ФАУНИСТИЧЕСКИЙ КОМПЛЕКС IXODIDAE КАРАЧАЕВО-ЧЕРКЕССКОЙ РЕСПУБЛИКИ
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
«КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
(ФГБОУ ВПО «КубГУ»)
Кафедра зоологии
|
|
ДОПУСТИТЬ К ЗАЩИТЕ В ГАК «_____»________________2014 г. Заведующий кафедрой – _______________ Т.Ю. Пескова «_____»________________2014 г.
|
ВЫПУСКНАЯ КВАЛИФИКАЦИОННАЯ РАБОТА
ФАУНИСТИЧЕСКИЙ КОМПЛЕКС IXODIDAE КАРАЧАЕВО-ЧЕРКЕССКОЙ РЕСПУБЛИКИ
Работу выполнила _____________________________________________Е.Л. Стрельцова
(подпись, дата)
Факультет биологический
Направление 06.03.01 Биология
Научный руководитель, канд. биол. наук, доцент __________________С.В. Островских
(подпись, дата)
Нормоконтролёр, канд. биол. наук___________________________________В.В. Гладун
(подпись, дата)
Краснодар 2014
РЕФЕРАТ
Работа изложена на 51 странице, состоит из введения, 4 глав, заключения, содержит 12 рисунков. Список цитированной литературы включает 46 источника, в том числе 2 иностранных.
Ключевые слова: ИКСОДОВЫЕ КЛЕЩИ, КАРАЧАЕВО-ЧЕРКЕССКАЯ РЕСПУБЛИКА, ВИДЫ ИКСОДИД, ФАУНА ИКСОДОВЫХ КЛЕЩЕЙ, СТЕПНАЯ И ЛЕСОСТЕПНАЯ ЗОНА.
Целью работы является изучение фауны иксодовых клещей степной и лесостепной зон Карачаево-Черкесской республики.
В ходе работы определен видовой состав иксодид степной и лесостепной зон КЧР, определен индекс доминирования найденных видов клещей, выявлена экстенсивность и интенсивность клещевой инвазии по зонам исследования.
СОДЕРЖАНИЕ
Введение…………………………………………………………………………. 4
1 Аналитический обзор………………………………………………………..... 6
1.1 Иксодовые клещи мировой фауны …………………………………..….. 6
1.2 Иксодовые клещи Северного Кавказа и сопредельных территорий ..... 8
2 Физико-географическая характеристика района исследования………..… 17
3 Материал и методы исследования………………………………………...... 20
4 Фаунистический комплекс Ixodidae Карачаево-Черкесской республики .. 24
4.1 Состав фауны иксодид……………………………………………….…... 24
4.2 Интенсивность и экстенсивность клещевой инвазии………….………. 27
Заключение…………………………………………………………….…….…. 45
Библиографический список…………………………………………...….….… 47
ВВЕДЕНИЕ
Последние годы в большинстве стран мира, в том числе и в РФ характеризуются значительным ростом заболеваемости инфекционными болезнями с природной очаговостью, среди которых особое место занимают инфекции, передаваемые иксодовыми клещами [Онищенко и др., 2001]. Изучение иксодовых клещей является одной из существенных задач, имеющей не только теоретическое, но и практическое значение. Исключительное практическое значение иксодид, как переносчиков возбудителей многих природно-очаговых инфекций человека и сельскохозяйственных животных, давно привлекло внимание исследователей [Померанцев, 1950]. Многочисленными исследованиями, выполненными отечественными и зарубежными авторами, доказана способность иксодид сохранять в своем теле и передавать прокормителям патогенных простейших, бактерий, вирусов, риккетсий и других возбудителей [Коренберг, 1999].
В последние десятилетия произошли существенные изменения условий ведения сельскохозяйственного производства. Это привело к увеличению биотопов, благоприятных для иксодовых клещей и ухудшению эпидемической и эпизоотической ситуации по ряду заболеваний передаваемых клещами. В частности, на юге России осложнилась эпидобстановка по Крымской геморрагической лихорадке (КГЛ), по пироплазмидозам и ряду других инфекций [Онищенко и др., 2001; 2005; Котти, Гудиев, 1997].
Изучение и уточнение видового состава иксодовых клещей, их взаимосвязь с прокормителями, позволяет прогнозировать численность и своевременно проводить мероприятия, направленные на профилактику эпидемических осложнений ряда инфекционных заболеваний в настоящее время представляет одну из актуальных задач.
Целью работы является изучение фауны иксодовых клещей степной и лесостепной зон Карачаево-Черкесской республики.
Задачи исследования:
1) Установить таксономический состав иксодовых клещей Карачаево-Черкесской республики по степной и лесостепной зоне и провести его сравнение с фауной иксодид Северного Кавказа.
2) Определить индекс доминирования клещей по зонам исследования.
3) Выявить экстенсивность и интенсивность клещевой инвазии степной и лесостепной зон.
- Аналитический обзор
- Иксодовые клещи мировой фауны
Иксодовые клещи (семейство Ixodidae, отряд Parasitiformes) распространены на всех материках земного шара, обитают в разнообразных ландшафтных зонах. Паразиты млекопитающих, птиц и рептилий. Служат переносчиками большого количества возбудителей (вирусов, бактерий, простейших) различных болезней человека и животных.
Мировая фауна Ixodidae составляет 14 родов (около 680 видов), из них 6 широко распространены и многочисленны. Максимальное число видов (250) включает род Ixodes [Балашов, 1998; Филиппова, 1997]. Род встречается во всех зонах вплоть до побережий полярных морей. Несмотря на широкое распространение, виды рода Ixodes повсеместно приурочены к местам с достаточной влагообеспеченностью. Виды, подстерегающие прокормителей на растительности (I. ricinus, I. рersulcatus), приурочены к различным типам леса при постоянно высокой относительной влажности в приземном слое воздуха. Виды, встречающиеся в жарком климате, обычно обитают в норах (гнездово-норовый тип паразитизма, I. laguri, I. subterraneus). Некоторые виды приспособились к жизни в норах с избыточным увлажнением – I. apronophorus в Палеарктике, I. ornithorhynchi из нор утконосов в Австралии или I. aulacodi, паразитирующий в норах тростниковой крысы в Африке. Подобные условия избыточной увлажненности характерны для местообитаний паразитов колониальных морских птиц – I. uriae, I. signatus.
Второй по количеству видов род Haemaphysalis (147 видов), включает 16 подродов, из которых 4 (17 видов) считаются эволюционно примитивными [Hoogstraal, Kaiser, 1960]. В Индо-Малайской области обитает 90 видов, в Эфиопской – 19, на Мадагаскаре – 12, в Австралии – 12, в Палеарктике – 10. Клещи рода связаны с мезофильными лесными ландшафтами. Некоторые виды, такие как H. concinna, имеют обширные ареалы, а ряд видов приспособилось к существованию в редколесье и кустарниках (H. рunctata), в степных или полупустынных участках (H. sulcata). На юго-западе бывшего СССР встречаются в дубняках, по долинам рек, в кустарниковых и лугово-степных местообитаниях [Емчук, 1960; Успенская, 1987].
Род Dermacentor насчитывает 32 вида и обладает обширным ареалом. В Палеарктике обитает 15 видов, в Северной и Центральной Америке – 11, в тропической Азии – 4 и в Африке – 2 вида. Для палеарктических видов характерна приуроченность к степным или луговым ландшафтам, редколесьям, вырубкам, опушкам. Некоторые виды, например D. pavlovskyi, заходят на высоту до 3500–4000 м над уровнем моря [Филиппова, Панова, 1989].
Из 73 видов рода Rhipicephalus 55 встречается в Африке, 8 – в Палеарктике и 4 – в тропической Азии. Подавляющая масса видов заселяет разные типы степей, саванн и реже пустынь. Некоторые виды могут заселять редколесья и кустарники (R. leporis). Род Boophilus включает 5 видов. С домашним скотом проник из Средиземноморья в Австралию, Северную и Южную Америку. Род Hyalomma включает 30 видов, из которых половина встречается в саваннах и тропических редколесьях Африки, Южной Европе, Центральной и Средней Азии [Ouhelli, 1994].
Большинство видов иксодид обладают полигостальностью. Круг хозяев клещей насчитывает десятки и сотни видов различных таксономических групп – млекопитающих, птиц, рептилий. Специфичность к хозяевам среди иксодид отмечается редко, причем обычно к крупным таксономическим группам. Наибольшее число видов иксодид (более 350), по крайней мере, на неполовозрелых фазах, питается на грызунах. Второй значительной группой прокормителей служат парнокопытные (в том числе крупный и мелкий рогатый скот), на которых паразитируют 190 видов иксодовых клещей (главным образом, имаго). Представители отряда хищных служат хозяевами 49 видов клещей, главным образом родов Haemaphysalis и Rhipicephalus. Многие виды иксодид обнаружены на насекомоядных, для 25 видов клещей они служат основными прокормителями. Особенно велика роль в прокормлении иксодид различных видов ежей. В условиях антропогенной трансформации ландшафтов и резкого сокращения численности диких копытных доминирующую роль в пропитании имаго многих видов иксодид повсеместно играет крупный и мелкий рогатый скот (КРС и МРС). Личинки и нимфы 3-хозяинных видов иксодид питаются на мелких и средних млекопитающих и на собирающих корм с земли птицах. Они в большей степени сохранили трофические связи с дикими видами.
На территории бывшего СССР известно 86 видов иксодовых клещей, относящихся к 8 родам, в России – 55 видов 6 родов [Филиппова, 1984]. Большого разнообразия фауна клещей достигает в южных регионах России, в которых теплый климат и сочетание разнохарактерных ландшафтов с многообразием естественных прокормителей создают широкий диапазон условий для жизнедеятельности иксодовых клещей.
1.2 Иксодовые клещи Северного Кавказа и сопредельных территорий
Фауна иксодид Карачаево-Черкесской республики изучена недостаточно. Чтобы составить полное представление об уже известных видах и тех, которые еще могут быть обнаружены на территории республики, мы рассматриваем фауну иксодид Северного Кавказа и других сопредельных территорий со сходными природными условиями. В России это Краснодарский [9 видов, Костюковский и др., 2005] и Ставропольский края [15 видов, Тохов, 2004], Волгоградская области [11 видов, Денисов и др., 2005], республик Ингушетия [8 видов, Онищенко и др., 2005;], Калмыкия [8 видов, Подсвиров и др., 2005] и Дагестан [15 видов, Климова и др., 2005], Армения [12 видов, Манукян и др., 2006], Азербайджан [19 видов, Кулиева, Соколова, 1976], Грузия [7 видов, Балашов, 1998] и Крым [17 видов, Евстафьев, 2001]. Всего, по литературным данным [Асваров, 2002; Балашов, 1998; Боженко, Шевченко, 1956; Григорьев и др., 2005; Денисов и др., 2005; Евстафьев, 2001; Емельянова, 2005; Климова и др., 2005; Колонин, 1984; Костюковский, и др., 2005; Котти, Гудиев, 1997; Котти др., 2001; Кузнецова и др., 2006; Кулиева, Соколова, 1976; Манукян и др., 2006; Омариева и др., 2005; Онищенко и др., 2001; Онищенко др., 2005; Подсвиров и др., 2005; Теплова и др., 1997; Тохов, 2004; Тохов и др., 2001; Филиппова, 1997], на Северном Кавказе и в прилегающих регионах обитает 26 видов иксодовых клещей (таблица 1).
Таблица 1 – Иксодовые клещи Северного Кавказа и прилегающих регионов (по данным литературных источников)
|
№
п.п.
|
Виды клещей
|
КЧР
|
Краснодарский край
|
Ставропольский край
|
Ростовская обл.
|
Волгоградская обл.
|
Ингушетия
|
Дагестан
|
Калмыкия
|
Азербайджан
|
Крым
|
Грузия
|
Армения
|
|
1
|
B. annulatus
|
+
|
+
|
+
|
|
+
|
+
|
+
|
|
+
|
+
|
|
+
|
|
2
|
D. marginatus
|
+
|
+
|
+
|
+
|
+
|
|
+
|
|
+
|
+
|
|
+
|
|
3
|
D. reticulatus
|
+
|
+
|
+
|
|
+
|
|
|
|
|
+
|
|
+
|
|
4
|
D. niveus
|
|
|
+
|
|
|
|
+
|
+
|
|
|
|
|
|
5
|
Н. concinna
|
+
|
+
|
|
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
|
6
|
H. punctata
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
|
7
|
H. inermis
|
|
|
+
|
|
|
|
+
|
|
+
|
+
|
+
|
+
|
|
8
|
H. parva
|
+
|
|
+
|
|
+
|
+
|
+
|
|
|
+
|
+
|
+
|
|
9
|
H. sulcata
|
|
|
|
|
|
|
+
|
|
+
|
+
|
|
+
|
|
10
|
H. pospelovashtromae
|
|
|
|
|
|
|
+
|
|
+
|
|
+
|
+
|
|
11
|
H. caucasica
|
|
|
|
|
|
|
+
|
|
+
|
+
|
+
|
|
|
12
|
H. asiaticum
|
|
|
|
|
|
|
|
|
|
|
|
+
|
|
13
|
H. aegyptium
|
|
|
|
|
|
|
|
|
+
|
|
|
|
|
14
|
H. marginatum
|
+
|
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
|
|
|
15
|
H. anatolicum
|
|
|
+
|
|
|
|
+
|
|
|
|
|
|
|
16
|
H. scupense
|
+
|
+
|
+
|
|
+
|
|
+
|
|
+
|
+
|
|
|
|
17
|
H. detritum
|
|
|
+
|
|
|
|
+
|
|
+
|
|
|
|
|
18
|
R. bursa
|
|
|
|
|
|
+
|
+
|
+
|
+
|
+
|
|
+
|
|
19
|
R. sanguineus
|
|
+
|
+
|
|
|
|
|
|
+
|
+
|
|
|
|
20
|
R. pumilio
|
|
|
+
|
|
|
|
|
|
+
|
+
|
|
|
|
21
|
R. schulzei
|
|
+
|
|
|
+
|
+
|
|
+
|
+
|
|
|
|
|
22
|
R. rossicus
|
+
|
|
+
|
+
|
+
|
+
|
|
+
|
+
|
+
|
+
|
|
|
23
|
R. turanicus
|
|
|
|
|
|
|
|
+
|
+
|
|
|
|
|
24
|
I. trianguliceps
|
|
|
|
|
|
|
|
|
|
+
|
|
|
|
25
|
I. redikorzevi
|
|
|
|
|
|
|
|
|
+
|
+
|
|
|
|
26
|
I. ricinus
|
+
|
+
|
+
|
+
|
+
|
|
|
|
|
|
|
|
Boophilus annulatus. Проник из Средиземноморья в Австралию, Северную и Южную Америку, в Россию. Известны находки его в Краснодарском и Ставропольском краях, Волгоградской области, Ингушетии, Азербайджане, Крыму, Грузии и Дагестане, Армении. Однохозяинный вид. Основной хозяин – КРС. Является переносчиком многих кровепаразитарных заболеваний сельскохозяйственных животных [Балашов, 1998].
Dermacentor reticulatus. Встречается в Великобритании, Испании, Франции, Германии, Бельгии, Швейцарии, Польше, Венгрии, бывшей Югославии, Румынии [Колонин, 1984]. Найден в Краснодарском и Ставропольском краях, Волгоградской области, в Армении и Крыму. Обитает в лиственных лесах, лесостепи и степи [Тохов, 2004]. Треххозяинный вид. Взрослые клещи питаются на крупных и средних млекопитающих, а также на собаках, зайцах и ежах. Цикл развития одногодичный, но может растянуться на 2 года. Является переносчиком возбудителей туляремии, клещевого энцефалита и кровепаразитарных заболеваний скота [Балашов, 1998].
Dermacentor marginatus. Широко распространен в Европе, в Африке, в Азии. Также широко распространен в странах бывшего СССР. На Северном Кавказе и прилегающих регионах встречается часто. Обитает в лесостепной, степной и полупустынной зонах. Треххозяинный вид. Является переносчиком возбудителя Крымской геморрагической лихорадки, туляремии и возбудителей многих кровепаразитарных заболеваний скота [Балашов, 1998].
Dermacentor niveus. Встречается в Турции, Иране, Афганистане, Китае, Монголии. На территории бывшего СССР не встречается. Из прилегающих к Северному Кавказу территорий обитает в Дагестане. Обнаружен в Ставропольском крае. Обитатель зоны пустынь, где населяет долины рек, оазисы, плавни. В горы до высоты 3500 м поднимается по опустыненным участкам. Обычный, а в долинах и плавнях – многочисленный вид. Треххозяинный вид. Является переносчиком Крымской геморрагической лихорадки, клещевого энцефалита и пироплазмозов [Балашов, 1998].
Haemaphysalis concinna. Область распространения лежит между Тихим и Атлантическим океанами и представляет собой несколько изолированных обширных территорий. На сопредельных Северному Кавказу территориях обнаружен: в Краснодарском крае, Волгоградской области, Крыму, Ингушетии, Калмыкии, Дагестане, Азербайджане, Армении и в Грузии. Треххозяинный вид с пастбищным типом паразитизма. Переносчик возбудителя бруцеллеза [Балашов, 1998].
Haemaphysalis inermis. Распространен в Центральной и Южной Европе, Турции. Из республик бывшего СССР – в Молдавии. На Северном Кавказе и сопредельных территориях обнаружен в Ставропольском крае и Дагестане [Климова и др., 2005], Армении [Манукян и др., 2006], Азербайджане [Кулиева и др., 1976], Грузии [Балашов, 1998] и Крыму [Евстафьев, 2001]. Населяет широколиственные и смешанные горные и равнинные лесные массивы, обитает в степных биотопах и на осветленных склонах с разреженной древесно-кустарниковой растительностью. Треххозяинный вид с пастбищным типом паразитизма. Является переносчиком Крымской геморрагической лихорадки [Балашов, 1998].
Haemaphysalis punctata. Встречается почти по всей Европе. На Кавказе и сопредельных территориях эти пятна включают: Ставропольский и Краснодарский края, Ростовскую и Волгоградскую области. Этот вид распространен преимущественно в островных дубовых лесах, по долинам рек, в кустарниковых и открытых лугово-степных местообитаниях [Емчук, 1960; Успенская, 1987]. Треххозяинный вид с пастбищным типом паразитизма. Переносчик возбудителей Крымской геморрагической лихорадки, лихорадки Ку, спонтанный носитель возбудителей Лайм-боррелиоза, туляремии [Балашов, 1998].
Haemaphysalis sulcata. Встречается в Южной Европе, на Ближнем и Среднем Востоке, в Северной Африке. На юге России встречается в Дагестане. Обитатель предгорных и горных полупустынных и сухостепных биотопов. По межгорным долинам поднимается до 2500 м над ур. м. [Гребенюк, 1966]. Треххозяинный вид с пастбищным типом паразитизма. Является переносчиком возбудителя анаплазмоза овец [Балашов, 1998].
Haemaphysalis parva. Встречается в Европе, Северной Африке, Передней Азии. В бывшем СССР – в Молдавии, на Украине. На Кавказе и прилегающих территориях – в Армении, Грузии, в Крыму, Ингушетии, Дагестане, в Ставропольском крае, Волгоградской области. Встречается на пастбищах, близ сельхозугодий. Треххозяинный вид с пастбищным типом паразитизма. В Ставропольском крае установлено спонтанное носительство этим видом возбудителя туляремии. Зарегистрирован как переносчик пироплазмоза овец [Балашов, 1998].
Haemaphysalis pospelovashtromae. Из стран бывшего СССР встречается в Казахстане. На Кавказе – Дагестане, Грузии, Армении, Азербайджане. Треххозяинный вид с пастбищным типом паразитизма. В эксперименте показана возможность инфицирования бруцеллами всех фаз жизненного цикла при питании на зараженных морских свинках [Гребенюк, 1966].
Haemaphysalis caucasica. Встречается на Кавказе и прилегающих территориях: в Дагестане, Крыму, Грузии, Азербайджане и Карачаево-Черкесии. Населяет предгорные и горные островные леса (до 2200 м над ур. м.), заросли кустарника, перемежающиеся со степями и лугами. Треххозяинный вид с пастбищным типом паразитизма. Медицинское значение не изучено [Балашов, 1998].
Hyalomma marginatum. Имеет обширный ареал, включающий Южную Европу, Казахстан, Переднюю и Среднюю Азию, Индостан и большую часть Африки, Кавказ. В России его распространение ограничено полупустынными и степными ландшафтами на юге Европейской части. Встречается по всему Предкавказью и в предгорьях Большого Кавказа. Численность клещей нарастает от предгорных и лесостепных ландшафтов к полупустынным. H. marginatum не характерен для среднегорий и высокогорий [Емельянова, 2006]. Двуххозяинный вид. Взрослые клещи паразитируют, главным образом, на крупном и мелком рогатом скоте, лошадях, собаках и свиньях; активно нападают на людей. Является основным переносчиком Крымской геморрагической лихорадки.
Hyalomma scupense. Ареал – Африка, Европа, Азия, Индия. На Кавказе и прилегающих территориях обитает в Дагестане, Азербайджане, Крыму, Ростовской и Волгоградской областях, Ставропольском и в Краснодарском краях. Однохозяинный вид. Основными прокормителями являются КРС, овцы, козы, лошади, косуля. Из клещей этого вида выделены возбудители Крымской геморрагической лихорадки [Балашов, 1998].
Hyalomma aegyptium. Встречается в Средиземноморье, Закавказье, Иране и средней Азии [Филиппова, 1997]. Половозрелые особи паразитируют, в основном, на черепахах, а личинки и нимфы – на мелких млекопитающих и птицах [Hoogstraal, Kaiser, 1960]. Треххозяинный вид с одногодичным циклом развития. Весной, на активизировавшихся после зимней спячки черепахах последовательно сменяются питающиеся личинки, нимфы и в начале лета – имаго. Медицинское значение не известно.
Hyalomma asiaticum. Этот вид широко представлен от Сирии на западе до Монголии и Центрального Китая на востоке, от Казахстана на севере до побережья Персидского залива на юге. На территории бывшего СССР обнаружен в Казахстане, Закавказье, Армении [Манукян и др., 2006]. Обитатель пустынных ландшафтов. Треххозяинный вид. Тип паразитизма пастбищный. Массовое паразитирование имаго на скоте наблюдается в апреле-мае и сентябре-октябре. Неполовозрелые фазы паразитируют на мелких млекопитающих, личинки – с марта по сентябрь с максимумом в июне-июле, нимфы – с февраля по ноябрь с максимумом в августе-сентябре.
Hyalomma anatolicum. Ареал охватывает Средиземноморье и Северную Африку почти дл экватора, Аравийский полуостров, Иран, Пакистан, Индию и Среднюю Азию [Балашов, 1998]. На территории Кавказа встречается в Ставропольском крае [Емельянова, 2006] и Дагестане [Климова и др., 2005]. Двуххозяинный вид, все активные фазы развития паразитируют на КРС. Неполовозрелые фазы питаются также на хищниках, ежах, зайцах и мелких грызунах. Продолжительность развития – 1 год. Является одним из переносчиков Крымской геморрагической лихорадки.
Hyalomma detritum. Имеет сходный с H. anatolicum ареал [Балашов, 1998]. Встречается в Ставропольском крае, Дагестане [Климова и др., 2005] и Азербайджане [Кулиева и др., 1976]. Тяготеет к орошаемой зоне, долинам рек. Двуххозяинный вид. Прокормителями служат парнокопытные и непарнокопытные, главным образом, КРС. Медицинское значение неизвестно.
Rhipicephalus sanguineus. Ареал включает следующие страны: Португалия, Испания, Франция, Югославия, Греция, Болгария, Румыния, Турция, Сирия, Израиль, Ливан, Тайвань, Аргентина, Гавана, Бразилия, страны Африки. На Северном Кавказе и прилегающих территориях встречается в Ставропольском и Краснодарском краях, Дагестане, Армении, Азербайджане, Крыму. Треххозяинный вид. Переносчик возбудителя Крымской геморрагической лихорадки, Марсельской лихорадки, пироплазмоза собак [Балашов, 1998].
Rhipicephalus rossicus. Из всех представителей данного рода дальше всех проникает на север. Встречается на Северном Кавказе и прилегающих регионах: в Ростовской и Волгоградской областя, Дагестане, Ингушетии, Калмыкии, Грузии, Азербайджане, Крыму. Является обитателем степей и полупустынь. Треххозяинный вид. Цикл развития – 2–3 года. Переносчик возбудителей Крымской геморрагической лихорадки, лихорадки КУ и туляремии [Балашов, 1998].
Rhipicephalus pumilio. Обитает в Китае. На территории России обнаружен в Ставропольском крае, Волгоградской области. На Кавказе – в Дагестане. Взрослые клещи паразитируют на разнообразных млекопитающих, как крупных, так и мелких. Могут нападать на человека. Переносчик возбудителей Крымской геморрагической лихорадки, Ку-лихорадки и туляремии [Балашов, 1998].
Rhipicephalus schulzei. Обитает в Китае и Иране. На Кавказе и прилегающих территориях обнаружен в Краснодарском крае, Волгоградской области, Ингушетии, Калмыкии, Дагестане, Азербайджане. Обитатель полупустынь, пустынь, сухих степей. Из клещей выделены возбудители чумы, туляремии и лихорадки Ку [Балашов, 1998].
Rhipicephalus bursa. Встречается в Португалии, Испании, Франции, Италии, Швейцарии, Югославии, Албании, Болгарии, Греции, Марокко, Алжире, Турции, Иране, Сирии, Израиле, Китае и Монголии. На Кавказе и прилегающих регионах обнаружен в Армении, Ингушетии, Калмыкии, Дагестане, Азербайджане, Крыму. Населяет равнины, нижний и средний пояс гор. Двуххозяинный вид. Цикл развития одногодичный. Переносчик возбудителей Крымской геморрагической лихорадки, КЭ и кровепаразитарных заболеваний скота [Балашов, 1998].
Rhipicephalus turanicus. Найден в Португалии, Испании, Франции, Италии, Югославии, Албании, Греции, Сицилии, Кипре, Марокко, Алжире, Тунисе, Египте, Турции, Сирии, Израиле, Иране, Афганистане. На Кавказе обнаружен в Азербайджане и Дагестане. Населяет полупустыни, сухие редколесья, заросли кустарников, заходит в пустыни. В горы поднимается до 3000 м над ур. м. Треххозяинный вид. Переносчик возбудителей гемоспоридиозов домашнего скота и Крымской геморрагической лихорадки [Балашов, 1998].
Ixodes ricinus. Ареал – от Англии и Ирландии на западе до Волги на востоке. На Юге России I. ricinus проникает за границы широколиственных лесов и многочисленен в лесостепи и в разных типах искусственных лесонасаждений степной зоны. Широко распространен в Предкавказье. Треххозяинный вид с пастбищным типом паразитизма. I. ricinus – важнейший переносчик и резервуар возбудителей [вирусов, бактерий, простейших] многочисленных инфекционных болезней человека, в том числе клещевого энцефалита, Лайм-боррелиоза [Балашов, 1998].
Ixodes trianguliceps. Распространен в Палеарктике от Великобритании до реки Енисей на востоке в разных типах равнинных и горных лесов. Обнаружен в Крыму [Евстафьев, 2001]. Продолжительность жизненного цикла варьирует от 2 до 5 лет. К зимовке способны напитавшиеся и голодные особи на всех фазах развития, включая яйца. Свойственен «примитивный» тип пастбищного паразитизма, при котором активные перемещения клещей ограничены верхним слоем подстилки и припочвенным ярусом растительности. Медицинское значение не установлено.
Ixodes redikorzevi. На Кавказе обнаружен в Ставропольском крае [Тифлова и др., 1970], Азербайджане [Кулиева и др., 1976] и Крыму [Евстафьев, 2001]. Клещи паразитируют на грызунах, млекопитающих и птицах. Носитель возбудителя туляремии в природных очагах инфекции.
По ландшафтно-экологическим зонам клещи Северного Кавказа и сопредельных территорий распределяются следующим образом. На пустынных и полупустынных территориях ведущее место занимает H. marginatum. В долинах рек встречаются: H. anatolicum, H. detritum, H. marginatum, B. annulatus и R. turanicus. В степях обитают: D. marginatus, D. niveus, H. sulcata, H. punctata, R. rossicus, H. marginatum, H. scupense, R. turanicus, R. bursa, R. sanguineus, R. pumilio, H. anatolicum, H. aegyptium, B. annulatus, I. apronophorus.
2 Физико-географическая характеристика района исследований
В физико-географическом отношении Кавказ с севера на юг условно делится на три крупных региона: Предкавказье, Большой Кавказ и Закавказье. На северном склоне Западного Кавказа размещается Карачаево-Черкесская республика. Протяженность с севера на юг составляет около 400 км, с запада на восток – около 360 км. Площадь республики – 14,2 тысяч км2. На севере республика граничит с Кочубеевским и Андроповским районами, на северо-востоке и востоке – с Предгорным районом Ставропольского края и Кабардино-Балкарской республикой; на юге – с Грузией (Абхазия), на западе – с Краснодарским краем [Эколого-географический атлас, 2001].
В республике выделено 8 административных районов: Адыге-Хабльский, Прикубанский, Усть-Джегутинский, Хабезский, Зеленчукский, Карачаевский, Малокарачаевский и Урупский. Имеется 4 города: г. Черкесск – столица республики, находится на территории Прикубанского района, г. Усть-Джегута, расположен на территории Усть-Джегутинского района, г. Теберда и Карачаевск – на территории Карачаевского района.
Рельеф республики разнообразен. Поверхность региона гористая, самая низкая точка лежит на высоте 400 над уровнем моря. По рельефу в республике можно выделить три зоны: в северной части – предгорная, в средней – низкогорная, в южной – высокогорная. В высокогорной части республики население не живет.
По данным государственного учета земель, земельный фонд республики на 01.01.1999 г. составил 1418 тыс. га. В предгорной равнинно-холмистой части особенно на водоразделах, находятся так называемые черноземы предкавказские, в поймах рек – аллювиально-луговые почвы, на водоразделах предгорья – горные черноземы. В лесном поясе гор в основном горно-лесные почвы, а в субальпийском и альпийском поясах – луговые бурые почвы. Анализ качественного состава земель показывает, что в республике прогрессируют процессы засоления, заболачивания почв, ухудшая их качество. Большой ущерб плодородию наносит эрозия почв, которой затронуто 548426 га. Современное состояние земель продолжает ухудшаться, кислотность почв увеличивается. Агроклиматические ресурсы республики весьма благоприятны для сельского хозяйства и, в первую очередь, для животноводства. Почти вся хозяйственная деятельность сосредоточена на территории, занимающей менее 30% площади республики, в основном в речных долинах.
Современное оледенение Кавказа занимает площадь 1800 км2. Несмотря на то, что ледники в настоящее время переживают стадию отступления (таяния), общий их объем составляет 142 км3, или 120 км3 воды. В бассейне реки Кубани ледниками покрыто 220 км3. В республике насчитывается более 200 крупных озер, главным образом ледникового происхождения, 419 больших и малых рек с их притоками. Гидрографическая сеть представлена крупными горными реками – Кубань, Теберда, Аксаут, Маруха, Большой Зеленчук, Уруп, Большая Лаба, Кума, Подкумок и их притоки [Канонников, 1977]. Они относятся к группе высокогорных рек со смешанным питанием – ледниковые, грунтовые, дождевые и снеговые.
Реки на территории республики расположены неравномерно. Имеются пространства бедные или вообще лишенные водотоков. На территории республики располагаются 3 водохозяйственных водоема питьевого назначения – водохранилища Эшкаконское (Малокарачаевский район), головное (Усть-Джегутинский район) и Кубанское (Прикубанский район), а также Большой Ставропольский канал.
Лес – один из главных компонентов ландшафта как равнинных, так и горных районов КЧР от северных границ республики до субальпийского пояса на юге, от 400 до 2400 м над ур. м. Лесистость в среднем составляет 30,1%, площадь лесов – 416201 га. В регионе произрастает более 1000 видов деревьев, кустарников и полукустарников.
Территорию республики по рельефу, климату, растительному и животному миру условно делят на 3 зоны – степную предгорную, лесостепную низкогорную и горную.
Степная предгорная зона занимает северную часть республики с высотами от 400 до 700 м, в основном, территорию Адыге-Хабльского и Прикубанского районов. Сплошной степной зоны в КЧР нет, она представляет собой территорию возделывания многих сельскохозяйственных культур с изолированными степными участками. Среди равнинной местности имеются холмы разной высоты, покрытые порослью фруктовых и диких деревьев, заброшенными садами. Большая часть равнины распахана и оконтурена лесополосами.
Лесостепная низкогорная зона включает Усть-Джегутинский, Хабезский, Зеленчукский, Урупский, Карачаевский и Малокарачаевский районы. Высота 800–1500м над уровнем моря с постепенным повышением к югу. В пределах зоны возвышаются Пастбищный и Скалистый хребты, высотой до 1500 м. Поверхность пересечена балками, короткими ущельями, по которым протекают реки. Из растений встречаются многие виды лекарственных, злаковых, одно - и двулетники, многолетники. Имеются леса (ольха, ива, тополь-белолистка и черный ясень). В нижнем ярусе произрастают кустарники (боярышник, бузина, калина, терновник, ежевика, облепиха). Из вьющихся растений можно встретить хмель, плющ и дикий виноград.
В высокогорной зоне (3200–4047 м) население не проживает, поэтому интереса для нашего исследования она не представляла.
- Материал и методы исследований
Исследования проводили в период с марта по май 2013 г. в 2 климатогеографических районах Карачаево-Черкесской республики – Прикубанском (степном) и Зеленчукском (лесостепном) (рисунок 1).
1 –Абазинский район; 2 – Адыге-Хабльский район; Ногайский район;
а – п. Кавказский; б – п. Новый; в – п. Мичуринский; г – ст. Зеленчукская;
д – с. Кардоник; е – п. Даусуз.
Рисунок 1 – Административно-территориальные районы Карачаево-Черкесской республики (А, Б), входящие в зону проведения исследований фауны Ixodidae в 2013 г. и пункты сбора материала (а–е).
Сбор, учет и подготовка к лабораторным исследованиям кровососущих членистоногих проводили по стандартным методикам [Методические указания …, 2002; Методические рекомендации…, 2003].
Проводили сбор и учет численности иксодовых клещей с крупных млекопитающих. В период максимальной активности клещей проводили учеты численности. Особое внимание при сборе клещей уделяли местам их концентрации на прокормителе: шея, подгрудок, ушные раковины, веки, подмышечные впадины, вымя, основание и конец хвоста. Присосавшихся иксодовых клещей снимали жестким пинцетом, ухватив за основание хоботка или руками в тонких резиновых перчатках. Отрывали клещей при любом способе не резко, а осторожно, расшатывающими движениями или вращали клеща вокруг продольной оси тела, чтобы не оторвать хоботок. Место прикрепления клеща смазывали маслом, вазелином, это способствует его отпадению. Клещей, собранных с животных, помещали во флаконы, для дальнейшего определения видового состава.
В исследуемых нами районах было выбрано по 3 населенных пункта, в каждом из которых выделены контрольные группы ежедневно выпасаемого крупного рогатого скота (КРС), по одной группе, состоящей из 3 коров. Осмотры коров проводились каждые две недели с марта по июнь.
В природе голодных иксодовых клещей во всех фазах развития можно обнаружить на траве, ветках кустарников, поверхности почвы, где они сосредотачиваются в ожидании прокормителя. Сборы клещей в солнечную погоду проводили в утренние и вечерние часы при отсутствии росы и при слабом ветре. В пасмурные дни хорошие сборы давали дневные часы. Учеты на маршрутах позволили оценить видовой состав и распределение в зоне обследования. Клещей собирали на флаг, который представляет собой кусок материи длиной 1 м и шириной 60 см прикрепленный узкой стороной к палке. Развернутый флаг протаскивали сбоку, ведя подсчет шагов по 25-м отрезкам (обычно 16–18 пар шагов), в промежутках между которыми осматривали флаг и одежду. На каждом маршруте при учетах было набрано не менее 1000 м расстояния и 2 ч времени. Обилие клещей выражают числом особей, собранных с флага и одежды учетчика на 1 км маршрута.
В настоящее время в паразитологии широко применяются такие показатели численности паразитов как: экстенсивность инвазии, интенсивность инвазии, индекс обилия и индекс доминирования, которые, являются показателями зараженности хозяина.
Экстенсивность инвазии, или встречаемость, есть процент зараженных хозяев (в нашем случае коров и маршрутов) конкретным видом или группой паразитов:
, (1)
где Np – число зараженных хозяев;
n – общее число хозяев.
Интенсивность инвазии – среднеарифметический показатель числа паразитов, приходящийся на одну зараженную особь хозяина:
, (2)
где Par – число обнаруженных паразитов;
Np – зараженные хозяева.
Индекс обилия – средняя численность определенного вида или группы паразитов у всех особей хозяина (включая незараженных):
, (3)
где Par – число обнаруженных паразитов;
n – обследованные животные.
Индекс доминирования – доля (в %), которую составляет обилие исследуемого вида по отношению к суммарному обилию всех сравниваемых между собой видов в изучаемом материале:
(4)
где n – число особей какого-либо вида;
N – общее число видов в биоценозе.
Всего за этот период было собрано 1656 экз. иксодовых клещей, из которых – 721 снято с крупного рогатого скота, 935 собрано на флаг.
Таблица 2 – Объём материала полевых исследований
|
Виды исследований
|
Количество
|
|
Основные учеты
|
|
Число обследованных населенных пунктов (с учетом кратности)
|
6 (36)
|
|
Количество фл./км (с учетом кратности)
|
6 (36)
|
|
Число контрольных животных (с учетом кратности)
|
18 (108)
|
|
Собрано и определено имаго, нимф и личинок клещей
|
|
с КРС
|
721
|
|
на флаг
|
935
|
|
Всего:
|
1656
|
Определение собранных клещей проведено с помощью определителей «Иксодовые клещи (Ixodidae)» [Померанцев, 1950] и «Фауна России и сопредельных стран» [Филиппова, 1997] и при участии энтомолога Санэпидемстанции г. Черкесска – Плясуновой С.Б.
4 Фаунистический комплекс Ixodidae Карачаево-Черкесской республики
4.1 Состав фауны иксодид
Таксономический состав фауны клещей степной и лесостепной зон Карачаево-Черкесии следующий: в степной зоне обнаружены в п. Кавказский (D. marginatus, D. reticulatus, H. punctata, I. ricinus, H. parva, H. сoncinna, H. scupense), п. Мичуринский (D. marginatus, D. reticulatus), п. Новый (D. marginatus, D. reticulatus). Видовой состав представлен 7 видам клещей из 4 родов: 3 вида из рода Haemaphysalis, 2 вида из рода Dermacentor, по 1 виду из родов Ixodes и Hyalommа.
В лесостепной зоне обнаружены в ст. Зеленчукская (D. marginatus, D. reticulatus), п. Даусуз (D. marginatus, D. reticulatus, I. ricinus), с. Кардоник (D. marginatus, D. reticulatus). Видовой состав представлен 3 видами клещей из 2 родов: 2 вида из рода Dermacentor, 1 вид из рода Ixodes. Клещи D. marginatus. Всего нами обнаружено 7 видов клещей из 4 родов, систематическое положение которых представлено ниже.
Тип Arthropoda
Класс Arachnida
Подкласс Acari
Надотряд Parasitiformes
Отряд Ixodida
Надсемейство Ixodoidea
Семейство Ixodidae
Род Dermacentor
Dermacentor marginatus
Dermacentor reticulatus
Род Нaemaphysalis
Нaemaphysalis concinna
Нaemaphysalis punctata
Нaemaphysalis parva
Род Hyalomma
Hyalomma scupense
Род Ixodes
Ixodes ricinus
Мы изучили по литературным источникам фауну иксодид Северного Кавказа и других сопредельных территорий со сходными природными условиями и сравнили с полученными нами данными (таблица 3).
Таблица 3 – Иксодовые клещи Северного Кавказа и прилегающих регионов
|
№
п.п.
|
Вид
|
КЧР (по нашим данным)
|
Краснодарский край
|
Ставропольский край
|
Ростовская обл.
|
Волгоградская обл.
|
Ингушетия
|
Дагестан
|
Калмыкия
|
Азербайджан
|
Крым
|
Грузия
|
Армения
|
КЧР (по данным источников)
|
|
1
|
Boophilus annulatus
|
|
+
|
+
|
|
+
|
+
|
+
|
|
+
|
+
|
|
+
|
+
|
|
2
|
Dermacentor marginatus
|
+
|
+
|
+
|
+
|
+
|
|
+
|
|
+
|
+
|
|
+
|
+
|
|
3
|
Dermacentor reticulatus
|
+
|
+
|
+
|
|
+
|
|
|
|
|
+
|
|
+
|
+
|
|
4
|
Dermacentor niveus
|
|
|
+
|
|
|
|
+
|
+
|
|
|
|
|
|
|
5
|
Нaemaphysalis concinna
|
+
|
+
|
|
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
|
6
|
Нaemaphysalis punctata
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
|
7
|
Нaemaphysalis inermis
|
|
|
+
|
|
|
|
+
|
|
+
|
+
|
+
|
+
|
|
|
8
|
Нaemaphysalis parva
|
+
|
|
+
|
|
+
|
+
|
+
|
|
|
+
|
+
|
+
|
+
|
|
9
|
Нaemaphysalis sulcata
|
|
|
|
|
|
|
+
|
|
+
|
+
|
|
+
|
|
|
10
|
Нaemaphysalis pospelovashtromae
|
|
|
|
|
|
|
+
|
|
+
|
|
+
|
+
|
|
|
11
|
Нaemaphysalis caucasica
|
|
|
|
|
|
|
+
|
|
+
|
+
|
+
|
|
|
|
12
|
Hyalomma asiaticum
|
|
|
|
|
|
|
|
|
|
|
|
+
|
|
|
13
|
Hyalomma aegyptium
|
|
|
|
|
|
|
|
|
+
|
|
|
|
|
|
14
|
Hyalomma marginatum
|
|
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
|
|
+
|
|
15
|
Hyalomma anatolicum
|
|
|
+
|
|
|
|
+
|
|
|
|
|
|
|
|
16
|
Hyalomma scupense
|
+
|
+
|
+
|
|
+
|
|
+
|
|
+
|
+
|
|
|
+
|
|
17
|
Hyalomma detritum
|
|
|
+
|
|
|
|
+
|
|
+
|
|
|
|
|
|
18
|
Rhipicephalus bursa
|
|
|
|
|
|
+
|
+
|
+
|
+
|
+
|
|
+
|
|
|
19
|
Rhipicephalus sanguineus
|
|
+
|
+
|
|
|
|
|
|
+
|
+
|
|
|
|
|
20
|
Rhipicephalus pumilio
|
|
|
+
|
|
|
|
|
|
+
|
+
|
|
|
|
|
21
|
Rhipicephalus schulzei
|
|
+
|
|
|
+
|
+
|
|
+
|
+
|
|
|
|
|
|
22
|
Rhipicephalus rossicus
|
|
|
+
|
+
|
+
|
+
|
|
+
|
+
|
+
|
+
|
|
+
|
|
23
|
Rhipicephalus turanicus
|
|
|
|
|
|
|
|
+
|
+
|
|
|
|
|
|
24
|
Ixodes trianguliceps
|
|
|
|
|
|
|
|
|
|
+
|
|
|
|
|
25
|
Ixodes redikorzevi
|
|
|
|
|
|
|
|
|
+
|
+
|
|
|
|
|
26
|
Ixodes ricinus
|
+
|
+
|
+
|
+
|
+
|
|
|
|
|
|
|
|
+
|
Ранее в КЧР описаны 10 видов клещей из 6 родов [Новикова, 2008]. Нами обнаружено 7 видов клещей из 4 родов. В сборах не найдены: H. marginatum, R. rossicus, B. annulatus. По имеющимся данным клещи H. marginatum и R. rossicus относятся к крайне редким видам в республике, кроме этого, клещи H. marginatum, встречаются в сборах не каждый год, в зависимости от протекания морозного периода, т.е. данный вид весьма чувствителен к низким температурам. В нашем случае зима была достаточно холодной, что повлияло на отсутствие клеща в сборах. Отсутствие B. annulatus объясняется проведением ветеринарной службой в этот период обработки скота акарицидными препаратами, что повлекло гибель всех фаз B. annulatus, и т.к. он является однохозяйным видом, его распространение стало невозможно. Что касается клещей прилегающих территорий, не обнаруженных нами в сборах, можно отметить H. аegyptium, который является паразитом средиземноморской черепахи, которая в КЧР не водится.
В Краснодарском крае обнаружено 9 видов клещей, в Ставропольском крае – 15 видов, в Волгоградской области – 11 видов, в республике Ингушетия – 8 видов, в Калмыкии – 8 видов, в Дагестане – 15 видов, в Армении – 12 видов, в Азербайджане – 19 видов, в Грузии – 7 видов, в Крыму – 17 видов.
Таким образом, видовой состав клещей в этих регионах в зоне степей и лесных низкогорных зонах не отличается от наших данных, а большее видовое разнообразие в прилегающих к КЧР регионах соотносится с большим числом климатических зон.
По ландшафтно-экологическим зонам клещи Северного Кавказа и сопредельных территорий распределяются следующим образом: в степях обитают D. marginatus, H. punctata, H. scupense, B. annulatus, основу населения лесных участков составляют I. ricinus, D. reticulatus, H. сoncinna, в широколиственных лесах обитает Н. concinna, в смешанных лиственных и широколиственных лесах (редколесьях, вырубках) – D. reticulatus. В искусственных лесонасаждениях, парках – встречаются I. ricinus и D. reticulatus.
4.2 Интенсивность и экстенсивность клещевой инвазии
Данные по сборам клещей в степной и лесостепной зонах Карачаево-Черкесской республики отражены в таблице 4, которая демонстрирует интенсивность общей клещевой инвазии.
Таблица 4 – Интенсивность клещевой инвазии животных в степной и лесостепной зонах Карачаево-Черкесии (01.03–09.05. 2013)
|
Живот-ное
|
Дата
|
Интенсивность инвазии, экз./животное
|
|
|
01.03
|
15.03
|
29.03
|
12.04
|
26.04
|
08.05
|
Всего
|
|
|
|
Степная зона (Прикубанский район КЧР)
|
|
|
1
|
2
|
2
|
5
|
11
|
29
|
9
|
58
|
9,7±4,14
|
|
2
|
1
|
3
|
5
|
9
|
18
|
12
|
48
|
8,0±2,58
|
|
3
|
1
|
4
|
7
|
14
|
21
|
10
|
57
|
9,5±2,95
|
|
4
|
1
|
3
|
6
|
9
|
11
|
6
|
36
|
6,0±1,51
|
|
5
|
2
|
2
|
6
|
8
|
19
|
11
|
48
|
8,0±2,62
|
|
6
|
1
|
2
|
4
|
9
|
25
|
13
|
54
|
9,0±3,70
|
|
7
|
3
|
3
|
3
|
10
|
16
|
7
|
42
|
7,0±2,14
|
|
8
|
2
|
4
|
4
|
13
|
30
|
6
|
59
|
9,8±4,32
|
|
9
|
0
|
3
|
4
|
8
|
41
|
4
|
60
|
10,0±6,29
|
|
Итого
|
13
|
26
|
44
|
91
|
210
|
78
|
462
|
–
|
|
|
Лесостепная зона (Зеленчукский район КЧР)
|
|
|
10
|
0
|
2
|
2
|
6
|
12
|
7
|
29
|
4,8±1,80
|
|
11
|
0
|
2
|
3
|
6
|
14
|
6
|
31
|
5,2±2,01
|
|
12
|
0
|
1
|
4
|
5
|
11
|
6
|
27
|
4,5±1,61
|
|
13
|
0
|
1
|
4
|
7
|
10
|
9
|
31
|
5,2±1,70
|
|
14
|
0
|
2
|
3
|
9
|
6
|
10
|
30
|
5,0±1,63
|
|
15
|
0
|
2
|
5
|
5
|
11
|
4
|
27
|
4,5±1,52
|
|
16
|
0
|
3
|
2
|
8
|
9
|
6
|
28
|
4,7±1,45
|
|
17
|
0
|
2
|
2
|
7
|
9
|
6
|
26
|
4,3±2,19
|
|
18
|
0
|
0
|
3
|
5
|
14
|
8
|
30
|
5,0±1,43
|
|
Итого
|
0
|
15
|
28
|
58
|
96
|
62
|
259
|
–
|
|
Всего
|
13
|
41
|
72
|
149
|
306
|
140
|
721
|
–
|
|
Примечание:
п. Кавказский № 1–3, п. Новый № 4–6, п. Мичуринский № 7–9,
ст. Зеленчукская № 10–12, с. Кардоник №13–15, п. Даусуз №16–18.
|
Сравнение интенсивности общей клещевой инвазии животных из степной и лесостепной зоны за период исследования с использованием критерия Стьюдента показало наличие достоверных различий данного показателя (Tst = 7,8; p < 0.001). Интенсивность общей клещевой инвазии в степной зоне (51,3±2,79 клещей на группу животных) практически в 2 раза выше таковой в зоне лесостепи (28,8±0,62), что мы связываем с особенностями природно-климатических зон, в которых проводили исследования.
Посёлки Кавказский, Новый, Мичуринский – относятся к степной предгорной зоне, занимающей северную часть республики с высотами от 400 до 700 м над уровнем моря. Травянистая растительность степных участков включает до 100 и более видов травянистых растений на 100 км – бобово-разнотравно-злаковых (роды кострец, коротконожка, тимофеевка, трясунка, пырей, шалфей, овсюг) и разнотравно-кустарниковых ожиновых. В этой зоне в настоящее время сохранились хомяки, полевые мыши. В лесополосах и глухих балках можно встретить лису, барсука, зайца, кабана. Из птиц отмечаются грачи, вороны, галки, сороки и другие [Ашибокова, 2007].
В летний период наиболее жарко на севере республики в ее степном Прикубанском районе. Этот район находятся в зоне с умерено-континентальным климатом. Самый холодный месяц в году – январь, среднемесячная температура которого составляет – 4С. Устойчивый снежный покров наблюдается лишь половину зимы. Наибольшая глубина промерзания почвы составляет 30–70 см. Зимой часто бывают оттепели (до 60 дней за сезон). В первой декаде марта происходит устойчивый переход среднесуточных температур через 0 С в сторону повышения. Средняя скорость ветра составляет 3 м/сек. Зимой и весной часто господствуют восточные и северо-восточные ветры, которые достигают 30 м/сек и более. Безморозный период – с середины апреля до октября. Лето сравнительно нежаркое. Годовое количество осадков в среднем составляет 500–550 мм.
Населенные пункты Зеленчукского района находятся в лесостепной низкогорной зоне. Высота 800–1500 м над уровнем моря с постепенным повышением к югу. В пределах зоны возвышаются Пастбищный и Скалистый хребты, высотой до 1500 м. Из растений встречаются многие виды лекарственных, злаковых, одно- и двулетники, многолетники. Здесь водятся кабаны, косули, лисы, волки и шакалы (единичны), ежи, зайцы, белки, хомяки, сони, лесная и домовая мыши, полевки обыкновенные, ящерицы, ужи [Ашибокова, 2007]. По мере продвижения к югу и увеличения высоты местности температура заметно понижается, причем в зимнее время меньше, чем в теплый период. Зеленчукский район расположен в умерено-теплом климате низкогорной части республики. Он свойственен высотам от 800 до 1200 м. Среднегодовая температура воздуха здесь 6–7 С. Зима длится 90–95 дней. Осадков в год выпадает 650–700 мм [Полтараус, 1972].
Таким образом, температура в степной зоне выше, чем в лесостепной, количество осадков меньше, зимний период мягче, сопровождается оттепелями, что сказывается на биологии клещей и их встречаемости.
В ходе наблюдений весной 2013 г. выявлена тенденция динамики клещевой инвазии. В конце апреля наблюдали всплеск численности клещей, в обеих зонах (рисунок 2). Данный факт мы объясняем концом морозного периода и выходом имаго клещей в активную фазу – кровососание. Далее, численность снижается, так как остается все меньше голодных клещей в природе. Тенденцию инвазии по датам мы объясняем увеличением температуры воздуха, и, соответственно, активности клещей.
Всего в степной зоне, по учетам на контрольных группах животных, обнаружено 462 клеща, в лесостепной зоне – 259 клещей. В степной зоне средняя интенсивность инвазии клещей на 1 животное составила 8,6±0,47 клещей (минимальное количество – 6, максимальное – 10), а в лесостепной зоне – 4,8±0,11 (минимальное количество – 4,3, максимальное – 5,2). Сравнение интенсивности клещевой инвазии в пересчёте на 1 животное из степной и лесостепной зоны с использованием критерия Стьюдента показало наличие достоверных различий данного показателя (Tst = 7,9; p < 0,001). Причины выявленных различий, на наш взгляд, аналогичны таковым для общей клещевой инвазии, что указано выше. Экстенсивность инвазии для обеих зон – 100% (рисунок 3)
Рисунок 2 – Весенняя динамика клещевой инвазии животных (контрольные группы) в степной и лесостепной зонах Карачаево-Черкесии
Рисунок 3 – Поражённость животных степной и лесостепной зон Карачаево-Черкесии клещами в весенний период
Достоверные различия интенсивности инвазии клещами в контрольных группах животных в марте-апреле выявлены для степной и лесостепной зон (таблица 5).
Таблица 5 – Сравнение интенсивности клещевой инвазии животных (контрольные группы) в степной и лесостепной зонах Карачаево-Черкесии по датам наблюдения
|
Дата наблюдения
|
Интенсивность инвазии,
особей/животное
|
Tst
|
|
|
степная зона
|
лесостепная зона
|
|
|
|
min–max
|
M±m
|
min–max
|
M±m
|
|
|
01.03.2013
|
0–3
|
1,4±0,29
|
–
|
0,0±0,00
|
4,9*
|
|
15.03.2013
|
2–4
|
2,9±0,26
|
0–3
|
1,7±0,29
|
3,14**
|
|
29.03.2013
|
3–7
|
4,9±0,42
|
2–5
|
3,1±0,35
|
3,23**
|
|
12.04.2013
|
8–14
|
10,1±0,72
|
5–9
|
6,4±0,48
|
4,27*
|
|
26.04.2013
|
11–41
|
23,3±3,00
|
4–14
|
10,7±0,85
|
4,06*
|
|
08.05.2013
|
4–13
|
8,7±1,03
|
4–10
|
6,9±0,61
|
1,49
|
|
Примечание: * – p < 0,001; ** – p < 0,01.
|
Отсутствие достоверных различий по 08.05.2013 в сравниваемых зонах, мы объясняем возрастающей долей сытых самок, некоторые из которых уже находились в стадии яйцекладки.
Отсутствие клещей H. scupense, H. concinna, Н. punctata и H. parva в сборах лесостепной зоны и уменьшение численности D. marginatus и D. retiсulatus, по сравнением со степной зоной мы объясняем более холодной зимой в лесостепной зоне (таблица 6). Выживает меньшее количество клещей, а также прокормителей (кровь лесных грызунов является пищей для личинок и нимф иксодового клеща).
Таблица 6 – Экстенсивность (%) и интенсивность (клещей на животное) клещевой инвазии по природно-климатическим зонам
|
Вид
|
Степная зона (Прикубанский район КЧР)
|
|
|
Животное
|
Всего
|
ЭИ, %
|
ИИ
|
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
|
|
|
|
D. marginatus
|
47
|
27
|
19
|
19
|
23
|
41
|
25
|
40
|
48
|
289
|
100
|
32,1±3,94
|
|
D. retiсulatus
|
8
|
18
|
35
|
15
|
23
|
12
|
16
|
19
|
12
|
158
|
100
|
17,6±2,63
|
|
H. scupense
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
11
|
0,1±0,11
|
|
H. concinna
|
0
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
11
|
0,1±0,11
|
|
Н. punctata
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
11
|
0,1±0,11
|
|
H. parva
|
0
|
0
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
11
|
0,1±0,11
|
|
I. ricinus
|
2
|
2
|
2
|
2
|
1
|
1
|
1
|
0
|
0
|
11
|
78
|
1,2±0,28
|
|
Вид
|
Лесостепная зона (Зеленчукский район КЧР)
|
|
|
Животное
|
Всего
|
ЭИ,
%
|
ИИ
|
|
|
10
|
11
|
12
|
13
|
14
|
15
|
16
|
17
|
18
|
|
|
|
|
D. marginatus
|
15
|
19
|
19
|
11
|
13
|
21
|
13
|
4
|
11
|
126
|
100
|
14,0±1,7
|
|
D. retiсulatus
|
13
|
12
|
8
|
20
|
17
|
6
|
15
|
22
|
19
|
132
|
100
|
14,7±1,8
|
|
H. scupense
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
–
|
–
|
|
H. concinna
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
–
|
–
|
|
Н. punctata
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
–
|
–
|
|
H. parva
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
–
|
–
|
|
I. ricinus
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
11
|
0,1±0,11
|
Для клещей D. marginatus и I. ricinus отмечены различия интенсивности инвазии ими животных в степной и лесостепной зонах (рисунок 4, 5). Интенсивность инвазии для обоих видов достоверна выше в степной зоне (Tst.= 4,20; p < 0,001 и Tst.= 3,71; p < 0,01). Мы связываем это с тем, что верхняя и нижняя границы жизни D. marginatus от -10 до 44 °С, а I. ricinus от -13 до 41°С (для взрослых особей) [Балашов, 1998], что и соответствует степной зоне.
Рисунок 4 – Интенсивность инвазии КРС клещом D. marginatus в степной и лесостепной зонах КЧР в весенний период
Рисунок 5 – Интенсивность инвазии КРС клещом I. ricinus в степной и лесостепной зонах КЧР в весенний период
Результаты получены нами в процессе сборов иксодовых клещей в обеих зонах КЧР на растительности, отраженные в таблице 7 и рисунке 6. В степной зоне, так же как и при сборах с животных нами было собрано большее число клещей (603 и 332). По данным источников, в степных районах КЧР количество клещей в сборах было выше, чем в лесостепной, причем это характерно для обоих методов [Новикова, 2008]. Нами получены аналогичные результаты.
Таблица 7 – Количество клещей в степной и лесостепной зонах по результатам учетов на флаг (01.03–09.05. 2013)
|
Маршрут
|
Дата
|
Количество клещей, ос./км
|
|
|
01.03
|
15.03
|
29.03
|
12.04
|
26.04
|
08.05
|
Всего
|
|
|
|
Степная зона (Прикубанский район)
|
|
|
1
|
10
|
14
|
35
|
57
|
91
|
14
|
221
|
36,8±13,05
|
|
2
|
11
|
19
|
38
|
64
|
63
|
13
|
208
|
34,7±9,92
|
|
3
|
5
|
14
|
41
|
49
|
56
|
9
|
174
|
29,0±9,08
|
|
Итого:
|
26
|
47
|
114
|
170
|
210
|
36
|
603
|
100,5±31,40
|
|
|
Лесостепная зона (Зеленчукский район)
|
|
|
4
|
0
|
11
|
23
|
33
|
65
|
8
|
140
|
23,3±9,59
|
|
5
|
0
|
6
|
15
|
18
|
43
|
11
|
93
|
15,5±6,09
|
|
6
|
0
|
21
|
14
|
11
|
38
|
15
|
99
|
16,5±5,14
|
|
Итого:
|
0
|
38
|
52
|
62
|
146
|
34
|
332
|
55,3±20,08
|
|
Всего:
|
26
|
85
|
166
|
232
|
356
|
70
|
935
|
|
|
Примечание:
Маршрут №1 – п. Кавказский, №2 – п. Новый, №3 – п. Мичуринский, № 4 – ст. Зеленчукская, № 5 – с. Кардоник, №6 – п. Даусуз.
|
Сравнение средних показателей численности клещей в степной и лесостепной зонах Карачаево-Черкесии по датам с использованием критерия Стьюдента показало наличие достоверных различий для 1 марта (Tst = 4,7; p < 0,01), 29 марта (Tst = 6,2; p < 0,01)и 12 апреля (Tst = 4,6; p < 0,01). Достоверные различия средних значений количества клещей по отдельным маршрутам и для зон в целом отсутствуют, причины нами не установлены.
Всего в степной зоне, по учетам на флаг, обнаружено 603 клеща, в лесостепной зоне – 332 клеща. В степной зоне среднее количество клещей на 1 км маршрута составило 33,5±2,33, а в лесостепной зоне – 18,4±2,45 клещей. Различия средних значений данного показателя для зон достоверны – Tst = 4,5; p < 0,05. Выживаемость клещей в более теплой степной зоне выше, чем в лесостепной. Наши данные, также совпадают с данными других авторов: для Кавказа и прилегающих территориях [Климова и др., 2005] Азербайджана [Кулиева и др., 1976], Крыма [Евстафьев, 2001], Ростовской и Волгоградской областей [Денисов и др., 2005] областях, Ставропольского и Краснодарского краёв [Костюковский и др., 2005].
Рисунок 6 – Численность клещей (ос./км) в степной и лесостепной зонах Карачаево-Черкесии в весенний период (по результатам учётов на флаг)
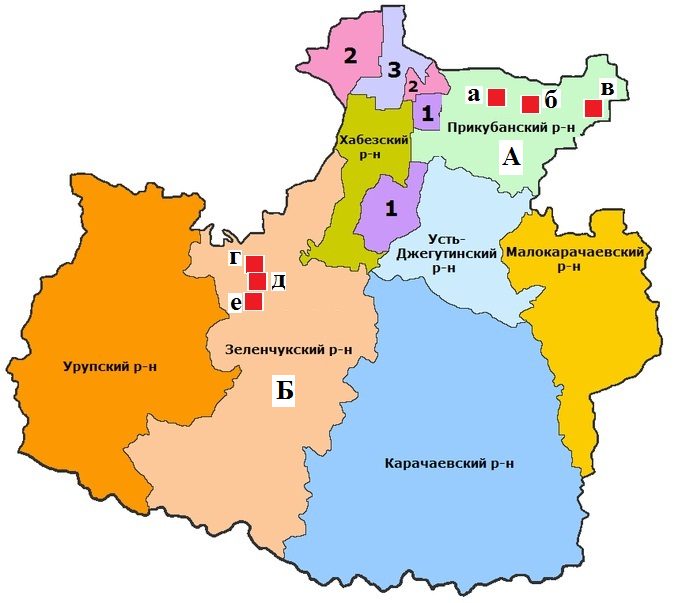
Данные о числе клещей, обилию и индексе доминирования в степной и лесостепной зоне по учетам на контрольных группах животных отображены в таблице 8. Нами установлено, что наиболее часто встречаемый вид в наших сборах – D. marginatus: индекс обилия в степной зоне составил 5,3, а в лесостепной зоне – 2,3, соответственно распределился и индекс доминирования по зонам: 62,5 и 48,4%. Меньший по численности вид D. retiсulatus, у которого индексы обилия и доминирования равны по степной зоне 2,9, а лесостепной 2,4. Индексы доминирования распределены следующим образом – 34,3 и 50,8%.
Таблица 8 – Обилие клещей в степной и лесостепной зонах по учетам на контрольных группах животных
|
Виды
|
Степная зона,
54 осмотренных животных с учетом кратности
|
Лесостепная зона,
54 осмотренных животных с учетом кратности
|
Всего
|
|
|
А
|
Б
|
В
|
А
|
Б
|
В
|
А
|
|
D. marginatus
|
289
|
5,3
|
62,5
|
126
|
2,3
|
48,4
|
415
|
|
D. retiсulatus
|
158
|
2,9
|
34,3
|
132
|
2,4
|
50,8
|
290
|
|
H. scupense
|
1
|
0,02
|
0,2
|
0
|
0
|
0
|
1
|
|
H. concinna
|
1
|
0,02
|
0,2
|
0
|
0
|
0
|
1
|
|
Н. punctata
|
1
|
0,02
|
0,2
|
0
|
0
|
0
|
1
|
|
H. parva
|
1
|
0,02
|
0,2
|
0
|
0
|
0
|
1
|
|
I. ricinus
|
11
|
0,2
|
2,4
|
1
|
0,02
|
0,8
|
12
|
|
Примечания:
А – число клещей, Б – индекс обилия, В – индекс доминирования, %.
|
Клещ I. ricinus – третий по численности вид клещей в степной и лесостепной зонах по учетам на контрольных группах животных, имеет индексы обилия по степной зоне – 0,2, а по лесостепной – 0,002%. Индекс доминирования по степной зоне составляет 2,4, а по лесостепной зоне 0,8. Клещи H. scupense, H. concinna, Н. punctate и H. parva обнаружены нами только в степной зоне. Индекс обилия для H. scupense – 0,2, индекс доминирования – 0,002%, для H. concinna – 0,2, индекс доминирования – 0,002%, для Н. punctata– 0,2, индекс доминирования – 0,002%, для H. parva – 0,2, индекс доминирования – 0,002%.
Наглядно, количественную тенденцию по результатам сборов в обеих зонах можно проследить на рисунке 7 и 8.
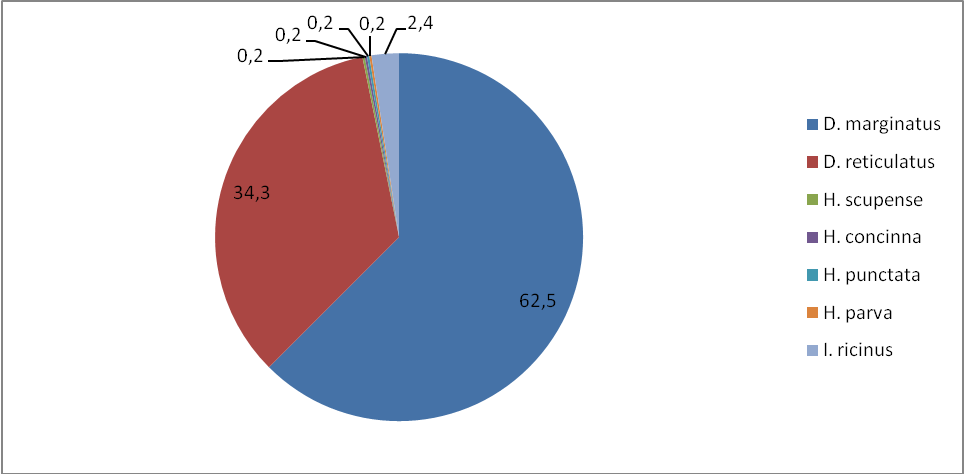
Рисунок 7 – Доля видов (в %) (индекс доминирования) для степной зоны по учетам на контрольных группах животных.
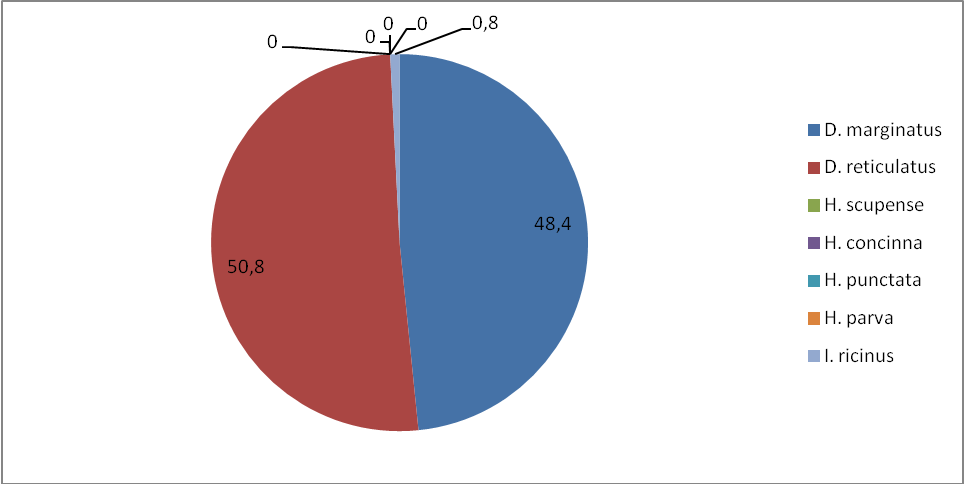
Рисунок 8 – Доля видов (в %) (индекс доминирования) для лесостепной зоны по учетам на контрольных группах животных.
Данные о числе, обилию, индексе по учетам на флаг на контрольных маршрутах отображены в таблице 9.
Таблица 9 – Обилие клещей в степной и лесостепной зонах по учетам на флаг
|
Виды
|
Степная зона,
18 фл/км
|
Лесостепная зона,
18 фл/км
|
Всего клещей
|
|
|
А
|
Б
|
В
|
А
|
Б
|
В
|
|
|
D. marginatus
|
254
|
14,1
|
42,1
|
183
|
10,2
|
55,1
|
437
|
|
D. retiсulatus
|
314
|
17,4
|
52
|
130
|
7,2
|
39,2
|
444
|
|
H. scupense
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
H. concinnа
|
1
|
0,05
|
0,2
|
0
|
0
|
0
|
1
|
|
Н. punctata
|
3
|
0,2
|
0,5
|
0
|
0
|
0
|
3
|
|
H. parva
|
1
|
0,05
|
0,2
|
0
|
0
|
0
|
1
|
|
I. ricinus
|
30
|
1,7
|
5
|
19
|
1,05
|
5,7
|
49
|
|
Примечания:
А – число клещей, Б – индекс обилия, В – индекс доминирования, %.
|
Установлено, что наиболее часто встречаемый видом, при сборе на флаг, также является – D. marginatus, маршруты проходили по местам обитания данного вида – увлажненные места: индекс обилия в степной зоне составил 14,1, а в лесостепной зоне – 10,2, индекс доминирования по зонам: 42,1 и 55,1%. Меньший по численности вид D. retiсulatus, у которого индексы обилия и доминирования равны по степной зоне 17,4, а лесостепной 7,2, индексы доминирования распределены следующим образом – 52 и 39,2%.
Клещ I. ricinus – третий по численности вид, имеет индексы обилия по степной зоне 1,7, по лесостепной – 1,05, доминирование 5 и 5,7 по степной и лесостепной зонам, соответственно. Клещи H. concinna, Н. punctata и H. parva обнаружены нами только в степной зоне. Индексы обилия – 0,05, 0,2, 0,05 и доминирования – 0,2, 0,5, 02. H. scupense – не обнаружен.
Наглядно, количественную тенденцию по результатам сборов в обеих зонах можно проследить на рисунке 9 и 10.
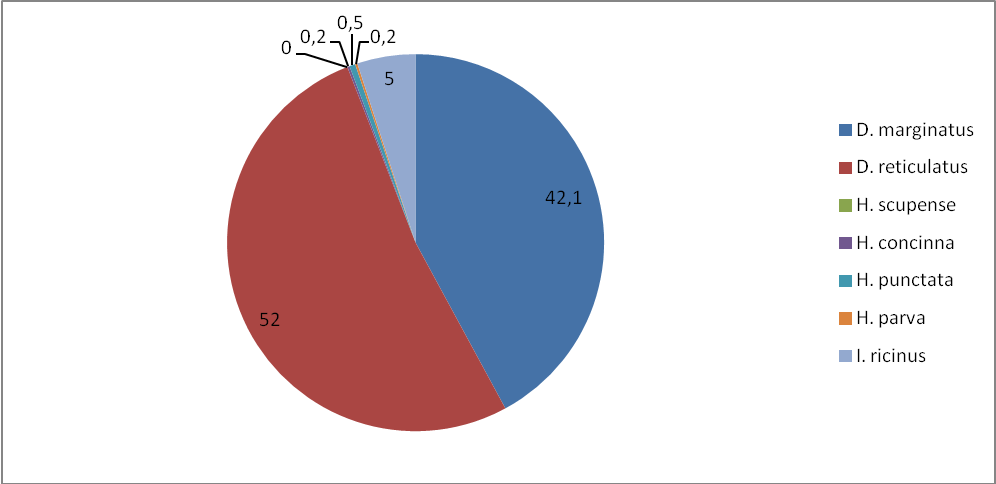
Рисунок 9 – Доля видов (в %) (индекс доминирования) для степной зоны по учетам на контрольных маршрутах.
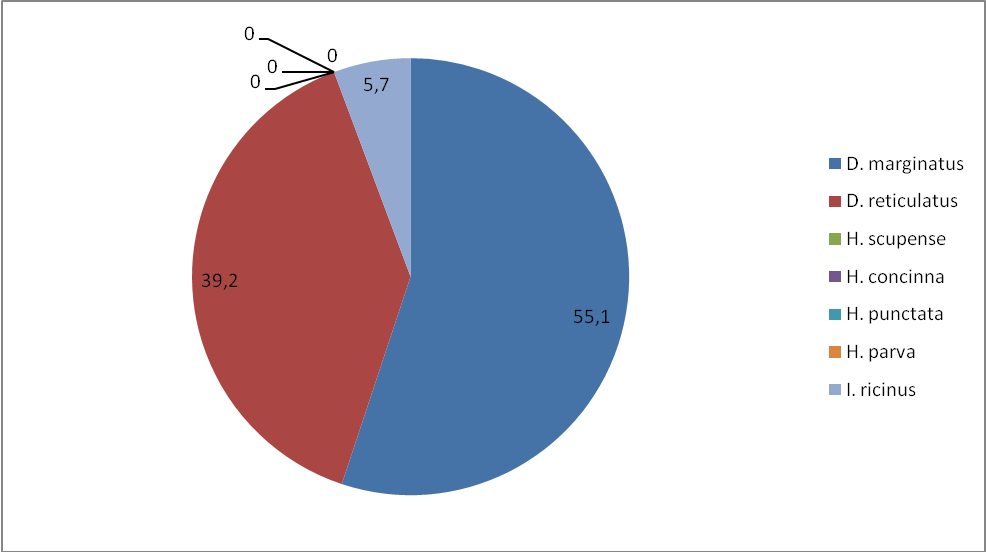
Рисунок 10 – Доля видов (в %) (индекс доминирования) для лесостепной зоны по учетам на контрольных маршрутах.
Данные о числе клещей, обилию и индексе доминирования в степной и лесостепной зоне по учетам на контрольных маршрутах и сборах на флаг отображены в таблице 10.
Таблица 10 – Обилие и доминирование клещей на контрольных группах животных и контрольных маршрутах
|
Виды
|
Сборы
|
|
|
Контрольные группы животных, 108 осмотренных животных
|
Контрольные маршруты,
36 фл/км
|
Всего
|
|
|
А
|
Б
|
В
|
А
|
Б
|
В
|
А
|
|
D. marginatus
|
415
|
3,8
|
57,6
|
437
|
12,1
|
46,7
|
852
|
|
D. retiсulatus
|
290
|
2,7
|
40,2
|
444
|
12,3
|
47,5
|
734
|
|
H. scupense
|
1
|
0,009
|
0,1
|
0
|
0
|
0
|
1
|
|
H. concinna
|
1
|
0,009
|
0,1
|
1
|
0,02
|
0,1
|
2
|
|
Н. punctata
|
1
|
0,009
|
0,1
|
3
|
0,08
|
0,3
|
4
|
|
H. parva
|
1
|
0,009
|
0,1
|
1
|
0,02
|
0,1
|
2
|
|
I. ricinus
|
12
|
0,1
|
1,8
|
49
|
1,4
|
5,3
|
61
|
|
Примечания:
А – число клещей, Б – индекс обилия, В – индекс доминирования, %.
|
Наглядно, количественную тенденцию по результатам сборов в обеих зонах можно проследить на рисунке 11 и 12.
Таким образом, согласно полученным нами данным D. marginatus наиболее широко распространенный и многочисленный вид в республике и это согласуется с данными предшествующих исследований [Котти и др., 2001; Батчаев и др., 2006]. Встречается повсеместно как в степной, так и в среднегорной зоне, достигая высоты 2275 м над ур. м. Отмечается слабо выраженное предпочтение степной зоны. При сборе на флаг встречался в биотопах с густой растительностью в непосредственной близости к водоемам. Индексы обилия этого вида на животных и на контрольных маршрутах значительно превышают показатели других видов. Доминирование его в среднем составляет 46,7–57,6% (таблицы 8–10). D. marginatus обнаружен во всех обследованных населенных пунктах. В большинстве районов D. marginatus доминирует.
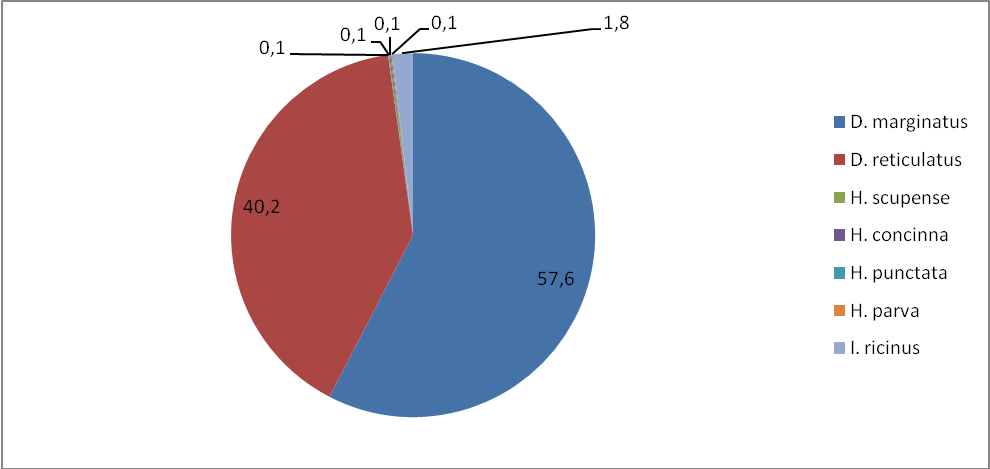
Рисунок 11 – Доля видов (в %) (индекс доминирования) по учетам на контрольных группах животных.
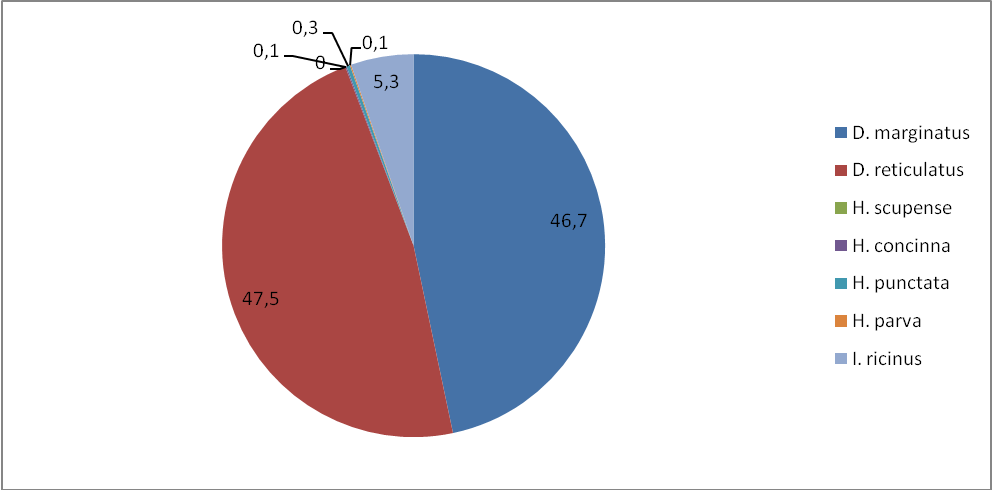
Рисунок 12 – Доля видов (в %) (индекс доминирования) на контрольных маршрутах.
D. reticulatus на территории республики встречается в степной и лесостепной зонах почти так же широко во всех 6 населенных пунктах как D. marginatus. Доминирование его в разных зонах при учетах разными методами очень сходно и колеблется в пределах 40,2–47,5%, хотя отмечается выраженная приуроченность к степной зоне (таблицы 4, 7–10) Индексы обилия на контрольных животных составляют 2,9 в степной зоне и 2,4 – в лесостепной. На контрольных маршрутах индекс обилия в степной зоне значительно превышает (17,4) индекс обилия в лесостепной зоне (7,2) (таблицы 8–10). Сезонная активность сходна с D. marginatus.
I. ricinus встречается в обеих зонах республики, однако, распространение его не столь широко, как у предыдущих видов. Он обнаружен во всех обследуемых нами населенных пунктах (таблицы 7, 8) Приуроченность к лесостепной зоне выражена слабее, чем к степной, что одинаково проявляется при обоих способах учетов. Индексы обилия на животных в степной зоне составляют 0,2 экз./животное, в лесостепной – 0,02. Доминирование – в степной зоне на животных 2,4 а в лесостепной зоне – 0,8 (таблицы 8–10). При учетах на флаг индекс обилия в степной зоне составляет 1,7, а в лесостепной зоне – 1,05. Доминирование в степной зоне составило 5, а в лесостепной – 5,7%. Активизация (при сборах с контрольных групп животных) I. ricinus начинается с середины марта в степной зоне, а в лесостепной нами собран всего один клещ. Активизация (при сборах на флаг) начинается с конца марта в степной зоне, а в лесостепной с середины апреля.
Н. scupense на территории республики относится к группе видов с низкой численностью. Обнаружен 1 клещ только в степной зоне (таблицы. 4, 7–10) и только при сборе с животных.
H. parva обнаружен только в зоне степей в конце апреля (таблицы 4, 7–10). Индексы обилия – 0,02 экз./животное и 0,05 экз./км, доминирования – 0,2%.
H. punctata обнаружен нами только в степной зоне при сборе на флаг и с контрольных групп животных, в конце апреля (таблицы 4, 7–10). Индексы обилия – 0,02–0,2 экз./животное, доминирования – 0,5–0,2%.
Н. concinna обнаружен нами только в степной зоне при сборе на флаг и с контрольных групп животных, в конце апреля (таблицы 4, 7–10). Индексы обилия - 0,05–0,02 экз./животное, доминирования – 0,2%.
Среди видов клещей, собранных в степной зоне преобладает D. marginatus - типичный представитель степных биотопов, в меньшей степени представлены клещи того же рода – D. reticulatus, обитатели более увлажненных мест среди кустарников. Клещи же I. ricinus – обитатели лесных массивов встречаются крайне редко из-за малого количества лесных участков предназначенных для выпаса скота. Крайне редки в этой зоне представители рода Haemaphysalis: parva, punctata и сoncinna. В этой зоне также редко встречается H. Scupense.
Клещи обнаружены во всех 6 исследуемых нами населенных пунктах. По численности фауну иксодид можно разделить на 3 группы. К первой группе очень высокой численности относится вид D. marginatus и D. retiсulatus. Индекс обилия D. marginatus на контрольных животных составляет в среднем 3,8 экз./животное, на контрольных маршрутах – 12,1 экз./км. Индекс обилия D. retiсulatus на контрольных животных составляет в среднем 2,7 экз./животное, на контрольных маршрутах – 12,3 экз./км. Высокая численность (вторая группа) характерна для I. ricinus. Индекс обилия для I. ricinus на контрольных животных составляет в среднем 0,1 экз./животное, на контрольных маршрутах – 1,4 экз./км. К третьей группе низкой численности относится большая часть иксодид – H. scupense, H. concinna, Н. punctata, H. parva, (0,009 экз./животное, 0,02-0,08 экз./км. Следует отметить, что при сборах двумя разными методами – с КРС и на флаг – видовой состав и порядок доминирования видов во многом совпадают, что свидетельствует о сходстве результативности этих методов. Не улавливаются на флаг H. scupense, очевидно, благодаря специфике биологии (однохозяинные виды). Степная и лесостепная зоны по видовому составу клещей несколько различаются. Из 7 выявленных нами видов 3 встречаются в обеих зонах (D. retiсulatus, D. marginatus, I. ricinus), а 4 вида (H. scupense, H. concinna, Н. punctata, H. parva) найдены только в степной зоне. Численность видов в обеих зонах также различна. Превышение индексов обилия и доминирования H. marginatum и D. retiсulatus на животных и на маршрутах в степной зоне в 1,2–2,4 раза выше, чем в лесостепной. У клеща I. ricinus отмечено выраженное предпочтение степной зоны (индексы обилия при учетах на контрольных животных в 10 раз выше, чем в лесостепной зоне и при учетах на флаг индекс обилия выше в 1,6 раза).
ЗАКЛЮЧЕНИЕ
По результатам работы сделаны следующие выводы:
- В весенний период в степной и лесостепной зоне КЧР обнаружены виды клещей из 4 родов. Клещи обнаружены во всех 6 (п. Кавказский, п. Новый, п. Мичуринский, ст. Зеленчукска, с. Кардоник, п. Даусуз) обследованных населенных пунктах. В районе исследования отмечено 7 видов иксодид (D. marginatus, D. reticulatus, I. ricinus, H. scupense, Н. concinna, H. punctata, H. parva), что составляет 27% от обнаруженных видов на территории Северного Кавказа и прилегающих районов.
- Доминирующими видами иксодид как в степной, так и в лесостепной зоне являются виды D. marginatus, D. reticulatus и I. ricinus. Индекс доминирования для D. marginatus составил для степной и лесостепной зоны 62,5% и 48,4%, соответственно, при сборе с контрольных групп животных, и 42,1% и 55,1 для степной и лесостепной зоны, соответственно, при сборе на флаг; для D. reticulatus эти показатели 34,3% и 50,8% для степной и лесостепной зоны, соответственно, при сборе с контрольных групп животных, и 52% и 39,2% для степной и лесостепной зоны, соответственно, при сборе на флаг; для I. ricinus – 2,4% и 0,8% для степной и лесостепной зоны, соответственно, при сборе с контрольных групп животных, и 5% и 5,7% для степной и лесостепной зоны, соответственно, при сборе на флаг. Для остальных видов клещей – H. concinna, Н. punctata, H. parva, H. scupense – индексы доминирования, для степной зоны от 0 до 0,2% и 0 для лесостепной.
- Наибольшая интенсивность инвазии в степной и лесостепной зонах характерна для D. marginatus (32 экз./животное и 14 экз./животное, соответственно) и для D. reticulatus (18 экз./животное и 15 экз./животное, соответственно), при 100% пораженности исследуемых животных. Интенсивность инвазии клещами H. scupense, Н. concinna, H. punctata, H. parva в степной зоне составила 0,1 экз./животное, при 11% экстенсивности инвазии. А в лесостепной зоне эти виды отсутствуют.
Выражаю благодарность сотрудникам Санэпидемстанции г. Черкесска (Балатчиеву К.Х. – заведующему эпидемиологическим отделом, Плясуновой С.Б. – энтомологу и др.), доценту кафедры внутренних болезней медицинского института КЧР Новиковой В.П. за предоставленную возможность совместно обследовать территорию, осуществлять необходимые для работы сборы и помощь в определении клещей и оказание помощи в обработке статистических данных.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
- Асваров Б.М. Проблемы арбовирусных инфекций в республике Дагестан // Сб. науч. тр. Махачкала, 2002. С. 152–153.
- Атшабар Б.Б., Айкимбаев А.М., Матаков М.И. и др. Экологические аспекты существования эндемии Конго-Крымской геморрагической лихорадки на юге Республики Казахстан // Пробл. особо опасных инфекций. Саратов, 2002. Вып. 84. С. 48–51.
- Ашибокова Л.Р. Исторический опыт рационального использования кормовых ресурсов в отгонном скотоводстве. //Сборник научн. Трудов по материалам н.п.к. Молочное и мясное скотоводство: состояние и перспективы развития в Южном Федеральном округе. Пос. Нижний Архыз 23-25 мая 2007 г. С.142-144.
- Балашов Ю.С. Иксодовые клещи – переносчики инфекций. СПб., 1998. 290 с.
- Боженко В.П., Шевченко С.Ф. К экологии клеща Ixodes laguri laguri в связи с его значением в поддержании некоторых природных очагов туляремии // Зоол. журн. 1956. Т. 35, № 6. С. 837–824.
- Вышемирский О.И., Петров В.А., Бутенко А.М. и др. Выделение вируса Крымской-Конго геморрагической лихорадки от больного человека в Волгоградской области летом 2000 г. // Вопр. вирусол. 2001. № 4. С. 21–22.
- Гальцева Г.В., Юничева Ю.В., Малай и др. Некоторые аспекты клещевого боррелиоза в Краснодарском крае // Матер. науч.-практ. конф. «Клещевые боррелиозы». Ижевск, 2002. С. 102.
- Гребенюк Р.В. Иксодовые клещи (Parasitiformes, Ixodidae) Киргизии (эколого-биологическая характеристика, вредоносное значение и основы борьбы с ними). Фрунзе, 1966. 328 с.
- Григорьев М.П., Евченко Ю.М., Шапошникова Л.И. и др. Изменение факторов природной очаговости Крымской геморрагической лихорадки в результате паводка // Мед. паразитол. и паразитарные болезни. 2005. № 4. С. 113–115.
- Денисов А.А., Савченко С.Т., Лазаренко В.В. и др. Носители и переносчики КГЛ в Волгоградской области // ЖМЭИ. 2005, № 4. С. 119–121.
- Доброхотов Б.П., Мещерякова И.С. Выявление энзоотичных территорий и изучение эпизоотий туляремии в различных типах природных очагов этой инфекции путем серологического исследования погадок птиц и помета хищных млекопитающих // ЖМЭИ. 1980. Т.1., № 24. С. 97–103.
- Евстафьев И.Л. Клещевой энцефалит в Крыму (Итоги двадцатилетнего изучения), Крымская противоч. станц. Симферополь, 2001. № 2. С. 53–57.
- Емельянова И.Н. Иксодовые клещи рода Hyalomma Koch, 1844 (Acari Ixodidae) Центрального Предкавказья и сопредельных территорий. Автореф. канд. биол. наук. Ставрополь, 2006. 16 с.
- Емчук Е.М. Иксодовые клещи. Киев, 1960. 162 с.
- Канонников А.М. Природа Кубани и Причерноморья. Краснодар: Краснодарское кн. изд-во, 1977. 112 с.
- Климова Л.И., Казакова Т.И., Газиев С.Г. и др. Изучение ареала и численности Hyalomma marginatum в республике Дагестан // ЖМЭИ. 2005. № 4. С. 123–124.
- Колонин Г.В. Распространение иксодовых клещей. М, 1984. 94 с.
- Коренберг Э.И., Горелова Н.Б., Постик Д., Котти Б.К. Новые для России виды боррелий – возможные возбудители иксодовых клещевых боррелиозов // ЖМЭИ, 1999. № 2. С. 3–5.
- Костюковский В.М., Пиликова О.М., Малай В.И., Юничева Ю.В. Изучение природной очаговости Крымской геморрагической лихорадки на территории Краснодарского края // ЖМЭИ. 2005. № 4. С.48–51.
- Котти Б.К., Гудиев О.Ю. Иксодовые клещи Ставрополя и его окрестностей // Мед. паразитол. и паразитарные болезни. 1997. № 4. 36–37.
- Котти Б.К., Новикова В.П., Емельянова И.Н. Фауна иксодовых клещей в Карачаево-Черкесской республике // Матер. 1 междунар. конф. посв. 110-летию со дня открытия проф. Виноградовым К.Н. сибирской двуустки у человека. Томск, 2001. С. 143–144.
- Кулиева Н.М., Соколова ЭИ. Фауна семейства (Ixodoidea) в природных областях Азербайджана // Тез. докл. на 3 Всесоюз. совещ. по теоретич. и прикл. акарологии (4–6 октября 1976 г.). Ташкент, 1976. С. 149–150.
- Львов Д.К. Значение вновь возвращающихся инфекций в биобезопасности // Тез. Докл. на 3 Всесоюзном совещании по теоретической и прикладной акарологии 4–6 октября 2002. Ташкент, 2002. № 5. С. 4–7.
- Манукян Д.В., Оганесян А.С., Шахназарян С.А., Алексанян Ю.Т. Видовой состав комаров и клещей в Армении // Мед. паразитол. и паразит. болезни. 2006. № 1. С. 31–33.
- Методические указания 3.1.1027-01. Сбор, учет и подготовка к лабораторному исследованию кровососущих членистоногих – переносчиков возбудителей природно-очаговых инфекций. М., 2002. 55с.
- Методические рекомендации. Профилактика инфекционных болезней. Организация и проведение мероприятий против Крымской геморрагической лихорадки на территории природных очагов России / Под ред. Г.Г. Онищенко. М., 2003. 30 с.
- Новикова В.П. Иксодовые клещи Карачаево-Черкесской республики и их эпидемиологическое значение. автореф. дисс канд. биол. наук. Черкесск, 2008. С. 19.
- Омариева Э.Я., Чернышев С.Н., Исаев А.Н., Омаров А.Ш. Эпидемиологическая обстановка по Крымской геморрагической лихорадке в республике Дагестан // ЖМЭИ. 2005. № 4. С. 59–61.
- Онищенко Г.Г., Марков В.И., Меркулов В.А. и др. Выделение и идентификация вируса Крымской-Конго гемморагической лихорадки в Ставропольском крае // ЖМЭИ. 2001. № 6. С. 7–11.
- Онищенко Г.Г., Ефременко В.И., Бейер А.П. и др. Обстановка по Крымской геморрагической лихорадке в Южном федеральном округе // ЖМЭИ. 2005. № 4. С. 5–7.
- Подсвиров А.В., Дмитриенко В.В., Подсвирова В.В., Санджиев В.Б-Х. Эпидемиологическая характеристика заболеваемости Крымской геморрагической лихорадкой на территории Республики Калмыкии // ЖМЭИ. 2005. С. 64–67.
- Полтараус Б.В. Климат Домбая. М. Изд. Моск. университета. 1972.-110 с.
- Померанцев БИ. Иксодовые клещи (Ixodoidea) // Фауна СССР. Паукообразные. Вып. 2. М., 1950. 223 с.
- Теплова Е.И., Прохорова В.Г., Гудиев О.Ю. Луговой клещ на Ставрополье // Фауна Ставрополья. Вып. 7. Ставрополь, 1997. С. 56–57.
- Теплова Е.И., Прохоров В.Г., Гудиев О.Ю. Фауна Ставрополья. Ставрополь, 1997. Вып. 7. С. 56–57.
- Тифлова Л.А., Резник П.А., Попова Е.В. Переносчики особо опасных инфекций и борьба с ними. Ставрополь, 1970. С. 459–471.
- Тохов Ю.М. Иксодовые клещи Ставропольского края (распространение, особенности паразитизма, меры борьбы). автореф. дисс канд. сельск. наук. Ставрополь, 2004. С. 18.
- Тохов Ю.М., Сысолятина Г.В., Чумакова И.В. и др. Особенности паразитарной системы Крымской геморрагической лихорадки на территории Ставропольского края в эпидсезон 2000 г. // ЖМЭИ. 2001. № 6. С. 98–99.
- Успенская И.Г. Иксодовые клещи Днестровско-Прутского междуречья. Кишинев, 1987. С. 143.
- Филлипова Н.А. Таксономический состав клещей семейства (Acarina, Parasitiformes) в фауне СССР и перспективы его изучения // Паразитол. сб. ЗИН АН СССР. 1984. Т. 32. С. 61–68.
- Филлипова Н.А. Фауна России и сопредельных стран. Паукообразные. Спб., 1997. 436 с.
- Филлипова Н.А., Панова И.В. Ревизия рода Dermacentor Koch фауны СССР и сопредельных территорий (Ixodoidea, Iхodidae) // Паразитол. сб. ЗИН АН СССР. 1989. Т. 35. С. 49–95.
- Эколого-географический атлас Карачаево-Черкесской республики / Под ред. Ю.П. Хрусталевой. Ростов-на Дону, 2001. 89 с.
- Юничева Ю.В., Оробей В.Г., Гальцева Г.В. Лайм-боррелиоз – новая эпидемиологическая проблема на Юге России // Сочинское ПЧО ГУ ППЧС ЦГСН. Сочи, 2004. С. 97–101.
- Hoogstraal H., Kaiser M.N. Observations on ticks (Ixodoidea) оf Libya // Ann. Entomol. Soc. Amer. 1960. Vol. 53, N 4. P. 445–457.
- Ouhelli H. Comparative develoment of Hyalomma marginatum, H. detritum, H. anatolicum excavatum, H. lusitanicum and H. dromedarii under laboratory conditions // Асta Parasitol. 1994. Vol. 39, N 3. P. 153–157.
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
«КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
(ФГБОУ ВПО «КубГУ»)
Кафедра зоологии
ДОПУСТИТЬ К ЗАЩИТЕ В ГАК
Заведующий кафедрой — д-р биол.
наук, проф._________ Т.Ю. Пескова
«____»_________________ _2014г
ДИПЛОМНАЯ РАБОТА
ФАУНИСТИЧЕСКИЙ КОМПЛЕКС IXODIDAE
КАРАЧАЕВО-ЧЕРКЕССКОЙ РЕСПУБЛИКИ
Работу выполнила ___________________________________Е.Л. Стрельцова
(подпись, дата)
Факультет биологический
Специальность 050102 Биология
Научный руководитель, канд. биол. наук, доцент._________ С.В. Островских
(подпись, дата)
Нормоконтролер, канд. биол. наук, доцент____________________В.В. Гладун
(подпись, дата)
Краснодар 2014
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
«КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
(ФГБОУ ВПО «КубГУ»)
Кафедра зоологии
ДОПУСТИТЬ К ЗАЩИТЕ В ГАК
Заведующий кафедрой — д-р биол.
наук, проф._________ Т.Ю. Пескова
«____»____________________2014г
ДИПЛОМНАЯ РАБОТА
ФАУНИСТИЧЕСКИЙ КОМПЛЕКС IXODIDAE
КАРАЧАЕВО-ЧЕРКЕССКОЙ РЕСПУБЛИКИ
Работу выполнила ______________________________________________Е.Л. Стрельцова
(подпись, дата)
Факультет биологический
Специальность 050102 Биология
Научный руководитель, канд. биол. наук, доцент.___________________ С.В. Островских
(подпись, дата)
Нормоконтролер, канд. биол. наук, доцент_______________________________В.В. Гладун
(подпись, дата)
Краснодар 2014
ФАУНИСТИЧЕСКИЙ КОМПЛЕКС IXODIDAE КАРАЧАЕВО-ЧЕРКЕССКОЙ РЕСПУБЛИКИ