр. и доп. — М.: Высш. шк., 2003. - 334 с.: ил.
ISBN 5-06-004589-7 (кн. 2)
В книге (1-й и 2-й книгах) освещены основные свойства жизни и эволюционные процессы последовательно на молекулярно-генетическом, онтогенетическом (1-я книга), популяционно-видовом и биогеоценотическом (2-я книга) уровнях организации. Изложены особенности проявления общебиологических закономерностей в онтогенезе и популяциях людей, их значение для медицинской практики. Уделено внимание биосоциальной сущности человека и его роли во взаимосвязях с природой.
Учебник отражает современные достижения биологической науки, играющие большую роль в практическом здравоохранении.
Для студентов медицинских специальностей вузов.
УДК 574/578
ББК 28.0
Учебное издание
Ярыгин
Владимир Никитич, Васильева
Вероника Игоревна,
Волков
Игорь Николаевич, Синелыцикова
Валерия Васильевна
БИОЛОГИЯ
В 2 книгах
Книга2
Редактор Т.С. Костян.
Художник К.Э. Семенков.
Художественный редактор Ю.Э. Иванова.
Технические редакторы Н.В. Быкова, В.М. Романова.
Корректор В.О. Бродская.
Компьютерная верстка Г.А. Шестакова.
Лицензия ИД № 06.236 от 09.11.01.
Изд. №|ХЕ-258. Подп. в печать 05.11.02. Формат 60х88 1
/16
.
Бум. газетн. Гарнитура «Таймс». Печать офсетная.
Объем 20,58 усл. печ. л. 20,58 усл. кр.-отт. 23,06 уч.-изд. л.
Тираж 8 000 экз. | Заказ № 259.
ФГУП «Издательство «Высшая школа»,
127994, Москва, ГСП-4, Неглинная ул., д. 29/14.
Тел.: (095) 200-04-56,
E-mail: info@v-shkola-ru, http: // www.v-shkola.ru
Отдел реализации:
(095) 200-07-69, 200-59-39, факс (095) 200-03-01.
E-mail: sales@v-shkola-ru
Отдел «Книга-почтой»:
(095) 200-33-36.E-mail: bookpost@v-shkola-ro
Набрано на персональных компьютерах издательства
Отпечатано в ОАО «Оригинал», 101990, Москва, Центр, Хохловский пер., 7—9, стр. 1—7.
1
Объем генетической информации, которой располагает вид или популяция, обусловливается совокупностью наследственных задатков во всех аллельных формах. Таким образом, более полно объем наследственной информации отражает термин «аллелофонд»,
но более употребим —«генофонд».
1
Определение справедливо для видов с половым размножением.
1
Число локусов (генов) у человека превышает эту цифру в 30—50 раз.
1
Наличие в популяций аллея с частотой менее 1% может быть объяснено только мутациями и комбинативной изменчивостью, без влияния естественного отбора.
1
Биопсия — метод гистологического исследования обычно окрашенных тканей, извлеченных из организма больного
|