СИНАПТИЧЕСКОЕ ВЗАИМОДЕЙСТВИЕ НЕЙРОНОВ
Контрольная работа
СИНАПТИЧЕСКОЕ ВЗАИМОДЕЙСТВИЕ НЕЙРОНОВ
Содержание
Введение
1 Строение химических синапсов
2. Медиаторы
3. Электрические процессы на постсинаптической мембране
4. Электрические синапсы
Литература
Введение
Взаимодействие нейронов между собой и между нервными клетками и исполнительным аппаратом (мышечными и железистыми клетками) осуществляется через контакты, синапсы. Термин «синапс» был впервые предложен выдающимся английским нейрофизиологом Ч.Шерригтоном в 1897г. хотя сведения о гистологической структуре контактов между нервными и другими клетками были известны ещё задолго до этого. Было очевидным, что в области контакта прекращается непрерывное распространение электрического потенциала и передача возбуждения связана с особыми механизмами, отличными от закономерностей движения возбуждения по нервным волокнам. Многочисленными исследованиями, начиная с работ О.Леви (1921), было показано, что передача возбуждения осуществляется с помощью особых химических веществ, названных медиаторами или трансмиттерами. Фармакологические, гистохимические исследования, изучение распространения возбуждения в нервных центрах выявили многочисленные факты которые легли в основу теории химической природы синаптической передачи. Длительное время химическая теория не находила всеобщего признания.
В связи с усовершенствованием методов морфологических исследований, появлением высокочувствительных усилителей биопотенциалов, в период 30-40 годов ХХ века были описаны многие закономерности синаптической передачи – односторонность проведения – от пресинаптического нейрона к постсинаптическим структурам, различия в процессах, протекающих в пре- и постсинаптических структурах. Были выявлены особенности формирования и распространения быстрых и медленных электрических потенциалов, описаны постсинаптические потенциалы, , показана связь между медленными постсинаптическими потенциалами и потенциалами действия, идентифицированы некоторые медиаторы, и др. (Ллойд, Леви, Экклс, Дейл, Барон и Мэтьюз, Геницинский ). Безусловной победой химической теории передачи возбуждения в синапсах следует считать факт присуждения в 1936 году Дейлу и Леви Нобелевской премии за работы в этом направлении. Выявление тонких механизмов работы синапсов стало возможным с введением в арсенал исследователей микроэлектродной техники и электронной микроскопии. Начиная с конца 40-х годов ХХ века развернулись интенсивные исследования процессов, протекающих в синапсах, включая процессы электрические, химические, фармакологические и информационные. Применение внутриклеточных микроэлектродов позволило наблюдать и изучать электрические процессы в отдельных частях клетки, в том числе и в области синаптического контакта, кроме того, через стеклянные микроэлектроды – микропипетки оказалось возможным электрофоретически вводить в область регистрации различные химические агенты, проводя, таким образом, точный анализ физико-химических процессов, лежащих в основе работы синапса (Экклс, Катц, Миледи, Костюк). Применение микроэлектродов позволило выявить и изучить ещё один вид контактного взаимодействия – влияние полем потенциала в местах плотного контакта конечной терминали аксона с электровозбудимыми структурами других клеток. Способ подобного взаимодействия получил название электрического синапса.
В последние десятилетия ХХ века исследования синаптического взаимодействия были направлены на детальное изучение работы различных синапсов – особенностей биосинтеза различных медиаторов и выделение их в синаптическое пространство, влияние медиаторов на структуры постсинаптической мембраны, движение и роль химических веществ разной природы в синаптических мембранах , поиск новых медиаторов и исследование механизмов их действия. Детально изучается распределение нейромедиаторов – нейропептидов в отдельных структурах центральной нервной системы.
1 Строение химических синапсов
В зависимости от того, какие структуры нейрона участвуют в образовании синапса, выделяют аксосоматические, аксодендритные, аксоаксональные и дендродентритные синапсы. Синапс, образованный аксоном мотонейрона и мышечной клеткой называется концевой пластинкой (нервно-мышечное соединение, мионевральный синапс). Непременными структурными атрибутами синапса являются пресинаптическая мембрана, постсинаптическая мембрана и синаптическая щель между ними. Остановимся подробнее на каждой из них.
Пресинаптическая мембрана образована окончанием конечных ветвлений аксона (или дендрита в дендродендритном синапсе). Отходящий от тела нервной клетки аксон покрывается миэлиновой оболочкой, которая сопровождает его на всём протяжении, вплоть до разветвления на конечные терминали. Количество конечных ветвлений аксона может достигать нескольких сотен, а протяженность их, теперь уже лишенных миэлиновой оболочки – до нескольких десятков мкм. Конечные ветвления аксона имеют малый диаметр – 0,5-2,5 мкм, иногда больше. Окончания терминалей в месте контакта имеют разнообразную форму – в виде булавы, сетевидной пластинки, колечка, или могут быть множественными – в виде чашечки, кисти. Конечная терминаль может иметь несколько расширений, контактирующих по ходу движения с разными участками одной клетки или с разными клетками, формируя таким образом множество синапсов. Некоторые исследователи подобные синапсы называют касательными.
В месте контакта конечная терминаль несколько утолщается и часть её мембраны, прилегающая к мембране контактируемой клетки образует пресинаптическую мембрану. В зоне конечной терминали, прилегающей к пресинаптической мембране путём электронной микроскопии обнаружено скопление ультраструктурных элементов – митохондрий, число которых колеблется, достигая иногда нескольких десятков, микротрубочек и синаптических пузырьков (везикул). Последние бывают двух видов – агранулярные (светлые) и гранулярные (тёмные). Первые имеют размер 40-50 нм, диаметр гранулярных везикул, как правило, более 70 нм. Их мембрана подобна клеточным и состоит из фосфолипидного бислоя и белков. Большая часть везикул фиксируется на цитоскелете с помощью специфического белка - синапсина, образуя трансмиттерный резервуар. Меньшая часть везикул прикрепляется к внутренней стороне пресинаптической мембраны посредством белка мембраны везикулы - синаптобревина и белка пресинаптической мембраны - синтаксина. Существует две гипотезы относительно происхождения везикул. Согласно одной из них (Хаббард, 1973), они образуются в области пресинаптического окончания из так называемых окаймлённых пузырьков. Последние формируются в углублениях клеточной мембраны пресинаптического окончания и сливаются в цистерны, от которых и отпочковываются везикулы, заполняемые медиатором. Согласно другому взгляду, везикулы как мембранные образования формируются в соме нейрона, пустыми транспортируются по аксону в область пресинаптического окончания и там заполняются медиатором. После выброса медиатора опустошенные везикулы ретроградным аксонным транспортом возвращаются в сому, где деградируются лизосомами. Синаптические пузырьки наиболее плотно расположены вблизи внутренней поверхности пресинаптичесой мембраны и их количество непостоянно. Везикулы заполнены медиатором, кроме того здесь сосредоточены так называемые котрансмиттеры – вещества белковой природы, играющие существенную роль в обеспечении активности основного медиатора. Малые везикулы содержат низкомолекулярные медиаторы, а большие – белки и пептиды. Показано, что медиатор может находиться и вне везикул. Расчеты показывают, что в нервно-мышечном соединении человека плотность везикул достигает 250-300 на 1 мкм2, а общее их количество – около 2 - 3х млн в одном синапсе. В одном пузырьке сосредоточено от 400 до 4-6 тысяч молекул медиатора, что и составляет так называемый «квант медиатора», выделяющийся в синаптическую щель спонтанно или при приходе импульса по пресинаптическому волокну. Поверхность пресинаптической мембраны неоднородна – в ней имеются утолщения, активные зоны, где скапливаются митохондрии и плотность везикул наибольшая. Кроме того, в области активной зоны выявлены потенциалзависимые кальциевые каналы, по которым кальций проходит сквозь пресинаптическую мембрану внутрь пресинаптической зоны конечной терминали. Во многих синапсах в пресинаптическую мембрану встроены так называемые ауторецепторы. При их взаимодействии с выделенными в синаптическую щель медиаторами, выделение последних либо усиливается, либо прекращается в зависимости от типа синапса.
Синаптическа щель – пространство между пресинаптической и постсинаптической мембранами, ограниченная площадью контакта, размер которой для большинства нейронов колеблется в пределах нескольких мкм2. Площадь контакта может варьировать в разных синапсах, что зависит от диаметра пресинаптической терминали, формы контакта, характера поверхности контактирующих мембран. Так, для наиболее изученных нервно-мышечных синапсов показано, что площадь контакта одной пресинаптической терминали с миофибриллой может составлять десятки мкм2. Размер синаптической щели колеблется от 20 до 50-60 нм. За пределами контакта полость синаптической щели сообщается с межклеточным пространством, таким образом между ними возможен двухсторонний обмен разными химическими агентами.
Постсинаптическая мембрана представляет собой участок мембраны нейрона, мышечной или железистой клетки, контактирующей с пресинаптической мембраной. Как правило, область постсинаптической мембраны несколько утолщена по сравнению с соседними участками контактируемой клетки. В 1959 году Е.Грей предложил разделить синапсы в коре мозга на два типа. Синапсы 1-го типа имеют более широкую щель, постсинаптическая мембрана у них толще и плотнее, чем у синапсов 2-го типа, уплотненный участок более обширен и занимает большую часть обеих синаптических мембран.
В постсинаптическую мембрану встроены белково-гликолипидные комплексы, выполняющие роль рецепторов, способных связываться с медиаторами и образовывать ионные каналы. Так, ацетилхолиновый рецептор в мионевральном синапсе состоит из пяти субъединиц, которые образуют комплекс с молекулярной массой 5000-30000, пронизывающий мембрану. Расчетным способом показано, что плотность таких рецепторов может составлять до 9 тысяч на мкм2 поверхности постсинаптической мембраны. Головка комплекса, выступающая в синаптическую щель имеет так называемый «узнающий центр». При связывании с ним двух молекул ацетилхолина ионный канал открывается, его внутренний диаметр становится проходимым для ионов натрия и калия, при этом канал остаётся непроходимым для анионов из-за имеющихся на его внутренних стенках зарядов. Важнейшую роль в процессах синаптической передачи играет мембранный белок, названный G-белком, который в комплексе с гуанинтрифосфатом (ГТФ) активирует ферменты, включающие вторичные мессенджеры – внутриклеточные регуляторы.
Рецепторы постсинаптических мембран находятся в так называемых «активных зонах» синапсов и среди них различают два типа – ионотропные и метаботропные. В ионотропных рецепторах (быстрых) для открытия ионных каналов достаточно их взаимодействия с молекулой медиатора, т.е. медиатор непосредственно открывает ионный канал. Своё название метаботропные (медленные) рецепторы получили в связи с особенностями их функционирования. Открытие ионных каналов в этом случае связано с каскадом метаболических процессов, в которых участвуют различные соединения (белки, в том числе и G-белок, ионы кальция, циклические нуклеотиды – цАМФ и цГМФ, диацетилглицерины), играющие роль вторичных мессенджеров. Метоботропные рецепторы сами по себе не являются ионными каналами; они лишь модифицируют работу расположенных рядом ионных каналов, ионных насосов и других белков посредством непрямых механизмов. К ионотропным относятся рецепторы ГАМК, глицина, глутамата, Н-холинорецепторы. К метаботропным – рецепторы дофамина, серотонина, норадреналина, М-холинорецепторы, некоторые рецепторы ГАМК, глутамата.
Обычно рецепторы располагаются строго в пределах постсинаптической мембраны, поэтому влияние медиаторов возможно только в области синапса. Обнаружено, однако, что небольшое количество рецепторов чувствительных к ацетилхолину имеется и за пределами нервно-мышечного синапса в мембране мышечной клетки. В некоторых условиях (при денервации, отравлении некоторыми ядами) чувствительные к ацетилхолину зоны могут образовываться вне синаптических контактов на миофибрилле, что сопровождается развитием гиперчувствительности мышцы к ацетилхолину.
Рецепторы, чувствительные к ацетилхолину широко распространены также в синапсах ЦНС и в периферических ганглиях. Рецепторы возбуждающего действия разделены на два класса, различающиеся по фармакологическим признакам.
Один из них – класс рецепторов, на которые влияния, сходные с ацетилхолином оказывает никотин, отсюда их название - никотиночувствительные (Н-холинорецепторы), другой класс – чувствительные к мускарину (яд мухомора) названы М-холинорецепторами. В связи с этим синапсы, где основным медиатором служит ацетилхолин, разделяются на группы никотинового и мускаринового типа. Внутри этих групп выделяют много разновидностей в зависимости от месторасположения и особенностей функционирования. Так, синапсы с Н-холинорецепторами описаны во всех скелетных мышцах, в окончаниях преганглионарных парасимпатических, и симпатических волокон, в мозговом слое надпочечников , а мускариновые синапсы – в ЦНС, гладких мышцах (в синапсах, образованных окончаниями парасимпатических волокон), в сердце.
2. Медиаторы
Медиаторы - это вещества, отвечающие следующим требованиям: они должны синтезироваться в данном нейроне, должны накапливаться в синаптических везикулах и из них выделяться, иметь сопутствующую систему своей инактивации (разрушения ферментами или обратного всасывания), при аппликации на данный нейрон вещество должно действовать идентично предполагаемому медиатору и имитировать действие пресинаптического волокна (Куффлер, Николс). Медиаторы имеют разную природу, структуру, различаются по механизмам синтеза, действия и распада. Принято выделять три группы медиаторов. К первой относят «классические» медиаторы – ацетилхолин, норадреналин, адреналин, дофамин, гистамин, серотонин, АТФ, глутамат, гамма-аминомаслянную кислоту (ГАМК), глицин. Ко второй относится большая группа нейропептидов (свыше 40), найденных в центральной нервной системе. Среди них – нейропептид Y, бомбезин, группа опиоидных пептидов, секретин, соматостатин, вещество Р и др. В отдельную группу выделяют оксид азота (NO) и оксид углерода (CO). Наиболее изучены ацетилхолин, моноамины (норадреналин, дофамин, серотонин, гистамин), аминокислоты (глутамат, глицин, ГАМК), пуриновые нуклеотиды и некоторые олигопептиды, действующие как трансмиттеры или как модуляторы синаптических процессов. К последним относят энкефалин, эндорфин, динорфин, соматостатин, вещество Р, нейропептид Y и другие.
Вначале предполагали, что все медиаторы синтезируются в теле нейрона и в готовом виде транспортируются по аксону в область пресинаптического окончания. Однако, по мере изучения синаптических процессов, стало ясно, что подобный механизм весьма малоэффективен вследствие небольшой скорости аксонального транспорта, которая не может обеспечить доставку медиатора к месту его действия в условиях его постоянного расходования в синаптических структурах. Сегодня известно, что большинство медиаторов синтезируются в области пресинаптического окончания, где имеются и необходимые для этого компоненты, и ферментные системы, регулирующие их синтез. Понятно, что в этом случае скорость аксонного тока не является лимитирующим фактором, поскольку «сборка» медиаторов осуществляется на месте. Необходимые для синтеза ферменты непрерывно доставляются в синаптические окончания путем аксонного транспорта, создавая достаточный его резерв в пресинаптических окончаниях. Медиаторы, имеющие пептидную и белковую природу, очевидно синтезируются в клеточных телах и аксонным током транспортируются в пресинаптическое окончание. В течение длительного времени считалось, что один нейрон синтезирует один медиатор, который выделяется во всех синапсах, образуемых его конечными терминалями (принцип Дейла). Исследования последних десятилетий показали, что почти все пресинаптические терминали освобождают не один медиатор, а целый ряд биологически активных соединений – «синаптический коктейль». Он может включать в себя, например глицин, ГАМК, глутамат, вещество Р, энкефалин, АТФ, окситоцин и др. Таким образом, хотя названия некоторых синапсов образовались от названия их основного, хорошо изученного ранее медиатора (например, холинергический), следует иметь в виду определённый полихимизм большинства синапсов.
Ацетилхолин – уксуснокислый эфир холина является медиатором в нервно-мышечном соединении, синапсах моронейронов на клетках Реншоу спинного мозга, в ганглиях симпатической и парасимпатической нервной системы, в синапсах мозгового вещества надпочечников, в постганглионарных синапсах эффекторных органов. В последнее время холинэргические синапсы описаны во многих структурах ЦНС – в ядрах перегородки, преоптических ядрах, в ретикулярной формации, гиппокампе и в коре головного мозга. Синтез ацетилхолина осуществляется в пресинаптических терминалях с участием фермента холинацетилтрансферазы. С помощью переносчиков медиатор заполняет везикулы, в которых он концентрируется и из которых попадает в синаптическую щель (о механизме освобождения везикул от медиатора см. далее). Сразу же после связывания с узнающим центром на постсинаптической мембране (почти одновременно) медиатор гидролизуется находящимся здесь же ферментом ацетилхолинэстеразой на составляющие компоненты – ацетат (остаток ацетила) и холин. Последний посредством высокоспецифичного механизма транспорта захватывается пресинаптическим окончанием и используется для синтеза новых молекул ацетилхолина. Полагают, что около половины поглощаемого при этом холина образуется в результате гидролиза ранее выделившегося ацетилхолина, а остальной холин, по-видимому, поступает из плазмы крови. Следовательно, не весь попавший в синаптичесую щель ацетилхолин гидролизуется, часть его попадает в межклеточное пространство и в кровь, также подвергаясь распаду (рис. 1)
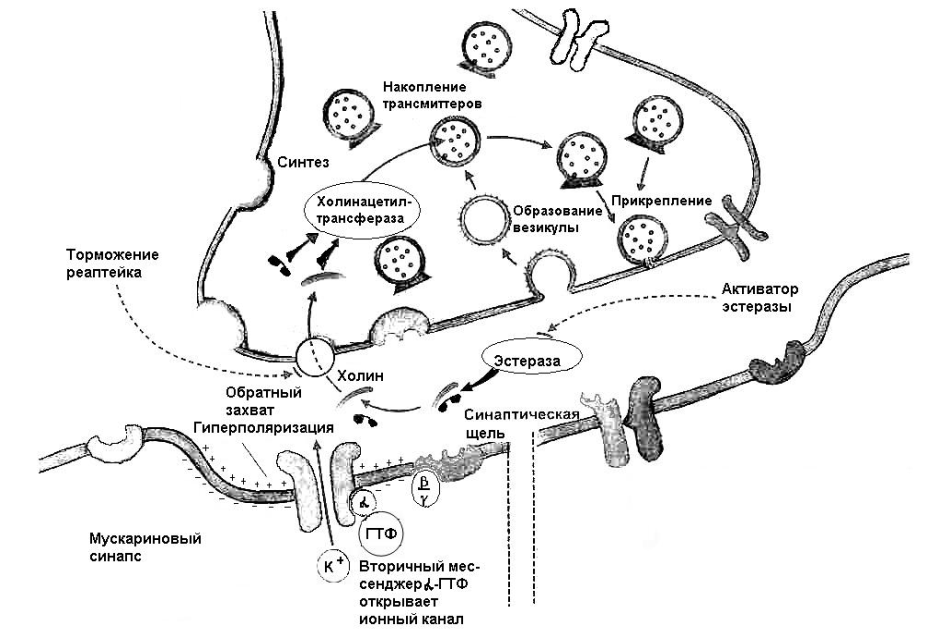
Рис. 1 Инактивация ацетилхолина в синапсе (Р.Клинке, 2004)
Глутамат – L-глутаминовая кислота является важнейшим медиатором для возбуждающих синапсов в центральной нервной системе. Глутамат имеется в крови, однако, в связи с тем, что это вещество плохо проходит через гематоэнцефалический барьер, в нервной ткани он синтезируется из глюкозы, приносимой кровью. Синапсы, в которых глутамат выступает в роли медиатора, составляют около половины всех межнейронных соединений в центральной нервной системе. Наибольшее их число определяется в переднем мозге, гиппокампе и мозжечке. Глутаматные рецепторы на постсинаптической мембране в большинстве своём являются ионотропными и подразделяются на три типа, различающиеся способностью связываться с экзогенными агонистами, каждый из которых в свою очередь делится на несколько подтипов. Один из типов связывается с N-метил-D-аспартатом (MNDA) и получил название MNDA-рецептор. Магний, находящийся в межклеточном пространстве, может выступать как неконкурентный блокатор сопряженного с этим рецептором ионного канала, таким образом, выделившийся медиатор становится неэффективным. Другой тип рецептора (AMPA-рецептор) своё название получил в связи с его способностью связываться с -амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислотой (AMPA). Считается, что активация этих рецепторов способствует длительному потенцированию, что создаёт основу для процессов обучения. Отмечено, что во многих синапсах вместо глутамата находится аспартат. Из синаптической щели глутамат удаляется либо путём обратного захвата в пресинаптическую зону конечной терминали, либо поглощается клетками астроглии.
Глицин и Гамма-аминомаслянная кислота (ГАМК) являются тормозными медиаторами. Они обнаружены в окончаниях многих нейронов – в клетках Рэншоу спинного мозга (в основном глицин, и около 30% - ГАМК), в клетках Пуркинье мозжечка и гиппокампе – здесь ГАМК играет роль нейромодулятора. ГАМК присутствует в синапсах локальных сетей интернейронов во многих структурах центральной нервной системы, в том числе и в коре больших полушарий. Выброшенная в синаптическую щель ГАМК после воздействия на рецепторы захватывается пресинаптическим окончанием и глиальными клетками, здесь трансформируется в глутамин, который, возвращаясь в нейроны, является предшественником новых молекул ГАМК. Таким образом, ГАМК представляет собой продукт декарбоксилирования глутамата с помощью фермента, специфичного для нервной ткани. ГАМК-рецепторы могут быть ионотропными и метаботропными. Известно минимум два различных постсинаптических механизма, запускаемых этими двумя группами рецепторов. Одни из них – ГАМКа-рецепторы и ГАМКс-рецепторы открывают непосредственно каналы для ионов хлора. что приводит к появлению входящего тока и гиперполяризации постсинаптической мембраны. Другие – ГАМКв-рецепторы являются метаботропными и запускают механизм подавления освобождения медиатора или активируют калиевые каналы, что также приводит к гиперполяризации постсинаптической мембраны. Глицинергические нейроны распространены в спинном и продолговатом мозге, где они выполняют функции тормозных элементов.
Катехоламины – родственные в химическом отношении медиаторы являются производными тирозина. К ним относятся дофамин, норадреналин и адреналин. Путь синтеза у них общий: 1 – гидроксилирование тирозина с образованием диоксифенилаланина (ДОФА); 2 – декарбоксилирование ДОФА, в результате чего получается дофамин; 3 – гидроксилирование дофамина с образованием норадреналина и, наконец, 4 – образование адреналина путём метилирования норадреналина. В дофаминергических нейронах процесс завершается на втором этапе, а в адренергических проходит полный цикл превращений. В последнем случае дофамин служит метаболическим предшественником медиатора.
Дофаминергические нейроны обнаруживаются в среднем мозге, гипоталамусе, базальных ганглиях, сетчатке глаза. Дофаминовые рецепторы являются метаботропными, а эффект действия дофамина может быть как возбуждающим, так и тормозным. Норадренергические клетки описаны в варолиевом мосте, промежуточном и продолговатом мозге. Самая значительная группа норадренергических нейронов концентрирована в голубом пятне. Синапсы, образуемые катехолергическими клетками находят практически во всех структурах центральной нервной системы, в ганглиях симпатической нервной системы. Катехоламины как медиаторы синтезируются в аксонных терминалях вне везикул, а затем проникают внутрь везикул и там концентрируются. Некоторая часть медиатора находится в аксоплазме. После выброса в синаптическую щель и воздействия на рецепторы, большая часть медиатора – до 80% обратным захватом возвращается в пресинаптическую терминаль, остальная либо рассеивается в межклеточном пространстве, либо инактивируется ферментом катехол-О-метилтрансферазой.
Адренорецепторы постсинаптической мембраны бывают двух видов - альфа- и бета-, разделяющиеся на множество подклассов. Все они относятся к метаботропным. Активация альфа-адренорецепторов сопровождается, в основном, возбуждением постсинаптических структур, а бета-адренорецепторов - преимущественно торможением (исключение касается только сердечной мышцы).
Серотонин (5-гидрокситриптамин) образуется путём гидроксилирования триптофана в синаптических терминалях и депонируется в везикулах. Из мембранной щели серотонин удаляется путем обратного захвата в пресинаптическую терминаль, а рассеянная часть разрушается в тканях моноаминоксидазой, содержащейся в митохондриях. Серотонинергические нейроны описаны в ядрах шва, в среднем мозге, в варолиевом мосту. Аксоны этих нейронов устанавливают контакты с обширными областями мозга – гиппокампом, миндалиной, ядрами гипоталамуса, корой больших полушарий, бледным шаром. Высокая концентрация серотонина обнаружена в эпифизе. Выделяется семь типов серотониновых рецепторов со множеством подтипов. Все они относятся к группе метаботропных. Активация рецепторов приводит к разнообразным эффектам - влияние на синтез цАМФ, активация калиевых и кальциевых каналов, к гиперполяризации мембраны и торможению клетки. Широкое распространение серотонинергических нейронов и синапсов в центральной нервной системе, особенности действия серотонина привели исследователей к формированию понятия «серотонинергическая система». Функции этой системы весьма разнообразны – это регуляция цикла сон-бодрствование, эмоционального поведения, двигательной активности, пищевого поведения и других сложных системных реакций. Во многих случаях серотонин выступает в роли регулятора некоторых иных медиаторов – дофамина, глутамата и др.
Нейропептиды – это олигопептиды, состоящие из 2 – 30 аминокислот, функционируют как медиаторы и комедиаторы или модулируют синаптические процессы. К ним относятся энкефалин, эндорфин и динорфин, играющие определённую роль в активации опиоидных рецепторов в лимбической системе, вегетативной нервной системе, в моторных системах. К другим нейропептидам, выполняющим роль нейромодуляторов, относятся вазоактивный интестинальный полипептид (ВИП), нейропептид Y и др. Большая часть из них были открыты как гормоны и только в последнее время была определена их роль в синаптических процессах. Нейропептиды находят в пресинаптической области, где они участвуют в синтезе медиаторов, и в крупных (тёмных) везикулах. Вместе с основным медиатором нейропептиды выбрасываются в синаптическую щель, модулируя его действие на рецепторы постсинаптической мембраны либо прямо, либо посредством вторичных мессенджеров.
Процесс выброса медиаторов в синаптическую щель наиболее подробно и ранее других описан в нервно-мышечном синапсе. Этому способствовало несколько обстоятельств – доступность визуального контроля подобных соединений (особенно в синапсах беспозвоночных), значительные размеры площади контакта, сравнительно несложный механизм запуска синаптических процессов, детальные сведения о медиаторе – ацетилхолине. Дальнейшие исследования показали, что основные синаптические процессы в мионевральных синапсах весьма сходны с основными процессами и в других синапсах. Проследим процесс выделения медиатора на примере холинергического мионеврального синапса.
Как уже указывалось, везикулы фиксируются на цитоскелете пресинаптической терминали и часть их расположена в области внутренней поверхности пресинаптической мембраны. Последние и представляют собой «оперативный резерв» медиатора для очередного выброса. Потенциал действия, распространяющийся вдоль аксона достигает аксонных терминалей и деполяризует пресинаптическую зону терминали. Во время этой деполяризации в нервном окончании возникает не только входящий ток натрия, как это наблюдается по всей длине аксона, но и включается специфический механизм пресинаптической мембраны, а именно – открытие потенциалзависимых ионных каналов для кальция. Эти каналы открываются на короткое время и ионы кальция устремляются внутрь терминали. Концентрация кальция в пресинаптической зоне возрастает на несколько порядков от 110-7 М в покое до 1·104 М при активации.
Одновременно ионы кальция дополнительно выходят из эндоплазматического ретикулюма. При достижении необходимой концентрации кальция в пресинаптической зоне здесь развивается два процесса (рис.2). Один из них – связывание кальция с белком мембраны везикул, прилегающих к пресинаптической мембране – синаптотагмином, в результате чего везикула раскрывается
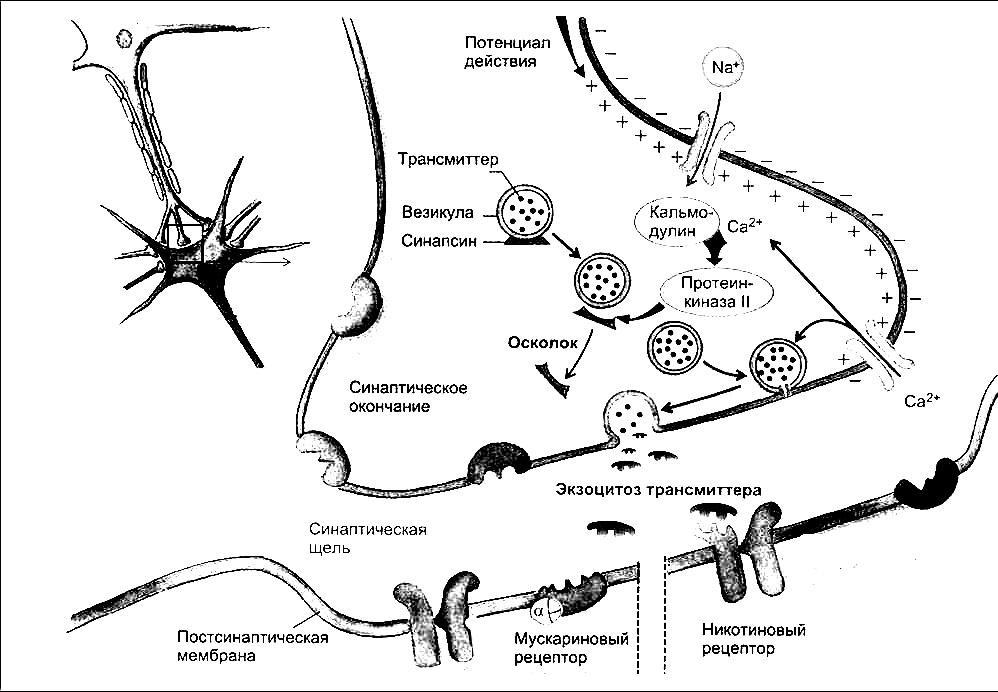
Рис. 2. Механизм освобождения медиатора (Р.Клинке, 2004)
Подробности в тексте
Одновременно происходит слияние мембраны везикулы и пресинаптической мембраны, возникает пора, через которую и осуществляетмя экзоцитоз, т.е. выброс содержимого везикулы (кванта медиатора) в синаптическую щель (рис.3). Кроме того, повышенная концентрация кальция приводит к активации фермента, фосфорилирующего синапсин, и везикулы трансмиттерного резерва освобождаются от цитоскелета, перемещаясь к пресинаптической мембране для осуществления очередного цикла экзоцитоза. Ионы кальция в данном случае выполняют роль вторичного мессенджера, запускающего процесс слияния мембраны везикулы с пресипаптической мембраной.
Предполагается участие в этом также небольших мембранных G-белков. После завершения пресинаптической деполяризации ионы кальция удаляются из пресинаптической зоны посредством переносчика с участием кальциевой
АТФазы, кроме того, включается механизм активного транспорта кальция в мито-
хондрии, так называемая секвестрация кальция митохондриями и связывание кальция внутриклеточными буферными системами.

Рис. 3. Экзоцитоз синаптических везикул. Увеличение в 200000 раз
(Р.Клинке, 2004)
«Время жизни» молекулы ацетилхолина в синаптической щели, по данным разных авторов, составляет от сотен микросекунд до нескольких миллисекунд. Судьба выделившегося ацетилхолина, а также других медиаторов прослежена при их описании (см. выше).
3. Электрические процессы на постсинаптической мембране.
Взаимодействие молекул медиатора с соответствующими рецепторами постсинаптической мембраны приводит, как уже было сказано, к открытию ионных каналов для натрия, калия и хлора. В соответствие с электрохимическим градиентом происходит быстрое движение ионов через мембрану (натрия и хлора внутрь клетки, калия – наружу), что приводит к сдвигу на некоторое время потенциала покоя мембраны на величину порядка 10-30мв. Этот процесс длится в течение 5-100мс и в зависимости от того, какие ионные каналы открыты, в зависимости от подвижности ионов заряд мембраны может либо уменьшаться (мембрана
деполяризуется), либо увеличиваться (мембрана гиперполяризуется) - рис.4).
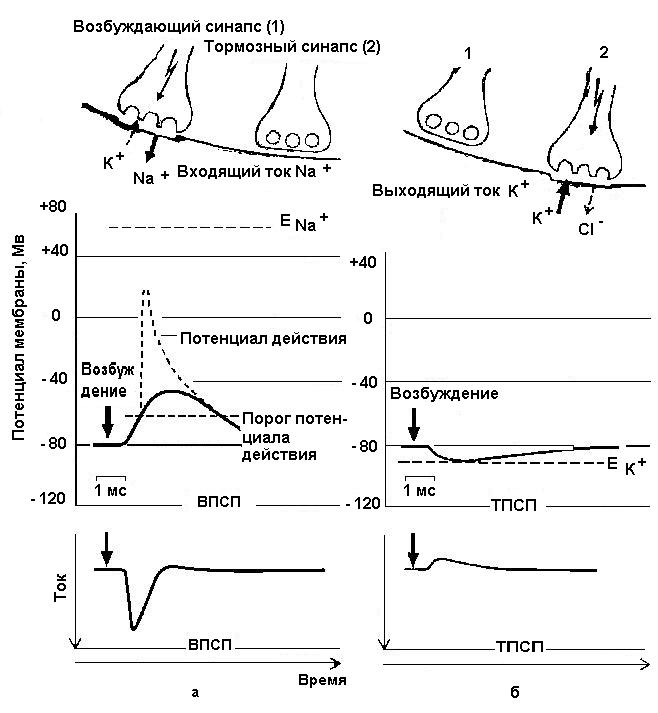
Рис. 4. Формирование постсинаптических потенциалов (Р.Клинке, 2004)
Подробности в тексте
В первом случае возникает возбуждающий постсинаптический потенциал (ВПСП), во втором – тормозный постсинаптический потенциал (ТПСП). Эти потенциалы различаются по амплитуде, среди них выделяют миниатюрные постсинаптические потенциалы (МинПСП), амплитуда которых находится в пределах от десятков мкВ до нескольких мв, а длительность меньше, чем у обычных ПСП.
ВПСП возникает в той зоне постсинаптической мембраны, где действие медиатора привело к открытию ионных каналов для натрия и калия. При этом амплитуда ВПСП прямо зависит от числа выделившихся квантов медиатора, концентрации медиатора в синаптической щели и, следовательно, от количества открытых ионных каналов. Наиболее подробно процесс генерации ВПСП описан для холинергических синапсов. Методами современной микроэлектродной техники и расчетным путём показано, что объём синаптической щели, в котором действует один квант ацетилхолина представляет сферу с диаметром около 1мкм, а площадь постсинаптической мембраны, на которой может быть реализован эффект одного кванта весьма приблизительно оценивается в несколько (до 5-8) мкм2. Учитывая плотность рецепторов на мембране, можно полагать, что для реализации эффекта одного кванта должно быть активировано не менее 2000 ионных каналов.
Следует отметить, что отдельные ВПСП являются локальными потенциалами, не распространяющимися за пределы не только синаптического аппарата, но и за пределы пункта возникновения, быстро угасая по мере удаления от точки возникновения. Амплитуда ВПСП нарастает в течение 2-20 мс (для разных нейронов), после чего начинается её медленный спад (в течение десятков, до 120мс) до исходного уровня заряда мембраны. Подобную несимметричность связывают с несколькими факторами, в частности, с постоянной времени мембраны, остаточным действием медиатора и разной подвижностью ионов, обеспечивающих входящий и выходящие токи. Во всяком случае, длительность ВПСП намного превышает время ионного тока, длящегося в пределах 2-5мс. Показано, что ВПСП в мотонейронах спинного мозга, как и в других клетках, могут развиваться «спонтанно» в случае редкой импульсации в пресинаптическом волокне, так и в результате случайных выбросов медиатора пресинаптическим окончанием. Такие редкие ВПСП носят название элементарных (или миниатюрных) ВПСП, поскольку они , как правило, не достигают пороговых значений для генерации потенциала действия, создавая при этом определённый фон для развития распространяющегося возбуждения в нейроне.
Возбуждающие ПСП возникают как результат перераспределения ионов относительно постсинаптической мембраны. Возникает сильный входящий ток ионов натрия и более слабый выходящий ток ионов калия. В конечном итоге в клетку входит больше положительно заряженных ионов, что снижает уровень поляризации мембраны, возникает локальная деполяризация (ВПСП). Роль отдельных ионов в формировании ВПСП выяснилась с использованием тончайших микроэлектродных исследований аппликацией различных ионов в среду по обе стороны постсинаптической мембраны реального нейрона (в том числе методом patch-clamp) и в модельных опытах на искусственных мембранах (Экклс, Ходжкин, Костюк, Хемил, Сакман, Неер и др.). Сильный ток ионов натрия внутрь клетки связан с тем, что равновесный потенциал для натрия равен +55мв, а потенциал мембраны находится в диапазоне от -60 до -80мв, таким образом открытие ионных каналов обеспечивает движение ионов по электрохимическому градиенту. Равновесный потенциал калия составляет -90мв, что с учётом указанной величины мембранного потенциала создаёт незначительный электрохимический градиент, направленный изнутри клетки наружу. Таким образом, встречные потоки ионов калия могут оказывать лёгкое противодействие деполяризации мембраны, вызванное активным потоком ионов натрия. Развивающаяся деполяризация приводит к снижению электрохимического градиента для натрия и его поток внутрь клетки снижается, ток калия увеличивается, что приводит к затуханию ВПСП.
В зоне синапса локально возникший ВПСП подпороговой силы пассивно (электротонически) может распространяться на всю постсинаптическую мембрану. В случае одновременного возбуждения во многих синапсах проявляется явление суммации, и амплитуда ВПСП может достигать порогового уровня, вовлекая в процесс деполяризации внесинаптические области клеточной мембраны. Как известно, в нейроне наиболее возбудимой зоной мембраны является область аксонного холмика, поэтому именно здесь зарождается нервный импульс, распространяющийся как на тело клетки, так и по аксону – возникает распростроняющееся возбуждение.
Подавление и торможение активности (ТПСП) в нейронах были впервые изучены на мотонейронах спинного мозга Экклсом вначале при внеклеточном, а позднее и при внутриклеточном микроэлектродном отведении потенциалов. Было обнаружено, что торможение мотонейронов мышц-антогонистов при раздражении мышечных афферентов сопровождается гиперполяризацией субсинаптической мембраны нейрона. Исследования выявили подобный сдвиг потенциала в постсинаптических нейронах, на которых оканчиваются аксоны клеток Пуркинье мозжечка, клеток Рэншоу спинного мозга и др. Подобная кратковременная гиперполяризация была названа тормозным постсинаптическим потенциалом (ТПСП). Дальнейшие эксперименты разных авторов показали наличие ТПСП в синапсах практически всех отделов центральной нервной системы и в периферических ганглиях. Среди ТПСП, также. как и для ВПСП, принято выделять миниатюрные ПСП, возникающие при спонтанном выделении медиатора в синаптическую щель. Длительность ТПСП близка к таковой для ВПСП, составляет единицы мс и характеризуется асимметричностью фаз нарастания и спада, представляя как бы зеркальное отражение последнего. Средняя продолжительность восходящей фазы для мотонейронов составляет около 1,5мс, время спада – около 10мс, амплитуда – до 5-8мв. Механизм выделения медиаторов в тормозных синапсах таков же, в основном, как и в возбуждающих. Связывание медиатора с субсинаптическими рецепторами приводит к открытию ионных каналов для хлора и калия, которые по электрохимическому градиенту движутся через постсинаптическую мембрану.
Ионы хлора, несущие отрицательный заряд, проникая внутрь клетки усиливают заряд мембраны, т.е. гиперполяризуют её. Ионы калия, выходя из клетки, оказывают на её заряд то же действие. Показано, что формирование ТПСП различных нейронов может быть связано с действием либо обоих ионов, либо в большей степени одним из них. Наряду с простой диффузией предполагается наличие в постсинаптической мембране специфичных хлор-калиевых насосов, также участвующих в обеспечении ионных токов.
4. Электрические синапсы
Структурно электрические синапсы представляют собой щелевой контакт – т.е. участок тесного прилегания клеточных мембран, промежуток между которыми составляет около 2 нм. Так же, как и для химических синапсов, этот контакт может образовываться между терминалями пресинаптического нейрона с разными отделами постсинаптического нейрона – сомой и отростками. Одна пресинаптическая терминаль может реализовывать как химический, так и электрический способ связи, так что химическая и электрическая передача может сосуществовать в одном синапсе. Определение «пресинаптическая» и «постсинаптическая» мембрана в случае электрического синапса условно, поскольку передача возбуждения в этом случае возможна в обоих направлениях. В данном случае как «пресинаптическая» обозначается мембрана или нейрон, находящийся в активном состоянии по отношению к неактивному. В области контакта во внутренней среде клеток отсутствуют структуры, характерные для химических синапсов – везикулы. заполненные медиатором. Вместе с тем, электрический синапс – это не просто тесное прилегание двух возбудимых мембран, что создаёт условия для взаимодействия электрических полей пре- и постсинаптических нейронов. (рис.5)
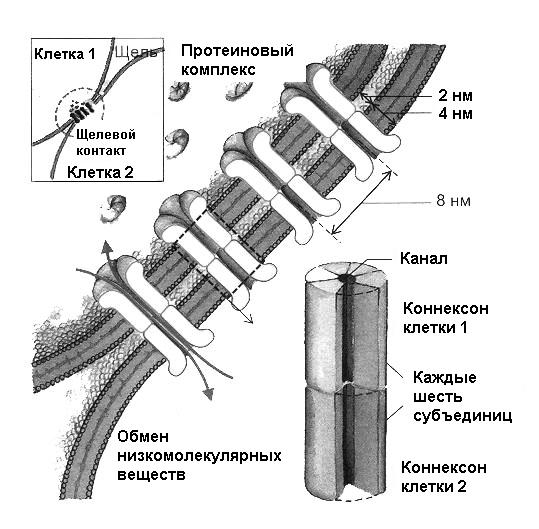
Рис. 5. Структура электрического синапса по Р.Клинке (2004).
Подробности в тексте
В месте сближения мембран имеются специфические белковые комплексы – коннексоны, встроенные в клеточную мембрану и пронизывающие её. Каждый комплексон состоит из шести субъединиц (коннексинов), образующих гексагональную структуру, в центре которой имеется канал диаметром около двух нанометров. Комплексоны каждой из контактирующих мембран плотно прилегают друг к другу, образуя сквозной канал, через который две клетки могут обмениваться ионами и водорастворимыми молекулами с молекулярным весом до 1,2-1,5 КД.
В частности, показано, что эти каналы проходимы для АМФ, АТФ, АДФ, цАМФ, ионов натрия, калия, хлора, сахаров, аминокислот. Открытие и закрытие каналов осуществляется путем изменения геометрии отдельных коннексинов с участием ионов кальция. Вне коннексонов клеточная мембрана по своему строению и функциональным особенностям соответствует характеристикам мембраны сомы и отростков нейрона, т.е. имеющиеся здесь ионные каналы являются потенциалзависимыми. Это во многом определяет механизм работы электрического синапса. Когда потенциал действия в пресинаптическом (активном) волокне достигает места контакта, пресинаптическая мембрана приобретает отрицательный заряд, следовательно, возникает разность потенциалов между деполяризованной пресинаптической и покоящейся постсинаптической мембранами. Положительные ионы по градиенту разности потенциалов устремляются внутрь постсинаптической клетки по открытым каналам комплексонов, а отрицательные – в обратном направлении. Вследствие этого участок постсинаптической мембраны частично деполяризуется, возникает локальный потенциал. В случае достижения порога локальный потенциал преобразуется в полноценный потенциал действия, который распространяется по постсинаптической мембране по законам распространения возбуждения в немиэлинизированном нервном волокне.
В отличие от химических синапсов, коэффициент связи в электрических синапсах близок к 4:1, т.е. в постсинаптической клетке изменения потенциала составляют четверть от такового в пресинаптической (активной) клетке.
Характер и величина коэффициента связи, как показано, зависит от соотношения диаметров контактирующих волокон. Так, если активное волокно тоньше, чем постсинаптическое, то результатом работы синапса будет лишь некоторое изменение потенциала последней, т.е. возникнут небольшие по амплитуде локальные потенциалы. При обратном соотношении размеров волокон достижение критического уровня деполяризации постсинаптической мембраны более вероятно. Однако и в этом случае, учитывая высокий градиент затухания потенциала действия по немиэлинизированнму волокну, эффект работы электрического синапса не может проявляться на большом расстоянии. Понятие синаптической задержки к электрическому синапсу неприменимо, поскольку ток немедленно проводится от одной клетке к другой.
Большинство исследователей склоняется к тому, что роль электрических синапсов в центральной нервной системе позвоночных сводится к повышению возбудимости постсинаптических нейронов, облегчая развитие возбуждения, вызванного действием химических синапсов, играя роль предусилителей для последних. Кроме того считается, что наличие электрической связи между нейронами в группе обеспечивает синхронизацию активности в нервных сетях. У беспозвоночных работа электрических синапсов обеспечивает срочные реакции защитного характера.
Центральные нейроны возбуждаются в результате синаптического действия возбуждающего медиатора, увеличивающего проницаемость хемозависимых каналов постсинаптической мембраны для ионов натрия и калия, что приводит к ее деполяризации. Возникающий при этом деполяризационный возбудительный постсинаптический потенциал (ВПСП) по своему фундаментальному механизму аналогичен локальному ответу. Однако в отличие от последнего, при генерации ВПСП входящий ток ионов натрия обусловлен открытием хемозависимых, а не потенциалзависимых, как при локальном ответе, каналов.
Отсутствие потенциалзависимых натриевых каналов в постсинаптической сома-дендритной мембране нейронов делает невозможным формирование здесь ПД. В соматической мембране потенциалзависимые натриевые каналы расположены лишь в аксонном холмике, где и возникает ПД под влиянием возбуждающего медиатора. Это триггерная зона. ВПСП, возникающие на разных расстояниях от триггерной зоны, распространяются по мембране только посредством местных круговых токов и при этом декрементно (т.е. с прогрессивным падением амплитуды).
Поэтому ПД возникает в аксонном холмике лишь тогда, когда амплитуда распространившегося в эту область ВПСП будет равной или превысит значение критической величины деполяризации мембраны.
ВПСП, в отличие от ПД, подчиняющегося закону «всё или ничего», этому закону не подчиняется. В отношении постсинаптических потенциалов (и возбудительных, и тормозных) возможна суммация и временная, и пространственная, а чаще – пространственно-временная, что и допускает превращение ВПСП в потенциал действия.
Литература
Анохин П. К. Очерки по физиологии функциональных систем. М.: Медицина, 2005.448 с.
Батуев А. С. Нейрофизиология коры головного мозга: Модульный принцип организации. Л., 2009.216 с.
Скок В. И., Шуба М. Ф. Нервномышечная физиология. К.:Вища школа, 2006. 224 с.
ШмидтНиельсен К. Физиология животных: приспособление и среда/Пер. с англ. М.:Мир, 2002.798 с.
Шульговский В. В. Основы нейрофизиологии Учебное пособие. М.:Аспект Пресс, 2000.с. 277
Волкова О. В. Эмбриональный гистогенез и постнатальное развитие органов человека. М., 2003
Костюк П. Г., Преображенский Н. М. Механизмы интеграции висцеральных и соматических афферентных сигналов. Л.: Наука, 2005.222 с.
Общая физиология нервной системы: Руководство по физиологии. Л.:Наука, 2003.254 с.
Физиология человека в 4х томах: Пер. с англ./Под ред. Р.Шмидта и Г. Тевса. Т.3М: Мир, 2006
Шульговский В. В. Физиология центральной нервной системы: Учебник. М.:Издво МГУ, 1997.368 с.
Батуев А. С. Нейрофизиология коры головного мозга: Модульный принцип организации. Л., 2009.216 с.
Куфлер С., Никольс Дж. От нейрона к мозгу. М.:Наука, 2009
Механизмы деятельности мозга человека. Ч.1:Нейрофизиология человека. Л.:Наука, 2008.677 с.
СИНАПТИЧЕСКОЕ ВЗАИМОДЕЙСТВИЕ НЕЙРОНОВ