ВЛИЯНИЕ АНТРОПОГЕННОЙ НАГРУЗКИ НА ФИТОПЛАНКТОН НОВОРОССИЙСКОЙ БУХТЫ
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное государственное бюджетное образовательное учреждение
высшего профессионального образования
«КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
(ФГБОУ ВПО «КубГУ»)
Кафедра биологии и экологии растений
ДОПУСТИТЬ К ЗАЩИТЕ В ГАК
Заведующий кафедрой – канд. биол. наук,
доцент______________М.В. Нагалевский
«____»________________________ 2015 г.
ДИПЛОМНАЯ РАБОТА
ВЛИЯНИЕ АНТРОПОГЕННОЙ НАГРУЗКИ НА ФИТОПЛАНКТОН НОВОРОССИЙСКОЙ БУХТЫ
Работу выполнил ______________________________________В.Н. Вартеваньян
(подпись, дата)
Факультет биологический
Специальность 02.08.03 Биоэкология
Научный руководитель,
канд. биол. наук,
доцент __________________________________________________О.В. Букарева
(подпись, дата)
Нормоконтролёр,
преподаватель___________________________________________А.М. Иваненко
(подпись, дата)
Краснодар 2015
РЕФЕРАТ
Дипломная работа выполнена на 67 страницах машинописного текста. Содержит введение, 4 главы, заключение, библиографический список, насчитывающий 45 литературных источников. Работа проиллюстрирована 5 таблицами и 6 рисунками в тексте работы и 22 таблицами и 4 рисунками в приложении.
Ключевые слова: ФИТОПЛАНКТОН, АНТРОПОГЕННАЯ НАГРУЗКА, БИОМАССА, ДИНАМИКА РАЗВИТИЯ, КОЛИЧЕСТВЕННЫЙ СОСТАВ.
Работа посвящена изучению влияния антропогенной нагрузки на фитопланктон Новороссийской бухты. В результате проведённых исследований составлен список фитопланктонных водорослей, содержащий 95 видов, проведён химический анализ воды, выявлено влияние антропогенной нагрузки на фитопланктон, изучена сезонная динамика биомассы и количественного состава фитопланктона Новороссийской бухты.
СОДЕРЖАНИЕ
Введение……………………………………………………………………..……... 4
1 Обзор литературы…….…………………………………………………….......... 6
2 Описание района исследования………………………………...……………... 11
2.1 Географическое положение………….……………………………….……. 11
2.2 Климат………………………………………………………..……………... 11
2.3 Гидрология…………………………..……………………….…...…...……. 13
2.4 Почвы………………………………………..…………..……..……………. 14
3 Материалы и методы исследования……………………………….………..…. 16
4 Влияние антропогенной нагрузки на фитопланктон Новороссийской бухты………………………………………………………………………......... 19
4.1 Систематический состав фитопланктона………………………..……...... 19
4.2 Сезонная динамика биомассы и количественного состава
фитопланктона……………………………………………………………... 24
4.3 Кислотность и уровень загрязнения вод в Новороссийской бухте…….. 33
4.4 Влияние антропогенной нагрузки на фитопланктон Новороссийской
бухты…………….………………………………………………………….. 34
Заключение……………………………………………………………..…….…… 37
Библиографический список…………………………………………..….………. 39
Приложение А Показатели численности и биомассы планктонных
водорослей Новороссийской бухты………………...…………. 44
Приложение Б Внешний вид некоторых представителей фитопланктона
Новороссийской бухты……………………..……………….… 66
ВВЕДЕНИЕ
В настоящее время любые вопросы, связанные с охраной, управлением и восстановлением различных водоёмов не могут быть решены без фундаментальных знаний о природных свойствах водных экосистем, их структуре и функционировании. Наиболее адекватно отражают природные свойства водоёмов процессы превращения вещества и энергии, которые замыкаются на круговороте органического вещества [Одум, 1975].
Скорость этого процесса обусловлена жизнедеятельностью гидробионтов во всей сложности их трофических отношений. Планктонные водоросли являются первым и основным биотическим звеном в трофических взаимоотношениях водных экосистем. Постепенное изменение любого из факторов в первую очередь находит свое отражение на качественных и количественных показателях фитопланктона. В этом отношении определённую ценность в целях мониторинга изменений состояния природных объектов несёт многолетнее изучение фитопланктона водоёмов [Сорокин, 1996].
Наибольший антропогенный пресс испытывают прибрежные районы моря, особенно закрытые и полузакрытые бухты. Большая изрезанность и протяженность береговой линии, малые глубины и ограниченный водообмен с морем способствует накоплению в бухтах загрязнения. Это связано с тем, что в бухтах имеются стационарные источники загрязнения (выпуски сточных вод городской канализации, стоянки судов в порту, стоки рек и др.) и эпизодически могут возникать аварийные ситуации (сброс балластных вод с судов, аварии с нефтеналивными судами, трубами транспортировки нефтепродуктов и др.). Последствия сбросов зависят не только от объёма (количества) и токсичности загрязняющих веществ, попавших в акваторию бухты, но и от целого комплекса физико-химических и биологических условий, результатом влияния которых является отклик фитопланктона [Сеничкина, 2002].
Основные изменения фитопланктонного сообщества в загрязнённых морских водах заключается в обеднении видового состава, увеличении численности и биомассы фитопланктона (возникновение «цветения»), изменении вклада основных групп водорослей в суммарную численность и биомассу фитопланктона и уменьшении среднего объёма (массы) и размера клеток водорослей. Такой отклик является следствием экологической адаптации водорослей планктона к условиям резкого увеличения концентраций биогенных элементов и токсических веществ в загрязнённых районах моря и является надёжным индикатором качества морских вод [Ясакова 2008a].
Эта проблема затронула и Новороссийскую бухту. Наибольший ущерб экологии бухты наносят нефтяные выбросы, поскольку Новороссийск является крупнейшим портом страны, да к тому же через его терминалы на экспорт отправляется треть всего добываемого в России чёрного золота. Помимо этого в гораздо большей степени акваторию загрязняют промышленные и бытовые канализационные стоки самого города. Следствием этого является обвальное снижение биомассы водорослей, морских животных, а также гибель видов с узким коридором толерантности к интоксикации нефтью (таковых видов в экосистемах Чёрного моря большинство). Исчезновение фитопланктона может привести к полному разрушению пищевых цепей бухты [Михайловская, 1903].
Целью данной работы является изучение влияния антропогенной нагрузки на фитопланктон Новороссийской бухты.
Для достижения цели исследований были поставлены следующие задачи:
Изучить систематический состав фитопланктона Новороссийской бухты;
Проследить сезонную динамику биомассы и количественного состава фитопланктона;
Определить кислотность и провести химический анализ воды;
Выявить влияние антропогенной нагрузки на фитопланктон Новороссийской бухты.
1 Обзор литературы
Планктонные водоросли являются начальным звеном трофической цепи в океане, от их качественных и количественных характеристик зависит развитие организмов других трофических уровней. Исследование фитопланктона даёт возможность судить о тенденциях изменений в структуре и функционировании
прибрежных экосистем. Сведения о состоянии планктонных водорослей могут быть использованы в системе экологического контроля и ранней диагностики санитарного состояния водоёма [Вершинин, Моргунов, 2003].
В акваториях, подверженных антропогенному воздействию, главным образом в виде бытовых и промышленных сбросов, прослеживаются изменения качественного и количественного состава фитопланктона. Чаще всего это проявляется в «цветении» воды, вызванном интенсивным развитием отдельных видов водорослей, некоторые из которых проявляют токсичные свойства. Это снижает рекреационную ценность черноморского побережья и существенно влияет на состояние всей экосистемы в целом [Виноградова, Василёва, 1992].
В российском секторе Чёрного моря негативным изменениям, связанным с повышением уровня эвтрофикации, в первую очередь, подвержен фитопланктон шельфовой зоны моря, в том числе бухт, расположенных на его побережье. Поэтому в современный период сведения о состоянии качественных и количественных характеристик планктонных микроводорослей, развивающихся как в открытых прибрежных водах Чёрного моря, так и в полузамкнутых акваториях бухт портовых и курортных городов представляют определённый интерес [Ясакова, 2005].
История изучения фитопланктона акватории Новороссийской бухты обусловлена актуальностью проблем, связанных с ростом индустриализации региона, усилением антропогенного воздействия в целом на водную систему и фитопланктона в частности [Болгова, Луговая, 2010].
Изучение фитопланктона северо-западной акватории Чёрного моря было начато в конце XIX в. С 1900-х гг. начались постоянные исследования фитопланктона в данном районе и в Новороссийской бухте. М.Б. Иванов [1982] обнаружил и опубликовал списки, включавшие 372 вида и внутривидовых таксонов водорослей, а затем пришёл к выводу о том, что в планктоне этой части моря насчитывается 461 вид.
Многолетние исследования, выполненные этим же исследователем, дали возможность по-новому пересмотреть районирование северо-западной части с учётом состава фитопланктона.
В литературе [Прошкина-Лавренко, 1955] описаны сезонные изменения видового разнообразия и биомассы фитопланктона, биомассы основных систематических групп, а также особенности вертикального распределения. Установлено, что показатели биомассы в этой части Чёрного моря выше, чем в других районах.
Возросшее антропогенное влияние на экосистему Чёрного моря и Новороссийской бухты вызвало заметные изменения в структуре и динамике развития фитопланктона [Киселёв, 1969].
Увеличение видового разнообразия фитопланктона отмечено в составе всех отделов, за исключением эвгленовых водорослей. Как и в прежние годы, его основу составляют диатомовые и динофитовые водоросли, соотношение которых изменилось. Вклад первых (35,7 %) уменьшился по сравнению с 1954–1960 гг., когда диатомовые составляли 48,3 % найденных видов, а динофитовых – увеличился, составляя соответственно 31,2 и 20,4 %. Изменения отмечены также в составе пресноводного комплекса видов: значение зелёных в разные периоды исследования возросло с 16,7 % в 1954–1960 гг. до 18,5 % в настоящее время, а сине-зелёных (5,7 %) несколько уменьшилось: последние составляли в этот период 6,4 % найденных видов [Нестерова, 2001].
В последнее десятилетие исследования фитопланктона регулярно проводились в прибрежной зоне, что привело к увеличению списка видов диатомовых в основном за счёт случайно-планктонных видов (45,6 % по сравнению с 22,8 % в 1954–1960 гг.), попадающих в водную толщу при вертикальном перемешивании водных масс [Теренько, 2004].
Увеличение видового разнообразия динофитовых наблюдалось в 1973–1993 гг., когда в их составе было найдено 36 новых для данного района видов. В дальнейшем обработка живых, нефиксированных проб фитопланктона позволила выявить в составе динофитовых водорослей много новых видов, так как при грубой фиксации проб формалином разрушаются многие беспанцирные или обладающие мягким панцирем виды [Георгиева, Сеничкина, 1996].
Фитопланктон в своём развитии проходит через ряд закономерных, направленных во времени изменений – сукцессии, представляющих собой процесс самоорганизации сообщества. Классическая схема сукцессии начинается с развития мелкоклеточных диатомовых водорослей (первая стадия), на смену которым приходят крупноклеточные диатомовые (вторая стадия), а заканчивается доминированием динофитовых водорослей (третья стадия). Описанная схема отмечалась для фитопланктона 1960–1970-х годов и включала две полных сукцессии, а именно: весеннюю, летне-осеннюю и, возможно, зимнюю [Виноградов, Шукшина, Сапожников, 2002].
Одним из первых «цветение» воды в северной части Чёрного моря описал П.И. Усачёв [1928]. За период 1973–2001 гг. в различных районах этой части моря и Новороссийской бухте зарегистрирован 151 случай «цветения» воды, вызванный развитием 41 вида и разновидностей водорослей. Среди возбудителей этого явления появились новые виды (например, Emiliania huxleyi), вспышки развития других усилились и участились [Нестерова, 2001].
В сезонной динамике биомассы фитопланктона Новороссийской бухты в 1973–1993 гг. было выделено два максимума весенний и позднелетний. Весенний максимум образовывали диатомовые водоросли, формировавшие «цветение» (Sceletonema costatum, Chaetoceros socialis), позднелетний наблюдался во время одновременного развития диатомовых (Сerataulina pelagica, Leptocylindrus danicus) и динофитовых (Protoperidinium conicum) водорослей. Из них наиболее сильным был весенний максимум. В случае тёплой зимы позднелетний максимум по абсолютным величинам превосходил весенний [Суханов, 1982].
В разные периоды исследований количественные показатели развития фитопланктона в северной части Чёрного моря и Новороссийской бухты были неодинаковы. Наибольшая биомасса фитопланктона отмечена в 1973–1980 гг., когда под влиянием эвтрофирования в его структуре и развитии наблюдались изменения, а средняя биомасса по сравнению с 1950–1960 гг. возросла в 17 раз [Селифонова, Ясакова].
С 1981 по 1993 г. биомасса фитопланктона стала постепенно уменьшаться. В 1981–1990 гг. отмечено меньшее количество (44) случаев «цветения» воды, также уменьшились площадь распространения и интенсивность развития отдельных видов (Сerataulina pelagica) [Сеничкина, 1983]. Вместе с тем среди возбудителей «цветения» воды появились мелкоклеточные сине-зелёные водоросли. В 1990–1993 гг. интенсивность развития фитопланктона уменьшилась, что отразилось на средней величине биомассы, которая была минимальной. Кроме того, именно в эти годы усилилась вегетация Sceletonema costatum, отнесённой к индикаторам гиперэвтрофных вод [Сорокин, 1982].
С 1980-х гг. до настоящего времени наблюдается постепенное уменьшение биомассы, сокращение случаев «цветения» воды и территорий, охваченных этим явлением, что может служить показателем ослабления влияния эвтрофирования на экосистему северо-западной части Чёрного моря. Вместе с тем его последствия продолжают отражаться на популяционном уровне [Ясакова, 2012].
Изучение состояния фитопланктона мелководных прибрежных акваторий Чёрного моря, а также изменений, происходящих в структуре этого сообщества, позволяет определить степень антропогенного влияния на него в настоящее время [Восконьян А.В., Восконьян В.Г., 2007].
В работе З.Л. Рейдгарда [1983] даётся оценка качественных и количественных изменений фитопланктонных сообществ в связи с повышением уровня антропогенной нагрузки. Публикации последних лет, написанные Н.В. Морозовой-Водяницкой [2004], О.Н. Ясаковой [2004], А.О. Вершининым [2005], затрагивают проблемы, связанные со структурной перестройкой планктонного альгоценоза, выражающейся в неоднородности видового состава и количественного развития, а также увеличении числа потенциально-токсичных видов, вселением их в экосистему Чёрного моря и массовым развитием.
2 Описание района исследования
2.1 Географическое положение
Новороссийская бухта является одной из лучших на Чёрном море по своему географическому положению и гидрографическим данным.
Эта бухта представляет собой природную гавань на Чёрноморском побережье Кавказа. Образована Суджукской косой и мысом Дооб. Длина 15 км, ширина у входа 9 км, в средней части 4,6 км, глубина 21–27 м. Западный берег бухты пологий, постепенно переходящий в Навагирский хребет. Вершины Навагирского Хребта – горы Сахарная Голова (540 м) и Колдун (Мысхако, 440 м). Гора Колдун – остаток прибрежного хребта, часть которого провалилась в результате геологических процессов и образовала вход в бухту. Продолжение этого хребта на другой стороне бухты у Геленджика – гора Дооб (452 м). Восточный берег бухты на всем протяжении высокий, местами обрывистый. Береговая линия почти не изрезана. Длина восточного берега 16,7 км, считая от мыса Дооб до восточного мола. К северо-западу от мыса Дооб расположены мысы Пенай и Шесхарис. В районе Мысхако берег на протяжении 2 км скалистый, затем сменяется галечным и переходит в пересыпь Суджукской косы. Последняя ограждает от моря мелководную Суджукскую лагуну, связанную с морем небольшим прорывом. В 1,6 км от основания Суджукской косы расположен мыс Любви. В бухту впадает река Цемес, именем которой иногда и называют бухту [Агроклиматический справочник … , 1961].
2.2. Климат
Климат в районе Новороссийской бухты относится к морскому климату умеренных широт с тёплым летом и умеренно мягкой зимой. Средняя температура января 2,6 С, августа 23,7 С, среднегодовая 12,7 С. Для района характерна значительная изменчивость термического режима [Агроклиматический справочник … , 1961].
Безморозный период составляет 232 дня в году. Зимой могут наблюдаться очень низкие, а летом – очень высокие температуры. По многолетним данным, абсолютный минимум составляет минус 24 С, абсолютный максимум 41 С. Вследствие посезонной смены термических уровней над Евроазиатским континентом происходит перестройка барических систем, поэтому атмосферная циркуляция имеет выраженный характер. Зимой при распространении Сибирского антициклона на восток Европы и на Чёрное море возникают устойчивые и сильные северо-восточные ветра, приносящие холодный и сухой континентальный воздух умеренных широт. При ослаблении Сибирского антициклона создаются условия для поступления теплового воздуха с Атлантики и Средиземного моря, что приводит к развитию циклонической деятельности. При этом происходит повышение температуры воздуха и смена ветров на западные и южные направления. Все указанные режимы наиболее интенсивно проявляются в северо-восточной части моря и районе Новороссийской бухты [Фёдоров, Радченко, Капков, 2010].
Весной, в марте-апреле, нагрев материка приводит к уменьшению барических градиентов между морем и сушей, к выравниванию атмосферного давления, при этом преобладают южные и юго-западные ветры, несущие большое количество влаги. Летом, начиная с мая, характерно пониженное давление, образующееся над материком. При этом ощущается влияние Азовского субтропического антициклона, отроги которого распространяются на северо-восточную и восточную части моря, создают длительные периоды спокойной погоды с большим количеством ясных дней. Осень является переходным периодом от летнего типа циркуляции атмосферы к зимнему [Агроклиматический справочник … , 1961].
Наибольшее количество осадков выпадает зимой, наименьшее – летом. Среднегодовое количество осадков по данным многолетних наблюдений 724 мм. Число дней с осадками в году по многолетним наблюдениям равно 139. Погодный мониторинг осуществляется стационарной метеостанцией на нефтегавани «Шесхарис», а также – с морских автоматических гидрометебуев. Для Новороссийской бухты характерен повышенный ветровой режим, среднегодовая скорость 4,6 м/с, преобладающее направление ветров – северо-восточное и южное, повторяемость этих ветров по многолетним наблюдениям 19,8 %. В среднем 50–60 дней в году сопровождаются сильными от 20 м/с до 40 м/с и более северо-восточными ветрами. При наступлении боры ветер усиливается очень быстро, холодные массы устремляются с вершин восточных гор вниз в бухту и на город. Температура понижается до минус 7–9 и до минус 20 С [Сикачинский, 1989].
Повторяемость южных и юго-восточных ветров для бухты составляет до 16 % в году, они достигают значительных скоростей (до 20 м/с) и распространяется в узком секторе вдоль береговых хребтов. Туманы для Новороссийской бухты редки, в среднем около 10–15 дней, в году, чаще в первой половине мая [Агроклиматический справочник … , 1961].
2.3 Гидрология
Течения Чёрного моря слабы, их скорость редко превышает 0,5 м/с, возникают в поверхностном слое в основном под воздействием речных стоков и под действием ветров. В Новороссийской бухте преобладает антициклональный тип циркуляции воды с отклонениями при действии северо-восточных ветров. Скорость течения 0,15–0,20 м/с, при действии сильных ветров увеличивается до 0,3–0,4 м/с [Фёдоров, 1979].
Волновой режим в Новороссийской бухте складывается из двух типов волнений.
Первое проникает в бухту со стороны моря, наиболее интенсивное при действии южных и юго-западных ветров. Так, согласно карте волновых полей, на входе в бухту при действии юго-западной синоптической обстановки для традиций ветра 10–15 м/с высота волны достигает 2,5 м, а длина волны – 40 м, для традиций ветра 15–20 м/с длина волны достигает 65 м [Агроклиматический справочник … , 1961].
Второе волновое поле возникает в самой бухте под действием юго-восточных, северо-западных и северо-восточных ветров. При сильном северо-восточном ветре в глубине бухты, на выходе из нее и под западным берегом возникает толчея с высотами волн до 2–2,5 м и зоной затишья со стороны северо-восточного берега. При ветрах остальных румбов бухта достаточно надежно укрыта от волнения [Сикачинский, 1989].
Среднегодовая температура воды на поверхности 14,4 С и колеблется от 12,8 до 15,3 С, максимальная температура воды 28 С, минимальная около 0 С. Количество дней в году с температурой воды на поверхности выше 20 С около 110 [Виноградов, Шукшина, Сапожников, 2002].
Солёность морской воды составляет в среднем 6,5 %. Зимой солёность уменьшается, летом и осенью возрастает. Максимальная амплитуда изменения среднемесячной солёности приходится на январь (7,3 %) минимальная наблюдается в октябре (2,0 %). Наибольшая наблюдаемая солёность воды в бухте составляла 19 %, наименьшая – 2,0 % [Сикачинский, 1989].
Основной массив значений плотности вод (1011–1016 кг/м3) вписывается в значения, прописанные в Лоции Чёрного моря (1014–1016 кг/м3) для зимнего времени, со снижением до 1011 кг/м3 для летнего времени, при этом значения 1017 кг/м3 объясняются погрешностью измерения [Сикачинский, 1989].
2.4 Почвы
В Новороссийском районе перегнойно-карбонатные почвы, наиболее благоприятные для выращивания винограда и других культур. Перегнойно-карбонатные почвы расположены в нижних частях склонов, выше по склонам – лесные бурые почвы. По понижениям рельефа почвообразование идёт по лесному типу. Цемесская долина покрыта почвами болотного типа с преобладанием луговых и лугово-болотных почв. Также в почвенном покрове Новороссийского района встречаются и коричневые почвы, хотя и значительно реже вышеописанных. Среди них более распространены средне- и маломощные почвенные разности различной щебневатости, каменистости и степени смытости. Далее следуют коричневые типичные и выщелоченные почвы (мощные, среднемощные и маломощные) [Ясакова, 2000].
3 Материалы и методы исследования
Объектом исследований является фитопланктон Новороссийской бухты. Материалом для проведения работы послужили литературные данные, записи полевых дневников и фотографии. Систематический состав фитопланктона был проведён на основе монографических сводок [Аннотированный каталог водорослей … , 1998], «Диатомовых водорослей СССР» под. ред. А.И. Прошкиной-Лавренко [1974], «Панцирных жгутиконосцев морей и пресных вод СССР» К.В. Киселёва [1950] и «Определителя низших растений. Водоросли» под. общ. ред. Л.И. Курсанова [1953].
Способ отбора (места отбора, пространственные и временные масштабы отбора, выбор оборудования, количество проб) определялся в зависимости от поставленной задачи.
Для количественного учёта фитопланктона сборы проб проводились при помощи батометра. Барометр, предложенный Робертом Хуком, представляет собой прямую призму, изготовленную из органического стекла, емкостью 5 л. Прибор снабжён верхней и нижней крышкой, которые открываются при опускании батометра в воду и закрываются при подъёме обратным током воды. К батометру снизу крепится грузик, сверху – трос, размеченный на требуемые интервалы. Данная процедура выглядела следующим образом. Батометр с открытыми крышками опускался в воду. При достижении необходимой глубины необходимо было дернуть за трос (можно 2 – 3 раза для подстраховки), чтобы крышки закрылись. Обратный ток воды удерживал крышки закрытыми. Вынув батометр из воды, следовало опустить его в ведро, осторожно приподнять нижнюю крышку и слить воду. Отбор проб фитопланктона батометрами позволяет учитывать водоросли всех размерных групп [Брянцева, Курилов, 2003].
Численность фитопланктона в морских водоёмах, за исключением отдельных видов во время их массового развития, очень низка, порядка нескольких клеток в миллилитре, поэтому для количественного учёта всех водорослей необходимо концентрирование проб. Существует несколько методов концентрирования фитопланктона, каждый из которых имеет свои достоинства и недостатки. В нашем случае использовался отстойный метод. Фиксированные пробы в стеклянных банках или цилиндрах в неподвижном состоянии в затемнённом месте отстаивают в течение нескольких часов или дней в зависимости от объёма пробы. После осаждения планктона пробы концентрируют путём декантирования (сливания) по каплям среднего слоя воды с помощью тонкого стеклянного сифона с концом, загнутым вверх на 2 см. Сконцентрированный таким образом материал годен для микроскопического исследования в счётной камере [Радченко, Капков, Фёдоров, 2010].
В водных пробах при пониженных температурном и световом режимах фитопланктон может сохранять жизнеспособность в течение некоторого времени. В сетных выборках или пробах, взятых во время цветения, период жизнеспособности может быть значительно короче из-за высокой концентрации водорослей. Для увеличения жизнеспособного периода рекомендуется снизить уровень физиологической активности гидробионтов. Водные пробы, как рекомендует Трондсен, желательно хранить в полной темноте при температуре 15 °С, если температура моря ниже 10 °С, или при 2 – 10 °С, если температура моря выше 15 °С. В качестве фиксаторов фитопланктона использовался формалин и раствор Люголя. Оба фиксатора использовались как самостоятельно, так и с различными добавками в зависимости от цели исследования [Фёдоров, Радченко, Капков, 2010].
Количественному учёту могут подвергаться только количественные пробы фитопланктона. Данные о численности водорослей являются исходными для определения их биомассы. Подсчёт численности водорослей осуществлялся на специальных счётных стёклах (разграфлённых на полосы и квадраты), на поверхность которых штемпель-пипеткой определённого объёма (большей частью 0,1 см3) наносят каплю воды из тщательно перемешанной исследуемой пробы [Брянцева, Курилов, 2003].
Линейные размеры клеток являются важным диагностическим признаком. Они использовались для расчёта объёма клеток. Измерения проводились с помощью окулярного линейчатого микрометра [Брянцева, Курилов, 2003].
Для определения биомассы водорослей счётно-объёмным методом необходимо располагать данными об их численности в каждой конкретной пробе для каждого вида отдельно и их средних объёмах (для каждого вида из каждой конкретной пробы). Существуют разные методы определения объёма тела водорослей. Мы использовали стереометрический метод, который считается наиболее точным. При использовании этого метода тело водоросли приравнивается к какому-нибудь геометрическому телу или комбинации таких тел, после чего их объёмы вычисляют по известным в геометрии формулам на основании линейных размеров конкретных организмов. Иногда пользовались готовые, вычисленные ранее средние объёмы тела для разных видов водорослей, которые приводятся в работах многих авторов. Биомассу рассчитывали для каждого вида отдельно, а затем суммировали [Иванов, 1965].
4 Влияния антропогенной нагрузки на фитопланктон Новороссийской бухты
4.1 Систематический состав фитопланктона
На протяжении весеннего, летнего и осеннего сезонов на четырёх станциях, таких как порт, Западный мол, мыс Любви и Южная Озереевка, (открытая часть бухты) производился отбор проб воды (рисунок 1).
В ходе дальнейшего исследования в Новороссийской бухте было обнаружено 95 вида планктонных водорослей, из которых диатомовые составляют 49 видов, динофитовые – 40 видов, эвгленовые – 3 вида, сине-зелёные – 2 вида, а золотистые – 1 вид. Так же стоит отметить наличие неидентифицированных мелких жгутиковых форм.

Рисунок 1 – Район исследования Новороссийской бухты
Видовой состав фитопланктона Новороссийской бухты:
Отдел BACYLLARIOPHYTA
Achnanthes brevipes Ag.
Amphiprora alata Kutz.
Amphora inflexa Greg.
Amphora hyalina Kutz.
Asterionella gracialis Greg.
Bacillaria paradoxa Gmel.
Berkeleya scopulorum Cl.
Cerataulina pelagica (Cl.) Hendey
Chaetoceros affinis Laud.
Chaetoceros compressus Laud.
Chaetoceros curvisetus Cl.
Chaetoceros danicus Cl.
Chaetoceros holsaticus Cl.
Chaetoceros Lorencianus Cl.
Chaetoceros rigidus Brigh.
Chaetoceros peruvianus Brigh.
Chaetoceros similis Laud.
Chaetoceros simplex Cl.
Chaetoceros socialis Laud.
Chaetoceros sp.
Chaetoceros Wighamii Brigh.
Cocconeis scutellum Ehr.
Coscinodiscus sp.
Cyclotella caspia Grun.
Cylindrotheta closterium Raberh.
Dactyliosolen flagilissima Ehr.
Detonula confervacea (P.T. Cleve) Grun.
Ditylum brightwellii (T.West) Grun.
Leptocylindrus danicus Cl.
Leptocylindrus minima Cl.
Licmophora Ehrenbergii (Kutz.) Grun
Licmophora gracilis Cl.
Melosira moniliformis Ehr.
Nitzschia tenuirostris Mer.
Nitzschia holsatica Mer.
Navicula forcipata (W. Smith) Ralfs
Navicula pennata A.S.
Proboscia alata Grun.
Pzeudonitzschia delikatissima (Cl.) Heid.
Pzeudonitzschia seriata(Cl.) Heid.
Pzeudozosolenia calcar avis Schultze
Pleurosigma elongatum W. Smith
Sceletonema costatum (Grev.) Cl.
Striatella unipunctata (Grev.) Cl.
Synedra crystallina Grun.
Thalassionema nitzschioides Grun.
Thalassiosira antiqua Grun.
Thalassiosira excentrica (Ehr.) CI.
Thalassiosira sp.
Отдел DINOPHYTA (Динофитовые)
Ceratium furca (Ehr.) Clap. et. Lachm
Ceratium fusus (Ehr.) Dujardin
Ceratium tripos (O. F. Muller) Nitzsch
Dinophysis acuminata Cl. et. Lachm.
Dinophysis acuta Ehr.
Dinophysis caudata Saville-Kent
Dinophysis sacculus Stein
Diplopsalis lenticula Ehr.
Glenodinium caspicum Pauls.
Glenodinium danicum Pauls.
Glenodinium pilula (Ostf.) Schill
Glenodinium rotundum Schil.
Glenodinium sp.
Gonyaulax digitale Lind.
Gonyaulax minima Marzen.
Gonyaulax spinifera (Clap. et. Lachm) Diesing
Gymnodinium blax Harris
Gymnodinium simplex (Lohm.) Kof. et. Sw.
Gymnodynium sp.
Gymnodynium wulffii Kof. et. Sw.
Gyrodinium fusiforme Kof. et. Sw.
Gyrodinium spirale (Bergh) Kof. et. Sw.
Lingolodinium polyedrum Ehr.
Peridinium guinguecorne (Ehr.) Stein
Prorocentrum compressum (Bail.) Abe ex Dodge
Prorocentrum lima (Ehr.) Stein
Prorocentrum micans Ehr.
Prorocentrum minimum (Pav.) Schill
Protoperidinium breve (Pauls.) Balech
Protoperidinium conicum (Gran.) Balech
Protoperidinium depressum (Bailey) Balech
Protoperidinium divergens (Ehr.) Balech
Protoperidinium globulus (Kof.) Balech
Protoperidinium granii (Ostf.) Balech
Protoperidinium pellucidum (Jorg.) Balech
Protoperidinium steinii (Jorg.) Balech
Protoperidinium subinerme (Jorg.) Balech
Protoperidinium sp.
Heterocapsa triguetra (Ostf.) Balech
Scrippsiella trochoidea (Stein) Balech
Отдел EUGLENOPHYTA (Эвгленовые)
Euglena sp.
Eutrepcia lanouwii Steuer.
Eutrepcia sp.
Отдел CHRYSOPHYTA (Золотистые)
Emiliania huxleyi (Lohm.) Hay et Mohler
Отдел CYANOPHYTA (Сине-зелёные)
Oscillatoria tenuis C. Agardh
Oscillatoria sp.
Таксономический анализ фитопланктона показал, что диатомовые водоросли, занимающие доминирующее положение, составляют 51,6 % от общего числа видов, динофитовые – 42,1 %, тогда как на долю видов из других отделов приходится 6,3 % (рисунок 2).
Рисунок 2 – Количественный состав фитопланктона (%)
Таблица 1 – Количество видов планктонных водорослей в Новороссийской бухте
|
Порт
|
Западный мол
|
Мыс Любви
|
Южная Озереевка
|
|
49
|
51
|
55
|
54
|
Каждая из исследуемых станций обладает богатым видовым составом, который колеблется в незначительных пределах. Наименьшее число видов планктонных водорослей встречается в районе порта и равняется 49, что отражает близость источника загрязнения. Наибольшее число видов было отмечено в открытой части бухты и на мысе Любви и составило 54 и 55 соответсвенно (таблица 1).
4.2 Сезонная динамика биомассы и количественного состава фитопланктона
Сезонная динамика биомассы и количественного состава планктонных водорослей бухты изучалась в весенний, летний и осениий периоды. Численность и биомасса фитопланктона во время исследования варьировала в значительных пределах.
По вкладу отдельных групп водорослей в общую численность и биомассу фитопланктон бухты можно отнести к диатомово-динофитовым (от 41 до 100 % от суммарного числа клеток), так как эти группы водорослей здесь разнообразны флористически и количественно преобладают.
Высокое биоразнообразие отмечается у диатомовых водорослей, в связи с их широкой эвритермностью и эвригалинностью. В течение двадцати дней качественный состав диатомовых водорослей менялся от 5 до 15 видов.
Среди них есть виды, которые практически постоянно присутствуют в составе планктона (Chaetoceros curvisetus, Sceletonema costatum, Cerataulina pelagica, Pzeudosolenia calcar avis), причём один из них – Sceletonema costatum является доминирующим видом на протяжении всего периода исследований, а другой – Thalassionema nitzschoides играет роль субдоминанта.
Наиболее богаты флористически роды Chaetoceros, Leptocylindrys, Licmophora, Pzeudonitzschia, Nitzschia. В составе сообщества также присутствуют литоральные и бентоcно-планктонные формы, такие как Cylindrotheta closterium, Licmophora Ehrenbergii и др.
Видовой состав динофитовых разнообразен, так как основная группа этих микроводорослей развивается в тёплое время года. В составе динофитового комплекса присутствуют как мелкие мезосапробные виды, характерные для эфтрофированных районов побережья, так и крупноразмерные формы, предпочитающие районы открытого побережья. Наиболее богаты роды Dinophysis, Protoperidinium, Glenodinium, Gonyaulax, Gymnodynium, Prorocentrum. Из них Prorocentrum micans и Protoperidinium brevipes являются практически постоянными в течение всего периода исследований, тогда как остальные виды присутствуют эпизодически.
Эвгленовые водоросли, представленные двумя видами Eutrepcia, постоянно отмечены в составе сообщества и их развитие приурочено к районам порта и Западного мола, что дает основание предполагать о присутствии органического загрязнения.
В середине весны наблюдалась активная вегетация мелкоразмерной колониальной диатомеи Leptoсylindrus danicus. В порту зафиксированы максимальные показатели её плотности (254,1 млн.кл/м3). На станциях вблизи Западного мола и мыса Любви численность клеток сократилась почти в 2 раза, но, тем не менее, оставалась довольно значительной. В районе Южной Озереевки эти показатели снизились более чем в 6,5 раза. Здесь к Leptoсylindrus danicus присоединилась Pzeudonitzschia delikatissima. Сопутствующим видом, типичным для данной акватории, являлась Sceletonema costatum. Максимальные значения этого вида приурочены к портовой части бухты, далее значения постепенно снижаются и в открытой части бухты показатели обилия минимальны. Основной вклад в показатели биомассы вносила крупная диатомея Cerataulina pelagica (62, 6%). На станциях, расположенных в самой бухте, видоразнообразие формировалось за счет диатомовой группы. На мысе Любви количество диатомовых видов возрастает до 15 и включает в себя бентосно-планктонные и литоральные формы (Bacillaria paradoxa, Berkeleya scopilorum, Cylindrotheta closterium, Licmophora gracilis, L. Ehrenbergii, Nitzschia tenuirostris, Synedra crystallina). В портовой части бухты при доминировании диатомей фиксировалось значительное количество мелких видов динофитового комплекса (Gymnodinium simplex, Gymnodinium sp.). В прибрежье посёлка Южная Озереевка основной вклад в видоразнообразие, напротив, вносили динофлагелляты (14 видов). В открытых водах лидирующими выступали виды из рода Prorocentrum (Prorocentrum minimum, P. compressum и P. micans). Сине-зелёные и эвгленовые водоросли в апреле зарегистрированы лишь в самой бухте, на открытом прибрежье эти группы микродорослей не входили в состав планктонного альгоценоза.
В мае активность ранее доминирующего вида (Leptoсylindrus danicus) заметно снизилась, что отразилась на суммарных показателях обилия фитопланктона. Динофитовые формы на всех трёх станциях в бухте были малочисленны. В основном в состав этой группы входили виды, отмеченные в апреле, но с более низкими количественными характеристиками. Относительно высокие показатели численности и биомассы обеспечивала крупная форма диатомовых – Cerataulina pelagica. На станции у Западного мола она входила в доминирующую группу. Здесь же активно развивались и другие диатомеи –Sceletonema costatum и Thalassionema nitzschoides. В самом порту зафиксировано активное развитие другой мелкоразмерной диатомовой формы – Pzeudonitzschia delicatissima (4,7 млн. кл/м3).
В июне планктонный альгоценоз находился в депрессионном состоянии, что, по-видимому, связано с повышением температуры воды и явными признаками усиления антропогенной нагрузки, выражающейся в обеднении таксономического состава микроводорослей и крайне низкими показателями биомассы при значительной численности, которую формировала Sceletonema costatum. В этот период в порту отмечено всего 13 видов водорослей, с высокой долей мелких колониальных форм, а также 2 вида эвгленовых – индикаторов хронического органического загрязнения. На станции у Западного мола видоразнообразие несколько увеличилось (до 18 видов). Численность основного доминирующего вида снизилась, и возросло количество динофитовых видов (8 видов) с высокой долей мелких миксотрофных форм Gymnodinium sp. У мыса Любви диатомовая Sceletonema costatum сохранила свой доминирующий статус, но численность её сократилась более чем в 2,5 раза. В планктоне присутствовали немногочисленные колонии сине-зелёных водорослей. Субдоминирующим видом, зафиксированным на всех станциях в этот временной период являлась мелкая кокколитина Emiliania huxleyi, развитие которой наиболее активно отмечено для станции у мыса Любви (28,1 млн. кл/м3). Здесь же развивались летние формы диатомовых микроводорослей (Chaetoseros curvisetus, C. Lorencianus, C. similis, C. simplex).
В июле численность Sceletonema costatum снизилась, хотя на станциях в акватории бухты она продолжала играть лидирующую роль. В порту развивались виды из рода Chaetoseros, а так же вид Cyclotella caspia. Видовой состав динофитовых обеднён и основной вклад в их численность и биомассу вносил Protoperidinium breve. На станции у Западного мола снижаются как качественные, так и количественные характеристики развития планктонного фитоценоза, а далее у мыса Любви они падают до минимальных показателей. В открытой части бухты, напротив, фитопланктон демонстрирует богатое видовое разнообразие (31 вид). Здесь не замечено резко доминирующей формы в составе растительного планктона. Из диатомовых наиболее активно вегетировали Pzeudonitzschia delicatissima и Thalassionema nitzschoides. Из динофитовой группы следует отметить Prorocentrum minimum и Scrippsiella trochoidea. Развитие отмечено и для крупных летних форм динофитовых водорослей, вносивших заметный вклад в биоразнообразие фитопланктона открытых вод (Protoperidinium depressum, P. granii, P. globulus, P. steinii, Heterocapsa triguetra).
В августе видовое разнообразие микроводорослей в бухте оставалось на невысоком уровне. В порту и у Западного мола численность формировали несколько видов из диатомовой и динофитовой групп (Cyclotella caspia, Sceletonema costatum, Chaetoseros rigidus, Gymnodinium sp., Prorocentrum micans), а также эвгленовые и мелкие жгутиковые клетки. На станции у Западного мола отмечено присутствие редкой диатомовой формы Asterionella gracialis. Этот неритический эвригалинный тропическо-бореально-арктический вид предпочитает развиваться в открытых водах и при более низких температурах. Чаще его развитие приходится на октябрь-ноябрь месяцы. В Новороссийской бухте накануне наблюдался умеренный нагонный ветер юго-восточного направления с элементами перемешивания водных масс и выносом придонных вод на поверхность, что, по-видимому, и зафиксировано присутствием данной водоросли в составе планктонного сообщества. Фитопланктон у мыса Любви, в основном, представляли эпифитные формы с лидирующим видом Cylindrotheta closterium. Активно развивались виды из рода Licmophora, а также Bacillaria paradoxa и Navicula pennata. Динофитовые отмечены здесь активностью Diplopsalis lenticula и Protoperidinium steinii. Количество жгутиковых клеток заметно снизилось, а эвгленовые отмечены единично. В составе сообщество присутствовали золотистые водоросли. На станции в районе Южной Озереевки флористическое разнообразие несколько снизилось, по сравнению с предыдущим, но, тем не менее, сообщество оставалось на высоком уровне с доминированием в его составе истинно летних, теплолюбивых форм динофитовой группы. В составе этой группы заметную роль играли виды из родов Ceratium, Gonyaulax, Prorocentrum и Protoperidinium. Лидирующими видами на этой станции были Thalassionema nitzschoides и Sceletonema costatum. В состав первых трёх видов в ранжированном по обилию ряду так же входила мелкоразмерная колониальная диатомея Pzeudonitzschia delicatissima.
Таблица 2 – Показатели численности фитопланктона Новороссийской бухты в весенний, летний и осенний периоды (млн. кл/м3)
|
Дата
|
Район
|
Отделы
|
Всего
|
|
|
|
|
Диато-мовые
|
Дино-фитовые
|
Эвгле-новые
|
Сине-зеленые
|
Золо-тистые
|
Мелкие
жгути-ковые
|
|
|
Апрель
|
П
|
248,0
|
2,9
|
0,4
|
1,0
|
-
|
-
|
252,3
|
|
|
З
|
152,4
|
5,5
|
-
|
0,8
|
-
|
-
|
158,7
|
|
|
М
|
167,4
|
8,6
|
0,1
|
1,4
|
-
|
-
|
177,5
|
|
|
Ю
|
74,0
|
11,0
|
-
|
-
|
-
|
-
|
85,0
|
|
Май
|
П
|
13,4
|
1,0
|
0,4
|
0,5
|
-
|
-
|
15,3
|
|
|
З
|
20,4
|
2,1
|
0,6
|
0,3
|
-
|
-
|
23,4
|
|
|
М
|
13,9
|
2,3
|
-
|
-
|
-
|
-
|
16,2
|
|
Июнь
|
П
|
164,6
|
1,6
|
0,9
|
-
|
15,6
|
-
|
182,7
|
|
|
З
|
114,4
|
3,5
|
1,9
|
-
|
21,9
|
-
|
141,7
|
|
|
М
|
78,1
|
0,5
|
0,1
|
4,9
|
28,1
|
-
|
111,7
|
|
Июль
|
П
|
30,1
|
3,0
|
0,5
|
-
|
-
|
-
|
33,6
|
|
|
З
|
13,8
|
1,1
|
0,3
|
-
|
-
|
-
|
15,2
|
|
|
М
|
22,0
|
0,3
|
-
|
-
|
-
|
-
|
22,3
|
|
|
Ю
|
18,6
|
8,9
|
-
|
2,6
|
-
|
-
|
30,1
|
|
Август
|
П
|
6,6
|
3,9
|
1,0
|
1,3
|
-
|
3,9
|
16,7
|
|
|
З
|
11,6
|
4,2
|
3,9
|
-
|
0,9
|
4,2
|
24,8
|
|
|
М
|
9,8
|
4,6
|
0,1
|
0,8
|
0,8
|
1,2
|
17,3
|
|
|
Ю
|
8,9
|
3,6
|
-
|
-
|
3,1
|
-
|
15,6
|
|
Сентябрь
|
П
|
31,1
|
-
|
-
|
-
|
-
|
1,8
|
32,9
|
|
|
З
|
17,0
|
0,5
|
-
|
-
|
-
|
0,5
|
18,0
|
|
|
М
|
16,8
|
0,6
|
-
|
-
|
-
|
0,3
|
17,7
|
|
|
Ю
|
15,8
|
1,1
|
-
|
-
|
-
|
-
|
16,9
|
|
Примечания
1 П – Порт
2 З – Западный мол
3 М – мыс Любви
4 Ю – Южная Озереевка
|
В сентябре показатели численности фитопланктона по-прежнему оставались на невысоком уровне. Кроме диатомовых и динофитовых водорослей представители из других отделов не были обнаружены.
Таблица 3 – Показатели биомассы фитопланктона Новороссийской бухты в весенний, летний и осенний периоды (мг/м3)
|
Дата
|
Район
|
Отделы
|
Всего
|
|
|
|
|
Диато-мовые
|
Дино-фитовые
|
Эвгле-новые
|
Сине-зеленые
|
Золо-тистые
|
Мелкие
жгути-
ковые
|
|
|
Апрель
|
П
|
211,73
|
19,79
|
0,16
|
0,36
|
-
|
-
|
232,04
|
|
|
З
|
168,26
|
22,62
|
-
|
0,27
|
-
|
-
|
191,15
|
|
|
М
|
60,92
|
35,29
|
0,05
|
0,50
|
-
|
-
|
96,76
|
|
|
Ю
|
66,06
|
149,11
|
-
|
-
|
-
|
-
|
215,17
|
|
Май
|
П
|
108,60
|
11,81
|
0,16
|
0,18
|
-
|
-
|
120,75
|
|
|
З
|
338,48
|
21,78
|
0,27
|
0,09
|
-
|
-
|
360,62
|
|
|
М
|
151,08
|
37,62
|
-
|
-
|
-
|
-
|
188,70
|
|
Июнь
|
П
|
91,15
|
4,94
|
0,59
|
-
|
6,28
|
-
|
102,96
|
|
|
З
|
50,40
|
14,38
|
1,43
|
-
|
8,79
|
-
|
55,76
|
|
|
М
|
64,06
|
1,75
|
0,09
|
0,17
|
11,30
|
-
|
77,37
|
|
Июль
|
П
|
41,56
|
104,18
|
0,22
|
-
|
-
|
-
|
145,96
|
|
|
З
|
45,33
|
25,18
|
1,09
|
-
|
-
|
-
|
71,60
|
|
|
М
|
80,83
|
0,26
|
-
|
-
|
-
|
-
|
81,09
|
|
|
Ю
|
197,58
|
81,69
|
-
|
0,95
|
-
|
-
|
280,22
|
|
Август
|
П
|
26,98
|
34,20
|
0,44
|
0,09
|
-
|
0,05
|
61,76
|
|
|
З
|
50,14
|
46,47
|
1,69
|
-
|
0,35
|
0,06
|
98,71
|
|
|
М
|
100,59
|
96,08
|
0,05
|
0,03
|
0,30
|
0,02
|
197,07
|
|
|
Ю
|
136,99
|
148,08
|
-
|
-
|
1,26
|
-
|
286,33
|
|
Сентябрь
|
П
|
321,16
|
-
|
-
|
-
|
-
|
0,02
|
321,18
|
|
|
З
|
193,44
|
12,88
|
-
|
-
|
-
|
0,01
|
206,33
|
|
|
М
|
175,32
|
7,15
|
-
|
-
|
-
|
0,00
|
182,47
|
|
|
Ю
|
203,38
|
18,19
|
-
|
-
|
-
|
-
|
221,57
|
|
Примечания
1 П – Порт
2 З – Западный мол
3 М – мыс Любви
4 Ю – Южная Озереевка
|
Стоит отметить в осеннем сезоне наличие мелких жгутиковых на всех исследуемых станциях за исключением открытой части бухты. Средние показатели биомассы в этом месяце намного превышают показатели августа, в основном за счет таких диатомовых водорослей как Cerataulina pelagica, Pzeudosolenia calcar avis и Thalassionema nitzschioides.
В порту в мае показатели численности были равны 15,5 млн. кл/м3, а в апреле достигали 252,3 млн. кл/м3, что является соответственно наименьшим и наибольшим показателями численности в весенний сезон (таблица 2).
В летний период наименьшие показатели численности были отмечены в июле на станции Западный мол и составили 15,2 млн. кл/м3, наибольшие – в июне на станции порт и достигали 182,7 млн. кл/м3.
В осенний период наименьшие показатели численности были зафиксированы в открытой части бухты и составили 16,9 млн. кл/м3, наибольшие – в порту и достигали 32,9 млн. кл/м3.
Показатели биомассы на мысе Любви в апреле были равны 96,76 мг/м3, на Западном моле в мае – 360,62 мг/м3, что является соответственно наименьшим и наибольшим показателями биомассы в весенний период (таблица 3).
В летний период наименьшие показатели биомассы были отмечены в июле на станции Западный мол и составили 55,76 мг/м3, наибольшие – в августе в открытой части бухты и достигали 286,33 мг/м3.
В осенний период наименьшие показатели биомассы были зафиксированы на мысе Любви и составили 182,47 мг/м3, наибольшие – в порту и достигали 321,18 мг/м3.
Сезонная динамика численности фитопланктона в Новороссийской бухте представлена на рисунке 3. Из представленной диаграммы следует, что пик численности планктонных водорослей приходится в весенний период, так как именно в этот сезон были зафиксированы самые высокие показатели. Максимальный показатель был отмечен в порту и составил 133,8 млн. кл/м3. В летний сезон показатели численности планктонных водорослей значительно снижаются и продолжают падать в осенний период. На протяжении последних двух сезонов самые высокие показатели по-прежнему были отмечены на станции порт.
Рисунок 3 – Сезонная динамика численности фитопланктона (млн. кл/м3)
Сезонная динамика биомассы фитопланктона в Новороссийской бухте представлена на рисунке 4. Пик биомассы планктонных водорослей приходится на осенний сезон, так как здесь зафиксированы довольно высокие показатели. Наибольший из них был отмечен в порту и составил 321,2 мг/м3. Наименьшими показателями биомассы фитопланктона характеризуется летний сезон. Исключение составляется Южная Озереевка: на этой станции был отмечен второй из наибольших показателей (283,3 мг/м3).
Рисунок 4 – Сезонная динамика биомассы фитопланктона (мг/м3)
4.3 Кислотность и уровень загрязнения вод Новороссийской бухты
Анализ на определение водородного показателя (pH) воды в местах отбора проб показал, что значение pH в акватории Новороссийской бухты варьирует от 7,6 до 8,1. Воды относятся к слабощелочным (таблица 4).
Таблица 4 – Водородный показатель (pH) воды
|
Район (место отбора проб)
|
Значение pH
|
|
Порт
|
8,1
|
|
Западный мол
|
8,1
|
|
Мыс Любви
|
8,0
|
|
Южная Озереевка
|
7,6
|
Анализы, проведённые на количественное определение в воде нитратов, хлоридов и сульфатов, дали следующие результаты. Количественные показатели нитратов в воде во всех районах исследования не превышают нормы ПДК (45 мг/л). Такая же благоприятная обстановка сложилась и с содержанием в воде сульфатов: ни на одной из станций их количественные показатели не превышают ПДК (500 мг/л).
Таблица 5 – Показатели количественного содержания в воде нитратов, хлоридов и сульфатов (мг/л)
|
Район
|
Нитраты
|
Хлориды
|
Сульфаты
|
|
Порт
|
20
|
710
|
53
|
|
Западный мол
|
10
|
444
|
50
|
|
Мыс Любви
|
5
|
431
|
47
|
|
Южная Озереевка
|
5
|
178
|
47
|
|
ПДК
|
45
|
350
|
500
|
Однако количественные показатели хлоридов в воде превышают нормы ПДК (350 мг/л) на трёх станциях: в порту – более чем в 2 раза, на Западном моле – в 1,3 раза, на мысе Любви – в 1,2 раза (таблица 5).
4.4 Влияние антропогенной нагрузки на фитопланктон Новороссийской бухты
Водоросли фитопланктона являются индикаторами экологического состояния морской среды. Они чётко отображают изменения экологического состояния района их обитания, произошедшие за определённый промежуток времени. Показателями качества воды являются: количество видов водорослей-индикаторов загрязнения, их численность и биомасса.
Рисунок 5 – Средние показатели численности фитопланктона
на исследуемых станциях (млн. кл/м3)
Наименьшие средние показатели численности планктонных водорослей были зафиксированы в открытой части бухты и составили 36,9 млн. кл/м3, наибольшие – в порту и были равны 88,9 млн. кл/м3. Такой высокий показатель был достигнут за счет мелкоразмерной диатомеи Sceletonema costatum, встречающейся здесь в значительном количестве (рисунок 5).
Рисунок 6 – Средние показатели биомассы фитопланктона
на исследуемых станциях (мг/м3)
Наименьшие средние показатели биомассы фитопланктона в Новороссийской бухте были отмечены на мысе Любви и составили 137,24 мг/м3, наибольшие – в Южной Озереевке и были равны 249,29 мг/м3. В основном, этот высокий показатель был достигнут за счёт наличия в водах бухты двух крупноразмерных водорослей Ceratium tripos и Cerataulina pelagica (рисунок 6).
В портовой части бухты встречается большое количество динофлагеллят мелких и крупных размеров (Glenodinium danicum, Glenodinium pilula, Gymnodinium sp., Prorocentrum minimum, Prorocentrum micans), характерных для загрязненных районов, испытывающих постоянный антропогенный пресс. Важную роль в этой группе играют Prorocentrum minimum и P. micans, являющиеся активными -мезосапробами. Здесь же отмечено присутствие эвгленовой водоросли Eutrepcia lanouwii, которая является видом-возбудителем «цветения» воды и служит индикатором сапробности водоёма, поскольку часто развивается вблизи сброса сточных вод и в местах активного разложения органического вещества.
В районе Западного мола в составе планктонного сообщества, так же как и в портовой акватории, отмечено значительное количество видов-индикаторов органического загрязнения (Cerataulina pelagica, Pzeudozosolenia calcar avis, Thalassionema nitzschioides, Glenodinium lenticula, Glenodinium pilula, Gonyaulax spinifera, Gymnodynium sp., Prorocentrum micans, Prorocentrum minimum, Eutrepcia lanouwii).
Станция, расположенная на мысе Любви, находится на значительном удалении от района порта и Западного мола. Видовой состав схож с ранее описанными станциями. Следует отметить активное развитие диатомового комплекса и, в первую очередь, вида Pzeudozosolenia calcar avis. Список видов-индикаторов схож с ранее перечисленным, но в количественном отношении развитие этих видов несколько снизилось.
На фоновой станции, расположенной в открытой части Новороссийской бухты, планктонный альгоценоз значительно отличался по своему составу и показателям обилия. Доминирующее положение занимали крупноразмерные формы (Cerataulina pelagica, Pzeudozosolenia calcar avis, Ceratium furca, Ceratium tripos, Protoperidinuim depressum), развитие которых, в большинстве своём, приурочено к относительно чистым водам.
ЗАКЛЮЧЕНИЕ
По результатам данной работы сделаны следующие выводы:
1. Видовой состав фитопланктона Новороссийской бухты включает в себя 93 вида, относящихся к 5 систематическим отделам. Наибольшее число видов насчитывают отделы Bacyllariophyta (49 видов) и Dinophyta (38 видов), 3 вида относятся к отделу Euglenophyta, 2 вида – к отделу Cyanophyta, 1 вид – к отделу Chrysophyta. Диатомовые водоросли составляют 52,7 % от общего числа видов, динофитовые – 40,8 %, на долю видов из других отделов приходится 6,5 %.
2. В порту в мае показатели численности были равны 15,5 млн. кл/м3, а в апреле достигали 252,3 млн. кл/м3, что является соответственно наименьшим и наибольшим показателями численности в весенний сезон. В летний период наименьшие показатели численности были отмечены в июле на станции Западный мол и составили 15,2 млн. кл/м3, наибольшие – в июне на станции порт и достигали 182,7 млн. кл/м3. В осенний период наименьшие показатели численности были зафиксированы в открытой части бухты и составили 16,9 млн. кл/м3, наибольшие – в порту и достигали 32,9 млн. кл/м3.
Показатели биомассы на мысе Любви в апреле были равны 96,76 мг/м3, на Западном моле в мае – 360,62 мг/м3, что является соответственно наименьшим и наибольшим показателями биомассы в весенний период. В летний период наименьшие показатели биомассы были отмечены в июле на станции Западный мол и составили 55,76 мг/м3, наибольшие – в августе в открытой части бухты и достигали 286,33 мг/м3. В осенний период наименьшие показатели биомассы были зафиксированы на мысе Любви и составили 182,47 мг/м3, наибольшие – в порту и достигали 321,18 мг/м3.
3. Анализ на определение водородного показателя (pH) воды в местах отбора проб показал, что значение pH в акватории Новороссийской бухты варьирует от 7,6 до 8,1. Воды относятся к слабощелочным. При помощи анализов, проведённых на количественное определение в воде нитратов, хлоридов и сульфатов, было выяснено, что количественные показатели нитратов и сульфатов в воде во всех районах исследования не превышают нормы ПДК. Однако количественные показатели хлоридов в воде превышают нормы ПДК на трёх станциях, в порту – более чем в 2 раза.
4. В районе порта и Западного мола отмечено значительное количество видов-индикаторов (Glenodinium danicum, Gymnodynium blax, Prorocentrum minimum, Protoperidium brevives и др.) органического загрязнения, что свидетельствует о значительной антропогенной нагрузке на данную акваторию. На мысе Любви число видов-индикаторов снизилось, тем не менее, их присутствие в этом районе обусловлено близостью источника органического загрязнения. В открытой части бухты (Южная Озереевка) количество индикаторных видов минимально, сообщество стабильно и при невысоких количественных характеристиках обладало высоким показателем биомассы и богатым видовым составом.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
1. Агроклиматический справочник по Краснодарскому краю / под ред. В.П. Гаврилова. Краснодар, 1961. 367 с.
2. Аннотированный каталог водорослей / под ред. Ю.Е. Петрова. М.,1998. 224 с.
3. Болгова Л.В., Селифонова Ж.П., Плайпа А.С. Функционирование экосистемы Новороссийской бухты в условиях растущего техногенного пресса. Ростов н/Д, 2004. 144 с.
4. Болгова Л.В., Луговая И.М. Фитопланктон акватории порта Новороссийской бухты как показатель состояния экосистемы // Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий: матер. ХХIII Межресп. науч.-практ. конф. с Междунар. участием. Краснодар, 2010. С. 33–35.
5. Брянцева Ю.В., Курилов А.В. Расчет объёмов клеток микроводорослей и планктонных инфузорий Чёрного моря. Севастополь, 2003. 20 с.
6. Вершинин А.О. Потенциально токсичные водоросли в составе прибрежного фитопланктона северо-восточной части Чёрного моря // Океанология. М., 2005. Т. 45. № 2. С. 240–246.
7. Вершинин А.О., Моргунов А.А. Потенциально-токсичные водоросли в прибрежном фитопланктоне северо-восточной части Чёрного моря // Экология моря. М., 2003. Вып. 64. С. 45–49.
8. Виноградов М.Е., Шукшина Э.А., Сапожников В.В. Экосистема Чёрного моря. М., 2002. 224 с.
9. Виноградова Л.А., Василёва В.Н. Многолетняя динамика и моделирование состояния экосистем прибрежных вод северо-западной части Чёрного моря. Санкт-Петербург: Гидрометеоиздат, 1992. 106 с.
10. Восконьян А.В., Восконьян В.Г. Помочь Чёрному морю // Успехи современного естествознания. М., 2007. № 12. С. 91–93.
11. Георгиева Л.В., Сеничкина Л.Г. Фитопланктон Чёрного моря: Современное состояние и перспективы исследований // Экология моря. М., 1996. Вып. 45. С. 6–12.
12. Диатомовые водоросли СССР / под. ред. А.И. Прошкиной-Лавренко. Л., 1974. 246 с.
13. Зернов С.А. Общая гидрология. М., 1934. 349 с.
14. Иванов М.Б. Зоопланктон и фитопланктон. М., 1982. 342 с.
15. Иванов А.И. Характеристика качественного состава фитопланктона Чёрного моря // Исследования планктона Чёрного и Азовского морей. Киев: Наук. думка, 1965. С. 17–35.
16. Киселёв И.А. Панцирные жгутиконосцы морей и пресных вод СССР. М.: Наука, 1950. 280 с.
17. Киселёв И.А. Планктон морей и континентальных водоёмов. М.: Наука, 1969. Т. 1. 439 с.
18. Нестерова Д.А. Районирование северо-западной части Чёрного моря по составу фитопланктона // Экология моря. М., 2001. Вып. 55. С. 22–23.
19. Михайловская Н.А. Панцирные жгутиконосцы. М., 1903. 280 с.
20. Морозова-Водяницкая Н.В. Фитоплантон в Чёрном море и его количественное развитие // Тр. Севаст. биол. станции. М., 2004. С. 11–99.
21. Одум Ю. Основы экологии. М., 1975. 54 с.
22. Определитель низших растений. Водоросли / под общ. ред. Л.И.Курсанова. М.: Советская наука, 1953. 396 с.
23. Прошкина-Лавренко А. И. Диатомовые водоросли планктона Чёрного моря. М., 1955. 270 с.
24. Рейдгард З.Л. Фитопланктон Чёрного моря, Керченского пролива, Босфора и Мраморного моря // Тр. об-ва испыт. прир. Астрахань, 1983. С. 1–8.
25. Селифонова Ж.П., Ясакова О.Н. Фитопланктон акваторий портовых городов северо-восточного шельфа Чёрного моря // Морской биологический журнал. Мурманск, 2012. Т. 11. № 4. С. 67–77.
26. Сеничкина Л.Г. Фитопланктон северо-западной части Чёрного моря в зимний период // Сезонные изменения черноморского планктона. М., 1983. С. 55–65.
27. Сеничкина Л.Г. Численность и видовой состав фитопланктона Новороссийской бухты // Геоэкологические исследования и охрана недр: науч.-практ. информ. сб. М., 2002. Вып. 2. С. 50–57.
28. Сикачинский Г.И. Край дальний и наш: Век Океана. М., 1989. 320 с.
29. Сорокин Ю.И. Чёрное море. М., 1982. 256 с.
30. Сорокин Ю.И. Биологические процессы. М., 1996. 342 с.
31. Суханова И.Н. Современные методы количественной оценки распределения морского планктона. М., 1983. 232 с.
32. Теренько Г.В. Видовое разнообразие фитопланктона Чёрного моря как характеристи�ка состояния экосистемы // Экология моря. М, 2001. Вып. 58. С. 53–56.
33. Усачёв П.И. К методике планктонных исследований // Дневник всесоюзного съезда ботаников. М., 1928. С.174–175.
34. Фёдоров В.Д. О методах изучения фитопланктона и его активности. М.: Изд-во МГУ, 1979. № 2. С. 71–76.
35. Фёдоров В.Д., Радченко И.Г., Капков В.И. Практическое руководство по сбору и анализу проб морского фитопланктона. М., 2010. 60 с.
36. Ясакова О.Н., Селифонова Ж.П., Ермакова Е.П. Современное состояние пелагической экосистемы Новороссийской бухты // Экология моря. М, 2000. Вып. 52. С. 22–25.
37. Ясакова О.Н. Фитопланктон Новороссийского порта в условиях антропогенного воздействия // Проблемы экологии Азово-Чёрноморского бассейна: современное состояние и прогноз. Понт Эвксинский II: тез. докл. конф. молодых учёных. Севастополь, 2001. С. 60–68.
38. Ясакова О.Н. Развитие фитопланктона в Новороссийской бухте весной 2004 г. // Проблемы репродукции и раннего онтогенеза морских гидробионтов: тез. докл. Междунар. науч. симпозиума. Мурманск, 2004. С. 151–153.
39. Ясакова О.Н. Состояние фитопланктона в Новороссийской бухте в весенний период 2004 года // Матер. XXIII конф. молодых учёных, посвящённой 70-летию МБС – ММБИ. Апатиты, 2005. С. 152–157.
40. Ясакова О.Н. Развитие фитопланктона в Новороссийской бухте Чёрного моря в условиях антропогенного эвтрофирования вод // Материалы II ежегодной науч. конф. студентов и спирантов базовых кафедр Южного научного центра РАН. Ростов н/Д, 2006. С. 49–50.
41. Ясакова О.Н. Многолетние исследования фитопланктона Новороссийской бухты Чёрного моря // Тепловодная аквакультура и биологическая продуктивность водоёмов аридного климата: тез. докл. Междунар. симпозиума. Астрахань, 2007а. С. 3–17.
42. Ясакова О.Н. Токсичные и инвазионные виды в составе фитопланктона Новороссийской бухты // Естественные и инвазионные процессы формирования биоразнообразия водных и наземных экосистем: тез. докл. Междунар. науч. конф. Ростов н/Д, 2007б. С. 345–346.
43. Ясакова О.Н. Воздействие антропогенного эвтрофирования на состояние планктонного альгоценоза Новороссийской бухты // Морские прибрежные экосистемы. Водоросли, беспозвоночные и продукты их переработки: тез. докл. III Междунар. науч.-практ. конф. Ростов н/Д, 2008а. 442 с.
44. Ясакова О.Н. «Красные приливы» и связанные с ними проблемы развития потенциально-токсичных водорослей в акватории Новороссийской бухты // Современные проблемы альгологии: матер. Междунар. науч. конф. и VII школы по морской биологии. Ростов н/Д, 2008б. С. 395–397.
45. Ясакова О.Н. Фитопланктон акваторий курортных городов северо-восточного побережья Чёрного моря // Экологическая безопасность приморских регионов: порты, берегозащита, рекреация, марикультура: матер. Междунар. науч. конф., посвящённой 150-летию Н.М. Книповича. Мурманск, 2012. С. 288–291.
ПРИЛОЖЕНИЕ А
Показатели численности и биомассы планктонных водорослей
Новороссийской бухты
|
Виды
|
Размер клетки,
мкм
|
Объём клетки,
мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
2750
|
145,71
|
|
Chaetoceros compressus
|
22*28*8
|
3868
|
250
|
0,97
|
|
Cocconeis scutellum
|
48*22*15
|
4145
|
125
|
0,52
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
125
|
0,01
|
|
Leptocylindrus danicus
|
4*4*10
|
128
|
234375
|
29,438
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
1250
|
0,18
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
250
|
33,33
|
|
Sceletonema costatum
|
12*6*3
|
170
|
7125
|
1,21
|
|
Thalassionema nitzschioides
|
65*2*2
|
201
|
1750
|
0,36
|
|
Итого
|
|
|
248000
|
211,73
|
|
Dinophyta:
|
|
|
|
|
|
Glenodinium danicum
|
30*31*15
|
7301
|
250
|
1,83
|
|
Gymnodynium sp.
|
30*31*15
|
7301
|
500
|
3,65
|
|
Gymnodynium simplex
|
11*4*4
|
92
|
1750
|
0,16
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
125
|
0,30
|
|
Peridinium guinguecorne
|
52*34*18
|
31824
|
125
|
4,37
|
|
Protoperidinium subinerme
|
54*52*27
|
75816
|
125
|
9,48
|
|
Итого
|
|
|
2875
|
19,79
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
375
|
0,16
|
|
Cyanophyta sp.
|
60*3*3
|
377
|
1000
|
0,36
|
|
Всего:
|
|
|
252250
|
232,04
|
Таблица А.1 – Численность и биомасса фитопланктона на станции порт в апреле 2014 г.
Таблица А.2 – Численность и биомасса фитопланктона на станции Западный мол в апреле 2014 г.
|
Виды
|
Размер
клетки, мкм
|
Объём
клетки,
мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
375
|
0,46
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
2750
|
145,71
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
1000
|
2,75
|
|
Cylindrotheta closterium
|
52*3*2
|
88
|
125
|
0,01
|
|
Leptocylindrus danicus
|
4*4*10
|
128
|
137500
|
17,27
|
|
Navicula pennata
|
38*5*4
|
597
|
250
|
0,15
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
875
|
0,13
|
|
Sceletonema costatum
|
12*6*3
|
170
|
4500
|
0,76
|
|
Thalassionema nitzschioides
|
65*2*2
|
201
|
5000
|
1,02
|
|
Итого
|
|
|
152375
|
168,26
|
|
Dinophyta:
|
|
|
|
|
|
Glenodinium danicum
|
30*31*15
|
7301
|
1625
|
11,86
|
|
Glenodinium pilula
|
24*24*12
|
3617
|
875
|
3,17
|
|
Gymnodynium simplex
|
11*4*4
|
92
|
1375
|
0,13
|
|
Gyrodinium spirale
|
30*10*10
|
1570
|
250
|
0,39
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
250
|
1,71
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
125
|
0,80
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
625
|
1,50
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
125
|
1,93
|
|
Protoperidinium sp.
|
32*28*17
|
132187
|
250
|
1, 13
|
|
Итого
|
|
|
5500
|
22,62
|
|
Cyanophyta sp.
|
60*3*3
|
377
|
750
|
0,27
|
|
Всего
|
|
|
158625
|
191,15
|
Таблица А.3 – Численность и биомасса фитопланктона на станции мыс Любви в апреле 2014 г.
|
Виды
|
Размер
клетки,
мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
750
|
0,91
|
|
Berkeleya scopulorum
|
156*6*5
|
3674
|
125
|
0,46
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
2500
|
13,25
|
|
Chaetoceros compressus
|
22*28*8
|
3868
|
750
|
2,90
|
|
Chaetoceros peruvianus
|
24*24*15
|
6782
|
125
|
0,85
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
125
|
0,02
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
153125
|
19,23
|
|
Licmophora Ehrenbergii
|
90*15*5
|
2250
|
875
|
1,97
|
|
Licmophora gracilis
|
62*10*2
|
413
|
125
|
0,05
|
|
Nitzschia tenuirostris
|
30*2*2
|
31
|
375
|
0,01
|
|
Pzeudonitzchia delikatissima
|
70*4*2
|
147
|
2625
|
0,39
|
|
Pzeudozosonelia calcar avis
|
575*38*17
|
133321
|
125
|
16,67
|
|
Sceletonema costatum
|
12*6*3
|
170
|
2250
|
0,38
|
|
Synedra crystallina
|
176*12*6
|
12672
|
250
|
3,17
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
3250
|
0,66
|
|
Итого
|
|
|
167375
|
60,92
|
|
Dinophyta:
|
|
|
|
|
|
Dinophysis acuta
|
38*32*17
|
10818
|
125
|
1,35
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
125
|
0,90
|
|
Glenodinium danicum
|
30*31*15
|
7301
|
125
|
0,91
|
|
Gonyaulax minima
|
22*16*11
|
2026
|
125
|
0,25
|
|
Gymnodynium simplex
|
11*4*4
|
92
|
4750
|
0,44
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
625
|
4,27
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
2000
|
4,81
|
|
Prorocentrum lima
|
42*32*25
|
17584
|
125
|
2,20
|
|
Protoperidinium conicum
|
88*90*45
|
94318
|
125
|
11,80
|
|
Protoperidinium globulus
|
56*54*28
|
44312
|
125
|
5,54
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
125
|
1,93
|
|
Heterocapsa triguetra
|
30*20*15
|
2944
|
125
|
0,37
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
125
|
0,52
|
|
Итого:
|
|
|
8625
|
35,29
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
125
|
0,05
|
|
Cyanophyta sp.
|
60*3*3
|
377
|
1375
|
0,50
|
|
Всего
|
|
|
177500
|
96,76
|
Таблица А.4 – Численность и биомасса фитопланктона на станции Южная Озереевка в апреле 2014 г.
|
Виды
|
Размер клетки,
мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
625
|
33,12
|
|
Chaetoceros curvisetus
|
35*35*10
|
2748
|
1250
|
3,43
|
|
Dactyliosolen fragilissima
|
56*10*10
|
4306
|
500
|
2,20
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
35750
|
4,50
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
30000
|
4,39
|
|
Pzeudonitzschia seriata
|
70*4*2
|
147
|
1000
|
0,45
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
125
|
16,66
|
|
Sceletonema costatum
|
12*6*3
|
170
|
750
|
0,13
|
|
Striatella unipunctata
|
88*12*6
|
1658
|
250
|
0,41
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
3750
|
0,77
|
|
Итого
|
|
|
74000
|
66,06
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
40*15*170
|
6431
|
125
|
5,44
|
|
Ceratium tripos
|
110*15*380
|
718929
|
125
|
89,87
|
|
Dinophysis acuta
|
38*32*17
|
10818
|
250
|
2,70
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
250
|
8,99
|
|
Gymnodynium sp.
|
22*12*10
|
1382
|
125
|
0,17
|
|
Gonyaulax spinifera
|
22*16*16
|
2947
|
250
|
0,74
|
|
Gyrodinium fusiforme
|
52*28*16
|
6096
|
500
|
3,05
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
1000
|
6,83
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
875
|
4,15
|
|
Prorocentrum minimum
|
25*18*9
|
2120
|
5500
|
13,24
|
|
Protoperidinium divergens
|
62*55*30
|
27554
|
125
|
3,44
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
250
|
3,86
|
|
Protoperidinium sp.
|
32*28*17
|
4537
|
375
|
1,70
|
|
Heterocapsa triguetra
|
30*20*15
|
2944
|
250
|
0,74
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
1000
|
4,19
|
|
Итого
|
|
|
11000
|
149,11
|
|
Всего
|
|
|
85000
|
215,17
|
Таблица А.5 – Численность и биомасса фитопланктона на станции порт в мае 2014 г.
|
Виды
|
Размер клетки,
мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
1625
|
86,10
|
|
Chaetoceros affinis
|
18*18*12
|
3052
|
750
|
2,29
|
|
Chaetoceros socialis
|
12*8*8
|
603
|
1625
|
0,98
|
|
Cocconeis scutellum
|
34*20*10
|
5338
|
250
|
1,33
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
1750
|
0,22
|
|
Leptocylindrus minimus
|
4*4*10
|
126
|
1125
|
0,14
|
|
Nitzschia holsatica
|
38*2*2
|
40
|
625
|
0,03
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
4750
|
0,70
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
125
|
16,66
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
750
|
0,15
|
|
Итого
|
|
|
13375
|
108,60
|
|
Dinophyta:
|
|
|
|
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
125
|
4,50
|
|
Glenodinium pilula
|
24*24*12
|
3617
|
125
|
0,45
|
|
Glenodinium rotundum
|
42*44*21
|
20310
|
250
|
5,07
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
250
|
1,19
|
|
Prorocentrum minimum
|
25*18*9
|
2120
|
250
|
0,60
|
|
Итого
|
|
|
1000
|
11,81
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia lanowii
|
32*6*2
|
435
|
375
|
0,16
|
|
Cyanophyta sp.
|
60*3*3
|
377
|
500
|
0,18
|
|
Всего
|
|
|
15250
|
120,75
|
Таблица А.6 – Численность и биомасса фитопланктона на станции Западный мол в мае 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
5500
|
291,43
|
|
Chaetoceros affinis
|
18*18*12
|
3052
|
500
|
1,53
|
|
Chaetoceros compressus
|
22*28*8
|
3868
|
2000
|
7,38
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
625
|
0,08
|
|
Licmophora Ehrenbergii
|
90*15*5
|
2250
|
250
|
0,56
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
250
|
33,33
|
|
Sceletonema costatum
|
12*6*3
|
170
|
4125
|
0,70
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
6500
|
1,33
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
625
|
2,14
|
|
Итого
|
|
|
20375
|
338,48
|
|
Dinophyta:
|
|
|
|
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
375
|
13,50
|
|
Glenodinium danicum
|
30*31*15
|
7301
|
125
|
0,91
|
|
Glenodinium pilula
|
24*24*12
|
3617
|
1000
|
3,62
|
|
Glenodinium rotundum
|
42*44*21
|
20310
|
125
|
2,54
|
|
Gymnodinium simplex
|
11*4*4
|
92
|
250
|
0,02
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
250
|
1,19
|
|
Итого
|
|
|
2125
|
21,78
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
500
|
0,22
|
|
Eutrepcia lanowii
|
32*6*2
|
435
|
125
|
0,05
|
|
Итого
|
|
|
625
|
0,27
|
|
Cyanophyta sp.
|
60*3*3
|
377
|
250
|
0,09
|
|
Всего
|
|
|
23375
|
360,62
|
Таблица А.7 – Численность и биомасса фитопланктона на станции мыс Любви в мае 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Achnanthes brevipes
|
87*25*17
|
36975
|
750
|
27,73
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
1750
|
2,14
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
2250
|
119,22
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
2000
|
0,25
|
|
Navicula pennata
|
38*5*4
|
597
|
875
|
0,52
|
|
Sceletonema costatum
|
12*6*3
|
170
|
1750
|
0,30
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
4500
|
0,92
|
|
Итого
|
|
|
13875
|
151,08
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
45*20*170
|
65482
|
125
|
5,54
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
125
|
4,50
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
125
|
0,59
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
250
|
0,60
|
|
Prorocentrum lima
|
42*32*25
|
17584
|
875
|
15,39
|
|
Protoperidinium divergens
|
62*55*30
|
27554
|
250
|
6,89
|
|
Protoperidinium pellucidum
|
50*40*25
|
14719
|
125
|
1,84
|
|
Protoperidinium sp.
|
32*28*17
|
4537
|
500
|
2,27
|
|
Итого
|
|
|
2275
|
37,62
|
|
Всего
|
|
|
16125
|
188,70
|
Таблица А.8 – Численность и биомасса фитопланктона на станции порт в июне 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
3375
|
9,27
|
|
Chaetoceros Lorencianus
|
17*10*7
|
934
|
1125
|
1,05
|
|
Melosira moniliformis
|
30*30*18
|
17309
|
1875
|
32,46
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
125
|
16,67
|
|
Sceletonema costatum
|
12*6*3
|
170
|
156250
|
26,49
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
375
|
0,08
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
1500
|
5,13
|
|
Итого
|
|
|
164625
|
91,15
|
|
Dinophyta:
|
|
|
|
|
|
Gymnodinium sp.
|
22*12*10
|
1382
|
250
|
0,17
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
625
|
2,97
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
750
|
1,80
|
|
Итого
|
|
|
1625
|
4,94
|
|
Euglenophyta:
|
|
|
|
|
|
Euglena sp.
|
45*7*2
|
789
|
625
|
0,49
|
|
Eutrepcia lanowii
|
32*6*2
|
435
|
250
|
0,10
|
|
Итого
|
|
|
875
|
0,59
|
|
Chryzophyta:
|
|
|
|
|
|
Emiliania huxleyi
|
8*8*8
|
402
|
15625
|
6,28
|
|
Всего
|
|
|
182750
|
102,96
|
Таблица А.9 – Численность и биомасса фитопланктона на станции Западный мол в июне 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
3375
|
9,27
|
|
Chaetoceros socialis
|
12*8*8
|
603
|
750
|
0,45
|
|
Cocconeis scutellum
|
34*20*10
|
5338
|
125
|
0,67
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
125
|
16,67
|
|
Sceletonema costatum
|
12*6*3
|
170
|
109375
|
19,39
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
500
|
0,10
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
125
|
3,85
|
|
Итого
|
|
|
114375
|
50,40
|
|
Dinophyta:
|
|
|
|
|
|
Dinophysis sacculus
|
52*30*15
|
12246
|
250
|
3,06
|
|
Gymnodinium sp.
|
22*12*10
|
1382
|
1875
|
3,87
|
|
Gonyaulax spinifera
|
34*30*18
|
9608
|
250
|
0,74
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
375
|
2,56
|
|
Prorocentrum micans
|
24*32*16
|
6431
|
250
|
1,19
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
250
|
0,60
|
|
Protoperidinium pellucidum
|
50*40*25
|
14719
|
125
|
1,84
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
125
|
0,52
|
|
Итого
|
|
|
3500
|
14,38
|
|
Euglenophyta:
|
|
|
|
|
|
Euglena sp.
|
45*7*2
|
789
|
1750
|
1,38
|
|
Eutrepcia lanowii
|
32*6*2
|
435
|
125
|
0,05
|
|
Итого
|
|
|
1875
|
1,43
|
|
Chryzophyta:
|
|
|
|
|
|
Emiliania huxleyi
|
8*8*8
|
402
|
21875
|
8,79
|
|
Всего
|
|
|
141625
|
75,00
|
Таблица А.10 – Численность и биомасса фитопланктона на станции мыс Любви в июне 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
4250
|
11,68
|
|
Chaetoceros Lorencianus
|
17*10*7
|
934
|
750
|
0,70
|
|
Chaetoceros similis
|
10*8*18
|
1130
|
1250
|
1,41
|
|
Chaetoceros socialis
|
12*8*8
|
603
|
625
|
0,38
|
|
Licmophora Ehrenbergii
|
90*15*5
|
2250
|
250
|
0,11
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
625
|
0,09
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
250
|
33,33
|
|
Sceletonema costatum
|
12*6*3
|
170
|
68750
|
11,66
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
1375
|
4,70
|
|
Итого
|
|
|
78125
|
64,06
|
|
Dinophyta:
|
|
|
|
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
125
|
0,85
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
375
|
0,90
|
|
Итого
|
|
|
500
|
1,75
|
|
Euglenophyta:
|
|
|
|
|
|
Euglena sp.
|
45*7*2
|
789
|
125
|
0,09
|
|
Chryzophyta:
|
|
|
|
|
|
Emiliania huxleyi
|
8*8*8
|
402
|
28125
|
11,30
|
|
Cyanophyta:
|
|
|
|
|
|
Oscillatoria sp.
|
5*3*3
|
35
|
4875
|
0,17
|
|
Всего
|
|
|
111750
|
77,37
|
Таблица А.11 – Численность и биомасса фитопланктона на станции порт в июле 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Chaetoceros compressus
|
22*28*8
|
3868
|
1625
|
6,27
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
2875
|
18,89
|
|
Chaetoceros danicus
|
18*15*10
|
2120
|
125
|
0,27
|
|
Chaetoceros peruvianus
|
24*24*15
|
6782
|
125
|
0,85
|
|
Chaetoceros socialis
|
12*8*8
|
603
|
3000
|
1,80
|
|
Chaetoceros sp.
|
21*15*10
|
2473
|
2625
|
6,50
|
|
Cocconeis scutellum
|
48*22*15
|
4145
|
250
|
1,04
|
|
Cyclotella caspia
|
6*6*6
|
170
|
4250
|
0,72
|
|
Sceletonema costatum
|
12*6*3
|
170
|
14500
|
2,46
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
250
|
0,05
|
|
Thalassiosira antiqua
|
24*24*12
|
5426
|
500
|
2,71
|
|
Итого
|
|
|
30125
|
41,56
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
45*20*170
|
65482
|
125
|
5,44
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
250
|
1,80
|
|
Gymnodinium simplex
|
11*4*4
|
92
|
500
|
0,05
|
|
Gymnodinium blax
|
12*7*6
|
264
|
375
|
0,09
|
|
Gyrodinium spirale
|
30*10*10
|
1570
|
125
|
0,20
|
|
Protoperidinium breve
|
82*60*40
|
59451
|
1625
|
96,60
|
|
Итого
|
|
|
3000
|
104,18
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
500
|
0,22
|
|
Всего
|
|
|
33625
|
145,96
|
Таблица А.12 – Численность и биомасса фитопланктона на станции Западный мол в июле 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Amphora inflexa
|
80*40*23
|
57776
|
125
|
7,22
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
375
|
19,87
|
|
Chaetoceros compressus
|
22*28*8
|
3868
|
1250
|
4,84
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
3125
|
8,59
|
|
Cyclotella caspia
|
6*6*6
|
170
|
1000
|
0,17
|
|
Navicula forcipata
|
56*20*10
|
8792
|
125
|
1,09
|
|
Sceletonema costatum
|
12*6*3
|
170
|
6250
|
1,06
|
|
Striatella unipunctata
|
88*12*6
|
1658
|
1500
|
2,49
|
|
Итого
|
|
|
13750
|
45,33
|
|
Dinophyta:
|
|
|
|
|
|
Glenodinium pilula
|
24*24*12
|
3617
|
250
|
0,90
|
|
Gonyaulax minima
|
22*16*11
|
2026
|
125
|
0,25
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
125
|
0,85
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
125
|
0,59
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
125
|
0,30
|
|
Protoperidinium breve
|
82*60*40
|
59451
|
375
|
22,29
|
|
Итого
|
|
|
1125
|
25,18
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
250
|
1,09
|
|
Всего
|
|
|
15125
|
71,60
|
Таблица А.13 – Численность и биомасса фитопланктона на станции мыс Любви в июле 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
1000
|
1,23
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
500
|
26,49
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
2250
|
6,18
|
|
Chaetoceros socialis
|
12*8*8
|
603
|
3250
|
1,96
|
|
Cyclotella caspia
|
6*6*6
|
170
|
1500
|
0,25
|
|
Navicula pennata
|
38*5*4
|
597
|
1000
|
0,60
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
250
|
33,33
|
|
Sceletonema costatum
|
12*6*3
|
170
|
10000
|
1,69
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
500
|
0,10
|
|
Thalassiosira antiqua
|
24*24*12
|
5426
|
1500
|
8,14
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
250
|
0,86
|
|
Итого
|
|
|
22000
|
80,83
|
|
Dinophyta:
|
|
|
|
|
|
Gonyaulax minima
|
22*16*11
|
2026
|
125
|
0,25
|
|
Gymnodinium simplex
|
11*4*4
|
92
|
125
|
0,01
|
|
Итого
|
|
|
250
|
0,26
|
|
Всего
|
|
|
22550
|
81,09
|
Таблица А.14 – Численность и биомасса фитопланктона на станции Южная Озереевка в июле 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
250
|
0,30
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
375
|
19,87
|
|
Chaetoceros affinis
|
18*18*12
|
30521
|
1000
|
3,05
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
1875
|
5,15
|
|
Chaetoceros holsaticus
|
22*13*7
|
1572
|
500
|
0,79
|
|
Licmophora Ehrenbergii
|
90*15*5
|
2250
|
250
|
0,56
|
|
Nitzschia tenuirostris
|
30*2*2
|
31
|
250
|
0,01
|
|
Proboscia alata
|
832*34*12
|
95330
|
125
|
11,91
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
3750
|
0,55
|
|
Pzeudonitzschia seriata
|
70*4*2
|
147
|
2500
|
0,37
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
1125
|
149,99
|
|
Sceletonema costatum
|
12*6*3
|
170
|
1375
|
0,23
|
|
Striatella unipunctata
|
88*12*6
|
1658
|
2250
|
3,73
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
3125
|
0,64
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
125
|
0,43
|
|
Итого
|
|
|
18625
|
197,58
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
40*15*170
|
43502
|
125
|
5,44
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
625
|
22,49
|
|
Glenodinium pilula
|
24*24*12
|
3617
|
125
|
0,45
|
|
Gonyaulax minima
|
22*16*11
|
2026
|
250
|
0,51
|
|
Gymnodinium blax
|
12*7*6
|
264
|
250
|
0,07
|
|
Gymnodynium wulffii
|
22*12*11
|
1520
|
500
|
0,76
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
500
|
3,42
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
375
|
1,78
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
3375
|
8,13
|
|
Protoperidinium depressum
|
82*60*41
|
62460
|
375
|
23,42
|
|
Protoperidinium granii
|
45*32*17
|
5823
|
125
|
0,73
|
|
Protoperidinium globulus
|
56*54*28
|
44312
|
125
|
5,54
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
125
|
1,93
|
|
Heterocapsa triguetra
|
30*20*15
|
2944
|
250
|
0,74
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
1500
|
6,28
|
|
Итого
|
|
|
8875
|
81,69
|
|
Cyanophyta sp.
|
60*3*3
|
377
|
2625
|
0,95
|
|
Всего
|
|
|
30125
|
280,22
|
Таблица А.15 – Численность и биомасса фитопланктона на станции порт в августе 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Amfora inflexa
|
80*40*23
|
57776
|
375
|
21,67
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
500
|
0,61
|
|
Cocconeis scutellum
|
48* 22*15
|
4145
|
500
|
2,07
|
|
Cyclotella caspia
|
6*6*6
|
170
|
2250
|
0,38
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
1000
|
0,09
|
|
Licmophora Ehrenbergii
|
90*15*5
|
2250
|
875
|
1,97
|
|
Sceletonema costatum
|
12*6*3
|
170
|
1125
|
0,19
|
|
Итого
|
|
|
6625
|
26,98
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
40*15*170
|
43502
|
125
|
5,44
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
500
|
17,99
|
|
Gymnodinium sp.
|
22*12*11
|
2904
|
1375
|
3,99
|
|
Gyrodinium spirale
|
30*10*10
|
1570
|
375
|
0,59
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
250
|
1,71
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
625
|
2,97
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
625
|
1,51
|
|
Итого
|
|
|
3875
|
34,20
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
1000
|
0,44
|
|
Cyanophyta:
|
|
|
|
|
|
Oscillatoria tenuis
|
4*4*6
|
113
|
625
|
0,07
|
|
Oscillatoria sp.
|
3*3*5
|
35
|
625
|
0,02
|
|
Итого
|
|
|
|
0,09
|
|
Мелкие жгутиковые
|
3*3*3
|
14
|
3875
|
0,05
|
|
Всего
|
|
|
16625
|
61,76
|
Таблица А.16 – Численность и биомасса фитопланктона на станции Западный мол в августе 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Asterionella glacialis
|
56*10*2
|
879
|
625
|
0,66
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
875
|
46,36
|
|
Chaetoceros affinis
|
48* 22*15
|
4145
|
375
|
1,15
|
|
Chaetoceros rigidus
|
8*10*4
|
251
|
4000
|
1,01
|
|
Cyclotella caspia
|
6*6*6
|
170
|
4500
|
0,76
|
|
Sceletonema costatum
|
12*6*3
|
170
|
1250
|
0,21
|
|
Итого
|
|
|
11625
|
50,14
|
|
Dinophyta:
|
|
|
|
|
|
Dinophysis acutа
|
38*32*17
|
10818
|
125
|
1,35
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
250
|
8,99
|
|
Glenodinium caspicum
|
42*52*21
|
24002
|
125
|
3,00
|
|
Glenodinium pilula
|
24*24*12
|
3617
|
125
|
0,45
|
|
Gonyaulax minima
|
22*16*11
|
2026
|
500
|
1,01
|
|
Gymnodinium sp.
|
22*12*11
|
2904
|
1375
|
3,99
|
|
Gyrodinium spirale
|
30*10*10
|
1570
|
250
|
0,39
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
375
|
1,78
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
250
|
0,60
|
|
Protoperidinium brevipes
|
82*60*40
|
59451
|
375
|
22,29
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
625
|
2,62
|
|
Итого
|
|
|
4250
|
46,47
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
3875
|
1,69
|
|
Chryzophyta:
|
|
|
|
|
|
Emiliania huxleyi
|
8*8*8
|
402
|
875
|
0,35
|
|
Мелкие жгутиковые
|
3*3*3
|
14
|
4250
|
0,06
|
|
Всего
|
|
|
24875
|
98,71
|
Таблица А.17 – Численность и биомасса фитопланктона на станции мыс Любви в августе 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Bacillaria paradoxa
|
78*5*4
|
1225
|
1500
|
1,84
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
250
|
13,25
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
3875
|
0,34
|
|
Licmophora gracilis
|
62*10*2
|
413
|
1750
|
0,72
|
|
Navicula pennata
|
38*5*4
|
597
|
1000
|
0,60
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
625
|
83,33
|
|
Striatella unipunctata
|
88*12*6
|
1658
|
250
|
0,41
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
500
|
0,10
|
|
Итого
|
|
|
9750
|
100,59
|
|
Dinophyta:
|
|
|
|
|
|
Dinophysis acutа
|
38*32*17
|
10818
|
125
|
1,35
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
1375
|
49,47
|
|
Gymnodinium simplex
|
11*4*4
|
92
|
250
|
0,02
|
|
Gyrodinium spirale
|
30*10*10
|
1570
|
125
|
0,19
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
250
|
1,71
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
500
|
1,20
|
|
Protoceratium reticulatum
|
36*32*20
|
12058
|
125
|
1,51
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
1250
|
19,31
|
|
Peridinium guinguecorne
|
52*34*18
|
31824
|
625
|
21,86
|
|
Итого
|
|
|
4625
|
96,08
|
|
Euglenophyta:
|
|
|
|
|
|
Eutrepcia sp.
|
32*6*2
|
435
|
125
|
0,05
|
|
Cyanophyta:
|
|
|
|
|
|
Oscillatoria sp.
|
3*3*5
|
35
|
750
|
0,03
|
|
Chryzophyta:
|
|
|
|
|
|
Emiliania huxleyi
|
8*8*8
|
402
|
750
|
0,30
|
|
Мелкие жгутиковые
|
3*3*3
|
14
|
1250
|
0,02
|
|
Всего
|
|
|
17250
|
197,07
|
Таблица А.18 – Численность и биомасса фитопланктона на станции Южная Озереевка августе 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Chaetoceros compressus
|
22*28*8
|
3868
|
625
|
2,42
|
|
Pzeudonitzschia delikatissima
|
70*4*2
|
147
|
2625
|
0,39
|
|
Pzeudonitzschia seriata
|
70*4*2
|
147
|
1375
|
0,20
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
1000
|
133,32
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
3250
|
0,66
|
|
Итого
|
|
|
8875
|
136,99
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
40*15*170
|
43502
|
250
|
10,88
|
|
Ceratium fusus
|
140*40*15
|
88836
|
125
|
11,10
|
|
Ceratium tripos
|
110*15*380
|
718929
|
125
|
89,87
|
|
Diplopsalis lenticula
|
50*55*25
|
35979
|
250
|
8,99
|
|
Gonyaulax minima
|
22*16*11
|
2026
|
250
|
0,51
|
|
Gonyaulax spinifera
|
22*16*16
|
2947
|
250
|
0,74
|
|
Gonyaulax digitale
|
45*35*22
|
18134
|
250
|
4,53
|
|
Gymnodinium simplex
|
11*4*4
|
92
|
125
|
0,01
|
|
Gyrodinium spirale
|
30*10*10
|
1570
|
125
|
0,20
|
|
Lingulodinium polyedrum
|
45*40*232
|
21666
|
125
|
2,71
|
|
Prorocentrum compressum
|
34*24*16
|
6833
|
125
|
0,85
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
125
|
0,59
|
|
Prorocentrum minimum
|
23*20*10
|
2407
|
750
|
1,81
|
|
Protoperidinium divergens
|
62*55*30
|
27554
|
125
|
3,44
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
375
|
5,79
|
|
Protoperidinium globulus
|
56*54*28
|
44312
|
125
|
5,54
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
125
|
0,52
|
|
Итого
|
|
|
3625
|
148,08
|
|
Chryzophyta:
|
|
|
|
|
|
Emiliania huxleyi
|
8*8*8
|
402
|
3125
|
1,26
|
|
Всего
|
|
|
15625
|
286,33
|
Таблица А.19 – Численность и биомасса фитопланктона на станции порт в сентябре 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Amphiprora alata
|
87*28*34
|
65017
|
125
|
8,13
|
|
Chaetoceros affinis
|
18*18*12
|
3052
|
2625
|
8,01
|
|
Cocconeis scutellum
|
48* 22*15
|
4145
|
125
|
0,52
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
500
|
0,04
|
|
Pzeudonitzschia seriata
|
70*4*2
|
147
|
500
|
0,07
|
|
Pleurosigma elongatum
|
130*20*10
|
6803
|
125
|
0,85
|
|
Proboscia alata
|
832*34*12
|
95330
|
250
|
23,83
|
|
Sceletonema costatum
|
12*6*3
|
170
|
14000
|
2,37
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
250
|
0,05
|
|
Thalassiosira excentrica
|
62*62*10
|
30175
|
8750
|
264,04
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
3875
|
13,25
|
|
Итого
|
|
|
31125
|
321,16
|
|
Мелкие жгутиковые
|
3*3*3
|
14
|
1750
|
0,02
|
|
Всего
|
|
|
32875
|
321,18
|
Таблица А.20 – Численность и биомасса фитопланктона на станции Западный мол в сентябре 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Amfora hualina
|
50*36*20
|
28260
|
125
|
3,53
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
625
|
0,05
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
250
|
13,25
|
|
Chaetoceros affinis
|
18*18*12
|
3052
|
1750
|
5,34
|
|
Ditylum Brightwellii
|
12*12*110
|
12434
|
250
|
3,11
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
1125
|
0,14.
|
|
Navicula pennata
|
38*5*4
|
597
|
875
|
0,52
|
|
Pzeudonitzschia seriata
|
70*4*2
|
147
|
1125
|
0,16
|
|
Proboscia alata
|
832*34*12
|
95330
|
375
|
35,75
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
250
|
33,33
|
|
Sceletonema costatum
|
12*6*3
|
170
|
5250
|
0,89
|
|
Thalassiosira excentrica
|
62*62*10
|
30175
|
3000
|
90,53
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
2000
|
6,84
|
|
Итого
|
|
|
17000
|
193,44
|
|
Dinophyta:
|
|
|
|
|
|
Dinophysis caudata
|
58*44*35
|
46744
|
250
|
11,69
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
250
|
1,19
|
|
Итого
|
|
|
500
|
12,88
|
|
Мелкие жгутиковые
|
3*3*3
|
14
|
500
|
0,01
|
|
Всего
|
|
|
18000
|
206,33
|
Таблица А.21 – Численность и биомасса фитопланктона на станции мыс Любви в сентябре 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Cylindrotheta closterium
|
56*3*2
|
88
|
250
|
0,02
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
625
|
33,18
|
|
Chaetoceros peruvianus
|
24*24*15
|
6782
|
250
|
1,70
|
|
Chaetoceros rigidus
|
8*10*4
|
251
|
1125
|
0,28
|
|
Leptocylindrus danicus
|
4*4*10
|
126
|
2500
|
0,31
|
|
Proboscia alata
|
832*34*12
|
95330
|
250
|
23,83
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
375
|
49,99
|
|
Sceletonema costatum
|
12*6*3
|
170
|
7625
|
1,29
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
500
|
0,10
|
|
Thalassiosira excentrica
|
62*62*10
|
30175
|
2000
|
60,35
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
1250
|
4,27
|
|
Итого
|
|
|
16750
|
175,32
|
|
Dinophyta:
|
|
|
|
|
|
Prorocentrum micans
|
36*28*9
|
4748
|
375
|
1,78
|
|
Protoperidinium divergens
|
62*55*30
|
27554
|
125
|
3,44
|
|
Protoperidinium steinii
|
56*25*27
|
15451
|
125
|
1,93
|
|
Итого
|
|
|
625
|
7,15
|
|
Мелкие жгутиковые
|
3*3*3
|
14
|
250
|
0,00
|
|
Всего
|
|
|
17625
|
182,47
|
Таблица А.22 – Численность и биомасса фитопланктона на станции Южная Озереевка в сентябре 2014 г.
|
Виды
|
Размер клетки, мкм
|
Объём клетки, мкм
|
Численность, млн.кл/м3
|
Биомасса, мг/м3
|
|
Bacyllariophyta:
|
|
|
|
|
|
Cerataulina pelagica
|
30*30*75
|
52988
|
1375
|
72,86
|
|
Chaetoceros affinis
|
18*18*12
|
3052
|
625
|
1,91
|
|
Chaetoceros curvisetus
|
35*10*10
|
2748
|
500
|
1,37
|
|
Chaetoceros rigidus
|
8*10*4
|
251
|
1000
|
0,03
|
|
Chaetoceros peruvianus
|
24*24*15
|
6782
|
500
|
3,39
|
|
Chaetoceros Wighamii
|
18*15*10
|
2120
|
875
|
1,86
|
|
Coscinodiscus sp.
|
63*63*23
|
71660
|
125
|
8,96
|
|
Detonula confervacea
|
22*22*7
|
2660
|
750
|
1,99
|
|
Proboscia alata
|
832*34*12
|
95330
|
125
|
11,92
|
|
Pzeudosolenia calcar avis
|
575*38*17
|
133321
|
500
|
66,66
|
|
Sceletonema costatum
|
12*6*3
|
170
|
7625
|
1,29
|
|
Thalassionema nitzschioides
|
65*2*2
|
204
|
500
|
0,10
|
|
Thalassiosira excentrica
|
62*62*10
|
30175
|
1000
|
30,18
|
|
Thalassiosira sp.
|
22*22*9
|
3419
|
250
|
0,86
|
|
Итого
|
|
|
15750
|
203,38
|
|
Dinophyta:
|
|
|
|
|
|
Ceratium furca
|
40*15*170
|
43502
|
250
|
10,88
|
|
Dinophysis acuta
|
38*32*17
|
10818
|
125
|
1,35
|
|
Protoperidinium divergens
|
62*55*30
|
27554
|
125
|
3,44
|
|
Protoperidinium granii
|
45*32*17
|
5823
|
125
|
0,73
|
|
Scrippsiella trochoidea
|
22*18*20
|
4187
|
250
|
1,05
|
|
Heterocapsa triguetra
|
30*20*15
|
2944
|
250
|
0,74
|
|
Итого
|
|
|
1125
|
18,19
|
|
Всего
|
|
|
16875
|
221,57
|
ПРИЛОЖЕНИЕ Б
Внешний вид некоторых представителей фитопланктона Новороссийской бухты
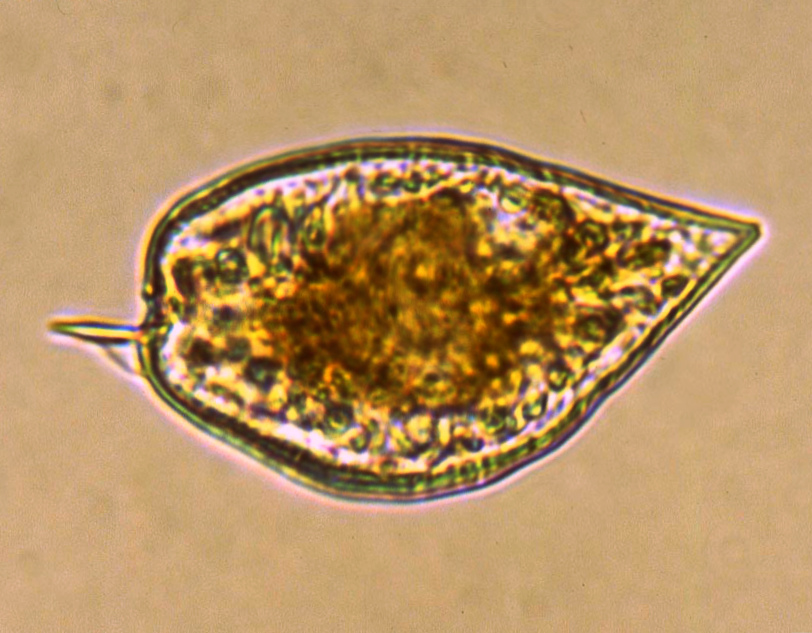
Рисунок Б.1 – Prorocentrum micans
Рисунок Б.2 – Prorocentrum minimum
Рисунок Б.3 – Sceletonema costatum

Рисунок Б.4 – Ceratium tripos
ВЛИЯНИЕ АНТРОПОГЕННОЙ НАГРУЗКИ НА ФИТОПЛАНКТОН НОВОРОССИЙСКОЙ БУХТЫ