НИТРИФИЦИРУЮЩИЕ ПРОКАРИОТЫ
МИНИСТЕРСТВО ОБРАЗОВАНИЯ РЕСПУБЛИКИ БЕЛАРУСЬ
Учреждение образования
«ПОЛЕССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
БИОТЕХНОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
КАФЕДРА БИОТЕХНОЛОГИИ
НИТРИФИЦИРУЮЩИЕ ПРОКАРИОТЫ
Курсовая работа
Специальность 1-31 01 01 Биология (по направлениям)
направление специальности 1-31 01 01-01 Биология (научно-производственная деятельность)
Исполнитель:
студент _5_ курса 1032411 группы
Климчук Сергей Анатольевич
заочной формы обучения _________
Научный руководитель:
канд. биол. наук
Юрченко Евгений Олегович _________
ПИНСК 2014
Реферат
Курсовая работа: 52 с., 2 табл., 1 рис., 1 фото, 31 источников.
НИТРИФИЦИРУЮЩИЕ БАКТЕРИИ, АЭРОБНЫЕ ХЕМОЛИТОТРОФЫ, ГРАМ-ОТРИЦАТЕЛЬНЫЕ БАКТЕРИИ, НИТРИФИКАЦИЯ, КРУГОВОРОТ АЗОТА, ПИТАТЕЛЬНАЯ СРЕДА, ГНИЕНИЕ.
Объектом и предметом исследования являлись доступные из публикаций сведения описывающие особенности строения и функционирования нитрифицирующих бактерий, процесс нитрификации и влияющие на него факторах, значение нитрифицирующих бактерий в природе и жизни человека.
Цель работы: собрать и обобщить доступные из публикаций сведения по данной теме, сделать аналитические выводы.
Задачи: сделать обзор и обобщить доступную информацию касающуюся истории открытия нитрифицирующих бактерий, их научной классификации, строении и свойствах, условиях существования; рассмотреть химический процесс последовательного окисления аммиака до азотистой и азотной кислот.
Автор работы подтверждает, что все заимствованные из литературных и других источников теоретические, методологические и методические положения и концепции сопровождаются ссылками на их авторов.
Оглавление
Стр.
ВВЕДЕНИЕ 4
ГЛАВА 1 НАУЧНАЯ КЛАССИФИКАЦИЯ
1.1 Принципы классификации прокариот 5
1.2 Научная классификация нитрифицирующих бактерий 7
ГЛАВА 2 ОБЩАЯ ХАРАКТЕРИСТИКА
2.1 История открытия 9
2.2 Краткая характеристика хемолитотрофных бактерий 11
ГЛАВА 3 ХАРАКТЕРИСТИКА БАКТЕРИЙ СЕМЕЙСТВА NITROBACTERIACEAE
3.1 Особенности строения и функционирования 14
3.2 Бактерии первой фазы нитрификации 16
3.3 Бактерии второй фазы нитрификации 19
ГЛАВА 4 ОПИСАНИЕ ПРОЦЕССА НИТРИФИКАЦИИ
4.1 Химизм процесса нитрификации 22
4.2 Условия, влияющие на процесс нитрификации 26
4.3 Влияние адсорбентов на процесс нитрификации 28
ГЛАВА 5 НИТРИФИЦИРУЮЩИЕ ОРГАНИЗМЫ В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА
5.1 Значение хелимолитоавтотрофов в круговороте веществ 31
5.2 Круговорот азота и роль в нем нитрифицирующих бактерий 35
5.3 Процессы нитрификации в почве 41
5.4 Применение нитрифицирующих бактерий в биоиндикации 44
ЗАКЛЮЧЕНИЕ 48
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 50
ВВЕДЕНИЕ
Биосфера заселена огромным числом живых существ. Одни из них составляют макромир, другие - микромир. К макромиру относятся животные, птицы, растения, насекомые и другие видимые невооруженным глазом существа; микромиру принадлежат не видимые невооруженным глазом мельчайшие живые существа, имеющие как растительное, так и животное происхождение; это главным образом бактерии, грибы, простейшие и вирусы. По ориентировочным подсчетам, в биосфере обитает не менее 1030 бактерий, а число вирусов, простейших не поддается учету.
Микроорганизмы в зависимости от молекулярно-биологической организации подразделяют на прокариотов и эукариотов. Прокариоты (от греч. karyon - ядро) доядерные простейшие одноклеточные формы жизни, не имеющие ядерной мембраны и высокоорганизованных органелл.
Особое место среди прокариотов занимают нитрифицирующие бактерии (лат nitro — азот и faceo — делаю).
ГЛАВА 1 НАУЧНАЯ КЛАССИФИКАЦИЯ
- Принципы классификации прокариот
В настоящее время в микробиологии существует два различных подхода к систематике, обуславливающие два принципа классификации. Одни исследователи считают, что классификация должна отражать историю развития организмов и строиться на филогенетической основе. Это естественная классификация. В 1936-1950 гг. исследователи голландской школы А. Юпойвер, К. ван Ниль и Р. Стейниер разрабатывали филогенетические подходы к систематике бактерий. Второй подход к систематике основан на учете легко определяемых признаков, удобных с точки зрения практики. Это искусственная, или традиционная классификация. Она чаще всего заранее подчинена конкретным задачам. При такой классификации признаки, определяющие принадлежность организма к той или иной таксономической единице, часто выбираются произвольно. Иногда микроорганизмы объединяются в группы на основании одного ведущего признака. Поэтому группы микроорганизмов, классифицированные по данной систематике, могут иметь большое практическое значение, но не соответствуют естественной классификации. Искусственная классификация положена в основу определителя Берги, предназначенного для идентификации бактерий по фенотипическим признакам и содержит сведения о видах бактерий [8].
Основой определения систематического положения являются: морфология и тинкториальные свойства клеток (форма, размеры, взаимное расположение, спорообразование, окраска по методу Грама и другими методами), культуральные, биохимические, антигенные характеристики, а также чувствительность к различным антимикробным воздействиям и степень генетического родства с представителями других таксонов (по процентному соотношению содержания гуанина и цитозина в геноме, гомологии нуклеиновых кислот и способности к обмену генетической информацией), патогенность и вирулентность, степень опасности для человека [5, 20].
Различают следующие таксономические категории: царство, подцарство, отдел, класс, порядок, семейство, род, вид и подвид и др. В рамках той или иной таксономической категории выделяют таксоны – группы организмов, объединенные по определенным однородным свойствам. Названия микроорганизмов регламентируются Международным кодексом номенклатуры бактерий. Название вида соответствует бинарной номенклатуре, т.е. состоит из двух слов. Например, возбудитель брюшного тифа пишется как Salmonella typhi. Первое слово - название рода начинается с прописной буквы, второе слово (название вида) – со строчной. При повторном написании, родовое название сокращается до начальной буквы, например S.typhi [12].
Наиболее употребительными во всем мире были классификации Кауфмана [Kauffmann F., 1963-1966], позднее Юинга [Ewing W., 1967-1972] [30].
Однако в 1974г. Вышел капитальный труд, созданный ведущими бактериологами разных стран 8-е издание Bergeys Маnual оf Determinative Bacteriology «Определитель бактерий Берги», где главными критериями для разделения бактерий на крупные таксоны служили морфология, физиология и биохимические свойства, отчасти генетический анализ. Для дифференциации более мелких категорий некоторых бактерий использованы особые признаки — серологические, отношение к бактериофагам и колицинам [30]. Первое издание Определителя было выпущено в 1923 г. группой американских бактериологов под руководством Д.X.Берги (D. Н. Bergey, 1860—1937) [9, 16].
Принципы идентификации, изложенные в Определителе бактерий Берги, 9-е издание которого вышло в 1994 г., нашли наибольшее распространение и признание в практической бактериологии. В настоящее время бактерии классифицируют в соответствии с Определителем Берги, выпускаемом периодически Международным Комитетом по тематике прокариот с привлечением крупных специалистов разных стран. Ценность Определителя Берги в том, что он представляет собой наиболее полную сводку известных бактерий и содержит необходимые тесты для их идентификации [8]. В СССР был составлен его сокращенный вариант на русском языке – «Краткий определитель бактерий Берги» (1980) [30].
- Научная классификация нитрифицирующих бактерий
В «Определителе бактерий-9» (1984-1989) прокариоты в зависимости от строения клеточной стенки разделены на 17 частей. Нитрифицирующие бактерии относятся к Части 12 – Аэробные хемолитотрофные бактерии и близкие к ним организмы. Группа разделена на 3 подгруппы в зависимости от химической природы окисляемых неорганических соединений [9].
Подгруппа 1: Бесцветные сероокисляющие бактерии. В подгруппе объединены бактерии, способные окислять неорганические восстановленные соединения серы, образуя в качестве конечного продукта сульфат. Семейство Thiobacillaceae. Обитатели почвы, воды, серных ключей или серных месторождений. Их можно обнаружить невооруженным глазом по наличию обильных белых отложений серы, взвесей, тяжей, розеток или пленок.
Подгруппа 2: Железо- и марганец-окисляющие и/или осаждающие бактерии, получают энергию в процессе окисления двухвалентного железа или марганца кислородом воздуха. Семейство Siderocapsaceae и несколько других родов. Они способны откладывать оксиды железа или марганца вне клетки. В основном обитают в железосодержащих водах.
Подгруппа 3: Нитрифицирующие бактерии, получают энергию в процессе окисления аммиака до нитрата. В подгруппу включены бактерии семейства Nitrobacteriaceae.
Процесс окисления аммиакиа до нитрата происходит в два этапа. На первом этапе происходит окисление аммимка до нитритов, а на втором нитриты окисляются до нитратов. Эти два процесса требуют различных наборов нитрифицирующих ферментов и выполняются двумя различными группами бактерий. В зависимости от того, какую фазу процесса осуществляют нитрифицирующие бактерии их причисляют к одной из двух секций. Секция А: окисление аммиака до нитритов производят бактерии родов Nitroso- : роды Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira и др. Секция В: окисление нитритов до нитратов производят бактерии родов Nitro- : Nitrobacter, Nitrospira, Nitrococcus [21].
Таким образом, научная классификация нитрифицирующих прокариот выглядит следующим образом:
Царство: Procariotaе
Подцарство (домен): Bacteria
Отдел: Gracilicites
Класс: Scotobacteria
Группа Proteobacteria
Семейство: Nitrobacteriaceae
Секция А
Род: Nitrosomonas
Род: Nitrosospira
Род: Nitrosococcus
Род: Nitrosolobus
Секция В
Род: Nitrobacter
Род: Nitrococcus
Род: Nitrospira
Так же можно придерживаться другой классификацией, по которой нитрифицирущие бактерии относятся к двум типам Nitrospira и Proteobacteria, пяти классам Nitrospira, Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria и Deltaproteobacteria [32]:
Домен Bacteria
Тип Nitrospira или Nitrospirae
Класс Nitrospira
Порядок Nitrospirales
Семейство Nitrospiraceae
Род Nitrospira
Тип Proteobacteria
Класс Alphaproteobacteria
Порядок Rhizobiales
Семейство Bradyrhizobiaceae
Род Nitrobacter
Класс Betaproteobacteria
Порядок Nitrosomonadales
Семейство Nitrosomonadaceae
Род Nitrosolobus
Род Nitrosospira
Род Nitrosomonas
Класс Gammaproteobacteria
Порядок Chromatiales
Семейство Ectothiorhodospiraceae
Род Nitrococcus
Семейство Chromatiaceae
Род Nitrosococcus
Класс Deltaproteobacteria or Deltabacteria
Порядок Desulfobacterales
Семейство Nitrospinaceae
Род Nitrospina
ГЛАВА 2 ОБЩАЯ ХАРАКТЕРИСТИКА
2.1 История открытия
Биологическая природа образования в почве нитратов была установлена во второй половине прошлого века. Предположение об участии микроорганизмов в этом процессе было высказано Л. Пастером. Биологическая сущность процесса нитрификации была доказана работами Т. Шлезинга и А. Мюнца в 1879 г [6]. В своей работе ни медленно пропускали сточные воды, содержащие аммиак, через длинную трубку, наполненную песком и калием, и установили, что через 20 дней в конце трубки выходил нитрат, а не аммиак; хлороформ или кипяток уничтожали эту способность аммиака к окислению, но при дополнительном внесении почвы этот процесс возобновлялся. Р. Уорингтон (во многих сообщениях 1878 и 1891 года) подтвердил эти открытия и пошел дальше, а именно он доказал участие двух организмов в этом процессе [26].
Однако выделение нитрифицирующих бактерий в чистую культуру и их описание являются заслугой Виноградского (1891 год). Для получения плотной минеральной питательной среды он использовал вместо агара силикагель. Он обнаружил, что в окислении аммония в нитриты и далее нитритов в нитраты принимают участие два различных организма - Nitrosomonas и Nitrobacter. Обе бактерии облигатно автотрофны. Изучением роста этих экзотических бактерий занимался О. Мейергоф (1916, I917). Последующие опыты по выделению бактерий убедили многих почвенных микробиологов в том, что в окислении аммиака в природе литоавтотрофные нитрификаторы занимают монопольное положение [12].
Новые роды, и среди них факультативные автотрофы и миксотрофы были описаны самим Виноградским (1892, 1933) и далее С. В. Уотсоном (в 1965-1975 годах) и Э. Боком с сотрудниками (1962-1975) [3,9].
Сообщение об образовании N2O нитрифицирующими микроорганизмами опубликовано Корбе в 1935 г. Автор обнаружил выделение N2O культурой микроорганизмов при окислении аммония или гидроксиламина в нитрит и предположил, что закись азота образовывалась либо при химическом разложении гипонитрита, либо при химическом взаимодействии гидроксиламина с нитритом.
Иошида и Александер в 1970 г. описали образование N2O суспензиями клеток Nitrosomonas europaea при окислении аммония и гидроксиламина; при этом до 95% окисленного азота гидроксиламина выделялось в форме N2O. Авторы сделали вывод, что N2O образуется из промежуточного продукта нитрификации, дальнейшее превращение которого в нитрит ингибируется аммонием, гидразином и температурой выше 35°С. Следует отметить, что такое значительное образование N2O было получено только в этой работе для клеток N.europaea, выдержанных в течение нескольких дней на холоду, и в дальнейшем не наблюдалось авторами, создававшими для клеток N.europaea физиологические условия [10].
Отмечено образование N2O культурой Nitrosospira briensis, растущей в присутствии аммония (Веденина, Заварзин, 1979) [15].
Блэкмер с соавторами (1980) описали выделение N2O в аэробных условиях из стерильных почв после внесения в них аммония и инокулирования культурами Nitrosomonas, Nitrosospira и Nitrosolobus. В одинаковых условиях наибольшее количество N2O было получено из почв, инокулированных двумя штаммами Nitrosomonas, и значительно меньшее в случае Nitrosospira и двух штаммов Nitrosolobus. Однако судить о родовых различиях в способности образовывать N2O нельзя, так как третья из исследованных бактерий - Nitrosomonas sp.- образовывала ничтожные количества N2O.
В совместных работах лабораторий Ватсона и Мак-Элроя (1980-1981) показана способность к образованию N2O у представителей 4 родов хемоавтотрофных нитрификаторов - Nitrosomonas europaea, Nitrosococcus oceanus, Nitrosolobus multiformis и Nitrosospira briensis. В условиях низкой аэрации скорость образования NO2- жидкой культурой Nitrosomonas sp. значительно уменьшается, а выделение N2O увеличивается. Обнаружено, что суспензии клеток N.europaea наряду с NO2- и N2O образуют NO. С уменьшением концентрации О2 увеличивался выход обоих газов и росло отношение NO/N2O, составляя в среднем 1,5 моля NO - N на моль N2O - N. Следует отметить, что у всех исследованных организмов азот N2O составлял не более 0,5% от суммы окисленного азота, что более чем на порядок ниже продукции N2O у Nitrosomonas europaea, наблюдавшейся Иошида и Александером (1970) [4].
Роль нитрификаторов в выветривании горных пород, в повреждении памятников и облицовки сточных каналов позволяет прогнозировать новый период в исследованиях этой группы бактерии.
2.2 Краткая характеристика хемолитотрофных бактерий
Хемолитотрофия – способ существования, обнаруженный только у прокариот, при котором источником энергии служат реакции окисления неорганических соединений.
Существование хемоавтотрофов было открыто С.Н. Виноградским. Началом послужили его работы (1885—1889) по изучению нитчатых микроорганизмов, называемых серобактериями (Beggiatoa) и железобактериями (Leptothrix ochracea). В результате проведенных наблюдений Виноградский пришел к выводу, что жизнедеятельность указанных форм связана с окислением соответственно сероводорода и серы до серной кислоты или закисного железа в окисное и оба процесса имеют энергетическое значение [3].
Свою гипотезу Виноградский блестяще доказал, выделив (1890—-1892) чистые культуры нитрифицирующих бактерий (Nitrosomonas и Nitrobacter), которые росли на минеральных средах, окисляя аммонийный азот или нитриты и фиксируя при этом углекислоту [3].
Хемолитотрофы могут существовать как в аэробных, так и анаэробных условиях и использовать довольно широкий круг неорганических соединений в качестве источников энергии. На основании специфичности хемолитотрофов в отношении субстратов их можно разделить на пять основных групп:
• нитрифицирующие бактерии окисляют восстановленные неорганические соединения азота;
• бактерии, окисляющие серу, используют в качестве источника энергии H2S, молекулярную серу (S0) или ее частично восстановленные окислы;
• железобактерии окисляют восстановленное железо или марганец;
• водородные бактерии используют в качестве источника энергии молекулярный водород;
• у карбоксидобактерий единственный источник углерода и энергии – СО2.
К хемолитотрофным можно отнести сульфатвосстаналивающие и метанобразующие бактерии. Сульфатвосстанавливающие бактерии получают энергию окислением в анаэробных условиях молекулярного водорода, используя в качестве конечного акцептора электронов сульфат (SO). Метаногенные бактерии используют CO2 в качестве конечного акцептора электронов при окислении молекулярного водорода.
Кроме того, к хемоавтотрофам, видимо, принадлежит недавно обнаруженный Н. Н. Ляликовой-Медведевой микроорганизм Stibiobacter, окисляющий окислы трехвалентной сурьмы (Sb2O3) до пятивалентной (Sb2О5) [11].
Все представители грамотрицательные. Морфологически разнообразны: палочки, кокки, спириллы. Подвижны и неподвижные. Аэробы. Группа разделена на 3 подгруппы в зависимости от химической природы окисляемых неорганических соединений.
Таким образом, выявлены хемоавтотрофы, способные получать энергию в результате окисления минеральных соединений пяти элементов: Н, N, S, Fe и Sb [16].
ГЛАВА 3 ХАРАКТЕРИСТИКА БАКТЕРИЙ
СЕМЕЙСТВА NITROBACTERIACEAE
3.1 Особенности строения и функционирования
Процесс последовательного окисления аммиака до азотистой и азотной кислот называется нитрификацией, а вызывающие его бактерии — нитрифицирующими. Сущность этого процесса была раскрыта и изучена С. Н. Виноградским [3].
Выделяют две группы нитрозных и нитратных бактерий. Первую группу нитрозных бактерий составляют роды Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosolobus, Nitrosovibrio, вторую — нитратных бактерий — Nitrobacter, Nitrospina, Nitrococcus.
Группа нитрифицирующих прокариот представлена грамотрицательными организмами, различающимися формой и размером клеток, способами размножения, типом жгутикования подвижных форм, особенностями клеточной структуры, молярным содержанием ГЦ-оснований ДНК, способами существования. При этом они имеют ряд общих свойств. Все организмы, помещенные в семейство Nitrobacteraceae, не образуют спор, красятся по Грамму отрицательно, облигатные аэробы и не нуждаются в органических факторах роста, оптимум развития лежит при температуре 25-30° и рН 7,5-8,0. Представители различных родов нитрифицирующих бактерий различаются морфологически [27]. Клетки палочковидные, эллипсоидальные, сферические, спиральные и дольчатые клетки без эндоспор. Жгутики субполярные или перитрихальные, часто отсутствуют. Клетки получают энергию от окисления аммиака и нитрита и удовлетворяют свои потребности в углероде путем фиксации CO2. Только для некоторых штаммов Nitrobacter показана способность к медленному росту в среде с органическими соединениями в качестве источника углерода и энергии. Не паразитические. Обычно встречаются в почве, пресной воде и морской воде. Клетки богаты цитохромами, но никаких других пигментов обнаружено не было [24].
Нитрифицирующие бактерии относятся к типичным хемосинтезирующим автотрофам; они очень чувствительны к наличию в среде органических соединений. Растут на простых минеральных средах, содержащих окисляемый субстрат в виде аммония или нитритов и углекислоту. Источником азота в конструктивных процессах могут быть, кроме аммония, гидроксиламин и нитриты [3].
Таблица 1.
Описание типовых культур нитрифицирующих бактерий (Горленко, 1977).
|
Бактерии |
Форма клетки |
Размер, мкм |
Способ деления |
Тип внутренних цитомембран |
Жгутикова-ние |
Окис-ление NO2-- NO3- |
Окис-ление NH4+- NO2- |
|
Nitrosolobus multiformis |
Лопаст-ная |
1,0*1,5 |
Дроб-ление |
В виде перегородок |
Пери-трихи-альн |
- |
+ |
|
Nitrosococcus oceanus |
Кокк |
1,8-2,2 |
Деле-ние |
Пластинчатый |
» |
- |
+ |
|
Nitrosococcus mobilis |
» |
1,5-1,8 |
» |
То же + тубулярные |
2 субпо-ляр-ных |
+ |
- |
|
Nitrospina gracilis |
Тонкая палочка |
0,3-0,4 *2,7-6,5 |
» |
Отсутствуют |
Нет |
+ |
- |
|
Nitrosovibrio tenuis |
Вибрион |
0,3-0,4 *1,1-3,0 |
» |
Отсутствуют или тубулярные |
1-4 субпо-ляр-ных |
_ |
+ |
|
Nitrobacter winogradskii |
Груше-видная |
0,6-0,8 *1,0-2,0 |
Почкование |
Перифери- ческие-Пластин- чатые |
1 |
+ |
- |
|
Nitrosomonas europeus |
Палочка |
0,8-0,9 *1,0-2,0 |
Деле-ние |
То же |
1-2 |
- |
+ |
|
Nitrosospira briensis |
Спи-ральная |
0,2-0,4 *1,0-5,0 |
» |
Отсутствуют |
Перит рих |
- |
+ |
По строению клеток исследованные нитрифицирующие бактерии похожи, на другие грамотрицательные микроорганизмы. У некоторых видов обнаружены развитые системы внутренних мембран, которые образуют стопку в центре клетки (Nitrosocystis oceanus), или располагаются по периферии параллельно цитоплазматической мембране (Nitrosomonas europaea), или образуют чашеподобную структуру из нескольких слоев (Nitrobacter winogradskyi). Видимо, с этими образованиями связаны ферменты, участвующие в окислении нитрификаторами специфических субстратов [13].
3.2 Бактерии первой фазы нитрификации
Бактерии первой фазы нитрификации представлены четырьмя родами: Nitrosomonas, Nitrosocystis, Nitrosolobus и Nitrosospira.
Род Nitrosomonas. Клетки эллипсоидальные или короткие палочки, подвижные или неподвижные, одиночные, в парах или коротких цепочках. Грамотрицательные. Обладают цитомембранами, образующими уплощенные пузырьки в периферических областях цитоплазмы. Клетки растут свободно в среде или заключены в слизистую зооглею. Богаты цитохромами, которые придают клеточным взвесям окраску от желтоватой до красноватой; других пигментов нет.
Облигатные хемолитотрофы; окисляют аммиак до нитрита и фиксируют СО2 для получения энергии и углерода. Растут в пресной или морской воде, обогащенной аммиаком и неорганическими солями, не требуют органических факторов роста. Строгие аэробы. Температурные границы для роста 5-30 C, пределы pH 5,8-8,5. Обитают в почвах, пресной и морской воде [24].
ФОТО 1 Stan Watson,
Woods Hole Oceanographic Institute
Nitrosomonas euroраеа
Наиболее изучен вид Nitrosomonas euroраеа, хотя получение чистых культур этих микроорганизмов, как и других нитрифицирующих хемоавтотрофов, до сих пор остается достаточно сложным. Клетки N.europaea обычно овальные (0,6-1,0) (0,9-2,0) мкм, размножаются бинарным делением. В процессе развития культур в жидкой среде наблюдаются подвижные формы, имеющие один или несколько жгутиков, и неподвижные зооглеи [9].
Среди N. europaea известны штаммы, имеющие температурный оптимум при 26 или около 40° C, и штаммы, довольно быстро растущие при 4° C.
Род Nitrosococcus. Клетки от сферических до эллипсоидных. Подвижные или неподвижные. Клеточная стенка типичная для грамотрицательных бактерий, но у морских видов обнаружен дополнительный наружный слой клеточной стенки. Одиночные, в парах или в тетрадах, свободно взвешенных в жидкой среде, или заключены в слизь с образованием агрегатов, которые либо прикреплены к стенкам сосуда, либо находятся во взвешенном состоянии в жидкой среде [21].
Облигатные хемолитотрофы; окисляют аммиак до нитрита и фиксируют СО2 для удовлетворения потребностей в энергии и углероде. За исключением типового вида, выделенного из почвы, виды Nitrosococcus облигатно галофильные и их распространение ограничено морскими и солоноватоводными средами. Растут в пресной или морской воде, обогащенной аммиаком и неорганическими солями; не нуждаются в органических факторах роста. Строгие аэробы. Температурные границы для роста 2-30 C, пределы pH 6,0-8,0. Типовой вид: Nitrosococcus nitrosus [21].
Род Nitrosospira. Клетки в виде туго скрученных спиралей шириной 0,3-0,4 мкм, с 3-20 витками. Грамотрицательные. Развитой системой внутрицитоплазматических мембран не обладают, но часто наблюдаются инвагинации цитоплазматической мембраны в протоплазму. Неподвижные или движутся с помощью перитрихальных жгутиков. Растут свободно в жидкой среде. Клетки богаты цитохромами, которые придают суспензии окраску от желтой до красноватой; других пигментов нет. Облигатные хемолитотрофы; окисляют аммиак до нитрита и фиксируют СО2 для удовлетворения потребностей в энергии и углероде. Растут в пресной воде, обогащенной аммиаком и неорганическими солями; или почве. Строгие аэробы, использующие кислород в качестве конечного акцептора электронов. Температурные границы для роста 15-30 C, пределы pH 6,5-8,5. Выделены из почв Бри (Франция), острова Крит, вершины горы Пилат (Швейцария) и храма Парфенон в Афинах.
С.Н.Виноградский разделил на два вида: Nitrosospira briensis и Nitrosospira arctica. Наиболее изученный предствитель Nitrosospira briensis клетки палочковидные и извитые (0,8-1,0) (1,5-2,5) мкм, имеют от одного до шести жгутиков [21].
Род Nitrosolobus. Клетки плеоморфные и лопастные, 1,0-1,5 мкм в поперечнике; делятся перетяжкой. Грамотрицательные. Клетки внутри частично подразделены впячиваниями плазматической мембраны и других частей клеточной оболочки в цитоплазму с образованием везикулярных участков. Подвижные, движутся с помощью перитрихальных жгутиков. Растут в свободном состоянии в жидкой среде. Клетки богаты цитохромами, которые придают клеточной суспензии окраску от желтоватой до красноватой; других пигментов нет. Облигатные хемолитотрофы; окисляют аммиак до нитрита и фиксируют СО2 для удовлетворения потребностей в энергии и углероде. Выделены только из почв. Строгие аэробы. Температурные пределы для роста 15-30 C, пределы pH 6,0-8,2. Найдены в почвах Суринама (Южная Америка), Галапагосских островов, юго-восточной Африки, странах СНГ [9,12,13].
Клетки вида Nitrosolobus multiformis составляют 1,0-1,5 1,0-2,5 мкм. Форма этих бактерий не совсем правильная, так как клетки разделены на отсеки, дольки (-lobus, отсюда и название Nitrosolobus), которые образуются в результате разрастания внутрь цитоплазматической мембраны.
Род Nitrosovibrio. Изогнутые тонкие палочки. В начале стационарной фазы роста наблюдаются сферические формы. Подвижные клетки обладают 1-4 жгутиками, полярными и субполярными. Развитые внутрицитоплазматические мембраны отсутствуют, однако наблюдаются инвагинации мембраны в протоплазму. Все изоляты Nitrosovibrio выделены из почв или камней зданий.
Род Nitrococystis. Отличается тем, что бактерии этого рода способны образовывать зооглеи - кокковидные формы микробов, окруженные общей капсулой [21].
Такой микроорганизм, как Nitrosocystis oceanus, выделенный из Атлантического океана, относится к облигатным галофилам и растет на среде, содержащей морскую воду. Имеет округлые клетки, диаметром 1,8-2,2 мкм, но бывают и крупнее (до 10 мкм). Способен к движению благодаря наличию одного жгутика или пучка жгутиков. Образует зооглеи и цисты [9].
2.3.3 Бактерии второй фазы нитрификации
Среди бактерий второй фазы нитрификации различают три рода: Nitrobacter, Nitrospina и Nitrococcus.
Род Nitrobacter. Клетки - короткие палочки, часто клиновидные или грушевидные. Как показано Г. А. Заварзиным, размножение Nitrobacter происходит путем почкования, причем дочерняя клетка бывает обычно подвижна, так как снабжена одним латерально расположенным жгутиком. Клетки обладают полярной шапкой цитомембран. Обычно неподвижные. Стадии покоя неизвестны. Грамотрицательные. Клетки богаты цитохромами, придающими клеточным суспензиям желтоватый цвет; других пигментов нет. Содержат внутриплазматические мембраны в виде полярных уплощенных везикул в периферической части протоплазмы. По данным электронной микроскопии тонких срезов клеточная стенка отличатся от таковой большинства грамотрицательных бактерий. Она состоит из трех слоев (электроноплотный, электронопрозрачный, электроноплотный), причем внутренний слой более электроноплотный, чем наружный [18].
Отмечают также сходство Nitrobacter с почкующимися бактериями рода Hyphomicrobium по составу жирных кислот, входящих в липиды.
Некоторые штаммы - облигатные хемолитотрофы, они окисляют нитриты до нитратов и фиксируют СО2 для удовлетворения потребностей в энергии и углероде. Строгие аэробы, использующие кислород в качестве конечного акцептора электронов. Пределы pH для роста 6,5-8,5; температурные пределы 5-40 C. Место обитания: почвы, пресная вода, морская вода [14].
Род Nitrospira. Клетки от слабо закрученных спиральных до вибриоидных, шириной 0,3-0,4 мкм, с круглой амплитудой 0,8-1,0 мкм. Подвижности не наблюдается. Внутрицитоплазматические мембраны отсутствуют. Клетки содержат цитохромы, других пигментов нет. Периплазматическое пространство необычно широко, в два раза шире, чем у других грамотрицательных бактерий [9,21].
Облигатные хемолитотрофы окисляют нитриты до нитратов и фиксируют СО2 для получения энергии и углерода. Растут в морской воде, обогащенной нитритами и неорганическими солями; в органических факторах роста не нуждаются. Также, как и род Nitrobacter, являются строгими аэробами. Температурные пределы для роста 20-30 C, пределы pH 7,8-8,0.. Единственным источником энергии служит нитрит, но рост в миксотрофных условиях лучше, чем в литоавтотрофных. Для роста необходима морская вода. Оптимальная среда, содержащая 70-100% морскую воду плюс нитрит, пируват, глицерол, а также дрожжевой экстракт или пептон. Не растут в среде из дистиллированной воды с минеральными солями, даже при включении NaCl. Выделены из различных морских местообитаний, таких как океанические воды и морские осадки. Обитают в южной части Атлантического океана. В единственном случае описано выделение накопительной культуры из проб почвы, отобранных в Намибии. Типовой (и единственный) вид: Nitrospira marina [21].
Род Nitrococcus. Клетки сферические, 1,5 мкм или больше. Грамотрицательные. Движутся при помощи одного или двух субтерминально прикрепленных жгутиков. Клетки богаты цитохромами, придающими взвесям желтовато- красноватую окраску; других пигментов нет. Облигатные хемолитотрофы; окисляют нитриты до нитратов и фиксируют СО2 для получения энергии и углерода. Внутрицитоплазматические мембраны в виде трубочек расположены по всей цитоплазме. Растут в морской воде, обогащенной нитритами и неорганическими солями; в органических факторах роста не нуждаются, являются строгими аэробами.
Температурные пределы для роста 15-30 C, пределы pH 6,8-8,0. Обитают в южной части Тихого океана.
Представитель Nitrococcus. mobilis обладает подвижностью. Клетки его округлые, диаметром около 1,5 мкм, с одним-двумя жгутиками [21].
ГЛАВА 4 ОПИСАНИЕ ПРОЦЕССА НИТРИФИКАЦИИ
4.1 Химизм процесса ниртификации
Процесс нитрификации представляет собой яркий пример метабиоза, когда одни микроорганизмы начинают развиваться после других на продуктах жизнедеятельности первых.
Процесс нитрификации локализован на цитоплазматической и внутрицитоплазматических мембранах. Ему предшествует поглощение NH4+ и перенос его через цитоплазматическую мембрану с помощью медьсодержащей транслоказы. При окислении аммиака до нитрита атом азота теряет шесть электронов. Предполагается, что на первом этапе аммиак окисляется до гидроксиламина с помощью монооксигеназы, катализирующей присоединение к молекуле аммиака одного атома О2; второй взаимодействует с НАД*Н2 , что приводит к образованию Н2О:
NH3 + О2 + НАД*Н2 NH2OH + Н2О + НАД+
Гидроксиламин далее ферментативно окисляется до нитрита:
NH2OH + О2 NO2- + Н2О + Н+
Электроны от NH2OH поступают в дыхательную цепь на уровне цитохрома «с» и далее на терминальную оксидазу. Их транспорт сопровождается переносом двух протонов через мембрану, приводящим к созданию протонного градиента и синтезу АТФ. Гидроксиламин в этой реакции, вероятно, остается связанным с ферментом [25].
Вторая фаза нитрификации сопровождается потерей двух электронов. Окисление нитрита до нитрата, катализируемое молибденсодержащим ферментом нитритоксидазой, локализовано на внутренней стороне цитоплазматической мембраны и происходит следующим образом:
NO2- + Н2О NO3- + 2Н+ 2е
Электроны поступают на цитохром «а1» и через цитохром «с» на терминальную оксидазу «аа3», где акцептируются молекулярным кислородом. При этом происходит перенос через мембрану 2Н+. Поток электронов от NO2- на О2 происходит с участием очень короткого отрезка дыхательной цепи. Так как Ео пары NO2/NO3- равен +420 мВ, восстановитель образуется в процессе энергозависимого обратного переноса электронов. Большая нагрузка на конечный участок дыхательной цепи объясняет высокое содержание цитохромов «с» и «а» у нитрифицирующих бактерий [27].
Схематически вторую фазу нитрификации можно представить следующим образом:

Таким образом, суммарно процесс окисления аммиака можно представить в следующем виде:
NH3NH2OH[HNO]NO2NO3
Таблица 2 Нитрифицирующие бактерии

Энергетически выгодными являются только стадии окисления гидроксиламина в нитрит и нитрита в нитрат, так как в результате происходит образование молекул АТФ.
Поскольку субстраты (аммиак, нитриты), которые окисляют нитрифицирующие бактерии, обладают сильно положительным окислительно- восстановительным потенциалом, который для пары NH4+/ NH2OH составляет +899 мВ, для пары NO2-/ NO3– - +420 мВ, а окислительно-восстановительный потенциал НАД / НАД · Н2 имеет отрицательную величину -320 мВ, то окисление NH4- или NO2- по термодинамическим причинам не может быть прямо связано с восстановлением НАД. Образование НАД · Н2 в таком случае происходит за счет функционирования обратного транспорта электронов, который имеется у нитрифицирующих бактерий наряду с прямым транспортом по дыхательной цепи. Обратный транспорт электронов сопровождается затратой энергии [20].
Схематически перенос электронов у нитрифицирующих бактерий можно представить следующим образом:

Многие хемоорганогетеротрофные бактерии, принадлежащие к родам Arthrobacter , Flavobacterium , Xanthomonas , Pseudomonas и др., способны окислять аммиак, гидроксиламин и другие восстановленные соединения азота до нитритов или нитратов. Процесс нитрификации этих организмов, однако, не приводит к получению ими энергии. Изучение природы этого процесса, получившего название гетеротрофной нитрификации, показало, что, возможно, он связан с разрушением образуемой бактериальными культурами перекиси водорода с помощью пероксидазы. Образующийся при этом активный кислород окисляет NH3 до NO3- [27].
4.2 Условия, влияющие на процесс нитрификации
В первых работах с нитрификаторами Виноградский отметил, что для их роста неблагоприятно присутствие в среде органических веществ, таких, как пептон, глюкоза, мочевина, глицерин и др. Отрицательное действие органических веществ на хемоавтотрофные нитрифицирующие бактерии неоднократно отмечалось и в дальнейшем. Сложилось даже мнение, что эти микроорганизмы вообще не способны использовать экзогенные органические соединения. Поэтому их стали называть «облигатными автотрофами». Однако в последнее время показано, что использовать некоторые органические соединения эти бактерии способны, но возможности их ограничены. Так, отмечено стимулирующее действие на рост Nitrobacter в присутствии нитрита дрожжевого автолизата, пиридоксина, глутамата и серина, если они в низкой концентрации вносятся в среду. Показано также включение в белки и другие компоненты клеток Nitrobacter 14С из пирувата, а-кетоглутарата, глутамата и аспартата. Известно, кроме того, что Nitrobacter медленно, но окисляет формиат. Включение 14С из ацетата, пирувата, сукцината и некоторых аминокислот, преимущественно в белковую фракцию, обнаружено при добавлении этих субстратов к суспензиям клеток Nitrosomonas europaea. Ограниченная ассимиляция глюкозы, пирувата, глутамата и аланина установлена для Nitrosocystis oceanus. Есть данные об использовании 14С-ацетата Nitrosolobus multiformis [17].
Недавно установлено также, что некоторые штаммы Nitrobacter растут на среде с ацетатом и дрожжевым автолизатом не только в присутствии, но и в отсутствие нитрита, хотя и медленно. При наличии нитрита окисление ацетата подавляется, но включение его углерода в разные аминокислоты, белок и другие компоненты клеток увеличивается. Имеются, наконец, данные, что возможен рост Nitrosomonas и Nitrobacter на среде с глюкозой в анализируемых условиях, которые обеспечивают удаление продуктов ее метаболизма, оказывающих ингибиторное действие на данные микроорганизмы. На основании этого делается вывод о способности нитрифицирующих бактерий переключаться на гетеротрофный образ жизни. Однако для окончательных выводов необходимо большее число экспериментов. Важно прежде всего выяснить, как долго нитрифицирующие бакте рии могут расти в гетеротрофных условиях при отсутствии специфических окисляемых субстратов.
Некоторые из условий, влияющих на рост нитрифицирующих бактерий, например необходимость углекислых солей кальция или магния, подавляющее действие многих органических соединений и солей аммония, были отмечены уже Виноградским и его школой. Впоследствии эти данные пополнил Мейергоф, подробно изучавший оба типа нитрифицирующих бактерий. Он считал, что влияние углекислых солей зависит от их буферного действия, и показал необходимость, установив чрезвычайную чувствительность этих микроорганизмов к изменениям концентрации водородных ионов [5].
На процесс нитрификации положительно сказывается присутствие кислорода. В обрабатываемых почвах процесс нитрификации протекает более интенсивно.
Нитрификация замедляется или прекращается в том случае, если почва является слишком холодной, слишком сухой или при загрязнении.
В загрязненных почвах бурно развиваются микроорганизмы, осуществляющие распад гнилостных продуктов, а нитрификация подавлена. При накоплении в почве аммиака, образующегося при распаде органических загрязнителей, начинается развитие нитрификаторов. Интенсивный процесс нитрификации говорит о завершении распада органических загрязнителей и активно идущем самоочищении почвы.
В почвах азотистая кислота не накапливается, поскольку и Nitrosomonas и Nitrobacter встречаются в одной среде, находятся в своеобразном симбиозе. Поскольку высокие концентрации аммиака в щелочных почвах оказывают на Nitrobacter токсическое действие, Nitrosomonas, используя аммиак и образуя кислоту (т. е. переводя катион в анион), тем самым улучшает и условия существования для Nitrobacter.
Нитрификаторы чувствительны к кислой среде. Для окисления аммиака оптимальное течение рН лежит в пределах 8,5-8,7, а для окисления нитрита - в пределах 8,3-9,3. Для 0,2М раствора двууглекислого натрия рН равно 9,15; поэтому очевидно, что одновременное присутствие углекислого натрия и свободный доступ углекислоты обеспечивают поддержание кислотности, наиболее благоприятствующей развитию этих бактерий. С точки зрения почвенного режима важно отметить, что микроорганизмы, выделенные из торфяников (рН 4,6), оказываются более устойчивыми к кислотам и продолжают нитрифицировать до тех пор, пока рН не упадет ниже 4,1 [26].
Высокая оптимальная щелочность и быстрое падение активности по обе стороны от этого оптимума заставляют предположить, что как в кислой, так и в щелочной области начинают действовать вторичные подавляющие факторы. Согласно предположению Мейергофа, быстрое падение в щелочной области, вероятно объясняется проникновением в клетку свободного аммиака [5].
Как и многие другие автотрофные бактерии, нитрифицирующие микроорганизмы строго специфичны в отношении окисляемого ими материала: кроме того, что Nitrobacter не оказывает никакого действия на аммиак, а Nitrosomonas - на нитрит, ни тот, ни другой не могут использовать в качестве источника энергии ни сульфиты, ни фосфиты, ни какие-либо углеродные соединения [33].
В результате жизнедеятельности нитрифицирующих бактерий на 1 га почвы может накапливаться за год до 300 кг азотной кислоты [34].
4.3 Влияние адсорбентов на процесс нитрификации
Давно замечено, что нитрификаторы при культивировании в жидких средах в большом количестве располагаются на поверхности частиц мела, песка, окиси алюминия, талька и других адсорбентов, вносимых в среду. Культивирование нитрификаторов в средах без адсорбентов обычно оказывается затруднительным. При очистке питательной среды от твердых взвешенных частиц скорость нитрификации уменьшается. В присутствии адсорбентов нитрификаторы лучше сохраняются как в жидких средах, так и в высушенном состоянии.
Еще Альбрехт и Мак-Кала установили, что адсорбированный почвами и глинистыми минералами, аммоний оказывается вполне доступным для микроорганизмов [26].
Изучение процесса нитрификации в перфузионном аппарате показало, что в почве этот процесс протекает на поверхности твердой фазы, где находится адсорбированный аммоний: бактерии также располагаются на поверхности твердых частиц. Скорость нитрификации возрастает с увеличением количества поглощенного аммония и почти не меняется с изменением его концентрации в растворе. Добавление к почве веществ, обладающих большой способностью адсорбировать катионы, в том числе и ион аммония, увеличивает скорость нитрификации. Добавление твердых веществ, не обладающих катионо-обменной способностью, не влияет на скорость процесса. Если из перфузионного аппарата после начала интенсивной нитрификации убрать почву, процесс почти полностью прекращается, в то время как в тех сосудах, где почва сохранилась, нитрификация идет очень интенсивно. Эти опыты подтверждают положение о том, что нитрификация происходит на поверхности почвенных частиц. Подобные данные были получены и другими исследователями. Установлено, что нитрификаторы покрывают поверхность прерывистым или сплошным монослоем, но не дают многослойной адгезии.
Сейферт показал, что интенсивность нитрификации была наибольшей в почвенной фракции с частицами размером меньше 1 мм и сокращалась с увеличением размеров частиц, что коррелирует с количеством адсорбируемого этими фракциями аммония [2].
Улучшению прикрепления клеток нитрификаторов к частицам может способствовать наличие фимбрий.
Некоторые исследователи подробно изучали действие на процесс нитрификации добавления к культурной среде глинистых минералов, которые насыщались различными катионами. Они устанавливали довольно сложные закономерности влияния минералов на нитрификацию. Часть глинистых минералов может фиксировать аммоний. Он не обменивается или с трудом обменивается на другие катионы. Фиксации аммония способствует добавление калия. Аллисон и др. показали, что ион аммония фиксируется на глинистых минералах и, по крайней мере частично, недоступен нитрификаторам. Вельч и Скот изучали нитрификацию на вермикулите, иллите, бентоните, насыщенных аммонием. По их данным, интенсивность нитрификации от добавления больших количеств калия сильно сокращается. Это объясняется тем, что калий блокирует освобождение аммония [9,13].
Гольдберг и др. установили, что добавление минералов в среду для культивирования нитрификаторов может повышать и понижать скорость процесса, а может и не оказывать на нее заметного влияния. Введение в среду минералов, обладающих большой емкостью поглощения, стимулировало нитрификацию. Однако введение мусковита и каолинита, обладающих небольшой емкостью поглощения, не стимулировало процесс и даже могло замедлить его. Внесение в почву монтмориллонита повышало скорость минерализации глицина и нитрификации образующегося при этом аммония [14].
В некоторых случаях отмечено значительное угнетение нитрификации адсорбентами (почвами, ионообменными смолами, илом), что может быть отнесено за счет недостаточной аэрации среды, а также токсического действия некоторых адсорбентов.
В опытах Шревена обменный аммоний был вполне доступен для нитрификаторов, а фиксированный почвами аммоний почти не использовался ими. Обменный аммоний идентифицировался в течение 2 недель, а фиксированный — сохранялся без изменений в течение 16 недель. Внесение калия делало фиксированный аммоний особенно мало доступным. Аналогичные данные были получены Акслей и Лег [23,30].
Кунц и Стотцкий показали, что внесение монтмориллонита повышает скорость минерализации глицина в почве и нитрификации образующегося при этом аммония [18].
Природа глинистых минералов оказывает влияние на нитрификацию. Так, при значении рН жидкой среды, равном 5,5, на аллофане нитрификация осуществлялась, а на каолините и бентоните не проявлялась.
Таким образом, адсорбированные клетки нитрификаторов могут успешно развиваться и использовать обменный аммоний. Адгезированное состояние, видимо, является естественным для этих бактерий при их развитии в почве. Обычно введение адсорбентов благоприятствует нитрификации, но в некоторых случаях может оказывать и отрицательное действие. Причина различного действия адсорбентов определяется рядом специфических условий, которые возникают в среде при внесении того или иного адсорбента и сильно зависят от природы катионов, насыщающих адсорбент, К сожалению, исследователи, изучающие влияние адсорбентов на процесс нитрификации, часто не учитывали степень адсорбции клеток, рН среды, степень аэрации и другие факторы. Вероятно, положительное действие адсорбентов проявлялось в тех случаях, когда клетки адсорбировались на поверхности частиц и оказывались вблизи от адсорбированного обменного иона аммония, причем рН и другие показатели этой микрозоны были благоприятными для развития бактерий, осуществляющих первую фазу нитрификации. Если аммоний находился в адсорбированном состоянии, а клетки — в свободном, то внесение адсорбента не оказывало положительного действия. Безусловно, фиксированный аммоний слабо доступен для микроорганизмов.
Активность бактерий, осуществляющих вторую фазу нитрификации и использующих не адсорбирующиеся почвенными частицами и глинистыми минералами ионы NО2, при адсорбции уменьшается. Причем на их развитие большое влияние оказывает спедифическое рН, создающееся на границе раздела твердой и жидкой фаз. Если судить о рН по его значению в жидкой фазе, то оптимум развития оказывается смещенным в щелочную сторону. Однако это явление кажущееся, так как клетки в действительности развиваются при более кислом рН, характерном для поверхности раздела жидкой и твердой фаз. Вероятно, рН на поверхности этих минералов было разным [5].
Довольно подробно в ряде работ изучена на колонках из почв и других адсорбентов кинетика процесса нитрификации, вызываемого адгезированными клетками. В колонках практически все бактерии находятся в адсорбированном состоянии. Обычно нитрифицирующие бактерии располагаются только в верхней части колонки. В силу своих адгезионных способностей клетки не образуют на поверхности адсорбента нескольких слоев и располагаются только сплошным монослоем. В нижнем слое колонки уже нет и сплошного монослоя, а обнаруживаются отдельные участки, занятые клетками, или единичные клетки. Монослойное расположение клеток дает основание для построения более простых математических моделей.
Ряд работ, выполненных в последнее время, посвящен изучению кинетики процесса нитрификации в ферментерах с инертной загрузкой или ферментерах с вращающимися дисковыми биофильтрами. Отмечается, что в таких условиях процесс идет более интенсивно, чем в аэротенках. Причем увеличение интенсивности процесса достигается именно путем увеличения адсорбирующей поверхности. Внутри прикрепленной биопленки реакция нитрификации описана с учетом процесса молекулярной диффузии [28].
ГЛАВА 5 НИТРИФИЦИРУЮЩИЕ ОРГАНИЗМЫ
В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА
5.1 Значение хелимолитоавтотрофов в круговороте веществ
Жизнь на Земле в значительной мере определяется биологическими процессами. Микроорганизмы обнаружены повсюду, где есть хотя бы какие-то источники энергии, углерода, азота. В основе разнообразия их метаболизма лежит способность использовать любые, даже самые минимальные возможности для своего существования. В то же время гигантская роль микроорганизмов в круговороте веществ в природе имеет исключительное значение для сохранения динамического равновесия всей биосферы, нарушение которого может привести к катастрофическим последствиям.
Под круговоротом веществ в природе понимают циклические превращения химических элементов, которые входят в состав живых существ. Эти превращения происходят вследствие разнообразия и гибкости метаболизма микроорганизмов. В окружающей среде, по-видимому, нет таких органических веществ, которые не разрушались бы теми или иными микроорганизмами. Круговороты химических элементов могут замыкаться в границах определенной экосистемы, то есть не включать обмен промежуточными продуктами цикла между экосистемой и окружающей средой. Но большинство экосистем – это открытые системы; между ними и окружающей средой происходит обмен химическими веществами. Такие экосистемы, как озеро, океан, участок почвы выступают как источники или как акцепторы химических соединений, обычно переносимых воздухом или током воды. Везде и всюду микроорганизмы сосуществуют в виде сложных ассоциаций – биоценозов, представленных многими и разнообразными видами, между которыми складываются своеобразные взаимоотношения. Особенности этих взаимоотношений таковы, что они обеспечивают существование всех многочисленных видов бактерий и в конечном счете всего царства прокариот, которое, в свою очередь, сосуществует с другими царствами жизни на Земле.
Учёные предполагают, что до появления живых организмов атмосферные газы на нашей планете находились в восстановленном состоянии: углерод – в форме метана (CH4), азот – в форме аммиака (NH3), кислород – в составе воды (H2O). Современное состояние этих химических элементов в виде окисленных форм – углерода в виде оксида (СО2), а азота и кислорода в форме простых газов (N2 и O2) – является следствием в основном активности микроорганизмов. Газовый состав атмосферы Земли и количественное содержание в ней химических элементов, необходимых для живых организмов, являются результатом равновесия между их образованием и использованием в биологических и геологических процессах. Автотрофные организмы, в том числе одноклеточные водоросли, и цианобактерии продуцируют органическое вещество из СО2 и других неорганических соединений, а также из воды как источника электронов и выделяют молекулярный кислород (процесс фотосинтеза). Это органическое вещество используется частично самими продуцентами при дыхании и частично животными и человеком (консументами), которые окисляют его до СО2 (потребление). Затем остатки растений и экскременты животных разлагаются микроорганизмами, в первую очередь бактериями и грибами (редуценты): в присутствии кислорода до СО2, в анаэробных условиях до СО2 и метана (биодеградация, минерализация). Азот, сера и фосфор, входящие в состав биомассы, высвобождаются в основном в форме аммиака, сероводорода и фосфата соответственно, затем претерпевают дальнейшие превращения. Следовательно, основная роль микроорганизмов в биосфере состоит в минерализации органического вещества, и большинство известных прокариот – гетеротрофы [30].
Другим важным фактором, определяющим роль микроорганизмов в природе, является высокая скорость их размножения при благоприятных условиях. Автотрофные прокариоты участвуют в первичной (цианобактерии) или вторичной продукции (большинство аноксигенных фототрофных бактерий). Они осуществляют фотосинтез органического вещества, но при этом используют в качестве доноров электронов (Н2S, H2, Fe2+) органические соединения, образование которых обычно связано с трансформацией органических веществ, синтезируемых первичными продуцентами. То же можно сказать и о хемолитотрофных микроорганизмах: они окисляют доноры электронов (например H2, NH3, H2S, Fe2+ и CO), в большинстве случаев восстановленные при окислении органического вещества, и зависят от восстановителей, действующих как промежуточные носители электронов в процессе разложения органического вещества. Поэтому некорректно считать, что экосистемы в зонах морских гидротермальных вод, которые полностью основаны на первичной продукции хемолитотрофных микроорганизмов, не зависят от фотосинтеза, так как хемосинтезирующие организмы используют в качестве акцептора электронов молекулярный кислород – продукт фотосинтеза, осуществляемого фитопланктоном в поверхностном слое морской воды, наземными растениями и цианобактериями [13].
Хемоавтотрофные нитрифицирующие бактерии имеют широкое распространение в природе и встречаются как в почве, так и в разных водоемах. Осуществляемые ими процессы могут происходить весьма в крупных масштабах и имеют существенное значение в круговороте азота в природе. Раньше считали, что деятельность нитрификаторов всегда способствует плодородию почвы, поскольку они переводят аммоний в нитраты, которые легко усваиваются растениями, а также повышают растворимость некоторых минералов. Сейчас, однако, взгляды на значение нитрификации несколько изменились. Во-первых, показано, что растения усваивают аммонийный азот и ионы аммония лучше удерживаются в почве, чем нитраты. Во-вторых, образование нитратов иногда приводит к нежелательному подкислению среды. В-третьих, нитраты могут восстанавливаться в результате денитрификации до N2, что приводит к обеднению почвы азотом.
Нитрифицирующие бактерии обнаружены в водоемах разного типа и в почвах, где они, как правило, развиваются совместно с бактериями, жизнедеятельность которых приводит к образованию исходного субстрата нитрификации - аммиака.
Нитрификаторы способны осуществлять хемосинтез, то есть создавать органическое вещество из углерода диоксида и воды за счет химической энергии окисления аммиака до азотистой кислоты и азотистой до азотной кислоты [30].
Процесс нитрификации, являясь важным звеном в круговороте азота в природе, имеет как положительные, так и отрицательные стороны. Переведение азота из аммонийной формы в нитратную способствует обеднению почвы азотом, поскольку нитраты легко вымываются из почвы. В то же время нитраты - хорошо используемый растениями источник азота. Связанное с нитрификацией подкисление почвы улучшает растворимость и, следовательно, доступность некоторых жизненно необходимых элементов, в первую очередь фосфора и железа.
5.2 Круговорот азота и роль в нем
нитрифицирующих бактерий
При самом активном, широком участии микроорганизмов в природе, главным образом в почве и гидросфере, постоянно осуществляется два противоположных процесса: синтез из минеральных веществ сложных органических соединений и, наоборот, разложение органических веществ до минеральных. Единство этих противоположных процессов лежит в основе биологической роли микроорганизмов в природе. Среди различных процессов превращения веществ, в которых микроорганизмы принимают активное участие, важнейшее значение для осуществления жизни растений, животных и человека на Земле имеют круговорот азота, углерода, фосфора, серы, железа. Важнейший элемент, входящий в состав белков, а, следовательно, имеющий исключительное значение для жизни — это азот.
Запасы азота в природе очень велики. Он входит в состав всех организмов на Земле. Общее содержание его в организмах составляет более 25 млрд т, большое количество азота находится также в почве. Но еще более грандиозен запас азота в атмосфере: над каждым гектаром почвы поднимается столб воздуха, содержащий около 80000т молекулярного азота. Ежегодно на образование вновь вырастающих растений требуется около 1,5 млрд т азота в форме, доступной для усвоения растениями [14].
Специфическая роль азота в биологических процессах обусловлена необычно большим числом степеней окисления, т.е. валентностей. Валентность – это свойство атома данного элемента присоединять или замещать определенное число атомов другого элемента. В организме животных и растений большая часть азота присутствует или в виде иона аммония, или в виде аминосоединений. В обеих формах азот сильно восстановлен: соединившись с тремя другими атомами, он принял от них три электрона, т. е. Имеет валентность –3. В другой сильно окисленной форме (нитрат-ион) пять внешних электронов атома азота участвуют в образовании связей с атомом кислорода, приобретая при этом валентность +5. Нитрат-ион – главная форма, в которой азот присутствует в почве. При переходе иона аммония или аминокислот в почвенные нитраты валентность азота должна меняться на 8 единиц, т. е. Атом теряет 8 электронов. И, наоборот, при переходе нитратного азота в азот аминогруппы атом приобретает 8 электронов.
В круговороте азота в природе с участием микроорганизмов различают четыре этапа:
- усвоение атмосферного азота,
- аммонификацию,
- нитрификацию,
- денитрификацию.
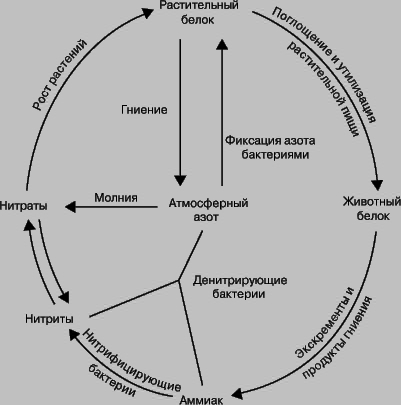
Рис. 1 Круговорот азота в природе
Среди процессов, от которых зависит биологическая продуктивность на земном шаре, одним из важнейших является фиксация микроорганизмами азота атмосферы. Ни человек, ни животные, ни растения не могут потреблять молекулярный азот, которым изобилует воздушный океан. Однако растениям нужен азот минеральных соединений, и, «купаясь» в молекулярном азоте, они могут испытывать «азотный голод».
Проблема биологической азотфиксации относится к числу основных проблем сельскохозяйственной и биологической науки. Перед учеными стоит задача изыскать возможности управления процессом азотфиксации и на этой основе увеличить урожайность сельскохозяйственных культур. Биологический азот может служить существенным дополнением азотного фонда почвы, способствуя повышению ее плодородия и обеспечивая тем самым более экономное расходование технического азота — азота удобрений [10].
Существуют две группы фиксирующих атмосферный азот микроорганизмов. Одна из них находится в симбиозе с высшими растениями, образуя клубеньки на корнях. К этой группе относятся клубеньковые бактерии. Микроорганизмы другой группы обитают в почве независимо от растений. К ним относятся азотобактер, клостридиум, бейеринкия и другие свободно-живущие микроорганизмы. Потенциальные возможности симбиотических азотфиксаторов значительно выше, чем свободноживущих [15].
Азотфиксирующие бактерии имеют ферменты, обладающие способностью связывать свободный азот с другими химическими элементами. Эти микроорганизмы синтезируют сложные органические соединения, обогащая, таким образом, почву связанным азотом и способствуют ее плодородию.
Имеющегося в воздухе и почве азота хватило бы для обеспечения урожая, даже при одностороннем использовании, на несколько миллионов лет. Однако растения часто дают низкие урожаи именно из-за недостатка азота в почве. Это объясняется тем, что только небольшая группа азотистых соединений может быть быстро усвоена растениями. Не только свободный азот, но и многие формы связанного азота не могут служить источником азотного питания для растений. Азот, поступающий в виде белковых веществ в почву вместе с остатками растений и животных, совсем не годится для этих целей, он должен быть подвергнут минерализации, а образующийся при этом аммиак должен быть окислен в соли азотистой и азотной кислот [12].
Микробиологический процесс, при котором под воздействием гнилостных микроорганизмов происходит гидролитическое расщепление белков, поступающих в почву с трупами животных и отмирающими растениями называется аммонификация белков или гниение, с образованием промежуточных продуктов (альбумоз, пептонов, амино– и амидокислот), а также дурнопахнущих веществ – индола, сероводорода, меркаптана, летучих жирных кислот. Конечным продуктом гидролиза белков и дезаминирования аминокислот является NН3, поэтому этот процесс и называется аммонификацией белка. Таким образом, при гниении происходит минерализация белковых веществ, которая в зависимости от химического состава белков субстрата, вида гнилостных бактерий и условий их жизнедеятельности может быть полной или не доведенной до конца. При полной мин реализации белка образуются Н2О, СО2, NH3, H2S и минеральные соли. Гниение – преимущественно анаэробный процесс. Гнилостные микробы широко распространены в почве, воде, воздухе, в животных и растительных организмах. Поэтому любой продукт, не защищенный от них, быстро подвергается гниению. Его вызывают как анаэробные, так и аэробные микроорганизмы, причем они могут действовать и преемственно, и одновременно.
Наиболее энергичными возбудителями гниения, сопровождающегося глубоким распадом белка и образованием азотистых и безазотистых соединений (индола, скатола, жирных кислот, NH3, H2, H2S и др.), являются Bacillus mycoides, B.subtilis, B.mesentericus, бактерии семейства Enterobacteriaceae (Proteus, Escherichia и др.), а также Clostridium putrificum, C.sporogenes. Последние два – анаэробы, содержатся в кишечнике и после смерти вызывают зловонное разложение трупов [15].
Процессы гниения протекают только при наличии условий, благоприятных для жизнедеятельности их возбудителей (влажность, температура и т. п.). В сухой песчаной почве трупы подвергаются мумификации (высушиванию без гниения). Гнилостные процессы происходят и в организме человека, в частности в кишечнике; причиной их являются E.coli и другие микробы. По мнению И. И. Мечникова, продукты гниения (скатол, индол и др.), постоянно образующиеся в организме, вызывают хроническую интоксикацию и являются одной из причин преждевременного старения [30].
Гнилостные процессы протекают также при газовой гангрене: ткани, омертвевшие под влиянием образуемых возбудителями этой болезни экзотоксинов, заселяются гнилостными аэробными и анаэробными бактериями и подвергаются распаду. Некоторые гнилостные процессы используются в промышленности с полезной целью, например при выработке кожи для отделения от нее шерсти – швицевании. Исключительное значение процессов гниения заключается в том, что они играют важную роль в естественном самоочищении почвы и воды. Этим пользуются для строительства специальных очистных сооружений (полей ассенизации, орошения и т. п.), для биологической переработки и обезвреживания фекальных нечистот и сточных вод, содержащих много мертвых белковых субстратов.
Гниение ведет к обогащению почвы азотистыми продуктами. Большое количество связанного азота поступает в почву также в виде мочевины (диамида угольной кислоты) – NH2-CO-NH2. Ежегодно люди и животные выделяют ее около 20 млн т. Но мочевина не может быть непосредственно использована в качестве азотного продукта для питания растений. Она подвергается также аммонификации, которую вызывают различные уробактерии. При этом вначале образуется нестойкая углеаммиачная соль, которая далее расщепляется с образованием NH3, СО2 и Н2О. Мочевая кислота, выделяемая в почву птицами и рептилиями, также быстро минералиэуется особыми группами микроорганизмов с образованием NH3 и СО2 [23].
Следующим важным этапом круговорота азота, вслед за образованием NH3, является процесс нитрификации, или окисления, аммиака в нитриты, а затем в нитраты осуществляют почвенные бактерии, в результате растения получают питательные вещества. Сначала бактерии (нитрозомонас) окисляют аммиак в азотистую кислоту, получая при этом энергию, необходимую для своей жизни. (NH2+1,5O2=NO2+H2O+2H). В присутствии кислорода аммиак дает нитритный ион и воду; выход энергии в этой реакции составляет 65 ккал/моль. Nitrosomonas относится к группе так называемых автотрофов – организмов, которые не потребляют энергию, запасенную в органических веществах. Фотоавтоторфы используют энергию света, а хемоавтотрофы, подобные Nitrosomonas, получают энергию получая ее из неорганических соединений [30].
На втором этапе нитратные бактерии (нитробактер) окисляют азотистую кислоту в азотную. (NO2+О2 = NO3). Процессы денитрофиксации иду при наличии в почве денитрофиксирующих бактерий, которые восстанавливают нитраты до молекудярного азота. NO3 = NO2 = NO = N2.Эти процессы протекают на глубине 10-15см в почве в анаэробных условиях и ведут к понижению плодородия почвы, уменьшая в ней запасы нитритов. При окислении нитритного иона в нитратный высвобождается около 17 ккал/моль – немного, но вполне достаточно для того, чтобы поддержать существование Nitrobacter.
Нитрифицирующие бактерии – строгие аэробы, хемолиттрофы. Энергию окисления они используют для восстановления СО2 в гексозу. Благодаря нитрифицирующим бактериям в почве могут образовываться огромные скопления солей азотной кислоты в виде селитры (в Чили, Перу). Завершая процесс минерализации белковых веществ, нитрифицирующие бактерии играют исключительно важную роль и в процессах самоочищения почвы и воды, и в санитарно-гигиенических устройствах (поля орошения и т. п.).
Следует также отметить, что наряду с нитрифицирующими хемоавтотрофными бактериями известны гетеротрофные микроорганизмы, способные вести близкие процессы. К гетеротрофным нитрификаторам относятся некоторые грибы из рода Fusarium и бактерии таких родов, как Alcaligenes, Corynebacterium, Achromobacter, Pseudomonas, Arthrobacter, Nocardia [17].
Показано, что Arthrobacter sp. окисляет в присутствии органических субстратов аммоний с образованием гидроксиламина и далее нитритов и нитратов. Кроме того, может образовываться гидроксамовая кислота. У ряда бактерий выявлена способность осуществлять нитрификацию органических азотсодержащих соединений: амидов, аминов, оксимов, гидроксаматов, нитросоединений и др.
Размеры гетеротрофной нитрификации в некоторых случаях бывают довольно большие. Кроме того, при этом образуются некоторые продукты, обладающие токсичным, канцерогенным, мутагенным действием и соединения с химиотерапевтическим эффектом. Поэтому исследованию данного процесса и выяснению его значения для гетеротрофных микроорганизмов сейчас уделяют значительное внимание.
Денитрифицирующие бактерии (в частности, некоторые виды Pseudomonas) в анаэробных условиях используют денитрификацию как основную форму дыхания. Для них соли азотной и азотистой кислот служат источниками азота. Энергию для своей деятельности денитрифицирующие бактерии получают из органических веществ, которыми богата почва. Денитрифицирующие бактерии наносят вред сельскому хозяйству, так как приводят к обеднению почвы минеральным азотом и переходу свободного азота в атмосферу. Особенно энергично процессы денитрификации развиваются в слежавшейся, плохо аэрируемой почве. Однако утрата азота почвой из-за активности денитрифицирующих бактерий компенсируется деятельностью свободноживущих аэробных, анаэробных и клубеньковых азот- фиксирующих бактерий. Более 90 % азота связывают азотфиксирующие бактерии, т. е. на каждый гектар почвы ежегодно от 25 до 300 кг азота привносят только они.
5.3 Процессы нитрификации в почве
Прокариоты играют огромную роль в сельском хозяйстве, так как они участвуют в биогехимических циклах важнейших химических элементов, поддерживают плодородие и структуру почв, взаимодействуют с растениями. Прокариоты составляют наибольшую фракцию почвенной микробной биомассы (от 450 до 7000 кг/га при массе пахотного слоя почвы на площади 1 га 2400 – 2700 т). Распределение микроорганизмов в почве неравномерно, потому что они образуют микроколонии, прикрепляясь к частицам минеральных или органических субстратов. В некоторых почвах плотность бактерий может достигать 108 колониеобразующих единиц (КОЕ) на 1 г сухой почвы. В конце ХХ в. для повышения урожайности культур и плодородия почв в сельском хозяйстве стали применять живые чистые культуры микроорганизмов в качестве инокулянтов. С каждым годом использование бактериальных инокулянтов увеличивается на значительных площадях, и их роль будет возрастать в особых системах землепользования, таких как рациональное сельское хозяйство или биологическое земледелие, в мероприятиях по очистке почв от загрязнений и восстановлению ландшафтов. В качестве инокулянтов будут создаваться новые генетически модифицированные штаммы и, естественно, разрабатываться методы контроля за ними [13,16].
Для синтеза белков растениям необходим азот. Азотсодержащие минеральные удобрения подразделяются на аммиачные, нитратные и амидные. Нитратная группа удобрений - селитры: натриевая NaNO3, калиевая KNO3 и кальциевая Ca(NO3)2 .
Недостаток удобрений в следующем: они взаимодействуют с водой постепенно, поэтому и питательные вещества поступают из них к растениям также постепенно. К микробам, использующим энергию, освобожденную при окислении минеральных веществ, относятся также бактерии-нитрификаторы. Они способны превращать аммиак в селитру. Эти бактерии, создают органические вещества из воды, углекислого газа и минеральных солей. Нитрификаторы, как и серобактерии, не нуждаются в солнечной энергии. Их можно встретить даже в бесплодных песках, в трещинах скал, в темных ущельях, лишенных каких-либо признаков жизни. В природных условиях они образуют огромное количество селитры из аммиака, выделяющегося при разложении животных и растительных остатков. В хорошо проветриваемой почве за год может быть образовано на гектаре более четверти тонны селитры — ценного азотного удобрения. В некоторых местах, где растительность скудна или ее совсем нет, а дождей почти не бывает, накапливающаяся селитра не вымывается из почвы. Здесь образуются залежи селитры [7].
В хорошо аэрируемой почве ионы NH+, освобождающиеся при минерализации азотсодержащих веществ, подвергаются быстрому окислению. Перевод катиона в анион ведет к подкислению почвы и тем самым к повышению растворимости минералов (солей калия, магния, кальция и фосфорной кислоты). Поэтому в нитрифицирующих микроорганизмах видели ранее важный фактор плодородия почв. Однако теперь эти представления изменились. Выяснилось, что ионы аммония задерживаются в почве гораздо лучше нитрата, особенно если они адсорбируются на глинистых минералах и более или менее прочно связываются с частицами гумуса; нитрат же легко вымывается. В связи с этим появилась тенденция к ограничению нитрификации в почвах сельскохозяйственных угодий. Ведутся поиски веществ, способных специфически подавлять рост нитрифицирующих бактерий и служить своего рода «стабилизаторами» почвенного азота (к таким веществам относится, например, 2-хлор-6-(трихлорметил)-пиридин).
Процесс нитрификации, являясь важным звеном в круговороте азота в природе, имеет как положительные, так и отрицательные стороны. Оказалось, например, что переведение азота из аммонийной формы в нитратную способствует обеднению почвы азотом, так как нитраты, как весьма растворимые соединения, легко вымываются из почвы. В то же время известно, что нитраты — хорошо используемый растениями источник азота. Кроме того, связанное с нитрификацией подкисление почвы улучшает растворимость и, следовательно, доступность некоторых жизненно необходимых элементов, в первую очередь фосфора и железа.
Нитрифицирующие бактерии - аэробы и требуют для развития постоянного притока кислорода. Они более активны в почвах, хорошо аэрируемых, близких к нейтральным, содержащих большое количество органических соединений. Поэтому нитрификация протекает интенсивно в дерново-подзолистых почвах, достаточно унавоженных и хорошо обрабатываемых, а также в черноземных и каштановых почвах, имеющих благоприятный водный режим. В процессе нитрификации образующаяся азотная кислота, соединяясь с основаниями, находящимися в почве, дает нитраты, которые используются растениями в качестве азотной пищи. При активной нитрификации в почве может накопиться в течение года до 300 кг нитратов на 1 га. Нитраты почвы хорошо потребляются растениями, но будучи легко- растворимыми в воде, могут легко вымываться в более глубокие слои почвы или за ее пределы.
Протекающие в почве реакции, в которых азот восстанавливается, дают значительно больше энергии, чем окислительные реакции, в результате которых у атомов азота отнимаются электроны. Энергия, выделяющаяся при окислении аммиачного азота до нитратного, используется бактериями для ассимиляции углекислого газа и для других эндотермических процессов.
В целом, нитрифицирующие бактерии способствуют повышению урожайности почвы благодаря накоплению в ней азотнокислых солей. Однако в почве происходят и противоположные процессы, т.е. процессы денитрификации, или восстановления микроорганизмами солей азотной кислоты в соли азотистой кислоты и в другие простые азотистые соединения, вплоть до свободного азота, который уходит в атмосферу. Способностью восстанавливать нитраты в нитриты обладает большое количество видов бактерий и грибов.
Нитрифицирующие бактерии косвенно участвуют в разрушении разного рода сооружений, для которых строительным материалом служат известь и цемент (т. е. различных зданий, автострад и т. п.). Это связано с тем, что нитрифицирующие бактерии окисляют аммиак, присутствующий в атмосфере или выделяющийся из фекалий животных, до азотной кислоты.
5.4 Применение нитрифицирующих бактерий в биоиндикации
По современным представлениям биоиндикаторы - организмы, присутствие, количество или особенности развития которых служат показателями естественных процессов, условий или антропогенных изменений среды обитания. Биоиндикация - метод, который позволяет судить о состоянии окружающей среды по факту встречи, отсутствия, особенностям развития организмов - биоиндикаторов.
Условия, определяемые с помощью биоиндикаторов, называются объектами биоиндикации. Ими могут быть как определенные типы природных объектов (почва, вода, воздух), так и различные свойства этих объектов (механический, химический состав и др.) и определенные процессы, протекающие в окружающей среде (эрозия, дефляция, заболачивание и т.п.), в том числе происходящие под влиянием человека (Экологический мониторинг, 2006).
Биоиндикационный метод исследования с успехом применяется для оценки состояния и функционирования почвы и последовательности биоценотических изменений в экосистеме, поскольку загрязняющие вещества влияют, прежде всего, на протекание жизненных процессов. Наибольший интерес в этом плане представляют почвенные животные (обладают повышенной чувствительностью к внешним факторам) - многочисленная группа животных, имеющая экологическое и экономическое значение.
Советскими авторами Никитина 3. И., Мамитко А. В., Мамитко В. Г. научно обоснована возможность использования микробиологических показателей для определения степени доступности техногенной нагрузки на экосистему [18].
В основе принципа биологической диагностики почв лежит представление о том, что почва как среда обитания составляет единую систему с населяющими ее популяциями разных организмов. В зависимости от сочетания природных факторов, определяющих почвообразовательный процесс, разные почвы различаются по составу своей биоты, направленности биохимических превращений и содержанию тех химических компонентов, которые являются продуктами этих превращений или их агентами.
Существуют специальные функциональные группы микроорганизмов, которые являются показателями загрязнения окружающей среды различными веществами. Так, например, некоторые авторы (Чегринец, Безбородько, Воронова, Никула, 1980) предлагают оценивать состояние биоценоза почвы, загрязненной медь - и хромсодержащими веществами, по жизнедеятельности наиболее важных групп микроорганизмов нитрификаторов, целюлозоразрушающих и по общему количеству микроорганизмов с учетом удельного веса споровых форм [18,25].
В качестве индикаторов фенольного загрязнения используются евтрофные микроорганизмы, которые наиболее чувствительны к данному соединению. По мере увеличения концентрации фенола отмечено падение численности этой группы микроорганизмов [11].
Хайниш отмечает тенденции преимущественного уменьшения численности отдельных групп микроорганизмов при внесении определенных пестицидов. Например, каптан снижал преимущественно численность патогенных грибов, гептахлор - бактерий, цинеб - спорообразующих бактерий, эптам, дикват, атразин - грибов и т.д. Однако, согласно Марфениной О.Е., не ясно, насколько это воздействие сохраняется в разных экологических условиях [19,13].
Существуют определенные группы микроорганизмов, уменьшение численности которых свидетельствует об антропогенном воздействие на почву. Данные физиологические группы реагирует на изменение многих показателей. Это, как правило, облигатные аэробные организмы, негативно реагирующие на увеличение органики в среде. К таким группам относятся нитрификаторы и целлюлозоразрушающие микроорганизмы.
Нитрификация замедляется или прекращается в том случае, если почва является слишком холодной, слишком сухой или при загрязнении.
В загрязненных почвах бурно развиваются микроорганизмы, осуществляющие распад гнилостных продуктов, а нитрификация подавлена. При накоплении в почве аммиака, образующегося при распаде органических загрязнителей, начинается развитие нитрификаторов. Интенсивный процесс нитрификации говорит о завершении распада органических загрязнителей и активно идущем самоочищении почвы [10].
Численность микроорганизмов - нитрификаторов снижается при внесении в почву медь- и хромсодержащих веществ, при загрязнении почвы серой, ПАВ. Среди микроорганизмов цикла азота автотрофные нитрифицирующие бактерии наиболее чувствительны к действию нефти. Снижение их активности связано с наличием в почве легкодоступных органических соединений в виде углеводородов нефти и продуктов их частичного окисления. Поэтому снижение численности нитрификаторов, как и целлюлозоразрушающих микроорганизмов, может служить индикатором нефтяного загрязнения почв [1,18, 19].
В качестве показателя активности самоочищения почвы используют определение энергии нитрификации. Усиленному размножению нитрифицирующих бактерий способствует наличие в почве большого количества аммиака — продукта белкового распада. Чем больше минерализовано органических веществ в почве, тем интенсивнее протекает процесс нитрификации и, следовательно, процесс самоочищения почвы.
ЗАКЛЮЧЕНИЕ
Целью данной работы было собрать и обобщить доступные из публикаций сведения о нитрифицирующих прокариотах, особенности и функционирования бактериальной клетки в связи с процессом нитрификации.
Нитрифицирующие прокариоты – грамотрицательные микроорганизмы, различающиеся формой и размером клеток. В эту группу входят бактерии с палочковидной, сферической, спиралевидной, грушевидной формой клеток. Все нитрифицирующие бактерии, кроме представителей рода Nitrobacter, размножаются бинарным делением. Бактерии, принадлежащие к роду Nitrobacter, размножаются почкованием.
Среди нитрифицирующих бактерий есть подвижные (с полярным или перитрихиальным жгутикованием) и неподвижные формы. Все нитрифицирующие бактерии – облигатные аэробы; большинство – облигатные автотрофы, рост которых ингибируется органическими соединениями в концентрациях, обычных для гетеротрофных прокариот.
Ассимиляция СО2 осуществляется в цикле Кальвина.
Размеры клеток разных видов Nitrobacteriaceae колеблются от 0,3 до 1 мкм в ширину и от 1 до 6,5 мкм в длину.
Почти у всех нитрификаторов имеется хорошо развитая система внутрицитоплазматических мембран, значительно различающихся по форме и расположению в клетках разных видов. Эти мембраны подобны мембранам фотосинтезирующих пурпурных бактерий.
Оптимальные условия для роста нитрифицирующих бактерий – температура 25–30 °С при рН 7,5–8,0.
Процесс нитрификации происходит в два этапа. На первом этапе происходит окисление аммиака до нитритов:
NH4++ 1/2 О2NO2-+H2O+2H+
а на втором нитриты окисляются до нитратов:
NO2-+1/2 O2NO3-
Нитрифицирующие прокариоты обнаружены в водоемах разного типа (озера, моря, океаны) и в почвах, где они, как правило, развиваются совместно с бактериями, жизнедеятельность которых приводит к образованию исходного субстрата нитрификации – аммиака.
Процесс нитрификации, являясь важным звеном в круговороте азота в природе, имеет как положительные, так и отрицательные стороны. Переведение азота из аммонийной формы в нитратную способствует объединению почвы азотом, так как нитраты как весьма растворимые соединения легко вымываются из почвы. В то же время известно, что нитраты – это хорошо используемый растениями источник азота. Кроме того, связанное с нитрификацией подкисление почвы улучшает растворимость и, следовательно, доступность некоторых жизненно необходимых элементов, в первую очередь фосфора и железа.
Нитрифицирующие бактерии косвенно участвуют в разрушении разного рода сооружений, для которых строительным материалом служат известь и цемент (т. е. различных зданий, автострад и т. д.). Это связано с тем, что нитрифицирующие бактерии окисляют аммиак, присутствующий в атмосфере или выделяющийся из фекалий животных, до азотной кислоты.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ:
- Асонов Н.Р. Микробиология / Н.Р. Асонов - 2-е изд., перераб. и доп. - М.: Агропромиздат, 1989. – 175 с.
- Бабьева М.А. Биология почв / М.А. Бабьева, Н.К. Зенова - М.: Изд-во МГУ, 1989. – 263 с.
- Виноградский С.Н. Микробиология почвы / С.Н. Виноградский - М.: Л., 1952. - 792 с.
- Воробьева А.А. Микробиология и иммунология: Учебник / А.А. Воробьева - М.: Медицина, 1999.— 464 с.: ил.
- Голубева И.В. Энтеробактерии: руководство для врачей / И.В. Голубева, В.А Килессо, Б.С. Киселева и др.; под ред. В.И. Покровского. – М.: Медицина, 1985. – 321 с.
- Горленко В.М. Экология водных микроорганизмов / В.М. Горленко, Г.А. Дубинина, С.И. Кузнецов – М: Издательский центр «Академия», 1977 – 137с.
- Гусев М.В. Микробиология: Учебник для студ. биол. специальностей
вузов / М. В. Гусев, Л. А. Минеева. 4-е изд., стер. — М.: Издательский центр «Академия», 2003. - 464 с. - Емцев, В.Т. Микробиология: учебник для вузов / В.Т. Емцев, Е.Н. Мишустин. – 7-е изд., стереотип. – М.: Дрофа, 2008 – 444 с.: ил.
- Колешко О. И., Микробиология с основами вирусологии: Учебник / Колешко О. И., Завезенова Т. В. - Иркутск: Изд-во Иркут. ун-та, 1999. - 452 с., 94 ил.
- Коротяев А. И. Медицинская микробиология, иммунология и вирусология : учебник для мед. вузов / А. И. Коротяев, С. А. Бабичев - 4-е изд., испр. и доп. - СПб.: СпецЛит, 2008. - 767 с.: ил.
- Лысак, В. В. Микробиология: учеб. пособие / В. В. Лысак. - Минск : БГУ, 2007. – 426 с.
- Марфенина О.Е. "Микробиологические аспекты охраны почв" / О.Е. Марфенина - М.: Изд-во МГУ, 1991. - 78 c.
- Методические указания по определению тяжелых металлов в почвах сельхозугодий и продукции растениеводства - Утв: 10.03.1992. - М.: ЦИНАО. - 62 с.
- Мишустин Е.Н. Микроорганизмы и продуктивность земледелия / Е.Н. Мишустин – М: Изд-во «Наука», 1972. - 312с.
- Мишустин Е.Н. Микроорганизмы как компоненты биогеоценоза / Е.Н. Мишустин - М: Изд-во «Наука», 1984. - 134с.
- Мурадова, Е.О. Микробиология / Е.О. Мурадова, К.В. Ткаченко. – М.: Эксмо, 2009. – 336 с.
- Нетрусов, А.И. Общая микробиология: учебник для студ. высш. учеб. заведений / А.И. Нетрусов, И.Б. Котова. – М.: Издательский центр «Академия», 2007. – 288 с.
- Никитина 3. И. Мониторинг загрязнения наземных экосистем по микробиологическим показателям. / 3. И. Никитина, А. В. Мамитко, В. Г. Мамитко – Изд-во: «Географические и природные ресурсы», 1980. – 167 с.
- Никитина З.И. Микробиологический мониторинг наземных экосистем. / З.И. Никитина. - Новосибирск: Наука, 1991. - 219 с.
- Никитина Е.В.Микробиология: Учебник / Е.В. Никитина, С.Н. Киямова, О.А. Решетник – СПб.: ГИОРД, 2008. – 368с.: ил.
- Определитель бактерий Берджи / Под ред. Дж. Хоулта, Н. Крига, П. Смита и др. - Пер. с англ. М.: Мир, 1997т В 2-х т.
- Орлов Д.С. Экология и охрана биосферы при химическом загрязнении / Д.С. Орлов, Л.К. Садовникова, И.Н. Лозановская - М. Высшая школа, 2002. – 352 с.
- Перспективные методы обработки осадков сточных вод - М.: ЦИНИС, 1995.- 100 с.
- Роль микроорганизмов в круговороте газов в природе / Под ред., Г.И. Заварзина - М., Наука, 1979. - 287c
- Руководство к практическим занятиям по микробиологии: Учеб. пособие / Под ред. Н. С. Егорова. — 3-е изд., перераб. и доп. — М.: Изд-во МГУ, 1995. — 224 с.: ил.
- Сидоренко О.Д. Микробиология: Учебник для агротехнологов / О.Д. Сидоренко, Е.Г. Борисенко, А.А. Ванькова, Л.И. Войно – М.: ИНФРА-М, 2005. – 293 с.: ил.
- Стефенсон М. Метаболизм бактерий, / М. Стефенсон - Изд-во Иностр. лит-ры, Москва, 1951. – 576 с.
- Чегринец Г.Я. Состояние биологической активности почвы как показатель при нормировании экзогенных химических веществ в почве // Г.Я. Чегринец, М.Д. Безбородько, Г.Ф. Воронова, Р.Г. Никула - Изд-во «Киев», 1980. Вып. 19. - 283 с.
- Чурбанова И.Н. Рациональное использование водных ресурсов и обезвреживание промешенных стоков / И.Н.Чурбанова. - М. : Высш. шк., 1987. – 238 с.
- Шведова JI.B. Миграция тяжелых металлов в системе «почва-растение» / JI.B. Шведова, Т.А. Чеснокова, А.В. Невский - Инженерная экология. 2004. № 6. - 227 с.
- Хотько Э. И. Справочник вредителей плодовых и ягодных культур / Э. И. Хотько, Т. П. Панкевич, Р. В. Молчанова, С. И. Ярчаковская - Изд-во: «Белорусская Энциклопедия», 1982. – 264 с.
Stanley W. Watson, Woods Hole Oceanographic Institute, 1995 [Электронный ресурс] / режим доступа:
http://www.whoi.edu/main/obituaries/archive?tid=3622&cid=853
- LPSN: List of prokaryotic names with standing in nomenclature
[Электронный ресурс] / режим доступа: http://www.bacterio.net/index.html
PAGE \* MERGEFORMAT 2
НИТРИФИЦИРУЮЩИЕ ПРОКАРИОТЫ