Застосування інокулянту та дія інокуляції на огірки
ЗМІСТ
Вступ…………………………………………………………………………….… 3
Розділ 1. Віруси – збудники хвороб овочевих рослин………………………. 5
1.1 Номенклатура і класифікація фітопатогенних вірусів……………5
1.2 Патологічний вплив вірусних інфекцій на овочеві рослини……...9
1.3 Локалізації вірусів в рослині та основні хвороби овочевих
культур……………………………………………………………...16
Розділ 2. Методика дослідження……………………………………………...21
Розділ 3. Результати досліду та їх обговорення ……………………………32
3.1 Народно-господарче і харчове значення культури огірка……….32
3.2 Ботанічна і біологічна характеристика огірка…………..………..34
3.3 Захворювання огірка, які передаються насінням…………..…….38
3.4 Захворювання огірка, поширювані попелицями…………………44
3.5 Індикація вірусу та вплив інокуляції збудника на ріст, розвиток та
врожай огірка……………………………………………………....45
3.6 Боротьба із вірусними хворобами рослин………………..……….51
Розділ 4. Охорона праці………………………………………………..............54
Висновки….……………………………………………………………………..60
Список використанних джерел………………………………………………61
ВСТУП
Протягом останнього часу в фітовірусології, як і в інших молекулярно-біологічних дисциплінах, спостерігається значний інтерес до розвитку досліджень на популяційному та еколого-популяційному рівні, і ця тенденція є загальною для багатьох традиційних молекулярно-біологічних досліджень. Значні успіхи у фітовірусології на молекулярно-біологічному рівні, такі як розшифровка структури генів, білок-білкової та білок-НК взаємодії, розшифровка АК-послідовностей та тривимірної будови структурних та регуляторних вірусних білків та багато іншого контрастує з нашими знаннями відносно поведінки вірусів рослин в природних умовах біоценозів
[ 3]. Поява в останні десятиріччя нового покоління діагностичних методів для визначення наявності фітовірусів (таких як ІФА, RIA, RIPA, ПЛР, ЛЛР, та ін.) дали змогу на новому рівні вивчати розповсюдженість фітовірусів в навколишньому середовищі [1]. Актуальність даного питання полягає в його як фундаментальності, так і в практичному значенні. Визначення ареалу розповсюдженості, механізмів передачі, кола рослин-господарів, реакції на зміни оточуючого середовища дає можливість не тільки більш повно характеризувати того чи іншого представника царства Vira, але і дасть змогу прогнозувати появу та розвиток вірусних хвороб і правильно виробляти стратегію і тактику боротьби з ними — запровадження стійких сортів, цілеспрямована боротьба з резерватами та переносниками, отримання безвірусного посадкового матеріалу [7, 10].
Предмет дослідження - збудники вірусних хвороб овочевих рослин.
Об`єкт дослідження - застосування інокулянту та дія інокуляції на огірки.
Мета роботи - з`ясувати вплив вірусних інфекцій на рослинні організми на прикладі огірка.
Основні завдання:
1) охарактеризувати сучасний стан епідеміології вірусних захворювань рослин на Україні;
2) розглянути шляхи розповсюдження вірусів рослин в природі та роль екофакторів у цьому процесі;
3) дати стислу характеристику найбільш поширених в Україні вірусних захворювань овочевих рослин;
4) проаналізувати вплив вірусної мозаїки на огірок;
5) розглянути основні заходи боротьби з вірусами овочевих рослин.
Практичне значення одержанних результатів:
Визначення ареалу розповсюдженості, механізмів передачі, кола рослин-господарів, реакції на зміни оточуючого середовища дає можливість не тільки більш повно характеризувати того чи іншого представника царства Vira, але і дасть змогу прогнозувати появу та розвиток вірусних хвороб і правильно виробляти стратегію і тактику боротьби з ними — запровадження стійких сортів, цілеспрямована боротьба з резерватами та переносниками, отримання безвірусного посадкового матеріалу
РОЗДІЛ 1.
ВІРУСИ – ЗБУДНИКИ ХВОРОБ ОВОЧЕВИХ РОСЛИН
1.1 Номенклатура і класифікація фітопатогенних вірусів
Спроби систематизувати віруси були зроблені багатьма вченими. У різний час в літературі знаходили відображення різні системи та номенклатури вірусів, що послідовно змінюють один одного, або (частіше) вживаються одночасно ( паралельно) . Однією з перших була номенклатура, запропонована Д. Джонсоном в 1927 р. [ 7, 27]. Він запропонував іменувати віруси за назвою рослини - хазяїна , в якому вони вперше були виявлені, і присвоювати їм певний порядковий номер (у тій послідовності , в якій вірус був виявлений на даній культурі ) . Так , вірус тютюнової мозаїки ( ВТМ) , по Джонсону , отримав назву « тютюновий вірус 1 » , наступні виявлені на тютюні віруси отримували відповідно назва тютюнового вірусу 2 , 3 і т. д.
К. Сміт в 1937 р. дещо видозмінив систему і номенклатуру Джонсона. За К. Сміту , вірус тютюнової мозаїки отримав назву Nicotiana virus 1 , вірус огіркової мозаїки - Cucumis virus 1 і т. д. (номенклатура К. Сміта широко увійшла в спеціальну літературу , в багатьох фітопатологічних виданнях вона вживається і до теперішнього часу) [3,7.10].
Ф. Холмс (1939 , 1948) , а потім і інші автори запропонували використовувати для вірусів так звану бінарну номенклатуру , в якій враховуються як характер викликаних вірусом симптомів , так і тип природних переносників . Так , за Д. Холмсу , вірус тютюнової мозаїки отримав назву Магтог tabaci . Перша спроба створити систему фітопатогенних вірусів належить Ф. Холмсу (1939). Він виділив віруси в новий світ ( царство) Vira з двома підрозділами - віруси, що паразитують в рослинах (тип » Phytophagi ), і віруси , що паразитують в тваринах (тип Zoophagi ) . В 1948 р., Ф. Холмс кілька видозмінив цю систему , представивши віруси не як царство , а як порядок Virales . В. М. Жданов (1953) запропонував систематику , за якою тип віруси ( Vira) поділяється на три класи - фаги , віруси рослин і віруси людини і тварин. Принципово ця класифікація майже не відрізнялася від системи Ф. Холмса. Обидві вони будувалися за аналогією з систематикою вищих рослин [12,15].
За системою, запропонованою В. Л. Рижковим (1952 , 1954), тип віруси (названий їм Ivanovskya ) поділяється на п'ять класів , при цьому враховуються вже окремі властивості самих вірусів ( форма і розміри вірусних частинок , їх хімічний склад , тип переносу та ін . ) .
Відомі також і інші системи ( Ф. Боуден , 1943 , 1948; К. С. Сухов , 1956; А. Є. Проценко , 1966; А. Гіббс і Б. Харрісон , 1968 , 1981, і ін.) [ 45,50 52].
На Міжнародному мікробіологічному конгресі , який відбувся в 1966 р. в Москві , найбільш доцільною була визнана система і номенклатура , запропонована О. Львовим , Р. Хорном і П. Турньє (система ЛХТ ) . За цією системою віруси поділяють на підтипи , класи , порядки , сімейства і пологи не тільки за формою і будовою віріонів , а й на підставі їх властивостей
[ 7,10,11 ].
Дещо пізніше рядом вірусологів ( Гіббс , Харрісон та інші) був рекомендований принципово новий ( неієрархічному ) підхід до класифікації вірусів [24, 25 ].
Кожна криптограма складається з чотирьох пар символів , в яких літерами або цифрами зашифровані певні властивості вірусу.
За класифікацією , прийнятою в даний час, всі рослинні віруси розділені на 20 основних груп , в кожну з яких включені віруси, подібні за низкою властивостей (розмір , форма , тип будови і загальний склад віріона ; тип , число ланцюгів і кількість фрагментів нуклеїнової кислоти; антигенні властивості вірусу , ставлення його до температури ; коло рослин - господарів ; переважаючі шляхи і механізм переносу; симптоми ураження та інші - всього близько 50 ознак) [15, 19 ]. Назви цих груп дано за скороченими назвами типового представника , що входить до групи .Так , до групи неповірусов об'єднані віруси , переносниками яких є нематоди,а віріони багатогранні діаметром 30 нм ( nematode polyhedra virus) , група кукумовірусів названа по типовому представнику групи - вірусу огіркової мозаїки - Cucumber mosaic virus , група потівірусов - за назвою У- вірусу картоплі ( Potato Y-virus ) і т. д.
Віруси , що вражають рослини, не можуть розмножуватися поза живої клітини рослини - хазяїна або комахи - переносника . Багато вірусів зберігаються поза живої рослини [ 1,34 ]. До таких вірусів відносяться насамперед циркулятивні віруси , тобто потребують біологічної
( персистентної ) передачі. Місцем їх перезимівлі можуть бути живі зимуючі частини рослини - коренеплоди , цибулини , бульби і т. д. Таким шляхом зберігаються віруси вегетативно розмножуючих рослин - картоплі , буряків , цибулі , гладіолуса і т. д. Так , у бульбах картоплі перезимовує більшість вражаючих його вірусів , в коренеплодах буряка - віруси мозаїки і жовтяниці цукрового буряка. Зберігаючись з посадковим матеріалом і передаючись з року в рік потомству віруси призводять до різкого зниження продуктивності рослин , викликаючи так зване явище виродження сорту [34,45].
З рослинними залишками і в ґрунті можуть зберігатися тільки найстійкіші віруси - вірус мозаїки тютюну ( ВТМ) , огірковий вірус 2 і ін.
Значна частина мозаїчних вірусів , як вже зазначалося вище , може зберігатися і передаватися з насінням ( огірковий вірус 2 , вірус мозаїки сої , вірус мозаїки квасолі , кільцеві віруси кісточкових ) .
Спеціалізація вірусів може бути як вузькою , так і широкою. Так , Cucumis virus 2 (вірус зеленої мозаїки огірка ) вражає тільки огірок і небагато інших рослин із сімейства гарбузових , a Cucumis virus 1 , крім гарбузових , заражає рослини з інших ботанічних сімейств ( гречку посівну , шпинат , томат, фіалку рогату , молочай , Амарантус , айстру китайську , примулу та ін.) [ 23,45,49,52 ].
Віруси - найдрібніші ( субмікроскопічні ) збудники інфекційних хвороб людини , тварин і рослин , що не мають клітинної будови та здатні розмножуватися тільки в живих клітинах організму - господаря. Відомо близько 600 вірусів , здатних паразитувати на рослинах . Вірусні хвороби широко поширені на злакових і бобових культурах , багатьох видах плодових і ягідних рослин. Дуже великої шкоди завдають віруси рослинам з сімейства пасльонові - картоплі , томату , тютюну. Саме на тютюні при вивченні так званої мозаїчної хвороби , яка характеризується нерівномірним забарвленням листя , були в 1892 р. відкриті віруси.
Пріоритет цього відкриття належить російському вченому Д. І. Івановському , який встановив не лише надзвичайно малі розміри вірусів , але і такі важливі їх властивості , як заразність ( інфекційність ), здатність до розмноження (накопиченню) і утворення специфічних внутрішньоклітинних включень (тепер їх називають кристалами Іванівського).
Слідом за Івановським аналогічне дослідження мозаїки тютюну було проведено голландським мікробіологом М. Бейерінком (1898), який і дав назву новому виду збудника фільтрівний вірус від латинського virus - отрута Пізніше фільтрівні віруси стали називати просто вірусами. До 1906 було описано вже 29 вірусних хвороб рослин. Ці факти поклали початок розвитку нової галузі біології - вірусології [ 3,7 ].
У 1935 р. американський біохімік і вірусолог У. Стенлі виділив вірус тютюнової мозаїки в чистому , кристалічному вигляді , а в 1937 р. англійці Ф. Боуден і Н. Пірі встановили , що вірус тютюнової мозаїки складається з 95 % білка і 5% нуклеїнової кислоти [21,36] .
Детальне вивчення будови вірусних частинок - їх форми , розмірів - стало можливим лише після винаходу електронного мікроскопа.
У нашій країні серйозну увагу на вірусні хвороби рослин було звернено в двадцяті роки , коли А. А. Ячевським було виявлено та описано декілька вірусних хвороб на картоплі [ 34 ]. У тридцяті роки в СРСР була організована перша вірусологічна лабораторія . Велику роль у становленні вітчизняної фітовірусології зіграли радянські вчені: В.Л.Рижков, М.С.Дунін,
С.Н Московець , К.С.Сухов , Л.П.Петерсон , А.Є.Проценко , М.І.Гольдін ,
А.Л.Амброуз , І.Г.Атабеков , а також В.Г.Рейфман , Ю.А.Леонтьєва ,
Т.Д.Вердеревський , Ю.І.Власов і багато інших фахівців
1.2 Патологічний вплив вірусних інфекцій на овочеві рослини
Віруси, віроїди і фітоплазми викликають різні патологічні зміни в уражених рослинах , які зачіпають (особливо при системних інфекціях ) практично всі процеси , що регулюють їх ріст і розвиток [ 24,26,31 ].
Зовнішні симптоми захворювань. Візуально вплив вірусної інфекції на рослину проявляється у вигляді різних ознак місцевого та системного характеру - мозаїк , некрозів , кільцевих плямистостей , затримки або , навпаки , стимуляції ростових процесів . Симптоми не завжди специфічні. Їх прояв варіює залежно від виду збудника або рослини , дії екологічних факторів , супутніх інфекцій. Реакція на зараження може мати безсимптомний (латентний ) характер. При гострій формі захворювання можливий летальний результат. Деякі прояви вірусної інфекції схожі з ознаками , індукованими збудниками грибний або бактеріальної природи , токсинами членистоногих і нематод , а також фізіологічними порушеннями
[ 13, 14].
Виділяються такі типи зовнішніх ознак прояву вірусних захворювань: порушення росту і розвитку рослин , зміни форми і розмірів різних органів , зміни забарвлення листя, квіток , плодів , утворення некрозів .
Мозаїки - відносяться до найбільш поширених і характерних симптомів вірусних захворювань. Залежно від розташування і форми малюнка , його колірної гами відзначають цілий ряд їх різновидів (жовта , біла , жилкова , смугаста мозаїки , кільцева плямистість , ряболиста , сітчастість , лінійний візерунок і т.д.). Для всіх типів мозаїк характерна зміна забарвлення листя , пелюсток квітки або плодів , насіння , пов'язані з порушеннями пігментації.
Хлороз . Ця ознака , подібно мозаїці , також представлена різними типами прояви - від пожовтіння , наприклад , тільки верхніх кінчиків листя
( верхівковий ) , або їх країв (крайовий) , тканин між жилками
( міжжилковий ) або всієї листової пластинки (загальний хлороз ) .
Некрози . Можуть проявлятися як самостійна ознака, що свідчить про поразки вірусами , або представляти собою заключну фазу патологічного процесу після появи симптомів мозаїки або хлорозу . Некрози можуть мати чорний , бурий , червоний або білястий колір , набувати округлий , неправильний або незграбний вигляд , форму штрихів , смуг , рисок або концентричних плям [ 39, 42,51 ].
В'янення . Ознаки виявляються , як правило , у випадках ураження судинної або кореневої систем , а також як результат вторинної дії стресових факторів (знижені або підвищені температури середовища , передозування мінеральних елементів живлення , супутні інфекції або шкідники ) .
Деформація . Симптоми обумовлені різними фізіолого- біохімічними порушеннями , що настали внаслідок інфікування рослин. Найбільш типовими є деформації пагонів , листя, квіток , плодів , прояви морщинистости , кучерявості , ниткоподібні , скручування листових пластинок . Своєрідний симптом , деформуючий орган , представляють новоутворення на коренях , стеблах і листках із зворотного боку ( енаціі ) . Деякі віруси викликають розростання або збільшення окремих органів
( верхівкової бруньки , жилок , чашолистків , листя) [11,31,39].
Затримка росту . При ослабленні або повне припинення росту рослин виникають ознаки карликовості . Інгібування росту і розвитку окремих органів призводить , наприклад, до дрібноплідних , безсім’янності .
Стовбуріння . Характерна ознака при фітоплазменних інфекціях. Зустрічається і як результат вірусних поразок. При цьому часто спостерігається проліферація вегетативних та генеративних органів. У хворого рослини утворюється велика кількість бічних пагонів ( " відьомський мітли " ) . З одного вузла може формуватися кілька стебел. Частини квітки перетворюються на листкоподібні утворення , а їх загальне число збільшується ( філлодіямі ) [ 39, 42 ].
Типовими симптомами при фітоплазменних захворюваннях , крім зазначеного раніше стовбуріння , є затримка росту і прояв карликовості , внаслідок інгібування ростових процесів . Поширені різні види деформацій на листі ( наприклад курчавість ) , пагонах , плодах і квітках . Пагони при ураженнях фітоплазми стають крихкими , число їх може зростати , міжвузля коротшають . Досить поширені такі характерні , специфічні ознаки, як позеленіння і проліферація квіток , а також тотальний хлороз рослин.
Візуальний прояв ознак при вірусних захворюваннях дуже різноманітний. Він варіює в динаміці розвитку інфекційного процесу. Так , звичайно віруси найбільш чітко і різко проявляються на молодих листях у період активного росту рослин. На більш старих листях вони менш помітні або повністю пропадають. За оптимальних умов живлення і, навпаки, при дефіциті тих або інших елементів може спостерігатися як пряма , так і зворотна кореляція зі ступенем прояву симптомів , тип та інтенсивність яких великою мірою визначаються характером метаболізму рослини-хазяїна і рівнем репродукції вірусу. Вплив температури і освітленості також залежить від виду господаря і вражаючого його збудника. Наприклад , на багаторічних рослинах , зокрема , плодових і ягідних , при підвищених температурах (25-30С) може спостерігатися маскування зовнішніх ознак захворювання. Саме тому в регіонах з теплим кліматом часто можна навіть спостерігати ефект масового тимчасового " одужання" . Разом з тим різке підвищення температури , як стресовий фактор , може призвести до летального результату. Оптимальною для прояву ознак вірусного ураження вважається діапазон температур в межах 16-25.
Рівень освітленості здатний також змінювати прояв морфологічних ознак хвороби , посилюючи або послаблюючи їх. Так , ВТМ тільки в умовах слабкої освітленості викликає на томаті специфічні ознаки [ 17, 39 ]. Типові ознаки захворювань утворюються при освітленні оптимальному для життєдіяльності самої рослини -хазяїна. Однак при штучному зараженні з метою підвищення ефективності інфікування рослини на першому етапі попередньо слід навіть притіняти. Ознаки хвороби можуть видозмінюватися і при комплексній поразці [19,21].
Фізіологічні зміни. Віруси, будучи облігатними внутрішньоклітинними паразитами рослин, позбавлені власної функції обміну речовин. При репродукції вони використовують ферменти , субстрат і енергію інфікованої клітини , прямо або побічно впливаючи на більшість фізіологічних процесів рослини-хазяїна (обмін речовин , активність ферментів , дихання , проникність мембран , зміст і реалізація функцій регуляторів росту , фотосинтез ) . Наприклад , віруси , використовуючи білоксинтезуючу систему господаря , здатні змінювати процеси формування клітинних органел (зокрема , рибосом і полісом ) , беручи участь у перерозподілі кількісних співвідношень між ними , а також впливаючи на їх стан [3,7,19].
При вірусній інфекції відбуваються такі зміни метаболізму у рослин , які зазвичай спостерігаються при їх старінні . Впливи вірусів можуть відрізнятися за специфічності та тривалості залежно від виду збудника , рослини і фази його розвитку , а також стадії захворювання.
Вплив вірусів на активність дихання вражаючого рослини визначається особливостями їх репродукції і захисними властивостями господаря. Як правило , максимальний рівень репродукції збігається з підвищенням інтенсивності дихання . Вже через кілька годин після зараження рослин спостерігається його активізація , яка збільшується при появі видимих патологічних ознак . У міру стабілізації концентрації вірусних частинок відбувається зниження рівня дихання. При ряді хронічних , повільно протікаючих, латентних інфекціях можна спостерігати навіть зниження рівня дихання в порівнянні з нормою. Найбільша стимуляція дихання відзначається при розвитку місцевої некротичної реакції після інфікування
[ 38, 42 ].
Вірусна інфекція призводить і до порушень ферментативних систем рослини -хазяїна. Збудники здатні викликати підвищення активності окисних ферментів ( оксидаза , пероксидаза , поліфенолоксидаза ) , рибонуклеази , придушення каталази. Їх активність особливо виявлялася безпосередньо в знебарвленого ділянках . В уражених тканинах значно підвищується і рівень вмісту дегідрази .
При ураженнях вірусами змінюється гормональний баланс у рослин . Такі симптоми вірусних захворювань , як пригнічення росту , карликовість , стовбуріння, пов'язані зі зміною змісту ростових речовин ( ауксини , цитокініни , гібереліни ) . При мозаїці і хлорозі патологічні зміни, що зачіпають хлоропласти , призводять до руйнування хлорофілу , зміст якого може знижуватися на 5-50% , в хлоротичних ділянках листової пластинки навіть у 20 разів. У результаті падає фотосинтетична активність [14,45].
До впливу вірусів схильні також синтез і транспорт вуглеводів. Спостерігаються кількісні та якісні зміни у складі вуглеводів в різних тканинах і органах. Відзначаються порушення азотного обміну , що призводять до руйнування білків господаря і придушенню їх нормального синтезу . З початком репродукції вірусу падає вміст небілкового азоту. Постійних відмінностей у змісті окремих амінокислот не спостерігалося. Найбільш виражено збільшення концентрації аспарагіну і глютаміну . Зростає також вміст фосфору нуклеїнових кислот , змінюється концентрація аскорбінової кислоти , деяких пігментів і мінеральних речовин [42, 51]. При вірусних ураженнях спостерігаються і значні порушення водного режиму. Залежно від характеру інфекції та фази захворювання швидкість транспірації і вміст води в тканинах знижується або підвищується. Таким чином , віруси викликають первинні порушення метаболізму , пов'язані з їх реплікацією , і вторинні зміни в організмі господаря , подібні з прискореними процесами його старіння [ 14 ].
Анатомічні та цитологічні зміни . До основних типів анатомічних змін можна віднести некротизацію (відмирання) тканин , гіперплазію (розростання тканин при надлишковому діленні клітин) , гіпоплазію (придушення диференціації і росту клітин ) . Наприклад , пригнічення функції диференціації клітин паренхіми спостерігається при мозаїчних захворюваннях в хлоротичних ділянках . Вони набувають форму , схожу з клітинами губчастої паренхіми , і товщина листової пластинки зменшується. Такий збудник , як вірус кучерявості верхівки цукрового буряку викликає гіперплазію клітин флоеми і ксилеми , утворення надмірної кількості ситовидних трубок. Редукція органів і клітин спостерігається при інфікуванні рослин вірусом закуклювання вівса. Цілий ряд вірусів здатний стимулювати формування пухлин і різних новоутворень ( опуклості , бородавки , вирости, енаціі , галли ) .
Відомі збудники , які надають патологічний вплив на судинну систему. Так , при ураженні вірусом коротковузля винограду в судинах ксилеми з'являються бар'єри ; при інфікуванні вірусом жовтої карликовості ячменю спостерігається накопичення каллоза в ситовидних трубках , відмирання і руйнування клітин флоеми .
При деяких вірусних захворюваннях спостерігається варіювання числа, форми , розміру і структури ядер. Наприклад , при ураженнях махорки вірусом бронзовості томатів у клітинах паренхіми число ядер збільшується в 3-5 разів у порівнянні з нормою [ 26, 34 ].
Однак найбільш характерним цитопатичним ефектом при інфікуванні є утворення включень , що виявляються в різних частинах рослин , гемолімфі і епітеліальних клітинах переносників у світловому або електронному мікроскопах.
В даний час відомо більше 100 вірусних захворювань , при яких утворюються різні види включень. Для кожного вірусу характерний певний їх тип ( табл. 1.2 ) . Як правило , зустрічаються включення у двох формах - кристали і аморфні освіти (Х- тіла).
Вони поділяються на 4 умовних типи : включення , видимі в світловому мікроскопі ( кристалічні утворення , що складаються з впорядковано розташованих вірусних частинок; аморфні утворення, що містять вірусні частинки і деякі компоненти клітини , утворення, що складаються з видозмінених клітинних органел ) , і структури, які виявляються тільки в електронному мікроскопі ( НК , структурний білок вірусу і інші білки). Включення зазвичай локалізуються в цитоплазмі , проте їх можна виявити і в ядрах різних органів ( коренях , листі , стеблах і квітках ) і тканинах ( клітинах епідермісу , мезофілу , паренхіми , флоеми ) . Вони варіюють за розмірами. Наприклад , при зараженні Х- вірусом картоплі в рослинах тютюну включення по величині не перевищують розміри клітинного ядра , тоді як у самому картоплі вони займають близько половини всієї клітини.
Таблиця 1.2
Типи включень при деяких вірусних захворюваннях
|
Збудник
|
Тип включень
|
Локалізація
|
|
1
|
2
|
3
|
|
Вірус мозаїки цибулі
|
голковидні кристали
|
Клітини епідермісу і паренхіми листя
|
|
Вірус тютюнової мозаїки
|
Гексагональні призми
Х-тіла
|
Клітини епідермісу і паренхіми
|
|
Вірус кільцевої плямистості тютюну
|
Гексагональні призми Х-тіла
|
Клітини епідермісу листя
|
|
Вірус гравіювання тютюну
|
Зернисті Х-тіла
8-гранні біпірамідні пластинчаті кристали
|
Цитоплазма і ядра клітин епідермісу
|
|
Вірус мозаїки буряків
|
Веретеноподібні паракристали
|
Клітини флоеми
|
|
Вірус мозаїки капусти
|
Амебоподібні Х-тіла
|
Клітини епідермісу листя
|
|
Вірус мозаїки салату
|
Гранулярні Х-тіла без вакуолей
|
Клітини епідермісу листя
|
|
Вірус аспермії томата
|
Склоподібні Х-тіла
|
Клітини епідермісу листя
|
Продовження табл..1.2
|
Вірус бронзовості томата
|
Вакуолярні Х-тіла
|
Епідерміс листя і пелюстків
|
|
У-вирус картофеля
|
Округлі Х-тіла, циліндричні і округлі паракристали
|
Різні тканини
|
|
Вірус Х-картоплі
|
Вакуолярні зернисті Х-тіла
|
Клітини флоеми
|
Один і той же вірус навіть в одній клітині може формувати включення різних типів. Наприклад , вірус мозаїки томатів ( томатний штам ВТМ) утворює гексагональні кристалічні пластинки , веретеноподібні паракристали або плоскі структури, які з паличок , тоді як при ураженнях вірусом гравіювання тютюну в цитоплазмі спостерігаються зернисті включення , а в ядрі тих же клітин - кристали. Форма включень залежить і від виду инфицируемого рослини. Так , вірус закуклювання вівса утворює в клітинах вівса голчасті кристали , а в тканинах пирію повзучого - веретеновідние . При цілому ряді захворювань зустрічаються включення у вигляді аморфних Х- тел. Вони можуть мати вакуолі. За допомогою електронного мікроскопа в їх структурі , зокрема при ураженнях ВТМ виявляються елементи ендоплазматичний мереж , рибосоми , вірусні частинки і широкі нитки. Розміри і число Х- тіл значно варіює залежно від штаму збудника і виду клітини [ 24,31].
1.3 Локалізації вірусів в рослині та основні хвороби овочевих культур
Поширення збудників по рослині . Найбільш повно механізми переміщення досліджені у вірусів , вони просуваються в рослинах по проводним тканинам і від клітини до клітини в паренхімних тканинах по плазмодесмам . При цьому їх поширення може здійснюватися як зрілими віріонами , так і за допомогою ПК. Ще в 1959 р. було показано , що розміри плазмодесм достатні для проходження по них зрілих вірусних частинок. Однак прямі докази вдалося отримати лише через 8 років за допомогою електронної мікроскопії. Зокрема , в плазмодесмах клітин паренхіми Beta vulgaris був виявлений вірус жовтяниці цукрового буряка. Згодом у плазмодесмах різних рослин виявлені й інші віруси. Швидкість поширення їх таким шляхом значно варіює залежно від типу клітин , розміру плазмодесм, виду збудника та напрямки його переміщення. Так , показано , що ВТМ переміщається з верхнього епідермісу в нижній зі швидкістю 8 мкм / год. Загалом на підставі даних багатьох досліджень був зроблений висновок про те , що відстань в один шар клітин вірус долає протягом 4-7 ч. [ 47 ].
Більшість вірусів викликають системне ураження у сприйнятливих рослин. Це передбачає досить швидке поширення інфекції по різних тканин і органів . Пересування вірусів на великі відстані відбувається по провідній системі - флоемі або ксилемним тканинам. Класичні експерименти , результати яких свідчать про можливість транспорту вірусів по судинній системі , проведені Самуелем ще на початку 40 -х р. Так , було показано , що жовтий штам ВТМ після локальної репродукції поблизу точки інокуляції переміщається по середній жилці листа і викликає системну інфекцію. Тест на інфекційність виявив напрямок руху ВТМ в рослинах томата від верхівкового листа спочатку до коріння і потім до молодих листків . Більш старі листки інфікувалися пізніше . При цьому швидкість розповсюдження вірусу перебувала в прямій залежності від віку рослини [21, 37 ].
Дані про можливість пересування вірусів по флоемі були отримані при експериментах по штучному блокування її шляхом окільцьовування стебел. У цих випадках відзначалося уповільнення або повне припинення переміщення збудників. Разом з тим частинки різних вірусів виявлені на зрізах ситовидних трубок і ексудатах з флоеми .
Швидкість пересування вірусів по флоемі не перевищує для ВТМ 1,5 см / год, тоді як у вірусу кучерявості верхівки цукрового буряка вона досягає 150 см / ч. Таким чином , поширення по ситовидним елементам флоеми відбувається в сотні разів швидше , ніж від клітини до клітини . Воно здійснюється з током поживних речовин в напрямку руху протоплазми судинної системи.
Пересування ж по ксилемі було вперше продемонстровано на прикладі вірусу південній мозаїки квасолі після інокуляції рослин методом щеплення. Відзначено , що збудник поширювався як вгору , так і вниз по ксилемі [15,26].
Наявність різних транспортних форм у вірусів і здатність їх до пересування забезпечує можливість системного ураження рослин.
Зміст збудників в різних тканинах. Концентрація різних патогенів в органах і тканинах рослин та їх переносників значно варіює. Наприклад , відомо , що в листках , бруньках і квітках віруси накопичуються в значно більшій концентрації , ніж в інших їх частинах , а в насінні більшості видів можуть бути відсутні зовсім. Вона залежить від особливостей репродукції збудника в конкретному хазяїні . На накопичення того чи іншого вірусу , безсумнівно , впливають вік рослини , спосіб його інфікування і складаються екологічні умови для розвитку. Наприклад , вміст вірусу жовтої карликовості ячменю становить від 25 до 100 мкг / л клітинного соку , тоді як ВТМ накопичується в листі тютюну до концентрації більше 2 г / л. У той же час в рослинах томата його кількість падає в 5-10 разів.
У коріння більшість вірусів міститься у меншій (порівняно з іншими органами рослин) концентрації [10,26]. Найбільше накопичення віріонів при системної інфекції спостерігається в молодих листках. Їх концентрація представляє змінну величину залежно від часу формування листя після зараження. Навіть на окремих пластинках листя або пелюстках , що розрізняються по інтенсивності прояву симптомів музичних захворювань , спостерігається значне варіювання концентрації збудників. В цілому , наприклад , рівень вмісту вірусу жовтої мозаїки турнепсу в стеблах , коренях і черешках листя в 10-20 разів менший , ніж у самих листках.
Віруси приурочені до певних тканин . Так , вірус трістеци цитрусових в основному локалізований під флоемними тканинами. Як правило , в найбільш високій концентрації вони містяться в паренхімних і флоемних тканинах , і меншою мірою - в ксилемі . В апікальній меристемі збудники практично відсутні. Однак розмір вільної від вірусу зони змінюється при різних поєднаннях " вірус - рослина " і може становити величини порядку 0,1-1,0 мм. Причини , що визначають відсутність або низьку концентрацію вірусів в меристематичних тканинах , до кінця не з'ясовані. Виникнення таких безвірусних ділянок пояснюють різною швидкістю росту апексів і поширення вірусів , інтенсивністю поділу клітин і відсутністю диференційованої провідної системи . Це практично виключає можливість потрапляння інфекції в апікальну меристему , оскільки перші її фіксуються елементи розташовані у відриві від основного тяжа провідної тканини . В якості чинників, що перешкоджають проникненню вірусів у меристему , можуть виступати і малі розміри її плазмодесм або низька швидкість руху протоплазми.
За зоною апікальної меристеми концентрація вірусів може зростати в напрямку більш віково- зрілої тканини. Для ряду комбінацій " вірус -хазяїн " зона , аналогічна апексу за змістом віріонів , виявляється у кінчиків коренів заражених рослин.
Значно нижчі концентрації вірусів характерні і для калюсних культур. Наприклад , ВТМ виявлявся лише в 40 % тестованих клітин.
Таким чином , розподіл інфекції по рослині не є рівномірним . Особливо це характерно для багаторічних і, зокрема , деревних видів рослин.
На овочевих культурах зареєстровано значне число вірусних та фітоплазменних інфекцій як у відкритому , так і закритому грунті , які відрізняються великою шкідливістю (табл. 1.3 ) . Так , на томатах , особливо в південних районах , поширене таке захворювання , як стрік , що викликається комплексної інфекцією (ВТМ , Х- і У- вірус картоплі , ВОМ і Раттл - вірус в різних поєднаннях ) , масовий розвиток якого визначається наявністю вогнищ серед сміттєвої і дикоростучої рослинності і чисельністю переносників. Вірус мозаїки огірка ( ВОМ ) широко поширений у всьому світі , переважно в районах з помірним кліматом. Зареєстровано на багатьох овочевих , технічних , плодових , цитрусових і квіткових культурах , засмічених і інших рослинах .умовах [ 8,13,31 ]
Таблиця 1.3
Основні хвороби овочевих культур
|
Збудник
|
Симп-томи
|
Переносники
|
Хозяїн
|
|
Віруси
|
|
Вірус огіркової мозаїки
|
М, ДЛ, ЗР
|
Різні види тлі
|
Sysymbrium officinale, Medicago sp., Cirsium sp.
|
|
Вірус мозаїки цибулі
|
ПМ, ДЛ
|
Aceria tulipae
|
Цибуля
|
Вірус некрозу табака
|
ЗР, М, Х
|
Гриби Olpidium
|
Огірки, картопля, квасоля та ін..
|
Вірус бронзовості томатів
|
БП, БК, ЗР
|
Thrips tabaci
|
Близько 200 видів з більш як 40 сім-в
|
Вірус аспермії томатів
|
ПЖ, К, ДЛ, Н
|
Різні види тлі
|
Томати, огірки, види табака та ін.
|
Вірус мозаїки буряку
|
М, СЛ
|
Myzus persicae, Macrosiphum euphorbiae, Aphis fabae
|
Буряк, Chenopodium sp.
|
Вірус табачної мозаїки
|
М, ШЛ, НЛ, НП, ЗР
|
Не виявлений
|
Більше 500 видів
|
Вірус мозаїки цвітної капусти
|
М, ПЖ
|
Brevicorine brassicae
|
Види капусти, редиски , Barbarea sp.
|
|
Фитоплазми
|
|
Стовбура
|
Х, НЛ, АО, РЦ
|
H.obsoletus, H.mlokosieviczi, Aphrodes bicinctus, Euscelis plebejus, M.laevis
|
Томати, перець, і різні види бур’янів
|
ШЛ - штриховатість листя, НП - внутрішній некроз плодів,
БП - бура плямистість, БК - бурі кільця, К - карликовість,
ДЛ - деформація листків.
РОЗДІЛ 2. МЕТОДИКА ДОСЛІДЖЕННЯ
Методи діагностики вірусних хвороб рослин [1,19,29,31,38]. В основі діагностики вірусного захворювання лежать в першу чергу зовнішні симптоми. Але діагноз захворювання тільки за зовнішніми ознаками не завжди точний. Зовнішніми ознаками при визначенні вірусного захворювання можна обмежитися в тих випадках , коли вони дуже чіткі і характерні тільки для даного виду захворювання .
У практиці дуже часто доводиться стикатися з такими явищами , коли одні й ті ж або дуже схожі ознаки ураження викликаються абсолютно різними причинами (явище “ Фітопатологічної конвергенції “ ) . Приклад такої конвергенції - хлороз плодових дерев або ягідних культур.
Він може бути інфекційним , викликаним вірусами , і фізіологічним , пов'язаним з недостатнім надходженням в рослину азоту , заліза , сірки і т. д. [27,39]. Мозаїчне забарвлення листя на огірку може бути наслідком і вірусного ураження та дефіциту марганцю. Білу мозаїку на огірку часто плутають з дефіцитом магнію. Іноді подібними ознаками виявляються і різні вірусні хвороби. Так , через схожість симптомів під одною і тою же назвою « мозаїка пшениці » були описані три зовсім різні хвороби. Різними вірусами , що істотно розрізняються за особливостями поширення, збереження та іншим властивостям , викликаються звичайна і зелена краплиста мозаїки на огірку , мають також схожі ознаки прояву . При випадковому попаданні гербіцидів на рослини винограду листя їх приймають потворну форму , дуже нагадує прояв вірусного коротковузля [1,19] .
Тому при визначенні захворювання поряд із зовнішнім оглядом необхідні додаткові методи , уточнюючі ( або спростовуючі ) попередній діагноз .
Застосовують наступні методи діагностики вірусних захворювань і викликаючих їх вірусів :
1) встановлення інфекційності захворювання ,
2) серологічний метод ;
3) метод рослин - індикаторів ;
4 ) електронна мікроскопія ;
5 ) метод внутрішньоклітинних включень ;
6 ) метод люмінесцентного аналізу;
7 ) анатомічний метод ;
8 ) хімічний метод
Основними з них є перші чотири методи .Для всіх вірусів , які вдається передати соком на трав'янисті індикатори , дуже важливими показниками при діагностиці вірусів є визначення таких фізичних властивостей вірусу , як температура інактивації вірусу ( гранично висока температура , при якій вірус інактивується in vitro протягом 10 хв) ; кордону розведення (ступінь граничного розведення соку хворого рослини , при якій ще зберігається його інфекційність ) ; тривалість збереження інфекційності соку при 20 ° С.
Так , для огіркового вірусу 2 ( Cucumis virus 2 ) температура інактивації 90 ° С , граничне розведення соку 1 : 1 000 000 , тривалість збереження інфекційності соку при кімнатній температурі більше року , а для огіркового вірусу 1 ( Cucumis virus 1 ) температура інактивації 60 - 73 ° С , межа розведення соку 1: 1000 , тривалість збереження інфекційності соку 3-6 днів.
Встановлення інфекційності захворювання.
Всі вірусні захворювання заразні, інфекційні. Залежно від виду вірусу як причини захворювання і його властивостей (стійкості вірусу , способів його розповсюдження в природних умовах і т. д.) для встановлення інфекційності застосовують такі способи зараження рослин [ 14 ].
Інокуляція соком хворої рослини . Цей метод придатний для передачі на здорові рослини тільки тих вірусів , які можуть поширюватися контактно - механічним способом.
Для приготування соку використовують листя хворих рослин з чіткими симптомами захворювання. Дрібно нарізані листя розтирають у ступці з додаванням невеликої кількості води і абразиву ( мелене скло , карборунд , дрібний пісок і ін ) , потім з розтертої маси віджимають сік через подвійний шар марлі і втирають його в лист здорової рослини пальцем , ватним тампоном або шпателем , обгорненим тонким шаром вати. Заражені рослини поміщають під ізолятори і спостерігають за появою через певний час симптомів захворювання.
Передача вірусної інфекції щепленнями. Хворий прищеп прищеплюють на здоровий підщеп або навпаки.
Передача вірусу за допомогою комах-переносників . Використовують види комах , які є найбільш ймовірними переносниками вірусу , найчастіше персикову тлю ( Myzodes persicae ) . Комах (завідомо вільних від вірусу особин) витримують протягом певного часу на хворих рослинах , а потім переносять на здорові [38,41] .
Всю роботу проводять в умовах суворої ізоляції рослин від зараження ззовні.
Передача вірусної інфекції за допомогою берізки .
Пересадка рослин. У деяких випадках для підтвердження або спростування вірусної природи захворювання вдаються до пересадки рослини на нове місце. Так чинять , наприклад , при визначенні характеру хлорозу малини . Якщо причиною хлорозу були погані грунтові умови , то у рослин , пересаджених на свідомо сприятливий грунт , відновлюється через певний час ( іноді вже через 3-4 тижні) нормальний зелений вигляд. При вірусному хлорозі рослина від такої пересадки не видужає [12,15,21] .
Серологічний метод заснований на тому , що віруси , як і бактерії , гриби, а також білки тварин і рослин , будучи введені в кров теплокровної тварини ( кролика , барана , коня і т. д.) , здатні викликати накопичення в плазмі крові ( сироватці ) тварини специфічних видозмінених білків , що одержали назву антитіл ( речовини, що вводяться в організм тварини і викликають у ньому утворення антитіл , називають антигенами ) [ 38, 41 ].
Отримана з крові тварини сироватка і що містяться в ній антитіла є суворо специфічними , тобто мають здатність реагувати (давати позитивну реакцію) тільки з тим антигеном , який був введений в тіло тварини і по відношенню до якого отримана антивірусна сироватка.
Реакція між антигеном і специфічними антитілами , що містяться в сироватці , носить назву серологічної реакції (від латинського serum - сироватка) .
У практиці широко застосовують крапельний серологічний метод, розроблений М. С. Луніним і Н. Н. Попової (1937 р.). Сутність цього методу полягає в наступному.
На предметне скло , ближче до його країв , наносять дві краплі випробуваного соку. Сік вичавлюють з листя затискачем Пеана через подвійний шар марлі. В одну з цих крапель (зазвичай ліву) чистою піпеткою додають одну - дві краплі сироватки , специфічної по відношенню до того вірусу , на присутність якого в рослині хочуть провести аналіз , а в іншу (праву) така ж кількість так званої нормальної , або контрольної , сироватки , отриманої від тварини , не піддавався введенню яких би то не було антигенів ( неімунізованних , контрольних тварин). Краплі соку та сироватки перемішують (куточками чистого предметного скла) , спочатку краплю з нормальною (контрольної) сироваткою , а потім зі специфічною до вірусу . При перемішуванні треба намагатися при дати краплям округлу форму діаметром 1,5-2 см. Якщо в соку випробуваного рослини міститься вірус , відповідний антитілам , що містяться в сироватці , то через 1-3 хв у краплі утворюються видимі неозброєним оком пластівцевидні згустки , або осад - преципітат (позитивна реакція).
Якщо ж в аналізованому соку такого вірусу немає , крапля залишається такою ж , як і контрольна , тобто рівномірно каламутною.
Крапельним методом можна користуватися як в лабораторних , так і в польових умовах. Для його застосування необхідні предметні скельця , затискачі або пінцети ( для віджимання соку ) , шматочки марлі , піпетки та відповідні антизбудливі сироватки.
У науково -дослідних інститутах і лабораторіях користуються також постановкою реакції преципітації в агарових гелях . Метод зручний тим, що дозволяє фіксувати результати за допомогою фотозйомки. Однак він більш громіздкий і тривалий (утворення ПЛС преципітації спостерігається зазвичай лише через 24-62 год) і чітку реакцію дає 1 - реакція негативна лише по відношенню до сферичним (рідина рівномірно каламутна ) вірусів , які мають невеликі розміри ; паличкоподібні і ниткоподібні віруси слабко проникають в гель . Новою, більш досконалою та високочутливою модифікацією серологічної діагностики вірусів є розроблений в медицині (Scott , 1963) і широко використовуваний останнім часом у рослинництві імуноферментний метод , або ензімозв’язанний імуносорбентний тест ( ELISA - TЕCT ) . Суть методу полягає у використанні антитіл , пов'язаних з молекулами ферменту ( пероксидази або фосфатази ) . Результат серологічного аналізу визначається за кольоровою ферментативною реакцією [29,31].
Розроблений також метод віробактеріальної аглютинації ( АБВ- тест).
Метод рослин - індикаторів заснований на використанні рослин, що дають дуже чітку реакцію , суворо специфічну по відношенню до певного виду вірусу. Так , для вірусу тютюнової мозаїки хорошою індикаторною рослиною є Nicotiana glutinosa . На листках цієї рослини в місцях зараження вірусом тютюнової мозаїки вже через 2-3 дні утворюються виразні некротичні плями. Для Х - вірусу картоплі як индикаторну рослину використовують гомфрену ( Gomphrena globosa ) ; зараження проявляється також у вигляді місцевих некротичних плям. Для вірусу огіркової мозаїки 1
( Cucumis virus 1) в якості індикатора можна використовувати тютюн
( Nicotiana tabacum ) . Для більшості вірусів, що вражають садову суницю , в якості індикаторної рослини використовують дику лісову суницю ( Fragaria vesca ) . Для вірусів плодових часто використовують види марі
( Chenopodium ) , огірок ( Cucumis sativus ) , а також сіянці і певні сорти плодових .
Зараження здійснюють , натираючи листя рослини -індикатора соком хворої (досліджуваної) рослини . Натирати лист можна ватним тампоном , скляним шпателем або навіть пальцем , змоченим в соку випробуваного рослини. Для отримання соку лист розтирають у ступці з додаванням невеликої кількості води (2-4 мл). Рослини - індикатори вирощують в умовах суворої ізоляції. Для інокуляції ( зараження) використовують молоді , інтенсивно зростаючі рослини у фазі 3-4 листків . Метод рослин - індикаторів кілька громіздкий , але досить точний . Для економії місця , матеріалу , зручності спостережень , а також підвищення його точності ( зменшення небезпеки випадкового занесення інфекції) можна користуватися не цілими рослинами , а відокремленими листям , поміщеними в чашки Петрі [31,34].
Метод електронної мікроскопії . Вивчення форми і будови вірусних частинок стало можливим завдяки винаходу електронного мікроскопа.
Встановлено , що близько 70 % « зрілих » , або покояться , форм фітопатогенних вірусів мають паличкоподібну або ниткоподібну форму і близько 30 % - сферичну
Паличкоподібну форму мають вірус тютюнової мозаїки ( ВТМ) , вірус зеленої мозаїки огірка ( ВМО -2); ниткоподібну - більшість вірусів картоплі ( X , У, N , 5 ) , вірус « шарки » злив; сферичну ( ізометричну ) - огірковий вірус I ( ВМО - l ) , віруси кільцевих плямистостей кісточкових , вірус аспермії томата , більшість вірусів суниці.
Електронний мікроскоп дозволив визначити не тільки форму і будову , а й розміри вірусних частинок.
Метод включень. Розвиток більшості вірусів у клітині супроводжується утворенням в ній специфічних внутрішньоклітинних відкладень , або , як їх називають , вірусних включень .В одних випадках це скупчення ( кристали , паракристали і т. п. ) самих вірусних частинок , в інших - утворення, що складаються з елементарних частинок вірусу і аморфної речовини клітини. Кожному виду вірусу властива своя форма вірусних включень . Наприклад , вірус тютюнової мозаїки утворює в клітинах голковидні і гексагональні кристали , аморфні тіла , огірковий вірус 2 - паракристали у вигляді коротких голок , вірус мозаїки буряків - веретеноподібні паракристали , вірус жовтяниці буряка - ниткоподібні або стрічкові включення. В інших випадках включення мають В Д сферичних і аморфних тіл .
Внутрішньоклітинні вірусні включення утворюються в основному в клітинах волосків або епідермісу листя і можуть бути виявлені в звичайному біологічному мікроскопі. Для кращого їх виявлення препарати зі зрізами рекомендується фарбувати кислим фуксином .
Хімічний метод діагностики розроблено по відношенню до зеленої і білої мозаїки огірка ( Cucumis virus 2 і Cucumis virus 2 N ) , деяких хвороб плодових культур.
Для аналізу на огірковий вірус 2 беруть зрілий плід ( сім’яник ) , очищають шкірку і розтирають у ступці. З отриманої кашкоподібної маси віджимають через подвійний шар марлі сік і кип'ятять його протягом однієї хвилини. Відфільтровують сік через паперовий фільтр і додають до нього
3 %-ний CuSO4 ( на три частини соку одну частину купоросу ) . Сік з хворих плодів через кілька хвилин стає жовтувато - зеленим ( або блакитним , якщо для аналізу було взято незрілі плоди) . Сік з здорових рослин буде зеленим .
В своїй роботі ми приділили увагу такій овочевій культурі як огірок.
Експеримент проводили на присадибній ділянці, засіяною насінням огірків сорту “ Нежинський“. Знайшли рослини, уражені мозаїчною хворобою ( рис. 2.1 )


Рис. 2.1 Мозаїки огірків
Відповідно методиці :[ 31 ]
1. Відібрали та зважили 500 мг листя ураженої рослини, перенесли в ступку та додали 3мг фосфатного буферного розчину рН 7,4 та розтерли рослинний матеріал.
2. Через марлю відфільтрували рослинний сік у пробірку.
3. Перенесли матеріал у центрифужну пробірку, урівноважили.
4. Відцентрифугували сік у режимі 4000 обертів 20 хвилин.
5. Надосад перенесли в еппендорф, підписали та помістили в холодильник для подальших досліджень.
Ми отримали інокулянт, який потрібний для нашого експерименту.
Інокулянт- інфекційний вірусний матеріал, який застосовується для ураження- інокуляції [ 6 ].
Готували розчини, центрифугували сік на базі лабораторії КЗ
“ХМКЛ “.
Фосфатний буфер(0,1М, рН 7,4)
NCl-8,0г; KH2PO4-0,2г; Na2HPO4-12г; H2O-2,8 KCl-0,2г. Об’єм розчину довели до 1л дистильованою водою.
1. Провели маркування дослідних та контрольних рослин.
2. Зробили укол інокулянта в центральну жилку листка 14.06.2013, що показано на рис. 2.2

Рис. 2.2 Укол іннокулянта
3. Проводили візуальні спостереження за індикацією вірусу на протязі 50 днів. Одержані дані обробляли статистично за методикою Ойвіна, Нью Соколова, 1960 р. і Урбаха, 1963 р. [62].
Обробка проводилася на ЕОМ в середовищі Microsoft Office 2007 у програмі Excel.
При цьому розраховували слідуючі показники:
Середнє арифметичне ; [1]
Помилка середнього [2]
Ступінь достовірності різниці між двома середніми показниками варіантів досліду
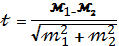 [3]
[3]
В формулах прийняті такі позначення:
- середнє арифметичне,
- сума варіантів досліджуваної сукупності,
- число повторень або вимірів в досліді,
- помилка середнього,
- сума квадратів відхилів від середнього арифметичного,
- ступінь достовірності,
- абсолютна величина різниці між двома середніми,
- квадрати помилок середніх арифметичних.
Цифрові дані виражені в роботі середніми значеннями досліджуваного показника в межах .
Достовірність відмінності порівнювальних показників встановлена при значенні Р<0,05 по таблиці Стьюдента (Урбах, 1963.)
-ступінь свободи , тобто (ступінь свободи =4)
Достовірність досліду визначається по таблиці Стьюдента при ступеню свободи (де Р). Р<0,05.
Точність досліду в %
Якщо %=1-2 (достовірність відмінна)
Якщо% =3-5(достовірність добра)
Якщо% =5-7(достовірність задовільна)
Знаходимо середнє арифметичне для контрольних та дослідних огірків:
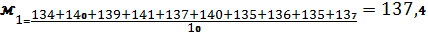
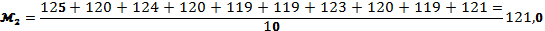
Знаходимо - суму квадратів відхилів від середнього арифметичного: контроль=62,12 ; дослід=44,0
Знаходимо m - помилку середнього, що показано в табл.2.1
Таблиця 2.1
Помилка середнього
Знаходимо t- ступінь достовірності:
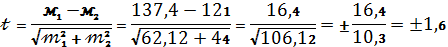
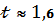
По таблиці Стьюдента при ступені свободи 2 і (1,6)
Р =0,2
% достовірності:
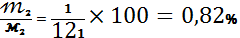
РОЗДІЛ 3. РЕЗУЛЬТАТИ ДОСЛІДУ ТА ЇХ ОБГОВОРЕННЯ
3.1 Народно-господарче і харчове значення культури огірка
Культурна рослина огірок відомий більше 6 тис. років. Його широко обробляли в Китаї , Індії , Єгипті . У III -IV ст. н . е. . З Азії він проник в європейські країни і слов'янську землю. Наприкінці XX - початку XXI століть найбільше у світі огірок вирощується в Росії та Україні . Під його посіви відводиться близько 120 тис. га землі ( 10-12% загальної площі посіву овочевих ) . Це воістину всенародний улюбленець , його вирощують на всіх широтах , від субтропічних до полярних [4]. Вчені - медики прямо пов'язують здоров'я і тривалість життя з якістю харчування , а саме з 1600 г продуктів , споживаних людиною на добу , 1000-1200 г повинно бути рослинного походження, у тому числі овочі - не менше 600 м. [ 4,8 ].
Огірки високо цінуються за хороші смакові якості і лікарські властивості , які проявляються завдяки його дивовижного складу. За кількістю органічної води (95-97 %) вони перевершують всі інші овочі. Плоди містять 4-5% сухої речовини , в тому числі цукру -2 , білкових речовин - 1 , жиру - 0,1 , клітковини - 0,7 , золи - 0,4 , аскорбінову кислоту , провітамін А , вітаміни групи В , біотин , фолієву , нікотинову та пантотенову кислоти. Крім вітамінів, огірки містять безліч мікроелементів: калій, фосфор, магній, кальцій, нікель, натрій, залізо, мідь, кремній, марганець, цинк, титан, цирконій, срібло і кобальт [25,33].
У зв'язку з цим , свіжі плоди огірка мають велике значення в харчуванні населення [ 9,28 ].
Огірки, як і редьку, називають санітарами людського організму. Комплекс солей, що у огірках, сприяє уповільненню процесів старіння, заважає відкладенню каменів в нирках і печінці. Клітковина, якою багаті огірки, регулює роботу травного тракту і виводить зайвий холестерин. Огірки містять тартронову кислоту, яка утримує утворення жиру з вуглеводів. Ця властивість робить їх ідеальним продуктом для тих, хто прагне схуднути [ 33 ] .
Вітаміну B1 в огірках більше, ніж в буряках, в 2 більше, ніж у редисці, а йоду - більше, ніж у будь-яких овочах. Молоді огірочки містять більше аскорбінової кислоти, ніж стиглі. Серед асортименту овочів, вирощуваних у нашій країні, особливе місце займають огірки, площа під якими щорічно складає більше 12% посівних площ овочів. Широке розповсюдження цієї овочевої культури пояснюється, перш за все, традиційними особливостями харчування народу, високими смаковими якостями плодів, що йдуть в їжу як в свіжому, так і в переробленому вигляді.
Харчове значення огірка не в його поживності, а вмісті ферментів і мінеральних солей необхідних для кращого засвоєння іншої їжі. Свіжі і перероблені огірки рекомендують при повищеній кислотності шлункового соку, а також при захворюваннях зобу, печінки, нирок.
Приємний, освіжаючий смак огірків залежить від наявності в них вільних органічних кислот, а характерний запах обумовлюється присутністю в плодах ефірного масла. Ці властивості роблять огірки ідеальним продуктом [ 33 ]. Мабуть, за позитивне значення плодів огірків цій культурі приділяють значну увагу. Так, наприклад:
У місті Шклові , на батьківщині Олександра Лукашенка , у 2007 році відкрито пам'ятник огірку [ 61 ].
У місті Лохвиця (Московська область) встановлено пам'ятник « Огірку - годувальнику » [ 60 ].
У місті Ніжині також встановлено пам'ятник ніжинському огірочки
[ 60 ].
В Ірані огірок вважається фруктом і подається на стіл разом з солодощами .
У селі Істобенське багато років проводиться свято огірка. Даний населений пункт в Кіровській області називають огіркової столицею [ 59 ].
27 липня відзначається Міжнародний День Огірка , саме в цей день вперше в Суздалі було організоване Свято Огірка [ 59 ].
Однак слід зазначити , що всіма перерахованими вище якостями володіють тільки здорові плоди , з чого випливає важливість питання захисту огірка від шкідливих організмів , в першу чергу збудників різних захворювань , вплив яких негативно позначається і на кількості , і на якості врожаю огірка [ 25 ].
Вірусні хвороби цієї культури на 10.90% знижують врожай і значно погіршують якість продукції [ 18 ].
3.2 Ботанічна і біологічна характеристика огірка. Вимогливість культури до умов зовнішнього середовища.
Грунтові сорти огірка (Cucumis sativus L.) представлені сланкими ліанами різної довжини. Стебло (батіг) п'ятигранне, борозенчасте, опушене. У залежності від довжини стебла розрізняють довгоплетинні сорти (> 150 см), короткоплетинні (<60 см) і середньоплетинні (61 ... 150 см)
Відомі також кущові та карликові сорти, довжина стебла у яких не перевищує декількох сантиметрів. Галуження у більшості сортів моноподіальне. Відомий детермінантний тип росту, що характеризується утворенням термінальної квітки і переходом до сімподіального розгалуження. Сорти розрізняються за силою розгалуження. Поряд з що сильно розгалужуються- більше 8 гілок, виділяють слабко-і средньорозгалуженні, які утворюють відповідно 1 ... 4 і 5 ... 8 бічних пагонів. Деякі сорти не гілкуються; найбільш часто це спостерігається у слаборослих детермінантних сортів.
Листя черешкові, які варіюють в межах рослини за разміром і формою. Розташування листя чергове. Нижнє листя відрізняється від наступних меншими розмірами і відносно округлою формою. У пазухах третього-четвертого і наступних листків утворюються вусики, в пазухах формуються чоловічі та жіночі квітки.
Огірок - рослина однодомна, що утворює чоловічі та жіночі квітки . Число жіночих квіток у вузлі може бути різним - 1, 2 і декілька (щиток). Чоловічі квітки утворюються в більшій кількості.
Чоловічі і жіночі квітки можуть формуватися в окремих вузлах (чоловічі та жіночі вузли) або в одних і тих же (змішані) [ 32,33 ]
У пазухах нижніх листків утворюються переважно чоловічі квітки. У міру руху вгору по стеблу співвідношення чоловічих і жіночих вузлів зміщується в бік останніх. Самі верхні вузли головного стебла, як і вузли гілок вищого порядку, можуть бути тільки жіночими [ 27,28 ]
Огірок - ентомофільна перехресноопилююча рослина. Створені партенокарпічні сорти, що утворюють плоди без опилення, що сприяє більш тривалому збереженню товарних якостей плодів та менш частішої вибірки. Партенокарпічні гібриди широко використовують в тепличному виробництві.
Плоди (зеленці) розрізняються за розмірами (5 ... 70 см), масі (20 ... 3000 г), формою, будовою і забарвленням [8, 51 ]. Поверхня плоду - важлива сортова ознака, варіює від неопущенної гладкої до ребристою і горбкуватою з шипами. Опущення (шипи) може бути простим, складним і змішаним. Забарвлення плоду коливається від білого до темно-зеленого. Забарвлення шипів може бути білим, коричневим або чорним. Плоди з чорним опушенням відносно швидко жовтіють. Важлива ознака - здатність до накопичення кукурбітацинів, що обумовлюють гіркий смак, чому сприяють також зовнішні умови (висока температура і недолік вологи, тривалі похолодання, довжина дня).
У одних сортів гіркими можуть бути плоди і листя, у других - тільки листя, і є сорти, не здатні до накопичення кукурбітацинів .
Рослини огірка утворюють розгалужену кореневу систему, що розташовується в основному в орному горизонті. Окремі корені можуть досягти глибини 70 ... 90 см і більше. Огірок легко утворює додаткове коріння з підсім’ядольного коліна і вузлів стебла. З початку проростанню насіння характерно значне випередження формування кореневої системи в порівнянні з надземної. Швидкий ріст коренів пов'язаний з необхідністю хорошої аерації грунту [ 27,43 ].
Огірок відноситься до групи теплолюбних культур, відрізняється високою тепловимогою і не переносить заморозків. Нижня температурна межа для проростання насіння знаходиться на рівні 12 - 13 ° С. Найбільш швидко (через 4- 7 днів) сходи з'являються при температурі грунту
25 - 30 ° С. При середньодобовій температурі повітря 11 - 27 ° С поява сходів розтягується на 7 - 16 днів. [Оптимальна денна температуру в період сходу - цвітіння знаходиться в межах 24 - 28 ° С в ясну і 18 - 22 ° С в похмуру погоду. Бажано, щоб нічна температура в цей час була не нижче 12 - 15 ° С.
Плодоутворення інтенсивніше за все йде при денній температурі 24 - 30 ° С і нічний - вище 16 ° С. При температурі повітря вдень 12 - 15 ° С слабшає зростання, погіршується утворення пилку, зменшуються розміри листя. Подальше зниження температурири в залежності від тривалості веде до пошкодження органів і загибелі рослин [ 2 ].
Для огірка небажані різкі переходи від високої до пониженої температури і, навпаки, від низької до високої, що часто призводить до загибелі посівів. Велике значення має температура грунту. Якщо вона падає нижче 16 ° С, затримується поява сходів, слабшає поглинання води і елементів мінерального живлення, активується патогенна мікрофлора (Pythium та ін), що призводить до загибелі проростків і рослин. При сприятливій температурі грунту рослини огірка значно краще переносять пониження температури повітря [ 4,11,21 ].
Сорти розрізняються по відношенню до тепла, що пов'язане з їх реакцією на екстремальну температуру, коливання температури і тривалість вегетаційного періоду. Далекосхідні сорти мають більш високу, ніж європейські і американські, здатністю адаптуватися до пониженнях і різких коливанням температури.
Температурні умови вегетаційного періоду - основні чинники врожайності культури. Особливо велике значення температури у північних кордонів зони обробки огірка. Непрямий вплив температури виявляється в інтенсивності ураження рослин грибними та вірусними хворобами. Рослини, ураженні вірусом огіркової мозаїки ВОМ-1, втрачають холодостійкість і гинуть при зниженні температури до 15 ° С.
Світло не є лімітуючим фактором при культурі огірка у відкритому грунті. Недолік світла спостерігається лише при дуже загущених посівах. Огірок - рослина короткого дня. Проте різко вираженою фотоперіодичної реакцією володіють лише сорти тропічного і субтропічного походження, не утворюють при літньому вирощуванні в середніх широтах не тільки жіночих, але часто і чоловічих квіток. Вони можуть з'явитисяся лише при температурі нижче 16 ° С.
Селекцією створені весняно-літні сорти огірка для відкритого грунту і тепличної культури, рясно плодоносять в умовах довгого дня, в тому числі майже цілодобового в північних районах. Особливість сортів - інтенсивне використання високої кількості сонячної радіації літнього сезону. У представників цієї екологічної групи сортів можуть спостерігатися невеликі зсуви в проявлення статі в чоловічу сторону в умовах довгого дня і високою температури, що не мають практичного значення в товарному овочівництві, але важливі в гібридному насінництві [24, 25,32 ].
Огірок вимогливий до вологості грунту і відносній вологості повітря (оптимальна вологість грунту близько 80% ПВ і відносна вологість повітря близько 90%). Недолік вологи в грунті призупиняє зростання, веде до втрати тургору в денний час, сприяє зсуву статі в чоловічу сторону і появленню гіркоти в плодах. Перезволоження грунту призводить до загибелі коренів.
Огірок - культура, яка дуже сильно реагує навіть на короткочасне затоплення [ 28 ].
Суттєво впливає на продуктивність фотосинтезу і зростання відвідносна вологість повітря. В умовах перегріву і низької відносної вологості повітря ефективні освіжаючі поливи дощуванням (50 м3 / га).
З урожаєм огірків виноситься порівняно невелика кількість поживних елементів, однак темпи споживання їх рослинами дуже високі, що знаходиться у відповідності з ходом на ¬ ростания всієї маси рослини. До особливостей харчування огурків слід віднести його чутливість до концентрації грунтового розчину і реакції грунту (оптимальне значення рН 6 ... 6,5) [ 2,8 ]
Вельми ефективне внесення під огірок поряд з мінеральними органічних добрив у високих дозах. Вони не тільки поліпшують режим мінерального живлення, але і оптимізують фізичні властивості грунту, її тепловий режим, активізують грунтову мікрофлору, значно збільшують виділення СО2 [ 8,28 ]
Разом з тим, нажаль, рослини огірків вражаються вірусами і найбільш пошириними вірусними хворобами є такі: вірус зеленої крапчастий мозаїки огірка, біла мозаїка огірка, плямистість листя огірка, мозаїка огірка і тд.
3.3 Захворювання огірка, які передаються насінням

Рис. 3.1 Cucumber green mottle mosaic tobamovirus (CGMM) / вірус зеленої крапчастої мозаїки огірка ВЗКМО
Біологія патогена
Симптоми зеленої крапчастий мозаїки
Вірус зеленої крапчастий мозаїки огірка (CGMM) (рис. 3.1) вражає невелике число видів рослин. Патоген відносять до РНК вірусів (група тобамовірусов), що викликає системне ураження рослин сімейства гарбузових. Цей вид вражає тільки огірок, диню і кавун і не відомий на гарбузі і шинку. Цим він відрізняється від вірусу огіркової мозаїки, який має широке коло господарів [ 42 ].
Збудник надзвичайно стійкий до дії екстремальних чинників зовнішнього середовища: віріон залишається життєздатним при нагріванні до 90 ° С, при висушуванні і заморожуванні. Патоген легко передається від хворої рослини до здорової, здатний зберігатися в сухому листі протягом року.
Біла мозаїка огірка
Симптоми білої мозаїки, викликаної агресивнішим штамом вірусу,
на молодих рослинах виявляють через 20-30 днів після висадки розсади на постійне місце, причому, частіше після різкого підвищення температури повітря до 30 ° С. На уражених рослинах розвиваються зморшкуваті, редуковані листя, кількість жіночих квіток і відповідно плодів зменшується. Зав'язані плоди уповільнено розвиваються, часто деформуються або набувають мозаїчного забарвлення, їх якість значно погіршується.
Найбільш патогенний штам (CGMM 2А) викликає симптом білої мозаїки. Перші ознаки цієї форми хвороби проявляються на молодих зростаючих листках: уздовж жилок утворюються просвітління, жовті кільця і плями зірчастої форми. Кільця і плями швидко розростаються і набувають білий, рідше жовтий колір. Через деякий час плями зливаються, в результаті чого більша частина листової пластинки стає білою. Подібний білий мозаїчний малюнок може спостерігатися і на плодах. (рис. 3.2)


Рис.3.2 Прояви білої мозаїки на листях та плодах.
Джерела інфекції: рослинні залишки, насіння (віруси знаходяться як в шкірці, так і в зародку) і грунт [ 23 ]. Вторинне зараження відбувається при попаданні інфікованого соку на здорову рослину в процесі збору плодів. Зараження можливе навіть при зіткненні заражених і здорових рослин. Віруси можуть зберігати інфекційність у висушеному вигляді, перебуваючи на інвентарі. Рослини-резерватори - представники сімейства гарбузові. Біла мозаїка знижує урожай на 50-90%.
Агротехнічні заходи
Використання стійких гібридів F1: ОФІКС, ОКТОПУС, Пасамонте, Пасадена, ПАСАЛІМО. Дотримання сівозміни. Використання вільних від інфекції насіння. У насінні з терміном зберігання більше 2-х років зараженість помітно знижується[ 49,52 ].
Термічне знезаражування насіння за такою схемою:
нагрівання при 70 ° С протягом 3 діб;
нагрівання протягом 3 діб при 50-52 ° С з подальшим нагріванням протягом 24 годин при 78-80 ° С.
Ця процедура повністю звільняє насіння від збудника. Просторова ізоляція посадок гарбузових культур знижує темпи розвитку епіфітотії. Поширення захворювання можна обмежити обприскуванням 10% розчином знежиреного молока або розчином мікроелементів [ 38,41 ]
Зелена крапчаста мозаїка огірка (ВЗКМО).
Вірус зустрічається повсюдно в захищеному грунті.Одним із штамів ВЗКМО є збудник білої мозаїки огірка - ВМО-2А [ 48,52 ].
Ураження рослин огірка ВМО-2А відзначається, як правило, в літній час, коли денна температура в теплицях перевищує 30 ° С.
Поразка огірка вірусом зеленої крапчастий мозаїки у виробничих умовах проявляється спочатку на молодих листках - у вигляді просвітления тканини уздовж жилок. Потім з'являється чітка мозаїка - чергування світло-зелених і жовтуватих ділянок листка з темно-зеленими міхуровидними роздутими ділянками. Листя хворих рослин зморшкуваті і дрібні. Ріст рослин пригнічений.
Симптоми на плодах проявляються у вигляді яскравого мозаїчного забарвлення, зазвичай без деформації.
Першоджерелами вірусу є насіння, зібране з хворих рослин, заражений грунт або штучний субстрат [ 47,49,51]
Відсоток поширення вірусу з насінням коливається в значних межах: від десятих часток відсотка до 15%. Ступінь передачі насінням залежить від штаму вірусу, що викликав ураження рослин, віку рослин у момент зараження, тривалості зберігання насіння, сорту, умов вирощування рослин та інших факторів. Найвищий відсоток передачі спостерігається при посіві свіжозібранного насіння. Вірус не передається через насіння дворічного терміну зберігання.
ВЗКМО має дуже високу інфекційність, легко передається соком від хворих рослин при щепленні, підв'язці, зборі плодів та інших виробничих операціях.
ВЗКМО (у тому числі ВМО-2А) зберігаються в грунті в рослинних залишках, хоча з часом частково втрачають інфекційність. При посадці рослин через 4-6 місяців після видалення старих заражаються тільки окремі рослини.
На жаль, багато тепличних господарств країни перейшли на монокультуру огірка, і якщо раніше ВЗКМО частіше діагностували в кінці вегетації, то тепер його можна виявити і виділити вже в розсадному відділенні.
При вирощуванні огірка на штучних субстратах ВЗКМО з'являється раніше і розвивається більш інтенсивно, ніж при вирощуванні в грунті.
ВЗКМО може зберігати інфекційність на внутрішній поверхні теплиці, шпалері, з якими стикалися хворі рослини, а також на тарі, інвентарі та одязі робітників.
Гібридів огірка, стійких до вірусу зеленої крапчастої мозаїки, поки немає, тому необхідний комплекс профілактичних заходів, спрямованих на зниження його шкодочинності і обмеження розповсюдження.
У зв'язку з тим, що першоджерелом ВЗКМО на культурі тепличного огірка є насіння, основна увага в боротьбі з цим вірусом має бути приділена питанням використання здорового або знезараженого насіннєвого матеріалу.
Найбільш ефективно знезараження насіннєвого матеріалу від ВЗКМО комбінованим методом, що поєднує термічну обробку з передпосівною обробкою в 15% розчині тринатрійфосфата (хімічно чистого) протягом 1 години з наступним промиванням у проточній воді (не менше 50 хвилин) . Це дозволяє знезаразити не тільки шкірку, а й плівчату оболонку насіння, тобто саме ті його частини, в яких локалізується вірус [ 29,34,38 ]
Ще одне джерело ВЗКМО - заражений грунт або гідропонний субстрат, куди інфекція потрапляє з виділеннями коренів хворих рослин, а також з рослинними залишками. Слід враховувати, що фунгіциди, ефективні проти збудників грибних хвороб огірка типу плямистостей, вірус не інактивують.
У теплицях з високим рівнем поширення зеленої крапчастий мозаїки необхідно ретельно пропарювати грунт протягом 1-2 годин при температурі 90 ° С або замінити його на новий, вільний від вірусу.
Щоб запобігти перенесення вірусу з внутрішньої поверхні теплиць, рекомендується проводити випал шпалер газовими пальниками [ 38,41].
Необхідно проводити фітопрочистки, починаючи з розсадного відділення, знезаражувати руки, ножі, рукавички 1% розчином KMnO4, 5% тринатрійфосфатом (Na3PO4) або 10% обратом. Під час вегетації рослин важливо підтримувати оптимальні умови температури і освітленості, не допускаючи різких перепадів нічних і денних температур. Рекомендується обмежувати дози азотних добрив, 1-2 рази на тиждень проводити некореневі підгодівлі з мікроелементами, що підвищує неспецифічну стійкість рослин до несприятливих факторів середовища.
При високому фоні ВЗКМО неприпустимо ведення монокультури (повернення культури огірка в теплиці, де було виявлено вірус зеленої крапчастої мозаїки, можливо не раніше ніж через 1-2 роки) [ 2,4 ].
Плямистість листя огірка (ВПЛО)
Збудник - Cucumber leaf spot virus. Інфекційність соку in vitro зберігається протягом 20 днів [20,48].
На молодих листках рослин спочатку з'являються світло-зелені або жовтуваті з нечіткими межами плями з коричневим некротичним центром. У більшості випадків вони спочатку виникають на краю листа і надалі поширюються на всю листкову пластинку. На плодах симптоми не виявлені. Хвороба спостерігається тільки при осінній та зимовій посадці огірків[47,49].
Вірус легко передається з соком хворих рослин методом механічної інокуляції, а також насінням. Попелиці переносниками вірусу не є. Він виявлений у грунтових пробах, взятих у районі природно інфікованих рослин огірка, і, мабуть, може інфікувати здорові рослини огірка без переносників. Основний господар - гарбузові [ 38, 46 ].
3.4 Захворювання огірка, поширювані попелицями
Збудник - Cucumber mosaic virus. Вірус залишається інфекційним in vitro при кімнатній температурі від 18 годин до 14 діб, а при температурі від -5 до 15 ° С - до 2-5 місяців. У сухих мозаїчних листках огірка він зберігає інфекційність до 50-60 днів, а в мозаїчних плодах огірка при 3 - 5 ° С
до 30-ти [ 30,51 ].
При ураженні рослин огірка ВМО на листках з'являються жовто-зелені плями, що розташовуються в основному між жилками, але іноді захоплюючі і жилки. Ці плями не так чітко відокремлені від темно-зелених ділянок листка, як при ураженні ВЗКМО. У молодих листків відзначаються деформація і зменшення розмірів листя. Черешки листя і міжвузля стебел коротшають.(рис. 3.3)

Рис.3.3 Мозаїка огірка (ВМО)
На плодах в більшості випадків симптоми не спостерігаються, іноді з'являються світлі плями.
Насінням огірка вірус не передається. Вірусна інфекція зменшує продуктивність насіннєвих рослин.
За останніми даними список рослин - господарів ВОМ включає 775 видів , що відносяться до 365 родів і 85 родин. Стійка циркуляція ВВМ в природі здійснюється за допомогою попелиць - переносників вірусу. Відомо більше 20 видів попелиць , здатних передавати вірус з дикорослих рослин на культурні . У більшості випадків епіфітотійного ситуацію визначають такі види, як Aphis gossipii , A. nasturtii і Myzus persicae . Найбільш часто епіфітотійного ситуацію провокує висока чисельність M. persicae .
При ураженні тепличних огірків ВМО втрати врожаю досягають 50-96% і залежать від сорту та віку рослин у момент зараження [ 15,17,23 ].
3.5 Індикація вірусу та вплив інокуляції збудника на ріст, розвиток та врожай огірка.
Індикація вірусу-визначення наявності чи відсутності вірусу у досліджуванному матеріалі.
Для експерименту було іннокульовано 10 рослин огірка та взято для контролю 10 здорових культур. Інокуляцію рослини провели 14.06.13р.
Починаючи з 21.06.2013, через 7 діб після інокуляції, ми брали для дослідження проби кожні 7 днів. Під час візуальних спостережень ми констатували факти зміни: контрольні рослини залишились не вражені, а на дослідних спочатку спостерігалось зав’ядання листків, а потім зміна забарвлення – поява жовто-зелених плям, а потім смугастість. По мірі досліду рослини відставали в рості і з’являлись пошкодженні плоди.
Результати, отримані в ході спостережень, піддавалися дисперсійному аналізу за алгоритмами, запропонованими ( Ойвін, Нью, Соколов,1960)
В ході досліду враховували зовнішні показники: температуру повітря та відносну вологість, що відображено в табл..3.1
Таблиця 3.1
Температура та відносна вологість повітря
|
Дати
|
21.06
|
28.06
|
05.07
|
12.07
|
19.07
|
26.07
|
01.08
|
03.08
|
05.08
|
07.08
|
|
Температура в 0С
|
21
|
26
|
25
|
23
|
22
|
25
|
26
|
23
|
25
|
23
|
|
Відносна вологість повітря в %
|
74
|
78
|
70
|
68
|
72
|
74
|
80
|
74
|
68
|
70
|
В період експерименту ми проводили візуальні спостереження, заміри рослин і ваги плодів. Спостерігали: появу жовто-зелених плям, вкорочення живців листя, пагонів, білі плями на плодах. Заміри висоти рослин проводили кожні 7 днів. Результати представлені в таблиці 3.2
Таблиця 3.2
Вплив вірусної хвороби на ріст і приріст рослин (висота в см)
|
Дата
|
Висота рослин
|
Приріст
|
|
|
I варіант
|
II варіант
|
I варіант
|
II варіант
|
|
21.06
|
14,0±1,8
|
14±1,8
|
–
|
–
|
|
28.06
|
19,0±1,8
|
16,4±1,8
|
5,0
|
2,4
|
|
05.07
|
24,0±2,0
|
19,4±1,8
|
5,0
|
3,0
|
|
12.07
|
39,2±2,3
|
24.3±2,0
|
15,2
|
5,0
|
|
19.07
|
58,8±2,1
|
30.5±2,2
|
19,6
|
6,0
|
|
26.07
|
78,4±2,5
|
40,5±2,1
|
19,6
|
6,0
|
|
01.08
|
98,0±2,5
|
53.2±2,4
|
19,6
|
6,7
|
|
03.08
|
113.8±2,4
|
63,5±2,6
|
15,8
|
10,3
|
|
05.08
|
130,2±2,6
|
74,2±2,5
|
16,4
|
10,7
|
|
07.08
|
136,0±2,5
|
83,0±2,6
|
5,8
|
8,8
|
Більш наглядніше це видно на рис. 3.1
Рис. 3.1 Вплив вірусної хвороби на ріст рослин(висота в см)
Отже, з наведеної діаграми ми бачимо, що за період часу з 21.06.2013 по 07.08 2013 контрольні здорові рослини виросли на 53,0 ±2 см більше ніж дослідні інокульовані рослини огірка.
Підраховували кількість плодів, що показано в табл..3.3
Таблиця 3.3
Вплив вірусної хвороби на кількість плодів рослин огірків
|
|
Кількість плодів, шт
|
|
Дата
|
Контрольні рослини
|
Дослідні рослини
|
|
1.08
|
10±1,5
|
5±1,1
|
|
3.08
|
14±1,7
|
7±1,3
|
|
5.08
|
19±1,6
|
13±1,2
|
|
7.08
|
21±2
|
15±1,4
|
Рис. 3.2 Вплив вірусної хвороби на кількість плодів під час розвитку рослин огірків
Отже, з наведеної діаграми ми бачимо, що за період часу з 01.08.2013 по 07.08.2013 кількість плодів з здорових рослин перевищила кількість плодів з уражених рослин на 21±1,7 огірка.
Визначали вагу плодів, що наведено в табл..3.4
Таблиця 3.4
Вплив вірусної хвороби на вагу плодів огірків
|
|
Вага плодів, в грамах
|
|
Дата
|
Контрольні рослини
|
Дослідні рослини
|
|
1.08
|
98,7±0,9
|
36,0±0,8
|
|
3.08
|
112,0±1,0
|
47,7±0,7
|
|
5.08
|
116,7±1,2
|
56,2±0,6
|
|
7.08
|
120,3±1,1
|
67,8±0,8
|
Рис. 3.3 Вплив вірусної хвороби на вагу плодів огірків
Отже, з наведеної діаграми ми бачимо, що за період часу з 01.08.2013 по 07.08.2013 вага плодів з здорових рослин перевищила вагу плодів з уражених рослин в середньому на 57,8±0,8 грам.
Вплив вірусної хвороби на врожай огірків загалом показаний в табл..3.5
Таблиця 3.5
Вплив вірусної хвороби на врожай огірків
|
|
Варіанти
|
|
|
Контроль (здорові рослини)
|
Дослідні рослини, вражені, Cucumis virus 1
|
|
Вага плоду огірка 07.08.2013 в грамах
|
120,3±0,9
|
67,8±0,8
|
|
Кількість плодів на рослині 07.08.2013
|
21±2
|
15±2
|
|
Загальна вага плодів з рослини за вегетаційний період
|
7294±20,1
|
2231,5±18,3
|
|
Кількість уражених(пошкоджених) плодів на рослині
|
0
|
2±2
|
Рис. 3.4 Вплив вірусної хвороби на врожай огірків
Вірусна хвороба огірка ВМО знизила врожай на 32,68%.
Таким чином, ми можемо констатувати, що мозаїка огірка викликає зміни росту, ваги, кількості плодів, врожаю загалом у рослини. Крім того, плоди огірків втрачають свій товарний вигляд (рис. 3.5) і харчові якості.
Рис. 3.5 Інфіковані плоди огірка
Але з цими недоліками можна боротися. [5]
3.6 Боротьба із вірусними хворобами рослин
Вибір та застосування різних заходів боротьби з вірусними хворобами залежить насамперед від виду хвороби і видів рослин, на які вона поширюється. Загальними для боротьби з вірусними хворобами рослин є такі заходи:
1) виведення і впровадження у виробництво стійких проти вірусів сортів рослин;
2) знищення комах та інших переносників вірусів;
3) ліквідація джерел поширення вірусів у природі (це передусім своєчасне лущення стерні і зяблева оранка, знищення бур'янів на полях, межах, узбіччях доріг, які є джерелом інфекції; дотримання оптимальних строків сівби і проведення противірусного прополювання насіннєвих посівів);
4) використання для сівби здорового безвірусного насінного матеріалу.
Останніми роками серед профілактичних заходів, які запобігають араженню рослин вірусами, поширення набуває :
1) вакцинація рослин (переважно закритого ґрунту) ослабленими штамами вірусів. Інтенсивно проводяться також роботи з
2) оздоровлення рослин методами термотерапії (з використанням спеціальних термокамер), щоб отримати безвірусні клони,
3) хіміотерапії,
4) культур верхівкових меристем тощо.
З появою терміну « нанотехнологія » медики задумалися про можливість створити такий тип лікарських препаратів , які немов би точний бомбовий удар проникали у вогнище захворювання і усували його причину[78]. Так виникла концепція адресної доставки медичних препаратів - воістину проблема третього тисячоліття , - і з самого моменту появи вона стала щорічно зазнавати новий виток розвитку. Один з таких витків стався на початку 2013 року. І пов'язаний він , як це не дивно , з вірусом ... мозаїки огірка. Славне минуле , інтригуюче теперішнє , багатообіцяюче майбутнє - все є у цього вірусу. У чому зв'язок між зараженим вірусом огірком і протипухлинним препаратом доксорубіцином? Як вони разом можуть стати «чарівною кулею» проти онкологічних захворювань? [ 35 ].
А недавно, навесні 2013 року, китайські вчені з Південного Медичного Університету відкрили нову сторінку в вірусології та медицині під назвою «Drug Delivery». Китайський рецепт простий: взяли вірус мозаїки огірка, протипухлинний препарат доксорубіцин, змішали і - на наступний день отримали готову систему вірус-ліки [ 54 ].
Вірус мозаїки огірка ( Cucumber mosaic virus , CMV ) - 29- нанометровий ікосаедричний вірус сімейства Bromoviridae , геном якого представлений п'ятьма одноланцюговою РНК з позитивною полярністю ( має властивості інформаційної РНК) , кожна з яких оточена своєю білковою оболонкою. CMV - класичний приклад вірусу з ікосаедріческой симетрією , що складається з Пентон і гексоній [ 52 ] . Пентамірні і гексамірні капсомери формують отвори ( пори) , і в підсумку білкова оболонка вірусу пронизана наскрізними каналами.
Це не заважає вірусу нормально існувати , а дослідникам ця пористість тільки на руку !
Рис. 3.6 Електронна мікрофотографія вірусу мозаїки огірка.
Стрілками відзначені Пентон і гексоній для збільшення ймовірності потрапляння препарату в пухлину вчені застосували принцип «активного націлювання » , а як « прицілу » вибрали класичну в онкології молекулу фолієвої кислоти. Причина - підвищений рівень експресії рецептора фолієвої кислоти ( folic acid receptor , FR) на пухлинних клітинах у порівнянні зі здоровими клітинами. Таким чином , фолієва кислота взаємодіє зі своїм рецептором , викликаючи так званий « рецептор -опосередкований ендоцитоз » вірусу ( простіше кажучи , його поглинання клітиною ) . Примітно , що клітини , на які вчені націлили свою нову зброю , відрізняються ще й тим , що на них рецептора FR дуже багато - це культура пухлинних клітин OVCAR - 3 (клітини раку яєчника людини ) .
«Завантаження» доксорубіцину в капсид здійснюється через ті самі пори, які так добре видно на електронних мікрофотографіях на малюнку 3.6.
Вільний доксорубіцин викликає розширення всіх камер серця, появу пристінкових тромбів, розрізненість кардіоміоцитів і т.п. При використанні досягнень спрямованої доставки ліків пухлини втрачали характерний дольчатий вигляд і набували EPR-пошкодження. І пухлина
переможена! [ 52 ].
І тут постає питання: вірус друг чи ворог?...
РОЗДІЛ 4. ОХОРОНА ПРАЦІ