ФИЗИОЛОГИЯ СЕРДЦА
ФИЗИОЛОГИЯ СЕРДЦА
- Автоматия сердца.
- Возбудимость сердца.
- Сократимость сердца.
- Гемодинамическая функция сердца.
- Регуляция сердечной деятельности.
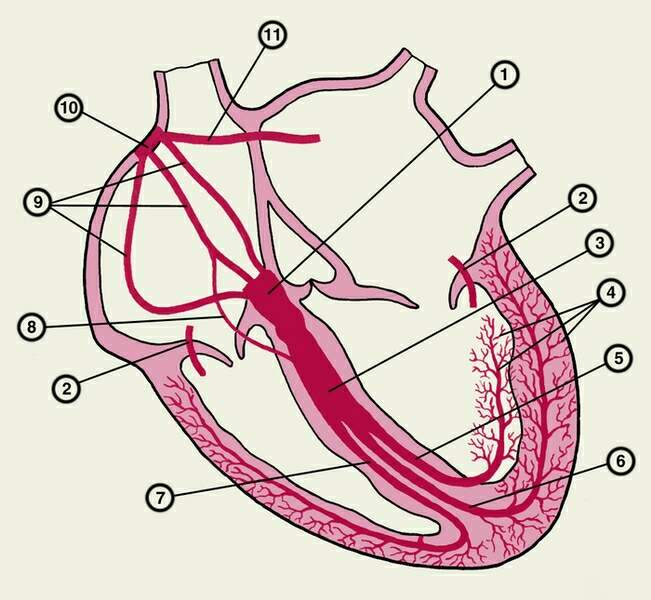
Общие принципы строения. Сердце - полый орган, расположенный в грудной полости. У высших позвоночных сердце состоит из двух половин: левой (системной) и правой (легочной). Их функциональное разделение происходит только после рождения. В каждой половине находятся предсердие и желудочек. Предсердие и желудочек соответствующей половины соединены между собой предсердно-желудочковым (атриовентрикулярным) отверстием, снабженным в левой половине двустворчатым, в правой — трехстворчатым клапанами.
Со стороны желудочков к клапанам прикрепляются сухожильные хорды (нити), что позволяет клапанам открываться только в сторону желудочков. От левого желудочка отходит аорта, которой начинается большой круг кровообращения, а от правого желудочка — легочная артерия, являющаяся началом малого, или легочного, круга кровообращения. Отверстия, которыми начинаются эти сосуды, закрыты полулунными клапанами, открывающимися только во время сокращения желудочков.
Поперечный срез, сделанный через середину обоих желудочков, указывает на значительно большую толщину левого. Такие различия в строении отражают функциональные особенности, т. е. те усилия, которые развиваются каждым из желудочков.
Стенка сердца состоит из трех слоев: эндокарда, миокарда и эпикарда. Основную массу составляет миокард, имеющий наиболее сложное строение. Его образуют отдельные мышечные волокна, каждое из которых является функциональной единицей (кардиомиоцит). Миокард представляет собой цепочку соединенных последовательно (конец в конец) клеток, имеющих общую мембрану. Ткань миокарда, сохраняя сходство с поперечнополосатой скелетной мышечной тканью, существенно отличается от нее рядом признаков, в том числе особой насыщенностью кардиомиоцитов митохондриями, что отражает высокий уровень метаболизма ткани, обладающей непрерывной активностью.
Волокна рабочего миокарда соединяются друг с другом с помощью вставочных дисков — нексусов, обладающих незначительным электрическим сопротивлением. Они служат местом перехода возбуждения между клетками, обеспечивая функциональное единство миокарда.
В миокарде кроме сократительных, или рабочих, волокон имеется особая система мышечных единиц, обладающих способностью к генерации спонтанной ритмической активности, распространению возбуждения ко всем мышечным слоям и координации последовательности сокращения камер сердца. Эти специализированные мышечные волокна составляют проводящую систему сердца.
Основными свойствами сердца являются автоматия, возбудимость и сократимость.
1. Автоматия сердца.
Автоматия сердца – это его способность к ритмическому сокращению без всяких видимых раздражений под влиянием импульсов, возникающих в самом органе.
Изолированное сердце при снабжении его питательным раствором способно сокращаться вне организма продолжительное время. У плода человека первые сокращения сердца возникают на 19-й или 20-й день внутриутробного развития, когда парные закладки сердца сливаются в одну сердечную трубку, все клетки которой способны к самовозбуждению. По мере формирования эмбрионального сердца в его ткани происходит разделение на сократительный миокард и проводящую систему сердца. Способность генерировать автоматический ритм закрепляется за узловой тканью проводящей системы, образующей узлы автоматии — синусно-предсердный и предсердно-желудочковый (так называемые водители ритма сердца, или пейсмекеры).
Так, в области ушка правого предсердия находится ведущий центр автоматизма — синусно-предсердного (синоатриального) узла. От него по рабочим клеткам миокарда и проводя�щим волокнам предсердий возбуждение достигает предсердно-желудочкового (атриовентрикулярного) узла, расположенного в стенке правого предсердия вблизи перегородки между предсердиями и желудочками. Далее возбуждение переходит на миокард желудочков по волокнам пучка Гиса (предсердно-желудочкового пучка) и достигает волокон Пуркинье (сердечных проводящих миоцитов).
В обычных условиях частоту активности миокарда всего сердца в целом определяет синоатриальный узел (пейсмекер первого порядка). При нарушении автоматизма синусно-предсердного узла ритмические сокращения сердца могут продолжаться благодаря импульсам, возникающим в атриовентрикулярном узле. Этот узел является пейсмекером второго порядка. Однако частота и сила сокращений при этом вдвое меньше, чем до нарушений. В случае невозможности передачи возбуждения к желудочкам они начинают сокращаться в ритме пейсмекеров третьего порядка – клеток пучка Гиса и волокон Пуркинье. При повреждении всех водителей ритма сердце останавливается.
Синусно-предсердный узел подчиняет себе все нижележащие образования проводящей системы, навязывая им свой ритм. Поэтому все отдельные части проводящей системы, хотя и имеют собственную активность, начинают работать в едином ритме. Явление, при котором структуры с замедленным ритмом генерации потенциалов усваивают более частый ритм других пейсмекерных участков называют усвоением ритма. Исходя из этого Гаскелл установил Закон градиента автоматизма сердца – у всех позвоночных степень автоматии тем выше, чем ближе расположен участок проводящей системы к синоатриальному узлу.
Теории автоматизма. Существует несколько теорий, объясняющих происхождение автоматизма (нейрогенная, эндогенная и др.). Наиболее популярна теория диастолического поля, в соответствии с которой после очередного возбуждения и сокращения (систолы) сердца в начальную фазу расслаблении (диастолы) в проводящих миоцитах регистрируется мембранный потенциал, равный -90 мВ. В течение диастолы метаболизм сердечной мышцы изменяется, при этом уменьшается проницаемость ее мемебраны для ионов К+ и увеличивается для Na+ и Са2+. В результате проникновения внутрь клетки Na+ и Са2+ и уменьшения скорости выхода из клетки К+ возникает медленная спонтанная диастолическая деполяризация. Когда уровень потенциала уменьшится по сравнению с исходным приблизительно на 2 мВ, наступает резкое увеличение проницаемости мембраны вначале для Na+, а позже для Са2+. В результате этих процессов МП приближается к критическому уровню деполяризации, при достижении которого в клетках синусно-предсердного узла возникает новый ПД. Все остальные отделы сердца подчиняются возникшему ПД — возбуждению, генерируемому в водителе ритма.
2. Возбудимость сердца.
Возбудимость — свойство отвечать на раздражение электрическим возбуждением в виде изменений мембранного потенциала с последующей генерацией ПД. Под действием электрических, химических, термических и других раздражителей сердце способно переходить в состояние возбуждения. В основе процессов возбуждения лежит появление отрицательного электрического потенциала в первоначально возбужденном участке, который затем распространяется на соседние клетки. В итоге кардиомиоциты сокращаются.
Механизмы возникновения сокращения в сердечной мышце принципиально не отличаются от таковых в скелетном мышечном волокне и нервной клетке. Как и в скелетной мышце, при возникновении ПД происходит увеличение проницаемости мембран для ионов кальция. Это вызывает последовательную цепь событий, завершающуюся укорочением составляющих их миофибрилл. Но поскольку сократительные кардиомиоциты (точнее, их эндоплазматическая сеть) имеют ограниченный запас ионов кальция, ведущая роль принадлежит внеклеточному кальцию, который определяет скорость и амп�литуду сокращения.
Ионный механизм ПД волокон сердца позвоночных состоит в быстрой акти�вации деполяризующим стимулом натриевых и натрий-кальциевых каналов при параллельной инактивации части калиевых каналов (это особое свойство К-каналов данного объекта). Последующая инактивация каналов входящих то�ков (Na и Ca-каналов) происходит медленно, активация реполяризующих К-каналов также задержана. Эти процессы обеспечивают так называемую фазу плато в процессе реполяризации на уровне, близком к пику ПД. Такая организация ПД клеток желудоч�ка сердца обеспечивает его значительную продолжительность (250-300 мс, что в 150 раз больше, чем в скелетной мышце).
При нанесении электрических раздражений на работающее сердце в разные фазы его цикла оказывается, что независимо от величины и силы раздражения сердце не ответит, если это раздражение будет нанесено в период систолы, т. е. во время периода абсолютной рефрактерности. Период рефрактерности длится столько же времени, сколько продолжается систола.
С началом расслабления возбудимость сердца начинает восстанавливаться и наступает период относительной рефрактерности. Нанесение в этот момент интенсивного стимула способно вызвать внеочередное сокращение — экстрасистолу. При этом пауза, следующая за экстрасистолой, длится больше времени, чем обычно, это так называемая компенсаторная пауза. После фазы относительной рефрактерности наступает период повышенной возбудимости. По времени он совпадает с диастолическим расслаблением и характеризуется тем, что импульсы небольшой силы могут вызвать сокращение сердца. Период этот непродолжителен, и вскоре наступает восстановление уровня возбудимости.
Длительный абсолютный рефрактерный период, т. е. полная невозбудимость сердечной мышцы, предохраняет ее от быстрого повторного возбуждения до тех пор, пока не закончилась предыдущая волна деполяризации. Тем самым предотвращается нарушение нагнетательной функции сердца. Наличие фазы рефрактерности также препятствует возникновению кругового движения возбуждения по миокарду. В противном случае нарушилось бы ритмическое чередование систолы и диастолы. Длительная абсолютная рефрактерность исключает возможность тетанического сокращения сердца, заставляя работать его в режиме одиночных сокращений.
3. Сократимость сердца.
Сократимость — способность сердца сокращаться, реализуя тем самым насосную функцию.
Несмотря на то, что миокард состоит на большого числа мышечных элементов, он всегда функционально реагирует как единое целое. В отличие от скелетной мышцы миокард не обнаруживает зави�симости между силой раздражения и величиной реакции. На подпороговые раздражения сердце вообще не отвечает, но как только сила раздражения достигает порогового уровня, возникает полное сокращение миокарда. Даль�нейшее нарастание силы раздражающего тока не изменяет величины сокраще�ния. Таким образом, пороговое раздражение является одновременно и макси�мальным (закон «все или ничего»).
Подчинение сердечной мышцы закону «все или ничего» объясняется ее струк�турной организацией. В сердечной мышце отдельные мышечные волокна соеди�нены друг с другом вставочными дисками — нексусами с очень малым электрическим сопротивлением. Поэтому при достижении раз�дражающим импульсом пороговой величины возбуждение распространяется, как по синцитию, и обязательно синхронно охватывает всю мышцу в целом.
Вместе с тем закон «все или ничего» имеет исключения (см. ниже Внутрисердечные механизмы регуляции работы сердца).
4. Гемодинамическая функция сердца.
Работа сердца проявляется последовательными рит�мическими сокращениями предсердий и желудочков, чередующимися с их расслаблениями. Сокращение любого отдела сердца называется систолой, расслабление — диастолой, общий покой — паузой. Систола предсердий происходит на фоне диастолы желудочков, вслед за тем возникает систола желудочков, а предсердия находятся в диастоле. Далее вся мышца сердца приходит в состояние покоя. После паузы наступает новое чередование его работы в том же порядке. Каждое повторение работы сердца и покоя называется одиночным циклом сердечной деятельности.
4.1. Одиночный цикл сердечной деятельности
В норме сердце человека совершает в среднем 70 уд/мин. Это означает, что один сердечный цикл длится 0,8 с.
Систола предсердий. Систола предсердий начинается при распростра�нении возбуждения от синусно-предсердного узла и длится 0,1 с. В процесс сокращения вовлекаются все миокардиоциты — и правого, и (чуть позже) левого пред�сердия. В результате сжимаются устья полых вен, впадающих в предсердия, повышается внутрипредсердное давление. Вся кровь, которая за время диастолы предсердия накопилась в нем, изгоняется в желудочки. Благодаря этому, во-первых, возрастает кровенаполнение желудоч�ков и, во-вторых создается сила, которая вызывает дополнительное растя�жение сократительных кардиомиоцитов желудочков.
Систола желудочков. Систолу желудочков принято делить на два пе�риода — фаза напряжения и фаза изгнания крови. Она занимает 0,33 с. В фазу напряжения повышается давление внутри желудочков, закрываются атриовентрикулярные клапаны. Это происходит в том случае, если давление в желудочках становится чуть выше, чем в предсердиях. При этом происходит быстрое повышение внутрижелудочкового давения, т.к. полулунные клапаны еще не открыты. Из-за несжимаемости крови и неподатливости стенок желудочков в результате продолжающе�гося сокращения миокардиоцитов в полостях желудочков сердца возрастает давление. Когда давление в желудочках становится больше, чем в аорте и легочном стволе - откры�ваются полулунные клапаны, что создает возможность изгнания крови в аорту и легочный ствол.
В остальное время систолы желудочков — 0,25 с — происходит изгна�ние крови. В начале процесс изгнания совершается быстро — давление в выходящих из желудочков сосудах (аорте, легочном стволе) сравнительно небольшое, а в желудочках продолжает нарастать. По мере заполнения аорты и легочного ствола выходящей из желудочков кровью сопротивление выходящему потоку крови увеличивается и фаза быстрого изгнания сменяется фазой медленного изгнания.
Диастола желудочков занимает около 0,47 с, ее разделяют на протодиастолический период, период изометрического расслаб�ления и период наполнения. Протодиастола – это промежуток времени от начала снижения дав�ления внутри желудочков до момента закрытия полулунных клапанов, т.е. до того момента, когда давление в желудочках станет меньше давления в аорте и легочном стволе. Далее давление в желудочках продолжает очень быстро падать. Как только оно снижается почти до нуля, открываются атриовентрикулярные клапаны и желудочки наполняются кровью, которая накопи�лась в предсердиях.
Наполнение кровью желудочков начи�нается с момента открытия атриовентрикулярных клапанов: вся кровь (около 33 мл) в фазу быстрого наполнения устремляется в желудочки. Затем наступает фаза медленного пассивного наполнения; в этот период кровь протека�ет «транзитом» сразу из вен через предсердие в желудочки. В завершение наступает систола предсердий, которая за 0,1 с «выжимает» дополнительно около 40 мл крови в желудочки. Эту фазу называют пресистолической.
Итак, длительность систолы предсердий составляет 0,1 с, длительность диастолы — 0,7 с, у желудочков соответственно 0,33 и 0,47 с. Эти цифры указывают на то, что 40 % времени миокардиоциты желудочков находятся в активном состоянии и 60 % — «отдыхают».
При учащении сердечной дея�тельности, например во время мышечной работы, при эмоциональном на�пряжении длительность сердечного цикла укорачивается прежде всего за счет сокращения времени общей паузы. Дальнейшее увеличение нагрузки приводит к укорочению продолжительности систолы.
4.2. Тоны сердца
Выслушивание (аускультация) стетофонендоскопом левой половины грудной клетки позволяет услышать два тона сердца: I тон и II тон. I тон связан с закрытием АВ-клапанов в начале систолы, II — с закрытием полулунных клапанов аорты и лёгочной артерии в конце систолы. Причина возникновения тонов сердца — вибрация напряжённых клапанов тотчас после закрытия совместно с вибрацией прилежащих сосудов, стенки сердца и крупных сосудов в области сердца.
· I тон образуется при сильном и быстром сокращении желудочков, в процессе которого давление в полости желудочков резко увеличивается, что приводит к закрытию и дальнейшему прогибанию клапанов в сторону предсердий до тех пор, пока сухожильные хорды клапанов внезапно не остановят это прогибание. Эластическая упругость сухожильных хорд и клапанов создаёт обратную ударную волну крови, направленную соответственно в каждый желудочек. Это приводит кровь, стенку желудочков, упругие клапаны в состояние вибрации и порождает вибрирующую турбулентность крови. Вибрация через прилежащие ткани достигает стенки грудной клетки и воспринимается в виде звука, слышимого с помощью стетофонендоскопа.
· II тон — результат быстрого закрытия полулунных клапанов. В момент закрытия они прогибаются в направлении полости желудочков. Их эластическая отдача толкает кровь в артерии, вызывая короткий период реверберации крови между стенкой артерий и полулунными клапанами. Вибрация стенки артерий передаётся вдоль артерий. Вибрация сосудов или желудочка, достигающая стенки грудной клетки, воспринимается в виде звука.
Продолжительность I тона составляет 0,14 с, II — 0,11 с. II тон сердца имеет более высокую частоту, чем I. Помимо I и II тонов, иногда можно выслушать дополнительные тоны сердца — III и IV, в подавляющем большинстве случаев отражающие наличие сердечной патологии.
4.3. Гемодинамические показатели
Циркуляцию крови по сердечно-сосудистой системе обеспечивает насосная функция сердца. Сердце сокращается в течение жизни человека до 4 млрд. раз, выбрасывая в аорту и способствуя поступлению в органы и ткани до 200 млн л крови. В физиологических условиях сердечный выброс составляет от 3 до 30 л/мин. При этом кровоток в различных органах (в зависимости от напряженности их функционирования) варьирует, увеличиваясь приблизительно вдвое.
При каждом сокращении желудочков в аорту и легочный ствол выталкивается кровь, заполнившая желудочки во время их диастолы, так называемый ударный, или систолический, объем (СО). У человека в состоянии покоя СО равен 50—70 мл, во время мышечной работы он возрастает до 150—180 мл за счет усиления мощности сокращения сердечной мышцы.
Минутным объемом крови (МОК) называется количество крови, выбра�сываемое одним желудочком за 1 мин, МОК равен СО, умноженному на число сокращений сердца (ЧСС) в 1 мин. Например, если СО равен 60 мл, а ЧСС в 1 мин равно 72, то МОК составит 60-72=4,32 л. У разных людей МОК колеблется от 3 до 5 л. При напряженной физической работе сердце здорового тренированного человека может выбросить за 1 мин в аорту до 25—30 л крови. Такой высокий МОК достигается как за счет увеличения СО, так и за счет возрастания ЧСС в 1 мин.
5. Регуляция сердечной деятельности.
Регуляция сердечной деятельности подчинена интересам целого орга�низма в каждый определенный момент времени. Для удобства ана�лиза конкретных физиологических механизмов включения сердца в ту или иную деятельность целого организма выделяют несколько видов регуляции.
За счет регуляторных влияний осуществляются:
• урежение или учащение сердечных сокращений (отрицательный и положительный хронотропный эффект);
• изменение возбудимости сердечной мышцы (батмотропный эффект);
• изменение силы сокращений (инотропный эффект);
• изменение проводимости (дромотропный эффект);
• изменение тонуса сердечной мышцы (тонотропный эффект).
5.1. Внутрисердечные механизмы регуляции. К ним относят внутриклеточные механизмы, регуляцию межклеточных взаимодействий и собственно внутрисердечные нервные механизмы.
5.1.1. Внутриклеточная регуляция. Имеет в своей основе гетерометрический и гомеометрический механизмы, а также эффекты феномена Анрепа и Лестницы Боудича.
Гетерометрический механизм основан на гемодинамических процессах, связанных с изменением длины кардиомиоцитов. В основе гемодинамической регуляции силы сердечных сокращений лежит закон Франка—Старлинга: чем больше крови притекает к сердцу во время диастолы, тем сильнее растягиваются волокна сердечной мышцы и тем сильнее оно сокращается при следующей систоле. Механизм этого явления объясняют двумя причинами:
сократительный кардиомиоцит состоит из двух элементов — собст�венно сократительного и эластического. Сократительный элемент в возбужденном состоянии способен сокращаться, а последовательно соединенный с ним эластический элемент действует как обычная пружина.
во время диастолы увеличивается площадь контакта между мито�хондриями и миофибриллами, вследствие чего возрастают интен�сивность диффузии АТФ из митохондрий в миофибриллы и энерге�тическое обеспечение сократительного аппарата.
В целостном организме дей�ствие закона сердца ограничено рядом других условий.
Гомеометрический (метаболический) механизм не связан с изменением длины саркомера и основан на непосредственном действии биологически активных веществ. Метаболическая регуляция заключается в способности кардиомиоцитов при выполнении ими специфической сократительной функции синтезировать различные белки в соответствии с уровнем их разрушения. Особенность кардиомиоцитов заключается в цикличности их обменных процессов, связанных с ритмом сердечной деятельности. Наиболее быстрый распад АТФ и гликогена происходит в момент систолы, ресинтез и восстановление уровня этих веществ успевает полностью восстановиться за время диастолы. Поэтому в чрезвычайных условиях при усиленной работе сердца одним из компенсаторных механизмов, адаптирующих деятельность сердца к воздействиям, является удлинение фазы диастолы.
Кардиомиоциты способны также адсорбировать из циркулирующей крови различные БАВ, Так, адсорбированные из крови катехоламины (адреналин и норадреналин) увеличивают вход Са2+ в клетку в момент развития потенциала действия, вызывая тем самым усиление сердечных со�кращений.
Существует также явление, получившее название феномен Анрепа: при повышении давления в аорте или легочном стволе сила сердечных сокращений автоматически возрастает, обеспечивая тем самым возможность выброса того же объема крови, что и при исходной величине артериального давления, т.е. чем больше противонагрузка, тем больше сила сокращений. Механизмы, лежащие в основе феномена Анрепа, до сих пор не раскрыты. Предполагают, что с увеличением противонагрузки растет концентрация Са2+ в межфибриллярном пространстве и поэтому возрастает сила сердечных сокращений.
Другим проявлением гомеометрической регуляции является лест�ница Боудича. Раздражая электрическим током полоску сердца ля�гушки, утратившую способность к автоматизму, автор обнаружил, что каждое последующее сокращение в ответ на стимулы одной амплитуды выше по амплитуде предыдущего — и так до некоторого предела (внешне это напоминало лестницу). В настоящее время установлено, что чем чаще сердце сокращается, тем (до определенного предела) выше сила его со�кращений и наоборот. В основе этого явления лежит повышение уровня Са2+ в межфибриллярном пространстве при увеличении частоты сокра�щений — он не успевает полностью «откачиваться» из наружной среды, а уже появляется новый поток Са2+ из эндоплазматической сети, что и создает более высокий фон Са2+, чем при редком ритме сокращений сердца.
5.1.2. Межклеточная регуляция.
Связана с работой нексусов, обеспечивающих транспорт необходимы веществ, соединение миофибрилл, переход возбуждения с клетки на клетку. Межклеточная регуляция включает также взаимодействие кардиомиоцитов с соединительнотканными клетками, составляющими строму сердечной мышцы и источник пополнения необходимых органических соединений.
5.1.3. Рефлекторная внутрисердечная регуляция.
Осуществляется интракардиальными периферическими рефлексами. Интракардиальные рефлекторные дуги включают афферентные интрамуральные нейроны, дендриты которых образуют рецепторы растяжения миокарда и коронарных сосудов, а также эфферентные нейроны, аксоны которых иннервируют миокард и гладкую мускулатуру коронарных сосудов.
Скопления тел эфферентных нейро�нов представляют собой интрамуральные сердечные ганглии, их составляют не только холинергические, но и адренергические клетки, причем последние обладают большей возбудимостью по сравнению с холинергическими. Эффекты раздражения эфферентных интрамуральных нейронов могут быть противоположными в зависимости от степени кровенаполнения сердца. При чрезмерном кровенаполнении афферентация от рецепторов сердца ведет к возбуждению адренергических нейронов, а выделяемый ими медиатop норадреналин оказывает стимулирующее влияние на сердце (сердечный выброс увеличиается). При слабом наполнении камер сердца кровью и интенсивном раздражении рецепторов возбуждаются холинергические эфферентные нейроны, оказывая тормозное влияние на работу сердца.
Внутрисердечные рефлекторные дуги — часть метасимпатической нервной системы. Эфферентные нейроны являются также общими с дугой классического парасимпатического рефлекса.
5.2. Внесердечная регуляция
Внесердечная нервная регуляция сердечной деятельности осуществля�ется нервным и гуморальным путем.
Нервная регуляция осуществляется по рефлекторному механизму, где можно выделить афферентное звено (рецепторы), нервные центры и эффекторы.
Рецепторные образования расположены в рефлексогенных зонах кровеносных сосудов: дуге аорты, сонном синусе, верхней полой вене и правом предсердии. Кроме того, рефлекторные влияния на работу сердца осуществляются с механорецепторов, расположенных в брыжейке, кишеч�нике, желудке.
Торможение сердечной деятельности рефлекторным путем иллюстрируют следующие опыты:
Глазосердечный (Даньини–Ашнера) рефлекс. Надавливание на глазные яблоки вызывает раздражение рецепторов тройничного нерва; импульсы передаются по этому нерву в средний мозг, далее по вставочному нейрону (возможно, по волокнам ретикулярной субстанции) направляются к ядрам блуждающих нервов, от которых импульсы доходят до нервных сплетений сердца. Кроме того, надавливание на глаза приводит к изменению внутриглазного давления и раздражению барорецепторов полостей глаз, импульсы от которых передаются в ствол мозга к ядрам блуждающих нервов.
Синокаротидный рефлекс (Чермака). В области каротидного синуса (область бифуркации сонной артерии) расположена рецепторная зона, содержащая барорецепторы, реагирующие на изменение давления. При повышении давления в сосуде в результате надавливания в рецепторах возникают импульсы, которые по чувствительным волокнам поступают в продолговатый мозг. В результате увеличивается активность ядер блуждающих нервов, снижается частота сердечных сокращений.
Солярный рефлекс (Гольтца, Тома-Ру). При надавливании на эпигастральную зону в области солнечного сплетения происходит раздражение многочисленных его рецепторов, афферентные импульсы от которых по чревным нервам достигают спинного мозга, а затем ядер блуждающих нервов. Далее по эфферентным волокнам импульсы достигают сердца, в результате чего частота сердцебиений замедляется.
Дыхательно-сердечный рефлекс (Геринга). Терминальные и дыхательные бронхиолы легких содержат механорецепторы блуждающих нервов. При значительном их растяжении (глубокое форсированное дыхание) афферентация от легких в центры продолговатого мозга резко усиливается, что, вследствие повышения тонуса ядер блуждающих нервов, вызывает замедление работы сердца.
Существуют рефлекторные влияния и с других рецепторов организма человека. Всякого рода болевые, температурные, световые и дру�гие раздражители в той или иной степени изменяют сердечную деятельность.
Нервные центры и эффекторы внесердечной нервной регуляции принадлежат автономной нервной системе.
Парасимпатическая иннервация представлена ветвями блуждающих нервов, отходящими от общих стволов этих нервов в верхней части грудной полости. Преганглионарные волокна заканчиваются на интрамуральных ганглиях. Если на шее живот�ного перерезать один блуждающий нерв, а его периферический конец, идущий к сердцу, раздражать электрическим током, то при слабом раздраже�нии возникает урежение сокращений сердца и ослабевает их сила. Если раздражение усилить, может произойти полная остановка сердца во время диастолы желудочков. Действие ацетилхолина в первую очередь основано на повышении мембранной проницаемости для иона К+, что вызывает гиперполяризацию. В то же время еще в 30-е годы XX в. В. Кеннон установил, что при слабом раздражении блуждающего нерва может наблюдаться усиление сердечной деятельности. В настоящее время в интрамуральных ганглиях наряду с холинергическими обна�ружены адренергические нейроны, которые и обеспечивают сердечную деятельность и иннервируют главным образом предсердия.
Эффекты парасимпатической иннервации: сила сокращений предсердий уменьшается — отрицательный инотропный эффект, ЧСС снижается — отрицательный хронотропный эффект, предсердно-желудочковая задержка проведения увеличивается — отрицательный дромотропный эффект.
Симпатическая иннервация. Ветви симпатических нервов берут начало от грудного отдела спинного мозга и прерываются в верхнем, среднем шейных и звездчатых ганглиях. Постганглионарные волокна иннервируют весь миокард, но в основном предсердия. Л. Цион и И.П. Павлов обнаружили, что раздражение симпатических нервов оказывает влияние, противоположное действию блуждающих нервов: увеличиваются частота и сила сердечных сокращений, улучшается проводимость и повышается возбудимость.
Указанные процессы обусловлены взаимодействием медиатора норадреналина преимущественно с -адренорецепторами миокарда. Его действие связано с ростом мембранной проницаемости для ионов кальция, что сопровождается повышением степени сопряженности возбуждения и сокращения миокарда. Кроме того, адреналин способствует расщеплению гликогена и образованию глюкозы, служащей источником энергии для сокращения миокарда. Норадреналин разрушается значительно медленнее, чем ацетилхолин, поэтому его взаимодействие с адренорецепторами сердечных клеток, как и раздражение симпатических волокон, сопровождается более продолжительным эффектом. При чрез�мерном возбуждении сердечных симпатических нервов возбудимость мио�карда может возрасти настолько, что появляются эктопические очаги воз�буждения и нарушения ритма.
Эффекты симпатической иннервации: сила сокращений предсердий и желудочков увеличивается — положительный инотропный эффект, ЧСС возрастает — положительный хронотропный эффект, интервал между сокращениями предсердий и желудочков (т.е. задержка проведения в АВ-соединении) укорачивается — положительный дромотропный эффект.
Кроме того, И.П. Павлов открыл в составе симпатического нерва особую веточку, раздражение которой усиливает сокращение сердечной мышцы без учащения ритма. Этот нерв получил название усиливающего нерва Павлова. Влияние усиливающего нерва на деятельность сердца объясняется усилением процессов обмена веществ, их улучшением в тканях сердца, т.е. положительным трофическим влиянием.
Антагонизм и синергизм парасимпатических и симпатических влияний на сердце. В опытах на животных обнаружено, что перерезка блуждающих нервов вызывает учащение сердечной деятельности вследствие выпадения их тормозного влияния. Наоборот, перерезка симпатических ветвей приво�дит к некоторому (значительно меньшему) замедлению сердечного ритма. Эти опыты доказывают, что сердце находится под двойным влиянием воз�буждений, постоянно идущих по симпатическим и блуждающим нервам. Причиной указанных постоянных влияний служит некоторое умеренное возбуждение в нервных клетках ядер этих нервов, что получило название центрального тонуса. Блуждающий и симпатический нервы оказывают вли�яние на работу сердца обычно одновременно (взаимосвязанное влияние) с некоторым преобладанием тормозных вагусных эффектов.
При этом парасимпатические влияния могут вызывать и положитель�ные, и отрицательные эффекты на сердце, осуществляя текущую «опера�тивную» регуляцию. Симпатические же влияния всегда однозначны и ведут к мобилизации его функций. Следовательно, в условиях деятельности це�лостного организма можно говорить не только об относительном антаго�низме, но и о синергизме симпатических и парасимпатических влияний на сердце, что обеспечивает наилучшее функционирование сердца в различ�ных условиях.
Гипоталамическая регуляция.
Ядра гипоталамуса являются следующей ступенью в иерархии нервных центров, регулирующих сердечную деятельность. У млекопитающих посредством прямых связей гипоталамус соединяется с ядами блуждающих нервов в продолговатом мозге и боковыми рогами спинного мозга, т.е. с центрами симпатической и парасимпатической иннервации сердца. Непосредственное участие в регуляции работы сердца принимают паравентрикулярное, вентромедиальное и преоптические ядра, сосцевидные тела.
Гипоталамус – интегративный центр, который может изменять параметры сердечной деятельности для обеспечения текущих потребностей организма и всех его систем при различных поведенческих реакциях.
Корковая регуляция.
Кора головного мозга является высшей ступенью в иерархии механизмов целенаправленного управления деятельностью сердца. Они проявляются особенно отчетливо при раздра�жении моторной и премоторной зон коры, поясной извилины, орбитальной поверхности лобных долей, передней части височной доли. В то же время электрическая стимуляция практически любого участка коры мозга вызывает ответы сер�дечно-сосудистой системы. Возможность произвольной корковой регуляции подтверждается экспериментами с БОС.
Кора головного мозга является органом психической деятельности, обеспе�чивающим целостные приспособительные реакции организма. Многочислен�ные наблюдения показывают, что работа сердца изменяется при действии усло�вий, влияющих на деятельность коры мозга: боли, страха, радости, тревоги, ярости и т. п. Например, у спортсменов наблюдается так называемое предстар�товое состояние, проявляющееся учащением сердцебиения. Оно связано с активацией корой мозга гипоталамо-гипофизарной системы и надпочечников, выделяющих адреналин в кровь. Сигналы, непосредственно предвещающие возникновение этих ситуаций или возможность их наступления, способны по механизму условного рефлекса вызвать перестройку функций сердца, чтобы обеспечить предстоящую деятельность организма.
Корковая регуляция осуществляется посредством гипоталамических влияний и изменения активности лимбической системы, которые, в свою очередь, реализуют свои эффекты через автономную нервную систему.
Гуморальная регуляция
Осуществляется за счет химических веществ, находящихся в крови, гормонов, различных ионов (Са2+, К+, Mg2+) и биологически активных веществ различной химической природы, в том числе регуляторных пептидов. Например, гормон мозгового слоя надпочечников адреналин вызывает учащение сердечной деятельности и увеличивает силу сердечных сокращений. Сходное влияние на сердце оказывают ионы кальция. Ионы калия оказывают на сердце влияние, подобное действию блуждающих нервов (ацетилхолина): избыток калия в крови вызывает урежение ритма сердца, ослабляет силу сокращения, угнетает проводимость и возбудимость. Угнетающий эффект связан с тем, что избыток калия в межклеточной жидкости вызывает уменьшение или даже исчезновение концентрационного калиевого градиента. В результате уменьшается или прекращается калиевый отток из клетки, уменьшаются мембранный потенциал и возбудимость мембраны вплоть до полной невозбудимости.
Из других биологически активных веществ, содержащихся в крови, следует отметить действие глюкагона, ангиотензина II, вызывающих положительный инотропный эффект (увеличение силы). Гормоны щитовидной железы (тироксин и трийодтиронин) усиливают сердечную деятельность, способст�вуя более частой генерации импульсов, увеличению силы сердечных сокращений и усилению транспорта кальция. Простагландины увеличивают силу и частоту сердечных сокращений за счет усиления коронарного кровотока и метаболизма сердечной мышцы. Кинины (брадикинин, лизилбрадикинин) также вызывают учащение сердечной деятельности в результате высвобождения катехоламинов.
Также известно, что мышечные клетки предсердий секретируют специфический гистогормон – натрийуретический пептид. Его выделение стимулируется растяжением предсердий, возникающим в результате притока крови, изменением в ней уровня натрия, а также симпатическими и парасимпатическими влияниями и содержанием в крови вазопрессина. Натрийуретический гормон подавляет реабсорбцию ионов натрия и хлора в канальцах нефрона, снижает реабсорбцию воды, увеличивает клубочковую фильтрацию, расслабляет гладкие мышцы стенки пищеварительного тракта и мелких сосудов. Все это приводит к снижению системного артериального давления и опосредованно уменьшают кровонаполнение предсердий.
Проводящая система сердца: строение и частота разрядов
ПД клеток синоатриального узла 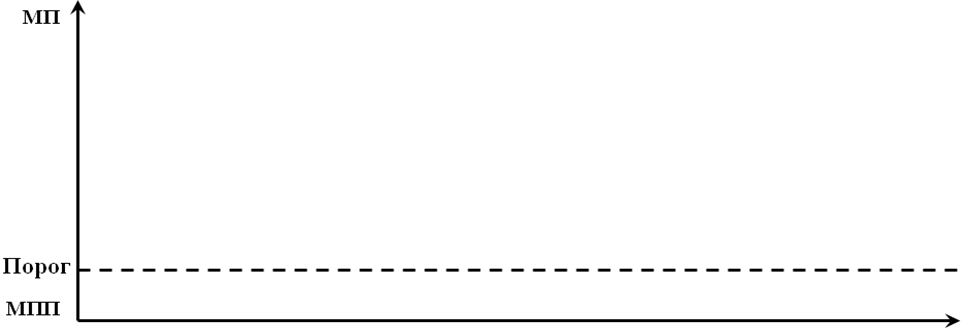
ФИЗИОЛОГИЯ СЕРДЦА