Морфологія і аутекологія деревно-чагарникових та багаторічних трав’яних рослин, що використовуються у топіарному мистецтві
РОЗДІЛ 2. Морфологія і аутекологія деревно-чагарникових та багаторічних трав’яних рослин,
що використовуються у топіарному мистецтві
2.1. Поняття про біоморфи
Велике різноманіття зовнішнього вигляду рослин можна звести до кількох найбільш узагальнених типових форм. У навчальній та спеціальній літературі наводиться велика кількість систем життєвих форм [література…], однак найбільш детальною і загальноприйнятою є класифікація біоморф К.Раункієра, перший варіант якої був опублікований у 1903 р. [Хржановський, 1976]. Він виділяв 5 основних біологічних типів рослин: 1) фанерофіти; 2) хамефіти; 3) гемікриптофіти; 4) геофіти; 5) терофіти (однорічники).
Більш детально ця система має такий вигляд.
А. Фанерофіти, або рослини з відкритими бруньками відновлення (Ph). Бруньки відновлення знаходяться вище 25 см над рівнем ґрунту.
1. Мегафанерофіти (дерева заввишки понад 30 м) і мезофанерофіти (дерева від 8 до 30 м заввишки).
2. Мікрофанерофіти (дерева і чагарники від 2 до 8 м заввишки).
3. Нанофанерофіти (чагарники до 2 м заввишки).
4. Епіфіти.
5. Сукуленти.
Б. Хамефіти, або рослини з низько розташованими бруньками відновлення (Ch). Бруньки відновлення знаходяться невисоко, не вище 25 см над рівнем ґрунту.
В. Гемікриптофіти, або рослини з приземними бруньками відновлення (H). Бруньки відновлення знаходяться на рівні поверхні ґрунту, захищені мертвим покривом або верхнім шаром ґрунту.
Г. Геофіти (G). Бруньки відновлення знаходяться під землею.
1. Геофіти кореневищні.
2. Геофіти бульбостеблові.
3. Геофіти бульбокореневі.
4. Геофіти цибулинні.
5. Геофіти з бруньками на коренях.
Д. Терофіти (Т). Однорічники. Переносять несприятливі пори року лише у вигляді насіння.
Дана система класифікує життєві форми покритонасінних і наглядно демонструє їх еволюцію: від дерев до трав.
2.2. Верхівковий і базисний розвиток деревних рослин
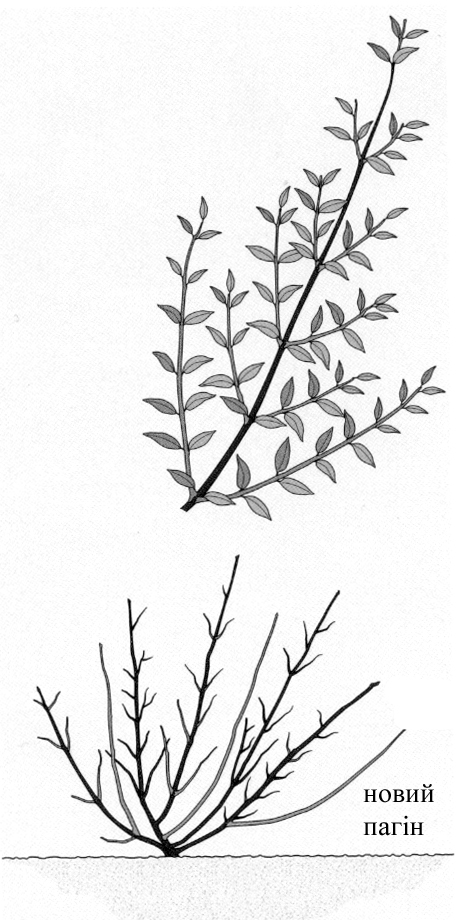
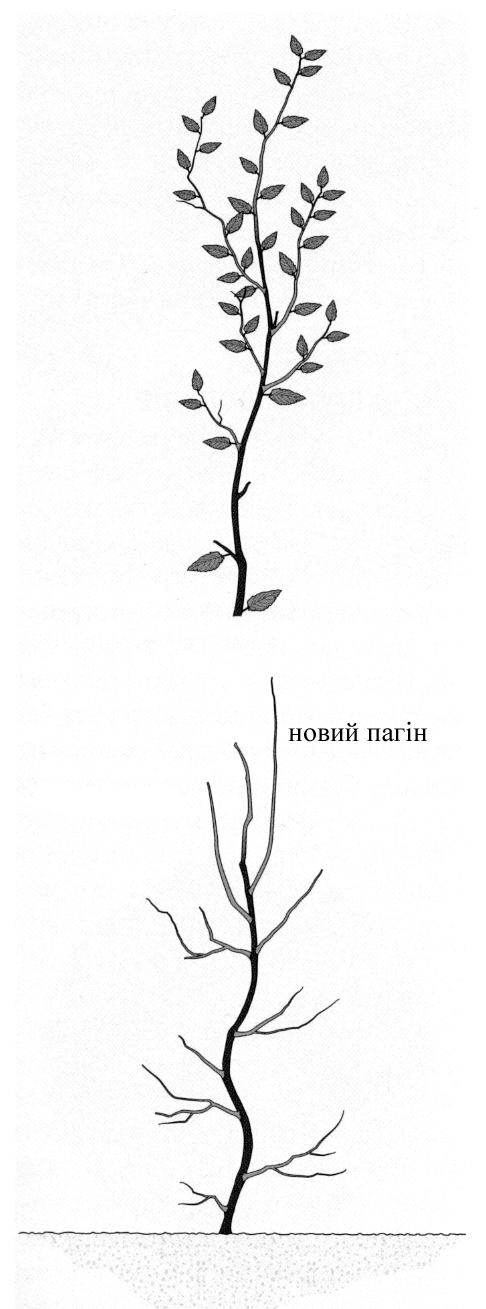
В природних умовах всі дерева та чагарники зростають по-різному. Залежно від процесу вигонки вони поділяються на два типи: базипетальні та акропетальні [Г.Бельц. Фигурная стрижка…].
Базипетальним (базисним) розвитком характеризуються чагарники, у яких найнижчі бруньки дають найбільш інтенсивно ростучі пагони, що в результаті утворюють найбільш довгі гілки (рисунок зправа). Така форма росту характерна для багатьох гарноквітучих чагарників – форзиції, вейгели, садового жасмину.
Дерева та деякі чагарникові рослини характеризуються верхівковим розвитком, тобто ростуть акропетально. В цьому випадку інтенсивніше розвиваються і утворюють найбільш довгі пагони бруньки, розташовані вгорі (рисунок зліва). У багатьох видів рослин, наприклад, у ялиці білої чи гіркокаштана звичайного, цей тип зростання яскраво виражений, у інших рослин, наприклад у самшиту вічнозеленого чи тису ягідного, дещо менше.
Форма розвитку рослин визначається на генетичному рівні: у акропетальних рослин верхівкові бруньки та пагони виділяють фітогормони (наприклад, ауксин), які стримують ріст бруньок та пагонів, розташованих нижче. Розташовані вище частини рослини розвиваються інтенсивніше нижніх, в результаті чого останні можуть частково втрачати листя та відставати в рості. Такі процеси регулюються за допомогою різних видів обрізування.
2.3. Морфологія вегетативних органів деревних рослин
2.3.1. Пагін
Спільне походження складових пагона (стебла, листків, бруньок), їх формування з конуса наростання в процесі онтогенезу, спільність їхньої провідної системи доводить цілісність і єдність його структури. Осьовою частиною пагона є стебло.
Стебло – осьовий орган вищих рослин, до якого належать два поздовжніх метамери – вузли та міжвузля. Стебло разом з розміщеними на ньому листками та бруньками утворюють пагін. Головний осьовий орган у деревних рослин називають стовбуром.
Стебло виконує різні функції, основними з яких є провідна, механічна (опорна) та запасаюча. Його морфологічна та анатомічна будови забезпечують виконання цих функцій. Двостороннє взаємопротилежне проведення речовин – мінеральних поживних речовин і продуктів фотосинтезу – забезпечують розвинені провідні тканини. Виконання опорної функції забезпечують механічні тканини у всіх основних рослинних органах. Розвиток запасаючих паренхімних тканин у корені та стеблі уможливлює відкладення органічних речовин.
Стебла за розміщенням у просторі (напрямком росту) розрізняють прямостоячі чи вертикальні (ортотропні); відхилені від вертикальної осі (плагіотропні), серед яких виділяють лежачі, сланкі, виткі, чіпкі, повзучі; анізотропні, що змінюють початковий горизонтальний напрямок росту на вертикальний. У ряду видів, зокрема верби, бука, берези, горобини та ін., є плакучі форми з повислими вниз пагонами.
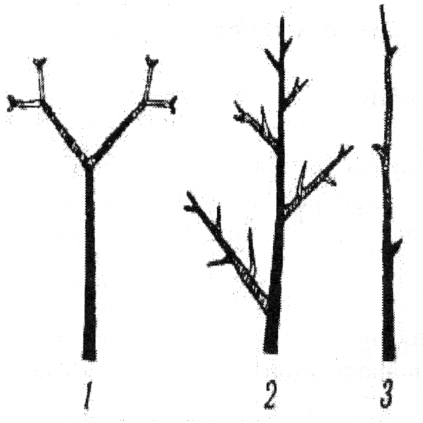
Характерною ознакою виду деревних рослин є тип галуження стебла чи пагона з почленуванням вегетативного тіла рослини для збільшення його поверхні. Розрізняють моноподіальне. симподіальне та несправжньо-дихотомічне галуження (рис. 1).
Під час моноподіального галуження головна вісь за рахунок щорічного росту верхівкової (термінальної) бруньки не припиняє росту у висоту протягом онтогенезу. Головна вісь переважає за швидкістю росту нижче розташовані бічні гілки. Моноподіальний тип галуження характерний для хвойних та значної частини квіткових рослин, зокрема, видів бука, дуба, ясена.
У ході симподіального галуження ріст головної осі в довжину відбувається з верхньої бічної бруньки, оскільки дерева, для яких характерний цей тип галуження, не закладають справжніх верхівкових бруньок при спіральному листкорозміщенні. Верхівку минулорічного приросту, що перетворюється на колючку чи підсихає, видно при основі нової верхівкової бруньки, а з другого її боку – листовий слід. Симподіальний тип галуження характерний для багатьох покритонасінних дерев і кущів, зокрема видів яблуні, груші, липи, вільхи, берези та ін.
Несправжньо-дихотомічне галуження є похідним від моноподіального та притаманне деревним рослинам з супротивним листкорозміщенням. Верхівкову бруньку, що припинила свій ріст через всихання чи перетворення у квітку, суцвіття чи колючку, замінюють дві найближчі бічні бруньки, розміщені супротивно. Такий тип галуження характерний для видів клена, бархата, гіркокаштана, калини, бузку, садового жасмину та ін.
У практичному використанні вживанішим є визначення пагона як частини стебла з розташованими на ній листками і бруньками. Місце розташування листка чи листків разом з бруньками на пагоні називають вузлом, а ділянку пагона між його вузлами – міжвузлям. Структурі пагона властиве чергування вздовж його осі вузлів та міжвузль. Можна вважати, що пагін складається з одиниць росту, до яких належить частина стебла, що завершується вузлом з листком і брунькою. Кут між стеблом і листком, в місці його кріплення, називають листковою пазухою.
Виділяють різні групи пагонів залежно від їх віку, розміщення, будови, швидкості росту та інших ознак.
Однорічними називають пагони, що розвинулись із бруньки відновлення за один вегетаційний період.
Ростовими називають розвинуті бічні пагони, що формуються на вершинах гілок (бічних утворень головної осі) та забезпечують збільшення об’єму крони.
Паросткові (водяні, жирові, вовчки) сильнорослі пагони з довгими міжвузлями з’являються на стовбурах і товстих гілках із сплячих бруньок та коренях із адвентивних бруньок (у видів тополі, малини, сливи, вишні, робінії звичайної). Причиною їх утворення є відмирання скелетних та бічних гілок, спричинене природним старінням чи пошкодженням (у видів дуба, липи, клена та ін.), а у плодових видів – обрізуванням чи обмерзанням. Цим швидкорослим пагонам властиві, порівняно з іншими, крупніші та іншої форми листки.
2.3.2. Брунька
Брунька – це зачаток нерозгорнутого пагона з укороченими міжвузлями та зачатковими листками, що знаходиться у стані відносного спокою для перенесення несприятливого сезону.
Складовими частинами бруньки є зачаткове стебельце із зачаткових метамерів, що закінчується конусом наростання, з зачатковими (примордіальними) листками різного віку, розміщеними один над одним (у вегетативних бруньок) чи з зачатковими квітками (у генеративних бруньок) чи тих і інших одночасно (у змішаних бруньок) (рис. 2).
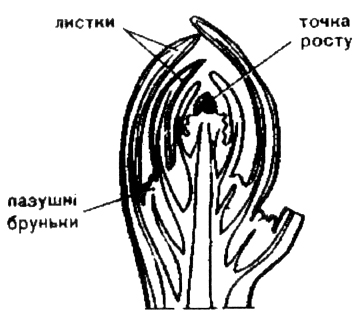
Переважно, вегетативні та генеративні бруньки чітко різняться між собою розмірами та формою. Форма, розміри, колір бруньок, наявність покривів, порядок розміщення та ін. є діагностичними ознаками видів деревних рослин.
Бруньки за походженням розрізняють екзогенні та ендогенні. Екзогенне походження за рахунок діяльності апікальної меристеми мають верхівкові та бічні бруньки. Бічні бруньки утворюються в пазухах листкових зачатків біля верхівки осьового пагона. Придаткові (адвентивні) бруньки мають ендогенне походження – утворюються на різних частинах рослини з різних груп живих клітин. Вони формуються на міжвузлях пагонів, коренях, рідше листках із внутрішніх рослинних тканин – перициклу, камбію, паренхіми серцевинних променів, травматичних меристем, епідерми та паренхіми листка. Адвентивні (додаткові) бруньки забезпечують утворення паросткових пагонів при вегетативному розмноження та відновленні пошкоджених органів, зокрема у видів тополі, малини, бузку та ін.
За особливостями розміщення бруньки поділяють на верхівкові (термінальні), які розташовані на верхівці стебла та забезпечують ріст рослини, та бічні (пазушні), що розмішені в пазухах листків та забезпечують формування розгалуженої крони. Бічні бруньки за розташуванням поділяють на почергові (у видів дуба, в’яза, яблуні та ін.), супротивні (у видів ясена, клена, бузку та ін.), кільчасті, що ідентично трьом типам листкорозміщення, а також серіальні та колатеральні. Серіальними називають кілька бруньок, сформованих в пазусі листка за рахунок діяльності пазушної меристеми, та розташованих вертикальним рядом. Формуються вони у видів жимолості, ожини, горіха, робінії, аморфи та ін. При розташуванні цих бруньок горизонтальними рядами у видів вишні, вовчеягідника та ін. їх називають колатеральними (рис. 3).
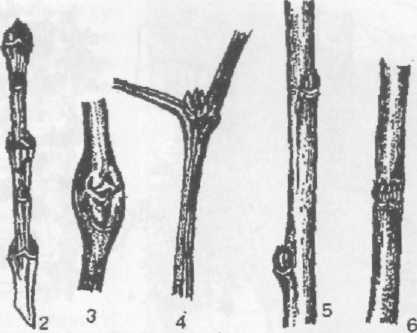
Рис. 3. Розташування бруньок на стеблі: 1 – чергове у в’яза, 2 – супротивне у клена,
3 – серіальне у хвилівника великолисткового, 4 – бісеріальне у хвилівника звичайного,
5 – колатеральне у вовчеягідника звичайного, 6 – кільчасте у вишні повстистої,
7 – листковий рубець з листковими слідами у дуба
За функціональними особливостями бруньки деревних рослин поділяють на вегетативні, генеративні, вегетативно-генеративні, бруньки регулярного відновлення, збагачення та сплячі.
Вегетативні та генеративні бруньки формуються окремо, переважно, у видів, у яких фази цвітіння та розпускання листків розмежовані у часі (види форзиції, дерену, магнолії та ін.). Вегетативно-генеративну бруньки властиві видам бузку, бузини, груші, гіркокаштана, свидини, яблуні та ін., які цвітуть після розпускання листків.
Бруньками регулярного відновлення називають такі, що під час несприятливого сезону року перебувають у стані спокою (спочиваючі чи зимуючі). До цієї групи відносять як верхівкові, так і бічні бруньки екзогенного походження і придаткові бруньки ендогенного походження, так і вегетативні, вегетативно-генеративні, генеративні бруньки майже всіх деревних рослин помірного поясу Землі.
Бруньками збагачення є частина бічних бруньок, що не мають періоду спокою. Закладаються вони у деревних рослин досить рідко, зокрема у видів берези, жимолості.
Частину бічних бруньок екзогенного походження та адвентивних бруньок ендогенного походження, що протягом тривалого часу не розвиваються, але залишаються життєздатними, називають сплячими. Сплячі бруньки залишаються на поверхні стебла за рахунок щорічного приросту нових зачаткових метамерів на ширину річного кільця.
Розвиток сплячих бруньок спричинюють пошкодження пагонів, старіння рослини, надмірне освітлення пагонів, рідше – грибкові зараження. Формування пагонів із сплячих бруньок забезпечує омолодження рослини, особливо кущів, та відновлення пошкоджених органів.
За способом кріплення до пагонів бруньки поділяють на черешкові, що з’єднані з пагоном за допомогою черешка, та сидячі, які прямо прикріплені до пагона без додаткових утворів.
Залежно від наявності покривів та стану захищеності бруньки поділяють на закриті та відкриті. У закритих бруньок зовнішні листки трансформувалися в брунькові луски, які запобігають висиханню внутрішніх тканин. Такі бруньки характерні, переважно, для всіх деревних рослин помірних та холодних кліматичних поясів. У деревних видів із закритою брунькою кількість брунькових лусок є типовою для кожного виду та виступає його діагностичною ознакою. їх кількість змінюється від однієї-двох (у видів платана, верби та ін.) до понад двадцяти (у видів дуба).
У відкритих (голих) бруньок відсутні спеціалізовані видозмінені брунькові луски. Здебільшого, у деревних рослин помірних та холодних кліматичних поясів це бруньки літнього періоду, за винятком небагатьох видів, зокрема калини-гордовини, лапини крилоплодої, крушини ламкої, відкриті бруньки яких зимують. Відкриті бруньки характерні для деревних рослин тропічного та субтропічних кліматичних поясів.
2.3.3. Листок
Листок є вегетативним органом рослин, що еволюційно сформувався як видозміна пагона. Будова та функції листка як елементарної одиниці тіла рослини та пагона взаємопов’язані.
Морфологічні особливості серединних листків є типовими для виду деревних рослин та виступають його діагностичними ознаками.
Складовими частинами листка деревних рослин, в основному, є листкова пластинка, черешок та прилистки. Інколи при основі листка чи на черешку утворюються лусочки, колючки (у видів шипшини). У черешкових листків листкова пластинка кріпиться до пагона за допомогою черешка. За його відсутності листок називають сидячим.
Прилистки – це парні вирости з обох боків листка, що симетрично розташовані. У більшості деревних рослин вони відсутні чи недорозвинені, або ж рано опадають після розпускання листків. Прилистки мають форму маленьких листочків (айва японська висока), лусок (у видів дуба, липи), плівок чи колючок (робінія звичайна). Розрізняють прилистки вільні (у видів глоду) чи зрослі з черешком (у видів шипшини).
Порядок розташування листків (листкорозміщення) на пагонах деревних рослин поділяють на типи: почергове (спіральне), супротивне та кільчасте.
При почерговому (спіральному) листкорозміщенні у вузлах розташовано тільки по одному листку, через основи яких можна провести спіральну лінію. Таке листкорозміщення властиве переважній більшості деревних рослин, зокрема видам дуба, тополі, яблуні, ліщини, смородини та ін.
При супротивному листкорозміщенні у вузлі формується по два листки, розташованих один навпроти одного. Причому у двох сусідніх вузлах пари супротивних листків розміщені перпендикулярно. Такий тип листкорозміщення властивий видам ясена, клена, бузку та Ін.
Для кільчастого листкорозміщення у вузлі характерне утворення по три і більше листків.
Діагностичними ознаками листкової пластинки є її форма, форми основи, верхівки, краю, жилкування, ступінь її розчленування, наявність опушення та ін.
Листкову пластинку за формою поділяють на округлу (тополя тремтяча), овальну (скумпія звичайна), продовгувату, широкояйцеподібну, яйцеподібну (бук лісовий), ланцетну (верба біла), оберненошироко-яйцеподібну (вільха чорна), оберненояйцеподібну (в’яз шорсткий), оберненоланцетну, лінійну, голчасту (сосна звичайна) та лускату (туя західна).
Типова форма листкової пластинки зумовлена у кожного виду спадково, проте за змінених умов Існування може варіювати. Форма листкової пластинки може змінюватися навіть у межах однієї рослини (гетерофілія), що пов’язано з екологічними умовами, віком пагонів, відмінностями у функціях листків. Серед деревних рослин явище гетерофілії характерне для тополі білої, багряника японського, шовковиці білої, смоківниці звичайної, плюща звичайного та ін.
За формою основи листкової пластинки розрізняють округлу (яблуня лісова), клиноподібну (верба біла), серцеподібну (липа серцелиста), ниркоподібну (церціс європейський), зрізану (ліріодендрон тюльпановий), асиметричну (в’яз граболистий), а також стрілоподібну, списоподібну.
За формою верхівки листкової пластинки розрізняють тупу, гостру, загострену, гострокінцеву (магонія падуболиста), виїмчасту (вільха чорна).
За формою країв листкові пластинки розрізняють цілокраї (магнолія оберненояйцеподібна), пилчасті (груша звичайна, липа дрібнолиста), двічіпилчасті (вільха сіра), зубчасті (в’яз шорсткий, ліщина звичайна, клен гостролистий), городчасті (гамамеліс віргінський) та ін. Виділяють також проміжні типи.
Листкова пластинка може бути цілісною чи розчленованою на частини, які розташовані перисто чи пальчасто залежно від типу жилкування. В першому випадку листки вважають перисто- чи пальчасто-цілісними. При незначному розчленуванні листки вважають перисто-чи пальчастонадрізаними.
При розчленуванні листкової пластинки до 1/4 її ширини листок вважають лопатевим (дуб скельний, клен польовий). При розчленуванні більше 1/4 ширини листок називають роздільним (дуб червоний, платан східний). При розчленуванні до середньої жилки чи основи пластинки листок називають розсіченим (розсіченолисті форми вільхи сірої, клена гостролистого). Перисторозсічений листок з найбільшою середньою частиною порівняно з боковими називають ліроподібним.
Листки називають простими при наявності однієї листкової пластинки на черешку. За наявності на одному черешку кількох листкових пластинок (їх називають листочками) листки вважають складними, а їх основну вісь – рахісом.
Розрізняють трійчастоскладні листки з трьох листочків (птелея трилиста, зіновать руська), пальчастоскладні листки з радіальним розходженням листочків від однієї точки (гіркокаштан звичайний, дикий виноград п’ятилисточковий) та перистоскладні листки.
Серед останніх виділяють парноперисті з попарно розташованими листочками з обох боків рахісу (карагана деревоподібна) та непарноперисті з одним листочком на верхівці (горіх грецький, горобина звичайна).
У топіарному мистецтві форма рослини, яка створюється за допомогою стрижки, виглядає ефектніше, якщо поверхня фігури має гладкий рівний вигляд. Такий ефект забезпечують рослини із дрібними листочками (самшит вічнозелений) чи хвоєю (ялина європейська, тис ягідний). Якщо ж у рослини крупні листки (наприклад, лавровишня лікарська), сформувати рівну фігуру дещо складніше. Крім того на крупних листках можуть бути помітні пошкоджені коричневі краї, які з’являються під час стрижки ріжучим інструментом. Тому розмір листя тісно пов’язаний із розмірами майбутньої фігури.
2.4. Основи екології деревних рослин
2.4.1. Класифікація екологічних факторів
Класифікація екологічних факторів на групи передбачає різні підходи. П.С. Погребняк (1968) екологічні фактори поділяє на три групи: абіотичні (фактори неживої природи), біотичні (фактори живої природи – ценотичні та зоологічні) та антропогенні (рис. _).
Абіотичні екологічні фактори поділяють на три категорії:
- кліматичні (атмосферні) – світло, тепло, опади, вологість повітря, вітер, випаровування, складові повітря, зокрема кисень та вуглекислий газ, атмосферне електричне поле та ін.;
- едафічні – фізичні властивості ґрунту (вологість, механічний склад, аерація, теплові властивості, потужність та ін.), хімічний склад, кислотність, наявність шкідливих речовин, засоленість, лісова підстилка та ін.;
- геологічні – рельєф місцевості, поверхневий стік, ерозія схилів, паводки, землетруси, морські припливи та відпливи, вулканічна діяльність та ін.
До біотичних екологічних факторів відносять сукупність елементів органічного світу (мікроорганізмів, тварин, рослин, а за деякими класифікаціями – і людей), які виявляють взаємний вплив та визначають умови існування різних видів організмів на певній території.
До антропогенних екологічних факторів відносять господарську діяльність людини і її вплив на органічний світ, що призводить до значних змін у природі.
Вивченням складного комплексу взаємозв’язків окремих видів рослинних організмів з навколишнім середовищем займається розділ екології, який називають аутекологією (від гр. сам та екологія).
2.4.2. Абіотичні екологічні фактори
Кліматичні екологічні фактори
Роль світла як надзвичайно важливого екологічного фактора вирішальна в процесі фотосинтезу. Сонячна фотосинтетична активна радіація (довжина хвилі від 380 до 750 нм) зумовлює освітлення, теплові умови та температурний режим. Світло за рахунок синтезу органічних сполук спричинюється до утворення та росту органів рослини.
С.С. П’ятницький (1960) за відношенням рослин до світла поділив їх на чотири групи: дуже світлолюбні, світлолюбні малотіневитривалі, відносно тіневитривалі, дуже тіневитривалі. Інші науковці (Нечитайло, 2000; Григора 2000) за вимогливістю до освітлення поділяють рослини на три екологічні групи: геліофіти – світлолюбні рослини, факультативні геліофіти – тіневитривалі рослини, сціофіти – тіневі рослини.
Узагальнена шкала тіневитривалості деревних рослин включає такі групи.
Дуже світлолюбні деревні рослини, які вимагають повного освітлення: саксаули (всі види), акації (всі види), тамарикси (всі види), евкаліпти (всі види), верби біла і ламка, тополі біла та чорна, дуби корковий і пухнастий та ін.
Світлолюбні деревні рослини, які погано витримують невелике затінення: модрини (всі види), робінія звичайна, гледичія колюча, софора японська, тополі тремтяча та сіріюча, ліріодендрон тюльпановий, береза повисла, айлант найвищий, сосна звичайна та ін.
Світлолюбні малотіневитривалі деревні рослини: горіхи грецький і чорний, оксамитник амурський, ясени (всі види), клен ясенелистий, в’яз граболистий, вільха чорна, дуби звичайний та скельний, сосни кримська та чорна, каштан їстівний, береза пухнаста, терен, шипшини (всі види), маслинки (всі види), обліпихи (всі види) та ін.
Відносно тіневитривалі деревні рослини: сосни веймутові (всі види), клени гостролистий, польовий, татарський та явір, дуб північний, в’яз шорсткий, платан східний, катальпи (всі види), черешня, горобини (всі види), яблуня лісова та ін. Ці рослини комфортно себе почувають і при повному освітленні, але можуть задовольнитися і його 30-70% рівнем.
Тіневитривалі деревні рослини: в’яз гладкий, псевдотсуга Мензіса, секвоя вічнозелена, сосни кедрові (всі види), ялини звичайна та колюча, дзелькова граболиста, липи (всі види), граб звичайний, буки лісовий і східний, вільха сіра, гіркокаштан звичайний, ліщини (всі види), свидини (всі види), бруслини (всі види), жимолость татарська, садовий жасмин звичайний, калини (всі види), бузини (всі види), глоди (всі види) та ін.
Дуже тіневитривалі (тіневі) деревні рослини: плющі (всі види), ялиця біла, тис ягідний, самшит вічнозелений та ін. Ці рослини, оптимально розвиваються при 10-30%-ому освітленні, а можуть підтримувати життєдіяльність навіть при однопроцентному освітленні (самшит вічнозелений).
Для життєдіяльності світлолюбних видів корисне верхнє освітлення чи бокове разом з розсіяним. Для розвитку тіневитривалих видів достатньо лише бокового чи розсіяного світла. Висока інтенсивність світла викликає у рослин важливі морфологічні зміни – прискорює розвиток та збільшує довжину коріння. Сонячне світло впливає на розвиток листків, галуження, розміри і форму крони, форму стовбура, величину приросту, якість деревини та насіння.
Освітленість, безпосередньо, визначає розташування генеративних органів в кроні і порядок розпускання квітів.
Для рослин важливим є як інтенсивність, так і тривалість освітлення. Реакцію рослинних організмів на співвідношення між тривалістю дня і ночі називають фотоперіодизмом. Він забезпечує рослинам перехід до цвітіння. Розрізняють такі основні фотоперіодичні групи рослин.
Короткоденні рослини (рослини короткого дня) потребують для утворення репродуктивних органів до 12 год. освітлення; ростуть в південних регіонах. Довгоденні рослини (рослини довгого дня) вимагають понад 12-14 год. тривалості дня для початку формування генеративних органів; ростуть в північних регіонах. У нейтральних рослин, що є, в основному, космополітами, цвітіння починається за будь-якої тривалості дня.
Тепло, як екологічний фактор, зумовлює географічне розповсюдження рослин.
П.С. Погребняк (1968) за ставленням до тепла деревні рослини поділив на чотири групи: дуже теплолюбні, теплолюбні, середньо-вимогливі до тепла, маловимогливі до тепла. С.С. П’ятницький (1960) за теплолюбністю (вимогливістю до тепла) деревних рослин запропонував деталізованішу шкалу: дуже теплолюбні, теплолюбні, відносно холодостійкі, холодостійкі та дуже холодостійкі.
Дуже теплолюбні види: кипариси (всі види), евкаліпти (всі види), кедри (всі види), секвоя вічнозелена, криптомерія японська, ялівці (окремі види), бамбуки (всі види), цитрусові (всі види), лавр благородний, дуб корковий, саксаули (всі види), сосни приморська, піцундська та ельдарська.
Теплолюбні види: каштан їстівний, айлант найвищий, смоківниця звичайна, евкомія в’язолиста, ясен білоцвітий, тиси (всі види), платани (всі види), каркас кавказький, дуб пухнастий, карія пекан, горіх грецький, софора японська, тополя біла, айва продовгувата, катальпи (всі види), маклюра яблуконосна, лапина крилоплода, самшит вічнозелений, клокичка периста, мигдалі (всі види), персики (всі види), ліріодендрон тюльпановий.
Відносно холодостійкі види: дуби звичайний, скельний, крупно-пиляковий та каштанолистий, граб звичайний, буки лісовий і східний, клени явір, польовий, цукристий, прирічковий, ясен звичайний, ялиці кавказька та біла, сосна кримська, туї західна та східна, ялівець віргінський, в’яз граболистий, оксамитник амурський. липи широколиста і срібляста, тополі пірамідальна і Болле, гледичія колюча, робінія звичайна, горіхи чорний, абрикос звичайний, каркас західний, вишня магалебська, ялина східна, шовковиця біла, черешня, алича, берека лікарська.
Холодостійкі види: дуби червоний і великоплодий, липа серцелиста, горіхи сірий і маньчжурський, ясени пухнастий і зелений, в’язи шорсткий і гладкий, груша звичайна, яблуня лісова, тополя чорна, верба біла, клени гостролистий і татарський, гіркокаштан звичайний, горобини звичайна і проміжна, вільха чорна, ліщина звичайна, калина звичайна, жовта акація, сосна чорна, ялиця бальзамічна.
Виключно холодостійкі види: тополі тремтяча і бальзамічна, берези повисла і пухнаста, ялини звичайна і сибірська, ялиці біла та сибірська, ялівець звичайний, сосна звичайна, сосни кедрові європейська, сибірська та сланка, модрини (всі види), вільха зелена.
Стовбур, крона та коренева система деревних рослин знаходяться в різних теплових умовах. Так, температура ґрунту залежить від кількості тепла, що надходить на поверхню ґрунту та проникає в його товщу. Під час найбільшої радіації о 13 годині температура на глибині 10 см, порівняно з поверхнею, нижча на І0-І5°С, а на 25 см — на 12-17°С.
Здатність деревних рослин витримувати без укриття низькі температури (нижче 0°С) повітря, що не призводить до їх ураження, називають морозостійкістю. Морозостійкість рослин зумовлюється грунтово-кліматичними умовами, спадковими особливостями, наявністю захисних покривів у рослин, утворенням в протоплазмі клітин цукрів, ліпідів, органічних кислот, глюкозидів та інших захисних речовин. Морозостійкість деревних рослин підвищується з віком, її можна покращити за допомогою агротехнічних заходів.
За ступенем морозостійкості деревні рослини поділяють на: дуже морозостійкі, що можуть витримувати без захисту тривалі пониження температури повітря до -35-50°С і нижче; морозостійкі, що здатні витримувати морози -25-35°С; відносно морозостійкі, що можуть витримувати морози до -15-25°С; неморозостійкі, здатні витримувати морози до -10-15°С; зовсім неморозостійкі, здатні витримувати лише короткотривалі пониження температури не нижче -10°С. Шкала морозостійкості деревних рослин є оберненою до шкали їх теплолюбності.
Вода – це складова частина для клітин і тканин: вона необхідна для життєдіяльності плазми, для засвоєння і пересування мінеральних речовин, увібраних корінням, із ґрунту в стовбур і крону, для нормального ходу фотосинтезу і транспірації. Джерелом води для рослини є атмосферні опади – дощ, сніг, а також роса, іній, туман, ґрунтові води та ґрунтова волога.
У період вегетації, особливо цвітіння і достигання плодів, інтенсивного росту пагонів і кореневої системи, рослиною витрачається значна кількість води. Тому волога, змінюючи до певних умов інтенсивність життєвих процесів, може проявляти істотний вплив на плодоношення рослин.
Вітер як кліматичний фактор може виявляти на деревні рослини пряму і опосередковану дію. Пряма дія вітру виражена у механічних впливах на рослини. Це, зокрема, розхитування дерев, оббивання гілок, зривання листків, квіток і плодів. Під впливом вітру посилюється інтенсивність втрати вологи ґрунтом, транспірація у рослин, відбувається поширення пилку для запилення квітів, розповсюдження плодів та насінин.
Опосередкованим впливом вітру на рослини є надмірне висушування повітря, що збільшує транспірацію. Також через насичення повітря вуглекислим газом вітер сприяє посиленню процесу фотосинтезу. Вітер часто спричинюється до ерозії ґрунтів, причому оголення кореневих систем рослин негативно позначається на їхньому рості.
У даному випадку незамінною є роль живоплотів та зелених стін у регулюванні вітрових потоків, які проявляється у зниженні швидкості вітру та створенні комфортних умов на ділянках перебування людей. Також важливою функцією живоплотів є регулювання снігових заметів у зимову пору року.
Едафічні екологічні фактори
Ґрунти слугують субстратом для закріплення рослин і джерелом отримання води та мінеральних елементів. Деревні рослини зазнають впливу фізичних властивостей та хімічного складу ґрунту та мікроорганізмів, що населяють його товщу.
Процентне співвідношення твердих кристалічних часток (каміння, щебеню, піску, пилу, мулу) визначає механічний склад ґрунту. У зв’язку з цим виділяють ґрунти щебенисті, піщані, супіщані, суглинкові, глинисті, мулуваті та ін. Рослини, що ростуть на цих ґрунтах, по-різному пристосувалися до умов місцезростань.
Механічний склад, структура і вологість ґрунту зумовлюють його пухкість і повітропроникність. Повітряний режим кращий у чорноземах і піщаних ґрунтах, гірший у глинистих і болотних.
За здатністю рости на різних субстратах рослини поділяють на такі групи.
Галофіти з низькорослими чи розпростертими на поверхні ґрунту стеблами ростуть на зволожених солончаках. Їх стебла та листки короткі, товсті, соковиті, мають добре розвинену паренхіму. Галофіти можуть нагромаджувати велику кількість води. Вони запасають (соленагромаджувальні) чи виділяють (солевидільні) надлишок солей. Галофіти є едифікаторами засолених ґрунтів.
Псамофіти ростуть на піщаних ґрунтах і закріплюються на них завдяки добре розвиненій кореневій системі. Псамофіти з безлистими пагонами в специфічних фізико-хімічних і гідрологічних умовах пристосувалися до прискореного використання вологи, швидкого проростання і вкорінювання насінин. Їх використовують для закріплення рухомих пісків. Серед псамофітів багато ксерофітів, зокрема відомі саксаули, піщані акації, деякі види верб.
Гелофіти – рослини, приурочені до торфових боліт з постійним чи періодичним надлишковим зволоженням. У таких умовах через відсутність кисню відмерлі органічні рештки, що не розклалися, відкладаються у вигляді торфу. В основному, ці рослини належать до гігрофітів. До вічнозелених і листопадних чагарничкових гелофітів належать андромеда багатолиста, багно звичайне, брусниця звичайна, журавлини болотна та дрібноплода, буяхи (лохина), верби чорнична та розмаринолиста, верес звичайний. Екземпляри деревних гелофітів – берези, сосни та ялини на торф’яниках відрізняються значно гіршими анатомо-морфологічним показниками.
Літофіти (петрофіти) – рослини, які ростуть на камінні та скелях, сприяючи руйнуванню гірських порід. Серед деревних рослин до цієї групи належать сосна гірська, ялівець сибірський, душекія зелена, дуб скельний та ін.
Для повноцінного розвитку деревних рослин, забезпечення нормального проходження фізіологічних процесів важливою є наявність азоту, кальцію, магнію, калію, фосфору, інших макро- і мікроелементів, що забезпечують високу родючість ґрунтів.
За вибагливістю до родючості ґрунту деревні рослини поділяють на: евтрофи (мегатрофи) – вибагливі до родючості ґрунту, мезотрофи – середньовибагливі до родючості ґрунту та оліготрофи – невибагливі до родючості ґрунту.
Евтрофи чи мегатрофи ростуть на родючих ґрунтах, потребуючи повноцінного мінерального живлення для забезпечення розвитку. Тут евтрофи вирізняються високою продуктивністю та сильним розвитком надземних і підземних органів. До цієї групи належать: ясени (всі види), горіх грецький, клени гостролистий, явір і польовий, граб звичайний, буки лісовий і східний, ялиці (всі види), тополя чорна, бархат амурський, верби біла і ламка, в’язи шорсткий і граболистий, малина звичайна.
Мезотрофи – середньовибагливі до родючості ґрунту рослини. До них належать види: дуби червоний, скельний, звичайний, каштан посівний, липи (всі види), яблуня лісова, груша звичайна, горобини звичайна і лопатева, сливи (всі види), вишня звичайна, черешня, верба козяча, тополя тремтяча, береза пухнаста, вільха чорна, ялини (всі види), сосна веймутова, модрини (всі види).
Оліготрофи – рослини невибагливі до родючості ґрунту, здатні рости на ґрунтах, бідних на органічні та мінеральні речовини. До оліготрофів належать сосни гірська, звичайна, Банкса та чорна, верес звичайний, береза повисла, робінія звичайна, ялівці (всі види).
Оптимальне забезпечення рослин вологою протягом вегетаційного періоду – важлива передумова їх інтенсивного росту, розвитку та плодоношення. Нестача води – один з факторів, що затримує ріст і знижує плодоношення рослини. Якщо її бракує, погано закладаються плодові бруньки, плоди формуються дрібні, легко обсипаються. Надлишкова кількість опадів сповільнює процес дозрівання плодів і насінин, а нестача – прискорює його, знижує якість насінин або спричинює загибель урожаю.
За вимогливістю до вологозабезпеченості ґрунту деревні рослини поділяють на такі групи.
Гігрофіти – це наземні рослини, що приурочені до постійно вологих та сирих умов місцезростання. До цієї групи відносять такі види: вільху чорну, ясен звичайний (болотний екотип), вербу сіру, вушкату і лапландську, болотні пальми, деякі епіфітні види.
Перехідною групою між гігро- та мезофітами є мезогігрофіти. До них належать: черемха звичайна, верби козяча, ламка, біла, тополі (всі види), береза пухнаста, в’язи гладкий, вільха сіра, крушина ламка, смородина чорна, жимолость звичайна, ясени пенсільванський і маньчжурський.
Мезофіти – наземні рослини, що приурочені до середніх за вологістю ґрунтів з помірним грунтово-повітряним зволоженням. Сюди належать: дуб червоний, липа серцелиста, граб звичайний, ясени звичайний (суходільний екотип) і зелений, бархат амурський, горіхи маньчжурський, сірий і чорний, модрина сибірська, каштан посівний, бук лісовий, ялиці (всі види), ялини (всі види), береза повисла, сосна веймутова, клени гостролистий І явір, в’яз шорсткий, ліщини (всі види), бузини (всі види), бруслина європейська, жимолость татарська.
Перехідною групою між мезо- та ксерофітами є мезоксерофіти. До них належать дуби звичайний та скельний, горіх грецький, клени цукристий, польовим і татарський, в’яз граболистий, черешня, яблуня (всі види), груша (всі види).
Ксерофіти – рослини, що ростуть в посушливих умовах місцезростання та можуть витримувати тривалий час атмосферну та ґрунтову посухи без шкоди для життєдіяльності. До ксерофітів належать: гледичія (всі види), робінія звичайна, граб східний, айлант найвищий, софора японська, скумпія звичайна, дуби пухнастий і корковий, фісташки (всі види), сосни кримська, звичайна, Банкса, платан східний, маслинка вузьколиста, тамарикси (всі види), ялівці (всі види).
Хімічний склад ґрунту суттєво впливає на процеси життєдіяльності рослин. Рослини часто виступають індикаторами наявності окремих елементів чи хімічних сполук у ґрунті. За кислотністю ґрунти поділяють на кілька категорій: кисла реакція ґрунтового розчину до 6,7 рН; нейтральна реакція середовища — 6,7-7,0 рН; лужна реакція — 7,0-14,0 рН.
Рослини за вибагливістю до кислотності ґрунту поділяють на кілька екологічних груп:
Ацидофільні (рН 2,8-6,7), що ростуть на кислих ґрунтах: модрини (всі види), ялини (всі види), ялиці (всі види), сосна звичайна, сосни кедрові, граб звичайний, каштан їстівний, берези (всі види), тополя тремтяча, горобина (всі види), рододендрони (всі види), журавлина дрібноплода, багно болотне, верес звичайний, чорниця звичайна, брусниця звичайна.
Нейтрофільні (рН 6,7-7,0), що ростуть на нейтральних ґрунтах, зокрема клен-явір, робінія звичайна, груша звичайна, в’яз граболистий, шипшини (всі види).
Базифільні (рН понад 7,0), що ростуть на лужних ґрунтах.
Індиферентні рослини невибагливі до кислотності ґрунтів, можуть рости при різній реакції ґрунтового розчину, зокрема дуб звичайний.
Деревні рослини можуть знезаражувати повітря, знищуючи шкідливі мікроорганізми, завдяки фітонцидним властивостям, виділяючи активні леткі речовини.
За фітонцидністю деревні рослини поділено на п’ять груп. До найфітонцидніших віднесено дуб звичайний та клен гостролистий. Чисельною є група сильнофітонцидних рослин – сосна звичайна, ялина звичайна, ялівець звичайний, берези повисла та пухнаста, тополя тремтяча, черемха звичайна, ліщина звичайна, малина звичайна та чорниця звичайна. Не менш чисельною є група середньофітонцидних рослин – модрини європейська, сибірська та Кемпфера, сосни кедрові європейська, сибірська та корейська, ясен звичайний, липа серцелиста, вільха чорна, горобина звичайна, карагана деревоподібна, бузок звичайний, жимолость татарська. Малочисельними є групи слабофітонцидних (в’язи гладкий і шорсткий, бруслина бородавчаста) та найменш фітонцидних рослин (бузина червона та крушина ламка).
2.4.3. Біотичні екологічні фактори
Як зазначає В.Я.Заячук (2008), серед біотичних факторів впливу на деревні рослини розрізняють вплив інших складових біогеоценозів – інших видів рослин (фітогенний), тварин (зоогенний), грибів (мікогенний) та мікроорганізмів (мікробогенний). Їх вплив буває прямим, що виражається в механічній дії, паразитизмі та симбіозі, а також опосередкованим, що полягає у конкуренції за елементи живлення, алелопатії та ін.
Між рослинами фітоценозу форми взаємозв’язків об’єднують у шість основних груп: біофізичні (створення мікроклімату та ін.), біохімічні (виділення хімічних сполук, алелопатія та ін.), біотрофні (харчування та кругообіг елементів), механічні (пошкодження, обламування, деформація та ін.), фізіологічні (зростання окремих органів у біогрупах), генеалогічні (взаємозапилення).
У практиці лісового та садово-паркового господарства вивчення особливостей конкуренції видів під дією екологічних факторів та ходу зрідження деревостанів допомагає моделювати структуру фітоценозів та почергово відбирати з нього відмираючі дерева, а також проводити профілактичні заходи із захисту насаджень від шкідників та хвороб.
2.4.4. Антропогенні екологічні фактори
Антропогенні екологічні фактори спричинюють чи не найзначніший вплив на рослинні організми. Антропогенний вплив на деревні рослини поділяють на прямо діючий (прямий), що виражається у їх вирубуванні, скошуванні, витоптуванні, зборі, штучному розведенні, охороні та ін., та непрямо діючий (опосередкований, непрямий), що відбувається через осушення боліт і заболочених лісових площ, зрошування, затоплення земель під час будівництва водосховищ, забруднення повітря, води і ґрунту та ін., тобто зміна природного середовища людиною (рис. _).
На садово-паркових об’єктах прямодіючим антропогенним фактором є рекреаційне навантаження на рослинне вкриття, яке проявляється механічною дією на рослини (пошкодження дерев, обламування пагонів, зривання квітів), їх витоптування, ущільнення ґрунту.
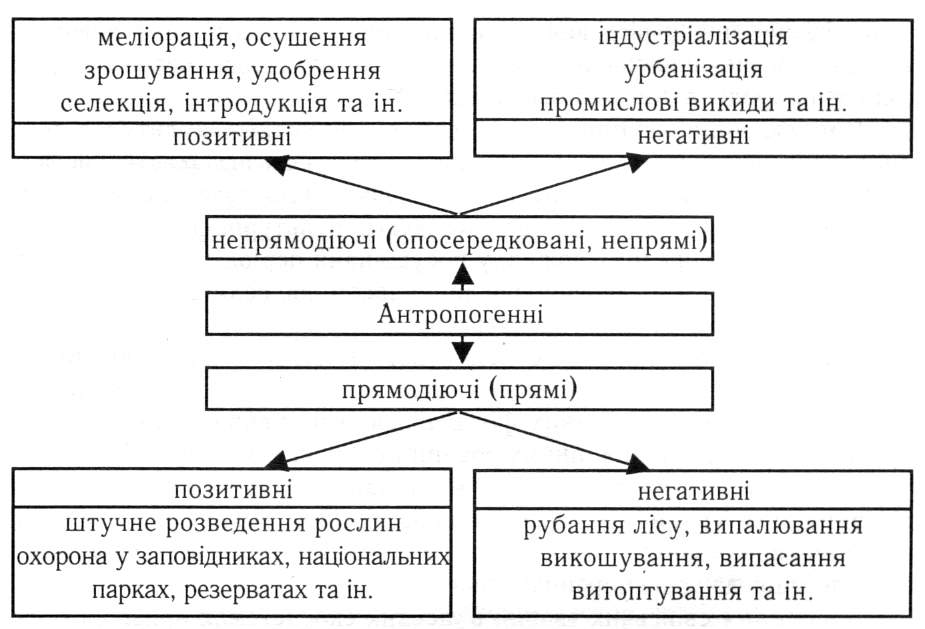
Рис. _. Класифікація антропогенних екологічних факторів
Ущільнення ґрунтів, переміщення підстилки спричинює у деревних рослин пошкодження кореневих систем, утворення кореневих гнилей, зараження грибами. Хаотичний рух пішоходів призводить до витоптування надземного чагарникового та трав’яного вкриття, що спричинюється, особливо на схилах, до водної ерозії ґрунту.
Розвиток промисловості та урбанізації – приклад непрямого впливу антропогенних факторів через зміну умов існування рослин, що спричиняє на деревні рослини надмірне навантаження. Ґрунти, повітря та воду забруднюють викиди автомобілів – вуглеводні та солі важких металів вздовж доріг. Токсичні речовини, перешкоджаючи фотосинтезу, призводять до ослаблення, всихання і загибелі рослин.
Бідніший вміст у ґрунтах міських парків і скверів елементів живлення, загазованість повітря призводять до скорочення тривалості життя деревних рослин. Підвищити вік і стійкість міських зелених насаджень можна шляхом відбору газостійких форм деревних рослин.
Морфологія і аутекологія деревно-чагарникових та багаторічних трав’яних рослин, що використовуються у топіарному мистецтві