Гістологія центральної нервової системи
Зміст
Вступ………………………………………………………………………2
Загальна анатомія нервової системи……………………………………3
Гістологія центральної нервової системи…………………………........6
Фізіологія центральних нейронів………………………………………22
Рецепція й поведінка комах…………………………………………….27
Висновок…...……………………………………………………………52
Список літератури………………………………………………………53
Вступ
Комахи — поширені на всьому суходолі, включаючи Антарктиду, тварини розміром від 0,2 міліметру до 33 сантиметри. Тіло комах складається з трьох відділів (тагм) — голови,грудей і черевця, кожнен з яких складається з сегментів (сегменти голови злиті) і вкритий хітинізованими покривами. На голові є пара членистих вусиків (антени), ротові придатки, органи зору. До грудей кріпляться 3 пари членистих ніг і 1—2 пари крил. На кінці черевця розташовані статеві і хвостові придатки (церки), іноді жало. Органи виділення — головним чином, мальпігієві судини. Кровоносна система незамкнена. Функцію серця виконує спинна судина, функцію крові — гемолімфа. Органи дихання — трахеї (у водних — зябро-трахеї), що відкриваються назовні дихальцями. Органи чуття дуже різноманітні, в основі будови багатьох з них лежать так звані сенсили, розкидані по різних частинах тіла поодиноко або зібрані у великі скупчення. Є органи зору — фасеткові очі та прості вічка, слуху — тимпанальний апарат, дотику — чуттєві волоски, нюху — ямки і волоски, рівноваги — хордотональний апарат та ін.
Майже всі комахи роздільностатеві. Розвиток відбувається з перетворенням (метаморфоз), яке буває повним (коли личинка різко відрізняється від дорослої особини (імаго) і перетворюється на лялечку, з якої виходить доросла комаха), або неповним (личинка, що вийшла з яйця, має вигляд, близький до імаго, стадії лялечки нема).
Відомо 1-3 млн видів, поширених від Антарктиди до 84° північної широти. Більшість комах — наземні тварини, багато видів живе в землі, деякі в прісних водах, окремі — в морях. Більшість видів комах — фітофаги, є всеїдні (таргани), хижаки (поїдають інших комах, молюсків, червів тощо), копрофаги (гнойовики), кровисисні (комарі, воші, блохи).
Є багато видів, які мають позитивне чи негативне значення для людини. Деякі комахи приносять користь як запилювачі рослин; деякі виробляють речовини, які використовуються як харчові продукти та сировина для промисловості (мед, віск, шовк, природні фарби). Багато комах — шкідники сільського та лісового господарств, окремі — переносники паразитів та збудників хвороб людини, тварин або рослин, де які самі є паразитами.
Загальна анатомія нервової системи
Живі створіння незалежно від ступеня складності їх організації реагують на оточуюче їх середовище. Таке реагування необхідне їм для існування : не тільки для уникнення небезпек та загрозливих їх благополуччю умов, але воно забезпечує їм процес харчування та розмноження.
Комахи мають центральну нервову систему, яка являє собою ряд нервових вузлів – гангліїв, які з’єднуються між собою продольними парними стволами – конективами. У вищих комах відбувається зменшення кількості гангліїв за рахунок їхнього об’єднання до 2–3 і навіть одного, що пов’язано з їхньою складною поведінкою.
У комах розрізняють головний мозок та тулубний (черевний нервовий ланцюжок). В свою чергу тулубний мозок може бути поділений на грудний (грудні ганглії) та черевний (черевні ганглії) відділи. Кількість гангліїв, які входять в нервовий ланцюжок. Кількість гангліїв може варіювати у різних видів, при чому у комах спостерігається тенденція до злиття гангліїв. Ця тенденція відображає більш високий рівень розвитку центральної нервової системи цих комах порівняно з іншими представниками ряду членистоногі. Проте дати функціональну оцінку такій тенденції в кожному окремому випадку у комах важко, тому що максимальний ступінь злиття гангліїв у цих тварин найчастіше не корелює з висотою їх організації.
Головний мозок комах представлений надглотковим та частково підглотковим гангліями. Часто під мозком комахи розуміють лише надглотковий ганглій, тоді як підглотковий повністю відносять до черевного ланцюжка. Проте ьаке ділення не оправдано з гістологічної точки зору, оскільки разом із загальними рисами, характерними для сегментарних гангліїв , «ганглій виявляє ознаки, які дозволяють роздивлятись його як орган більш високого порядку, який відповідає заднім відділам мозку хребтового ( його стовбурової частини). Функціонально підглотковий ганглій також відрізняється від гангліїв черевного нервового ланцюжка.
Надглотковий ганглії складається з трьох частин: протоцерембрума, дейтоцеребрума та тритоцеребрума. Протоцеребрум (який ще називають «зоровим мозком») здійснює зв’язки з очима та очками, дейтоцеребрум («нюховий мозок») отримує сенсорні волокна від антен, а також іннервує антенальні м’язи, тритоцеребрум подає нерви до верхньої губи, а також зв’язує мозок з вегетативною нервовою системою. Найскладніший – протоцеребрум, бо там знаходяться характерні утворення, у різному ступені розвинуті у різних форм і мають назву грибоподібних тіл.
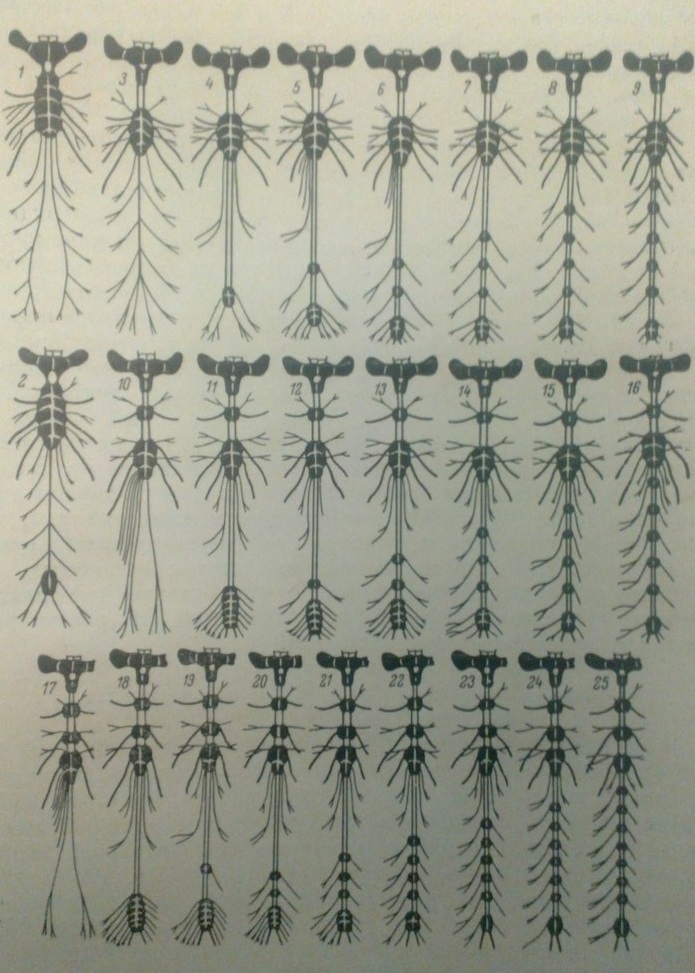
(Рис.1)Загальний вигляд нервового ланцюжка різних комах
Підглотковий ганглій складається з декількох злитих гангліїв. Він подає нерви до сенсорних органів та м’язам ротової частини, а також до рецепторів шиї. У ряду комах підглотковий ганглій може зливатися з грудними гангліями.
Грудний відділ черевного нервового ланцюжка включає проторакальний, мезоторакальний, та метаторакальний ганглії, які можуть лежати роздільно, чи зливатися разом, у деяких комах зливаються лише мезоторакальний та метаторакальний ганглії. Грудні ганглії винятково важливі в локомоторній діяльності комах: саме вони управляють рухами кінцівок і крил. Топографія і число нервів, що постачають ноги і крила, може варіювати у різних видів.
Значно менш великі і більш просто влаштовані ганглії входять до складу черевного відділу нервової ланцюжка. Максимальне число гангліїв 8, але в ряді випадків ганглії можуть зливатися як між собою, так і з грудними гангліями, і тоді число черевних гангліїв може варіювати від 1 до 7; у деяких комах вільні черевні ганглії можуть взагалі бути відсутні - вони повністю зливаються з метаторакальними.
Окрім центральної , у комах розрізняють вегетативну нервову систему , яка складається з трьох відділів: краниального , каудального і тулубного , причому тулубовий відділ відповідає симпатичної нервової системі хребетних тварин. Цей відділ представлений у комах так званим вентральним непарним нервом, що постачають м'язи дихальців , трахеї , серце , кишечник і деякі інші внутрішні органи.
Нарешті , периферична нервова система комах , як і інших тварин , складається з провідних відділів (як правило , змішаних нервів) , ефекторних закінчень і рецепторних клітин з допоміжним апаратом.
Гістологія центральної нервової системи
В основі сучасних уявлень про гістологічних особливості нервової системи комах лежить велика кількість робіт різних авторів . Не намагаючись перерахувати тут всі роботи в цій області , слід вказати на дослідження оптичних гангліїв , черевного нервового ланцюжка і чутливої нервової системи личинок бабок, мозку і оптичних центрів таргана, черевної нервового ланцюжка бабок і сарани, мозку і черевного нервового ланцюжка сарани , провідних шляхів мозку і черевного нервового ланцюжка сарани , бджіл , мух , личинок бабок і деякі інші.
Цікаво відзначити , що певну і все більшу допомогу у вирішенні низки нейрологических завдань надають морфологію нейрофізіологи . Це стосується , наприклад , таких питань, як простежування шляхів окремих інтернейронов або їх груп протягом кількох гангліїв , що здійснити за допомогою гістологічних методик виявляється надзвичайно важким.
На допомогу тут приходять методи « електрофізіологічної анатомії » - вивчення шляхів проведення нервових імпульсів , швидкості проведення цих імпульсів , визначення часу синаптичних затримок і т. п. Тому в розділі глави , присвяченій структурі центральної нервової системи комах , будуть зустрічатися і роботи деяких нейрофізіологів .
Незважаючи на велику кількість досліджень , провідні шляхи і тонка будова елементів центральної нервової системи комах вивчені ще недостатньо. Особливо це відноситься до гістологічним дослідженням головних гангліїв . Тому в даному розділі ми . Висвітлимо тільки той морфологічний матеріал , який стосується гістології черевної нервової ланцюжка і необхідний для розуміння і тлумачення наведених фізіологічних фактів .
Основною структурною одиницею центральної нервової системи комах , як і інших високоорганізованих г : тварин, є нейрон. У гангліях комах розташовуються тіла мото-і інтернейронов . Клітинні тіла сенсорних нейронів знаходяться на значній відстані від центральної нервової системи; ці нейрони фактично є первічночувствую -ські рецепторними клітинами ( Вінніков , 1946 , 1971).
Розглянемо основні структурні особливості різних нейронів комах.
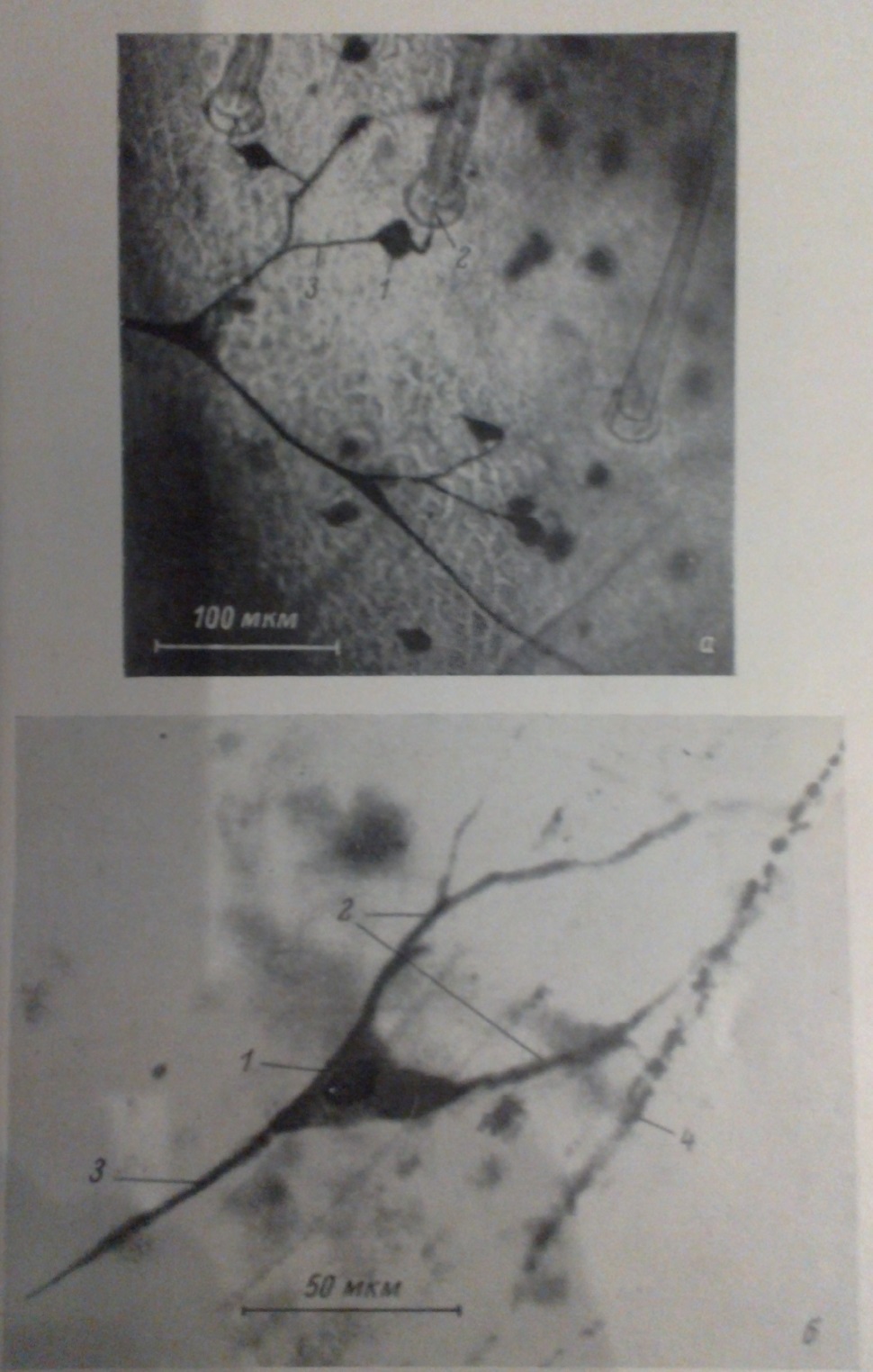
(Рис.2) Мікрофотографії сенсорних нейронів азіатської сарани
На допомогу тут приходять методи « електрофізіологічної анатомії » - вивчення шляхів проведення нервових імпульсів , швидкості проведення цих імпульсів , визначення часу синаптичних затримок і т. п. Тому в розділі глави , присвяченій структурі центральної нервової системи комах , будуть зустрічатися і роботи деяких нейрофізіологів .
Незважаючи на велику кількість досліджень , провідні шляхи і тонка будова елементів центральної нервової системи комах вивчені ще недостатньо. Особливо це відноситься до гістологічним дослідженням головних гангліїв . Тому в даному розділі ми . Висвітлимо тільки той морфологічний матеріал , який стосується гістології черевної нервової ланцюжка і необхідний для розуміння і тлумачення наведених фізіологічних фактів .
Основною структурною одиницею центральної нервової системи комах , як і інших високоорганізованих г : тварин, є нейрон. У гангліях комах розташовуються тіла мото-і інтернейронов . Клітинні тіла сенсорних нейронів знаходяться на значній відстані від центральної нервової системи; ці нейрони фактично є первинновідчуваючими рецепторними клітинами ( Вінніков , 1946 , 1971).
Розглянемо основні структурні особливості різних нейронів комах.
Сенсорні нейрони. Клітинні тіла цих нейронів мають звичайно біполярну або мультиполярними форму і лежать поблизу відповідного сенсорного органу або инервуємої тканини. Розміри клітинних тел сенсорних нейронів вкрай невеликі і складають в поперечнику всього кілька мікронів.
Відповідно до класифікації Заварзіна ( 1924а ) , всі сенсорні нейрони комах можна розділити на два типи. Нейрони I типу - це нервові клітини з неветвящихся дендритом, який набирає небудь сенсорний орган модифікованої кутикули (наприклад , сенсорний волосок або орган хімічного почуття) . Аксон нейрона йде в центральну нервову систему. Нейрони II типу - це нервові клітини з вільно розгалуженими дендритами , що закінчуються в якої тканини і не пов'язаними зі спеціалізованими сенсорними органами. Серед клітин II типу зустрічаються і полівалентні нейрони , дендрити яких можуть закінчуватися в один і той же час в різних тканинах , наприклад в епітелії і сполучної тканини , трахеях і м'язах. Аксони нейронів II типу також прямують в ганглії центральної нервової системи.
Сенсорні органи є частиною спеціалізованої кутикули і можуть бути представлені волосками , гаіпікамі , пластинками та іншими утвореннями , що продукуються гіподермальними клітинами ( тріхогенной - волоскообразующей н тормоген - мембранообразующей ) . Іннервація сенсорних органів може здійснюватися як однією , так і багатьма сенсорними клітинами. Вступаючи в центральну нервову систему ( у відповідний ганглії ) , сенсорні аксони закінчуються там у вигляді розгалужень , що утворюють синаптичні контакти з відростками центральних нейронів. Аксони деяких сенсорних нейронів можуть давати колатералі усередині даного ганглія , а самі проходити в наступний ганглій .
Моторні нейрони. Клітинні тіла цих нейронів лежать під оболонкою ганглія переважно з вентральної його боку і з боків і мають зазвичай Уніполярні форму. Разом з тим в центральній нервовій системі комах можуть зустрічатися і нейрони біполярного типу. Однак форма клітинних тіл нейронів - значною мірою чисто формальний показники н залежить від положення тіла нейронів стосовно нейропіля н своїм відростках ( Заварзін , 1916; Орлов , 1925).
Розміри клітинних тел мотонейронів у комах дуже невеликі і рідко досягають 40-50 мкм. Наприклад , в грудних гангліях американського таргана описані мотонейрони , сома яких не перевищує в діаметрі 30 мкм. Ще більш дрібні нейрони ( сома 15 мкм ) виявлені в грудному ганглії мухи uSarcophaga bullata Разом з тим у комах іноді зустрічаються і дуже великі клітинні тіла мотонейронів . Так , сома мотонейронів , що іннервують разгибатель гомілки пустельної сарани , може досягати в поперечнику 90 - 110 мкм ( найбільші тіла нейронів у метаторакальном ганглії комахи ).
Топографія нейронів комах відрізняється рядом важливих особливостей . Якщо у хребетних тварин відростки нейрона ( дендрити і аксон ) , як правило , відходять від тіла клітини , то у комах і дендрити і аксон центральних нейронів відходять немає від тіла нейрона , а від особливого мозкового відростка ( мозкового розгалуження ); це розгалуження у свою чергу пов'язано з тілом клітини клітинним відростком , який часто має відносно велику довжину ( Заварзін , 1924а ; Цвіленева , 1970 ; Плотнікова , 1973). У результаті область синаптичних контактів нейрона може бути віднесена на значну відстань від клітинного тіла. Важливо відзначити , що самі клітинні тіла нейронів комах позбавлені будь-яких синаптичних контактів .
До недавнього часу вважали , що тіла мотонейронів розташовуються в ганглії в тому ж самому сегменті , що і іннервіруемие їм м'язові волокна. Однак було виявлено, що у деяких комах поздовжні спинні м'язи , які знаходяться в одному сегменті і беруть участь у локомоції , можуть отримувати аксони від мотонейронів , розташованих не тільки в ганглії того ж сегменту , але і в сусідньому ганглії . Такі відносини були виявлені у деяких видів метеликів, сарани і тарганів . Не виключена можливість подібних же межсегментарних відносин і для інших м'язів комах. У гангліях личинки бабки Aeschna і таргана Periplaneta amerscana було виявлено контралатеральное положення тіл мотонейронів ( « перехрещені нейрони ») . У всіх цих випадках мозковий розгалуження нейрона , від якого відходять дендрити і аксон , також розташовується в тому ж ганглії або на тій же стороні ганглія , що і тіло нейрона. Однак Цвіленевой в грудному ганглії личинки бабки Aeschna були виявлені мотонейрони , тіла яких розташовані контралатерально стосовно відходить від мозкового розгалуження аксону , в той час як саме мозковий розгалуження було іпсилатеральний стосовно аксону ( « ложноперекрещенние нейрони » ) .
Таким чином , топографія мотонейронів комах може бути досить складною. Однак у всіх випадках клітинні тіла подібних за функції мотонейронів і в ще більшому ступені їх відростки розташовуються за певним « планом» , який строго витримується в кожному ганглії .
У нейропіля мотонейрони комах утворюють синаптичні контакти з різними інтернейронамі , будучи кінцевою ланкою полісінаптіческіх дуг різного ступеня складності. Двочленні ( моносинаптичні ) рефлекторні дуги , тобто синаптичні контакти мотонейронів безпосередньо з розгалуженнями сенсорних волокон , хоча і виявлені у комах морфологічно, мабуть , не типові для центральної нервової системи цих тварин .
Характер розгалуження дендритів мотонейронів може бути різним: дифузним , захоплюючим широкі області нейропіля , або ж порожнисте локальним. Нарешті , у комах можливі випадки, коли аксон мотонейрона , розташованого в одному з гангліїв , віддає коллатераль , що проходить в коннектіве в сусідній ганглій (так званий моторно- асоціативний тип нейрона ) . Нейрони такого типу, характерні для центральної нервової системи черв'яків, були виявлені в грудних гангліях азіатської сарани. У всіх випадках аксони мотонейронів йдуть на периферію через відходять від ганглія соматичні нерви.
Інтернейрони . За положенням клітинних тіл ( на периферії ганглія ) , за їх формою (зазвичай уніполярної ) , а також по загальній конфігурації нейрона (наявність мозкового розгалуження , від якого відходять дендрити і аксон ) інтернейрони аналогічні мотонейронам . Проте розміри клітинних тел інтернейронов ще дрібніше , ніж мотонейронів , і в гангліях деяких видів становлять всього 2-3 мкм. 1 Разом з тим у комах описані і інтернейрони , розміри клітинних тел яких перевищують розміри тіл мотонейронів . Подібні, дещо незвичні для комах відносини в гангліях мухи Sarcophaga bullata передбачає Чіародо , хоча автор і не призводить переконливих даних щодо ідентифікації нейронів.
Інтернейрони є сполучними ланками між сенсорними і моторними нейронами ( інтернейрони першого порядку) , причому число таких вставних елементів може бути і більшим , і тоді розрізняють інтернейрони 2 -го , 3-го і більш високих порядків . Терміни « нейрони першого » або « вищого порядку » використовуються , однак , не стільки для топографічного розрізнення інтернейронов , скільки для їх функціональної характеристики , і тому застосовуються по відношенню до якогось певного нервового шляху. Існує ще одна функціональна класифікація інтернейронов комах. При наявності більш ніж одного вставного нейрона між сенсорним і моторним нейронами інтернейрон , безпосередньо пов'язаний з сенсорним нейроном, називають іноді «сенсорним інтер нейроном», а інтернейрон, імпульсація якого передається мотонейронами, - «моторним інтернейрон».
Інтернейрони можуть бути сегментарними або мультісегментарнимі залежно від того, взаємодіють вони виключно с нейронами в одном ганглии или же в некотором числе последовательных ганглиев. Большинство интернейронов принадлежит к мультисегментарному типу. Пример такого интернейрона, имеющего дендритические разветвления по крайней мере в нейропилях трех ганглиев водного клопа-водомерки. Характерно, что даже в сравнительно просто устроенном IV брюшном ганглии личинки стрекозы Заварзин (1924а) смог продемонстрировать только два нейрона, которые могут быть отнесены к сегментарному типу.
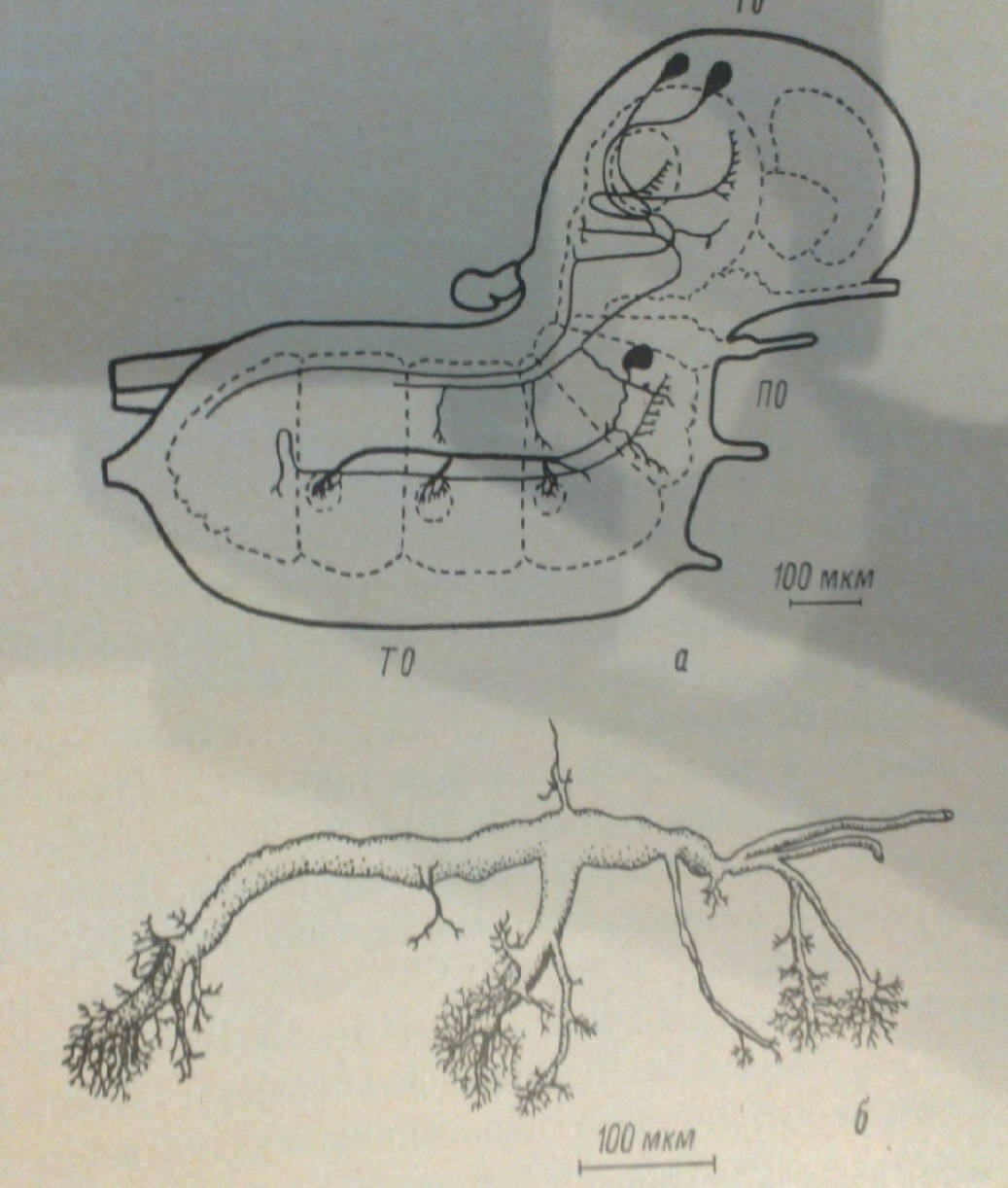
(Рис.3) Схема центральної нервової системи клопа, яка показує локалізацію мультисегментарного нейрона
Вже цей опис показує , наскільки складними і різноманітними можуть бути зв'язки інтернейронов комах. Дійсно, навіть інтернейрони сегментарного типу черевного ганглія личинки бабки , складові асоціативний апарат даного ганглія, здійснюють зв'язку не тільки між нейронами одного боку ганглія , але можуть пов'язувати також праву і ліву його половини ( комісурний зв'язок ). Ще більш складна топографія інтернейронов , що здійснюють поздовжні (коннективність) зв'язку між гангліями . Ряд аналогічних нейронів утворює у комах особливі тракти , або шляху. Шляхи ці можуть бути поперечними , що з'єднують бічні відділи ганглія один з одним , або поздовжніми , що з'єднують окремі пункти нейропіля в одному ганглії або в декількох сусідніх гангліях . З найбільш довгих поздовжніх зв'язків , призначених для передачі інформації на великі відстані , добре відома система так званих гігантських волокон.
Назва « гігантські » інтернейрони , складові ці системи , отримали не стільки внаслідок їх великої протяжності , скільки через дуже великого діаметра аксонів , що досягає у деяких комах декількох десятків мікрон. До теперішнього часу системи гігантських висхідних волокон , що беруть початок в останньому черевному ганглії і йдуть вгору по ланцюжку до грудних , а у деяких видів , можливо , навіть до головних гангліїв , досліджені у тарганів ( Roeder , 1948 ; Hess , 1958а ; Farley , Milburn , 1969 ; Harris , Smyth , 1971a ; Milburn , Bentley , 1971 ) , сарани ( Cook , 1951 ; Gheze , 1968 ; Seabrook , 1970 , 1971 ) , личинок бабок ( Hughes , 1953 ; Mill , 1964 ), деяких метеликів ( Тищенко , 1962 , 1963 ) , цвіркунів (Bentley , 1969а ) , і , мабуть , такі системи є типовими для всього класу комах.
Функціональна роль цих інтернейронов , що полягає і здійсненні « реакції тікання » ( escape reaction ) від небезпеки , що загрожує комасі ззаду , до останнього часу здавалася безсумнівною . Проте нещодавно з'явилися дані , що говорять про те , що принаймні у американського таргана гігантські інтер- нейроіи можуть виконувати зовсім іншу функцію , а саме брати участь у контролі спрямованих рухів антен.
Крім висхідних волокон , у комах існують спеціалізовані системи інтернейронов, які забезпечують спадні впливу ( Guthrie , 1964 ; Свідерський , 1967б ) . Функціональна роль цих систем може бути різною; багато інтернейрони беруть участь і контролі локомоторних реакцій комахи. Одна з таких систем , необхідна для запуску і підтримки роботи м'язів Крилової апарату , детально вивчена у азіатській сарани.
Кількість нейронів у комах . Підрахунки показують , що число центральних нейронів у комах вкрай невелика, причому навіть порівняно складні локомоторні акти цих тварин можуть забезпечувати часом лише десятки нейронів , а іноді і їх одиниці. Наприклад , всю мускулатуру 2 -й пари кінцівок личинки бабки Aeschna обслуговує всього 6 мотонейронів ( Заварзін , 1924а ) . Навіть таку складну м'язову систему , як крилова , можуть обслужити у азіатській і пустельної сарани , мабуть , не більше 80 мотонейронів в кожному грудному ганглії .
На жаль , в даний час є всього кілька підрахунків сумарного числа нейронів у гангліях комах. Один з таких підрахунків на тотальних препаратах провів Заварзін , який в II грудному ганглії личинки бабки Aeschna знайшов 140 нейронів (або , на його думку , приблизно 70 % ' загального числа нейронів , наявних в даному ганглії взагалі), з них близько 40 нейронів представляли собою мотонейрони , а решта 100 - інтернейрони. Для черевних гангліїв цього ж комахи загальне число нейронів становило 90.
Підрахунки кількості нейронів були здійснені і на зрізах гангліїв ряду комах. У личинки бабки Апах і палочника Carausius morosus в черевних гангліях було виявлено близько 600 недронов , хоча автори і не підрозділяли підраховані нейрони на мото-і інтернейрони . Значно більше число нейронів виявлено в останньому черевному ганглії , який представляє собою складну структуру і складається з декількох злилися гангліїв . Так , у личинки бабки Апах , за даними Філден і Хьюза ( Fielden , Hughes , 1962 ) , в цьому ганглії виявлено близько 1200 нейронів ( мото-і інтернедронов ) . Ще більша кількість нейронів ( більше 2100 , з них близько 2000 інтернейронов ) містить останній черевної ганглій цвіркуна Acheta domesticus . Нарешті , також в злившись метаторакальном ганглії ( що складається з останнього грудного і декількох злилися з ним черевних гангліїв ) американського таргана , за даними Кохена і Жаклета ( Cohen , Jacklet , 1967 ) , є близько 3000 нейронів , причому найбільш великих (з діаметром клітинного тіла більше 20 мкм) налічується 230 . З цієї кількості великих нейронів авторам вдалося ідентифікувати як мотонейрони близько 100 клітин , що , мабуть , не є повним числом мотонейронів усередині даного ганглія .
Серед центральних нейронів комах переважають клітини з відносно невеликим розміром клітинних тел. Про характер розподілу діаметрів соми нейронів проторакального ганглія палочника може дати уявлення гістограма .
Роблячи поправку на деякі обмеження існуючих методів підрахунку нейронів комах , ми можемо вважати, що всю складну моторну і інтегративну функцію , яку здійснюють ганглії цих тварин , виконує не більше 102 - 103 нейронів або в перерахунку на черевну нервову ланцюжок (без урахування надглоткового і подглоточного гангліїв ) не більше 103 – 104 нейронів. Для порівняння зазначимо , що загальне число нейронів у спинному мозку собаки становить 107 і , таким чином , число нейронів у комах в порівнюваних відділах центральної нервової системи виявляється меншим на 3-4 порядки (у кілька тисяч разів).
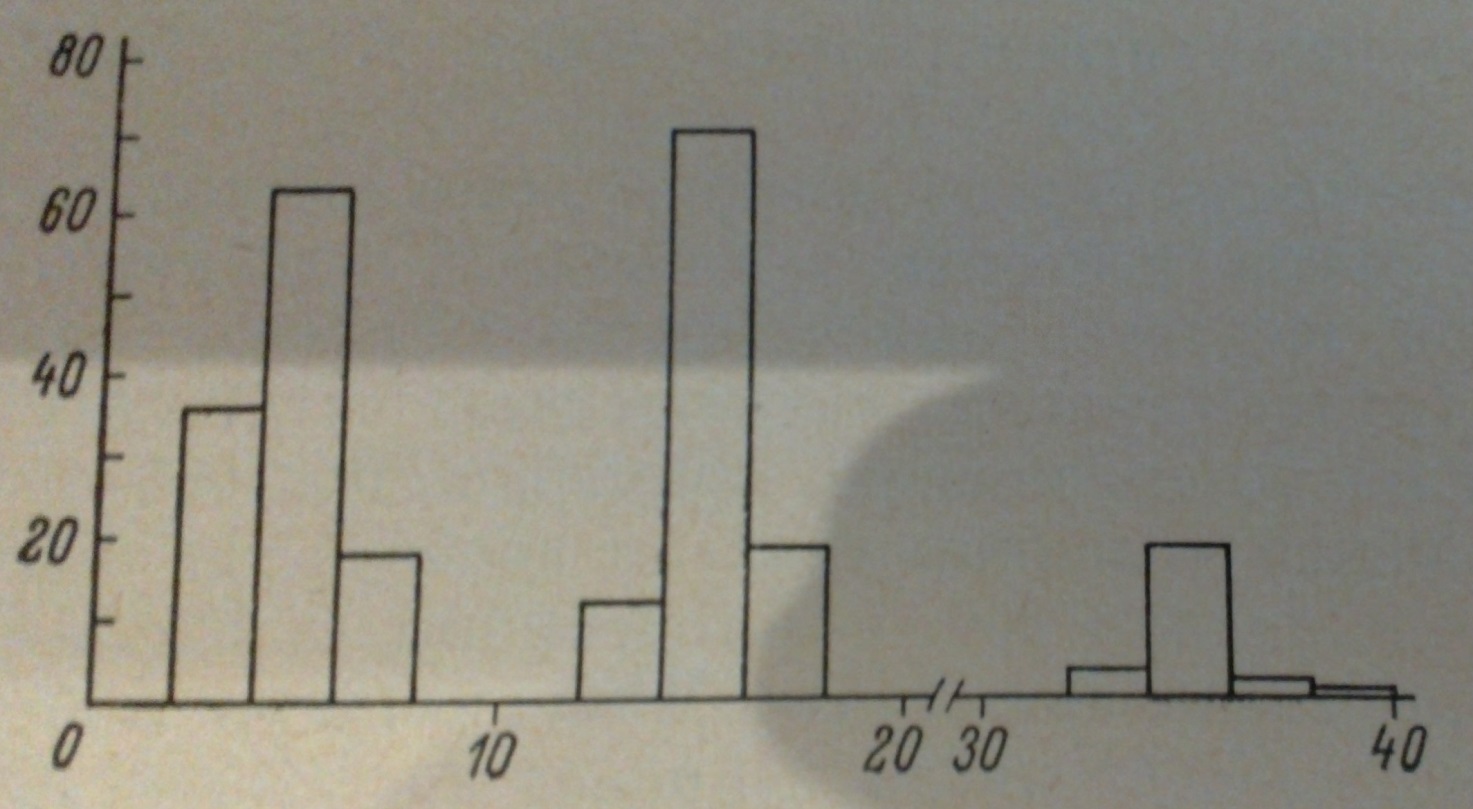
(Рис.4) Гістограма розподілення діаметрів соми нейронів проторакального ганглія палочника
У значно більшій кількості в порівнянні з центральними представлені у комах периферически розташовані сенсорні нейрони. Наприклад , на антені медоносної бджоли налічується більше 2000 сенсорних нейронів , на половині членика клопа - більше 420 клітин і т. п. Проте і ці кількості нейронів у комах виявляються у багато разів меншими порівняно з такими у хребетних тварин.
Нейрпропіль. Нейропіль являє собою місце нейрональної інтеграції , область , в якій генеруються і організовуються картини складних поведінкових реакцій комах. Саме в цій області здійснюються синаптичні зв'язки між сенсорними , моторними і проміжними елементами нервової системи.
Незважаючи на велику складність своєї будови , нейропіль комах може бути поділений на праву і ліву половини і на кілька зон , або областей. Велика заслуга у встановленні топографічних відносин розгалужень нейронів в нейропілі належить Заварзіну, який в гангліях личинки бабки виділив п'ять п'ять основних областей: 1) дорсальну, що є областю коміссурних або, точніше, коннективних волокон; 2) область дорсальних корінців; 3) область основного нейропіля (найзначніша по товщині область ганглія); 4 ) область вентральних корінців і 5) область вентральних (коннективність) волокон. У свою чергу в основному нейропіля Заварзін розрізняв дорсальну , середню і вентральную зони. Аналогічні області можуть бути виділені і в нейропіля гангліїв іншої комахи - азіатської сарани.
В області дорсальних корінців розташовуються дендрітіческіе розгалуження всіх рухових нейронів ганглія ( руховий нейропіля ) . Аксони цих нейронів виходять з нейропіля через дорсальні корінці бічних нервів. Область вентральних корінців відповідає області чутливого нейропіля . Вона утворена центральними відростками периферичних розташованих сенсорних нейронів , аксони яких входять до нейропіля через вентральні корінці.
Таким чином , у комах є чітке підрозділ на чутливий і руховий Нейропен , які відокремлені один від одного товщею основного нейропіля , що містить асоціативні елементи . Ця область найбільш складна за структурою. Тут розташовуються розгалуження інтернейронів, здійснюють часом дуже складні зв'язки . Можна виділити але принаймні три групи інтернейронов , розгалуження дедрітов і аксонів яких розташовуються в основному нейропіля . До першої групи належать інтернейрони , здійснюють зв'язку з руховою областю нейроліля , до другої - з чутливою , і, нарешті « до третьої - інтернейрони власного апарату , відростки яких не виходять за межі мозкової речовини ганглія і забезпечують зв'язки тільки всередині цього ганглія , т. е . інтернейрони сегментарного типу.
Область дорсальних конвективних волокон містить аксони , які можна розділити на дві групи : одні з них проходять з коннектіви одного ганглія в коніектнву іншого , інші ж , увійшовши в конвективную область через одну з Коннект , закінчуються в нейропіля термінальними розгалуженнями . Деякі з проходять волокон дають в руховій області нейропіля термінальні колатералі , інші - на всьому протязі таких терминалей , як правило , не дають. Нарешті , область вентральних коннективність волокон містить волокна як проходять , так і термінальні .
Вже наведені дані показують , наскільки складна і диференціювання структура нейропіля комах , особливо його асоціативного апарату . Не випадково Заварзін вказував , що внутрішня структура гангліїв комах порівнянна зі структурою туловищного мозку вищих представників хребетних тварин - птахів і ссавців. Разом з тим не можна не враховувати , що в різних представників комах , що знаходяться на різному рівні еволюційного розвитку і володіють різною мірою розвиненим локомоторним апаратом , внутрішня структура нейропіля певних гангліїв може виявитися більш диференційованої або . навпаки, менш диференційованою , ніж це було описано вище. Зокрема, дослідження черевних і грудних гангліїв іншої комахи - сарани - показало певний примітивізм структурних відносин у цієї комахи , що виражалося головним чином в більшій диффузности розподілу мозкових розгалужень в ненропіле . Цей факт можна пояснити більш слабким розвитком польоту в сарани порівняно з бабкою . Ймовірно , саме унікальний політ , удосконалювався в процесі еволюції , вплинув на диференціювання чутливого ненроший бабки і більш чітке відокремлення її локомоторних центрів. Однак у загальному плані будови гангліїв сарани і бабки відзначається глибоке і принципову схожість .
Ультраструктура нейронів та їх зв'язків і нейропіля. Перші спроби дослідити тонку структуру центральної нервової системи комах за допомогою електронного мікроскопа були зроблені в кінці 50-х років. В останні десятиліття інтенсивність електронномікроськопічного вивчення нейтральної нервової системи комах значно зросла. У результаті всіх цих робіт була досліджена ультраструктура ганліїв черевного нервового ланцюжка деяких представників тарганів , метеликів , мурашок , сарани , палочников , дорослих мух та їх личинок , а також личинок бабок , комарів і мошок.
Субмікроскопічна організація клітинних тіл нейронів комах виявилася схожою з такою хребетних тварин. Не було виявлено суттєвих відмінності і між структурою клітинних тел у комах різних видів: нейрони відрізнялися між собою в деталях - за кількістю цитоплазми , характеру і ступеня розвитку ЕПР , кількістю мітохондрій , кількістю та розмірами включень в цитоплазмі , а також по просторості інвагінацій плазматичної мембрани. Як цього і слід було очікувати на підставі гістологічних робіт , будь-яких синапсів на поверхні тіл нейронів комах виявлено не було. Було показано також , що сома нейронів комах покрита концентричними шарами гліальних клітин , що перешкоджають встановленню будь-яких синаптичних контактів на поверхні перікаріона .
У зв'язку з цим особливий інтерес для вивчення представляє ультраструктура розгалужень отробтков нейронів в нейропіля , особливо в синаптичних зонах. Потрібно відзначити , що область синаптичних перемикань виявилася найбільш складною для електронномікроськопічеського аналізу . На електронних мікрофотографіях важко було , наприклад , з достовірністю провести різницю між розгалуженнями дендритів і аксонів , і автори називають часто ці утворення невизначено - « нервові волокна ».
В результаті цілого ряду робіт било виявлено , що нервові волокна в нейропіля утворюють складну систему. Діаметр їх в цій області коливається від 0.1 мкм до кілька мікромікрон . Між нервовими волокнами розташовуються гліальні елементи , які є свого роду ізолюючим матеріалом. Однак є місця , де нервові волокна позбавлені гліального покриття , саме в цих областях вони приходять в зіткнення і здатні утворювати синаптичні контакти . У той же час деякі великі , мабуть , що проходять аксони не втрачають глиальной оболонки на всьому своєму протязі.
У всіх нервових волокнах нейропіля були виявлені типові органели , характерні і для волокон хребетних тварин : протонейрофібрілли ( діаметр яких у волокнах таргана Periplaneta americana становить 100-150 А. а у волокнах мухи Sarcophaga bullata - близько 200 А ) , мітохондрії ( які розташовуються уздовж по довжині волокон) , мікропухирці , елементи а шорсткогоЕПР . нейросекреторні включення та інші.
В даний час описано декілька типів синаптичних контактів між нейронами комах : поперечні ( перехресні ) контакти , поздовжні контакти та контакти типу термінального гудзичка.

(Рис.5) Типи синаптичних контактів в нейропілі комах
Ці види синаптичних зв'язків є, мабуть, загальними для нейроіпілей всіх комах. Контакти типу термінальній гудзички зустрічаються на кінцях нервових волокон; поздовжні контакти утворюються між волокнами, расположеннимй паралельно один одному, і, нарешті поперечні ~ між волокнами, розташованими під кутом.
Пресинаптичне нервове водокно у комах може утворювати контакти як з одним постінаптіческім волокном, так і з кількома такими волокнами одночасно. З іншого боку, іноді два пресинаптических волокна можуть мати синапси з одним і тим же постсинаптическим волокном. Крім того , в нейропіля деяких комах існують так звані послідовні ( serial ) синапси . Синапси в цьому випадку розташовуються уздовж але нервового волокна на деякій відстані один від одного , що дає можливість цьому волокну здійснювати синаптичні зв'язки з низкою сусідніх волокон.
Більшість пресинаптических нервових волокон комах містить синаптичні пухирці. Такі бульбашки в синапсах центральної нервової системи цих тварин вперше були описані Хесом ( Hess. 1958b ) . Вважають , що і у комах бульбашки містять кванти хімічного медіатора. Діаметр синаптичних бульбашок становить 200-600 A . В області синапсу бульбашки можуть утворювати скупчення , або грона.
Цікаво , що пресинаптичні волокна у комах зазвичай більше за діаметром , ніж постспнаптпческпе ( в середньому 0.5-4.0 і 0.1-0.25 мкм відповідно). Осборн ( Osborne , 1966 ) пояснює це тим . що саме в пресинаптическом волокні повинен знаходитися деякий резерв метаболічних речовин і речовини медіатора. Треба зазначити, однак , що синаптичні пухирці у комах зустрічаються іноді і в постсінаптіческнх волокнах , хоча і в значно менших кількостях.
Між пресінаітнческой і постсинаптичної мембранами в синапсі комах є щілину розміром 100-200 А , причому обидві мембрани , за даними Осборна ( Osborne , 1966 ) , тришарові . Більш широка синаптична щілину (близько 300 А ) описана нещодавно в синапсах центральної нервової системи домашньої мухи. Загальне уявлення про ультраструктурну організації центрального синапсу комахи дає електронна мікрофотографія .
Наведені вище матеріали свідчать про те , що ультраструктура синаптичних утворень в центральній нервовій системі комах має принципову схожість з такою синапсів хребетних , які здійснюють передачу нервового імпульсу за допомогою певних хімічних речовин ( медіаторів ) . Все це дає можливість припускати наявність подібних механізмів синаптичної передачі в цих синапсах і у комах .
Однак у комах , як ми бачили , зустрічаються і синапси , в яких синаптичні пухирці розташовуються по обидві сторони синаптичної щілини. Функціональна роль такого роду « неполяризованих » синапсів залишається поки неясною.
Крім щойно описаних синаптичних зв'язків , в яких в тій чи іншій формі здійснюється хімічна передача нервового імпульсу , між нервовими волокнами комах в нейропіля були виявлені дуже тісні контакти особливого роду.
Вони були утворені за допомогою пятислойной мембранної структури, яка була наслідком з'єднання двох (пре- і постсинаптичні ) тришарових мембран ( Osborne , 1966). Синаптичних бульбашок в зонах таких контактів виявлено не було. Не виключено , що зазначені контакти відповідають синапсам з електричною передачею нервового імпульсу.
Конективи . Конективи є парними структурними утвореннями ( тяжами ) , що зв'язують між собою ганглії нервової ланцюжка. У них проходять аксони інтернейронов як висхідного і низхідного типів , так і аксони сенсорних нейронів , що не обмежуються тими гангліями , в які ці аксони вступають по вентральним корінцях . Нарешті , в Коннект- вах можуть проходити також аксони нейросекреторну клітин , а у деяких комах - і колатералі розгалужень моторних аксонів. Проте бблипую частина волокон , що проходять в коннектівах , складають аксони інтернейронов .
Певний порядок в розташуванні нейрональних елементів , існуючий в нейропіля , зберігається також і в коннекивавах , волокна в яких . проходять строго орієнтовано по відношенню до поздовжньої осі коннектив . Так , в коннектівах між грудними гангліями личинки бабки Aeschna найбільш великі волокна розташовуються в дорсальній , частково у вентральній частинах коннективи, тоді як пучки більш тонких волокон проходять між цими товстими волокнами; нарешті , найтонші волокна знаходяться в середній частині коннектіви .
Те , що такий порядок розташування волокон не випадковий , доводить схожість в топографії волокон правої і лівої коннектив. На підставі аналізу топографії волокон на поперечних зрізах коннектив Заварзін приходить до висновку , що , як і в нейропіля гангліїв , в коннектівах в глибині розташовані волокна коротких шляхів , тоді як в поверхневих частинах - коннективність волокна довгих шляхів. Виняток , мабуть , становлять гігантські висхідні аксони у тарганів і сарани , що проходять ближче до середини коннектив .
Число аксонів , що проходять в коннектівах , може бути дуже великим . Так , у пустельної сарани в коннектівах , що пов'язують подглоточний і проторакальний ганглії , проходить близько 3100 аксонів , в коннектівах між про-і мезоторакальнимі гангліями - близько 4000 , мезо- та метаторакальним гангліями - близько 8000 , нарешті , між метаторакальним і 1- м черевним гангліями - близько 2400 аксонів ( Rowell , Dorey , 1967). При цьому приблизно 60 % всіх аксонів мають діаметр менше 1 мкм , 25 % - 1-2 мкм і тільки 5 % аксонів мають діаметр 5 мкм і більше. У черевних коннектівах гусениць Saturnia pyri і Antherаеа pernyi переважна кількість волокон становлять дрібні волокна , діаметр ж найбільших з них не перевищує 5 мкм ( Тищенко , 1962).
У коннектівах комах не були виявлені картини , які свідчили б про розгалуженні аксонів або про наявність будь-яких синаптичних контактів між ними ( Rowell , Dorey , 1967).
Оболонка . Як ганглії , так і коннектіви комах оточені оболонкою , в якій можуть бути виділені два шари - зовнішній неклітинний і лежить під ним клітинний шар ( Ash - hurst , 1959 , 1968 ; Wigglesworth , 1959). Зовнішній шар отримав назву нейральной пластинки , а внутрішній - перінейріума (Smith , Treherne , 1963).
Неклітинний шар - нейрального платівка - містить основну речовину і велике число колагенових волокон ( Richards , Schneider , 1958 ) , причому цей шар , як вважають , чисто механічно відмежовує нервову систему комах від оточуючих рідин і тканин і надає лише невеликий опір проходженню всередину ганглія різних речовин та іонів.
Основною частиною оболонки нервової системи, що регулює таке проходження, є клітинний шар - іерінейріум , який являє собою , таким чином , виборчий іонний бар'єр. Цікаво , що характер і вираженість неклітинного і клітинного шарів оболонки у тих чи інших комах можуть розрізнятися досить значно , що , по всій ймовірності , знаходиться в прямому зв'язку з хімічним складом гемолімфи у різних видів цих тварин.
Не виключено, однак, що певна бар'єрна функція належить і внутрішнім частинам ганглія .
Фізіологія центральних нейронів
До питання про електричної збудливості мембрани клітинних тіл нейронів . До недавнього часу недостатньо ясним залишалося питання про те, чи володіє чи ні мембрана клітинних тіл нейронів центральної нервової системи комах електричної збудливістю . Одні автори ( Hoyle , 1970 ) відстоювали точку зору про електричної НЕ збудливості мембрани , інші - наводили дані , що свідчать про те , що мембрана соми але принаймні деяких центральних нейронів комах є електрично збудливою (і функціонує як класична мембрана нейронів хребетних тварин .
Зараз , мабуть , можна вважати встановленим , що електричної збудливістю у комах володіє мембрана соми лише дуже невеликого числа центральних нейронів , які в грудних гангліях таргана й сарани розташовуються дорсально. Водночас мембрана центрально розташованих тіл нейронів (а таких , як відомо, в гангліях переважна більшість) електрично невозбудимі ,
В даний час ще важко сказати , яка функціональна роль дорсально розташованих нейронів з електрично збудливою мембраною клітинних тел. Мабуть , ці нейрони ідентичні раніше виявленим в нашій лабораторії Плотникової ( 19696 ) еффекторним нейронам з кількома симетричними аксонами , що йдуть в різні пари соматичних нервів. Можна думати , що такі нейрони виконують і організмі комахи регулюючу ( Плотникова , 19096 ) або , як вважають Гроссман та ін ( Groflsman el aJ . , 1972 ) , гальмівну функцію. Відповідь на це питання можуть дати лише подальші нейрофізіологічні дослідження .
Електрична активність нейронів нейтральної нервової системи , За допомогою внутріклетоних відведень було показано , що величина потенціалів спокою клітинних тел центральних нейронів комах досягає 50 -70 верб. Потенціали дії в сомі переважної більшості нейронів центральної нервової системи не реєструються. Тому найбільший інтерес для дослідження представляє область розгалужень відростків нейронів комах в нейропіля .
На жаль , мала товщина дендритів і аксонів комах ( від десятих часток мікрона до декількох мікрон) ускладнювала до останнього часу реєстрацію електричної активності цих утворень за допомогою внутрішньоклітинних мікроелектродів . Однак важливі відомості про характер імпульсної активності в нормі , а також на тлі різних впливів , як і про синаптичних зв'язках центральних нейронів , вдалося отримати за допомогою позаклітинних відведень . Такого роду дослідження , піонером яких слід вважати Редера ( Roeder , 1948 ) , були виконані в наступні роки за допомогою мікроелектродів на нейронах гангліїв личинок бабок , дорослої сарани , цикад , тарганів , гусениць , цвіркунів і ряду інших комах.
Всі ці дослідження , однак , практично не зачіпали питань тонких механізмів міжнейронної синаптичноїпередачі . Виняток становлять , мабуть , лише роботи японських ( Yamasaki , Narahashi , 1960 ) і французьких ( Pichon , Callec , 1970 ; Callec et al . , 1971 ) авторів . У першій з цих робіт , вивчаючи реакцію гігантських інтернейронов таргана у відповідь на стимуляції церкального нерва, авторі отримали електричні феномени, які вони інтерпретували як церкальний потенціал пресинаптичний потенціал , збудливий постеінаптнческій потенціал і потенціал гігантських волокон. В інших роботах методом так званої « масляної пробки» автори в цій же системі інтернейронов таргана реєстрували як збуджуючі , так н гальмівні постспнаптнческіе потенціали при електричному разараженіі церкальних нервів або тактильному подразненні рецепторів церок .
Крок вперед у вивченні механізмів синаптичної передачі у комах був зроблений у зв'язку з використанням внутрішньоклітинних мікроелектродів . Було показано , що нейрони комах , як і нейрони хребетних , активуються збудливими постсинаптическими потенціалами , а гальмуються гальмівними постсинаптическими потенціалами , причому як за формою , так і за амплітудою постсинаптические потенціали у комах і хребетних тварин є аналогічними. Про характер електричної активності в нейронах Крилової апарату польового цвіркуна при генерації і гальмуванні ритму польоту комахи, а також про потенціалах дії , зареєстрованих при внутрішньоклітинному відведенні в нейропіля азіатської сарани ( спонтанно нерозряджаються одиниця).
Цікаво відзначити , що амплітуда потенціалів дії , реєструються в нейропіля комах , виявилася надзвичайно вариабельной , причому перевищення потенціалу дії над потенціалом спокою спостерігати не вдавалося. Так , потенціали дії у волокнах нейропіля американського таргана досягали всього 2-5 мв при потенціалах спокою 20-50 мв , а в нейропіля польового цвіркуна - 5-40 мв при потенціалах спокою 20-40 мв ; перевищення було відсутнє. Амплітуда потенціалів дії в нейропіля пустельній і азіатської сарани в ряді випадків досягала 40 - 50 мв . Однак перевищення також не спостерігалося. Нарешті , в нейропіля пустельної сарани реєстрували потенціали дії, що становлять всього 3-25 мв ( при потенціалах спокою , що досягали 50-56 мв ) , а - в нейропіля гангліїв будинкового цвіркуна потенціали дії , рівні 12.4 + 4.05 мв ( при потенціалах спокою 37.7 +5.5 мв ) .
Приписати одержувані результати небудь методичним похибок (наприклад , пошкодження кінчиком мікроелектрода тонких волокон в нейропіля ) було важко , оскільки потенціали спокою цих волокон в дослідах виявлялися досить високими і, як правило , підтримувалися стабільними протягом тривалого часу. Було потрібно шукати інше пояснення , і воно було запропоновано в роботі Хойла ( Hoyle , 1970).
У цій роботі, виходячи з гістологічних і фізіологічних даних , Хойл представив цікаву уніфіковану модель мотонейрона , яка , мабуть , може бути поширена і на інтернейрони комах. Хойл обгрунтовано вважає , що місцем ініціації імпульсів в нейроні комахи є истонченная область аксона поблизу виходу останнього з нейропіля . Внутрішньоклітинні реєстрації в цій частині нейрона крайнє рідкісні і нестабільні через дуже малого діаметру волокна. Практично відведення можуть бути здійснені в наступних частинах нейрона : 1) у аксоні , після виходу (або при виході) його нейропіля ; 2 ) у мозковому розгалуженні («нейропілярном сегменті » , по Хойлу ), 3) в сомі нейрона.
Із зони ініціації імпульс йде по товщають аксону на периферію - ортодромной і в глиб нейропіля ( по истонченной частини аксона ) - антидромний . Антидромний імпульс поширюється електротоніческі і швидко згасає у зв'язку з малою товщиною (і, отже , великим опором ) провідних шляхів в нейропіля . У результаті в області мозкового розгалуження в залежності від того , наскільки близька зона регистрації до місця ініціацій імпульсу, можуть бути відводяться потенціали дії більшою чи меншою амплітуди, яка, однак, ніколи не буде перевищувати величини потенціалу спокою. І лише в аксоні після виходу його з Нейропен я можуть бути зареєстровані «повноцінні» потенціали дії з перевищенням. Навпаки, в сомі нейрона із зазначених вище причин реєструються тільки потенціали спокою.
Запропонована Хойлом модель нейрона комахи не може бути на даному етапі досліджень ні підтверджена, ні спростована.
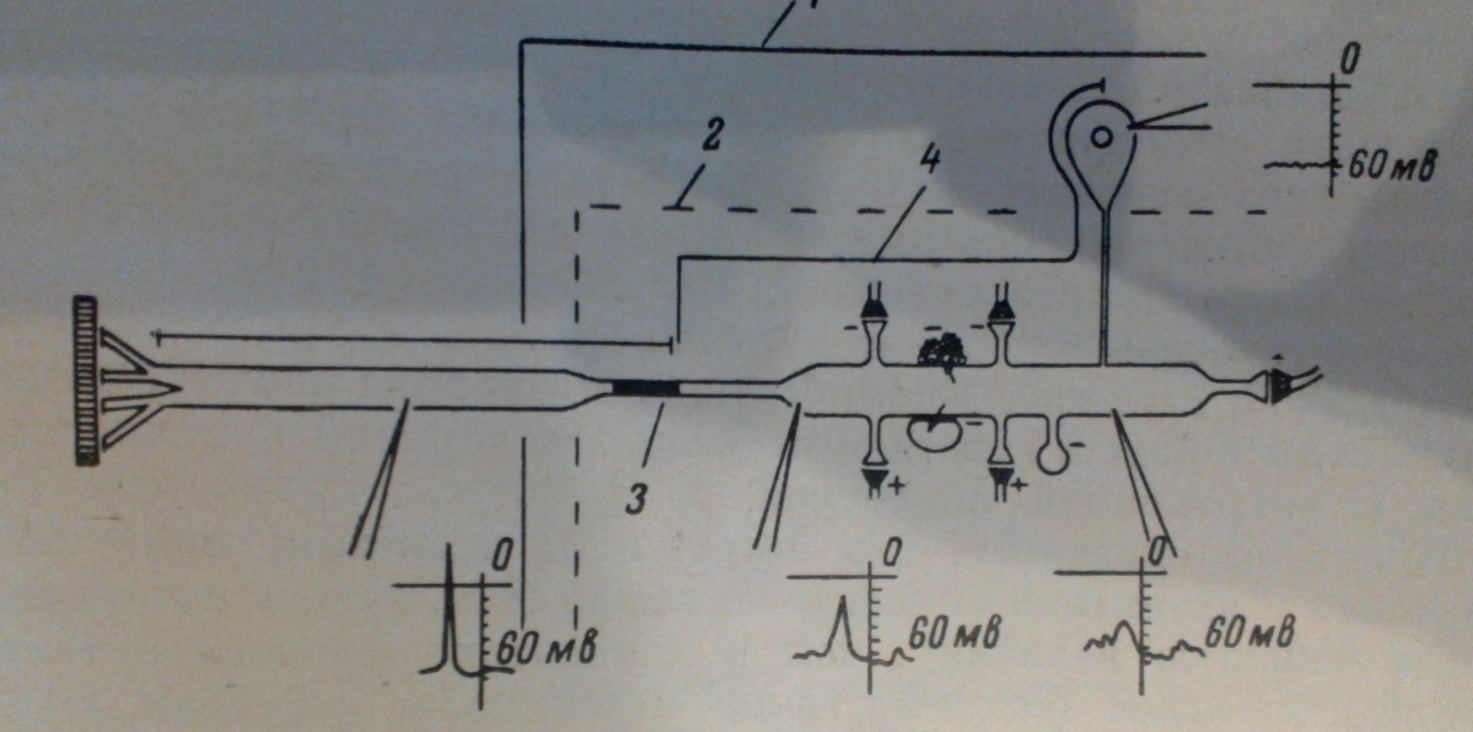
(Рис.6) Схема «типового нейрона» комахи
На закінчення отого розділу необхідно зазначити , що у комах не можна виключити можливості існування нейронів не з одним , а з декількома пунктами генерації нервового імпульсу ( Hughes , 1965). Такі нейрони виявлені, зокрема , в ганггліях черевної нервової ланцюжка інших членистоногих - ракоподібних ( Kennedy , Mellon , 1964). На жаль , будь-які фактичні дані про наявність у комах такого роду нейронів в даний час повністю відсутні.
Рецепція й поведінка комах
Рецептор - елементарний орган почуттів , утворений сенсорними нейроном або нейронами і допоміжними структурами. Налаштований на сприйняття адекватних подразнень , він забезпечує взаємодію їх носіїв зі специфічними білковими молекулами на плазматичній мембрані нейрона. В результаті цих взаємодій її проникність змінюється і виникають іонні потоки призводять до деполяризації мембрани , реєстрованої як постійно наростаючий рецепторний потенціал. Останній сприяє утворенню імпульсів збудження і їх поширенню у відповідні нервові центри. Таким чином , рецептор трансформує енергію дратівної стимулу в нервові імпульси , частоти яких відповідають силі роздратування.
Малоклітинна і мініатюрні рецептори комах відрізняються винятковою чутливістю і стійкістю до перешкод , надійністю і економічністю роботи . За багатьма параметрами вони перевершують вимірювальні прилади високої точності. Так , одні з них , сприймаючи леткі сполуки в зникаюче малих концентраціях , служать « лічильниками » окремих молекул , а інші реєструють поодинокі фотони або коливання субстрату з амплітудою , рівною радіусу атома водню.
Для комах характерні первинні рецептори , нейрони яких не тільки сприймають енергію стимулу , а й самі генерують і передають нервові імпульси. У цьому відношенні вони протиставляються вторинним рецепторам хребетних тварин , утвореним двома нейронами : сприймачем роздратування і передавальним виникає в нім збудження через синаптичну щілину на нейрон , що генерує імпульси в центральну нервову систему.
Після проведеної первинної обробки , кодування і передачі інформації її подальший аналіз , а потім і синтез цілісного образу подразника відбуваються в нервових центрах , які разом з органами почуттів і провідними шляхами об'єднуються в аналізатори .
У комах виявлено не менше дев'яти аналізаторів : зоровий , нюховий , смаковий , слуховий , тактильний , руховий , гравітаційний , вісцеральний і температурний . Є відомості про здатність комах реєструвати зміни вологості та електромагнітного поля , але відповідні нервові центри ще не досліджені. Тим часом перерахованого набору сприйманих відчуттів відповідають лише чотири класи рецепторів: механо- , термо- , хемо - і фоторецептори . Пояснення цьому невідповідності можна знайти в тому , що , наприклад , відмінності смакових і нюхових стимулів визначаються лише тим середовищем - рідкої або газоподібної , в якій сприймаються їх носії , а протиставлення тактильних , рухових , гравітаційних відчуттів або відчуттів звукових хвиль і стану внутрішніх органів ( висцеральное почуття ) зводяться до відмінностей в механічних подразненнях сенсорного нейрона.
Механорецепція .
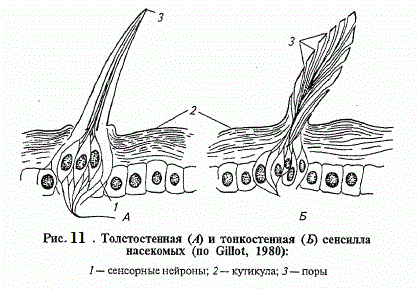
Опис механорецепторов доречно почати з найпростішого з них - осязательного волоска , іменованого тріхоід -ної сенсілли (рис. 119). Розташований на покривах , в сочленовной резіліновой мембрані , він пов'язаний з дендритом сенсорного нейрона , закінчення якого (перетворена вія ) одягнене кутику - лярні чехликом - сколопсом . Цей волосок настільки пружний, що будь-яке його відхилення відразу передається мембрані і , деформуючи сколопс , генерує рецепторний потенціал. Зазвичай сенсілли такого типу групуються в місцях найбільш частих контактів з механічними подразниками і диференціюються як органи дотику і ветрочувствітельние волоски. Останні, як правило , більш тонкі й рухливі .
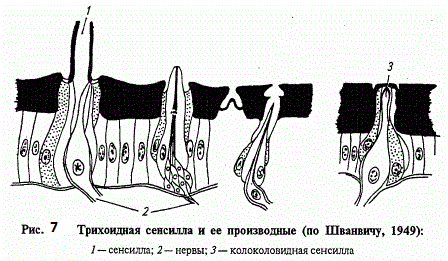
При стимуляції тріхоідной сенсілли поступово наростаючий рецепторний потенціал досягає порогового значення і збуджує незгасаючі нервові імпульси. Їх частоти визначаються силою впливу на рецептор. За сприйняттям впливу тонічним сенсілли протиставляються фазові . При сприйнятті фазовими сенсілли високі значення рецепторного потенціалу та частот відзначаються тільки на початку впливу і при тривалому подразненні придушуються , при сприйнятті тонічними - зберігаються незмінними весь час , поки діє подразник. Поряд з ними виділяють фазотоніческіе сенсілли , що реагують на початку роздратування сплеском нервових імпульсів , частоти яких , однак , різко знижуються при тривалій дії подразника. Таким чином , тонічні сенсілли передають у нервові центри аналізатора інформацію про силу і тривалості подразнення , фазові - про його початок , а фазотоніческіе об'єднують властивості тих і інших.
Зазвичай сенсілли спеціалізуються на певному режимі сприйняття стимулу , але в яйцеклада деяких мух присутні всі три їх типу. Мабуть , це дозволяє їм відрізняти короткочасні і випадкові зіткнення з субстратом при русі від більш тривалих контактів.
Особливо густі скупчення фазотоніческіх тріхоідних сенсілли ( волоскові пластинки) в зчленуваннях і на кордонах сегментів реєструють взаємні зміщення і положення останніх. Будучи пропріоцептори ( рецепторами власного становища ) , ці сенсілли нерідко набувають значення рецепторів гравітації і , сприймаючи орієнтацію тіла в гравітаційному полі , сприяють корекції його положення при польоті і плаванні .
Незважаючи на простоту будови , тріхоідние сенсілли беруть участь у рецепції досить різноманітних механічних стимулів . У цьому відношенні вони протиставляються кампаніформним ( дзвоноподібних ) сенсілли , що спеціалізуються як пропріоцептори пружних деформацій кутикули при м'язових скороченнях . Морфологічно вони відрізняються від тріхоідних сенсілли тільки тим , що позбавлені волоска , перетвореного тут в невисокий овальний ковпачок. Розташовуючись по лініях деформацій , вони передають в фазотоніческом режимі сигнали про силу і спрямованості згинань кутикули.
У ще більшій мірою спеціалізовані хордотональние сенсілли - сколопідіі , які поряд з функціями пропріоцепто - рів нерідко набувають здатність до реєстрації звуків. Дендрит сенсорного нейрона сколопідія утворює тонку , розширену посередині вію. Оточена кутикулярним сколопсом , виділеним обкладочной кліткою , вона простягається за його межі в центральний канал дзвоноподібних шапочки ( рис. 120 ) , до якої примикає шапочковая клітина. Численні тонофібрілли цієї клітини сприяють прикріпленню всього комплексу до епікутікуле покривів і при їх розтягуванні переводять роздратування на вію дендрита . Зазвичай сколопідіі групуються в більш-менш складні хордотональние органи. Натягнуті , як струни , вони реєструють найменші деформації і нерідко диференціюються на відносно дрібні тонічні і більші фазові і фазотоніческіе елементи.

Про конкретне призначення механорецепторов можна судити по наступних прикладів . Богомол Mantis religiosa L. , помітивши жертву , залишається нерухомим і лише злегка повертає слідом їй голову. Дратуючи при цьому волоскові пластинки в шийному зчленуванні , він легко перебудовує напрямок кидка. Однак вимикання цих рецепторів , не перешкоджаючи стеженню , скорочує точність кидків ( прицільність ) у кілька разів. Ще більш наочний тарзальной рефлекс , властивий багатьом комахою. Він полягає в наступному. Відрив лапок комахи від опори відразу викликає політ , який припиняється настільки ж швидко при поновленні контакту з субстратом.
Разом з тим для підтримки польоту необхідна стимуляція ветрочувствітельних сенсілли на голові і крилах , а для його контролю - пропріоцепторів антен: чим сильніше на них тиск зустрічних потоків повітря , тим менше амплітуда помахів крил. Не менш важливу роль відіграють механорецептори в підтримці рівноваги та реєстрації положення тіла в полі гравітації. Зіставляючи роздратування від різних частин тіла , відповідні аналізатори дуже тонко відрізняють найменші (до 2е ) відхилення від горизонталі. При цьому у водних форм роль провідників гравітаційних стимулів нерідко виконують бульбашки повітря. Прагнучи до поверхні і чинячи тиск на розташовані над ними групи сенсілли , вони сприяють також збереженню рівноваги і реєстрації глибини занурення під воду.
Всі ці настільки різноманітні функції здійснюються дотикальними волосками і розвиваються на їх основі більш складними кампаніформнимі і хордотональнимі сенсілли . Подальша спеціалізація останніх як віброрецептори призводить до оформлення вельми чутливих до щонайменших струсів субстрату підколінних органів коників і розташованих в антенах джонстонових органів. Виконуючи роль пропріоцепторів , вони одночасно набувають здатність до сприйняття звуків. На цій підставі їх можна протиставити всім раніше розглянутим контактним механорецепторів , що сприймає механічні стимули лише при безпосередньому контакті з їх носіями як рецептори дистантного дії , що працюють на відстані від джерела стимулу. Тому ми розглянемо їх разом зі спеціалізованими органами слуху , а зараз зупинимося на рецепторах розтягування.
На відміну від інших біполярних сенсорних нейронів мультиполярні нейрони рецепторів розтягування розпластуються своїми численними дендритами по поверхні внутрішніх органів і в порожнині тіла. Реєструючи натяг м'язів , вони генерують імпульси , частота яких зростає при посиленні та прискоренні розтягування. При цьому у відповідні аналізатори передається інформація про силу і швидкості скорочення м'язів. Поряд з реєстрацією стану мускулатури рецептори такого роду повідомляють в нервові центри про перистальтиці кишечника , битті спинного судини , рухах внутрішніх органів і активності вентиляції трахей .
Сприйняття і генерація звуків.
Здатність до сприйняття і генерації звукових хвиль у різних видів комах далеко не однакова. Не обмежуючись сприйняттям сигналів , комахи відповідають своєму партнеру , але набагато частіше безмовно реагують на них специфічними актами поведінки .
Про біологічному значенні акустичної комунікації можна судити по набору виявлених сигналів , серед яких розрізняються заклики , звернені до самок , і їхні відповіді , шлюбні пісні спаровуються особин , звукові сигнали самців , що охороняють обрану територію , і сигнали агресії , попередні поєдинків , сигнали протесту , лиха і тривоги.
Найбільшу акустичну обдарованість виявляють прямокрилі , особливо самці цвіркунів , які перевершують в цьому відношенні не тільки риб , амфібій і рептилій , а й багатьох співочих птахів . Загальне число сигналів , пов'язаних зі специфічними актами поведінки , досягає у комах 13 ( в окремих видів - 7).
Комахи володіють різними засобами і способами виробництва звуків. Вельми звичайні для них фрикційні органи , принцип роботи яких заснований на терті гострого кушкулярного канта про зазубрену поверхню. Наприклад , у коників і цвіркунів відповідні структури розташовані на передніх крилах : потираючи їх один про одного, вони видають характерне стрекотіння ( стрідуляцію ) . Схожий ефект досягається тертям про край крила внутрішньої поверхні стегон задніх ніг , яка в саранових несе дрібні шипики .
Властиві півчим цикад тімбальние органи мають жорсткі кутикулярний мембрани , які приводяться в дію сильними асинхронними м'язами. Вгібаясь і вигинаючись , як дно у жерстяної банки , вони генерують звуки , багаторазово підсилюється резонатором , функції якого виконує черевце , пусте у співаючих самців.
Звуки , що виникають при вібрації крил і стінок грудних сегментів , - побічний наслідок роботи крилових м'язів , але для багатьох двокрилих ці звуки набувають значення відоспеціфічен -них сигналів і акустичних орієнтирів.
Засмоктуючи повітря в кишечник і з силою виштовхуючи його назовні , бражник мертва голова ( Acherontia atropos L. ) шипить , як потривожена змія . Деякі таргани , перетинчастокрилі і двокрилі шиплять , випускаючи повітря через дихальца . Багато жуки- точильники закликають самок стуком голови про стінки прокладають у мертвій деревині ходів. Ці звуки , що нагадують тихе цокання годинника ( «годинник смерті» ) , нерідкі в старих дерев'яних будівлях . Подібним чином попереджають один одного про небезпеку терміти . Голодні личинки деяких ос, вимагаючи їжі , скрегочуть щелепами про краю осередків гнізда. Багато совки видають ультразвуки , дізнаючись наближається кажана з відстані до 30 м або сприймають її власні ультразвукові посилки з ще більших відстаней .
Звуки , вироблені комахами , варіюють за частотою від 5 до 2 кГц , а їх сила у найбільш громкоголосих коників і цикад досягає 110-115 дБ. Нерідко вони складаються в пісні , утворені елементарними звуковими посилками - пульсами і клацаннями , триваючими зазвичай 10-30 мс , серіями пульсов і безперервними трелями . При цьому набори пульсов в серіях і трелях варіюють від 5 до 800 за 1 с.
Частотні характеристики генеруються звуків в основному визначаються механічними властивостями звукопроізводящіх органів , тоді як тимчасової малюнок пісень контролюється роботою нервово -м'язового апарату і центральною нервовою системою. Однак перш ніж судити про значення тих чи інших параметрів звукових сигналів для акустичної комунікації , необхідно ознайомитися з властивостями органів слуху комах. При цьому зауважимо , що здатність звукових хвиль поширюватися в повітрі , а також у водному і твердої середовищах не дозволяє провести різких кордонів між слухом і віброрецепціей . Очевидно , мова може йти лише про протиставлення дистантной і контактної рецепції механічних стимулів .
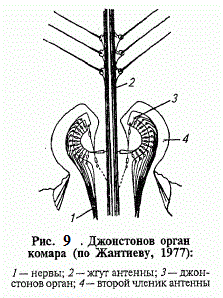
Один з найбільш складних хордотональних органів , здатних до сприйняття звукових хвиль в повітряному і водному середовищах , - Джонстон орган. Зазвичай він утворений групою з 1 - 3 - нейронних сколопідіев , які црікрепляются до сочленовной мембрані 2 -го і 3-го члеників антен. Сприймаючи коливання джгута , вони передають імпульси збудження безпосередньо в головний мозок. У мух - дрозофіл цей орган налаштований на видоспецифічі сигнали , породжувані вібрацією крил самців , а у жуків - вертячек поряд з функціями віброрецептори він реєструє викривлення поверхневої плівки води ( меніски ) на відстані до 15 мм від перепони.
Особливо складний і своєрідний Джонстон орган у комарів (рис. 121). Його основу складають тисячі нейронів , які групуються концентричними шарами біля основи джгута антен. Сприймаючи найменші вібрації і відхилення антен , вони генерують імпульси в фазовому і тоническом режимах. Якщо при цьому фронт звукових хвиль паралельний осі антени , то покривають її волоски починають вібрувати і , розгойдуючи джгут антени , викликають максимальне роздратування рецепторів. Відтворюючи за допомогою звукових генераторів специфічні для комарів сигнали , в деяких випадках вдається залучити цих надокучливих комах до засмоктуючим пасткам .
Спеціалізовані органи слуху комах забезпечені тимпанальной мембранами , реєструючими звуковий тиск і його градієнт . Зазначені в прямокрилих , цикад , деяких клопів , сетчатокрилих і метеликів , вони розрізняються становищем , набором рецепторів і допоміжними структурами. У коників , цвіркунів і медведок тімпанальние органи розташовані на гомілках передніх ніг і відкриваються назовні вузькими віконцями . Основу цих органів складають розмежовані стволами трахей дві тімпанальние мембрани і примикають до них групи сколопідіев підколінного органу , проміжного відділу і слухового гребеня.
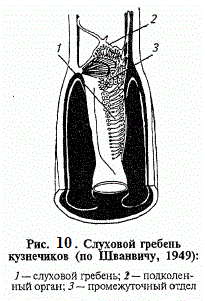
Підколінний орган , утворений віялом сколопідіев , служить високочутливим віброрецептори . Нечисленні сенсілли проміжного відділу , натягнуті між кутикулою й трахеями , ймовірно , виконують функції пропріоцепторів . Розташовані в один ряд сколопідіі слухового гребеня своєрідні в двох відношеннях: по-перше , вони примикають до трахеї і не скріплюються шапочковимі клітинами з кутикулою , по-друге , поступово зменшуються в розмірах , починаючи з самих великих проксимальних і кінчаючи дрібними дистальними . Не маючи контакту з тимпанальной мембраною , вони сприймають коливання , що передаються нею на трахейні стовбури .
У саранових дендрити сенсорних нейронів впритул прилягають до мембрани і порушуються її коливаннями безпосередньо . На відміну від органів коників тімпанальние органи саранових , а також метеликів і цикад розташовані в першому сегменті черевця або в заднегруди . У деяких боржників органами слуху служать заповнені трахейними мішками членики нижньогубні Щупика , дотичні з рудиментом верхньої губи - піліфером , а у сетчатокрилих і метеликів - бархатніца - заповнені гемолімфою здуття при підставах радіальних жилок крила.
Зазвичай в тімпанальние органи комах входять десятки , рідше - сотні нейронів , але у цикад їх число досягає 1500 . Реагуючи на роздратування , вони генерують залпи імпульсів , частоти яких незабаром скорочуються і стабілізуються на деякому рівні . Володіючи порівняно невисокою гостротою слуху ( 7-40 дБ) , вони разом з тим здатні розрізняти звукові посилки , наступні один за одним з частотою 100-300 на секунду. Незважаючи на рекордне число нейронів , тімпанальние органи цикад налаштовані лише на певний діапазон частот , відповідний максимальної енергії в спектрі призовних пісень самців. У саранових виявлено чотири типи різних нейронів , налаштованих на сприйняття низьких і високих частот. У здатних генерувати і сприймати ультразвук метеликів- совок виявлено три нейрона , причому тільки два з них збуджуються ультразвуком , а третій , навпаки , гальмує імпульсну активність. Ймовірно , це гальмування підвищує «пильність » всього органу до сприйняття ультразвукових посилок кажанів.
Таким чином , тімпанальние органи комах сприймають в першу чергу не частотні , а часові параметри звукових стимулів , передаючи в вищі нервові центри інформацію про тривалості імпульсів і про характер їх повторень , тобто тимчасової малюнок пісень.
Перетворена сенсорними нейронами інформація про характер звукових стимулів надходить в нервові центри грудних гангліїв , де піддається подальшій обробці. Розташовані тут асоціативні нейрони , працюючи в різних режимах , відповідають на одні й ті ж сигнали по- різному. Фазові нейрони реєструють початок пісень і на кожен сигнал генерують по 1-2 імпульсу , підкреслюючи ритмічну організованість звукових посилок. Одночасно тонічні нейрони реєструють тривалість окремих сигналів.
Таким чином , інтенсивність імпульсів , що продукуються сенсорними нейронами в залежності від інтенсивності подразника , визначає частоту імпульсації асоціативних нейронів і разом з тим загальна кількість реагуючих елементів в нервовому центрі . Деякі нейрони при підвищенні інтенсивності опосередкованих сигналів гальмуються і вимикаються , інші - набирають обробку акустичної інформації. У сарани Locusta migratoria L. відповідні нейрони можуть працювати або в фазовому , або в тонічному режимі. При звуках середньої і низької інтенсивності вони дають фазові відповіді , при звуках високої інтенсивності - тонічні . Це дозволяє розрізняти ритмічні посилки й обумовлює високу чутливість аналізатора до змін інтенсивності шумових « фонових » звуків.
Очевидно , що на рівні нервових гангліїв контрастируются тимчасові візерунки звукових стимулів за інтенсивністю , частоті повторення посилок і серій , спрямованості. Мабуть , саме тут відбуваються розрізнення і впізнання їх видовий специфічності. Одночасно йде фільтрація сторонніх шумів і перешкод. Зрештою асоціативні нейрони грудних гангліїв передають повідомлення в вищі нервові центри слухового аналізатора , розташованого в надглоточнимі ганглії .
Отже , обробка сприйнятої акустичної інформації проходить щонайменше в три етапи: на рівні сенсорних нейронів рецептора , на рівні асоціативних нейронів грудних гангліїв і , нарешті , в головному мозку , де тривають впізнання і локалізація джерела звуку і формується цілісний образ подразника.
У різноголосому хорі комах , птахів і амфібій , серед шелесту листя і трав самки коників роду Isophya не чують пісень цвіркунів , але відповідають на кожну звукову посилку самців свого виду одним - двома характерними клацаннями . Нерідко до виконуючого соло самцеві приєднуються інші , підпорядковуючи його пісням ритмічний малюнок своїх трелей .
Терморецепція і гігрорецепція .
Впливаючи на метаболічні процеси , осмотичні властивості цитоплазми , властивості мембран і розчинів , зміни температури і вологості могли б реєструватися організмом безпосередньо як зміни всіх цих процесів і властивостей. Більше того, проведення імпульсів збудження по нервах і синапсах і спонтанна активність нейронів також схильні до впливів температури і вологості , але не як рецепторних стимулів , а як екологічних факторів , порівнянних за значенням і характером дії з впливом корму , статевого партнера або хижака.
Навряд чи є сумніви в тому , що моментальна реакція на зміни температури і вологості в порівнянні з їх реєстрацією через зміни в метаболічних процесах біологічно доцільна. Іншими словами , що ввійшов у вогонь чи у воду здатність лише до опосередкованого сприйняття такого роду факторів могла б обійтися надто дорого.
Разом з тим не виключені можливі впливу вологості та температури на рецепторні мембрани сенсорних нейронів механо- або хеморецепторів . У всякому разі , комахи мають спеціалізовані термо- і гігрорецептори .
Тріхоідние сенсілли на антенах таргана Periplaneta americana L. реагують п'ятикратним збільшенням частоти імпульсів на зниження температури на 1 ° С. Дресировані бджоли розрізняють температуру відвідуваних субстратів з точністю до 2 ° С. Їх фазотоніческіе рецептори мають вигляд коротких волосків , занурених у кугі - кулу ( целоконіческіе сенсілли ) , і збуджуються при зниженні температури. Однак подібні сенсілли на антенах хлібного жука- кузьки Anisoplia austriaca L. генерують фазові залпи імпульсів лише з підвищенням температури, як , втім , і терморецептори клопа Oncopeltus fasciatus L. На цій підставі їх іменують тепловими на відміну від холодових рецепторів бджоли і таргана.
Особливою чутливістю відрізняються целоконіческіе сенсілли на кінчиках антен комара Aedes aegypti L. Маючи в своєму розпорядженні нейронами обох типів , вони генерують фазові відповіді при зміні температури на 0,05 ° С; при цьому збудження теплового нейрона гальмує активність холодового і навпаки. Однак вони не реагують на джерело інфрачервоного випромінювання , сприймаючи лише конвекційне тепло - енергію молекул нагрітого повітря.
Видалення антен у багатьох комах і реєстрація потенціалів дії (ПД) нейронів деяких лускокрилих дозволяють визнати рецепторами вологості тонкостінні базіконіческіе сенсілли . Останні відрізняються від тріхоідних сенсілли тим , що мають вигляд коротких конусів із закругленими вершинами. У відповідь на струмінь вологого повітря вони генерують імпульси , які, однак , гальмуються сухим повітрям. Подібні реакції відзначені в деяких нейронах механорецепторов кровосисних клопів пологів Triatorna і Rhodnius , але диференціювати їх від можливих впливів вологості на механічні (фізичні) властивості допоміжних структур не представляється можливим.
Хеморецепція .
Сприйняття хімічних стимулів припускає умовне протиставлення дистантної і контактної хеморецепции . У першому випадку стимуляторами служать леткі сполуки , в другому - розчини цих сполук.
В обох випадках джерелом стимулу служить потенційна енергія взаємного притягання і відштовхування елементарних частинок на рецепторних мембранах . Разом з тим очевидні відмінності в абсорбції газів і рідин визначають своєрідність допоміжних структур , обслуговуючих товстостінні смакові та тонкостінні нюхові сенсіли. Нерідко і ті й інші мають вигляд тріхоідних , базіконіческіх і стілоконіческіх сенсілли , але перші , як правило , несуть одну вершинну пору, другі - безліч пір по всій поверхні рецептора.
Контактні хеморецептори і смаковий аналізатор . Товстостінні смакові сенсілли найчастіше зосереджені на ротових придатках і на лапках комах , рідше - на їх антенах і яйцеклада . Зазвичай їм супроводжують 3-5 ( рідше - до 50) нейронів з дендритами , що досягають вершинної пори , яка здатна замикатися при механічних подразненнях .
Незважаючи на порівняно невелике число сенсорних елементів , смакові реакції і переваги комах далеко не однозначні. Наприклад , в трехнейронной лабеллярних сенсілли мухи Phormia regina Mg . кожен нейрон виконує певні і специфічні функції : один з них служить Механорецептори , а два інших диференційовані на сольовий ( L- нейрон ) і цукровий ( S- нейрон) . У складніших четирехнейронних сенсілли поряд з ними присутній W- нейрон , дратуємо дистильованою водою.
Стимуляція S- нейрона провокує позитивну харчову реакцію , засновану на лабеллярном рефлексі - розгортанні хоботка при контактах смакових рецепторів лапки із привабливим харчовим субстратом. При подразненні L- нейрона харчова реакція гальмується і комаха прагне покинути субстрат. При цьому частота генеруючих імпульсів пропорційна силі впливу , тобто концентрації розчинів Сахаров або солей.
Відрізняючись вельми низькою спонтанною активністю (1-4 імпульсу в секунду) , обидва нейрона володіють гальмують впливами : подразнення сольового рецептора гальмують активність цукрового і навпаки.
Таким чином , нейрони контактних хеморецепторов комах дозволяють розрізняти привертають і відкидаємо речовини і більш дрібно диференціювати їх за концентраціями і властивостями .
Інформація, що надходить в нервові центри аналізатора , піддається подальшій обробці, і виникають на її основі смакові реакції вельми різноманітні. Комахи диференціюють чотири первинних смакових якості , розрізняючи солодке , солоне , кисле та гірке , не поступаючись людині по чутливості до них. Наприклад , людина розрізняє цукор в концентрації 2-10 ~ 2 моль / л , а бджоли і метелик- адмірал ( Pyrameis atalanta L. ) - в концентрації б- 10 ~ 2 і Ю - 1 - 10 ~ 2 моль / л відповідно.
Однак при тривалому утриманні від Сахаров метелик- адмірал здатна знизити поріг чутливості до 8-10 ~ 5 моль / л. Комахи відрізняються своєрідністю смаків. Наприклад , позбавлена смаку для бджіл лактоза виявилася помірно солодкою для гусениць метеликів , що сприймають з їжею істотно меншу кількість Сахаров.
Відносно солей комахою властиві позитивні харчові реакції лише на малі концентрації , але найбільш « солені » для них катіони К, найменш - катіони Li .
Пороги чутливості бджіл до кислих речовинам – 10-3 моль / л. Відносно нестерпно гірких розчинів хініну , ацетілсахарози і настоїв полину комахи малочутливі .
Комахи сприймають смак дистильованої води , яка для людини позбавлена смаку і нерідко вони виявляють дивну пристрасть до отрут . Наприклад , живиться на звіробої листоед Chrysolina sp . має спеціальний рецептор , порушуваний тільки характерним для цієї рослини алкалоїдом - гіперізідом .

Дистантних хеморецептори і нюховий аналізатор . Дистантних хеморецептори , як правило , зосереджені на антенах ; зазвичай їх більше, ніж контактних (до 60 ТОВ у самців метелики Antheraea polyphemus L. ) , але все ж комахи значно поступаються за цим показником ссавцям. Наприклад , у кролика є до 10 ольфакторних нейронів.
Серед дистантних хеморецепторов комах нерідкі тріхоідние , базіконіческіе , целоконіческіе , а також плакоидной сенсілли . У одних видів вони розсіяні по всій поверхні антен , у інших - об'єднані групами під покривами в складні органи нюху.
Нюхові сенсілли зазвичай відрізняються численними порами , заповненими 10-20 трубочками , з якими пов'язані розгалужені відростки дендритів . Призначення цієї поровотрубчатой системи, мабуть , полягає в активній абсорбції та транспортуванні молекул ольфакторних подразників до рецепторних мембран .
Поки залишається неясним , яким чином деякі молекули летючих сполук ( феромонів ) , потрапляючи на антену , досягають рецепторів і вступають з ними у взаємодію . Неясний і характер цієї взаємодії. За вельми популярної свого часу стереохимической теорії нюху володіють певною просторовою конфігурацією молекули осідають на рецепторну мембрану , в якій є відповідні цій конфігурації ділянки - лунки. На зміну цій оригінальній , але погано узгоджується з фактами гіпотезі була висунута вібраційна теорія, що пов'язує здатність розрізняти запахи із сприйняттям коливань молекул , які реєструються за спектрами поглинання в інфрачервоній області. Однак при заміщенні водню тритієм в молекулах особливо привабливих речовин вони як і раніше охоче сприймалися комахами , незважаючи на різку зміну спектрів поглинання.
Тонкостінні нюхові сенсілли зазвичай містять по кілька нейронів ; зрідка їх число досягає 40-60 ; при цьому вони спеціалізуються на сприйнятті різних хімічних сполук.
У відповідь на нюхові стимули нейрони генерують імпульси , частоти яких залежать не тільки від концентрації речовин , але і від їх привабливості для комах.
Зазвичай при підвищенні концентрації залучають з'єднань частоти імпульсів збільшуються , а зростання концентрацій відкидала речовин призводить до її зниження .
Таким чином , на рівні дистантних хеморецепторів відбувається первинна диференціація подразників , чому неабиякою мірою сприяють два типи сенсорних нейронів : генералісти і фахівці. Перші з них відповідають на широкий набір різних летючих сполук , але малочутливі , другий реагують на одне або кілька близьких сполук і відрізняються підвищеною чутливістю. Наприклад , у жука - мертвоїди Necwphorus sp . нейрони цього типу забезпечують реєстрацію трупного запаху з відстані до 90 м , у самців шовкопряда Bombyx mori L. вони дозволяють розрізняти статевий аттрактант самок - бомбікол в концентрації 100 молекул на 1 см3. При цьому для роздратування одного нейрона -спеціаліста достатньо однієї молекули , а для генерації порушення - не менше 1 молекули в секунду в розрахунку на поверхню обох антен . За образним зауваженням Ж. А. Фабра , можливість рецепції самцем грушевої сатурнії запаху самки з відстані 5-10 км (рекордний випадок) можна порівняти зі спробою забарвити води озера однією краплею барвника карміну .
Настільки фантастична чутливість комах до специфічних запахів передбачає дуже строгу диференціацію ольфакторних стимулів . Аксони відповідних сенсорних нейронів переводять нервові імпульси в нюхові частки Дейтоцеребрум безпосередньо , без синаптичних перемикань. Мабуть , існування ізольованих шляхів для проведення збудження від нейрона -спеціаліста до асоціативних центрам відповідає особливому біологічному призначенням ольфакторних стимулів .
Тим часом по порогу чутливості до деяких запахів комах можна порівняти з людиною. Так , муха Phormia regina Mg . пари аміаку відчуває в концентрації 2-10 ~ б моль / л і бджола сприймає огидно пахне скатол в концентрації 2,3 * 10-8 моль / л. З запропонованого бджолі набору 1816 парних поєднань запахів вона при відповідній дресируванню з успіхом розрізняє майже всі з них ( 95,5 %).
Таким чином , комахи розрізняють найрізноманітніші запахи , серед яких одні належать улубленних корму , інші - субстратам для відкладання яєць , треті - орієнтирам гнізд або сім'ї . Серед них можна виділити запахи слідів фуражира термітів або мурашок , тривоги , потенційного статевого партнера і видавані бджолиної маткою для гальмування розвитку яєчників у робочих особин.
Фоторецепція .
В основі светоразліченія лежить реєстрація сенсорним нейроном фотоизомеризации зорового пігменту - родопсину . Під впливом поглиненого кванта світла він перетвориться в ізомер - метародопсін , який , відновлюючись в родопсин , забезпечує багаторазове повторення елементарних зорових актів.
Для сприйняття зорових стимулів комахи розташовують досконалими фоторецепторами - омматідія , згрупованими в складні фасеточні очі , дорзального очками і властивими личинкам Стемма . У омматідіі поряд з 4-12 уніполярними нейронами , званими тут ретінулярной клітинами , виділяються допоміжні структури рецептора - светопреломляющій апарат і апарат пігментного ізоляції. Перший з них утворений шестигранною кугікулярной лінзою рогівки , або кришталиком ( фасеткою ) , і лежачими під нею чотирма земперовимі клітинами , що формують кристалічний конус. Зовні кожна фасетка рогівки покрита мережею дрібних горбків. Знижуючи віддзеркалення світла і тим самим підвищуючи прозорість кришталика , ця найтонша мережу , видима лише під електронним мікроскопом , надає йому властивості просвітленої оптики і усуває блиск поверхні ока . Клітини кристалічного конуса проводять в глиб омматідія тонкі нитки світловодів , що надає светопреломляющие апарату очей комах гідності сучасних оптичних приладів.
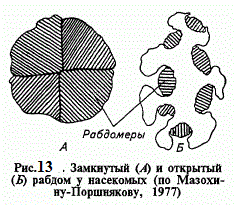
Сфальцьовані збиральними лінзами і проходять по световодам промені світла досягають розташованих в глибині ретина - лярних клітин. Групуючись навколо оптичної осі, як часточки апельсина , ці світлочутливі нейрони утворюють спрямовані до центру мікровілли - найтонші мікротрубочки , що складаються в рабдомер . Саме тут відбуваються елементарні акти світло- розрізнення. У мікровілли кожного рабдомера , тобто оптично активної частини світлочутливого нейрона , міститься до 107 молекул зорового пігменту. Рабдомери кожної ретинулярной клітини або об'єднуються один з одним у замкнутий рабдом , або , наприклад у мух , зберігаючи відособленість , шикуються по стінках центральної порожнини відкритого рабдома.
До складу апарату пігментного ізоляції омматідія входять дві корнеагенние ( формують кришталик ) клітини , не менше чотирьох ірисових клітин і часто ще кілька ретінальних пігментних клітин. У їх цитоплазмі розсіяні темні червонуваті гранули оммохроми , поглинаючі майже всі промені видимого світла , крім червоних , і світліші гранули птерінов , поглинаючі ультрафіолетові промені. Утворюючи світлонепроникний чохол навколо світлочутливих елементів очі у денних комах , ці клітини у сутінкових і нічних форм здатні концентрувати гранули пігменту поблизу кришталика , відкриваючи рабдом ретінулярних клітин для засвічення бічними променями сусідніх омматідієв . Скорочуючи гостроту сприйняття , це явище ( ретіномоторной реакція) багаторазово збільшує світлосилу очі. При низькій освітленості , у сутінках , таке підвищення світлосили дуже важливо. Проте якість зображення, що створюється оптичними структурами , не має великого значення , так як розглядаємим комахами об'єкт сприймається кожним омматідія окремо , як мозаїка плям різної яскравості.
Відповідно до зазначених відмінностями властивостей і набору пігментних клітин можна протиставити аппозиційного очам денних форм суперпозіціонного сутінкових і нічних комах.
Розрізняючись , крім того , довжиною рабдоме , ці два типи очей припускають або додавання ( аппозиція ) сприйманого зображення з окремих вловлюються омматідія плям , або накладення ( суперпозицію ) цих плям один на одного. Здатність пігментних клітин суперпозіціонного омматідія розосереджуватися по всій довжині рабдома приводить його в функціонально аппозиційного стан , що відбувається всякий раз при надлишку освітлення днем . Розосередження пігменту завершується в лічені хвилини , тоді як для пристосування очі до нічного бачення потрібно від 10 до 50 хв .
У двокрилих , що відрізняються відкритим рабдоме , орієнтація окремих рабдомеров омматідія така, що всі вони «дивляться » в різні точки простору , однак ті ж самі точки сприймаються шістьма рабдомерами , що належать різним омматідія . Іннер - вірующіе їх волокна сходяться в одному нервовому центрі , тому сигнали ретінулярних клітин різних омматі - Дієв підсумовуються в оптичних частках протоцеребрума . Таким чином , зберігаючи переваги аппозиційного бачення , нейросуперпозіціонние очі мух мають підвищену світлосилою . Однак у порівнянні з суперпозіціонного очима нічних метеликів , світлосила яких перевершує аналогічний показник аппозіціонних очей в десятки і сотні тисяч разів , нейросуперпозіціонние очі набагато менш чутливі в сутінках або вночі.
Очі денних форм комах також здатні пристосовуватися до сутінкового висвітлення , але тільки за рахунок порушення рівноваги фотоизомеризации зорового пігменту і зміни коефіцієнтів заломлення ретінулярних клітин.
При поглинанні рабдомерами фотона світла вони генерують мініатюрні імпульси , мабуть , пов'язані з виділенням медіаторів в ретінулярной клітини , які у відповідь на це генерують постійно наростаючий потенціал деполяризації мембрани. Такого роду безимпульсние сигнали характерні для світлочутливих елементів , що відрізняє їх від сенсорних нейронів інших типів. Досягаючи оптичних часткою протоцеребрума , ці сигнали перемикаються на асоціативні нейрони , що зв'язують один з одним різні гангліозні маси. Тут виділяються чотири відділи і нервові волокна більшості омматідієв схрещуються в оптичній платівці .
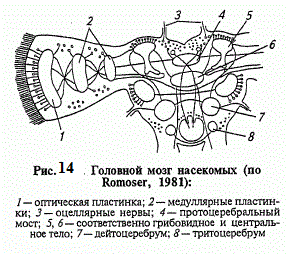
При замкнутому рабдоме волокна об'єднують аксони ретінулярних клітин одного омматідія , при відкритому рабдоме - аксони різних омматідієв , рабдомери яких мають збігається орієнтацію . Разом з тим окремі сенсорні нейрони передають сприйняту інформацію відразу в медулярний пластинку , пов'язану з оптичною платівкою нервовими волокнами зорового перехрещення. Подальші шляхи проведення нервового збудження ще не досліджені , але оптичні волокна відзначені і в центральних областях протоцеребрума , і в стебельчатих тілах.
Найважливіші робочі елементи зорових центрів - спеціалізовані нейрони , які представляють собою детектори , вибірково реагують на параметри зорових стимулів : їх розміри , форму , забарвлення і характер руху. Нерідко виділяються тільки ті з них , які рухаються в строго визначеному , улюбленому для сприйняття напрямку. Рухомі інакше гальмують імпульсну активність і , мабуть , не сприймаються комахами.
Найбільш прості детектори - нейрони тривоги , що реагують на затінення частини ока і будь-які переміщення об'єктів у полі зору. Генеруються ними імпульси зумовлюють включення рефлекторної реакції втечі . Дещо складніше організовані детектори ознак статевого партнера або спрямованості рухів , зокрема детектор рухається плями у бабок , що включає реакцію переслідування.
Своєрідний рефлекторний механізм латерального гальмування , що виявляється в тому , що активність омматідієв залежить не тільки від сили стимулу , а й від активності сусідніх омматідієв . Посилюючи перепади в освітленості споглядаємо об'єктів , ці механізми пригнічують оптично шкідливі наслідки перекривання полів зору сусідніми омматідія , не перешкоджаючи , однак , стереоскопічності бачення. Відзначимо , що подібні принципи реалізовані в декодуючих пристроях реставраторів і криміналістів , використовуваних для розшифровки нечітких фотографій.
Розподіл омматідієв по сфері складного очі і їх чисельність , що досягає 12000-17000 у метеликів і 10000-28000 у бабок , надає зору комах деякі особливі властивості .
Очевидно , що чим ближче зведені оптичні осі омматідієв , чим менше між ними кут , тим вище гострота зору - здатність до розрізнення двох зближених між собою точок . У комах вона в 3 рази вище , ніж у людини (30 і 100 мкм відповідно) , але разом з тим при видаленні об'єкту від ока гострота зору різко скорочується - комахи короткозорі.
Крім того , при найменших зсувах об'єкта його сприйняття відразу ж переходить до інших , сусідніх омматідія , повідомляючи комахою особливу чутливість до рухомих стимулам . Поряд з дуже малою інерційністю рецепторів це надає оку здатність до розрізнення швидко мигтючих предметів . Якщо для людини злите зображення на екрані виникає при зміні 16 кадрів в секунду , то для комахи - 250-300 . Цілком зрозуміло , що це властивість необхідно при швидкісному польоті серед рослин . Ні людина, ні комаха взагалі не бачать об'єктів , нерухомих відносно сітківки ока. Постійний тремор окорухових м'язів людини - одна з неодмінних умов бачення- протиставляється нерухомості очей комах , які для виконання цієї умови змушені вібрувати всім тілом. Зазвичай при величезному числі фасеток це не вимагає великих витрат енергії , але якщо їх кількість обмежена , як , наприклад , у гусениць метеликів , то необхідні більш інтенсивні рухи тіла. Бути може , саме цим пояснюється особливість гусениць відриватися час від часу від прийому їжі , підводячи і розгойдуючи передню частину тіла.
Подібним чином пояснюється розрізнення комахами форми об'єктів і перевагу ними тих , чий контур більш розсічений . Крім того , вже доведена стереоскопичность зору комах , що забезпечує сприйняття об'ємів тіл і глибини простору , а також здатність до сприйняття поляризації світла . Не тільки блакитне небо , а й поверхню водної гладі , і покритих восковим нальотом листя рослин , і покривів комах , поляризуючи світло , надають їм неразлічаемие людиною відблиски і тіні.
У ще більшій мірі збагачує зорові сприйняття комах здатність до колірного зору , доведена для багатьох з них. Вони не тільки можуть розрізняти спектральний склад світла , але і сприймають , як і людина , природне забарвлення предметів незалежно від кольору освітлення. Серед досліджених в цьому відношенні комах цветослепотой страждали лише печерні форми , терміти і великий борошняний хрущак { Tenebrio molitor L. ) .
Здатність до цветоразличению визначається щонайменше двома світлоприймачами з різною спектральною чутливістю. У людини їх три , у бджіл і багатьох денних метеликів - також три , у інших комах - зазвичай тільки два. Один з них , як правило , налаштований на сприйняття ультрафіолетових променів і володіє
підвищеною чутливістю , відрізняючи , наприклад , жовтий колір від жовтого з 1 % домішки ультрафіолетового світла . На малюнку 14 наведена схема різної спектральної чутливості очей людини , бджоли і мухи.
Відчуття, випробовувані людиною при сприйнятті різних стимулів , завжди суб'єктивні . Тому , якщо бджоли , наприклад , розрізняють жовтий і зелений кольори , ми не знаємо , як виглядають ці кольори для них , як вони відчуваються ними . Крім того , багато комахи володіють вродженою схильністю, наприклад, до синім і пурпурним (метелик -вітрильник Papillo troilus L. ) або жовтим кольорам ( мухи- журчалки роду Eristalis , багато попелиці ) і не відрізняють білий колір від суміші жовтого з синьо- фіолетовим.
Однак особливу своєрідність колірним сприйняттям надає ультрафіолетова забарвлення . Одноколірний для людини самець метелика - лимонниця Gonepteryx rhamni L. сприймається його самкою як двокольоровий, так як його передні крила сильно відбивають ультрафіолетові промені. Однотонно -білі капустяна білявка ( Pieris brassicae L. ) або метелик- бояришніца ( Aporia crataegi L. ) для комах досить різні за кольором і мають явні ознаки статевої приналежності.
Багато жовті або білі квіти розрізняються комахами за орієнтирами нектарників , а червоний мак для несприйнятливих до червоних променів мух « сяє » лише одним ультрафіолетовим « кольором ».
Поряд з омматідія складних очей в сприйнятті зорових стимулів беруть участь дорзальние оченята , а у позбавлених складних очей личинок вони заміщені Стемма . І ті й інші значно простіше по пристрою, але відрізняються деякими своєрідними функціями.
Светопреломляющая кутікулярная лінза дорзальних вічок відмежована шаром дрібних корнеагенних клітин від очного келиха , в якому зосереджені світлочутливі ретинальні клітини часто упереміж з пігментними . Відходять від вічок аксони сенсорних клітин досягають розташованих поруч оцеллярних гангліїв , переводячи сприйняту зорову інформацію до асоціативних нейронам .
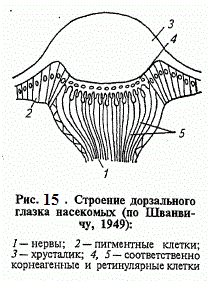
Таким чином , ганглії вічок знаходяться поза протоцеребрума , але тісно пов'язані з ним товстими оцеллярнимі нервами. Очки реєструють зміни освітленості і наступ темряви , розрізняють кольори і поляризацію світла , сприймають рух великих об'єктів і при цьому володіють підвищеною світлосилою . Крім того , надходять від них сигнали впливають на активність головного мозку , який отримує високу тонічне збудження навіть у темряві , регулюють спрямованість руху до джерела світла і ритміку биття спинного судини. Однак деякі вельми активні комахи позбавлені розвинених вічок.
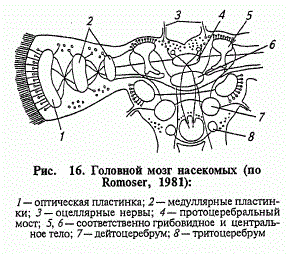
Личинкові Стемма різноманітні за будовою . Більш примітивні з них нагадують дорзальние оченята , досконаліші - окремі омматідіі . Відрізняючись високою світлосилою , вони здатні до цветоразличению і сприйняттю поляризації світла.
Узагальнення зорових образів. Про здатність комах до дресирування , запам'ятовування та узагальнення зорових образів можна судити за результатами експериментів , в яких використані умовах а- норефлекторние реакції на певні стимули.
Пропонуючи бджолам корм , розташований на пофарбованому столику , неважко вже через 2-3 прильоту привчити їх до певного кольору як до символу їжі. Формуванню умовного рефлексу на двокольорові стимули відповідає перевагу та інших двоцвітних поєднань , навіть якщо кожен з обраних кольорів раніше не використали при дресируванні .
Найбільш вражаючі демонстрації здібностей комах належать К. Фришу , що відкрив танець бджоли- розвідниці . Повернувшись у вулик , вона повідомляє членам своєї сім'ї про знайдений нею кормі і про те , де фуражирам його слід шукати. Пробігаючи деяку відстань по стінці вулика , бджола - розвідниця виляє черевцем і , згортаючи направо і наліво , описує кола . Орієнтація траєкторії прямого пробігу відносно напрямку сили тяжкості точно відповідає напрямку польоту при орієнтації за Сонцем , а частота виляння черевця - відстані до джерела корму. Якщо танець відбувається на горизонтальній площині камери з верхнім оглядовим віконцем , то орієнтація прямого пробігу відповідає напрямку пошуку , але тільки тоді , коли небо не затягнуте хмарами . В іншому випадку танець стає ненаправленим . Варто бджолі побачити хоча б шматочок блакитного неба в розриві хмар , танець знову стає орієнтованим.
Таким чином, поряд з розшифровкою сенсу танцю бджоли К. Фріш відкрив здатність комах до сприйняття поляризації світла і використання цієї здатності для наведення на ціль. По суті це ті ж способи орієнтації , якими користується людина при русі по азимуту . Очевидно , що компасом комах може бути Сонце чи напрямок поляризації променів світла блакитного неба .
Перетворення орієнтирів , в даному випадку кута напрямку на Сонце в вертикальний кут , утворений траєкторією прямого пробігу відносно сили гравітації , свідчить про більш глибокі здібностях комахи. Однак навіть ці здібності все ж недостатні для орієнтації багатогодинних нічних перельотів і міграцій на тисячі кілометрів. Виникаюча необхідність корекції напрямку польоту за астрономічними орієнтирам вимагає не тільки відчуття часу , але і синхронізації внутрішніх біологічних годин організму з добовим рухом Сонця на різних широтах і в різних часових поясах. Демонструючи якості досвідчених навігаторів , комахи справляються і з цими , далеко не простими завданнями. Можливо , вони реєструють геомагнітне поле , але для однозначної відповіді на ці питання потрібні більш глибокі дослідження .
Поведінка .
Сприйняті комахами стимули служать основою для формування доцільних актів поведінки як результату поєднання вроджених рефлексів , інстинктів і набутого досвіду.
Вроджені , або безумовні , рефлекси проявляються як однозначні відповіді на подразники. Приклади їх - описані раніше тарзальной і лабеллярний рефлекси , рефлекс рухається плями , що включає реакцію переслідування у бабок і кидок богомола . Серії безумовних рефлексів визначають стереотипи харчового і сексуальної поведінки комах , поведінки при виборі місць відкладання яєць і інстинкти при будівництві гнізд і будиночків , при завивці коконів і т. п.
Інстинкти і стереотипи поведінки являють собою ланцюги послідовних рефлексів , які виникають у відповідь на зміни фізіологічного стану організму. У цьому випадку їх спонукальними мотивами можуть бути і голодування , і дозрівання статевих продуктів або зміни в концентрації гормонів та інших фізіологічно активних речовин , відповідні досягнення певних фаз і стадій розвитку.
Подібну нейрофизиологическую природу мають таксиси і тропізми . Їх слід протиставити один одному як алгоритми наведення на ціль і способи орієнтації незалежно від природи ориентирующего стимулу ( таксиси ) і як власні відношення до цих стимулам , пов'язані з внутрішньою мотивацією їх вибору ( тропізми ) . Відповідно з набором стимулів і ставленням до них комах розрізняють позитивні і негативні фото- , гео- , хемо - , гигро - , термотропізми і т. д. , маючи на увазі позитивні і негативні реакції на джерела світла , гравітацію , джерела хімічних стимулів , вологості і тепла.
Механізми формування тропізмов досі залишаються неясними , але їх біологічний сенс часто цілком очевидний. Відомі численні приклади використання тропізмов комах (як і рефлексів , і інстинктів ) у практиці захисту від них рослин, зокрема при накладенні ловчих поясів на штамби плодових дерев (негативний геотропізм гусениць плодожерок ) , при знищенні рослинних залишків , в яких ховаються від світла багато шкідники , при організації вилову комах різними светоловушки і приманками з феромонами .
Що стосується таксисов , то найбільш явні з них - це фобо - , клино - , тропо - , тіло - і менотаксіс .
Фоботаксис , іноді званий методом проб і помилок , являє собою відносно примітивний , але і найбільш загальний алгоритм поведінки в екстремальних умовах , нерідко загрожують самому існуванню організму. Він проявляється в уповільненні або прискоренні рухів під впливом стимулу і в частоті змін напрямку руху. Наприклад , при підвищеній температурі багато комахи починають швидко бігати , постійно змінюючи напрямок руху , при цьому ймовірність виходу за межі зони небезпеки зростає. На основі фоботаксис можна пояснити формування скупчень комах в місцях із сприятливими для них режимами вологості , освітленості , температури.
Клинотаксис визначає цілеспрямованість руху комахи при послідовному виборі рівнів збудження відповідних рецепторів. Наприклад , личинки кімнатної мухи , що відрізняються негативним фототропізм , розгойдують передню частину тіла з розташованими на ній фоторецепторами , обираючи напрямок руху , при якому рецептори збуджуються мінімально .
Тропотаксіс являє собою більш досконалий алгоритм наведення на джерело стимулу , заснований на досягненні рівноваги в порушенні симетричних рецепторів. Наприклад , медоносна бджола , розставивши в сторони антени , легко сприймає пахучу приманку , розташовану в одному з колін Y -подібної камери. Якщо її антени закріпити на лобі в перехрещених стані , то вона завжди прямує в протилежне коліно камери на основі тропотаксіса . При порушенні симетрії рецепторів , зафарбовуючи , наприклад , одне око , ми змушуємо комаха до постійних Манежній ( круговим ) рухам навіть в паралельному пучку світла .
Телотаксіс передбачає наведення на стимул за рахунок того , що його сприймають рецептори , спрямовані вперед. При цьому навіть осліплене на одне око комаха зберігає вірну орієнтацію , не здійснюючи Манежній рухів. Мабуть , цілеспрямований кидок богомола грунтується на телотаксісе , прояви якого буває важко відрізнити від проявів тропотаксіса і менотаксіса .
Менотаксіс своєрідний у тому відношенні , що наведення за допомогою його на ціль передбачає фіксацію деякого кута між напрямком руху і ориентирующим стимулом. При цьому обрані орієнтири можуть бути несуттєвими для комахи , не завжди строго фіксованими по положенню щодо справжньої мети . Зокрема , залучені на світ метелики демонструють менотаксіс , який порівняти зі способами орієнтації по азимуту .
Таким чином , багато аспектів поведінки комах розшифровуються на основі вроджених , безусловнорефлекторного реакцій. Однак всупереч думку Ж. Фабра комахи не є рабами своїх інстинктів , свого роду автоматами , діючими за заздалегідь розробленою програмою . Наведені раніше приклади формування умовних рефлексів у комах , їх здібностей до узагальнення і дресируванню надають поведінці гнучкість , необхідну в мінливих умовах існування . Відомі приклади , коли формуються умовні рефлекси гальмують прояви інстинктів і тропізмів . Піддаючи тарганів дії слабких електричних розрядів всякий раз , коли з двох пропонованих їм камер - освітленій і затемненій - вони вибирають останню , можна незабаром придушити прояви негативного фототропізму і привчити їх до існування в незвичних умовах ( при освітленні) .
Отже , сенсорні системи комах , що розрізняються по складності організації допоміжних структур , служать провідниками інформаційних стимулів зовнішнього світу. Разом з тим вони реєструють стан внутрішнього середовища організму.
Енергія сприйнятих стимулів , перетворена сенсорними нейронами в специфічні процеси нервового збудження , піддається подальшій обробці і фільтрації в центральних відділах аналізаторів . Формуючи цілісний образ подразників на основі вроджених реакцій і набутого досвіду , комахи виявляють явні здібності до дресирування , запам'ятовування та узагальнення , не поступаючись в цьому відношенні хребетним тваринам. При цьому комахи зберігають своєрідність , незважаючи на всі подібності доцільних реакцій і актів поведінки з такими хребетних тварин , протиставляються їм як істоти, що живуть у світі інших масштабів і стимулів .
Висновок
Розгляд структурної та функціональної організації центральної нервової системи комах показує , що тулубний мозок цих тварин виступає як виключно складна система , порівнянна в багатьох відносинах з туловищного мозком вищих хребетних. Разом з тим у структурі та функції нервової системи комах є ряд важливих особливостей , що відрізняють її від інших високоорганізованих тварин.
Комахи мають центральну нервову систему , що складається з нервових вузлів - гангліїв , з'єднаних поздовжньо парними нервовими стовбурами - конективами . Клітинні тіла сенсорних нейронів комах винесені з центральної нервової системи та розташовуються поблизу іннервіруємих органів і тканин. Тіла центральних нейронів ( інтер- і мотонейронів ) знаходяться на периферії ганглія під оболонкою і позбавлені будь-яких синаптичних контактів. Встановлено , що мембрана соми більшості центральних нейронів комах електрично невозбудима .
Список літератури
1.Елтрингем
2. Свидерський В.Л.
3. http://www.museumkiev.org/zoology/insecta.html
4. http://www.entomologa.ru/book/22.htm
5. http://uk.wikipedia.org/wiki/%D0%9A%D0%BE%D0%BC%D0%B0%D1%85%D0%B8
6. http://webkonspect.ru/?room=profile&id=2687&labelid=26823
26.11.2013
PAGE \* MERGEFORMAT1
Гістологія центральної нервової системи