Физиология дыхания
Физиология дыхания
1. Общая характеристика процесса дыхания.
2. Дыхательный акт.
2.1. Отрицательное давление плевральной полости.
2.2. Механизм вдоха и выдоха.
2.3. Паттерны дыхания.
2.4. Характеристика легочной вентиляции.
3. Газообмен в легких
4. Транспорт газов кровью
4.1. Транспорт кислорода
4.2. Транспорт двуокиси углерода.
5. Транспорт кислорода и углекислого газа в тканях.
6. Регуляция дыхания.
7. Особенности дыхания в различных условиях
Морфо-функциональная характеристика дыхательной системы.
Дыхательная система состоит она из дыхательных (воздухоносных) путей и парных органов газообмена (легких). В дыхательных путях вдыхаемый воздух согревается (охлаждается), увлажняется, очищается от инородных частиц. В легких происходит газообмен между вдыхаемым воздухом и кровью. Из альвеол легких путем диффузии в кровь легочных капилляров поступает кислород, а в обратном направлении — из крови в альвеолы выходит углекислый газ.
Полость носа образована лицевыми костями, хрящами и разделена на две симметричные половины. Она выполняет двойную функцию — она является началом дыхательных путей и одновременно органом обоняния. Вдыхаемый воздух, проходя через полость носа, очищается oт чужеродных частиц, согревается, увлажняется. Находящиеся во вдыхаемом воздухе пахучие вещества раздражают обонятельные рецепторы, в которых возникают обонятельные нервные импульсы. В носовую полость открываются воздухоносные придаточные полости (пазухи) носа: лобная, верхнечелюстная (гайморова), клиновидная и решетчатые лабиринты. Воздухоносные придаточные пазухи не только уменьшают вес (массу) черепа, но и служат резонаторами звуков, голоса. В полость носа открывается носослезный канал, по которому выводится избыток слезной жидкости.
Из полости носа вдыхаемый воздух через хоаны попадает в носоглотку. Затем, пройдя через ротовую часть глотки, где пере�секаются дыхательный и пищеварительный пути, воздух попадает в гортань.
Гортань, являющаяся и органом дыхания, и органом голосообразования. Скелетом гортани служат хрящи, соединенные друг с другом при помощи суставов и связок. В голосовых складках гортанной полости расположены покрытые слизистой оболочкой голосовые связки. Узкое пространство между правой и левой голосовыми складками носит название голосовой щели. При про�хождении выдыхаемого воздуха через голосовую щель голосовые связки колеблются, вибрируют и воспроизводят звуки. Голосовые связки образуют только звук, сила и высота кото�рого зависят от натяжения голосовых связок и скорости прохож�дения воздушной струи через голосовую щель. В формировании членораздельной речи участвуют губы, язык, зубы, полость рта и полость носа с ее придаточными пазухами.
Трахея имеет скелет в виде 16-20 хрящевых полуколец, не замкнутых сзади и соединенных друг с другом короткими кольцевыми связками. Задняя стенка трахеи, прилежащая к пищеводу, где нет хрящей, перепончатая. Она построена из соединительной ткани и гладкомышечных пучков. Слизистая оболочка трахеи по�крыта мерцательным эпителием, содержит много желез и лимфоидных узелков.
На уровне V грудного позвонка трахея делится на правый и левый главные бронхи (бифуркация трахеи), каждый из которых направляется к воротам легкого. Многократное (до 23 раз) разветвле�ние бронхов в легком называют бронхиальным деревом.
Правое и левое легкие располагаются в грудной полости, спра�ва и слева от сердца и крупных кровеносных сосудов.
Каждое лег�кое покрыто серозной оболочкой — плеврой. Плевра имеет два листка - наружный и внутренний, между которыми замкнута заполненная небольшим количеством жидкости плевральная полость. Она снижает трение легкого при дыхании, и имеет решающее значение в механизме вдоха.
Структурно-функциональной единицей легкого является ацинус, куда входит одна концевая бронхиола с ее разветвлениями (дыхательными бронхиолами, аль�веолярными ходами и альвеолами). В альвеолах ацинуса происходит газообмен между протекающей по капиллярам кро�вью и воздухом, поступающим в легкие.
В обоих легких человека имеется 600-700 млн альвеол, дыхательная поверхность которых составляет от 40 м2 при выдохе и до 120 м2 при вдохе. Легкие весят 500-600 г, их плотность с момента первого вдоха меньше плотности воды. Ткань, образующая легкие, имеет розовый оттенок, однако они темнеют (от сероватого до темно-бурого) и тем больше, чем более загрязненным воздухом дышит человек.
Функции дыхательной системы: газообмен, теплорегуляция, голосообразование, обоняние, выделение, защитная функция.
1. Общая характеристика дыхания.
Дыхание – совокупность процессов, обеспечивающих поступление в организм кислорода, использование кислорода в биологическом окислении и удаление из него углекислого газа. Это позволяет организму поддерживать дыхательный гомеостаз — состояние, характеризующееся оп�тимальным для жизнедеятельности относительным постоянством газового состава крови и тканей.
Само по себе поступление кислорода в организм и удаление из него двуокиси углерода еще не может обеспечить поддержания оптимального для метаболизма тканевого обмена. Постоянно меняющиеся режимы дея�тельности организма (мышечная деятельность, эмоциональные реакции и др.), а также изменяющиеся состав атмосферного воздуха, атмосферное давление и т. д. требуют от организма способности под�держивать кислородно-углекислотный баланс организма в различных условиях и восполнять возникаю�щую газовую потребность
Дыхание осуществляется в несколько стадий.
Первая стадия — легочное, или внешнее, дыхание — связана с поступ�лением и удалением воздуха из легких, т.е. со вдохом (инспирацией) и выдохом (экспирацией). Во время вдоха в легкие поступает насыщенный кислородом атмосферный воздух, при выдохе в атмосферу возвращается воздух, обед�ненный О2 и обогащенный СО2.
Вторая стадия — газообмен в легких между альвеолярным воздухом и легочными капиллярами.
Третья стадия — транспорт газов кровью: кислород — из легких к тка�ням, двуокись углерода — из тканей в легкие.
Четвертая стадия — тканевое (внутреннее) дыхание — метаболические процессы утилизации кислорода клетками и образование двуокиси углерода. Эта стадия изучается в курсах биохимии и молекулярной биологии.
В процессе дыхания важен состав вдыхаемого воздуха:
- Атмосферное давление - это полное газа давление в атмосфере, которое возникает из-за веса атмосферы Земли. Давление газа традиционно сообщается на основе высоты столба жидкости, например, мм рт. ст. Атмосферное давление = 760 мм рт. ст. на уровне моря, оно уменьшается на больших высотах и увеличивается на больших глубинах.
- Количество конкретного газа может быть описано как парциальное давление, представляющее долю от общего давления. Парциальное давление - это часть общего давления смеси газов, приходящаяся на отдельный газ (если бы он занимал весь объем смеси).
- Атмосферный газ состоит из приблизительно 21% O2 и 78% N2 и очень небольшого количества других газов.
2. Дыхательный акт.
2.1. Отрицательное давление плевральной полости.
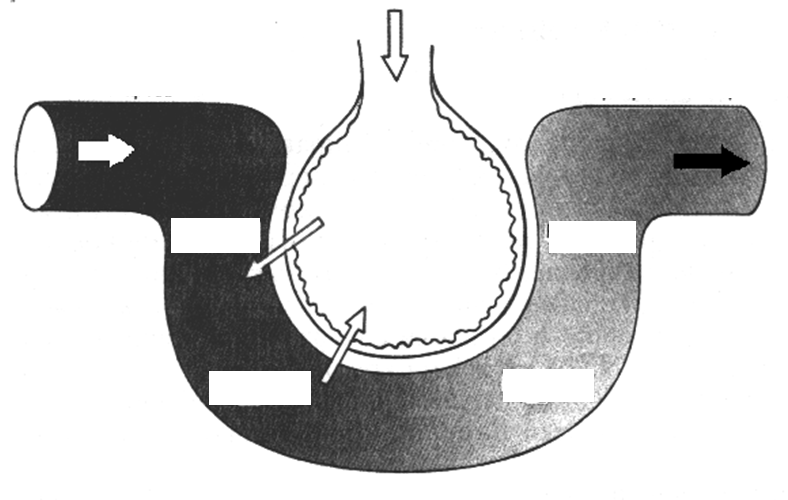
С момента рождения каждого млекопитающего легкие всегда находятся в более или менее растянутом состоянии. Это объясняется отрицательным давлением в плевральной полости, окружающей легкие (на 3-4 мм рт.ст. ниже атмосферного), которая полностью изолирована от атмо�сферного воздуха.
Благодаря наличию отрицательного давления в плевральной полости в обычных условиях спадания легких не происходит. В спавшемся состоянии легкие находятся только у плода до рождения и первого вдоха, а также в случае, если в плевральную полость войдет воздух или газ, - наступает пневматоракс. При одностороннем пневмотораксе дыхание только одним легким на неповрежденной стороне может обеспечить дыхательную потребность при отсутствии физической нагрузки. Двусторонний пневмоторакс делает не�возможным естественное дыхание, в этом случае единственным способом сохранения жизни является искусственное дыхание.
Растянутые легкие стремят�ся сжаться благодаря собственной эластической тяге и поверхностному натя�жению в альвеолах:
- Эластическая тяга обусловлена эластическими свойствами тканей легких и грудной клетки.
- Поверхностное натяжение в альвеолах формируется покрывающими их изнутри жидкостями. Оно существенно снижается поверхностно-активными веществами – сурфактантами. Они позволяют альвеолам оставаться в расправленном (не спавшемся) состоянии при уменьшении объема (во время выдоха) Сурфактант помогает лёгким усваивать кислород. Мода на безжировое питание приводит к возникновению гипоксий у людей, которые не употребляют в пищу качественные жиры (сурфактант примерно на 90% состоит из жиров).
2.2. Механизм вдоха и выдоха.
Вдох и выдох обеспечиваются дыхательными экскурсиями (движениями) грудной клетки и диафрагмы. За организацию этих движений отвечает дыхательный центр продолговатого мозга, который передает к ним импульсы через спинальные центры диафрагмальной и межреберных мышц.
Мышцы, осуществляющие дыхательный акт, подразделяют на:
1) инспираторные (вдоха) - диафрагма и наружные межреберные мышцы. При сокращении инспираторных мышц ребра поднимаются. В результате объем грудной клетки увеличивается, особенно в ее нижних отделах. Сокращение мышцы диафрагмы также вызывает увеличение объема грудной клетки. Во время вдоха диа�фрагма уплощается, а в покое и особенно во время выдоха купол ее подни�мается и перемещается в грудную клетку. При грудном типе дыхание осуществляет�ся за счет сокращения межреберных мышц, при брюшном типе в основном сокращается диафрагма, которая одновременно смещает органы брюшной полости.
2) экспираторные (выдоха) – внутренние межреберные и мышцы брюшной стенки
3) вспомогательные, которые включаются при форсированном дыхании. Это ряд мышц шеи, груди и спины.
Во время вдоха при увеличении объема грудной клетки в замкнутой плевральной полости давление еще больше падает. Вследствие различия между атмосферным давлением в альвеолах и плевральным давлением лег�кие растягиваются, в целом увеличиваясь в объеме, следуя за грудной клеткой. По�явившаяся разница между давлением в легких и атмосферным давлением приводит к тому, что воздух начинает поступать через воздухоносные пути трахея, бронхи, в альвеолы, заполняя их, при этом давление выравнивает�ся.
Выдох в основном происходит пассивно: как только инспираторная мускулатура расслабляется, возросшая в ход вдоха эластическая тяга легких возвращает их в исходное состояние. В результате объем грудной клетки уменьшается и давление в плевральной полости возрастает. Это дав�ление передается на легочную ткань, поэтому одновременно повышается давление воздуха в альвеолах. Теперь уже давление воздуха в легких стано�вится больше, чем в атмосфере, и воздух благодаря этому начинает выхо�дить из легких по воздухоносным путям наружу.
Помимо указанного выше эластического сопротивления легких, дыхательные мышцы вынуждены преодолевать неэластическое сопротивление дыханию. Оно зависит от просвета воздухонос�ных путей — особенно голосовой щели, бронхов. Так, во время вдоха голосовая щель несколько расширяется, на выдохе — сужается, увеличивая сопротивле�ние потоку воздуха, что служит одной из причин большей длительности экспи�раторной фазы. Подобным же образом циклически меняются просвет бронхов и их проходимость. Тонус гладкой мускулатуры бронхов (сила сужения просвета) зависит от активности парасимпатической и симпатической иннервации: вагусные холинэргические влияния усиливают тонус мышц и сужают просвет бронхов, симпатическая адренергическая иннервация оказывает расслабляющее влияние.
В целом сопротивление, а значит – и работа дыхательных мышц, усиливаются с возрастанием глубины дыхания (эластическое сопротивление) и – при чрезмерно частом дыхании (неэластическое сопротивление).
2.3. Паттерны дыхания.
Один цикл чередования вдоха и выдоха составляет дыхательный акт. Обычно вдох несколько короче выдоха, у человека их соотношение примерно 1:3.
Паттерн дыхания - это соотношение компонентов дыхательного акта (длительность фаз, глубина дыхания, динамика давления и потоков в воздухоносных путях). Организм избирает такой паттерн дыхания, при котором требуемый уровень альвеолярной вентиляции достигается с наименьшей затратой энергии на работу дыхательных мышц.
Ритмичность дыхательных циклов относительна. Даже в условиях покоя низкоамплитудные дыхательные движения периодически сменяются движениями большой амплитуды, так называемыми вставочными вдохами, после которых наступает удлиненная пауза. Такие высокоамплитудные вдохи способствуют вентиляции альвеол застойных участков легких.
При различных состояниях организма, фармакологических воздействиях или неадекватных окружающих условиях наблюдаются расстройства дыха�ния. Все формы нарушения нормального дыхания объединяются термином «диспноэ». Часто встречается так называемое периодическое дыхание Чейна-Стокса: дыхательные движения постепенно углубляются и учащаются, что приводит к развитию гипервентиляции и гипокапнии. В результате частота и глубина дыхания уменьшаются вплоть до апноэ. Затем концентрация углекис�лоты в крови увеличивается, что в свою очередь приводит к новому нарастанию частоты и глубины дыхания. Причиной могут быть функциональные изменения возбудимости центрального дыхательного механизма, наступающие в результате ги�поксии, либо органического поражения ды�хательного центра. Дыхание Чейна-Сток�са наблюдается иногда у детей младшего возраста, у практически здоровых людей во время сна, а также в горах.
При хронической гипоксии головного мозга или отделении перерезкой передней части моста среднего мозга дыхание напоминает "лягушачье" — после развития вдоха следует инспираторная задержка и короткий выдох. Такой тип дыхания называется апнейстическим. Он может быть вызван снижением тонического импульсного потока к нервным клеткам пневмотаксического центра, регулирующего соотношение фаз дыхательного цикла, или частичной блокадой сенсорной информации, поступающей по блуждающему нерву.
Если устранены все влияния, исходя�щие из ростральных отделов центральной нервной системы, то дыхание приобретает судорожный характер — «гаспинг-дыхание», с резким вдохом макси�мальной амплитуды, в котором принимают участие не только дыхательные мышцы, но и другая скелетная мускулатура. Выдох происходит активно. Ды�хательные движения становятся редкими, с большими экспираторными пауза�ми. «Гаспинг-дыхание» присуще черепахам и суркам во время спячки.
Особый характер дыхания отмечен при одышке, т. е. при нарушении часто�ты и глубины дыхательных движений, сопровождающихся субъективным ощущением удушья.
2.4. Характеристика легочной вентиляции.
Легочной вентиляцией называют объем воздуха, вдыхаемого за единицу времени. Легочную вентиляцию подразделяют на несколько компонентов.
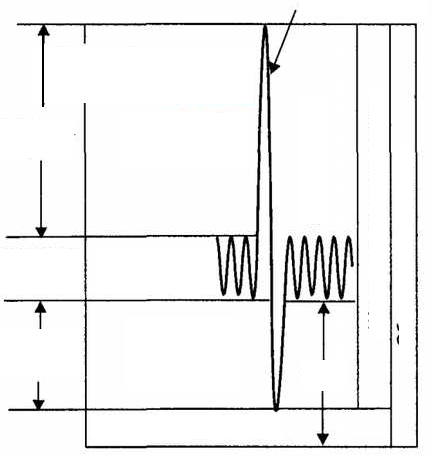
Дыхательный объем — количество воздуха, которое человек вдыхает и выдыхает в покое.
Резервный объем вдоха — количество воздуха, которое человек может до�полнительно вдохнуть после нормального вдоха.
Резервный объем выдоха — количество воздуха, которое человек может дополнительно выдохнуть после спокойного выдоха.
Остаточный объем — количество воздуха, оставшееся в легких после максимального выдоха.
Жизненная емкость легких — максимальное количество воздуха, кото�рое можно выдохнуть после наибольшего вдоха, состоящее из суммы дыха�тельного объема и резервных объемов вдоха и выдоха. Она зависит от многих факторов: конституции, возраста, пола, степени тренированности. С возрастом ЖЕЛ уменьшается, что связано со снижением эластичности легких и подвижности грудной клетки. У женщин ЖЕЛ в среднем на 25 % ниже, чем у мужчин. У мужчин ростом 180 см она в среднем составляет 4,5 л.
Общая емкость легких — максимальное количество воздуха, содержаще�гося в легких при наибольшем вдохе, является суммой жизненной емкости и общей емкости легких.
«Мертвое» воздушное пространство. Воздухоносные пути, включающие полости носа, рта, трахеи, бронхов, образуют так называемое «мертвое» пространство. Воздух, занимающий объем «мертвого» пространства, не уча�ствует в газообмене. Во время вдоха первая порция вдыхаемого воздуха по�ступает в альвеолы из «мертвого» пространства. Во время выдоха она воз�вращается последней в воздухоносные пути этого пространства, т.е. факти�чески один и тот же воздух «мертвого» пространства без обновления соста�ва поступает в легкие. Выдыхаемый воздух представляет собой смесь альвеолярного газа и воздуха мертвого пространства. В чистом виде альвеолярный газ выводится лишь с последней порцией воздуха.
3. Газообмен в легких
Происходящий в воздухоносных путях перенос газов направлен на поддержание постоянства парциального давления О2 и СО2 в легочных альвеолах, где идет непрерывный обмен газов с кровью, протекающей через легочные капилляры. В среднем парциальное давление кислорода при нормальных атмосферных условиях поддерживается в альвеолярном воздухе на уровне ~ 102 мм рт.ст., а дву�окиси углерода — на уровне около 40 мм рт.ст.
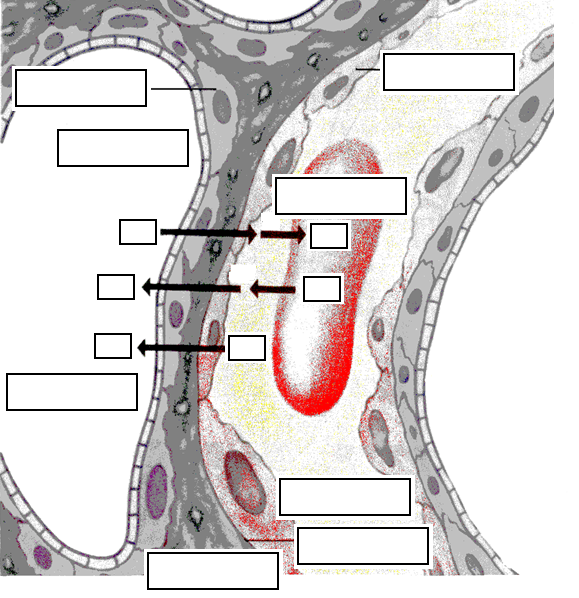
Перенос О2 из альвеолярного газа в кровь и СО2 из крови в альвеолярный газ происходит исключительно путем диффузии. Ее движущей силой служат разности (градиенты) парциальных давлений (напряжений) О2 и СО2 по обе стороны аэрогематического барьера (толщина 1 мкм). Кислород и углекислый газ диффундируют в растворенном состоянии, т.к. все воздухоносные пути увлажнены слоем слизи. Благодаря огромной общей поверхности альвеол, составляющей 80-100 м2, имеются условия для достаточно эффек�тивной диффузии. Тонкий слой легочной ткани, отделяющей кровь легочных капилля�ров от альвеолярного пространства, легко проницаем для газов. В процессе диффузии газы проходят через альвеолярный эпителий, интерстинальное пространство между основными мембранами, эпителий капилляров, плаз�му крови, мембраны эритроцитов во внутреннюю среду эритроцитов.
Вдыхаемый воздух имеет наибольшее парциальное давление кислоро�да (100 мм рт.ст.) и наименьшее парциальное давление двуокиси углерода (20 мм рт.ст.). В то же время парциальное давление кислорода в притекающей к альвеолам венозной крови не пре�вышает 40 мм рт.ст., а парциаль�ное давление двуокиси углерода составляет 50 мм рт.ст. Благодаря градиенту давлений происходит транспорт газов через стенку альвеол: двуокись углерода покидает венозную кровь и поступает в альвеолярный воздух, а кислород диффундирует в противопо�ложном направлении — из альвеолярного воздуха в кровь. Оттекающая от альвеол легких артериальная кровь имеет парциальное давление кислоро�да 100 мм рт.ст., а двуокиси углерода — 40 мм рт.ст.
4. Транспорт газов кровью
4.1. Транспорт кислорода
Кислород транспортируется кровью двумя способами:
- в связанном с гемоглобином виде — в форме оксигемоглобина (около 98% всего О2).
- за счет физического растворения газа в плазме крови. В плазме растворена лишь небольшая часть О2 (около 2%).
Большая часть кислорода переносится кровью в виде химических соединений с гемоглобином, который легко вступает с ним в непрочное соединение.
Hb + O2 HbO2
(дезоксигемоглобин) (оксигемоглобин)
Один моль гемоглобина может связать до четырех молей кислорода и в среднем 1 г гемоглобина способен связать 1,34—1,36 мл кислорода. Учитывая, что в крови человека содержится примерно 150 г/л гемоглобина, 100 мл крови могут переносить около 21 мл О2. Это так называемая кислородная емкость крови.
В эмбриональный период гемоглобин человека имеет особую форму – фетальный гемоглобин F. Он способен переносить на 20-30 % больше кислорода, обладает большей способностью связываться с ним (сродством к кислороду). К моменту рождения гемоглобин F составляет 50-80%, к 3 годам около 2 %, затем исчезает. Большая часть гемоглобина взрослого человека (95-98%) состоит из фракции А (взрослый гемоглобин), около 1-2% гемоглобин F (фетальный).
Гемоглобин легко соединяется с угарным газом – оксидом углерода СО, образуя устойчивое соединение - карбоксигемоглобин. Химическое сродство СО к гемоглобину почти в 300 раз выше, чем к О2, поэтому даже при небольших концентрациях СО в воздухе гемоглобин оказывается блокирован для кислорода (на 80% при концентрации СО 0,1 %, если концентрация около 1% - гибель через несколько минут).
Способность гемоглобина связывать и отдавать O2 зависит от напряжения кислорода, угольной кислоты в крови, рН крови, ее температуры и т.д. Графическое изображение зависимости процента насыщения гемоглобина кислородом от напряжения О2 называют кривой диссоциации оксигемоглобина. Участок кривой, соответствующий низким парциальным значениям кислорода, характеризует содержание оксигемо�глобина в капиллярах тканей, а фрагмент кривой, лежащий в области высо�кого парциального давления кислорода, соответствует крови в легочных капиллярах. Чем выше парциальное давление кислорода, тем больше содержание оксигемоглобина; при парциальном давлении 80-100 мм рт.ст. практически весь гемоглобин насыщается кислородом, за исключением незначительно�го количества (1—2 %), «занятого» двуокисью углерода. В области высокого парциального давления кислорода (в легких) кривая близка к насыщению кислородом крови, а в области низких значений парциального давления кислорода в тканях зна�чительная часть оксигемоглобина отдает кислород и превращается в вос�становленную форму.
Динамика кривой зависит от нескольких факторов. Кривая может сдвигаться относительно оси абсцисс вправо или влево (эффект Бора) в зависимости от температуры, парциального давления двуокиси углеро�да и величины рН. При увеличении содержания двуокиси углерода, температуры и закислении крови (ацидоз) кривая диссоциации оксигемоглобина сдвигается вправо. Это отражает по�вышение способности оксигемоглобина отдавать кислород тканям и тем самым высвобождаться для дополнительного связывания СО2 и переноса его избытка из тканей в легкие. Напротив, при снижении Рсо2 и защелачивании крови (алкалоз) кривая сдвигается влево.
4.2. Транспорт двуокиси углерода.
Двуокись углерода, образующаяся в тканях, транспортируется кровью тремя способами.
- в виде гидрокарбоната НСОз- плазмы крови и цитоплазмы эритроцитов, образующегося в результате диссоциации угольной кислоты:
Н2О + СО2 = Н2СО3 = Н+ + НСО3- Таким способом транспортируется около 4/5 всего углекислого газа.
- в виде химического со�единения с дезоксигенированным гемоглобином — карбогемоглобина (около 15 %).
- так же как и О2, СО2 переносится в фи�зически растворенном состоянии (3-6% общего количества СО2). Содержание физически растворенной двуокиси углерода в артериальной крови составляет 0,026 мл в 1 мл крови, что в 9 раз превышает количество физически растворенного кислорода.
Содержание СО2 и О2 в крови и тканях активно влияет на рН. Избыток двуокиси углерода ведет к увеличению содержания угольной кислоты и повышению концентрации водородных ионов (ацидоз). Сниже�ние двуокиси углерода вызывает обратную реакцию — развитие защелачивания (алкалоз).
5. Транспорт кислорода и углекислого газа в тканях.
Кислород проникает из крови в клетки тканей путем диффузии, обуслов�ленной разностью (градиентом) его парциальных давлений по обе стороны, так называемого гематопаренхиматозного барьера. Так, среднее Ро2 артериальной крови составляет около 100 мм рт. ст., а в клетках, где кислород непрерывно утилизируется, стремится к нулю.
Напряжение кислорода в тканях в среднем составляет 20-40 мм рт. ст. Однако эта величина в различных участках живой ткани отнюдь не одинакова. Наибольшее значение Ро2 фиксируется вблизи артериального конца кровенос�ного капилляра, наименьшая — в самой удаленной от капилляра точке («мерт�вый угол»).
Функция газотранспортной системы организма в конечном счете направлена на поддержание парциального давления кислорода на клеточной мембране не ниже критического, т. е. минимального, необходимого для работы ферментов дыхательной цепи в митохондриях. Для клеток, интенсивно потребля�ющих кислород, критическое Ро2 составляет около 1 мм рт. ст.
Вместе с тем следует иметь в виду, что напряжение О2 в тканях зависит не только от снабжения кислородом, но и от его потребления клетками. Наиболее чувствительны к недостатку кислорода клетки кардиомиоцитов и нейроны мозга, где окислительные про�цессы очень интенсивны (реанимация, инфаркт). В отличие от этих клеток, скелетные мыш�цы относительно устойчивы к кратковременному прекращению кислородного снабжения, т.к. они могут использовать анаэробные процессы получения энергии, а также содержат (особенно красные волокна) миоглобин.
Перенос СО2 из клеток тканей в кровь тоже происходит главным образом путем диффузии, т. е. в силу разности напряжений СО2 по обе стороны гематопаренхиматозного барьера. Среднее артериальное значение Рсо2 в среднем составляет 40 мм рт. ст., а в клетках может достигать 60 мм рт. ст. Локальное парциальное давление углекислого газа и, следовательно, скорости его диффузионного транспорта в значительной мере определяются продукцией СО2 (т. е. интенсивностью окислительных процессов) в данном органе.
По той же причине Рсо2 и Ро2 в различных венах не одинаковы. Так, в крови, оттекающей от работающей мышцы, напряжение О2 гораздо ниже, а напряжение СО2 гораздо выше, чем, например, в крови, оттекающей от соеди�нительной ткани.
6. Регуляция дыхания.
В механизмах регуляции дыхания можно выделить афферентное, центральное и эфферентное звенья.
6.2. Афферентное звено
Величина дыхательных показателей в организме определяется: периферическими хеморецепторами сосудистых рефлексогенных зон, центральными хеморецепторами, находящимися в области продолговатого мозга и рецепторами легких. Хеморецепторы воспринимают изменения парциального давления СО2 и О2, а также рН крови, а механорецепторы – это рецепторы растяжения легких. Так, повышение давления СО2 (явление гиперкапнии), снижение рН крови (ацидоз) и снижение содержания О2 (гипоксемия) вызывают стимуляцию бульбарных хемочувствительных структур, что приводит к увеличению легочной вентиляции. Снижение же давления CO2 (явление гипокапнии) и повышение рН крови (алкалоз) вызывают торможение бульбарных хемочувствительных структур, что приводит к снижению напряжения СО2.
6.3. Нервные центры
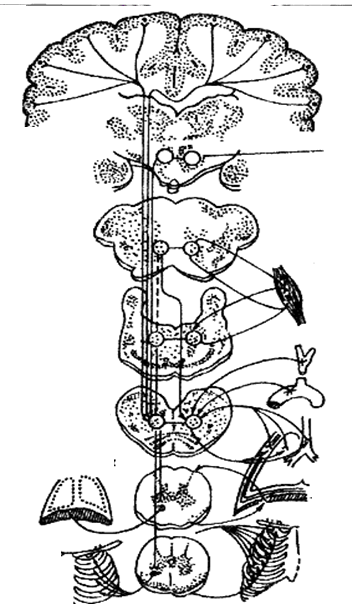
В широком смысле слова под дыхательным центром понимают сово�купность структур мозга, так или иначе участвующих в регуляции дыхания и в наиболее совершенном приспособлении его к изменяющимся дыха�тельным потребностям организма. Среди этих структур — кора большого мозга, гипо�таламус, мост мозга, а также продолговатый и спинной мозг.
6.3.1. Дыхательные нейроны. Основными нейрональными элементами, входя�щими в дыхательный центр, являются дыхательные нейроны. Все они обла�дают ритмической залповой активностью, возникающей в определенный период дыхательного цикла. В зависимости от того, в какой период разря�жаются нейроны, их относят к инспираторным или экспираторным нейро�нам. Инспираторные нейроны дают залповый разряд в фазу вдоха, экспира�торные нейроны разряжаются в период выдоха. Инспираторные и экспираторные нейроны рас�полагаются диффузно в непосредственной близости друг от друга. Между инспираторными и экспираторными нейронами в большинстве случаев проявляются реципрокные отношения. Для инспираторных нейро�нов характерна непрерывная импульсная активность, обусловленная биоэлектрохимическими процессами. Первично возникающая импульсная активность инспираторных нейронов активирует экспираторные нейроны, те в свою очередь тормозят активность инспираторных нейронов.
Ритмическое ды�хание обеспечивается в первую очередь периодической (фазной) деятельностью дыхательного центра продолговатого мозга. Это единственная структура из множества образований мозга, принимающих участие в регуляции дыхания, которая способна самостоятельно автоматически поддерживать ритмическое дыха�ние. Остальные структу�ры мозга модулируют ритм деятельности дыхательного центра, оказывая влияние на частоту и глубину дыхания и плавность смены вдоха на выдох.
Таким образом, функции дыхательных нейронов различных отделов мозга существенно отличаются.
Нейроны спинального уровня расположены в передних рогах шейного и грудного отделов спинного мозга, они рeгулируют деятельность диафрагмы и дыхательных мышц.
Нейроны продолговатого мозга расположены на дне 4-го желудочка, делятся на инспираторные и экспираторные, являются центрами реципроктной координации (согласование вдоха и выдоха).
Нейроны моста расположены в ядрах черепных нервов и ретикулярной формации – регулируют частоту дыхания.
Нейроны гипоталамуса обеспечивают зависимость дыхания от функционального состояния организма (сон, бодрствование, физическая нагрузка), условий внешней среды, согласование дыхания с работой сердца.
Корковые нейроны обеспечивают зависимость дыхания от психоэмоционального состояния, условно-рефлекторные реакции. Произвольный контроль дыхательных движений.
6.3.2. Легочно-вагусная регуляция дыхания
Установлено, что в альвеолах легких, стенках бронхиол имеются механорецепторы, которые реагируют на увеличение объема и растяжения ле�гочной ткани. При поступлении в легкие воздуха альвеолы растягиваются, при этом увеличивается частота импульсации, генерируемая механорецепторами. По афферентным волокнам блуждающих нервов им�пульсы от механорецепторов легких проводятся к дыхательному центру. Пропорционально степени расширения легких во время вдоха в афферент�ных волокнах блуждающих нервов постепенно нарастает импульсация, которая при определенной частоте тормозит вдох и вызывает выдох.
7. Особенности дыхания в различных условиях
Кроме афферентации, поступающей по блуждающим нервам, актив�ность дыхательного центра определяется рядом других факторов: темпера�турой крови, импульсацией, поступающей от сосудистых барорецепторов и от мышечных проприорецепторов, болевыми стимулами, а также влияния�ми, связанными с эмоциями и речью. В целом эти влияния отражают меж�системные взаимоотношения с другими функциональными системами ор�ганизма.
Дыхание при разных температурах среды. Повышение температуры тела вызывает учащение дыхания. У ряда животных, в частности у собак, учащение дыхания, спо�собствующее испарению влаги с языка, является одним из путей стабилиза�ции температуры тела. Охлаждение организма, вызывающее мышечную дрожь, также влияет на дыхание, нарушая его регулярность.
Дыхание на большой высоте. По мере подъема на высоту парциальное давление кислорода падает параллельно снижению атмосферного давления.
При вдыхании атмосферного воздуха с уменьшенным парциальным давлением кислорода в организме возникает гипоксия, дыхательным отве�том на которую является усиление легочной вентиляции. В результате ги�первентиляции из организма в избытке удаляется двуокись углерода, разви�вается гипокапния и появляется связанный с ним сдвиг рН в основную сторону (алкалоз). Этот процесс в определенной степени тормозит прирост легочной вентиляции и снижает участие внешнего дыхания в компенсации гипоксии. Дыхание становится поверхностным и частым. При нарастании гипоксии человек теряет сознание, а затем погибает.
Негативное влияние гипоксии на больших высотах (при подъеме в ус�ловиях высокогорья) усугубляется тем, что человек не в состоянии объек�тивно оценить опасность, так как развитие гипоксии сопровождается эйфорией.
Для предотвращения гипоксических нарушений необходимо контроли�ровать подъем и продолжительность пребывания человека на высоте. Эф�фективным средством профилактики гипоксических нарушений при подъ�еме на высоту является использование кислорода для дыхания. Для этого применяют специальные кислородные аппараты.
Дыхание при повышенном давлении (кессонная болезнь). Под водой человек вынужден ды�шать воздухом, подаваемым в легкие из баллона под избыточным давлени�ем, нарастающим по мере погружения. При повышении парциального давления газов физическая раствори�мость их в плазме крови и других жидкостях организма увеличивается. Осо�бенно это касается азота, который в наибольшем количестве содержится во вдыхаемом воздухе, следовательно, и парциальное давление будет при этом наивысшим. При пребывании человека в течение продолжительного времени на большой глубине азот, находящийся при высоком парциальном давлении во вдыхаемом воздухе, в избытке растворяется в плазме крови (явление компрессии). При бы�стром подъеме на поверхность растворенный в крови азот быстро возвращается в газообразное состояние, мелкие его пузырьки закупоривают кро�веносные сосуды, особенно капилляры и артериолы (явление декомпрессии). В результате газовой эмболии нарушается кровообращение в различных органах, которое сопро�вождается резкими болями в мышцах, головокружением, потерей созна�ния, нервными расстройствами, парезами и параличами; в первую очередь страдают функции ЦНС. Такие явления называют кессонной болезнью. Кессонная болезнь может развиваться также при быстром подъеме в горы.
Для предотвращения кессонной болезни необходим медленный подъем человека на поверхность для того, чтобы азот мог постепенно выйти из ор�ганизма без образования пузырьков газа. Сразу после подъема проводят по�степенную декомпрессию человека в барокамере, в которой создано повы�шенное давление, имитирующее давление вдыхаемого воздуха при погру�жении. Постепенно в течение нескольких часов производят снижение дав�ления, позволяющее медленно вывести из организма избыток растворенно�го газа. В ряде случаев при работе на больших глубинах используют газовые смеси, лишенные азота (например, гелио-кислородную), которые наряду со снижением опасности развития кессонной болезни снимают токсическое действие азота (азотное опьянение) на глубине.
Влияние артериального давления. Деятельность сосудодвигательного и дыхательного центров носит сопряженный характер и координируется влияниями блуждающих нервов. Депрессорная активность, возникающая в барорецепторных зонах, способна влиять на величину артериального давления и на характер дыхания, одновременно затрагивая деятельность сосудодвигательного и дыхательного центров. Повышение артериального давления может сочетаться с урежением дыхания.
Сигнализация от мышц. При изменении мышечного тонуса и при появ�лении моторной активности мышечные проприорецепторы (мышечные ве�ретена) посылают по у-афферентным волокнам импульсацию к дыхатель�ному центру, которая вызывает активацию дыхания. Благодаря этому меха�низму активация дыхания возникает задолго до того, как появляется нару�шение кислородно-углекислотного баланса в организме, например при мы�шечной работе.
Болевые реакции. Отчетливые изменения дыхания возникают при болевых реакциях. Как показали опыты, даже у наркотизированных нембуталом кошек незначительное раздражение в виде укола кожи живота вызывает изменение в ритмической деятельности дыхательного центра.
Эмоциональные влияния. Эмоциональное возбуждение, охватывающее структуры лимбико-ретикулярного комплекса и в первую очередь гипота�ламус, распространяется в нисходящем направлении и также вызывает из�менение деятельности дыхательного центра.
Речь и дыхание. Речь, относящаяся к высшим мозговым функциям человека, а также голосовые реакции животных возникают на основе дыха�тельных движений, вызывающих прохождение воздуха через голосовой аппарат. Внешнее дыхание становится эффекторной функцией сразу двух систем: функциональной системы дыхания, обеспечивающей оптимальный газовый баланс в организме, и функциональной системы речи, осущест�вляющей построение слов, фраз и контролирующей смысловое содержание речи. Поэтому во время речи к дыхательному центру приходят влияния, подстраивающие его деятельность для необходимых речевых реакций. В то же время дыхательный центр управляет тем объемом легочной вентиляции, который необходим для поддержания дыхательного гомеостаза. Поэтому дыхание в этих условиях становится апериодическим, одновременно удов�летворяя двум необходимым условиям.
Произвольный контроль дыхания в отдельных случаях может иметь самостоятельное значение в регуляции дыхательных показателей организ�ма. Например, предварительная гипервентиляция перед подводным погру�жением и задержка дыхания под водой могут служить компенсаторными механизмами для предотвращения неблагоприятных для организма послед�ствий. Гипервентиляция перед погружением в воду способствует увеличе�нию запаса кислорода в организме и уменьшению содержания двуокиси уг�лерода, что позволяет удлинить время задержки дыхания под водой.
Состав сухого воздуха:
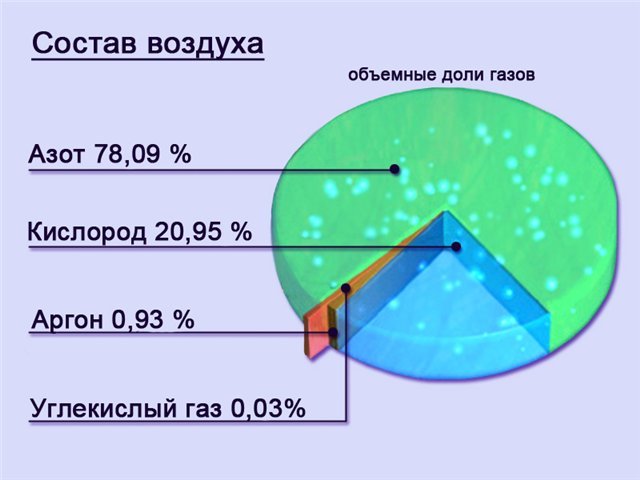
Содержание и парциальное давление (напряжение) кислорода и углекислого газа в различных средах
|
Среда
|
Кислород
|
Углекислый газ
|
|
|
%
|
мм рт.ст.
|
%
|
мм рт.ст.
|
|
Вдыхаемый воздух
|
|
|
|
|
|
Выдыхаемый воздух
|
|
|
|
|
|
Альвеолярный воздух
|
|
|
|
|
|
Артериальная кровь
|
|
|
|
|
|
Венозная кровь
|
|
|
|
|
|
Ткань
|
|
|
|
|
|
Около митохондрий
|
|
|
|
|
Плевральная полость:
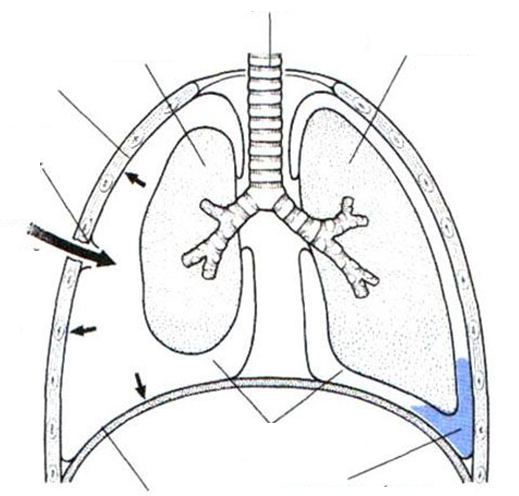
Механизм вдоха и выдоха

Фазы дыхательного акта (спирограмма)

Паттерны дыхания
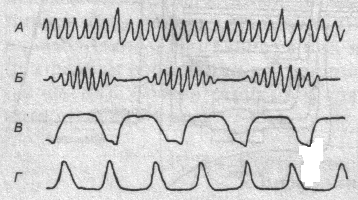
Схема легочных объемов и емкостей, их отражение на спирограмме
Газообмен в легких
Аэрогематический барьер
Кривая диссоциации оксигемоглобина
Дыхательный центр
Физиология дыхания