Динамика популяций
Лекция 4
5 Динамика популяций
Динамика популяции – это процессы изменений ее основных биологических показателей во времени. При этом особое значение в изучении этой динамики придается изменениям численности особей, биомассы и популяционной структуры. Динамика популяции – одно из наиболее значимых биологических и экологических явлений. Образно говоря, жизнь популяции проявляется в ее динамике.
Модели динамики и роста популяции. Любая популяция способна (теоретически) к неограниченному росту численности, если ее не лимитируют факторы внешней среды абиотического (прежде всего климат) и биотического (конкуренция, хищники, паразиты, болезни) происхождения. В таком случае, конечно, гипотетическом, скорость роста популяции будет определяться величиной ранее упоминавшегося биотического или репродуктивного потенциала.
Эта динамика описывается уравнением А. Лотки:
(1)
где N – численность особей;
- время;
r – биотический потенциал.
График этой функции есть экспонента (рисунок 1, кривая 1).
Значение биотического потенциала чрезвычайно различается у разных видов. Так, самка косули способна произвести за жизнь 10…15 козлят, а луна-рыба откладывает до 3 млрд. икринок.
В природе, однако, рост численности популяций любого вида никогда не бывает бесконечным, поскольку ресурсы, за счет которых существуют виды, на любой территории имеют пределы. Эти пределы называют емкостью среды для конкретных популяций. Например, еловый лес – более емкая среда для белок, нежели смешанный, с березами, поскольку основная пища этих зверьков – семена шишек.
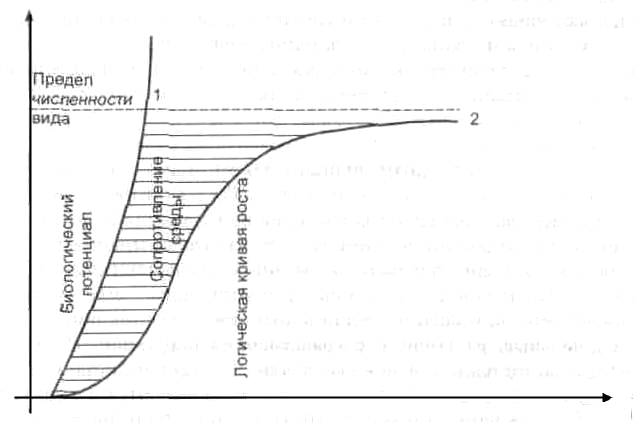
Рисунок 1 – Теоретические кривые роста популяции
Модель динамики численности популяции при ограниченных (лимитированных) ресурсах предложили Р. Пирл и А. Ферхюльст. В общем виде их уравнение записывается так:
(2)
где K – ёмкость среды.
Выражение- характеризует так называемое сопротивление среды. Под этим термином понимают совокупность всех «ограничителей» роста популяции (неоптимальная температура, кислотность, соленость, влажность, присутствие хищников, паразитов, нехватка пищи и т.п.).
Таким образом, рост, снижение или постоянство численности популяции зависит от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей). Уравнение Пирла-Ферхюльста лежит в основе практически всех математических моделей конкуренции, хищничества, симбиоза. Оно описывает наиболее реальный и универсальный тип роста популяций микроорганизмов, животных, растений и человека.
В природе характерные типы роста популяций могут иметь место, когда тех или иных животных вселяют в незанятые области или они сами распространяются в новые районы. Рост плотности этих популяций описывается логистической кривой 2 (рисунок 1).
Стабильные, растущие и сокращающиеся популяции. Популяция приспосабливается к изменениям условий среды обитания путем обновления и замещения особей. Последние появляются в популяции вследствие рождения и иммиграции (вселения пришельцев), а исчезают в результате смерти и эмиграции. Если интенсивности рождаемости и смертности сбалансированы, то формируется стабильная популяция, и ее численность и ареал обитания сохраняются на одном уровне. Однако в природе нет ни одной популяции, сохраняющейся неизменной на протяжении более или менее длительного периода.
Во многих случаях имеет место превышение, подчас существенное, рождаемости над смертностью; тогда численность популяции растет, иногда так быстро, что может наступить вспышка массового размножения. В качестве примера такой растущей популяции может служить колорадский жук, который за относительно короткий период пересек Атлантический океан, быстро расселился во Франции, достиг Украины, Беларуси и занял значительные территории России.
Однако при чрезмерном развитии популяции ухудшаются условия существования, что вызывается ее переуплотнением. Согласно правилу пищевой корреляции (Уинни-Эдварде), в ходе эволюции сохраняются только те популяции, скорость размножения которых скоррелирована с количеством пищевых ресурсов среды их обитания. Отступление от этого правила ведет к тому, что популяция остается без пищи и вымирает или снижает темпы размножения, т.е. она становится сокращающейся.
С другой стороны, согласно принципу В. Олли, агрегация (скопление) особей, как правило, усиливает конкурентную борьбу между ними за пищевые ресурсы и жизненное пространство, но приводит к повышению способности группы в целом к выживанию. Отсюда вытекает, что как «перенаселение» (т.е. повышенная агрегация особей), так и «недоселенность» (отсутствие агрегации) могут быть лимитирующим фактором.
Необходимо подчеркнуть, что безгранично популяция сокращаться не может. При достижении определенной численности популяции смертность начинает падать, а плодовитость повышаться. В определенный момент времени интенсивность смертности и рождаемости выравнивается, популяция переходит в стабильное состояние, а после становится растущей.
Гомеостаз популяции. В природных условиях численность популяций испытывает постоянные колебания, их амплитуда и период зависят от особенностей вида и от условий среды обитания. Так, у многих крупных позвоночных численность обычно колеблется в несколько раз, в популяциях насекомых – в 40…50 раз, а в особо благоприятных условиях возникают резкие вспышки численности, когда она возрастает в миллионы раз (саранча).
Кроме указанных нерегулярных колебаний у ряда организмов вы-явлены периодические колебания численности с относительно постоянной длительностью цикла, например связанные с периодическими колебаниями активности Солнца.
Н.В. Тимофеев-Ресовский ввел в 1928 г. термин «популяционные волны» для обозначения колебаний численности особей популяции, которые возникают под влиянием различных факторов биотической и абиотической среды. Будучи характерными для всех видов, популяционные волны (или «волны жизни») имеют определенное эволюционное значение, поскольку при резком сокращении численности какой-либо популяции среди оставшихся в живых особей могут оказаться редкие генотипы. В дальнейшем восстановление численности данной популяции будет идти за счет выживших особей, что приведет к изменению частот генов, а значит, и генофонда.
Факторы, которые влияют на численность популяции, разделяют на не зависящие и зависящие от ее плотности. Установлено, что к первым относятся прежде всего абиотические факторы. Продолжительная засуха, суровая зима, ураган и т.п. могут способствовать резкому снижению численности самых разных популяций, причем независимо от их первоначальной плотности.
К зависящим от плотности относится подавляющее большинство биотических факторов (конкуренция, хищники, обеспеченность кормом, инфекции и др.). Здесь имеет место в ряде случаев монотонная зависимость: с повышением плотности популяции сильнее влияют указанные факторы. Так, чем выше плотность популяции растений, тем сильнее они затеняют друг друга. Однако зависимость от плотности может быть и более сложной. Еще пример, связанный с внутривидовой регуляцией численности. Ученые обнаружили, что даже у таких медленно размножающихся животных, как африканские слоны, прослеживается связь особенностей размножения с численностью и плотностью популяций. Так, в некоторых заповедниках африканского континента, где насчитывалось по 2…3 особи на площади 2,5 км2, интервалы между родами у слоних составляли около трех лет, а возраст достижения молодыми половой зрелости – около 12 лет. После принятия более эффективных мер по охране слонов численность их возросла до 6…7 на ту же площадь. Однако самки стали приносить детенышей раз в 5…6 лет, а молодые стали размножаться только в 18 лет.
Особенностью зависящих от плотности факторов является то, что их воздействие обычно сглаживает колебания численности, способствуя при возрастании плотности популяций возвращению ее к среднему уровню. Следовательно, эти факторы действуют как еще один механизм регуляции численности, который способствует поддержанию ее на определенном уровне.
Способность популяции поддерживать определенную численность своих особей называется гомеостазом популяции. В основе этого важнейшего, эволюционно приобретенного свойства лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой эта особь принадлежит.
На рисунке 2 представлена схема поддержания гомеостаза животных, регулируемого доступностью пищевых ресурсов.
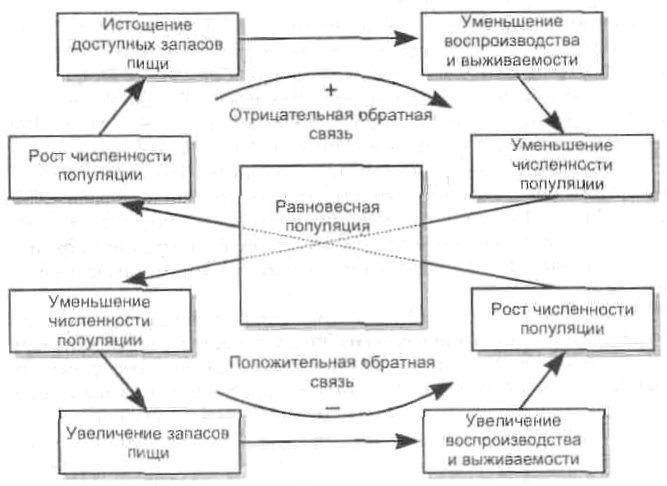
Рисунок 2 – Гомеостаз в популяции животных, регулируемый доступностью пищевых ресурсов (И.И. Дедю, 1989 г.)
При этом возникает так называемая отрицательная обратная связь: повышение плотности популяции усиливает действие механизмов, снижающих эту плотность.
Вышеизложенное говорит о том, что популяции присуще важнейшее свойство – саморегуляция. Она осуществляется действующими в природе двумя взаимно уравновешивающимися буферными силами: способностью к размножению и зависящей от плотности популяции реакции, напротив, ограничивающей воспроизводство.
В последнее время установлено, что важным механизмом регуляции численности, который срабатывает в переуплотненной популяции, является стресс-реакция. При воздействии на популяцию какого-то сильного раздражителя, она отвечает на него неспецифической реакцией, называемой стрессом. Многообразие живой природы порождает и много форм стресса: антропический (возникает под воздействием человека); нервно-психический (имеет место при несовместимости индивидуумов в группе или в результате переуплотнения популяции); тепловой; шумовой и др. Так, в результате переуплотнения популяции у отдельных особей возникают существенные физиологические изменения, приводящие к резкому сокращению рождаемости и увеличению смертности. У млекопитающих такое явление носит название стресс-синдрома. В стрессовом состоянии некоторые животные становятся настолько агрессивными, что у них почти полностью прекращается размножение. У самок часто нарушается овуляция, подчас происходит рассасывание эмбрионов, гаснет инстинкт заботы о потомстве. Тем не менее, важно подчеркнуть, что даже чрезвычайно высокая плотность или смертность не вызывают резких нарушений в структуре популяции. Этим гарантируется восстановление численности популяции в пределах оптимума в относительно короткие сроки.
Вопросы для самоконтроля
Динамика популяций.
PAGE 1
Динамика популяций