Анатомия центральной нервной системы
Содержание
Введение
Таламус (зрительный бугор)
Гипоталамус
Гипофиз: строение и функционирование
Гормоны передней доли гипофиза
Процессы в задней доле гипофиза
Функциональное значение ядер гипоталамуса
Особенности кровоснабжения гипоталамуса
Функциональное значение гипоталамуса
Заключение
Список литературы
Введение
Для современного психолога анатомия центральной нервной системы является базовым пластом психологических знаний. Не имея представлений о физиологической работе мозга, нельзя качественно изучать психические процессы и явления, а так же понимать их сущность.
Говоря о таламусе и гипоталамусе, следует прежде рассказать о промежуточном мозге (diencephalon). Промежуточный мозг располагается выше среднего мозга, под мозолистым телом. Он состоит из таламуса, эпиталамуса, метаталамуса и гипоталамуса. На основании мозга его граница спереди проходит по передней поверхности перекреста зрительных нервов, переднему краю заднего продырявленного вещества и зрительным трактам, а сзади - по краю ножек мозга. На дорсальной поверхности передней границей является терминальная полоска, отделяющая промежуточный мозг от конечного мозга, а задней границей - борозда, отделяющая промежуточный мозг от верхних холмиков среднего мозга. На сагиттальном срезе промежуточный мозг виден под мозолистым телом и сводом.
Полостью промежуточного мозга является III желудочек, который посредством правого и левого межжелудочковых отверстий сообщается с боковыми желудочками, расположенными внутри больших полушарий и посредством водопровода мозга - с полостью IV желудочка мозга. В верхней стенке III желудочка располагается сосудистое сплетение, участвующее наряду со сплетениями в других желудочках мозга в образовании спинномозговой жидкости.
Таламический мозг подразделяется на парные образования:
таламус (зрительный бугор);
метаталамус (заталамическая область);
эпиталамус (надталамическая область);
субталамус (подталамическая область).
Метаталамус (заталамическая область) образован парными медиальными и латеральными коленчатыми телами, расположенными позади каждого таламуса. В коленчатых телах располагаются ядра, в которых переключаются импульсы, идущие к корковым отделам зрительного и слухового анализатора.
Медиальное коленчатое тело находится позади подушки таламуса; вместе с нижними холмиками пластинки крыши среднего мозга оно является подкорковым центром слухового анализатора.
Латеральное коленчатое тело располагается книзу от подушки таламуса. Вместе с верхними бугорками четверохолмия оно образует подкорковый центр зрительного анализатора.
Эпиталамус (надталамическая область) включает шишковидное тело (эпифиз), поводки и треугольники поводков. В треугольниках поводков залегают ядра, относящиеся к обонятельному анализатору. Поводки отходят от треугольников поводков, идут каудально, соединяются посредством спайки и переходят в шишковидное тело. Последнее как бы подвешено на них и располагается между верхними бугорками четверохолмия. Шишковидное тело является железой внутренней секреции. Его функции полностью не установлены, предполагается, что оно регулирует наступление полового созревания.
Таламус (зрительный бугор)
Общее строение и расположение таламуса.
Таламус, или зрительный бугор, представляет собой парное образование яйцевидной формы объёмом около 3,3 см3, состоящее в основном из серого вещества (скопления многочисленных ядер). Таламусы образуются за счет утолщения боковых стенок промежуточного мозга. Спереди заостренная часть таламуса образует передний бугорок, в котором располагаются промежуточные центры сенсорных (афферентных) путей, идущих от ствола мозга в кору больших полушарий. Задняя, расширенная и закругленная часть таламуса - подушка - содержит подкорковый зрительный центр.

Рисунок 1. Промежуточный мозг на сагиттальном разрезе.
Толща серого вещества таламуса разделена вертикальной Y-образной прослойкой (пластинкой) белого вещества на три части - переднюю, медиальную и латеральную.
Медиальная поверхность таламуса хорошо видна на сагиттальном (сагиттальный - стреловидный (лат. "sagitta" - стрела), делящей на симметричные правую и левую половины) разрезе мозга (рис.1). Медиальная (т.е. располагающаяся ближе к середине) поверхность правого и левого таламусов, обращенные друг к другу, образуют боковые стенки III мозгового желудочка (полость промежуточного мозга) посередине они соединены между собой межталамическим сращением.
Передняя (нижняя) поверхность таламусов сращена с гипоталамусом, через нее с каудальной стороны (т.е. находящейся ближе к нижней части тела) в промежуточный мозг входят проводящие пути из ножек мозга.
Латеральная (т.е. боковая) поверхность таламуса граничит с внутренней капсулой - слоем белого вещества полушарий головного мозга, состоящего из проекционных волокон, соединяющих кору полушарий с нижележащими мозговыми структурами.
В каждой из этих частей таламуса находится несколько групп таламических ядер. Всего в таламусе содержится от 40 до 150 специализированных ядер [1; 37].
Функциональное значение ядер таламуса.
По топографии ядра таламуса объединяют в 8 основных групп:
1. переднюю группу;
2. медиодорсальную группу;
3. группу ядер средней линии;
4. дорсолатеральную группу;
5. вентролатеральную группу;
6. вентральную заднемедиальную группу;
7. заднюю группу (ядра подушки таламуса);
8. интраламинарную группу.
Ядра таламуса делят на сенсорные (специфические и неспецифические), моторные и ассоциативные. Рассмотрим основные группы ядер таламуса, необходимые для понимания его функциональной роли в передаче сенсорной информации в кору больших полушарий.
В передней части таламуса располагается передняя группа таламических ядер (рис.2). Наиболее крупные из них - передневентральное ядро и переднемедиальное ядро. Они получают афферентные волокна от сосцевидных тел - обонятельного центра промежуточного мозга. Эфферентные волокна (нисходящие, т.е. выносящие импульсы из мозга) от передних ядер направляются к поясной извилине коры больших полушарий.
Передняя группа таламических ядер и связанные с нею структуры являются важным компонентом лимбической системы мозга, управляющей психоэмоциональным поведением [5; 242].
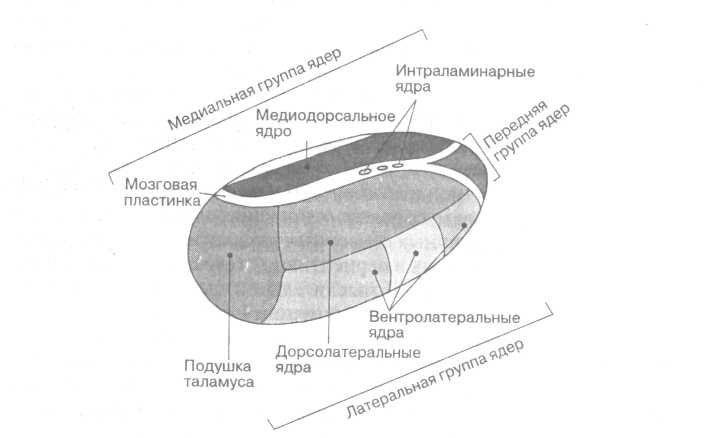
Рис. 2. Топография ядер таламуса
В медиальной части таламуса различают медиодорсалъное ядро и группу ядер средней линии.
Медиодорсальное ядро имеет двусторонние связи с обонятельной корой лобной доли и поясной извилиной больших полушарий, миндалевидным телом и переднемедиальным ядром таламуса. Функционально оно тесно связано также с лимбической системой и имеет двусторонние связи с корой теменной, височной и островковой долей мозга.
Медиодорсальное ядро участвует в реализации высших психических процессов. Его разрушение приводит к снижению беспокойства, тревожности, напряженности, агрессивности, устранению навязчивых мыслей.
Ядра средней линии многочисленны и занимают наиболее медиальное положение в таламусе. Они получают афферентные (т.е. восходящие) волокна от гипоталамуса, от ядер шва, голубого пятна ретикулярной формации ствола мозга и частично от спинно-таламических путей в составе медиальной петли. Эфферентные волокна от ядер средней линии направляются к гиппокампу, миндалевидному телу и поясной извилине больших полушарий, входящих в состав лимбической системы. Связи с корой больших полушарий двусторонние.
Ядра средней линии играют важную роль в процессах пробуждения и активации коры больших полушарий, а также в обеспечении процессов памяти.
В латеральной (т.е. боковой) части таламуса располагаются дорсолатералъная, вентролатеральная, вентральная заднемедиальная и задняя группы ядер.
Ядра дорсолатералъной группы относительно мало изучены. Известно, что они причастны к системе восприятия боли.
Ядра вентролатералъной группы анатомически и функционально различаются между собой. Задние ядра вентролатеральной группы часто рассматриваются как одно вентролатеральное ядро таламуса. Эта группа получает волокна восходящего пути общей чувствительности в составе медиальной петли. Сюда приходят также волокна вкусовой чувствительности и волокна от вестибулярных ядер. Эфферентные волокна, начинающиеся от ядер вентролатеральной группы, направляются в кору теменной доли больших полушарий, куда проводят соматосенсорную информацию от всего тела.
К ядрам задней группы (ядра подушки таламуса) идут афферентные волокна от верхних холмиков четверохолмия и волокна в составе зрительных трактов. Эфферентные волокна широко распространяются в коре лобной, теменной, затылочной, височной и лимбической долей больших полушарий.
Ядерные центры подушки таламуса причастны к комплексному анализу различных сенсорных раздражителей. Они играют значительную роль в перцептивной (связанной с восприятием) и когнитивной (познавательной, мыслительной) деятельности мозга, а также в процессах памяти - хранения и воспроизведения информации.
Интраламинарная группа ядер таламуса лежит в толще вертикальной Y-образной прослойки белого вещества. Интраламинарные ядра взаимосвязаны с базальными ядрами, зубчатым ядром мозжечка и корой больших полушарий.
Эти ядра играют важную роль в активационной системе мозга. Повреждение интраламинарных ядер в обоих таламусах приводит к резкому снижению двигательной активности, а также апатии и разрушению мотивационной структуры личности.
Кора больших полушарий благодаря двусторонним связям с ядрами таламуса способна оказывать регулирующее воздействие на их функциональную активность.
Таким образом, основными функциями таламуса являются:
переработка сенсорной информации от рецепторов и подкорковых переключающих центров с последующей передачей её коре;
участие в регуляции движений;
обеспечение связи и интеграции различных отделов мозга [5; 151].
Гипоталамус
Общее строение и расположение гипоталамуса.
Гипоталамус (hypothalamus) представляет собой вентральный отдел (т.е. брюшной) промежуточного мозга. В его состав входит комплекс образований, расположенных под III желудочком. Гипоталамус спереди ограничивается зрительным перекрестом (хиазмой), латерально - передней частью субталамуса, внутренней капсулой и зрительными трактами, отходящими от хиазмы. Сзади гипоталамус продолжается в покрышку среднего мозга. К гипоталамусу относят сосцевидные тела, серый бугор и зрительный перекрест. Сосцевидные тела располагаются по бокам средней линии кпереди от заднего продырявленного вещества. Это образования неправильной шаровидной формы белого цвета. Спереди от серого бугра располагается зрительный перекрёст. В нём происходит переход на противоположную сторону части волокон зрительного нерва, идущей от медиальной половины сетчатки. После перекрёста формируются зрительные тракты.
Серый бугор располагается кпереди от сосцевидных тел, между зрительными трактами. Серый бугор является полым выступом нижней стенки III желудочка, образованной тонкой пластинкой серого вещества. Верхушка серого бугра вытянута в узкую полую воронку, на конце которой находится гипофиз [4; 18].
Гипофиз: строение и функционирование
Гипофиз (hypophysis) - железа внутренней секреции, он располагается в специальном углублении основания черепа, "турецком седле" и при помощи ножки связан с основанием мозга. В гипофизе выделяют переднюю долю (аденогипофиз - железистый гипофиз) и заднюю долю (нейрогипофиз).
Задняя доля, или нейрогипофиз, состоит из нейроглиальных клеток и является продолжением воронки гипоталамуса. Более крупная доля - аденогипофиз, построена из железистых клеток. Благодаря тесному взаимодействию гипоталамуса с гипофизом в промежуточном мозге функционирует единая гипиталамо-гипофизарная система, управляющая работой всех эндокринных желез, а с их помощью - вегетативными функциями организма (рис.3).
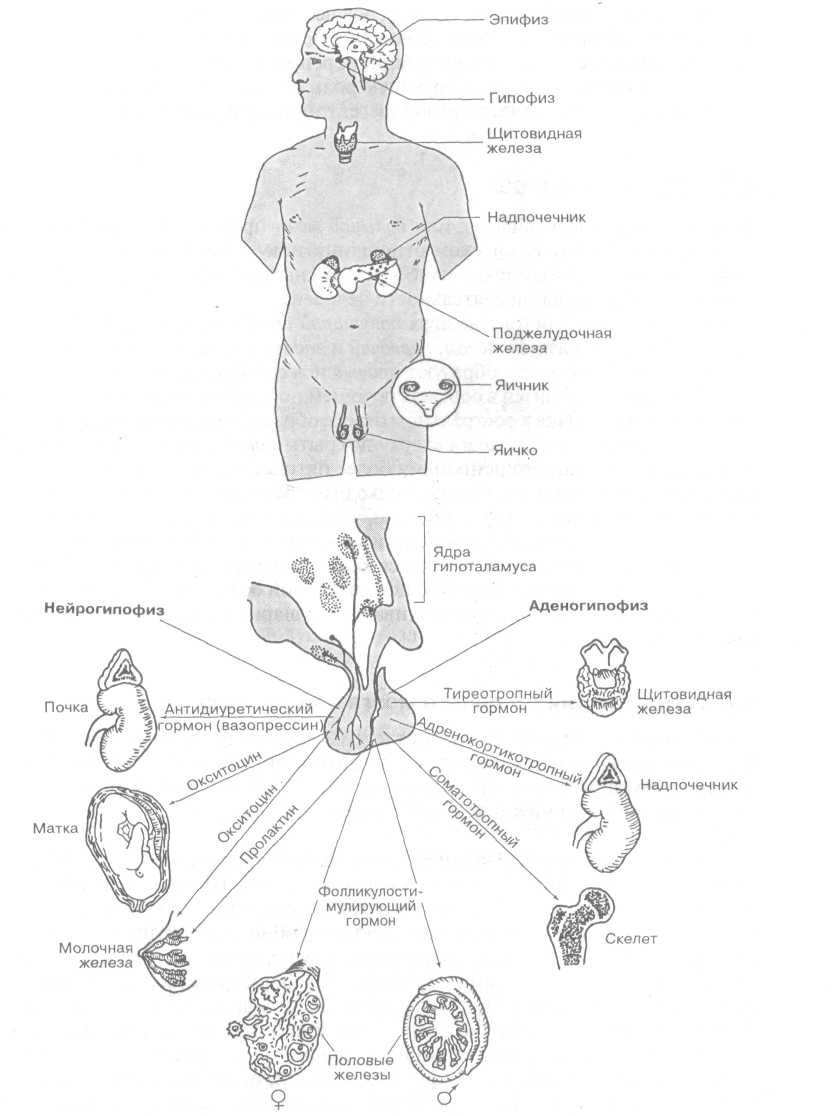
Рисунок 3. Гипофиз и его влияние на другие эндокринные железы
В сером веществе гипоталамуса выделяют 32 пары ядер. Взаимодействие с гипофизом осуществляется посредством выделяемых ядрами гипоталамуса нейрогормонов - рилизинг-гормонов. По системе кровеносных сосудов они попадают в переднюю долю гипофиза (аденогипофиз), где способствуют высвобождению тропных гормонов, стимулирующих синтез специфических гормонов в других эндокринных железах.
В передней доле гипофиза вырабатываются тропные гормоны (тиреотропный гормон - тиреотропин, адренокортикотропный гормон - кортикотропин и гонадотропные гормоны - гонадотропины) и эффекторные гормоны (гормоны роста - соматотропин и пролактин) [5; 139].
Гормоны передней доли гипофиза
Тропные:
Тиреотропный гормон (тиреотропин) стимулирует функцию щитовидной железы. Если удалить или разрушить гипофиз у животных, то наступает атрофия щитовидной железы, а введение тиреотропина восстанавливает ее функции.
Адренокортикотропный гормон (кортикотропин) стимулирует функцию пучковой зоны коры надпочечников, в которой образуются гормоны глюкокортикоиды. В меньшей степени выражено влияние гормона на клубочковую и сетчатую зоны. Удаление гипофиза у животных приводит к атрофии коркового слоя надпочечников. Атрофические процессы захватывают все зоны коры надпочечников, но наиболее глубокие изменения происходят в клетках сетчатой и пучковой зонах. Вненадпочечниковое действие кортикотропина выражается в стимуляции процессов липолиза, усилении пигментации, анаболическом влиянии.
Гонадотропные гормоны (гонадотропины). Фолликулостимулирующий гормон (фоллитропин) стимулирует рост везикулярного фолликула в яичнике. Влияние фоллитропина на образование женских половых гормонов (эстрогенов) небольшое. Этот гормон имеется как у женщин, так и у мужчин. У мужчин под влиянием фоллитропина происходит образование половых клеток (сперматозоидов). Лютеинизирующий гормон (лютропин) необходим для роста везикулярного фолликула яичника на стадиях, предшествующих овуляции, и для самой овуляции (разрыва оболочки созревшего фолликула и выхода из него яйцеклетки), образования желтого тела на месте лопнувшего фолликула. Лютропин стимулирует образование женских половых гормонов - эстрогенов. Однако для того чтобы этот гормон осуществил свое действие на яичник, необходимо предварительное длительное действие фоллитропина. Лютропин стимулирует выработку прогестерона желтым телом. Лютропин имеется как у женщин, так и у мужчин. У мужчин он способствует образованию мужских половых гормонов - андрогенов.
Эффекторные:
Гормон роста (соматотропин) стимулирует рост организма путем усиления образования белка. Под влиянием роста эпифизарных хрящей в длинных костях верхних и нижних конечностей происходит рост костей в длину. Гормон роста усиливает секрецию инсулина посредством соматомединов, образующихся в печени.
Пролактин стимулирует образование молока в альвеолах молочных желез. Свое действие на молочные железы пролактин оказывает после предварительного действия на них женских половых гормонов прогестерона и эстрогенов. Акт сосания стимулирует образование и выделение пролактина. Пролактин обладает также и лютеотропным действием (способствует продолжительному функционированию желтого тела и образованию им гормона прогестерона) [1; 73].
Процессы в задней доле гипофиза
В задней доле гипофиза гормоны не вырабатываются. Сюда поступают неактивные гормоны, которые синтезируются в паравентрикулярном и супраоптическом ядрах гипоталамуса.
В нейронах паравентрикулярного ядра образуется преимущественно гормон окситоцин, а в нейронах супраоптического ядра - вазопрессин (антидиуретический гормон). Эти гормоны накапливаются в клетках задней доли гипофиза, где они превращаются в активные гормоны.
Вазопрессин (антидиуретический гормон) играет важную роль в процессах мочеобразования и в меньшей степени в регуляции тонуса кровеносных сосудов. Вазопрессин, или антидиуретический гормон - АДГ (диурез - выделение мочи) - стимулирует обратное всасывание (резорбцию) воды в почечных канальцах.
Окситоцин (оцитонин) усиливает сокращение матки. Ее сокращение резко усиливается, если она предварительно находилась под действием женских половых гормонов эстрогенов. Во время беременности окситоцин не влияет на матку, так как под влиянием гормона желтого тела прогестерона она становится нечувствительной к окситоцину. Механическое раздражение шейки матки вызывает отделение окситоцина рефлекторно. Окситоцин обладает способностью стимулировать также выделение молока. Акт сосания рефлекторно способствует выделению окситоцина из нейрогипофиза и выделению молока. В состоянии напряжения организма гипофиз выделяет дополнительное количество АКТГ, стимулирующего выброс адаптивных гормонов корой надпочечников [1; 75].
Функциональное значение ядер гипоталамуса
В передне-боковой части гипоталамуса различают переднюю и среднюю группы гипоталамических ядер (рис.4).
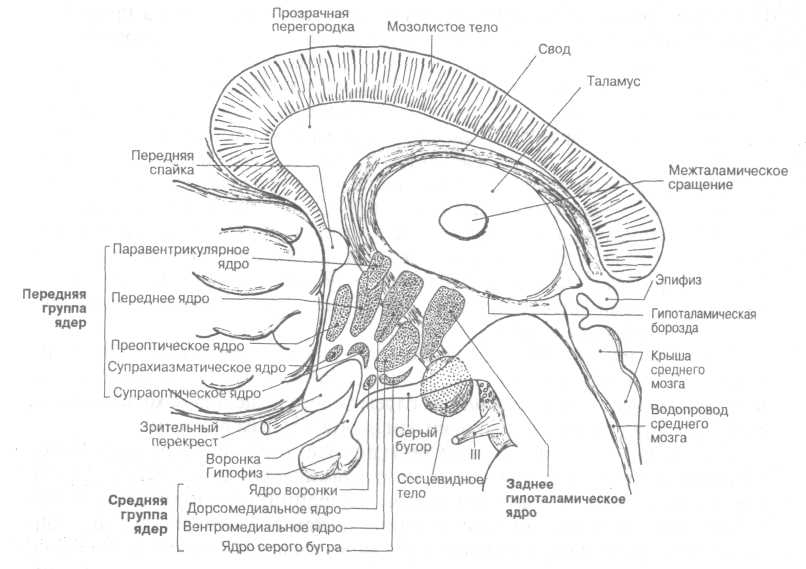
Рисунок 4. Топография ядер гипоталамуса
К передней группе относятся супрахиазматические ядра, преоптическое ядро, и самые крупные - супраоптическое и паравентрикулярное ядра.
В ядрах передней группы локализуются:
центр парасимпатического отдела (ПСНС) вегетативной нервной системы.
Стимуляция переднего отдела гипоталамуса приводит к реакциям парасимпатического типа: сужению зрачка, снижению частоты сокращений сердца, расширению просвета сосудов, падению артериального давления, усилению перистальтики (т.е. волнообразного сокращения стенок полых трубчатых органов, способствующего продвижению их содержимого к выходным отверстиям кишечника);
центр теплоотдачи. Разрушение переднего отдела сопровождается необратимым повышением температуры тела;
центр жажды;
нейросекреторные клетки, продуцирующие вазопрессин (супраоптическое ядро) и окситоцин (паравентрикулярное ядро). В нейронах паравентрикулярного и супраоптических ядер образуется нейросекрет, который по их аксонам перемещается в задний отдел гипофиза (нейрогипофиз), где высвобождается в виде нейрогормонов - вазопрессина и окситоцина, поступающих в кровь.
Повреждение передних ядер гипоталамуса приводит к прекращению выделения вазопрессина, вследствие чего развивается несахарный диабет. Окситоцин оказывает стимулирующее действие на гладкую мускулатуру внутренних органов, например матки. В целом от этих гормонов зависит водносолевой баланс организма.
В преоптическом ядре образуется один из рилизинг-гормонов - люлиберин, стимулирующий выработку в аденогипофизе лютеинизирующего гормона, контролирующего активность половых желез.
Супрахиазматические ядра принимают активное участие в регуляции циклических изменений активности организма - циркадианных, или суточных, биоритмов (например, в чередовании сна и бодрствования).
К средней группе гипоталамических ядер относят дорсомедиальное и вент-ромедиальное ядра, ядро серого бугра и ядро воронки.
В ядрах средней группы локализуются:
центр голода и насыщения. Разрушение вентромедиального ядра гипоталамуса приводит к избыточному потреблению пищи (гиперфагии) и ожирению, а повреждение ядра серою бугра - к снижению аппетита и резкому исхуданию (кахексии);
центр полового поведения;
центр агрессии;
центр удовольствия, играющий важную роль в процессах формирования мотиваций и психоэмоциональных форм поведения;
нейросекреторные клетки, продуцирующие рилизинг-гормоны (либерины и статины), регулирующие продукцию гипофизарных гормонов: соматостатин, соматолиберин, люлиберин, фоллиберин, пролактолиберин, тиреолиберин и др. Через гипоталамо-гипофизарную систему они оказывают влияние на ростовые процессы, скорость физического развития и полового созревания, формирование вторичных половых признаков, функции половой системы, а также на обмен веществ.
Средняя группа ядер контролирует водный, жировой и углеводный обмен, влияет на уровень сахара в крови, ионный баланс организма, проницаемость сосудов и клеточных мембран.
Задняя часть гипоталамуса расположена между серым бугром и задним продырявленным веществом и состоит из правого и левого сосцевидных тел.
В задней части гипоталамуса наиболее крупными ядрами являются: медиальное и латеральное ядра, заднее гипоталамическое ядро.
В ядрах задней группы локализуются:
центр, координирующий активность симпатического отдела (СНС) вегетативной нервной системы (заднее гипоталамическое ядро). Стимуляция этого ядра приводит к реакциям симпатического типа: расширению зрачка, повышению частоты сокращений сердца и артериального давления, учащению дыхания и уменьшению тонических сокращений кишечника;
центр теплопродукции (заднее гипоталамическое ядро). Разрушение заднего отдела гипоталамуса вызывает вялость, сонливость и снижение температуры тела;
подкорковые центры обонятельного анализатора. Медиальное и латеральное ядра в каждом сосцевидном теле являются подкорковыми центрами обонятельного анализатора, а также входят в лимбическую систему;
нейросекреторные клетки, продуцирующие рилизинг-гормоны, регулирующие продукцию гипофизарных гормонов [5; 148].
Особенности кровоснабжения гипоталамуса
Ядра гипоталамуса получают обильное кровоснабжение. Капиллярная сеть гипоталамуса по своей разветвлённости в несколько раз больше, чем в других отделах ЦНС. Одной из особенностей капилляров гипоталамуса является их высокая проницаемость, обусловленная истонченностью стенок капилляров и их фенестрированностью ("окончатостью" - наличие промежутков - "окон" - между смежными эндотелиальными клетками капилляров (от лат. "fenestra" - окно). В результате этого в гипоталамусе слабо выражен гематоэнцефалический барьер (ГЭБ), и нейроны гипоталамуса способны воспринимать изменения состава спинномозговой жидкости и крови (температуру, содержание ионов, наличие и количество гормонов и т.д.).
Функциональное значение гипоталамуса
Гипоталамус является центральным звеном, связующим нервные и гуморальные механизмы регуляции вегетативных функций организма. Управляющая функция гипоталамуса обусловлена способностью его клеток к секреции и аксональному транспорту регуляторных веществ, которые переносится в другие структуры мозга, спинномозговую жидкость, кровь или в гипофиз, изменяя функциональную активность органов-мишеней.
В гипоталамусе выделяют 4 нейроэндокринные системы:
Гипоталамо-экстрагипоталамная система представлена нейросекреторными клетками гипоталамуса, аксоны которых уходят в таламус, структуры лимбической системы, продолговатый мозг. Эти клетки выделяют эндогенные опиоиды, соматостатин и др.
Гипоталамо-аденогипофизарная система связывает ядра заднего гипоталамуса с передней долей гипофиза. По этому пути транспортируются рилизинг-гормоны (либерины и статины). Посредством их гипоталамус регулирует секрецию тропных гормонов аденогипофиза, определяющих секреторную активность желёз внутренней секреции (щитовидной, половых и др.).
Гипоталамо-метагипофизарная система связывает нейросекреторные клетки гипоталамуса с гипофизом. По аксонам этих клеток транспортируются меланостатин и меланолиберин, которые регулируют синтез меланина - пигмента, определяющего окраску кожи, волос, радужки и других тканей организма.
Гипоталамо-нейрогипофизарная система связывает ядра переднего гипоталамуса с задней (железистой) долей гипофиза. По этим аксонам транспортируются вазопрессин и окситоцин, которые накапливаются в задней доле гипофиза и выделяются в кровоток по мере необходимости [6; 92].
Заключение
Таким образом, дорсальный отдел промежуточного мозга представляет собой филогенетически более молодой таламический мозг, являющийся высшим подкорковым сенсорным центром, в котором переключаются практически все афферентные пути, несущие сенсорную информацию от органов тела и органов чувств к большим полушариям головного мозга. К задачам гипоталамуса относится также управление психоэмоциональным поведением и участие в реализации высших психических и психологических процессов, в частности памяти.
Вентральный отдел - гипоталамус является более старым в филогенетическом отношении образованием. Гипоталамо-гипофизарная система осуществляет контроль над гуморальной регуляцией водносолевого баланса, обменом веществ и энергии, работой иммунной системы, терморегуляцией, репродуктивной функцией и т.д. Выполняя и этой системе регулирующую роль, гипоталамус является высшим центром, управляющим автономной (вегетативной) нервной системой.
Список литературы
- Анатомия человека / Под ред. М.Р. Сапина. - М.: Медицина, 1993.
- Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум поведение. - М.: Мир, 1988.
- Гистология / Под ред. В.Г. Елисеева. - М.: Медицина, 1983.
- Привес М.Г., Лысенков Н.К., Бушкович В.И. Анатомия человека. - М.: Медицина, 1985.
- Синельников Р.Д., Синельников Я.Р. Атлас анатомии человека. - М.: Медицина, 1994.
- Тишевской И.А. Анатомия центральной нервной системы: Учебное пособие. - Челябинск: Изд-во ЮУрГУ, 2000.
Анатомия центральной нервной системы