Биологические особенности кеты в связи с его искусственным разведением
PAGE \* MERGEFORMAT - 1 -
КУРСОВАЯ РАБОТА
на тему: «Биологические особенности кеты в связи с его искусственным разведением»
Содержание
Введение ------------------------------------------------------------------------------------ 3
Глава 1. Биологическая характеристика кеты в связи со средой обитания и образом жизни ----------------------------------------------------------------------------- 9
- Общая биология вида ------------------------------------------------------------ 9
- Эмбриональное развитие -------------------------------------------------------- 18
- Постэмбриональное развитие -------------------------------------------------- 46
Глава 2. Влияние различных факторов среды на кету ( температура, pH, кислород, гидрохимические показатели)-------------------------------------------- 54
Глава 3. Управление половыми циклами у кеты ----------------------------------- 59
Глава 4. Биологические особенности кормления кеты --------------------------- 63
Глава 5. Транспортировка икры, личинок, молоди и взрослых особей кеты-
------------------------------------------------------------------------------------------------- 66
Глава 6. Биологические основы акклиматизации кеты --------------------------- 71
Список используемых источников----------------------------------------------------- 75
ВВЕДЕНИЕ
Рыбное хозяйство в Российской Федерации является комплексным сектором экономики, включающим широкий спектр видов деятельности - от прогнозирования сырьевой базы отрасли до организации торговли рыбной продукцией в стране и за рубежом.
Со многими государствами заключены межправительственные соглашения об организации использования водных биологических ресурсов в их исключительных экономических зонах, в районах действия международных конвенций по рыболовству и в исключительной экономической зоне Российской Федерации.
Предприятия рыбного хозяйства являются градообразующими во многих приморских регионах страны, обеспечивают занятость населения. Особое значение это имеет для районов Дальнего Востока и Крайнего Севера, где рыбный промысел является основным источником обеспечения жизнедеятельности населения, в том числе коренных малочисленных народов.
Сырьевая база рыбного хозяйства имеет ряд особенностей, связанных с сезонностью промысла, подвижностью водных биологических ресурсов, трудностью прогнозирования запасов водных биологических ресурсов, определения рациональной доли их изъятия без ущерба для воспроизводства. Изучение, добыча, сохранение и воспроизводство водных биологических ресурсов обеспечиваются специализированным научным, рыбопромысловым, рыбоохранным и вспомогательным флотами и объектами по воспроизводству рыбных запасов.
Целью развития рыбного хозяйства в Российской Федерации является достижение устойчивого функционирования рыбохозяйственного комплекса на основе сохранения, воспроизводства и рационального использования водных биологических ресурсов, развития аква- и марикультуры, обеспечивающего удовлетворение внутреннего спроса на рыбную продукцию, продовольственную независимость страны, социально-экономическое развитие регионов, экономика которых зависит от прибрежного рыбного промысла. При этом должны быть созданы условия для повышения эффективности экспорта рыбной продукции и ее конкурентоспособности и оптимизации структуры управления рыбохозяйственным комплексом.
В последние 10 лет Госкомрыболовством России проводились широкомасштабные мероприятия по развитию лососеводства на Дальнем Востоке, особенно в Сахалино-Курильском регионе. Построено 6 новых рыбоводных заводов, а также реконструировано 60% действующих заводов и в настоящий момент в России действует 59 рыбоводных предприятий по воспроизводству различных видов атлантических и тихоокеанских лососей.
Из всех объектов аквакультуры широко известная группа рыб - ло�сосевые (лососеобразные) давно освоена и имеет высокую ценность. Среди всех морских и пресноводных рыб лососевые занимают ведущее место в связи с высокими пищевыми качествами мяса и икры. Многие из них являются проходными рыбами, имеют сложный жизненный цикл развития. Другие, чисто пресноводные, населяют водоемы арк�тических, субарктических зон, зоны умеренного климата, реки и озера горных ландшафтов.
Среди представителей мировой ихтиофауны, насчитывающей око�ло 20 тыс. видов, лососевым принадлежит относительно скромное место. Однако в системе мировой аквакультуры они занимают особое положение, которое определяется, в первую очередь, особенностью биологии и высокой рыбохозяйственной ценностью. Лососевым рыбам свойственны резкие колебания численности, высокая репродуктивная способность, быстрый темп роста и пластичность по отношению к условиям индустриальных хозяйств аквакультуры. Они как источник высококачественной пищевой продукции являются традиционным объектом промысла. На мировом рынке эти рыбы пользуются неиз�менной популярностью, и, несмотря на высокую цену, спрос в полной мере удовлетворяется. В современном мировом улове, составляющем более 100 млн т, лососевые составляют не более 0,5 млн т, но эта отно�сительно скромная часть, менее 5%, достигает 10% общей стоимости улова.
К семейству лососевых (Salmonidae) относятся проходные и пре�сноводные . Сложный жизненный цикл повышает уязвимость лососей на про�тяжении онтогенеза под влиянием естественных и антропогенных факторов. Под влиянием интенсивного промысла численность проходных лососевых рыб в последние десятилетия продолжала снижаться. Помимо нерационального лова, существенный ущерб запасам лососей был нанесен ухудшением гидрологических и гидрохимических усло�вий нерестовых водоемов. Гидростроительство, лесосплав, загрязнение водоемов сточными водами явились основными факторами снижения эффективности естественного воспроизводства лососевых рыб. В результате загрязнения нерестовых рек особенно пострадали атлантические лососи p. Salmo. Многие ранее нерестовые реки США, Канады, Англии, Франции, Ирландии, Норвегии, Швеции и России полностью лишились этого лосося.
Меры охраны естественного воспроизводства лососевых дают ожи�даемые результаты только в районах, где нерестовый фонд находится в удовлетворительном состоянии и численность нерестовых популяций обеспечивает необходимый уровень пополнения запаса. Поскольку в большинстве районов мира эти условия нарушены, все большее раз�витие получает управляемое воспроизводство, в частности, искусст�венное разведение молоди и выпуск ее в естественные водоемы.
Искусственное разведение как способ пополнения запасов проход�ных лососей имеет определенные преимущества - возможность конт�роля на всех этапах эмбрионально-личиночного и малькового развития. Это позволяет выращивать потомство с определенными свойствами, нужного размера и качества, обеспечивает повышенную сохранность особей, эффективность селективного воздействия с целью повышения жизнестойкости покатной молоди.
Главной задачей искусственного воспроизводства тихоокеанских лососей является получение крупной, физиологически полноценной молоди, способной выжить не только после выпуска ее в естественные водоемы и в процессе катадромной миграции, но и дать промысловый возврат (Хованский, 2004; Запорожец, Запорожец, 2006). Зарубежный и отечественный опыт лососеводства показали, что размеры и физиологическая полноценность рыб во многом зависят от состава применяемых искусственных рационов (Halver, 1972; Watanabe et al., 1974; Остроумова, 1983; Канидьев, 1984; Фомин, 1996; Валова, 1999; Гамыгин, 2001; Воропаев и др., 2003, Пономарев, Пономарева, 2003).
Выживаемость молоди на рыбоводных заводах составляет обычно 70-85% от инкубируемой икры, в то время как в естественных условиях эта величина в несколько раз ниже. Это подтверждается величиной коэффициента ската, т.е. количеством мигрантов в процентах к общей плодовитости, как показателя эффективности нереста, эмбрионально-�личиночного развития и роста молоди в пресноводный период жизни. Коэффициент ската атлантических лососей преимущественно колеблется в пределах 3-10%, тихоокеанских лососей - от 0,04-8,5%. В на�стоящее время заводское разведение смолтов и выпуск в естественные водоемы обеспечивает большую часть промыслового запаса лососей и р. Salmo таким странам, как Швеция, Норвегия, и р. Oncorhynclms таким странам как Россия, Канада, США и Япония.
Разведение тихоокеанских лососей имеет более чем вековую историю. Первые отечественные лососевые рыбоводные заводы (ЛРЗ) появились в 1909 г. В странах Азиатско-тихоокеанского региона в настоящее время функционируют около 400 лососевых рыборазводных заводов. Искусственное воспроизводство тихоокеанских лососей наиболее развито в Японии, США, Канаде и России.
Основным регионом искусственного воспроизводства лососевых в Российской Федерации на сегодняшний день является Сахалинская область. В общем промысловом изъятии кеты Сахалинской области, кета искусственного воспроизводства составляет 95% (Затулякин, 2004). В последние годы наметилась тенденция по увеличению заводского воспроизводства кеты. Выпуск молоди кеты на ЛРЗ Российского Дальнего Востока, в том числе Сахалина, превышает 50% от выпуска всех лососей на Дальнем Востоке России (Смирнов и др. 2006; Марковцев, 2006), что свидетельствует о возрастании интереса к кете как к промысловому виду. Кета также является основным объектом разведения на японских рыбоводных заводах (Гриценко и др., 2001; 2001; Кловач, 2003; Шевляков и др., 2006).
Коэффициенты промыслового возврата кеты колеблются в широких пределах, так для ЛРЗ Сахалинской области коэффициент может изменяться в пределах от 0,02% для Соколовского ЛРЗ в 1997 г. и до 7,75% для Рейдового ЛРЗ в 1999 г. (Смирнов и др., 2006). На Калининском ЛРЗ из года в год коэффициент возврата варьирует от 0,1% до 1,6%. В годы низких промысловых возвратов ряд ЛРЗ Сахалинской области испытывают нехватку собственных производителей, в таком случае довольно часто практикуются перевозки оплодотворенной икры с других ЛРЗ.
В современных условиях, при интенсивном давлении антропогенных факторов не только на нерестовую, но и на нагульную часть видового ареала роль искусственного воспроизводства атлантического лосося существенно увеличилась. Без рыбоводных мероприятий тихоокеанский лосось может в короткое время потерять свое промысловое значение.
Цель данной курсовой работы – изучить биологические особенности кеты в связи с ее искусственным разведением. Поставленная цель определила следующие задачи:
- дать биологическую характеристику кеты в связи со средой обитания и образом жизни: общая биология вида, основные периоды онтогенеза;
- изучить влияние различных факторов среды на кету;
- выбрать один из методов стимулирования половых продуктов у производителей кеты и описать данный метод;
- описать биологические основы кормления кеты, определить спектр питания исследуемого объекта в ранние периоды онтогенеза;
- выбрать одни из методов транспортировки кеты на разных этапах онтогенеза;
- подобрать подходящий район акклиматизации, описать водоем ,в котором можно акклиматизировать кету.
Глава 1. Биологическая характеристика объекта в связи со средой обитания и образом жизни
1.1 Общая биология вида
Кета по современной систематике (Решетников, 2003) занимает следующее положение:
Домен Эукариоты – Eukaryota
Царство Животные – Animalia
Подцарство Многоклеточные – Eumetazoa
Раздел Двусторонне симметричные – Bilateria
Тип Хордовые – Chordata
Подтип Позвоночные – Vertebrata
Надкласс Челюстноротые – Gnathostomata
Группа (ряд) Рыбы – Pisces
Класс Костные рыбы – Osteichthyes
Подкласс Лучеперые – Actinopterygii
Надотряд Клюпеоидные – Clupeomorpha
Отряд Лососеобразные – Salmoniformes
Семейство Лососёвые – Salmonidae
Род Тихоокеанские лососи – Oncorhynchus
Вид Кета – Oncorhynchus keta ( рис.1)

Рисунок 1 – Кета
Кета - наиболее широко распространенный вид тихоокеанских лососей.
В пределах российского Дальнего Востока выделяют шесть районов воспроизводства кеты: Камчатка, Сахалин, северо-западное побережье Охотского моря, бассейн р. Амур, Приморье и бассейн р. Анадырь. В данной работе речь пойдет о кете, которая использует в качестве нерестилищ район бассейна реки Амур.
Являясь представителем рода тихоокеанских лососей, кета, естественно, имеет много общего со всеми остальными видами лососей, однако у нее есть и свои специфические черты, по которым может быть легко отличима от других видов. Для кеты характерно большое количество пилорических придатков (до 185), жаберных тычинок имеет 19 - 25, жаберных лучей - 12 - 15, чешуй в боковой линии - 125 - 150.
Кета по уловам среди тихоокеанских лососей занимает второе место, а в годы слабых подходов горбуши - первое. Средняя длина для половозрелых особей составляет 60-70 см, вес 3—4,5 кг; рекордные экземпляры вырастают более метра и весят свыше 14 кг. Этого лосося давно разводят на Дальнем Востоке. Во многих от�ношениях он изучен лучше родственных видов.
В течение лета лососи пере�мещаются в высокие широты, достигая северных берегов Охотского и Берингова морей. Небольшое количество кеты мигрирует в воды Ледовитого океана на западе до р. Лены, на востоке - до р. Макензи (Берг, 1948). Кета это хищная рыба. Молодняк кеты едят различный планктон, личинки, насекомых и червей. Позже рыба переходит на питание рыбой, ракообразными, моллюсками. В рацион часто входит молодняк сельди, песчанки, корюшки и другой не крупной рыбы. В период нереста кета прекращает питание. У созревающих особей нагульная миграция переходит в нерестовую, а рыба младших возрастов с осен�ним похолоданием движется к зоне действия теплых вод Куросио и Северотихоокеанского течения на зимовку. Сведения о местах зимовок, нагула, миграциях содержат многие источники (Hirano, 1953; Бирман, 1958, 1967, 1968; Семко, 19581964; Hartt, 1962 Sano1966). На юге распространение кеты совпадает примерно с изотермой 10°, на севере - с изотермой 1,0—1,5°. Кета, воспроизводящаяся в водоемах Азии, зимует и нагуливается в разных рай�онах. По имеющимся данным, из района, расположенного южнее западной группы Алеутских островов, миграции прослеживаются к р. Анадырь, восточному и западному побережьям Камчатки, к северу охотоморского побережья. Осенняя кета, меченная в Але�утском районе, вылавливалась в реках Сахалина, Хоккайдо, Хон�сю и даже в Амуре. Из района зимовок, лежащих юго-восточнее южных Курильских островов и о. Хоккайдо, в Амур, реки Саха�лина и Хоккайдо мигрирует летняя кета. Там же зимует какая-то часть охотской, западно- и восточнокамчатской кеты. В Японском море к востоку и юго-востоку от Корейского полуострова зимует и нагуливается приморская кета и некоторая часть летней кеты Амура, Сахалина и Хоккайдо. Ареал кеты шире родственных ви�дов. Особенно далеко от родных берегов уходит осенненерестующая кета.
Для размножения кета заходит почти во все реки Азиатского континента от северной границы распрост�ранения до р. Тумень-Ула и р. Раку то на юге. На Командорских островах встречается редко. В массе нерестится в реках Сахали�на и Курильских островов, в значительном количестве заходит в реки о. Хоккайдо; границей распространения служат северные реки о. Кюсю. В Северной Америке кета нерестится от р. Макен�зи до рек зал. Монтеней (Берг, 1948, Clemens, Wilby, 1961; Neave, 1966b; Sano, 1966).
Промысловики издавна вы�деляют летнюю и осеннюю кету. Л.С. Берг (1934, цит. по 1953, 1948) назвал их расами, или экотипами; в таком смысле эти тер�мины мы и будем применять. В качестве типичной формы - О. keta (Walbaum) он взял летненерестующую кету, причем не обо�собляя представителей разных регионов. Ей противопоставлялась осенняя амурская кета - О. keta infraspecies autumnalis Berg.
А.Г. Смирнов (1947) подчеркнул экологическую специфику се�зонных форм амурской кеты. В бассейне Амура осенняя кета не�рестится на местах выхода грунтовых вод, тогда как летняя раз�множается в руслах рек и протоках, и ее гнезда омываются преи�мущественно подрусловым потоком. Различия экологии отра�жаются на всей биологии. В. К. Солдатов (1912) отметил боль�ший размер, вес и плодовитость осенней кеты, которая растет быстрее. Данные о различиях размеров, веса, плодовитости и других признаков сезонных рас амурской кеты приводят многие авторы ((Кузнецов, 1928. 1937: Ловецкая, 1948 Бирман. 1952, 1956 и др). Они отличаются морфологически и кариотипами ( Кулякова, 1971, 1972).
В отличие от летней, осеняя кета заходит в Амур до основного паводка, имея слабо развитые гонады. Крупный размер и высокая жирность осенней кеты рассматриваются как приспособление к длительной миграции против течения (Никольский, 1963) По Амуру она поднимается до 2000 км и более Девятнадцать главнейших для нее нерестовых рек другие лососи не посещают Основная масса осенней кеты в бассейне Амура нерестует в малоснежных районах с суровыми зимами, и потомство успешно развивается лишь благодаря снабжению гнезд грунтовыми водами, сильно не остывающими зимой. Эта форма занимает южную часть ареала вида, совпадающую с границами древней системы Полеоамура (Бирман, 1952).
Летняя кета по Амуру поднимается значительно ниже - в промысловых количествах до р. Ульчи (около 600 км от устья). В бас�сейнах крупных рек летняя и осенняя кета предпочитаю небольшие притоки. При заходе в один и тот же приток нерестилища осенней кеты располагаются выше. Летняя кета созревает несколько раньше осенней и дружнее: в Амуре главным образом четырех летками, иногда трехлетками, тогда как в нерестовой популяции осенней кеты значительным удельный вес занимают пятилетки и. четырехлетки небольшое число особей созревает семилетками, а на юге ареала - двухлетками (Ловецкая, 1948; Бирман, 1951, 1956; Sano, 1959; Воловик, Ландышевская,1968).
Сроки нерестового хода кеты, как указано в цитированных выше источ�никах, растянуты. Наиболее ранние подходы отмечаются в восточнокамчатском районе, в начале второй декады нюня. В ана�дырском районе подходы поздние, с середины июля до конца августа. В западнокамчатском районе кета появляется в двадцатых числах июля. В реки залива Терпения производители заходят с конца июля - начала августа до конца сентября. В лимане Амура ход летней кеты начинается с первых чисел июля, через несколько дней становится массовым и остается таким около ме�сяца, а заканчивается - в середине или конце августа. Самки лет�ней кеты заходят в реки близко ко времени завершения в овоцитах процесса вителлогенеза (III, III-IV стадии зрелости). У самцов в это время активно идет процесс сперматогенеза. В ампулах се�менников, помимо сперматогоний, обнаруживаются сперматоциты и сперматиды (Иевлева, 1964).
В Амуре первые экземпляры осенней кеты появляются в ию�ле - середине августа, рунный ход приходится на конец августа - начало сентября, последние производители заходят в сере�дине сентября. При входе в Амур половые продукты у осенней ке�ты менее зрелы, нежели у летней. Нерест наблюдается с середи�ны сентября по декабрь включительно. Нерестилища осенней ке�ты имеются в небольших реках лимана н нижних притоках Аму�ра, но основная масса рыбы поднимается в притоки среднего тече�ния, главным образом в реки Гур, Тунгуска, Анюй, Бира, Биджан. Важнейшие нерестилища расположены в притоках р. Ус�сури (Хор, Б. Уссурка, Бикин). Значительное количество особей до�стигает р. Кумары а отдельные экземпляры поднимаются до рек Аргунь и Онон. В реки Сахалина осенняя кета заходит с конца ав�густа - начала сентября, в массе - со второй половины сентября или начала октября, заход длится до середины ноября. В бассейне р. Найба ход и нерест сильно растянуты. Первые экземпляры начинают заходить в конце августа, при среднесуточной температуре воды около 15°С. Затем примерно на 15-20 дней ход прекращается. Подход возоб�новляется с конца второй декады сентября, когда температура воды снижается до 12-13°С. В конце сентября суточные уловы до�стигают нескольких сот экземпляров. Обильные подходы совпадают с повышением уровня реки, ускорением течения и охлаждени�ем воды примерно до 10°С. В начале октября интенсивность хода несколько ослабевает, но с середины октября ход становится особенно обильным и сохраняется таким до начала декабря. Продвижение кеты к нерестилищам наблюдается до двадцатых чисел де�кабря. К декабрю температура воды снижается до 0.5—0.2° С. Рыба движется к местам выхода грунтовых вод, которые в это время имеют температуру в 5-7 °С, а местами и выше. В реках о. Хоккайдо ход кеты продолжается весь декабрь, наблюдается и в январе (Sano 1959, 1966).
В отличии от Амура и других крупных рек в небольших реках Южного Приморья, Сахалина, Хоккайдо во время раннего хода производители кеты заходят лошалыми, на (римскими 4-5 и 5) стадиях зрелости, а нерест наблюдается на первых километрах от моря.
В начале созревания чешуя утрачивает серебристый блеск. Тело принимает буровато-желтый оттенок, спинка темнеет. По мере продвижения по реке и созревания на боках появляются 6-7 и более поперечных светло-лиловых полос, перекрашивающихся затем в темно-лиловые или темно-малиновые. На брюхе они сливаются в сплошную продольную полоску. По бокам яркие цветные полосы чередуются с темными. Со временем брюхо, голова и спинка становятся почти черными. К концу нереста темнеют нёбо, язык, основания жаберных дуг. Одновременно с изменением окраски кожа утолщается, в не погружается чешуя, которая частично резорбируется. На челюстях, нёбе, сошнике и языке вырастают крючковатые зубы, более крупные у самцов. Челюсти удлиняются и у самцов изгибаются. Тело самцов уплощается с боков, на спине вырастает небольшой киль. Преднерестовые изменения больше затрагивают самцов, причем наиболее крупных. Замечено, что брачный наряд проявляется особенно ярко у особей, нерестящихся в слабопроточных ключах, вода которых содержит мало кислорода. Пропорции тела меняются в меньшей степени.
В пресной воде производители, как известно, не питаются; развитие гонад и продвижение рыбы в верховья идет за счет питательных веществ, накопленных в море. За время продвижения по Амуру на 1200 км самки расходуют 8%, а самцы 5,8% жира (Пентегов, и др, 1928)
Летняя кета в бассейне Амура и в сахалинских реках, как правило, в массе нерестится в нижнем и среднем течениях рек. Кета выбирает для нереста более глубокие места (от 60 до 100 см) и преимущественно ближе к берегам, где есть тень и лежат подводные карчи (Кузнецов, 1928). Летняя кета нерестится в удалении от верховьев, на более глубоких плесах. В таких местах течение спокойное: на р. Мы, впадающей в лиман Амура, наблюдалась скорость течения от 10-20 до 60-80см/сек. Грунт нерестилищ подвергается значительному заилению, ухудшающему водоснабжение нерестовых бугров (Никольский, 1952; Никольский, Соин, 1954; Васильев, Юровицкий, 1958; Стрекалова, 1963).
Летняя Амурская кета нерестится в самое теплое время, когда в устьях рек температура колеблется от 9,8 до 13,6С , а в верхнем районе нерестилищ снижается до 6-7С (Кузнецов, 1937). Икра, выметываемая в начале нереста, довольно долго развивается при температуре 12-14С и ее инкубация длится около месяца. В конце нереста, в сентябре, некоторое время сохраняется температура 9-10С, а в октябре она падает до 3С, а затем приближается к нулю (Соин, 1954).
Брачные игры лососей, процесс закапывания икры описывали многие авторы ( Rutter, 1904; Chamberlain, 1907; Солдатов, 1912; Кузнецов, 1928; Семко, 1954; Sano, 1959; Стрекалова, 1963; Барсуков, 1964). В избранном участке самка останавливается у дна головой против течения, поворачивается на бок и производит хвостом энергичные волнообразные движения, перемещаясь вперед. Мощные удары хвоста поднимают со дна ил, песок, мелкие камни. Течение реки способствует сносу поднятого со дна материала. Однако выбивание ям и устройство нерестовых бугров успешно осуществляется лососями и при отсутствии течения. Когда образуется ямка значительной величины, в нее вместе с самкой заходит самец (иногда два или даже три самца). Около нерестящихся самок держится по нескольку самцов. Самка и чаще самый крупный самец останавливаются рядом, сближаются, опускают хвосты ко дну и по их телу проходят волны конвульсивных движений, которые способствуют выметыванию икры и молок. Икру, политую молоками, самки тут же начинают закрывать грунтом, действуя как и при устройстве ямки. Летняя кета выметывает икру двумя» чаще тремя порциями, каждая из которых в общем нерестовом бугре располагается таким образом, что омывается свежей во�дой. Рассредоточение икры повышает шансы на выживание по�томства. Закладывается она на значительную глубину: в рыхлой грунте до 30—40, даже 50 см, в твердом - ближе к поверхности.
В процессе икрометания расчищается площадка длиной до 3 м и шириной 2,5 м. Чаще эти величины составляют соответственно 1,2 и 1,0 м. Грунт нерестовых бугров оказывается очищенным не только от ила и мелкого песка, но и определенным образом стра�тифицированным. На глубине закладки икры он крупнее и икринки попадают в щели между камнями. Затем следует мелкая галь�ка, а на поверхности оказываются камешки среднего размера. Икрометание обычно совершается ночью, реже в сумерки и днем. Процесс откладки икры длится в 50% случаев 3~5 дней, но мо�жет совершиться за сутки или растянуться на неделю. Гибель са�мок после икрометания наступает чаще через 9—14 дней, при варь�ировании этого срока от 4 до 22 суток (Кузнецов, 1928. 1937). Самцы после вымета икры одной самкой переходят к другим и участвуют в нересте с несколькими самками. Для кеты типичны массовые нерестилища.
Икра в гнездах летней амурской и сахалинской кеты омывается в основном так называемым подрусловым потоком, обновляю�щимся за счет поступления в него русловой воды. Отмечается не�большая примесь грунтовых вод поверхностного залегания (Лева�нидов, 1969). Разрыхление и очистка грунта нерестовых бугров улучшает, его проницаемость.
Осенняя кета крупнее летней и ее нерестовые бугры больше. На рыхлом грунте и при слабом течении выкапываются глубокие короткие ямы, немного превышающие длину самок. При крупном камне и быстром течении нерестовые бугры принимают удлинен�но-овальную форму. Их ширина обычно составляет 1,5 м, длина 2.5 м. Ширина бугров колеблется от 106 до 213 см, а длина - от 125 до 320 см. От поверхности грунта икра залегает на глубине до 30-40 см, иногда глубже 50 см (Кузнецов, 1929, 1937). Сред�няя площадь нерестовых бугров в сахалинских реках равна 1,6 м2 (Рухлов, 1969).
Длительное развитие в хорошо защищенных условиях обычно обеспечивает высокую выживаемость лососей и хорошее пополнение стад. Однако наблюдается и падение численности этих рыб, принимающее временами катастрофический характер. Анализ причин массовой гибели по�томства указывает на доминирующее влияние изменчивости усло�вий жизни в пресных водах; морская среда представляется более стабильной (Леванидов, 19646, 1968, 1969).
В теле отнерестившихся самок, как правило, остаются единич�ные икринки их количество редко превышает 1 - 1,5% плодовитости. Иначе обстоит дело с потерями во время нереста. В бассейне Амура они составляют 21- 31%. (Леванидов, 1954), на Камчатке достигают 68% (Семко, 1954), на Сахалине - 75% (Рухлов, 1969) от средней плодовитости. Основ�ные факторы смертности — промерзание, обсыхание в маловод�ные годы, заиление и снижение проточности гнезд. В Амуре у летней кеты и горбуши насчитывали 25- 50% гнезд с полностью погибшей икрой; в суровые годы в отдельных реках промерзает до 90—100% гнезд горбуши (Бирман, Леванидов, 1953). Посколь�ку в бугры летней кеты поступает некоторое количество грунто�вых вод, опасность их промерзания снижается.
Условия развития осенней кеты, стабильнее. У нее промерзает и заиливается от 10 до 20% гнезд
Развитие кеты.
Половые продукты.
Сперматозоиды кеты, так же как у родственных ей видов и других костистых рыб, жгутикового типа. Созревшие сперматозоиды лососи выделяют отдельными порциями в течении продолжительного времени .
Кета выметывает очень крупные икринки. Такой средний вес зрелых яиц колеблется от 183 до 300 мг. Диаметр набухших икринок осенней сахалинской кеты колеблется от 6,7 до 9,0 мм, модальная величина – 7-8 мм.
У осенней кеты икринки ярко-оранжевые. Сравнение цвета икры разных самок с цветовыми полями Атласа цветов (Рабкин. 1956) позволяет охарактеризовать вариации цветового тона длиной волны 589—592 мкм. Окраска обусловлена каротиноидным пиг�ментом. Оплодотворенные икринки камчатской кеты содержат 1,60-1,76% астаксантина (Яржомбек, 1970).
Время, в течение которого яйца кеты в воде сохраняют способность к оплодотворению (фертильность), указывается от 2 до 30 мин (Kusa, 1950; К. Yamamoto, 1951).В связи с этим важно подчеркнуть, что основная масса икринок после активация водой быстро утрачивает фертильность.
1.2 Эмбриональное развитие
В морфологии под стадией развития принято воспринимать любое произвольно взятое состояние организма. (Васнецов, 1953; Крыжановский 1953).
Группы этапов, характеризующиеся общими приспособления�ми, в свою очередь, объединяются в периоды развития.
B эмбриональный период онтогенеза зародыши питаются ма�теринскими запасами пищи, заключенными в желтке. Такое эн�догенное питание свойственно не только зародышам, находящим�ся под оболочкой икринок, или собственно эмбрионам, но сохра�няется определенное время и после их вылупления у свободных зародышей. По этой причине данный интервал онтогенеза можно также называть зародышевым периодом, или периодом эндоген�ного питания.
Важнейшее биологическое отличие личинок состоит в переходе к активному добыванию пищи из внешней среды и наличии провизорных личиночных органов. Задаче активного добывания пищи подчинены приобретаемые личинками морфологические, фи�зиологические и экологические особенности, сохраняющиеся в течение личиночного периода развития. Лососи подготавливаются к началу активного питания, имея еще большой остаток желтка, и для их личинок характерен продолжительный срок смешанного эн�догенно-экзогенного литания. При отсутствии доступной пищи вре�мя смешанного питания сокращается.
К мальковому периоду жизни провизорные личиночные орга�ну исчезают и появляются некоторые характерные для взрослых рыб органы (например, непарные и парные плавники со скелетом и мускулатурой, чешуя). Соответственно меняется среда обитания и поведение.
Переходим к характеристике половых продуктов и развития кеты.
В течение зародышевого периода развитие осуществляется за счет запасов, заключенных в желтке, и поэтому его называют также периодом экзогенного. Тот интервал развития, который эмбрионы проводят в икринке, часто называют инкубационным.
Существование дочернего организма начинается со слияния ядер родительских гамет и образования зиготы. Этому предшествуют сложные процессы, изученные у кеты достаточно подробно.
Осеменение, активация икры, образование зиготы.
Зрелые икринки кеты находятся на стадии метафазы второго деления созревания. Это типично для зрелых ооцитов костистых рыб. Дальнейшее продвижение мейотического процесса осуществляется после встречи сперматозоида с яйцом и его активации с водой. При температуре 7,5-9,0 ° С преобразование ядра яйциклетки завершается примерно через 3 часа после осеменения образуется зигота.
Преобразование ядерного аппарата родительских гамет совпадает по времени с обводнением яиц, образованием перивителлинового пространства и затвердением оболочки. Процесс обводнения начинается после активации яиц водой и выделения под оболочку осмотически активных в веществ, содержащих в альвеолах кортикального слоя цитоплазмы. Кортикальные альвеолы яиц кеты имеет диаметр от 3 до 30 мк и содержит осмотически активные кислые мукополисахариды, а так же небольшое количество белков.
У лососей полостная жидкость служит мощным активатором и сщ мии сохраняют в ней подвижность значительно дальше, нежели в воЗ В полостной жидкости и в изотоническом растворе спермии лосося способны проникать в яйца. Известно, что для вовлечения спермы в дальнейшее развитие необходима активация яиц водой. Указанным особенностям половых продуктов отвечает методика «сухого», называемого также русским, способа искусственного оплодотворения икры разработанного В.П. Врасским. В настоящее время он применяется в несколько измененном виде. Икра перемешивается с неразбавленной спермой, которая берется от нескольких самцов. Затем приливается немного воды и перемешивание повторяется. После смены воды икра выдерживается при полном покое для набухания. При температуре воды 5-7°С обводнение яиц завершается в течение от 40-50 до 60 мин. В результате обводнения масса яиц кеты увеличивается на 13,4-18%.1 Через 10-15 мин. после помещения в воду икринки приобретают слабую клейкость. Они могут соединяться в группы и прикрепляться к частицам грунта, что в какой-то мере предохраняет их от вымывания из гнездовых ям. При искусственном оплодотворении к икринкам могут прилипать погибшие сперматозоиды, сгустки крови, слизь и другие органические частицы, которые оказываются субстратом для развития сапролегнии. Клейкое вещество смывается при смене воды. Промывку икры следует производить в течение 5-10 мин. после осеменения, пока икра имеет низкую чувствительность к механическим воздействиям. После указанного срока чувствительность икры начинает повышаться.
До попадания в воду икра стойко переносит значительные механические воздействия. Этим обеспечивается сохранность икры при точках самок во время выбивания нерестовых ям, и закапывания. Это же качество позволяет успешно транспортировать зрелых наркотизированных или умерщвленных самок, как и неоплодотворенную икру, помещенную в специальную изотермич6скую тару.
У набухших икринок куполообразное выпячивание, на котором формируется цитоплазматический бугорок, или зародышевый диск. Благодаря полярности строения яйцо всегда располагается анимальным полюсом кверху.
Необходимо заметить, что в случаях передержки зрелых самок некоторые икринки могут начинать обводняться еще в полости тела, их оболочки затвердевают, жировые капли сливаются, укрупняются. Такую перезревшую икру рыбоводы образно называют горохом. Она не способна оплодотворятся, а ее присутствие указывает на общее снижение качества икры.
1-й этап. Образование цитоплазматического бугорка-зародышевого диска
За начало первого этапа онтогенеза принимают образование зиготы. Продолжается этап до начала дробления. При средней температуре 3,4 С. Этот этап длится около 20 часов, при 9,6 С у одних икринок начало дробления отмечено через 8 часов, у других - через 10 часов после осеменения.
В течении этапа на анимальном полюсе икринок, в месте скопления жировых капель формируется цитоплазматический бугорок. У зрелых яйцеклеток на анимальном полюсе выявляется лишь небольшое скопление цитоплазмы. В первые часы после осеменения икринок процесс концентрации цитоплазмы протекает медленно. Сформировавшийся зародышевый бугорок принимает форму, близкую к полушарию. Его контур четко очерчивается. Диаметр сформировавшегося зародышевого диска у осенней кеты варьирует от 0,9 до 1,3 мм.
К концу этапа нижняя поверхность желточного мешка приобретает способность прилипать к оболочке. Между поверхностью анимального полюса яйцеклетки и оболочкой образуется пространство, достигающее 0,4 – 0,8 мм.
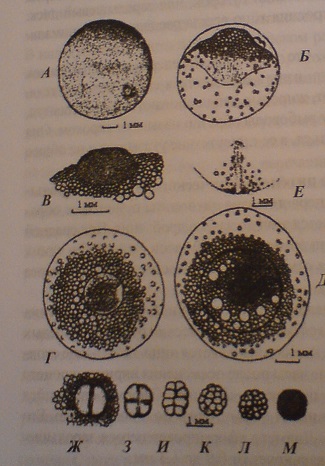
Рисунок 2 - 1-й этап. Образование цитоплазматического бугорка-зародышевого диска.
2-й этап. Дробление зародышевого листка
Этап начинается с появления первой борозды дробления и заканчивается образованием бластомерной бластулы, которая формируется при 3,3°С в возрасте 8 суток, а при 10,4°С – в возрасте 3 суток (около 3 градусодия). Образовавшиеся два бластомера по форме напоминают семена бобов. Вторая борозда дробление проходит перпендикулярно первой, образуя четыре плотно прилегающих друг к другу бластомера. Две третьих борозды дробления проходят параллельно первой и образуют 8 бластомеров, располагающихся двумя параллельными рядами. Зародышевый диск на этой стадии принимает форму удлиненного овала и его площадь несколько увеличивается. У кеты тепловского стада на этой стадии два, три или четыре бластомера лежат над другими, непосредственно прилегающими к желтку.
Борозды четвертого дробления проходят уже в горизонтальной (экваториальной) плоскости, и образованием 16 бластомеров располагаются двумя слоями. Нижние бластомеры крупнее верхних. При просмотре дробящихся икринок можно видеть не только 4, 8, 16, 32, но и также 3, 6, 12, около 30 и иное число бластомеров, что указывает на необязательность синхронного деления всех бластомеров. После 4-5-го дробления зародышевый диск принимает округлую форму.
В процесс дальнейшего дробления верхние бластомеры располагаются плотнее, они немного мельче и уплощены. Нижние бластомеры крупнее, делятся медленнее, расположены рыхло. Те из них, которые лежат на периферии бластодиска, сохраняют связь с краевой цитоплазмой. Часть цитоплазмы нижних бластомеров заходит между глыбками желтка, давая начало центральному участку перибласта.
Под зародышевым диском и в непосредственной близости от него расположены более мелкие и жировые капли. Если яйцо быстро перевернуть, они перемешаются вместе с бластодиском. Более крупные жировые капли, расположенные по периферии, перемещаются в желточном мешке легко, независимо от бластодиска.
С началом дробления повышается чувствительность икры к механическим воздействиям. Поэтому к началу дробления икра должна быть размещена в аппараты и инкубироваться при полном покое.
З-й этап. Бластула
Продолжается от образования бластомерио бластулы до начала гаструляции. При температуре 3,3°С этап длится 10-11 сут., при 10,4°С-2,5-3 сут. Одним из внешних признаков перехода к стадии бластулы служит начало увеличения бластодиска. У кеты диаметр бластодиска в это время составляет 1,3-1,4 мм. Он куполообразно выдается над поверхностью желточного мешка и охватывает зону мелких жировых капель.
На стадии бластулы хорошо выражен перибласт, формирующийся в результате деления бластомеров, прилегающих к желтку. При этом одно из новых ядер образует типичный бластомер, а второе ядро проникает в промежуточный слой цитоплазмы, отделяющей бластодиск от желтка.
Поверхность бластулы покрыта слоем плотно прилегающих друг к другу клеток полигональной формы, под которыми рыхло располагаются более крупные клетки. Начальную стадию бластулы лосося называют бластомерной бластулой. В дальнейшем наружный слой клеток уплотняется, и к нему прилегают нижележащие бластомеры наблюдается процесс эптилеизации. Поздняя бластула названа эпите�лиальной бластулой. Диаметр поздней бластулы у осенней амурской кеты достигает 2,3-2,6 мм. Яйца сахалинской осенней кеты крупнее, и у нее диаметр бластулы достигает 3,0-3,2 мм.
4-й этап. Образование зародышевых пластов
В течение этапа протекает процесс гаструляции. При средней температуре 10,4°С, образование зародышевых пластов начинается на исходе шестых суток. Этап заканчивается через двое суток, когда образуются первые мезодермальные сегменты. При 3,4°С, гаструляция начинается в возрасте 18-19 суток и завершается примерно через 6 суток.
Во время гаструляции зародышевый диск увеличивается, уплощается, в центральной части становится тонким. В одном из краевых участков бластодиска, будущем каудальном конце формирующегося зародыша, образуется утолщение - «краевой узелок». Методом прижизненной мар�кировки участков бластодисков установлено, что поверхностные клетки бластодермы у лососевых и других костистых рыб распространяются до поверхности желточного мешка исключительно путем эпиболии, не подворачиваются под зародышевый диск и не принимают участия в формировании гипобласта. В определенное время внутренние клетки разрастающейся бластодермы поодиночке или группами отделяются от нее, мигрируют в направлении зачатка тела и образуют гипобласт, а клетки, лежащие непосредственно под эпителиальной поверхностью бластодермы, смещаясь единым пластом, формируют эпибласт.
При диаметре зародышевого диска около 3,5 мм его края переходят зону расположения мелких жировых капель «Краевой узелок» удлиняется и превращается в «зародышевый язычок». За�родышевые пласты обособляются четче. По данным Манон и Хо�ра (1956), эпидермальный слой, зародыша представлен уплощен�ными клетками. Под ним лежат более крупные клетки эктодермы. Непосредственно под эктодермой, по продольной оси зародыша, плотным тяжем располагаются клетки мезодермы. Между ними и перибластом разбросаны клетки энтодермы, которые несколько позже уплощаются. В переднем конце зародыша эктодерму под�стилает выделяющийся слой клеток прехордальной пластинки. B ядрах центрального перибласта митоза уже не наблюдается, они делятся амитотически. В начале гаструляции Манон и Хор обнаруживали группу заметно увеличенных клеток с выростами, подобными псевдоподиям амебы. Эти амебообразные клетки пред�положительно были названы примордиальными зародышевыми клеткам (Персов, 1966, 1968) относит появление первичных половых клеток у кеты к несколько более позднему возрасту.
Когда диаметр зародышевого диска достигает 4 мм, зароды�шевая полоска увеличивается до 1,5 мм. На поперечных срезах такого зародыша, сделанных несколько впереди его каудального выступа, в центральной части выявляются концентрически рас�полагающиеся клетки зачатка хорды. По бокам от него лежит недифференцированная боковая мезодерма. Под зачатком хорды и немного сбоку образуется полость, окруженная цилиндрическими клетками, представляющая зачаток Купферова пузырька. Снару�жи по сторонам продольной оси эмбриона утолщенный слой эктодермальных клеток образует небольшие продольные выпячивания - нервную пластинку.
Зародышам рыб на ранних этапах развития свойственна протоплазматическая моторика, наблюдавшаяся многими исследователями. Движения яиц под оболочкой обусловлены сократительной активностью цитоплазмы, покрывающей желточный мешок. Вол�ны ее сокрушений вызывают деформацию и смещения центра тяжести яйца что приводит к изменению его положения в прост�ранстве. У лососей на ранних этапах развития анимальный полюс яйца описывает круговое движение по направлению или против движения часовой стрелку При низкой температуре уловить эти движения, наблюдая живое яйцо под микроскопом, трудно. На данной стадии движения становятся заметнее. Во время наблюдений бластодиск кеты описывал полный «круг» вращения за 4 мин. 40 сек. - 5 мин. В результате подобных дви�жений яйца, а позже и самого эмбриона перемешивается перевителлиновая жидкость и интенсифицируется внешнее дыхание зародышей. С началом гаструляции чувствительность икры к механиче�ским воздействиям возрастает и одинаковое воздействие на стадии хорошо выраженного «зародышевого узелка» вызывает примерно вдвое больший отход, чем после воздействия на стадиях дробления.
5-этап. Формирование головы и туловища зародыша
Этап охватывает интервал развития от начала сегментации туловищной мезодермы до образования 46—47 сегментов при средней тем�пературе 3,2 С он начинался на 24-25-е сут. и заканчивался на 35-36 сегментах.
Первые сомиты формируются у зародышей длиной 2 мм, когда диаметр зародышевого диска увеличивался да 4,5 мм. Образование трех сомитов и начало дифференцировки отделов головного мозга отмечено при длине эмбриона 2,5 мм. С достижением длины 3 мм число сомитов возрастало до семи (при 3,4°С на 26-е сутки). Бластодиск к этому времени охватывал до половины окружности яйца. У эмбрионов длиной около 3,5 мм число сегментов возрастало до 13.
При длине эмбриона 3,0-3,5 мм в зачатках глаз образуются полоски, превращаются в глазные пузыри. Вскоре дистальная часть их стенки инвагинирует, и образуются глазные бокалы. Над передним концом хорды благодаря инвагинации внутренних слоев эктодермы появляются зачатки слуховых пузырьков. В момент закладки они отдалены от переднего конца зародыша на значительное расстояние. В основании головного мозга на уровне заднего края глаз закладывается воронка (infundibulum). Несколько позади этой области эктодермальный зачаток подгибается внутрь, давая начало глоточным карманам. Из нижнего участка мезодермы начинает формироваться зачаток сердца. Мезодермальные боковые пластинки делятся на соматический и спланхнический слои, узкая щель между которыми указывает на образование псрикардиального целома. Продольное скопление энтодермальных клеток в средней части тела зародыша образует зачаток кишки.
У зародышей длиной 4 мм сомиты дифференцируются на дерматом миотом и склеротом. Формируется пронефрическая складка. Вдоль склеротома располагаются клетки, дающие начало осевым кровенос�ным сосудам и клеткам крови.
С того времени, когда бластодермой покрывается более половины поверхности желточного мешка, чувствительность икры к толчкам на некоторое время снижается.
6-й Этап. Обособление задней части тела зародыша от поверхности желточного мешка
Этап продолжается от формирования 46-48 до 65 сегментов. Существенная функциональная особенность этапа состоит в становлении нервномышечной моторике эмбрионов.
Внешний вид зародыша в начале этапа на рис 3. К этому времени процесс эпиболии (обрастание желточного мешка бластодермой) приближается к завершению. Начала дифференцироваться хвостовая почка. Впереди глаз выявились обонятельные плакоиды. Наметились зачатки жаберных крышек. Сердечная трубка становится изогнутой, наблюдаются редкие, слабые сокращения ее стенок. Частота пульсации сердца быстро нарастает, обеспечивая перемешивание полостной жидкости. Миотомы приобрели способность сокращаться. Потребление кислорода икрой заметно увеличивается, что отвечает возросшим потребностям эмбрионов, которые достигли значительного размера.
Заложились осевые кровеносные сосуды и в перикардиальной полости появились эмбриональные кровяные клетки. Вдоль средней линии тела выявляются зачатки органов боковой линии.
После отчленения хвостовой почки быстро нарастает число хвостовых сегментов. Хвост удлиняется, на нем появляется зачаток непарной плавниковой складки. С началом отделения от желтка задней части туловищного отдела развивается прианальная складка.
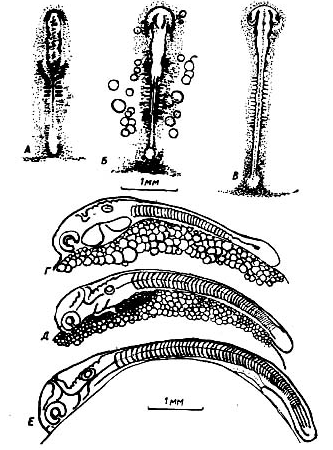
Рисунок 3- Стадия начала сегментации мезодермы и формирования головы и туловища зародыша осенней кеты
Во время закрытия желточной пробки икринки кеты чрезвычайно чувствительны к толчкам и погибают даже от слабых прикосновения. Эпиболия завершается у разных зародышей не одновременно, и пока этот процесс не закончится, икра должна находится в абсолютном покое.
Эмбрионы кеты имеют очень большой желточный мешок, и его обрастание затягивается до поздних стадий развития. В самых мелких икринках кеты эпиболия завершается при 52-53 сегментах.
После завершения процесса эпиболии чувствительность икры к травмам резко падает. У кеты это происходит раньше появления пигмента в глазах.
К моменту исчезновения желточной пробки, хрусталики отделяются от эктодермы и погружаются в глубь глазных бокалов. Значительно продвигается дифференцировка головного мозга: увеличиваются его желудочки, закладываются обонятельные доли, формируются крупный мозжечок, эпифиз и гипофиз. Намечается дифференцировка первых жаберных дуг. Энтодерма образует кишечную трубку. Пальцеобразный вырост средней кишки дает начало печени. Начинают формироваться почечные клубочки и пронефрический проток. На желточном мешке проявляются кровяные островки.
К концу этапа задняя часть туловищного отдела до 29-го сегмента отделяется от желточного мешка. В течение этапа увеличивается подвижность эмбрионов. Они проводят из стороны в сторону хвостом и временами совершают им волнообразные движениям
7-й этап. Развитие подкишечно-желточной системы кровообращения
Этап охватывает интервал от начала кровообращений формирования кардинальных вен. При средней температуре 9,6°С длится 1,5-2 сут., при 3,5°С - около 5 сут. К концу этапа эмбрионы достигают длины 7,3 - 7,8 мм и у них формируется до 71 сегмента. В начале кровообращения ток крови в сосудах слабый, эритроцитов мало и они имеют светлую окраску. Присутствие гемоглобина у лососей выявляется до начала тока крови.
В начале этапа от сердца кровь течет по парным мандибулярным дугам аорты. Над жаберной областью от них ответвляются сосуды, по которым кровь течет в голову и по спинной аорте в туловище. Позади ануса спинная аорта переходит в подкишечную вену, по которой кровь течет на протяжении 25-26 сегментов. Чуть впереди прианальной складки подкишечная вена выходит на левую сторону поверхности желточного мешка, давая начало желточной вене. На левой стороне желточного мешка поток крови описывает небольшую дужку, объединяется с кровью, идущей из передней кардинальной, попадает в предсердие.
К этому времени закладываются плавнички. Голова начинает отделяться от поверхности желточного мешка рис. Уже в начале этапа кровь текла по хвостовой вене до 10-го хвостового сегмента, и у заднего участка подкишечно - желточной вены имелись два или три капиллярных сосуда. Это более поздняя стадия дифференцировки. Задержку начала кровообращения можно объяснить меньшим размером икринок и лучшей аэрацией воды.
В течение этапа хвостовые артерия и вена удлиняются, подкишечно-желточная вена опускается ниже, число капилляров на желточном мешке увеличивается. Желточная вена смещается вниз следующим образом. Постепенно количество форменных элементов в кровяных островках, расположенных вблизи внешней стороны вены,увеличива�ется, они сливаются и включаются в кровоток, тогда как внутренний участок сосуда постепенно запустевает. Новые капилляры образуются также за счет слияния кровяных островков.
Развивающиеся зародыши двигаются все энергичнее и чаще. С началом кровообращения потребление икрой кислорода по сравнению с предшествующим этапом почти удваивается. Икра стойко переносит механические воздействия.
8-й этап. Возникновение кардинальных вен и смешанного подкишечно-желточного и печеночно-желточного кровообращения
Данный этап длится 5-14 суток. К его завершению эмбрионы достигают 10,5-11,0 мм.
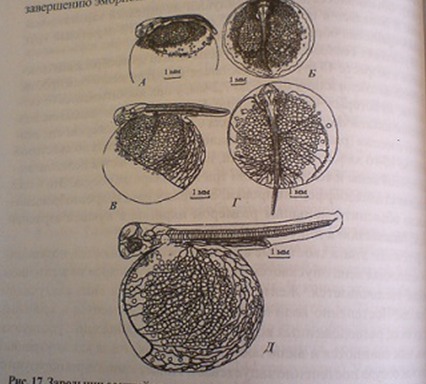
Рисунок 4 - Зародыши осенней кеты на разных стадиях развития подкишечно-желточной системы
К началу этапа сосуды обильно заполняются эритроцитами, имеющие интенсивную окраску. Начинается кровоснабжение глаз. Из головы кровь отводится передними кардинальными венами и образуются короткие кьюверовы протоки. В сосудах двух первых пар жаберных дуг появляются кровяные элементы, но тока крови в них еще нет. На значительной части задней половины желточного мешка образуется сеть каппилярных сосудов. Они имеются теперь не только на левой, как было ранее, но и на правой стороне.
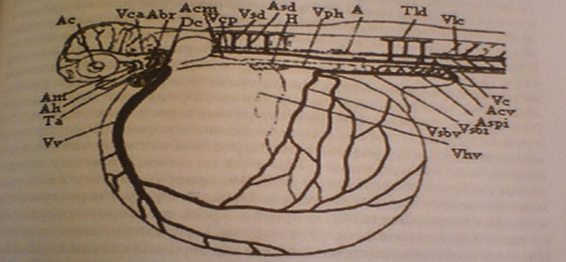
Рисунок 5 - Схема расположения основных кровеносных сосудов эмбриона лосося
У живых зародышей позади грудных плавничков хорошо просматриваются клубочки пронефроса.
К этому времени заметно вырастают грудные плавники. На спине непарная плавниковая складка разрастается до первых миотомов. Голова обособляется от желточного мешка. В связи с образованием изгибов головного мозга слуховые пузырьки сблизились с глазами и длина головы уменьшается. В слуховых пузырьках идет формирование полукружных каналов, образовалось по нескольку мелких отолитов. В глазах можно заметить зерна меланина. Формируются обонятельные ямки. Появился ток крови в первых парах жаберных дуг. На желточном мешке кровеносные сосуды заняли значительную площадь. Краевая желточная вена опустилась на нижнюю сторону желточного мешка. Печень достигла довольно крупного размера, расположена между 5-м и 9-м сегментами, причем сдвинута немного вправо. Возникло печеночно-желточное кровообращение. Первоначально из печени на левую сторону желточного мешка кровь течет по одному тонкому сосуду и впадает в краевую желточную вену. На правую сторону желточного мешка из печени выходят два сосуда. Первый образует короткую петлю и впадает в правый кьюверов проток, кровь из второго сосуда течет к краевой желточной вене. Эти сосуды вскоре начинают ветвиться. Подкишечно-желточная вена, снабжающая кровью сосуды каудального участка желточного мешка, после образования задних кардинальных вен и печеночного кровообращения постепенно запустевает. Вместе с тем на желточном мешке возрастает количество сосудов, снабжаемых кровью из печени. Таким образом, для описываемого этапа характерно смешанное кровоснабжение желточного мешка при постепенном возрастании роли печеночного кровоснабжения.
9-й этап. Функционирование печеночно-желточной системы кровообращения
Этап длится с переключением функции кровообращения желточного мешка полностью на печеночные вены до начала формирования конусов миотомов.
К началу этапа увеличивается высота головы зародышей. Слуховые капсулы приближаются к глазам. Глаза становятся темными. Многочисленные мелкие отолитики объединяются в крупные образования. Наметившаяся ранее воронка ротовой полости углубилась. Ускорился рост нижней челюсти. Заметно разрастаются жаберные крышки. Основания грудных плавников образуют с продольной осью тела угол около 30°. Удлинившийся хвостовой отдел превышает 1/3 общей длины эмбриона. Задний конец хорды изогнулся вверх, и хвостовой плавник уфатил симметричность. Начали формироваться опорные и мышечные элементы хвоста рис. Число хвостовых сегментов сокращается до 27-28. Аналогичный процесс свойственен и другим костистым рыбам. В хвостовом отделе идет обособление позвонков. Содержимое кишечника приобрело лимонно-желтую окраску, обусловленную, по- видимому, появившимися желчными пигментами. У эмбрионов четко различаются предсердие и желудочек. Обнаруживается движение крови в гиоидных дугах, в третьей, а немного позже и в четвертой парах жаберных дуг. Развивается сеть капилляров в покровах теменной области головы, затем над мозжечком и в заты�лочной области. Появляются сегментальные сосуды, формирование которых идет от головы к хвосту. Хвостовая вена утрачивает связь с подкишечной, а последняя – с сосудистой системой желточного мешка. Задний отдел кишки оплетают разветвления надкишечной артерии, а средний отдел – каппиляры кишечной артерии. Из этих каппиляров кровь поступает в подкишечную вену, по которой направляется к печени. На желточном мешке развивается мощная сосудистая сеть, снабжаемая кровью исключительно через печеночные вены.
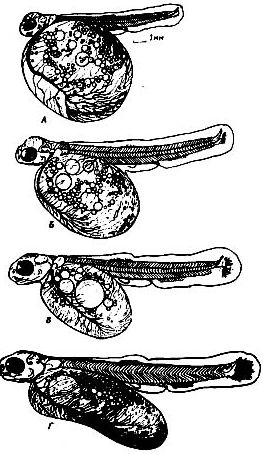
Рисунок 6 - Поздние эмбрионы осенней кеты
При длине эмбрионов около 13 мм в месте закладки анального и спинного плавников обнаруживается скопление мезенхимы и зачатки мускульных почек. Удлинившийся хвостовой отдел достигает 37% от общей длины эмбриона. В месте закладки опорных и мышечных элементов хвостового плавника появляются петли капилляров. Капилляры покрывают всю поверхность желточного мешка. У извлеченных из икринок зародышей желточный мешок принимает яйцевидную форму. Диаметр некоторых жировых капель достигает 1 мм.
10 этап. Дифференцировка верхних и нижних конусов миотомов
Верхние нижние конусы миотомов при средней температуре 3,3С начинали формироваться на 83-й день (около 270 градусодней). При температуре 3,3°С этап длился 12-17 сут., а при 10°С -5-6 сут.
В начале этапа эмбрионы имеют длину 14 мм и более. К этому времени жаберные крышки стали прикрывать 2-ю пару жабр. На 3-й, а затем 4-й и первой парах жаберных дужек появляются зачатки жаберных лепестков. Основания грудных плавников принимают почти вертикальное положение, и они приобретают подвижность. Под 25-30-й туловищными миотомами появляются небольшие продольные складочки зачатков брюшных плавников. В основания закладывающихся непарных плавников врастают выступы конусов миотомов, образующие впоследствии мышцы плавниковых лучей. Зачатки лучей первоначально представлены стержневидными сгущениями мезенхиматозных клеток. Процесс охрящевения приводит к образованию лепидотрихии, которые опираются на птеригофоры. У кеты дифференцировка анального плавника несколько опережает дифференцировку спинного. Постепенно разрастается брюшная лопасть хвостового плавника. В нем формируется до 8-10 лучей. Нижние остистые отростки последних позвонков образуют широкие пластинки - гипуралии.
В течение этапа густота капиллярной сети желточного мешка продолжает возрастать за счет увеличения числа анастомозов между сосудами. Краевая вена опускается на вентральную сторону желточного мешка. Капиллярные сосуды развиваются почти на всей верхней поверхности головы. Появляются каппиляры в зачатках псевдобранхий. Подключичная артерия образует кольцевые сосуды грудных плавников. Во всех хвостовых миотомах появляются сегментальные сосуды. От передних туловищных ответвляются сегментальные сосуды. От передних сегментальных артерий ответвляются сосуды идущие к спинному мозгу, в толщу мышечной ткани и к поверхности тела. В покровах тела образуется сеть каппиляров, несущая дыхательную функцию Ответвления сегментальных сосудов над миотомами образуют truncus longitudinalis dorsalis, у поверхности тела вдоль средней линии- truncus longitudinalis lateralis, они же питают кровяные каппиляры, проходящие у поверхности тела по межсегментальным миосептам. В начале этапа в покровах головы, а затем и туловища появляются меланофоры.
Развитие верхних и нижних конусов миотомов идет в направлении от хвоста к голове, в порядке, обратном возникновению миотомов. В такой же последовательности дифференцируются и позвонки. Более раннее усложнение формы миотомов хвоста обосновано его важностью для плавания, т.е. с функцией органа на более поздних этапах развития. Следует заметить, что движения хвоста важны для жизни эмбриона. Они благоприятствуют циркуляции крови и обеспечивают перемешивание перивителлиновой жидкости.
К концу этапа число хвостовых сегментов сокращается до 22 и их общее число уменьшается до 65-63. Непарная плавниковая складка в месте закладки непарных плавников расширяется, и в них формируются опорные элементы. В хвостовой лопасти развиваем довольно густая сеть капилляров. Передний край нижней челюсти достигает вертикали, опущенной от переднего края глаз. Выявляют контуры костных элементов верхней и нижней челюстей. Рот приобретает подвижность. Жаберные крышки прикрывают 3-ю пару жабр. Печень развивается в мощный орган, пронизанный густой сетью кровеносных сосудов. Заканчивается дифференцировка верхних нижних конусов миотомов. Обонятельные ямки приобретают удлиненную форму. Слуховые пузырьки приобретают удлиненную форму. Слуховые пузырьки принимают вид неправильных пирамидок. В головы появляются мелкие оранжево-красные липофоры. головы и под ней на передней части желточного мешка обознаются железки вылупления. К концу этапа длина зародышей увеличивается до 16-18 мм.
11-й этап. Развитие подвижности челюстей, жаберных крышек, завершение инкубации
Настоящим этапом завершается развитие эмбрионов под оболочкой. Длительность инкубации при разной температуре варьирует в широких пределах. В описываемой серии инку- икринок длилась 51-55 сут. (521-530 градусодней). Основная часть зародышей при вылуплении имеет длину 22-23 мм, а некоторые 24 мм.
В течение этапа продолжает формироваться ротовой аппарат. Ниж�няя челюсть вырастает за вертикаль переднего края глаз. Рот принимает полунижнее положение. Прорывается дно ротовой воронки. Жаберные крышки прикрывают все жабры. Намечается образование перемычки, разделяющей отверстие обонятельной ямки на переднее и заднее. Рот, жаберные крышки и жаберный аппарат приобретают подвижность. Дви�жения челюстей и жаберных крышек в икринке затрудняются тем, что крупная голова эмбриона оказывается вдавленной в желточный мешок.
Ко времени вылупления разрастаются лопасти грудных плавников и их вершины достигают 5-6-го миотомов. В спинном и анальном плавниках четко выделяются лучи. Плавники вычленяются из непарной плавниковой складки, которая заметно резорбируется. В месте складки жирового плавника намечается скопление мелких меланофоров и липофоров. Вершины увеличивающихся брюшных плавников приближаются к краю прианальной плавниковой складки. В хвостовом плавнике развивается 15-19 лепидотрихий.
Дорсальная поверхность зародышей заметно темнеет. Зерна пигмента в хроматофорах приобрели способность мигрировать, благодаря чему могут изменять окраску. На туловище меланофоры многочисленны до средней линии тела, а ниже единичны. Появились меланофоры между лучами хвостового плавника. Образуется ряд пигментных клеток над спинным мозгом (невральный ряд), вдоль крупных кровеносных сосудов (гемальный ряд), пигментируется выстилаясь по брюшной полости. Мелкие красно-оранжевые липофоры покрывают дорсальную поверхность головы и в небольшом количестве появляются на спине зародыша. Позади зачатков спинного и анального плавников липофоры проникают в основание непарной плавниковой складки.
К концу этапа капиллярная сеть на желточном мешке достигает максимальной густоты. Длина капилляров на 1 мм2 его поверхности достигает 7,5-8 мм. Усложняется сеть капилляров вблизи поверхности тела.
За несколько дней до вылупления в гонадах выделяются крупные зародышевые клетки диаметром 7,2-15,4 мк. В их ядрах наблюдаются фигуры митоза. Митотическое деление первичных половых клеток продолжается и после вылупления.
Эмбрионы способны производить хвостом энергичные волнообразные движения и при возбуждении начинают вращаться под оболочкой. Если такого зародыша извлечь из икринки, он способен подняться в толщу воды и продвинуться на несколько сантиметров вперед локомоции хвоста быстро ослабевают и эмбрион опускается на дно. При благоприятных условиях аэрации эмбрионы в икринке движутся редко, что помогает экономить запасы энергии. Перемешивание перивителлиновой жидкости в это время осуществляется в основном благодаря ритмичному движению грудных плавничков. Выполнение той же задачи служат движения рта и жаберного аппарата.
К концу инкубации весь мочеточник и мочевой пузырь заполняются накопленными и кристаллизовавшимися продуктами азотистого обмена. Во время инкубации икры через оболочку выделяется лишь небольшое количество аммиака. Для лососей, откладывающих икру кучно в грунт, где проточность слабая, отмеченная особенность имеет жизненно важное значение, так как благодаря этому не загрязняется вода нерестовых бугров.
Непосредственно перед вылуплением эмбрионы ведут себя беспокойно, энергично ворочаются. Под действием протеолитического фермента, выделяемого железками вылупления, яйцевые оболочки разрушаются ,и эмбрионы выходят наружу. Фермент вылупления выделяется в подоболочечное пространство за короткое время и быстро разрушает оболочку, подготавливает оболочку к действию фермента вылупления накопившаяся гиалуронидаза. Значение механических факторов в акте вылупления невелико.
Зародыши выходят в разные сроки из икринок, взятых даже от одной самки, и связано это с индивидуальными особенностями икринок. Вылупление длится 2-3 дня, но иногда оно растягивается до 7-10 дней.
12-й этап. Пассивное состояние свободных зародышей
Этот этап начинается после выхода зародышей из оболочек и длится при температуре 3,3°С - 5-7 дней, при температуре 9°С 3-4 дня. Вылупившиеся зародыши слабо реагируют на внешние раздражители и малоподвижны. Их длина варьирует в широких пределах. Размер зависит от величины икры и стадии развития. Так, из икринок среднего размера 8,7 мм зародыши выходят длиной 21,0-24,5 мм при модальной длине 22,5-23,0 мм, тогда как из икринок диаметром 7,9 мм вылупляются зародыши длиной 19-23 мм, при модальной длине 20,5-20,7 мм. Меньшие экземпляры выходящие из икринок сходного размера имели в хвосте на 1-3 луча меньше, нежели крупный зародыши.
При выходе из икринки кета имеет огромный желточный мешок рядной формы. В желтке содержится много мелких жировых капель и обычно одна крупная, диаметром около 3 мм. При изменении положения зародыша жировые капли в желтке по-прежнему могут перемешиваться.
Число туловищных сегментов осталось прежним, в хвосте к моменту вылупления оно сократилось до 22-21. Хвостовые миотомы стали шире вследствие чего длина постанального отдела составляла 36,5% от длины зародыша. Форма хвостовой лопасти асимметрична.
Непарная плавниковая складка продолжает резорбироваться. Из нее обособился спинной плавник, наметились контуры жирового плавничка. В спинном и анальном плавниках заложилось большинство эспидотрихии. Ко времени массового вылупления в хвостовом плавике обычно закладывается 17-19 лепидотрихий. Если же при вылуплении их количество менее 15это указывает на раннее вылупление. У зародышей образовался костный плечевой пояс. Грудные плавники крупные, округлой формы. В брюшных плавниках опорные элементы еще отсутствуют. Прианальная плавниковая складка ко времени вылупления достигает максимального развития.
Рот у вылупившихся зародышей занимает полунижнее положение. В слуховых капсулах четко различаются полукружные каналы и крупные отолиты. Статоакустические органы максимально сблизились с глазами. Жаберные крышки закрывают жабры. На первой жабре средние лепестки имеют длину 0,25 мм и становятся ветвистыми; в лепестках появились капиллярные сосуды. Закладываются жаберные тычинки, имеющие вид небольших бугорков.
В начале этапа желудочно-кишечный тракт представляет почти прямую трубку. Зачаток желудка, имеющий длину около 1мм, выделяется утолщенными стенками и некоторым сужением на границе с кишкой. На его будущем пилорическом отделе заметны почкообразные выпячивания - зачатки первых пилорических придатков. Длина кишки 7,5-8,0 мм - около 39% общей длины зародыша. Слизистая кишки приобрела складчатость. Зачаток плавательного пузыря вырастает до 1,1-1,2 мм.
Мочевой пузырь после вылупления начинает периодически опорожняться и заполняться вновь. Вместе с этим уменьшается количество кристаллов, обильно заполнявших мочеточник. Зародыши быстро освобождаются от накопленных продуктов обмена, и, если в питомниках проточность слабая, плотность посадки чрезмерно высокая, окружающая зародышей среда может ухудшиться.
Ко времени вылупления формируются темные пятна над глазами и слуховыми капсулами, меланофоры равномерно покрывают всю дорсальную половину тела, располагаются между лучами хвостового плавника. Усиливается и липофоровая пигментация головы. Липофоры появляются на всей дорсальной стороне тела и в хвостовом участке непарной плавниковой складки.
Органы боковой линии - невромасты имеют вид бугорков. Они образуют ряды: надглазничный, подглазничный, височный и преоперкуло-мандибулярный; появляются невромасты и около обонятельных отверстий. Вдоль средней линии тела чувствующие бугорки дифференцировались до середины хвостового отдела или немного дальше. Невромасты надвисочной и югальной групп еще не дифференцированы. Чувствующие бугорки у живых зародышей прокрашиваются водным раствором метиленовой сини, нервы же, иннервирующие их, еще не прокрашиваются, что указывает на недостаточную функциональную зрелость.
Выйдя из оболочек, зародыши остаются лежать между камнями нерестовых бугров. В питомниках с ровным дном они ложатся на бок. Грудные плавнички находятся почти в постоянном движении, что способствует смене воды у поверхности желточного мешка и тела и имеет дыхательное значение. Ритмичная работа рта жаберных крышек обеспечивает хорошую вентиляцию жабр. Потребление кислорода после выхода эмбрионов из оболочки резко возрастает.
В питомниках зародыши временами начинают активно двигаться, переворачиваются спиной кверху и устремляются вперед. Движутся они без отрыва от дна, наклонив голову книзу. Будучи помещенными в толщу воды, быстро опускаются ко дну. Их маневренности мешает большой желточный мешок. Опыты показали, что такие зародыши на струи воды реагируют еще нечетко, к прикосновению каким-либо предметом относятся безразлично, на стук не реагируют. На свет зародыши реагируют слабо отрицательно и при ярком освещении постепенно передвигаются в затененые участки. В гнездах вылупившиеся зародыши некоторое время остаются там, где лежала икра, а затем начинают передвигаться; щелям между камнями. В зависимости от режима водоснабжения нерестилищ они могут двигаться в разные стороны, со временем приближаются к поверхности грунта. В питомниках вылупившиеся зародыши проваливаются через ячею рыбоводных рамок и скапливаются под стопками. Вследствие большой скученности они закрывают друг друга поверхность желточного мешка, важную дыхания, и могут задохнуться даже при хорошем насыщении воды кислородом. Такие скопления надо своевременно рассеивать
13-й этап. Формирование непарных, брюшных плавников и плавательного пузыря
Переход к новому этапу развития четко выявляется по смене поведения. Зародыши начинают укрываться от света, активно двигаются навстречу течению. Появляется ярко выраженная положительная реакция на прикосновение посторонних предметов (положительное тактильное чувство).
В течение этапа формируются непарные и брюшные плавники. Очень быстро растет плавательный пузырь, удлиняется и дифференцируется пищеварительный тракт. К концу этапа личинки достигают длины 30-32 мм и начинают потреблять внешнюю пищу.
Ускоренно растет хвостовой отдел, и через 20 сут. после вылупления длина постанального отдела превышает 38,5% общей длины. Продолжается резорбция дорсальной и вентральной плавниковых складок. Прианальная складка сохраняет прежний размер. Увеличи�вается высота спинного, анального и жирового плавников. В спинном канальном плавниках закладываются все лучи. Хвост снова приобретает симметричную форму. Ускоренный рост верхних и нижних лучей приводит к образованию вырезки лопасти хвостового плавника. В его крупных лучах формируется до 3—4 члеников. Закладываются лучи в грудных и брюшных плавниках.
На челюстях появляются зачатки зубов, которые прорезаются к юнцу этапа. По краям жаберных крышек вырастают кожистые складки и в них формируются лучи. Кожистые складки достигают плечевого пояса и хорошо прикрывают жабры. Нижние челюсти и жаберные крышки совершают ритмичные дыхательные движения.
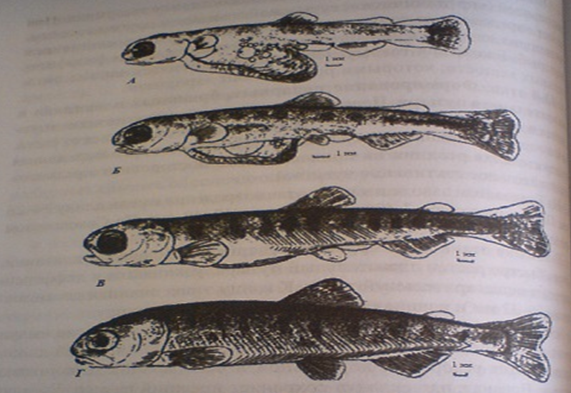
Рисунок 7- Молодь осенней кеты
Смене воды в жаберной полости способствует развившийся во рту парус. Смыкаются боковые выросты стенок обонятельных ямок, обособляется переднее и заднее обонятельные отверстия. Расстояние между глазами и слуховыми капсулами начало медленно увеличиваться. Возрастает абсолютный и относительный размер головы.
После вылупления зародышей резорбция желтка ускоряется. Со временем все большая часть желточного мешка закрывается вентральными выростами миотомов, мелкие кровеносные сосуды на его поверхности. Запустевают и все большую роль в обеспечении организма кислородом играют жабры. Дыхательную функцию несут также капилляры, расположенные в лопасти хвостового плавника и у поверхности тела.
За счет формирующихся изгибов удлиняется желудочно-кишечный тракт. Кишечник заполняется содержимым, заметным, заметны перистальтические движения его стенок. Увеличивается число пилорических придатков и их размер. Быстро растет плавательный пузырь. Васкуляризуется средняя область гонад, и продолжается размножение зародышевых клеток.
Меланофоры по бокам тела распространятся до нижних конусов миотомов, появляются в основном и вдоль переднего края спинного плавника, их становится заметно больше в хвостовом плавнике. Усиливается пигментация оболочек головного и спинного мозга, выстилки брюшной полости, кровеносной сосудов. Мелкие красно-оранжевые липофоры распространяются ниже средней линии тела.
Заканчивается дифференцировка чувствующих бугорков органов боковой линии на голове и по всему телу. Водным раствором метиленовой сини прокрашиваются не только невромасты, но и нервы, их иннервирующие, что указывает на функциональную зрелость этой системы органов чувств. Зародыши четко реагируют на ток воды и движутся навстречу ему, способны противостоять слабому течению. Они воспринимают токи воды, вызываемые движением других особей, и вслед за возбуждением нескольких экземпляров в движение приходит вся масса зародышей. Ярко выражена положительная реакция на прикосновение. Эта реакция возникла в связи со своеобразием каменисто-песчаных гнезд и препятствует преждевременному выходу свободных зародышей из грунта. Оказавшись по каким-либо причинам вне гнезда. Зародыши возбуждаются и активно плавают до тех пор, пока опять не попадут в расщелины между камнями. Преждевременному выходу личинок из гнезд препятствует также их светобоязнь.
Как известно, солнечный свет на икру и вылупившихся зародышей лососей действует отрицательно. При средней температуре 4,1 С действие света отмечается до одномесячного возраста после вылупления. Солнечный свет вызывает обесцвечивание эритроцитов и уменьшение их количества. Под его действием интенсивнее развивается черный пигмент, тогда как оранжевый разрушается и остается только на теменной поверхности головы. На желточном мешке появляются белесые островки отмирающей ткани. К разрушенным участкам присасываются планарии, которые поглощают ускоряют гибель зародышей.
С возникновением положительном реакции на течение, прикосновение к посторонним предметам, светобоязнь зародыши в с ровным дном начинают двигаться навстречу притока укрытия. Они скапливаются у мест поступления воды, в углублении дна, забиваются в щели. Подобные скопления, как говорилось для жизни зародышей, и на их разгонку рыбоводы вынуждены тратить много усилий. Во время перемещений в питомниках зародыши о шероховатое дно повреждают нежный покров желточного мешка, привести к гибели. В питомниках иногда наблюдается дугообразное искривление туловища, изменение формы желточного мешка, смешение жировой капли назад, где она может давить на кишечник. Taкие изменения связываются с выдерживанием свободных зародышем на ровном дне, то есть в несвойственных для них условиях. Указывается, правда, на нивелировку подобных отклонений в ходе дальнейшего развития. В связи с освещенностью питомников ускоряется развитие предметного видения, кета несколько раньше переходит на плав и может преждевременно покидать питомник. Естественным потребностям свободных зародышей отвечает покрытие питомников несколькими слоями крупной обкатанной гальки, которая должна омываться вон снизу, а также синтетический нерестовый субстрат. Этап завершает зародышевый период развития кеты.
1.3 Постэмбриональное развитие
Личиночный период
В начале личиночного периода развития зародыши приобретают способность добывать и переваривать внешнюю пищу. Кета при наличии доступной пищи и подходящей температуры переходит на внешнее питание при большом остатке желтка, достигающем почти половины его величины во время вылупления. Поэтому у личинок длительное время сохраняется смешанное питание и экзогенное питание. В качестве органов движения личинкам продолжают служить непарные плавниковые складки ,которые редуцируются к началу малькового периода. К личиночным чертам организации относятся также остатки эмбриональных органов дыхания, значение которых постепенно снижается и возрастает роль жаберного аппарата. В начале периода на боках тела личинок появляются темные пятнышки, усиливающиеся по мере развития.
Этап смешанного питания
Этап смешанного питания длится 24-40 суток, в зависимости от температуры воды. Длина личинок в этот период колеблется в районе 29-40мм.
Нижняя челюсть у ранних личинок почти достигает кончика рыла. Прорезаются челюстные зубы. Личинки совершают ртом хватательные движения. Желточный мешок утрачивает свисавший книзу задний участок ,и нижний контур его оказывается почти параллельным продольной оси тела. Форма тела становится обтекаемой. Возрастает подвижность молоди.
К началу личиночного периода развития большинство жировых капель желтка сливается и образуется очень крупная капля, которая вскоре приобретает вытянутую яйцеобразную, а иногда грушевидную форму. Желток становится вязким, в связи с чем перемещение в нем жировой капли затрудняется. Если до этого личинки располагались головой кверху, жировая капля оказывается спереди. Так как жировая капля несет функцию гидростатического органа, при ее переднем положении личинки хорошо маневрируют. В питомниках с ровным дном личинки при движении держатся головой вниз, и жировая капля смещается в задний участок желточного мешка. В таком положении она давит на кишку и затрудняет прохождение пищи. А при подъеме личинок в толщу воды сковывает их движение.
В начале этапа нижние конусы туловищных миотомов закрывают примерно половину желточного мешка, а в конце весь его остаток и, наконец, желток резорбируется полностью. С утратой дыхательных поверхностей желточного мешка основная роль в обеспечении личинки кислорода переходит жабрам.
В течении этапа заканчивается вычленение из плавниковой складки жирового плавника, резорбируется прианальная плавниковая складка, развивается вырезка лопасти хвостового плавника. Лепидотрихии хвостового плавника разветвляются, в них увеличивается число члеников. Выше и ниже длинных лучей появляются короткие, так называемые редуцированные лучи хвостового плавника. Возникает членистость лепидотрихий других плавников.
У ранних личинок заполняется воздухом плавательные пузырь, дифференцируется передний (железистый) и пилорический отделы желудка, закладывается много пилорических придатков. При развитии личинки в более высоких температурах воды обнаруживаются признаки перехода от оогоний к ооцитам и начало протоплазматического роста последних; происходит дифференцировка пола. Глаза приобретают подвижность. На голове и туловище увеличивается количество невмастов.
С возрастом пигментация личинок усиливается, особенно на спинной поверхности. На боках формируется темное овальные поперечные пятна, parr marks, который у кеты чаще бывает 10-12 ,изредка 14. Меланофорами обильно покрывается выстилка брюшной полости, головной и спинной мозг, крупные сосуды. Усиливается липофоровая пигментация тела. Оранжво0красную окраску плавников уже можно различать невооруженным глазом. На поверхности жаберных крышек, а по бокам тела появляются отложения гуанина. Свойственную таким личинкам окраску удачно характеризует термин «пестрятка».
В природе кета начинает добывать пищу - детрит, диатомовые водоросли, циклопов, изредка мелких личинок хирономид. Пища разыскивается с помощью органов осязания и вкуса. Переход к внешнему питанию осуществляется до наполнения воздухом плавательного пузыря. Возможность начала питания в гнездах связана с обилием кормовых организмов в грунте.
К концу пребывания в гнездах светобоязнь и продолжительная реакция личинок ослабевают. Постепенно они приближаются к поверхности. Первые выходы из гнезд совершаются ночью, а днем личинки снова прячутся под камнями. Яркий свет они еще избегают. Личинки уже подготовлены к обитанию в водоеме и выживают в случае разрушения гнезд весенними паводками. Однако значительная масса желтка отягощает личинок, связывает их движение, и такие личинки оказываются легкой добычей для хищников. Развитие лососей в грунте до поздней стадий повышает их выживаемость. Возможность продолжительного использования кетой защитных свойств гнезда сочетается с факультативностью внешнего питания в начале личиночного периода развития.
Выйдя из грунта, личинки поднимаются к поверхности воды, захватывают пузырьки воздуха и заполняют им плавательный пузырь. После этого они приобретают возможность устойчивость удерживаться в толще воды. Первое время личинки остаются в районе нерестилища, скапливаются в мелководных прибрежных участках со слабым течением, часто поросших высшей растительностью Их пища состоит из представителей фауны зарослей, обрастаний камней, обитателей грунта; планктонные организмы потребляются редко.
Личинки держатся стайно, в стаях они координируют свои движения, пользуясь преимущественно зрением. Зрение выполняет также важную роль при добычи пищи и ориентировке в пространстве. На течении личинки устойчиво придерживаются избранного участка. Снос потоком контролируется при помощи зрения и тактильно – путем периодических прикосновений брюшной поверхностью ко дну. Тактильная реакция имеет особенно важное значение для удержания личинок на избранном участке водоема ночью, когда зрительная ориентировка в пространстве утрачивается. Ранние личинки на движения предметов над водой не реагируют и вяло реагируют на колебания предметов. При прикосновении отплывают от раздражающего предмета. К концу этапа появляется верхнее поле зрения. Личинки начинают схватывать упавших в воду мелких насекомых. В это время на голове появляются зачатки канальцев органов боковой линии. Кета выходит на открытые, более глубокие места, образуя значительные скопления. В таких скоплениях нередко наблюдается характерное движение по кругу.
В конце этапа остаток желтка и жировая капля быстро резорбируются, но запасы, заключенные в них .не расходуются полностью, а происходит их перераспределение в организме, и в брюшной полости появляются отложения жира. Сковавший движения личинок желточный мешок осчезает, но часть заключавшихся в нем питательных веществ сохраняется в виде жировой ткани и представляет энергетический резерв на время ската, когда расходуется много энергии и молодь может попадать в малокормные участки водоема.
У личинок кеты в основании жаберных лепестков рано появляются особые крупные натрийэкскретирующие клетки Кейсейльмера, позволяющий им избавится от избытков натрия при переходе в морскую воду. Личинки рано начинают проявлять предпочтение к морской воде. Они переносят многократную пересадку из пресной воды в морскую и обратно. Это свидетельствует об одновременной существовании механизмов гипотонической и гипертонической осморегуляции. Указанное качество позволяет молоди продолжительное время находится в приустьевых пространствах рек и прибрежных участках моря, где соленость изменчива.
Молодь кеты из небольших рек в массе мигрирует в море на этапе смешанного питания, еще в состоянии желтка. 25 мая 1954 г. в р. Пугачевка самые мелкие из отловленных скатывавшихся особей имели длину 35мм и массу 276 мг, их средняя длина составила 39мм, а вес-335мг. Массовый скат начинается с наступлением глубокой темноты. Кета в реках может задерживаться до 3-4 месяцев и достигать значительной величины. Как правило, катадромная миграция проходит интенсивно в ночное время, с максимумом через 2-3 часа после захода солнца. При ярком лунном свете интенсивность миграции снижается. Наблюдается миграция кеты и в дневное время. Это обычно происходит в крупных реках, особенно при мутной воде. Иногда дневной скат может быть обильнее ночного. Миграция носит активный характер.
При сплывании по крупным рекам кета задерживается на хороших кормовых участках, заходит в заливы и озера, расположенные на пути движения ,в половодье расселяет по залитым площадям. Она активно питается и интенсивно растет. Задержка в реках и увеличение размера оценивается как фактор, благоприятствующий дальнейшему выживанию кеты.
При ранней миграции у кеты не отмечается сдвига в соотношении пропорций тела, интенсивного посеребрения, отмечаемых перед выходом в море у лососевых. Морфологические, физиологические и этологические отличия покатников кеты расцениваются как свидетельства отсутствия у них так называемой стадии smolt и процесса смолтификации. Когда в покровах личинок кеты начинает откладываться гуанин, обнаруживаются признаки активации щитовидной железы. На этой основе данное состояние аналогично процессу смолтификации. Между тем весной у покатников кеты активность щитовидной железы низкая. Jleтом же при вынужденной задержке в пресной воде, обнаруживаются признаки активизации это органа.
Мальковый период развития
К началу малькового периода развития исчезают произвольные личиночные органы, в том числе прианальная плавниковая складка, и появляются некоторые органы и функции, характерные для взрослых рыб. Закладывается чешуя мальков изменяются пропорции тела, форма плавников, окраска. Этот период кета может проводить как в реках, так и в приустьевых пространствах моря.
Первый мальковый этап. Н.Н. Дислер в мальковом периоде развития кеты выделил два этапа. Личинки кеты превращаются в мальков при достижении длины 38-40 мм. В течение первого этапа мальки достигают длины 50-55 мм. При средней температуре 6,5°С этап длится около месяца. В разных условиях начало закладки чешуи отмечается при длине от 27 до 40 мм
В начале этапа насчитывается 18-20 жаберных тычинок, число которых постепенно нарастает; жаберные лепестки удлиняются, становятся ветвистыми. Обильно снабжаются кровью.
Методом графического сопоставления изменений пропорций тела и отдельных органов кеты устанавливают переломный момент в из�менении относительного размера головы, глаз, спинного, анального и грудных плавников в конце резорбции желтка, т.е. при переходе из личиночного в мальковое состояние. У крупных мальков повышается упитанность, что рассматривается как подготовка к скату.
На боках тела мальков выше и ниже крупных поперечных темных пятен появляются дополнительные мелкие пятнышки, примерно равные диаметру зрачка. На анальном плавнике кеты имеются меланофоры; на жабрах кеты средние лепестки заметно длинные тычинки.
В начале малькового периода невромасты на туловище и хвосте остаются открытыми. На туловище появляются акцессорные чувствующие бугорки, отпочковывающиеся от основного ряда. Начинают формироваться надглазничный, подглазничный и височный каналы органов боковой линии, а также каналы на предкрышке и нижней челюсти. Мальки становятся более подвижными, они чувствительны к волнению. Глазами они улавливают движения предметов не только в воде, но и над водой. Мальки, как и личинки, держатся стайками.
Спектр питания молоди довольно широкий. Основными и предпочитаемыми пищевыми объектами являются личинки и куколки хирономид преимущественно из подсемейств Ortocladiinae и Diamesinae. В массе потребляются нимфы поденок родов Ephemerella, Siphlonurus и некоторых комаров, а также олигохет. В отдельных водоемах кета потребляет много мелких водяных осликов, личинок мух и других организмов. Такие мелкие планктонные формы, как циклопы и дафнии, в желудках встречаются редко и являются случайным кормом. У более крупных мальков пищевой спектр расширяется за счет наземных бес�позвоночных. Крупные мальки питаются реже мелких (отмечены ут�ренний и предвечерний максимумы), но с большей интенсивностью.
Установлено, что личинки хирономид - не только предпочитаемая, но и наиболее эффективная пища; их кормовой коэффициент равен 2,6, а поденок - 3,8. При смене пищи рост замедляется, и расход нового корма на единицу прироста резко возрастает. Суточные пищевые ра�ционы при температуре воды 7-10°С составляет 10-12% массы тела мальков и 17-20% при температуре 12-16°С. Суточный прирост массы в первом случае равен 3,5-3,9% массы тела. Соответственно изменяется интенсивность дыхания. Если при температуре 10С за час на 1 г живой массы малька потребляют 0,325 мг кислорода, то при 20С эта величина возрастает до 0,636. Мальки предпочитают температуру от 8 до 14 С. В Амуре они активно питаются при температуре, достигающей 25-26С. При 10С асфиксия наступает ,когда содержание кислорода понижается до 1,5 мг/л. Для сеголеток кеты пороговое содержание кислорода при 17С составляет 1,99 мг/л, а при 25С повышается до 3,36 мг/л.
Второй мальковый период
При достижении длины 5-5,5 см мальки уходят с мелководья на более глубокие участки рек и озер, а скатившиеся из рек ранее живут в приустьевых участках , эстуариях, прибрежных районах моря. Так как кета может проводить в реках первый и значительную часть второго малькового этапа развития, ее можно подращивать на заводах довольно продолжительное время.
Ко времени достижения длины 6-7 см заканчивается процесс дифференцировки плавниковых лучей. Мальки задерживаются в пресной воде, приобретают некоторую вальковатость. Число пилорических придатков достигает дефинитивного количества. При том же размере завершается формирование каналов органов боковой линии на голове, тулове и хвосте.
Молодь, выходящая в море в начале сезона ската, около 3-4 месяцев держится в близи берегов. Мелководные участки моря прогреваются рано и в конце апреля - начале мая здесь развивается много кормового зоопланктона. Мальки на отмелях интенсивно питаются, быстро рас�тут. Весной основным кормом служат разнообразные мелкие формы Harpacticoida. В июне-июле постепенно возрастает роль крупных форм Gammaridea и Musidacea. В пище более крупных мальков, которые отходят на приглубые участки, все чаще встречаются взрослые насекомые, крупные мизиды, десятиногие рачки, личинки рыб.
Глава 2. Влияние различных факторов среды на исследуемый объект
Температура воды является одним из важнейших факторов, оказывающих воздействие на отправление жизненных функций рыбы, определяющих ее рост и развитие. Этот фактор действует на рыбу как непосредственно – изменяя интенсивность ферментативных процессов, происходящих в организме, активность потребления пищи, характер обмена веществ, ход развития половой системы и прочие, так и косвенно, оказывая влияние на улучшение или ухудшение состояния кормовой базы.
Рассматривая лососевых рыб по отношению к температуре можно отметить, что это холодолюбивые, зачастую стенотермные формы.
Кета - моноцикличная рыба.
Зимой лососи круглосуточно держатся на глубинах, достигающих 300500 м. Вертикальные миграции, по-видимому, совершаются небольшие; их ограничивает концентрация кормового планктона на определенных горизонтах. С началом весеннего прогрева увеличивается количество кормовых организмов в поверхностных водах куда перемещаются и лососи. Ближе к поверхности они держатся ночью - весной в слое 0-50 м, летом в слое 0-13 м. На глубинах до 8-10 м в это время и осуществляется дрифтерный лов. Днем вслед за кормовыми организмами лососи опускаются на глубины в несколько десятков метров. Вертикальное перемещение рыбы наблюдается примерно в тех же температурных границах, что и горизонтальное расселение. На севере прогревается меньший поверхностный слой и вертикальные миграции здесь короче. Весеннее перемещение кеты с мест зимовок в более холодные районы совпадает с повышением температуры поверхностного слоя моря до 2,5-3° С. В период наиболее сильного прогрева океана, в августе — сентябре, распределение нагуливающейся кеты на юге ограничивается поверхностной изотермой около 14° С. В районах океана с температурой 15-17° С лососи во время нагула не встречаются (Бирман, 1958, 1967, 1968). В Охотском море лососи вылавливались при температуре до 18,7°С - более высокой, чем в океане. Такое различие А. И. Фролов (1964) связывает с изменением физиологического состояния рыбы при переходе от кормовой миграции к нерестовой. Оказываясь в зоне влияния речного стока, созревающие производители начинают положительно реагировать на опресненную воду и заходят в эстуарии и устья рек.
Температурный и гидрохимический режимы тех и других нерестилищ существенно различаются, о чем подробнее будет сказано ниже.
Первые экземпляры начинают заходить в конце августа, при среднесуточной температуре воды около 15° С. Затем примерно на 15-20 дней ход прекращается. Подход возобновляется с конца второй декады сентября, когда температура воды снижается до 12-13°С. В конце сентября суточные уловы достигают нескольких сот экземпляров. Обильные подходы совпадают с повышением уровня реки, ускорением течения и охлаждением воды примерно до 10°С. В начале октября интенсивность хода несколько ослабевает, но с середины октября ход становится особенно обильным и сохраняется таким до начала декабря. Продвижение кеты к нерестилищам наблюдается до двадцатых чисел декабря. К декабрю температура воды снижается до 0.5—0.2С. Рыба движется к местам выхода грунтовых вод, которые в это время имеют температуру в 5-7 °С, а местами и выше.
Летняя Амурская кета нерестится в самое теплое время, когда в устьях рек температура колеблется от 9,8 до 13,6С , а в верхнем районе нерестилищ снижается до 6-7С (Кузнецов, 1937).
Суточные колебания температуры в буграх меньше, чем на поверхности и обычно не превышают 1,5 - 2С. Однако зимой разницы в 0,2—0,5°С уже достаточно, чтобы предохранить икру от промерзания.
Кислорода в воде из нерестовых бугров содержится несколько больше, нежели в воде, взятой из грунта по соседству с буграми. Различие объясняется очисткой грунта нерестовых бугров от ила, активно поглощающего кислород. В сентябре в гнездах летней кеты на р. Мы отмечалось от 2 - 3 до 8 мг/л кислорода; его было на 3,5-4 мг/л меньше, чем в толще воды. Количество углекислоты колебалось примерно от 2 до 10 мг/л, а pH — в пределах 6,2-6,3. Скорость течения в буграх составляла 0,3—2,6 м/час (средняя величина —1,24).
В связи с осенним похолоданием воды и повышением растворимости в ней газов количество кислорода возрастало. Зимой падение уровня реки, заиление и уплотнение грунта вызывает снижение проточности и ухудшение газового режима гнезд. Особенно опасно промерзание грунта, при котором создаются экстремальные условия. Для развития летней амурской кеты благоприятно содержание кислорода от 4,5 до 7-8 мг/л (Васильев, Юровицкий, 1954; Васильев, 1957, 1958, 1956).
Камчатская летненерестующая кета тяготеет к выходам грунтовых вод. Скорость течения на ее нерестилищах составляет 12— 33 см/сек. В воде речных нерестилищ содержится кислорода 8 - 9 мг/л около 70% насыщения), свободной углекислоты от 5,80 до 13,75, в среднем 7—9 мг/л, pH колеблется от 6,4 до 6,9 (Кролем, Крогнус, 1976).
В Амуре, реках Приморья, Курильских островов, Сахалина, Хоккайдо, Хонсю осенняя кета размножается в водоемах, снабжаемых грунтовыми водами. В лимнокренах и нерестовых ключах бассейна Амура летние температуры не превышают 11-12С, зимой колеблются в пределах 2,5-5°С (на нерестилищах хоккайдских рек температура бывает и выше). Развивается этот лосось при температуре 1,0-11,5°С (Леванидов, 1956; Sano, 1959). В реках Британской Колумбии во время нереста кеты температура иногда достигает 16°С (Neave, 1966).
На перекатах, в местах нереста осенней кеты, скорость течения составляет 10-30 см/сек, а на плесах она совершенно незначительна. Скорость фильтрации воды в гнездах разных нерестилищ колеблется в среднем от 11 до 36,5 и может достигать 72 м/час. Поверхностные воды нерестилищ р. Хор и в оз. Теплом в октябре содержат кислорода в среднем 6,8-9,5 мг/л, в ноябре 5,8-8,8, в феврале на некоторых нерестилищах 1,6—6,2 мг/л (12,6—48,4% насыщения)с апреля аэрация грунтовых вод улучшается. В некоторых гнездах вода содержала кислорода около 2 мг/л (15% насыщения), тем не менее икра в них развивалась нормально.
Границы для успешного развития осенней кеты определяются содержанием кислорода около 2—8 мг/л (15—60% насыщения), а углекислоты 7—14 мг/л в начале, и 15—27 мг/л в конце эмбриогенеза (Леванидов, 1964,1969).
Об условиях развития того же лосося на Сахалине можно судить по Данным анализов грунтовых вод, ранее питавших нерестовый ключ, а в настоящее время один из цехов Соколовского завода. По 39 определениям, проведенным нашим отрядом в i960 г, поступающая на завод вода содержала кислорода 4,38— 6.52 мг/л (насыщение 39,3—53,3%, в среднем 45,1%). В 1952 г. отмечено максимальное содержание кислорода - 7,3 мг/л, а свободной углекислоты — от 21,6 до 30,8 мг/л. Вода постоянно имела слабую кислую реакцию; временами pH снижалось до 6,4—6,2.
Установлено, что личинки хирономид - не только предпочитаемая, но и наиболее эффективная пища; их кормовой коэффициент равен 2,6, а поденок - 3,8. При смене пищи рост замедляется, и расход нового корма на единицу прироста резко возрастает. Суточные пищевые рационы при температуре воды 7-10°С составляет 10-12% массы тела мальков и 17-20% - при температуре 12-16°С. Суточный прирост массы в первом случае равен 3,5-3,9% массы тела. Соответственно изменяется интенсивность дыхания. Если при температуре 10С за час на 1 г живой массы малька потребляют 0,325 мг кислорода, то при 20С эта величина возрастает до 0,636. Мальки предпочитают температуру от 8 до 14 С. В Амуре они активно питаются при температуре, достигающей 25-26С. При 10С асфиксия наступает ,когда содержание кислорода понижается до 1,5 мг/л. Для сеголеток кеты пороговое содержание кислорода при 17С составляет 1,99 мг/л, а при 25С повышается до 3,36 мг/л.
Одним из факторов, обеспечивающих выживаемость молоди лососей в
период катадромной миграции, является её способность адаптироваться к
условиям нарастающей солёности (Краюшкина,1977). Осморегуляторный
механизм начинает формироваться у молоди кеты ещё в пресноводный
период на стадии пресмолтов и достигает значительного уровня развития
на стадии смолтов, перед миграцией в море.
Глава3. Управление половыми циклами у рыб различными методами
В практике искусственного рыборазведения применяют три метода стимулирования созревания половых продуктов у производителей рыб: экологический, физиологический и эколого�физиологический.
Экологический метод разработан в 30 -е годы академиком А.Н. Державиным. Применяется, главным образом, при разведении осетровых, лососевых и реофильных карповых рыб при выдерживании производителей с целью получения от них зрелых половых продуктов. Сущность этого метода состоит в том, что производителей выдерживают в садках и бассейнах (иногда с управляемым термическим режимом) в обстановке соответствующей естественным условиям. При этом учитываются те факторы, которые способствуют развитию и созреванию половых клеток, овуляции и образованию спермы. Это, прежде всего температура, близкая к температуре нереста данного вида рыб, течение, кислородный режим и грунт (нерестовый субстрат).
Физиологический метод стимулирования созревания половых клеток у рыб разработан профессором Гербильским H. JI. Он провел гистологические исследования гипофиза рыб и раскрыл механизм его физиологического воздействия на созревание половых продуктов.
Гипофиз - железа внутренней секреции, расположенная у основания головного мозга. Гипофиз выделяет в кровеносную систему организма гормоны. Среди них гонадотропный гормон, который регулирует оогенез и сперматогенез, вызывает созревание половых клеток, овуляцию и образование спермы.
Н. JI. Гербильский установил гонадотропную активность гипофиза рыб в различные периоды годового цикла. В определенные сезоны года он накапливается в гипофизе. Это позволяет использовать гипофиз как источник гонадотропного гормона, при помощи которого можно получить зрелые половые продукты от производителей на рыбоводных предприятиях. При внутримышечных инъекциях производителям суспензии гипофиза рыб гонадотропный гормон поступает в кровь и стимулирует половой процесс. Это приводит к быстрому переходу половых желез производителей из IV в V стадию зрелости и получению от них зрелой, способной к оплодотворению и развитию икры у самок и доброкачественной спермы у самцов.
Введение ацетонированного препарата гипофиза в мышцы тела самки и самца, от которых хотят получить зрелую икру и сперму, называется гипофизарной инъекцией.
Эколого-физиологический метод стимулирования созревания половых клеток у рыб. В настоящее время для стимулирования половых продуктов у рыб применяют комбинированный метод. Он сочетает в себе два метода - экологический и физиологический и предусматривает стимулирование созревания половых продуктов у производителей путем комбинированного воздействия на организм рыбы экологических факторов среды и вводимых физиологически активных веществ. Это дает возможность рыбоводу получать в определенный день и даже час необходимое количество зрелой икры и спермы, что позволяет планировать работу рыбоводного предприятия.
Многие из разводимых видов рыб не созревают в неволе без специаль�ных гормональных воздействий, однако у большинства представителей лососевых гаметогенез завершается в условиях рыбоводных предприятий без дополнительной гормональной стимуляции. В этом случае процесс созревания рыб полностью зависит от условий внешней среды и часто растягивается на длительный период времени, что значительно усложняет рыбоводный процесс.
Заготовку производителей для рыбоводных заводов осуществляют в нерестовых реках путем установки заграждений с ловушками или отлова неводами производителей ниже района расположения нерестилищ. Из улова отбирают особей с хорошо выраженным брачным нарядом (IV стадия зрелости гонад) и отсаживают для окончательного созревания в садки. Период дозревания составляет от 1 до 15 суток. Производителей со слабо выраженными признаками брачного наряда пропускают вверх по реке на естественные нерестилища. Для рыбоводных целей можно использовать икру и молоки только от производителей с вполне зрелыми половыми продуктами.
Таким образом, для стимулирования созревания половых продуктов используют экологический метод стимулирования. Если же нужно получить половые продукты в короткие сроки, то используют эколого-физиологический метод.
Рассмотрим экологический метод выдерживания производителей: в садки для длительного выдерживания необходимо отсаживать только здоровых полноценных производителей. Размерно-массовые и экстерьерные показатели производителей предпочтительны средние для данной популяции. Производителей, доставленных из других районов или отловленных в других реках, необходимо содержать отдельно, в карантинных бассейнах или садках. Для кратковременного выдерживания и сортировки производителей в период нереста можно использовать имеющиеся пластиковые бассейны. Уровень воды в них должен поддерживаться не ниже 0,5 – 0,6 м, а полная ее смена 30-40 мин. Бассейны должны быть закрыты сверху крышками или делью.
При снижении температуры воды до 7-8°С и приближении сроков нереста необходимо пересаживать самок и самцов раздельно из стационарных в речные садки. После этого всех производителей регулярно, через каждые 4-5 суток, следует подвергать индивидуальной проверке на созревание.
Признаком созревания самцов является появление капли молок при легком нажиме у анального отверстия. О степени зрелости самок судят по размягчению и западанию стенок брюшной полости а заднем отделе тела при подъеме их за хвостовой стебель
Близких к зрелости производителей необходимо пересаживать в отдельные садки. В этих садках их просматривают через каждые 2-3 суток. При этом учитывают, что самцы созревают раньше самок.
Созревших самцов, у которых молоки выделяются струей, пересаживают с садки и используют в первую очередь. Сперма у самцов созревает и выделяется порциями (эякулятами), поэтому в течении нерестового периода каждого полноценного самца можно использовать не менее 4-6 раз с интервалом в 3-4 дня. Соответственно необходимо отдельно 3-4 садка для содержания зрелых самцов.
Созревших, текучих самок надо сразу после просмотра использовать для отцеживания икры. Таких самок легко отличить по сильному размягчению брюшка, набуханию и покраснению генитального отверстия. При подъеме созревших самок за голову и легком нажиме на брюшко икринки начинают струйками выделяться наружу.
Глава 4. Биологические основы кормления исследуемого объекта
Кета и другие тихоокеанские лососи переходят на внешнее питание, имея еще половину желточного мешка. В естественных условиях кормом личинкам служат бентосные организмы, преимущественно личинки хирономид, поденок, ручейников, веснянок. При экстенсивном методе выращивания молоди естественная кормовая база играет решающую роль в удовлетворении пищевых потребностей молоди. На современных лососевых заводах, где молодь, содержится при уплотненных посадках, основное значение приобретает искусственный корм, отвечающий потребностям рыбы в основных элементах питания. Для нормального роста и развития молоди корма должны содержать все необходимые питательные вещества. Молодь кормят стартовым кормом, который включает 45-50% протеина, 10-15% жира, 10-12 % минеральных солей, до 18 % углеводов и комплекс необходимых витаминов (табл. 1).
Таблица 1 - Состав полноценных гранулированных кормов для молоди проходных лососей р. Oncorhynchus, %
|
Ингредиенты
|
РГМ 8М
|
РГМ 6М
|
РГМ 7М
|
|
Мука рыбная
|
48,6
|
48,0
|
48,0
|
|
мясокостная
|
5,0
|
5,0
|
5,0
|
|
кровяная
|
5,0
|
5,0
|
5,0
|
|
пшеничная
|
1,0
|
5,1
|
5,1
|
|
водорослевая
|
1,0
|
1,0
|
1,0
|
|
Обрат сухой
|
5,5
|
5,5
|
-
|
|
Молочно белковый концентрат
|
-
|
-
|
12,0
|
|
Дрожжи гидролизные
|
6,0
|
6,0
|
6,0
|
|
Шрот соевый
|
16,0
|
16,0
|
9,5
|
|
Жир рыбный
|
10,0
|
7,1
|
7,1
|
|
Краситель рубиновый СК
|
0,6
|
0,3
|
0,3
|
|
Премикс ПФ-1М
|
1,0
|
1,0
|
1,0
|
|
Сырой протеин
|
45,7
|
45,8
|
50,1
|
|
Жир
|
15,0
|
11,5
|
11,5
|
|
Общие углеводы
|
15,7
|
18,4
|
17,0
|
|
Энергия с учетом переваримости, ккал/кг
|
3233
|
3019
|
3142
|
|
Стоимость 1 кг корма, руб.
|
0,69
|
0,62
|
0,66
|
Корма бывают пастообразные или гранулированные. Гранулированный корм для лососевых рыб включает в себя сухие мукообразные ингредиенты, при этом главную роль играет рыбная мука (до 50%). При использовании сухих гранулированных кормов отпадает необходимость в строительстве кормоцеха и холодильника, снижаются затраты на транспортировку и хранение корма, сокращаются трудовые затраты на раздачу корма.
Раздачу сухого гранулированного корма производят с применением специальных кормораздатчиков. Раннюю молодь кормят 12-24 раза в сутки, подросшую молодь-6-8 раз в сутки. Применение механических кормораздатчиков снижает трудовые - затраты и повышает эффективность кормления. Искусственные гранулированные корма особенно нужны при выращивании мелкой молоди до массы 5-10 г, выживаемость которой значительно ниже, чем подросшей. Личиночный период развития у кеты при температуре 6-7°С завершается через 22-25 сут, при температуре до 50 этот срок увеличивается до 30-35 сут. Период нагула молоди кеты в. реке зависит главным образом от температуры воды и обеспеченности пищей.
Разные локальные стада имеют неодинаковую продолжительность речного периода. В реках, где достаточно корма, молодь может нагуливаться до 3 мес., но основная масса скатывается через 1-2 мес.
Глава 5. Транспортировка икры, личинок, молоди и взрослых особей выбранного объекта
Транспортировка половых продуктов, посадочного материала и взрослой рыбы может быть внутрихозяйственной и межхозяйственной.
Непродолжительные по времени внутрихозяйственные перевозки осуществляют в полиэтиленовых мешках, живорыбных контейнерах, молочных флягах и живорыбных автомашинах (в зависимости от вида и возраста рыб, а также расстояния перевозки). Для межхозяйственных перевозок на большие расстояния используют авиацию, железнодорожный и автомобильный транспорт. Важным отличием между этими перевозками является то, что при межхозяйственных перевозках всегда необходимо разрешение ветеринарной службы о том, что хозяйство, откуда перевозится рыба, благополучно по заболеваниям, не закрыто на карантин и т.д. Так на межхозяйственные перевозки необходимо получать разрешение (проходное свидетельство) в Санэпидстанций области, а при межобластных перевозках - в республиканской СЭС.
Запрещается вывозить рыбу из прудов, в которых обнаружены краснуха, бранхиомикоз, фурункулёз, инфекционная анемия, вертеж и дискокотилоз. Нельзя в такие пруды и ввозить рыбу.
Не рекомендуется перевозить рыбу с пучеглазием, взъерошенной чешуей, раздутым брюшком, белыми или эродированными жабрами, черной сыпью по телу или с черным хвостом; вялую, травмированную и истощенную рыбу.
Емкости для перевозки рыбы дезинфицируют 10-20% раствором хлорной извести и затем тщательно промывают водой.
Перевозимую рыбу предварительно обрабатывают в соляных (5% NaCl) или аммиачных (0,2% NH4) ваннах.
После перевозки рыбу помещают на 20 суток в карантин. При выявлении болезни рыб лечат, или продают населению (в свежем или переработанном виде), либо перерабатывают на комбикорм, либо уничтожают (сжигают или закапывают).
Воду для перевозок используют свежую, чистую, с нейтральной или слабощелочной реакцией. При длительных перевозках в воду добавляют поваренную соль (5-6 кг/м для межой рыбы, 5-10 кг/м для крупной).
В связи с тем, что в семенной жидкости сперматозоиды находятся в активном состоянии, сперму рыб можно перевозить на любые расстояния в сухих стерильных пробирках, установленных в термосе со льдом. При этом необходимо учитывать сроки ее активности (табл. 2).
Таблица 2 - Длительность оплодотворяющей способности спермы разных видов рыб
|
Вид рыбы
|
Температура, °С
|
Срок активности
|
|
Лососи
|
2
|
2-3 суток
|
Хранению и перевозке подлежит свежеотобранная сперма, помещенная в сухие пробирки отдельно для каждого самца с плотно пригнанными пробками во избежание попадания воды. Пробирки с этикетками заворачивают 1 марлю и отпускают в термос, наполненный мелко наколотым льдом. К каждой пробирке со спермой должна быть прикреплена этикетка с указанием вида рыбы, номера производителя, даты и часа получения спермы, объема и качественной ее оценки. Для перевозки небольшого количества спермы широко используются сумки- холодильники.
Для перевозки неоплодотворенной ее икры закладывают в сухую банку, которую плотно закрывают пробкой и помещают в термос. Банка должна полностью заполняться икрой без свободного воздушного пространства. Перевозка оплодотворенной икры осуществляется на определенной стадии развития эмбриона. У кеты это первые сутки после оплодотворения, либо на стадии пигментации глаз у зародыша.
Как правило икру транспортируют в стандартных пластиковых контейнерах на деревянных рамках.
На каждую рамку кладут, марлевую салфетку, ПК которой в два раза больше рамки, и опускают в лоток с водой на глубину несколько сантиметров. Икру размещают равномерно на салфетке слоем в 3-4 ряда. Затем рамки вынимают из лотка, ставят на некоторое время в наклоном : положении для удаления избытка воды, обертывают краями салфетки и укладывают их в изотермные пенопластовые контейнеры. При их отсутствии стопки рамок с икрой помещают в деревянные ящики размером на 15 - 20 см больше рамок для укладки в промежутки изоляционного материала.
В случаях перевозки на небольшие расстояния икру можно перевозить без увлажнения; рамки с икрой завертывают в бумагу. При емкости бака 23 л запаса воды хватает на 12-14 часов. В один ящик вмещается 600-1 800 тыс. оплодотворенных икринок осетровых рыб.
Перед отправкой икру промывают от осевшего ила, из нее удаляют икринки, покрытые сапролегнией. После этого рамки с икрой закрывают влажными марлевыми салфетками и стопками укладывают в транспортировочные ящики. Сверху стопок устанавливают лотки со льдом. Температура в ящиках при транспортировке поддерживается 8-10°C. B отдельных случаях на непродолжительное время ее можно снизить до 5-6°С. Общее время транспортировки может продолжаться до 64 часов.
Перевозку икры при температуре воздуха выше 7°С осуществляют, как правило, в ящиках со льдом. Если же икру перевозят поздней осенью или зимой при низких температурах на транспорте, то в тару не только не закладывают лед, но, наоборот, ее утепляют, чтобы избежать промерзания икры. Стопку рамок (кюветов) обертывают плотной бумагой и перевязывают шпагатом. Для доступа воздуха бумагу, закрывающую рамки, в нескольких местах прорывают. Упакованную в бумагу стопку рамок опускают в пенопластовый изотермический ящик.
В каждый транспортировочный ящик вкладывают конверт с накладной, в которой указаны количество икры, дата оплодотворения и температура инкубации.
На крышке транспортировочного ящика делают надпись «Живая икра. Верх. Не кантовать». С боковой стороны ящика прикрепляют этикетку с адресом назначения и отправления.
Транспортировка личинок
Чаще всего для транспортировки личинок рыб используют стандартные полиэтиленовые пакеты объемом 40 л, длиной 65 см с шириной рукава 50 см. Перед перевозкой их упаковывают в стандартную картонную коробку размером 65x35x35 см. В каждый полиэтиленовый пакет наливают 10-12 л воды, помещают личинок, свободное пространство заполняют кислородом и закрывают ракет с помощью зажима Мора или резинового шланга.
Оптимальной температурой для перевозки в летнее холодолюбивых 6-8°С, а весной и осенью, соответственно, 6°С и 3-5°С.
В один пакет помещают до 166 тыс. лососевых. При расчете плотности посадки предличинок, личинок и молоди рыб в герметически закрытый полиэтиленовый пакет, заполняемый водой и кислород: необходимо учитывать следующее:
степень растворения кислорода в воде в зависимости от механического перемешивания в пути, температуры и парциального давления;
пороговое содержание кислорода для предличинок, личинок и молоди (1-3 мл/л);
потребление кислорода и выделение-свободной углекислоты предличинками, личинками и молодью в зависимости от температуры, их массы и видовой принадлежности;
предельно допустимые концентрации углекислоты для предличинок, личинок и молоди;
коэффициент растворения углекислоты в воде в зависимости от температуры;
фактор пространственного расположения предличинок, личинок и молоди (для рыбы до 1 г. принимается соотношение ее массы и воды от 1: 8 до 1: 10, а свыше 1 г.- от 1: 2 до 1: 6);
длительность перевозки предличинок, личинок и молоди.
Нормативы плотности посадки предличинок, личинок и молоди в полиэтиленовые стандартные пакеты представлены в таблицах 4-6.
Глава 6. Биологические особенности акклиматизации исследуемого объекта
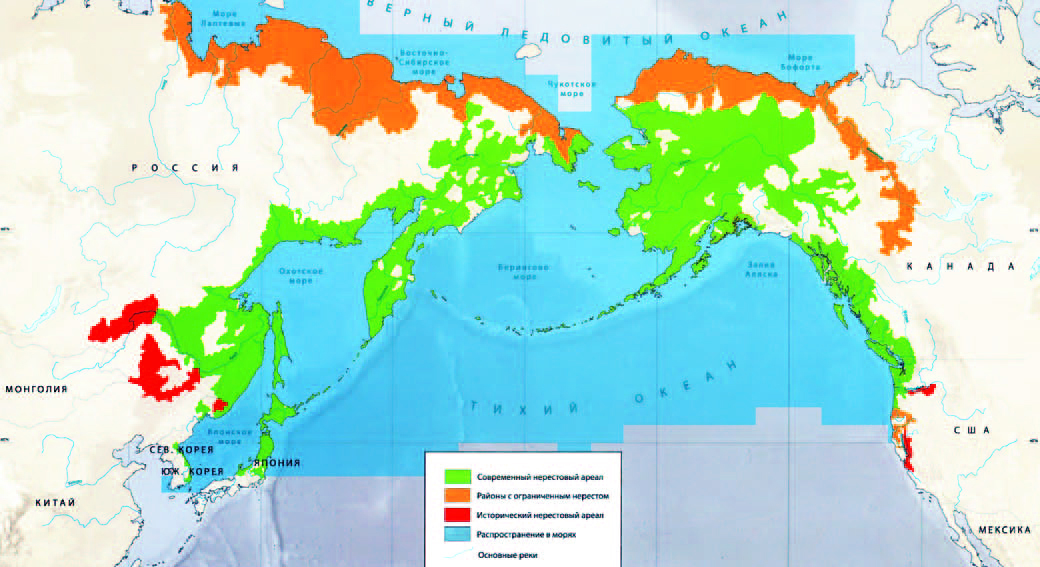
Рисунок 8 - Ареал обитания Кеты
Акклиматизация — процесс приспособления переселен�ных в другой водоем рыб к новым условиям среды, в результате чего из их потомства образуется популяция.
Этот процесс проте�кает медленно и связан с глубокой перестройкой, происходящей в организме рыбы под воздействием новых факторов среды.
Он завершается благополучно не всегда и не для всех переселенных особей.
Выживают только рыбы, наиболее приспособленные к данным условиям.
Эти рыбы, приспособившись к новым услови�ям, размножаются и в дальнейшем образуют популяцию, кото�рая будет существовать за счет естественного воспроизводства.
Под вселением понимается переселение рыб в новый во�доем, условия среды в котором мало или совершенно не отли�чаются от условий существования данного вида рыб в исходном (материнском) водоеме.
Вселенные рыбы успешно размножают�ся в новом водоеме без какой-либо предварительной внутренней перестройки организма.
Биологические и экологические особен�ности потомства переселенных рыб не изменяются.
Акклиматизация рыб и кормовых объектов является составной частью комплексных мероприятий по воспроизводству рыбных запасов и кормовых ресурсов водоемов.
Задачей акклиматизационных работ является повышение продуктивности и хозяйственной ценности водоемов, а также сохранение и увеличение численности ценных видов гидробионов за счет расширения их ареала обитания. Самым первым и, безусловно, важным этапом процесса акклиматизации является интродукция. В практике переселения водных организмов интродукция- это перенос организмов с целью введения их в новую область, биотоп, водоем. Но не всегда процесс интродукции заканчивается акклиматизацией. Поэтому главным шагом акклиматизационного процесса является выбор водоема- объекта заселения. Он не должен отличаться от материнского водоема по основным показателям (гидрохимический режим, кормовая база и так далее).
Кета в прошлом являлась объектом многочисленных, но безуспешных интродукций. До настоящего времени удачных случаев акклиматизации этого вида не было выявлено. Тем не менее, известно, что она великолепно выносит поэтапную акклиматизацию: первые этапы жизненного цикла (получение икры, ее оплодотворение и инкубация) поддерживаются на рыбоводных лососевых заводах, а нагул и созревание протекают в реках и морях.
План акклиматизационных работ.
1. Выбор объекта.
2. Выбор водоема.
3. Анализ гидрохимического режима водоема.
4. Вселение объекта – акклиматизанта.
Период работы с объектом акклиматизации составит 8 месяцев.
На данный момент запасы кеты истощены. Для пополнения численности этого ценного объекта целесообразно будет использовать реоакклиматизацию в водах р. Колыма, где условия среды приближены к привычным виду условиям обитания.
р.Колыма – одна из крупных рек Якутской области. Длина реки примерно 2150 километров, площадь бассейна 644 квадратных километра.
Колыма берет начало под 61°30` северной широты, с северной стороны Станового хребта, двумя вершинами, из коих одна близко подходит к вершине реки Кохтуя, впадающей в Охотское море у Охотска. После соединения обеих вершин река течет с юго-запада на северо-восток, до впадения в нее справа реки Шубиной, отсюда течет на запад-северо-запад — северо-восток и север извиваясь во все стороны до левого своего притока реки Шумихи, далее Колыма течет на север, делая значительное колено к юго-востоку, затем склоняется снова к северо-северо-западу и северо-западу, до впадения в нее слева реки Зырянки.
Колыма извилинами и большими коленами протекает на северо-северо-восток до урочища Вяткина, где река делает большие завороты в разные стороны, образует, наконец, дугу, заворачиваясь к восток-юго-востоку, затем круто поворачивает к западу и запад-северо-западу до города Средне-Колымска, от которого вновь поворачивает на северо-восток, протекая большими изгибами до реки Филиповой, откуда течет на северо-запад и, не доходя до главного своего притока реки Омолон. Затем русло вновь отворачивает на восток-северо-восток, протекая в таком направлении до впадения в нее справа реки Анюя, откуда отворачивает на север и затем на северо-запад, сохраняя это последнее направление до своего впадения в Колымский залив Северного Ледовитого океана, в который вливается тремя главными устьями, не считая мелких проток.
Водный режим в низовьях зарегулирован водохранилищами. Весеннее половодье начинается во второй декаде мая, заканчивается в 3 пентаде июля. Средняя продолжительность половодья 52 дня. Что подходит для выпускаемой заводской молоди кеты.
Температура воды в реке низкая — 10—15°C, и только на спокойных участках в конце июля — начале августа достигает 20—22°C Скорость течения 10 см/с. Кислород составляет 7-9 мг/л в половодье. В состав планктонных организмов р.Колыма входят большое кол-во личинок хиронамид, паденок, коловратки, зеленые водоросли.
Разведение дальневосточной кеты в реке Колыма может оказаться эффективным и для развития рыбного хозяйства Республики Саха (Якутия). Следует продолжить работы в направлении – управляемого лососевого хозяйства.
Список используемой литературы
- Баранникова И. А.; Дюбин В. П.; Боев Е. В.; Буковская О. С.; Васильева Е. В.; Травкин Б. Г.
Роль гормонального воздействия в регуляции созревания самок осенней кеты. // Вопросы ихтиологии. – 1990 г. – Т.30. Вып.4. – с 660 – 667.
- Бирман И. Б.
Приспособительные особенности нерестовой миграции амурской кеты. Известия Тихоокеанского НИ института рыбного хозяйства и океанографии, Т. 37, 1952. стр. 109.
- Бочаров Г. Д.
Материалы по приспособляемости молоди горбуши и кеты к морской воде. Тр. Мурманского биологического института. Вып. 571, 1964, стр 154.
- Гриценко О. Ф. ; Ковтун А. А.; Коеткин В. К.
Экология и воспроизводство кеты и горбуши. Москва. В О. «Агропромиздат». 1987.
- Зиничев В. В.; Зотин А. И.
Избирательная температура и оптимумы развития у предличинок и личинок кеты Oncorhynchus Keta. // Вопросы ихтиологии. – 1988. – Т. 28. – Вып. 1. – С. 164-166.
- Каев А. М.; Рослый Ю. С.
Мечение осенней кеты в лимане реки Амур. // Рыб. хоз. – 1987. №2 – С. 21-23.
- Канидьев А. И.
Степень выживания молоди кеты Oncorhynchus Keta ( Walbaum) в реке [Вопросы искусственного разведения]. Вопросы ихтиологии, Т. 6, вып 4. 1966, с. 708-7019.
- Карпенко В. И.; Николаева Е. Т.
Суточный ритм питания и рационы молоди кеты Oncorhynchus Keta в речной и ранний морской периоды жизни. // Вопросы ихтиологии. – 1989. – Т. 29. Вып. 2. С. 318 – 326.
- Ковтун А. А.
Возраст и линейный рост осенней кеты Oncorhynchus Keta ( Walbaum) Сахалина. – Вопросы ихтиологии, 1981. Т. 21, Вып 6, С. 1030-1038.
- Кольгаев А. М.
Зависимость выживаемость икры Амурской осенней кеты Oncorhynchus Keta от способа и условий обмывания ее водой . Вопросы Ихтиологии 1962, Т. 2, Вып. 4/25/, Стр 742.
- Павлов Д. А.
Лососевые: [ Биологическое развитие и воспроизводство] – М.: Изд-во МГУ, 1989 – 213 с.
- Пономарев С. В.
Лососеводство. Москва. Моркнига 2012 г.
- Пономарев С. В.; Пономарева Е. Н.
Технологические основы разведения и кормления лососевых рыб в индустриальных условиях. Монография. Астрахань. 2003 г.
- http://cyberleninka.ru/article/n/eksperimentalnye-raboty-po-akklimatizatsii-kety-v-vodoemah-yakutii
Биологические особенности кеты в связи с его искусственным разведением