Классификация Пиреномицетов. Хозяйственное значение и биология грибов семейства Нектриевые
Министерство сельского хозяйства РФ
ФГОУ ВПО «Кубанский Государственный Аграрный Университет»
Кафедра фитопатологии
Курсовая работа
по общей фитопатологии
Тема: «Классификация Пиреномицетов.
Хозяйственное значение и биология грибов семейства Нектриевые».
Выполнил: студентка
2 курса, ЗР – 1301,Галицкая О.В.
Проверил:
профессор Сокирко В.П.
Краснодар
2014
СОДЕРЖАНИЕ
ВВЕДЕНИЕ 3 1.ЦЕЛИ И ЗАДАЧИ КУРСОВОЙ РАБОТЫ 6
1.1ОБЩАЯ ХАРАКТЕРИСТИКА СЕМЕЙСТВА НЕКТРИЕВЫЕ 7
1.2 Внешние признаки поражения растений 7
1.3 Характеристика вегетативных и репродуктивных органов 9
1.4 Специализация грибов 13
2. СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ 15
3. ПРЕДСТАВИТЕЛИ 16
3.1 Отдел Euascomycota 16
3.2 Класс Ascomycetes 18
3.3 Saccharomycetes- Дрожжи 23
3.4 Pyreromycetes- пиреномицеты 24
3.5 Порядок Hypocreales 25
3.6 Род Nectria и Gibberella 26
4. БИОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ МЕР БОРЬБЫ 27
4.1 Агротехнические меры борьбы 27
4.2 Химические меры борьбы 28
5. ЗАКЛЮЧЕНИЕ 29
6. СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 30
ВВЕДЕНИЕ
Все более полное удовлетворение возрастающих потребностей населения в продуктах питания, и промышленности в сырье является одной из важнейших задач сельского хозяйства нашей страны. Научно-технический прогресс и интенсификация производства позволили значительно повысить урожайность почти всех сельскохозяйственных культур. Немаловажную роль в этом сыграли мероприятия по защите растений от вредителей, болезней и сорняков, внедрение в практику борьбы с ними интегрированных систем. Защита растений от вредителей, болезней и сорняков — неотъемлемая часть общего технологического комплекса по выращиванию культуры, важнейший резерв сохранения урожая. Средние ежегодные потери, которые несет мировое сельское хозяйство от вредителей, болезней и сорняков, до сих пор составляют 20, а иногда и 30% потенциального урожая. Вместе с тем правильная, научно обоснованная организация защитных мероприятий, своевременное их выполнение позволяют свести до минимума потери урожая от вредителей и болезней при высокой степени окупаемости этих мероприятий.
В СССР была создана специальная служба защиты растений, в составе которой работает около 30 тыс. человек, в том числе 16 тыс. специалистов по защите растений. Однако основами защиты растений от болезней, вредителей и сорняков должны владеть все специалисты сельского хозяйства, в первую очередь агрономы отраслевых хозяйств, в том числе плодоводы и овощеводы, так как решающе важными в защите сельскохозяйственных культур от болезней и других вредных объектов являются санитарно-гигиенические, организационно-хозяйственные и другие профилактические мероприятия, планирование и осуществление которых зависит от агронома. Наука о болезнях растений и мерах борьбы с ними называется фитопатологией (от греческих phyton — растение, pathos — болезнь, страдание и logos — слово, учение).В задачу фитопатологии входит изучение инфекционных болезней, вызываемых грибами, бактериями, вирусами, цветковыми растениями-паразитами и другими патогенными организмами, неинфекционных болезней, возникающих под влиянием неблагоприятных для растений условий окружающей среды, а также построение научно-обоснованной, высокоэффективной системы защитных мероприятий, которая снижала бы до минимума потери, вызываемые болезнями растений, или, в идеальном случае, вообще предупреждала бы их возникновение и развитие.
Для выполнения этой задачи необходимо знать причины (этиологию) болезней растений, уметь своевременно и правильно распознавать их (владеть методами диагностики), изучать закономерности возникновения и развития в связи с особенностями паразитических и других свойств возбудителя (источники инфекции, специализация патогена, влияние на него условий окружающей среды и т. д.), уметь прогнозировать сроки проявления болезни и интенсивность ее развития, изучать факторы иммунитета, направленно повышать болезнеустойчивость растений, применять своевременно санитарно-гигиенические, а также химические и другие специальные меры, активно внедрять в производство достижения науки и передовой производственный опыт. Возникновение и развитие фитопатологии как науки нельзя рассматривать в отрыве от истории агрономии и биологии в целом. Фитопатология — отрасль биологии и неразрывно связана с такими дисциплинами, как ботаники, микология, микробиология, анатомия и физиология растении, вирусология, химия и др.
Болезни растений известны давно. Но о вызывающих их причинах почти ничего не знали вплоть до середины прошлого столетия. Даже меры борьбы со многими болезнями были известны раньше, чем их причины. Первые практические защитные мероприятия по борьбе с болезнями растений были описаны еще Демокритом (460 лет до н. э.), который указывал, в частности, на возможность протравливания семян злаков. С течением времени накапливались сведения о болезнях — их проявлении, вредоносности, зависимости от условий произрастания, сорта и т. д.
Большая роль в развитии фитопатологии принадлежит академику Н. И. Вавилову (1887—1943), которого по праву можно считать основоположником фитоиммунологии. Его работы в этом направлении получили всеобщее признание и оказали влияние на дальнейшее развитие фитопатологии не только у нас, но и в других странах. Обобщая многочисленные исследования в области иммунитета, Н. И. Вавилов критически подходил к оценке частных его факторов — морфологических особенностей, кислотности, содержания Сахаров, дубильных веществ и т. д. Паразитической специализации гриба он придавал особое значение как фактору иммунитета. В работе «Учение об иммунитете растений к инфекционным заболеваниям» (глава «Среда и иммунитет») он писал: «Иммунитет или, обратно восприимчивость определяются, по существу, тремя категориями факторов: с одной стороны — наследственными особенностями сорта, с другой — избирательной способностью, или специализацией видов паразитов или их отдельных рас и с третьей стороны — также условиями среды». Его выводы основаны не только па обобщении литературных материалов, но и на экспериментах, проведенных с разными видами растений. Установление причин заболеваний растений позволило разработать методы и средства борьбы с ними. Первой попыткой применения химических средств борьбы с болезнями было использование медного купороса в качестве протравителя семян. В середине XIX в. для борьбы с оидиумом на винограде в Европе начали применять серу. В 1887 г. появилась бордоская жидкость — фунгицид, который и поныне— один из наиболее эффективных средств против милдью винограда, парши яблони и других болезней. С этого времени началось быстрое развитие химического метода борьбы с болезнями растений. Характерная черта современной фитопатологии — развитие ее как комплексной науки, обобщающей целую систему научных дисциплин: общую и сельскохозяйственную фитопатологию, фитовирусологию, физиологию больного растения, учение об иммунитете, эпифитотиологию, прогноз болезней растений. Главнейшая задача практической фитопатологии — свести до минимума потери урожая, вызываемые болезнями растении. Важнейшая роль в ее решении будет принадлежать дальнейшему совершенствованию всех методов борьбы с болезнями растении и разработке комплексных, интегрированных систем защиты растений, выведению и внедрению в сельскохозяйственное производство болезнеустойчивых сортов, быстрейшему использованию в производстве достижений фитопатологической науки. В условиях дальнейшей интенсификации сельского хозяйства на основе специализации и концентрации производства и создания крупных предприятий аграрно-промышленного типа роль защиты растений от болезней, вредителей и сорняков будет все более возрастать.
ЦЕЛЬ И ЗАДАЧИ КУРСОВОЙ РАБОТЫ
Курсовая работа является одной из важнейших форм учебного процесса, она направлена преимущественно на практическую подготовку и выполняется в соответствии с учебными планами. Курсовая работа является завершающим этапом изучения дисциплины. Цель ее выполнения состоит в углублении разработки одной из проблем курса, представляющейся актуальной и недостаточно исследованной, либо требующей переосмысления в новых условиях, овладеть навыками самостоятельной работы, подготовиться к более сложной задаче -выполнению дипломной работы. Тематика курсовых работ должна отвечать учебным задачам теоретического курса, быть увязана с практическими задачами народного хозяйства и науки, быть реальной. Структура курсовой работы должна способствовать раскрытию избранной темы и быть аналогична структуре дипломной работы: иметь титульный лист, реферат, содержание, введение, основную часть, заключение, список использованных источников и приложения. Курсовая работа носит учебно-исследовательский характер.
В процессе выполнения курсовой работы решаются следующие задачи:
- систематизация, закрепление и расширение теоретических и практических знаний по специальности;
- приобретение опыта применения этих знаний при решении конкретных задач;
- овладение элементами методики научного исследования;
- формирование интереса к научно-исследовательскому поиску;
- развитие и совершенствование самообразовательных потребностей и умений;
- выработка самостоятельной позиции, умений ее отстаивать и защищать.
ОБЩАЯ ХАРАКТЕРИСТИКА ГРИБОВ СЕМЕЙСТВА НЕКТРИЕВЫЕ
Внешние признаки поражения растений
Семейство нектриевых характеризуется поверхностными перитециями, образующимися на субстрате или на строме. Стромы обычно небольшого размера, подушковидные. Конидиальные стадии типа туберкулярия (Tubercularia), цилиндрокарпон (Cylindrocarpon), фузариум (Fusarium) и др.
Центральный род этого семейства — нектрия (Nectria). Для грибов этого рода характерно наличие двухклетных аскоспор, иногда с перетяжкой, в зрелости не распадающихся на отдельные клетки. Перитеции у нектрии образуются поодиночке или группами на субстрате или на маленьких подушковидных стромах.
Род Nectria (нектрия). Nectria cinnabarina(нектрия киноварно-красная) встречается повсеместно как сапротроф или раневой паразит. Наносит ущерб растениям в городских насаждениях и лесопитомнтках. На отмерших ветвях древесных растений и кустарников из-под покровной ткани появляются оранжево-розовые подушечки-стромы с поверхностным конидиальным спороношением,которое называется туберкулярия обыкновенная(Tubercularia vulgaris). Конидиеносцы простые или слабо ветвящиеся,конидии распространяются каплями дождя. Осенью по краю тех же стром или рядом с ними образуются плодовые тела-перитеции, которые погружены в строму лишь основанием. Стромы при этом меняют облик и выглядят как маленькие зернистые красные подушечки.

Рисунок 1- Нектрия киноварно-красная Nectria cinnabarina
[автор: http://wikigrib.ru /]
Род Gibberella (гибберелла). Сапротрофы или паразиты растений.Развита конидиальная стадия, известна под названием фузариум (Fusarium). Конидии бывают 2 видов-микроконидии и макроконидии серповидноизогнутой формы. На отмерших частях растений развиваются перетеции, которые образуются на субстрате или погружены в строму темно-синего цвета. В пораженных органах растений грибы выделяют ростовые вещества гиббереллины, что приводят к аномальному удлинению междоузлий. Растения становятся бледными и гибнут. Заболевание риса, сахарного тростника, кукурузы, сорго в Японии, Индии носит название баканэ, или болезнь дурных побегов. В настоящее время гибберллины получают при культивировании гриба в промышленных масштабах, используют в сельском хозяйстве как биостимулятор.

Рисунок 2- Заболевание винограда гиббереллином Аmylum [автор:http://wine.historic.ru/]
Наиболее важным считается собственно гибберелловая кислота.
Это вещество запускает и активизирует рост побегов, вызывает прорастание семян, помогает сформировать бессемянные плоды, и заставляет цвести растения длинного дня. Это естественное составляющее для растительных клеток, которое регулирует рост и развитие растения в целом. У бессемянных сортов винограда, именно из-за отсутствия семян, наблюдается дефицит этих гормонов. По этой причине урожай этой группы сортов обычно невелик.
Фузариум (Fusarium) — род анаморфных (несовершенных) плесневых грибов. Телеоморфы (совершенные, или половые стадии жизненного цикла) принадлежат родам Gibberella и Haematonectria, оба рода относятся к семейству Нектриевые (Nectriaceae) сумчатых грибов. У многих видов телеоморфа отсутствует или неизвестна. Представители рода имеют существенное значение как грибы, приносящие вред народному хозяйству и патогены, вызывающие заболевания или токсикозы у растений и животных, в том числе человека. Заболевания растений, вызываемые этими грибами, имеют название фузариозы.

Рисунок 3- конидиевая стадия Monilia fructigena, гниль яблоневая плодовая
[автор: http://berrylib.ru/]
Инфекция проникает в растение из зараженной почвы через корневые волоски или пораженные места, особенно около корневой шейки. Признаки поражения появляются на базальной части растения: она буреет или чернеет (отсюда и «черная ножка»), хотя такое течение болезни не всегда бывает достаточно ярко выражено. Внутри стебля сосудистые пучки начинают буреть. Иногда во влажную погоду на поверхности пораженных частей появляется ватообразный или суховатый налет из спор и грибницы. Грибница проникает в сосудистые переплетения ткани и выделяет токсины, вызывающие отмирание клеток. Поступление воды и растворенных в ней питательных веществ прекращается, растение начинает вянуть.
Характеристика вегетативных и репродуктивных органов грибов семейства Нектриевые
Вегетативное тело— разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки (септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. В центре септы остается пора , через которую происходит движение цитоплазмы. Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста. У некоторых мицелий может распадаться на отдельные клетки или почковаться. У дрожжей (порядок Endomycetales) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися, реже делящимися клетками, иногда образующими псевдомицелий. Дрожжеподобный рост наблюдается и у некоторых мицелиальных аскомицетов, например у грибов-дерматофитов (порядок Eurotiales), тафриновых (порядок Taphrinales), некоторых видов из рода цератостис (порядок Microascales).
В цикле развития большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды). Типы конидиальных спороношений и различные способы образования конидий подробно онисапы в главе о дейтеромицетах, или несовершенных грибах.
Конидиальные спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов — паразитов растений они обычно образуются на живом растении, а сумчатые спороношения, за немногими исключениями,— после отмирания растения или его частей, в конце периода вегетации или после перезимовки. Для большого числа грибов, встречающихся в природе в гаплоидной конидиальной стадии, половые стадии неизвестны. Такие грибы относят к классу дейтеромицетов, или несовершенных грибов. Половой процесс, типичный для аскомицетов,— гаметапгиогамия, т. е. слияние двух гаметангиев — специализированных клеток, не дифференцированных на гаметы. Для высших аскомицетов (подклассы Euascomycetidae и Loculoascomycetidae) характерны дифференциация и усложнение строения гаметангиев. Образуются
одноклеточный антеридий и аскогон, обычно с трихогиной. При оплодотворении содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются сразу, а объединяются попарно, образуя дикарионы. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Аскогенные гифы ветвятся и разделяются на двухъядерные клетки. На концах аскогенных гиф развиваются сумки.
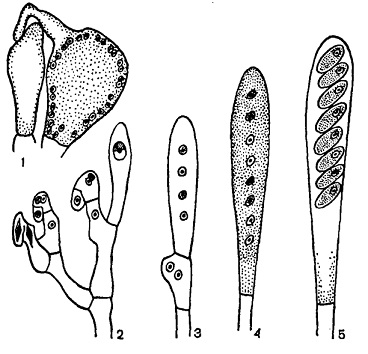
Рисунок 4- Половой процесс и развитие сумок у аскомицетов
[автор: http://berrylib.ru/]
1- аскогон с трихогиной и антередий; 2 - развитие сумок по способу крючка; 3 - молодая сумка после мейоза; 4 - молодая сумка с восемью гаплоидными ядрами; 5 - зрелая сумка с аскоспорами.
Плодовые тела аскомицетов
Различают следующие типы плодовых тел: клейстотеций – полностью замкнутое плодовое тело, перитеций – полузамкнутое, обычно кувшиновидное плодовое тело с отверстием на вершине, апотеций – открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз. Клейстотеции, перитеции, апотеции – настоящие плодовые тела, развивающиеся по аскогеминиальному типу. Перидий (оболчка) такого плодового тела образуется после плазмогамии.
Гаплоидные вегетативные гифы оплетают аскогенные гифы и сумки, формирую плотную покровную ткань. Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах – плотных сплетениях гиф различной формы, размера, консистенции. Аскострома развивается иначе. Сначала складывается строма из переплетающихся гиф. В ней образуются аскогоны и происходит половой процесс. Аскогонные гифы и появляющиеся на них сумки раздвигают или разрушают плектенхиму стромы, освобождая в ней полость (локулу). Каждая локула содержит одну или несколько сумок. Строма под локулой разрушается и образуется отверстие, через которое освобождаются аскоспоры.
СПЕЦИАЛИЗАЦИЯ ГРИБОВ
Специализация у грибов — полное физиолого-биологическое соответствие гриба-паразита и его растения-хозяина. Настоящие паразиты (например , виды аскомицетов семействе Erysiphaceae и базидиомицетов порядке Uredinales) строго приспособлены и способны поражать ограниченный круг растений. Специализация у грибов является свойством вида или более мелких систематических единиц — отдельных форм. Грибы, как гетеротрофные организмы нуждаются в готовых органических веществах. В природе они находят его или в виде разнообразных органических остатков растительного или животного происхождения, на которых они развиваются как сапрофиты, или в виде живых тканей других организмов, на которых они паразитируют.
Сапротрофный способ питания для грибов первичен, а паразитизм представляет собой один из путей специализации грибов. Существующие сейчас грибы-паразиты стоят на разных ступенях эволюции – от факультативных паразитов до облигатных паразитов. Факультативными паразитами называют грибы, обычно развивающиеся как паразиты, но в определенных условиях способных к сапротрофному питанию. Экологически облигатные паразиты могут развиваться как сапротрофы только при отсутствии конкурентов, в природных же условиях существуют только как паразиты на живых тканях, недоступных для других организмов.
Факультативные паразиты — это грибы, для которых обычным в природе является сапрофитный образ жизни. Однако при определенных условиях (поэтому их называют также условными паразитами) они способны переходить к паразитированию на живых, чаще всего ослабленных растениях. В отсутствие подходящих растений-хозяев они развиваются и размножаются на органических остатках в почве. Факультативные сапрофиты обладают более высокой степенью паразитической активности. Обычно они ведут паразитный образ жизни, развиваясь в течение вегетации на живых растениях и образуя на них бесполые спороношения. Однако завершают свой цикл развития они нередко как сапрофиты: половое спороношение этих грибов чаще всего формируется на отмерших частях растений (опавших листьях и хвое, засохших ветвях и т. п.). К этой группе относятся очень многие фитопатогенные грибы из числа сумчатых и несовершенных, например возбудители пятнистостей листьев, парши плодовых и др.
Облигатные паразиты (биотрофы) составляют группу грибов, характеризующихся наивысшей степенью паразитической активности. Они могут питаться только содержимым живых клеток растения-хозяина. Облигатные паразиты обычно поражают вполне жизнеспособные, без каких-либо признаков ослабления растения. Свойствами облигатных паразитов обладают мучнисторосяные, плазмодиофоровые, тафриновые, ржавчинные грибы.
Паразитические грибы, как и другие фитопатогенные организмы (бактерии, вирусы и пр.), характеризуются специализацией, т. е. приспособленностью к поражению определенного круга растений-хозяев или определенных органов и тканей либо растений и тканей только определенного возраста. Различают следующие типы специализации: филогенетическую, онтогенетическую, органотропную и гистотропную. Степень (широта) специализации любого типа у разных возбудителей неодинакова.
Филогенетическая специализация означает приспособленность патогена к паразитированию на одном или нескольких (иногда многих), но всегда определенных питающих растениях. Она является результатом длительной сопряженной эволюции (филогенеза) паразитов и их растений-хозяев. Филогенетическая специализация может быть широкой (когда возбудитель поражает растения разных родов и даже семейств) и узкой (когда он способен поражать растения лишь определенного рода, вида или сорта). Широкоспециализированных патогенов называют полифагами, а узкоспециализированных паразитов, отличающихся строгой избирательностью в отношении растений-хозяев, монофагами.
Онтогенетическая специализация патогенов означает их приуроченность к определенному возрастно-физиологическому этапу индивидуального развития (онтогенеза) растений или к определенному возрастно-физиологическому состоянию поражаемых органов.
Рассмотренные типы специализации (филогенетическая и онтогенетическая) имеют большое практическое значение, поскольку непосредственно связаны с устойчивостью растений к инфекционным болезням. Они могут быть использованы в селекционной работе, при обосновании конкретных мер борьбы и способов повышения устойчивости растений к болезням.
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
Систематика грибов, т.е. разделение этих организмов на отделы,
классы, порядки, семейства, роды и виды, основана на особенностях
их морфологии, физиологии, цитологии, экологии и других биоло-
гических свойствах.
На основе отсутствия или наличия плодовых тел и их типа класс Ascomycetes делят на три подкласса:
Подкласс гемиаскомицеты, или голосумчатые – Hemiascomycetidae, - плодовые тела отсутствуют, сумки образуются непосредственно на мицелии, прототуникатные.
Подкласс эуаскомицеты – Euascomucetidae – сумки образуются в клейстотециях, перитециях или апотециях, прототуникатные или уникатные.
Подкласс лакулоаскомицеты – Loculoascomucetidae – сумки образуются в аскостромах, битуникатные.
У подкласса эуаскомицетов (Euascomycetidae) сумки образуются в настоящих плодовых телах, развивающихся по аскогимениальному типу. Лишь у немногих примитивных представителей этого подкласса сумки формируются группами или пучками на мицелии и не окружены перидием или перидий плодового тела представляет рыхлое сплетение гиф, сквозь которое просвечивают сумки. Система эуаскомицетов основана на строении плодовых тел, типе развития центра перитеция (у пиреномицетов), строении сумок, наличии и характере строме. В современной системе эуаскомицеты для удобства часто подразделяют на группы порядков соответственно типу плодовых тел и строению сумок, обычно не присваивая этим группам ранга таксонов.
К подклассу эуаскомицеты относят сейчас около двадцати порядков. К порядоку гипокрейные (Hypocreales) относятся 3 семейства:
1. семейство Нектриевые (Nectriaceae)
2. семейство Гипомицетовые (Hypomycetaceae)
3. семейство Гипокрейные (Нуросгеасеае)
ПРЕДСТАВИТЕЛИ
Отдел Euascomycota
У эуаскомицетов сумки образуются в настоящих плодовых телах — аскокарпах, развивающихся по аскогимениальному типу. Образование перидия (оболочки) такого плодового тела происходит после плазмогамии. Гаплоидные вегетативные гифы оплетают развивающиеся аскогенные гифы и сумки, образуя плотную покровную ткань. Лишь у немногих примитивных представителей этого подкласса сумки образуются группами или пучками на мицелии и не окружены перидием или перидий плодового тела представляет очень рыхлое, просвечивающее сплетение гиф, например роды биссохламис (Byssochlamys, и аскодесмис Ascodesmis.
По строению различают три типа плодовых тел — клейстотеций, перитеций и апотеций.
Перитеции — полузамкнутые плодовые тела, большей частью округлые или кувшиновидные, с узким отверстием на вершине. Перидий обычно хорошо развит, плотный, с различными окраской (от яркой или светлой до черной) и консистенцией (от мягкой, мясистой до твердой, углистой). Со дна перитеция пучком или слоем поднимаются сумки, обычно булавовидные или цилиндрические. Между сумками развиваются стерильные элементы — парафизы, или апикальные парафизы. Настоящие парафизы врастают в полость перитеция между сумками от внутренних слоев его основания. Они свободны на концах, простые или ветвящиеся. У грибов порядка гипокрейные (Hypocreales) в перитециях образуются апикальные парафизы — стерильные гифы, врастающие в полость перитеция сверху (с «потолка») к его основанию. Сумки в этом случае врастают между апикальными парафизами. Кроме парафиз, в перитеции имеются перифизы — нитевидные короткие гифы, расположенные в носике перитеция и направленные к выходу. Сумки, парафизы и внутренние ткани перитеция называют его центром.
Развитие центра перитеция — важный систематический признак, используемый при разграничении порядков. Выбрасывание аскоспор из перитециев обычно происходит активно. Упорядоченное расположение сумок в перитеции позволяет им поочередно, удлиняясь за счет растяжения оболочки или роста, достигать отверстия перитеция и выбрасывать споры на расстояние до 20 см. У некоторых эуаскомицетов аскоспоры выбрасываются из сумки не все одновременно, а поочередно, например у грибов рода кордицепс (Cordyceps). Наконец, у диапортовых (Diaporthales) сумки имеют короткую, быстро растворяющуюся ножку и в зрелости свободно лежат в слизи в полости перитеция. При набухании сумок в перитеции создается давление и они поочередно выталкиваются к отверстию перитеция и выбрасывают аскоспоры. Такой механизм обеспечивает быстрое освобождение аскоспор из перитециев с длинной шейкой. У Endothia parasitica, например, за час из перитеция выбрасывается около 14 000 аскоспор. Лишь у немногих аскомицетов в перитециях образуются прототуникатные сумки. В этом случае зрелый перитеций содержит массу аскоспор, погруженных в слизь. При ее набухании эта споровая масса выдавливается через носик перитеция и образует у его устья капельку или длинную нить, например у грибов из родов хетомиум (Chaetomium), цератоцистис (Ceratocystis), микроаскус (Microascus).
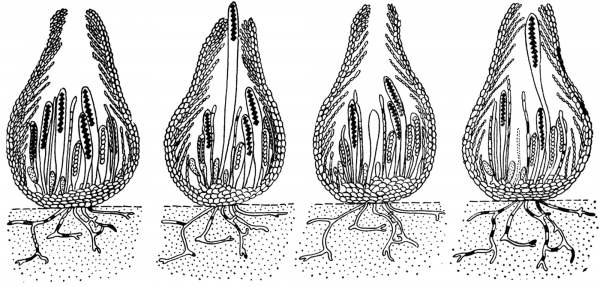
Рисунок 5- Выбрасывание аскопор из перитеция с ордарии.
[автор http://molbiol.ru/]
Плодовые тела эуаскомицетов обычно мелкие. Размеры клейстотециев и перитециев не превышают 1-2 мм. Апотеции — более крупные плодовые тела, их размеры — от долей миллиметра до 15-20 см, а у некоторых грибов и более крупные. У многих эуаскомицетов плодовые тела образуются не на мицелии, а на стромах различного размера, окраски и консистенции, состоящих из переплетения гиф. Однако и в этом случае каждое плодовое тело имеет собственный перидий, хотя бы на ранних стадиях развития. Система эуаскомицетов основана на строении плодовых тел, типе развития центра перитеция (у пиреномицетов), строении сумок, наличии и характере стромы. В современной системе эуаскомицеты часто для удобства делят на группы порядков, соответственно типу плодовых тел и сумок.
Класс Ascomycetes
Аскомицеты – один из самых больших классов грибов, он объе-
диняет около 30 тысяч видов (это около 30% всех грибов). Сумча-
тые грибы имеют многоклеточный (септированный) мицелий, кле-
точные стенки мицелия содержат хитин и глюканы. Основной при-
знак класса – формирование в результате полового процесса сумок –
одноклеточных структур, содержащих фиксированное количество
аскоспор, обычно 8. Сумки (или аски) образуются либо непосредст-
венно из зиготы (у низших аскомицетов), либо на развивающихся из
зиготы аскогенных гифах.
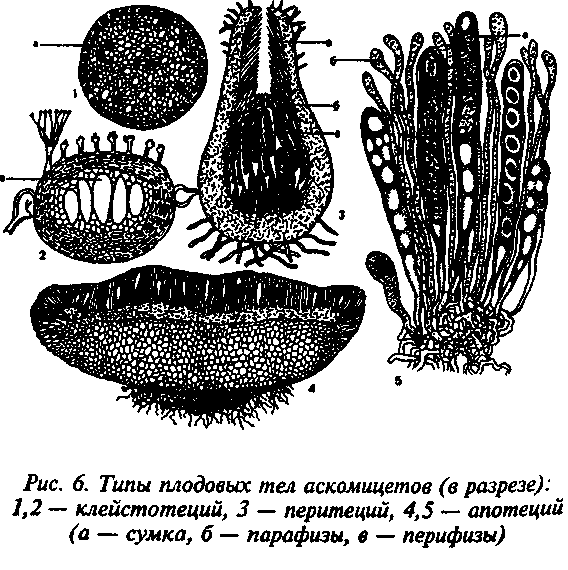
Рисунок 6- Типы плодовых тел аскомицетов Ascomycetes
[автор http://onlineslovari.com/]
Вегетативное тело аскомицетов – разветвлённый гаплоидный
мицелий, состоящий из многоядерных или одноядерных клеток.
Перегородки (септы) образуются в мицелии аскомицетов упорядо-
ченно, синхронно с делением ядер. В центре септы остаётся пора,
через которую передвигается цитоплазма, мигрируют органеллы
клетки и ядра. Скорость движения цитоплазмы у разных видов
варьирует – от 1-2 до 25-40 см в час. У некоторых аскомицетов ми-
целий может распадаться на отдельные клетки или почковаться.
Некоторые высокоспециализированные паразиты насекомых (лабульбениевые грибы) не имеют мицелия, а их тело – т. н. рецеп-
такул – состоит из настоящей ткани. В цикле развития аскомицетов
есть следующие этапы: 1) телеоморфа – это стадия полового раз-
множения (сумки с аскоспорами); 2) анаморфа – стадия бесполого
размножения, конидиеносцы с конидиями, которые образуются на
гаплоидном мицелии.
У аскомицетов, паразитирующих на растениях, конидиальные
спороношения обычно образуются на живом хозяине, а сумчатые –
чаще после отмирания растения или его частей, в конце вегетации
или после перезимовки.
Типичный для аскомицетов половой процесс – гаметангиогамия.
Сущность его состоит в слиянии гаметангиев – специализирован-
ных клеток, не дифференцированных на гаметы. По степени слож-
ности строения гаметангиев и особенностям полового процесса ас-
комицеты условно делят на низшие и высшие. У низших аскомице-
тов половой процесс сходен с зигогамией у зигомицетов. Гаметан-
гии разного пола морфологически сходны или малоразличимы – это
выросты или веточки мицелия. После их слияния сразу происходит
кариогамия, и непосредственно из зиготы развивается сумка. Но, в
отличие от зигомицетов, в многоядерных гаметангиях сливаются
только 2 ядра (множественной кариогамии нет), и зигота не перехо-
дит в состояние покоя, а сразу развивается в сумку. Для высших
аскомицетов характерны дифференциация и усложнение гаметанги-
ев. У них при половом процессе образуется одноклеточный муж-
ской гаметангий – антеридий. Женский гаметангий обычно состоит
из двух частей – аскогона и трихогины (воспринимающей гифы,
которая развивается на вершине аскогона). При оплодотворении
трихогина приклеивается к расположенному рядом антеридию, за-
тем мужские ядра переходят в аскогон и перемешиваются там с
женскими ядрами. Мужские и женские ядра не сливаются сразу, а
объединяются попарно, образуя дикарионы. Затем из вершины жен-
ского гаметангия вырастают многочисленные аскогенные гифы, в
которых ядра дикариона синхронно делятся. Аскогенные гифы вет-
вятся и септируются (разделяются на двухъядерные клетки). На
концах аскогенных гиф развиваются сумки. Происходит этот про-
цесс в несколько этапов (см. рис. 17):
1) Конечная клетка аскогенной гифы загибается крючком, ядра
дикариона располагаются в месте перегиба и одновременно делятся.
2) Пара ядер разного пола остаётся в месте перегиба крючка, од-
но ядро переходит в его кончик, а другое – в основание.
3) Затем образуются две перегородки, отделяющие одноядерные
конечную и базальную клетки крючка. В результате слияния этих
клеток восстанавливается дикарион, и может происходить повтор-
ное образование крючка.
4) Средняя двухъядерная клетка крючка развивается в сумку.
Она увеличивается в размерах, ядра дикариона сливаются. Образо-
вавшееся диплоидное ядро делится редукционно. После мейоза сле-
дует ещё одно – митотическое деление. Образуется 8 гаплоидных
ядер, вокруг которых формируются аскоспоры. Аскоспоры образу-
ются по способу так называемого “свободного образования клеток”
– часть цитоплазмы сумки обособляется вокруг ядер и одевается
оболочкой. Часть цитоплазмы, не использованная на формирование
аскоспор, называется эпиплазмой. К моменту созревания аскоспор в
эпиплазме происходит превращение гликогена в сахар, тургорное
давление в сумке возрастает (у некоторых видов – до 10-13 атмо-
сфер), и аскоспоры с силой выбрасываются.
Образование сумок по способу крючка типично для большинст-
ва аскомицетов, но встречаются также и другие способы полового
процесса и образования сумок. Помимо гаметангиогамии (о которой
речь шла выше) у аскомицетов могут быть другие способы оплодо-
творения, а именно:
1) Дейтерогамия – способ оплодотворения, при котором на три-
хогину попадают мужские клетки (фрагменты гиф, конидии или
спермации), но специализированого мужского органа нет.
2) Соматогамия, при которой сливаются клетки гиф, аскоспоры,
конидии или почкующиеся клетки; ни аскогона, ни антеридия нет.
3) Партеногамия – развитие сумок происходит без предвари-
тельного слияния мужской и жеской клеток, но внутри аскогона поперечная стенка. В результате получаются две дочерние клетки,
каждая занимает половину исходной осевой клетки и половину
пряжки. Обе образовавшиеся клетки – двухъядерные. Из апикаль-
ной клетки такой аскогенной гифы развивается первая сумка. При
этом оба ядра субапикальной клетки переходят в пряжку, которая
даёт либо тоже сумку, либо – после серии одновременных делений
по тому же, (пряжковому) типу – целые пучки сумок
происходит самооплодотворение, антеридия нет.
4) Апомиксис – аскоспоры образуются без слияния ядер, весь
жизненный цикл проходит в гаплоидной фазе (как, например, у Ascobolus equinus).
Развитие сумки может не только крючком, как было описано
выше, но и другими способами:
1) Пряжкой – когда деление клетки в аскогенной гифе сопрово-
ждается выпячиванием боковой стенки гифы (т.е. образованием
пряжки). Затем одно ядро переходит в это выпячивание (пряжку) и
делится – одновременно с оставшимся в клетке. Пряжка загибается
назад и снова соединяется с основной гифой. Между дочерними
ядрами – как в основной гифе, так и в пряжке – закладывается поперечная стенка. В результате получаются две дочерние клетки,
каждая занимает половину исходной осевой клетки и половину
пряжки. Обе образовавшиеся клетки – двухъядерные. Из апикаль-
ной клетки такой аскогенной гифы развивается первая сумка. При
этом оба ядра субапикальной клетки переходят в пряжку, которая
даёт либо тоже сумку, либо – после серии одновременных делений
по тому же, (пряжковому) типу – целые пучки сумок.
2) Цепочкой – аскогенные гифы расчленяются на двухъядерные
клетки, которые непосредственно преобразуются в сумки.
3) Почкованием – когда сумка вырастает прямо из аскогенной
клетки.
Обычно в сумках по 8 аскоспор, но может быть меньше или
больше. Меньшее количество аскоспор образуется, если: 1) было
меньше делений ядер; 2) если часть образовавшихся спор дегенери-
рует; 3) если в каждой аскоспоре по 2 ядра. Больше восьми аскоспор
в сумке получается, если: 1) больше было делений ядер; 2) если ас-
коспоры почкуются (как, например, у Taphrina); 3) если образуются
двухклеточные споры, которые потом распадаются (например, у
гипокрейных грибов из 8 двуклеточных спор образуется 16 одно-
клеточных).
Сумки у аскомицетов бывают разного строения – прототуникат-
ные и эутуникатные. Прототуникатные сумки имеют тонкую не-
дифференцированную оболочку, из которой аскоспоры освобожда-
ются пассивно при её разрушении. Эутуникатные сумки – с более
плотными оболочками и часто обладают специальными приспособ-
лениями для освобождения аскоспор. Эутуникатные сумки подраз-
деляют на унитуникатные и битуникатные.
Унитуникатные сумки имеют однослойную оболочку, битуни-
катные – две оболочки: наружная оболочка (экзоаск) у них жёсткая,
а внутренняя (эндоаск) – эластичная. При созревании аскоспор в
битуникатной сумке повышается давление, экзоаск разрывается.
Через разрыв эндоаск выпячивается наружу, и через его специаль-
ное выводное отверстие выбрасываются аскоспоры. Верхушка су-
мок бывает устроена по-разному. У одних аскомицетов вокруг вер-
шины сумки закладывается бороздка, а примыкающая к бороздке
стенка – утолщается. В результате на вершине сумки образуется так
называемый оперкулюм – “крышечка”, которая при созревании спор
открывается и выпускает споры наружу. Такие сумки называют
“оперкулятными”. У других аскомицетов – иноперкулятных – в
области вершины сумки, с её внутренней стороны, образуется кольцевой валик. Стенка сумки на верхушке становится неодинако-
вой толщины, и при созревании аскоспор под давлением разрывает-
ся в более тонком месте, через разрыв выбрасываются созревшие
аскоспоры.
По характеру формирования и размещения сумок класс Ascomycetes делится на 4 подкласса:
1. Hemiascomycetidae (голосумчатые) – грибы, у которых прими-
тивные прототуникатные сумки образуются непосредственно на
гифах мицелия.
2. Euascomycetidae (настоящие сумчатые грибы) – образуют сум-
ки в плодовых телах, сумки прото- или унитуникатные.
3. Loculoascomycetidae (локулоаскомицеты) – образуют битуни-
катные сумки в аскостромах.
4. Laboulbeniomycetidae (лабульбениевые) – специализированая
группа грибов, паразитирующих на насекомых. Вегетативное тело
лабульбениевых состоит из тканей, половой процесс – сперматиза-
ция, сумки – прототуникатные
Saccharomycetes — Дрожжи
Сахаромицеты (лат. Saccharomycetes) — класс сумчатых грибов (аскомицетов). Сахаромицеты — единственный класс подотдела Saccharomycotina, называемого также «гемиаскомицетами» или «голосумчатыми» (подотдел Hemiascomycotina или класс Hemiascomycetes). К классу сахаромицетов, в свою очередь, относят один порядок Сахаромицетовые (Saccharomycetales). Вплоть до конца XX века к группе голосумчатых относили аскомицеты примитивного строения, у которых не образуются плодовые тела, а сумки формируются на мицелии или из отдельных клеток (у дрожжевых форм).
Вегетативное тело сахаромицетов часто представляет собой отдельные почкующиеся клетки, то есть дрожжевую форму, в определённых условиях они могут образовывать псевдомицелий, есть и виды, имеющие настоящий мицелий. Некоторые виды диморфны — в зависимости от условий развиваются в мицелиальной или дрожжевой форме. К мицелиальным сахаромицетам относится, например, семейство Диподасковые (Dipodascaceae), виды которого встречаются в слизистых истечениях растений, на древесине, в почве, в ассоциациях с беспозвоночными животными (насекомые, кольчатые черви). Размножение — вегетативным, бесполым и половым способом, но вегетативное размножение (почкованием или фрагментацией гиф) у сахаромицетов часто трудно чётко отграничить от бесполого (образования конидий).
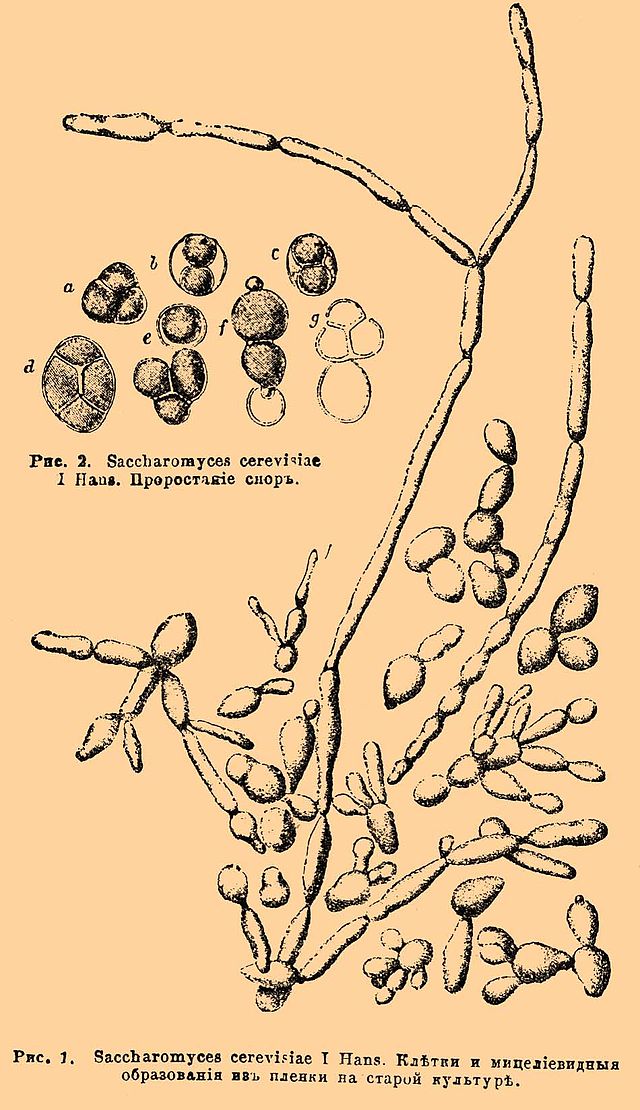
Рисунок 6- Saccharomyces cerevisiae. Иллюстрация из Энциклопедического словаря Брокгауза и Ефрона
Pyrenomycetes – пиреномицеты
У аскомицетов, объединяемых в эту группу порядков, плодовые
тела – перитеции, реже – клейстотеции, сумки – унитуникатные,
расположены в плодовом теле пучком или слоем. Освобождение
спор – активное. Систематика пиреномицетов запутана, существует
множество систем этой группы.
У этих грибов цилиндрические аски обычно находятся в плодовых телах, называемых перитециями, которые внешне напоминают колбу и открываются в окружающую среду отверстием на конце суженной шейки. По форме, окраске и консистенции перитеции сильно варьируют, они бывают одиночными или собранными в группы, иногда погруженными в особые компактные, образованные гифами структуры, называемые стромами. Так, у обычно встречающегося на навозе вида Sordaria fumicola перитеции одиночные, длиной ок. 0,5 мм, а у Daldinia concentrica сотни плодовых тел находятся по периферии разделенной на четкие концентрические зоны стромы диаметром иногда более 2,5 см.
Большинство микологов выделяет 5 порядков пиреномицетов:
1) Erysiphales – образуют сумки в виде упорядоченного слоя в
замкнутых плодовых телах – клейстотециях.
2) Sphaeriales (=Xylariales) – образуют сумки в тёмноокрашен-
ных перитециях, формирующихся в стромах или на мицелии.
3) Diaporthales – образуют тёмноокрашенные перитеции, при со-
зревании заполняющиеся слизью, в которой свободно плавают
сумки. 109
4) Clavicipitales – имеют нитевидные многоклеточные аскоспо-
ры, которые после выхода из сумки распадаются на отдельные
клетки.
5) Hypocreales – аскоспоры бывают одно-, двух- и многоклеточ-
ные, сумки формируются в яркоокрашенных стромах.
Порядок Hypocreales
Порядок объединяет грибы (более 100 родов) с мягкими мяси-
стыми перитециями яркой или светлой окраски. Перитеции образу-
ются на мицелии – на поверхности или внутри субстрата, на мице-
лиальном сплетении (субикулюме) или на хорошо развитых стромах.
Перитеции имеют хорошо развитый перидий и выводное отверстие
(остилу), выстланное перифизами. Настоящие парафизы в перите-
циях отсутствуют, но развиваются апикальные парафизы – стериль-
ные гифы, врастающие в полость молодого перитеция сверху. Сум-
ки цилиндрические или булавовидные, отходят от дна и боковых
стенок перитеция, врастая между апикальными парафизами. В зре-
лых перитециях апикальные парафизы часто полностью разрушают-
ся. У многих гипокрейных перитеции развиваются в стромах. Стро-
мы бывают следующих типов:
1) Субикулярные. Субикулюм – это довольно рыхлое сплетение
гиф гриба на поверхности субстрата.
2) Базальные – плотные сплетения гиф, которые закладываются
под корой на мицелии и прорывают кору по мере разрастания. Ба-
зальные стромы обычно мелкие, подушковидные. Перитеции обыч-
но погружены в такие стромы только основаниями. 127
3) Компактные стромы с погружёнными в них перитециями.
Компактные стромы могут быть разнообразной формы и достигают
довольно крупных размеров.
Стромы гипокрейных обычно имеют светлую (белую или желто-
ватую) либо яркую окраску – обычно жёлтую, оранжевую, красную,
реже фиолетовую или синюю. Иногда встречаются тёмно-зелёные,
синевато-чёрные и коричневые стромы. В цикле развития гипокрей-
ных грибов важную роль играет конидиальная стадия. Чаще всего
конидии гипокрейных являются фиалоспорами, у некоторых – алев-
риоспоры и бластоспоры. У грибов этого порядка в цикле развития
часто образуются две конидиальных стадии. Например, у родов
Nectria и Gibberella, имеющих конидиальные стадии Fusarium, об-
разуются макро- и микроконидии (фиалоспоры). По особенностям
строения стром, аскоспор и анаморф в порядке выделяют 3 семейст-
ва: Nectriaceae, Hypomycetaceae, Hypocreaceae.
род Nectria и Gibberella
У грибов этого порядка в цикле развития часто образуются две конидиальных стадии. Например, у родов Nectria и Gibberella, имеющих конидиальные стадии Fusarium, образуются макро- и микроконидии (фиалоспоры). По особенностям строения стром, аскоспор и анаморф в порядке выделяют 3 семейства: Nectriaceae, Hypomycetaceae, Hypocreaceae.
У грибов семейства Nectriaceae перитеции образуются на поверхности субстрата или на строме. Стромы обычно небольшого размера, подушковидные.
Центральный род семейства – Nectria. У грибов этого рода аскоспоры двухклеточные, не распадающиеся в зрелости на отдельные клетки. Повсеместно встречаются нектрия киноварно-красная (Nectria cinnabarina). Она обычно обитает на мёртвых сухих ветвях деревьев, но может развиваться как раневой паразит. Нередко этим грибом поражаются ягодные культуры –чёрная и красная смородина, крыжовник. Род Gibberella объединяет 13 видов.
У этих грибов мягкие кожистые перитеции чёрного или чёрно-коричневого цвета, они образуются прямо на субстрате или на тёмно-синих стромах. В тёплых и влажных районах умеренной зоны, тропиках и субтропиках широко распространён вид Gibberella fujikuroi (гибберелла Фуджикоро). Этот гриб поражает корни, стебли и семена многих растений, особенно часто – злаков (риса, сахарного тростника, кукурузы, сорго). У
больных растений риса вытягиваются междоузлия, листья становят-
ся более узкими, длинными, а при сильном поражении – хлоротич-
ными. Такие симптомы вызывают ростовые вещества – гибберелли-
ны, образуемые в поражённом грибом растении. Листья отмирают
один за другим, семена не образуются – метёлки остаются пустыми.
В итоге растения гибнут. В Японии эту болезнь называют “баканэ”
или “болезнь дурных побегов”, в Китае - "белый стебель”. В от-
дельные годы от этой болезни гибнет до 20% урожая. Инфекция
передаётся преимущественно через почву или с семенами. Гриб зи-
мует или переносит период засухи в почве либо на растительных
остатках в виде мицелия, конидий, а иногда в виде перитециев.Массовое развитие гриба происходит при повышенных температурах (30-35 °С).
БИОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ МЕР БОРЬБЫ
Агротехнические меры борьбы
Агротехнические мероприятия - совокупность научно обоснованных приемов обработки почв в целях воспроизводства плодородия земель сельскохозяйственного назначения.
Агротехнические меры борьбы с ядовитыми, вредными и сорными растениями на сенокосных и пастбищных угодьях предусматривают:
• уничтожение источников инфекции (заделка в почву, сгребание, и компостирование опавших листьев);
• оптимальную нагрузку скота на пастбище;
• подкашивание трав в загонах до обсеменения ядовитых и вредных растений, а также выкапывание корневищ этих растений;
• своевременное разбрасывание экскрементов животных на участках
• загонный и загонно-порционный выпасы животных;
• осушение заболоченных участков;
• поддержание высокой агротехники, своевременные и высококачественные химические обработки насаждений медьсодержащими, карбонатными и другими фунгицидами, особенно в период до цветения в первой половине лета.
• создание условий, неблагоприятных для развития возбудителя болезни (незагущенные, хорошо продуваемые и освещаемые солнцем насаждения и кроны растений);
Химические меры борьбы
• опрыскивание почвы приствольных кругов 10% аммиачной селитрой
• обработка семян перед посевом
• грибница проникает в проростки. Опрыскивание всходов ядохимикатами(формалин).
• выведение сортов, устойчивых к грибковым заболеваниям.
ЗАКЛЮЧЕНИЕ
Курс фитопатологии является важным звеном в подготовке специалиста сельского хозяйства. Знакомство с основами фитопатологии важно для студентов, прежде всего для того чтобы отчетливо можно было осознать тот вред, который причиняют фитопатогенные организмы сельскому, а также лесному хозяйству. Фитопатология исследует болезни конкретных культур, рассматривает применительно к каждой из них видовой состав возбудителей, симптомы заболеваний, приемы защиты. Развитие фитопатологии привело к возникновению новых наук – этиологии, фитоиммунологии, эпифитотиологии, защиты растений от болезней. Знание фитопатологии очень важно для правильного восприятия и усвоение студентами курсов защиты растений. Особое значение приобретает изучение фитопатологии в связи с широко развернувшейся химизацией народного хозяйства, решением продовольственной программы и проблемы окружающей среды. Специалист по сельскому и парковому хозяйству должен знать механизм действия на живые организмы веществ, применяемых в сельском хозяйстве с целью защиты от вредителей и болезней, увеличения производительности труда и повышения продуктивности. Основной задачей изучения курса фитопатологии является изучение болезней растений, вызываемых разными группами возбудителей, определение болезней и их возбудителей, изучение болезнетворных и других факторов, разработка рациональных мер борьбы с болезнями.
При изучении курса студент должен знать: Причины, приводящие к возникновению болезней, особенности их развития, возбудителей, методы защиты, закономерности развития патологических процессов и факторов, воздействующих на них. Изучить взаимодействие растения и возбудителя в определенных условиях среды, иммунитет растений, прогноз появления и распространения болезней, биологический метод защиты растений, карантин растений, породы деревьев и кустарников, устойчивых к болезням.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Белякова Г.А., Дьякова Ю.Т., Тарасов К.Л. Ботаника: в 4 т. Т.1. Водоросли и грибы: учебник для студ. Высш. учебн. заведений / Г.А. Белякова, Ю.Т. Дьякова, К.Л. Тарасов. – М: Издательский центр «Академия». 2006.- 320 с.
2. Букреев Д.Д. Модульный метод обучения в курсе «Общая фитопатология». Учебное пособие для студентов агрономического факультета, Курск: Издательство А.П. «Курск», 2002. – 143с.
3. Власов И.Ю., Ларина Э.И. Сельскохозяйственная вирусология. М.: «Колос» 1982
4. Горленко М.В. Бактериальные болезни растений. М.: «Колос», 1979
5. Егураздова А.С. Микоплазменные болезни сельскохозяйственных растений. М.: «Колос», 1990
6. Защита растений в устойчивых системах землепользования (в 4-х книгах). Под общ. ред. Д. Шпаар, книга 1, Торжок ООО «Вариант» , 2003, 390 с.
7. Левитин М.М. Генетические основы изменчивости фитопатогенных грибов (всесоюзн. акад. сельскохозяйственных наук. – Л.: Агропромиздат. 1986)
8. Мюллер Э., Леффлер В. Микология / Пер. с нем. М.: Мир, 1995. 343 с.
9. Поглушной И.С. Микофлора плодовых деревьев ССР. М.: Наука 1971
10. Переведенцева Л.Г. « Микология: грибы и грибоподобные организмы» Издательство «Лань» , 2012
11. Попкова К.В. Общая фитопатология. – М.: Агропромиздат, 2005. – 399с.
12. Соколов М.С. Биологизация и биобезопастность в защите растений в 21 веке в России// Актуальные вопросы в биологизации растений.- Пущино, 2000 – с. 26-32
13. Хоуксворт Д.Л. Общее количество грибов. Их значение в функционировании экосистем. Сохранение и значения для человека // Микология И фитопатология 1992. Т. 26, Вып.2 с. 152-166.
14. Хохряков М.К. Вредные и полезные грибы. Издание 2. Ленинград: Колос, 1969
PAGE \* MERGEFORMAT 1
Классификация Пиреномицетов. Хозяйственное значение и биология грибов семейства Нектриевые