Листок, будова листка, фотосинтез, листопад
МІНІСТЕРСТВО НАУКИ І ОСВІТИ УКРАЇНИ
ВІННИЦЬКИЙ ДЕРЖАВНИЙ ПЕДАГОГІЧНИЙ УНІВЕРСИТЕТ
ім. М. КОЦЮБИНСЬКОГО
На тему:
“Листок, будова листка, фотосинтез, листопад.”
виконала студентка природничо-географічного факультету
групи ЖБХ
Крисько Леся
Викладач: д. б. н., проф.
Курята Володимир Григорович
Вінниця 2014 р
Зміст
- Листок (вступ)
- Зовнішня будова
- Різноманітність морфології
- Розміщення на пагоні
- Гетерофілія
- Метаморфози (видозміни) листка
- Внутрішня будова
- Функції
- Фотосинтез
- Типи фотосинтезу
- Просторова локалізація
- Світлова (світлозалежна) стадія
- Фотохімічна суть процесу
- Світлозбиральні комплекси
- Основні компоненти електронно-транспортного ланцюжка
- Фотосистема II
- b6f або b/f-комплекс
- Фотосистема I
- Циклічний і псевдоциклічний транспорт електрона
- Темнова стадія
- С3 – фотосинтез, цикл Кальвіна
- С4 – фотосинтез, цикл Хетча-Слека-Карпілова
- САМ фотосинтез
- Аноксигенний фотосинтез
- Значення фотосинтезу
- Листопад
Список використаної літератури
Вступ
Листок – вегетативний орган вищих рослин, похідне пагону, що виконує функції фотосинтезу, транспірації та газообміну. Розміри листків найчастіше знаходяться в межах 3-10 см, однак є й цікаві винятки. Наприклад, Вікторія амазонська, з родини німфейних, має листки діаметром до 2 м, які можуть витримати вантаж більше 50 кг, а деякі тропічні пальми мають листки довжиною 20-25 м (разом із черешком). Це бічна, зазвичай плоска, зеленого кольору частина пагона. Першими листками насінних рослин є сім'ядолі зародка. Потім завдяки заросткам верхівкової меристеми формуються надземні або підземні листки. Листкова пластинка виконує основні функції листка.Черешок виконує опірну і провідну функції, регулює положення листкової пластинки відносно напрямку світла. По черешку проходять судинно-волокнисті пучки, які з'єднують стебло з листковою пластинкою. Прилистки також можуть мати різну форму й розміри.
�
Листок Victoria regia
Листок – бічний дорсивентральний орган пагона з детермінованим (обмеженим) ростом, пристосований до здійснення фотосинтезу, газообміну, транспірації, а також може виконувати запасальну функцію, слугувати для вегетативного розмноження, навіть бути ловильним апаратом у рослин-хижаків.
На одній рослині листки бувають анатомічно та морфологічно відмінними, що доводить їх пластичність. Певні сукупності листків (формації) різняться положенням, будовою та переважаючою функцією. Пластичність листка проявляється у формуванні різних формацій, а також у здатності змінюватися під впливом умов зовнішнього середовища (гетерофілія).
Зовнішня будова
Типовий листок складається з листкової пластинки (lamina), черешка (petiolus) та прилистків (stipulae). Проте наявність двох останніх частин зовсім необов'язкова. Листки без черешка називаються сидячими. Отже, листкова пластинка – основна частина листка. Якщо листкова пластинка одна, то листок називається простим, а якщо кілька – складним.
Наприклад, прості листки мають береза, липа, дуб, бузок, а складні – конюшина, акація, троянда. Листкові пластинки складного листа розміщуються на загальному черешку, який має назву рахіс. Цікаво, що при листопаді, кожен листочок складного листа відпадає окремо.
� �
Простий листок липи Складний листок троянди
Як правило на листку добре помітні жилочки, що являють собою провідні пучки, по яких листок отримує воду з мінеральними речовинами, а синтезовані в листку органічні речовини, відводяться до інших органів рослини. Жилки також виконують і опорну функцію для клітин листка.
�
Скелет жилкування листка
Виділяють 4 основні типи жилкування:
- паралельне – жилки розташовані вздовж листка паралельно одна одній (наприклад, злакові);
- дугове – жилки знизу і зверху листка зближуються (наприклад, подорожник, конвалія);
- сітчасте – чітко виділяється центральна жилка (одна або кілька), від якої відходять бічні, меншої товщини, які, в свою чергу, неодноразово галузяться, утворюючи цілу сітку дрібних жилок (наприклад, кропива, клен, дуб);
- дихотомічне – жилки галузяться вилчасто (наприклад, гінкго дволопатеве).
Інші типи жилкування показані на малюнку нижче.
Різноманітність морфології
�
Форми листкової пластинки, країв та жилкування листків
Різноманіття зовнішнього вигляду листя просто вражає, тому, природно, виникає потреба в їх класифікації. Малюнок дає досить вичерпну картину зовнішньої морфології листкової пластинки, розглянемо деякі підходи до її класифікації.
Прості листки залежно від форми листкової пластинки можна поділити на дві великі групи:
- з нерозчленованою пластинкою
- голчастий (хвойні)
- лінійний (злаки)
- ланцетний
- округлий
- яйцеподібний
- оберненояйцеподібний
- щитоподібний
- серцеподібний
- ниркоподібний
- стрілоподібний
- списоподібний тощо;
- з розчленованою пластинкою
- лопатевий – розчленування досягають 1/3-1/4 листкової пластинки
- трійчастолопатевий
- пальчастолопатевий
- перистолопатевий
- роздільний – глибина вирізів понад половину ширини листкової пластинки
- трійчастороздільний
- пальчастороздільний
- перистороздільний
- розсічений – розчленування доходять до центральної жилки чи основи листка
- трійчасторозсічений
- пальчасторозсічений
- перисторозсічений
Це була класифікація простих листків за формою листкової пластинки, на перший погляд вона може здатися складною, проте це не так. До цієї класифікації дещо «не вписується» листок картоплі. Він є своєрідним: за формою він перистий, але розчленування неоднакове, великі долі чергуються з маленькими. Такий листок матиме назву «непарнопереривчастоперисторозсічений».
Наступна класифікація простих листків за формою края листкової пластинки. Розрізняють:
- пилчастий
- двічіпилчастий
- зубчастий
- городчастий
- суцільний та ін.
Класифікація складних листків дещо нагадує класифікацію простих з розчленованою листковою пластинкою. Виділяють:
- трійчастоскладний
- пальчастоскладний
- перистоскладний
- парноперистоскладний (рахіс закінчується двома листочками)
- непарноперистоскладний (рахіс закінчується одним листочком)
Розміщення на пагоні
Листки розміщуються на пагонах у певному порядку, не затінюючи один одного. У більшості квіткових рослин (яблуня, груша, верба, осокір та ін.) розміщення листків спіральне, або чергове (вони прикріплені по одному у вузлі). Кількість листків, що замикають спіраль на стеблі, називається листковим циклом. Він може бути з одного або двох обертів спіралі на пагоні. У кожному оберті по два – п'ять і більше листків.
У бузку, глухої кропиви, гвоздики листки прикріплені парами і розміщені на стеблі один проти одного. Таке розміщення листків називають супротивним. Кожна пара відносно верхньої і нижньої пар розміщена навхрест, що сприяє кращому використанню світла.
Якщо до одного вузла прикріплені п'ять і більше листків, то таке розміщення називають кільцевим. Воно буває у хвощів, ялівцю, воронячого ока. У маренки запашної (Asperula odorata), підмаренника північного (Galium boreale) розміщення листків несправжньокільцеве. Крім двох листків, у їх кільцях є розрослі прилистки.
У рослин з прикореневими листками вони зібрані біля землі на дуже вкороченому стеблі з майже недорозвинутими меживузлями (у кульбаби, кок-сагизу), а в перший рік вегетації – у буряка, моркви та ін.
Гетерофілія
Гетерофілія (від грец. hteros – різний та phllon – листок), або різнолистність – це відмінності у формі, розмірах та структурі листків однієї рослини.
Справа в тому, що не всі листки на пагоні однакові. При проростанні рослини з насіння першими з'являються листки зародку – сім'ядолі. В середній частині пагона, пізніше розвиваються серединні листки. Вони зелені та характеризуються всією різноманітністю морфології, описаною вище. Верхівкові листкирозвиваються в області суцвіття. Це покривні листки квіток – приквітники. Вони недорозвинуті, слабко розчленовані, часто плівчасті.
Метаморфози (видозміни) листка
Вчення про метаморфози у рослин висунув Й. В. Гете у 1790 р. Він розвивав ідею єдності всіх органів рослини, які є видозмінами (метаморфозами) листка.
Метаморфози листка виникли в процесі еволюції не просто так. Вони є пристосуванням рослин до певних умов. Розглянемо це на конкретних прикладах. Цікаво, що багато видозмін листків, схожі з видозмінами пагона (стебла).
- Вусики характерні для лазаючих рослин, у яких частина листка або навіть весь листок перетворюються на нитковидні вирости. Чудовий приклад – горох посівний. У нього на вусики перетворена верхня частина рахісу та 3-7 листочків. Виноград теж має вусики, проте вони не листкового походження, оскільки, по-перше, відходять з пазухи листка, а по-друге – можуть галузитись, що властиво пагону, а не листку.
- Колючки виникають як пристосування до посушливих умов зростання, наприклад, у кактусів, таким чином зменшуються втрати води рослиною. Колючками можуть ставати закінчення жилок, рахіс, прилистки. Такі колючки захищають рослину від поїдання тваринами. Колючки пагоневого походження також легко відрізнити від листкових: вони знаходяться в пазусі листка та інколи галузяться (наприклад, колючки глоду, сливи, гледичії, барбарису – це все приклади видозмін пагону).
- Ловильні апарати, певно, найвищий ступінь спеціалізації листка. Вони властиві комахоїдним рослинам. Зростаючи на бідних ґрунтах, росичка, непентеста інші комахоїдні рослини, змушені були шукати додаткове джерело поживних речовин, а саме джерело азоту. В процесі еволюції в них виникли ловильні апарати, пристосовані для полювання на комах та інших дрібних тварин.
-
�
Вусики Гороху посівного
�
Колючки опунції
�
Комахоїдна рослина Росичка круглолиста
�
Комахоїдна рослина Непентес
Внутрішня будова
�
Схема внутрішньої будови листка
�
Устячко листа під мікроскопом
зазвичай, листок складається з таких тканин:
Епідерміс – шар клітин, що захищають від шкідливого впливу зовнішнього середовища та надлишкового випарування. Часто ззовні епідерміс листка вкритий захисним шаром воскоподібного походження (кутикулою).
Мезофіл, або паренхіма – внутрішня хлорофіловмісна тканина, що виконує основну фотосинтетичнуфункцію.
Мережа жилок, що утворені провідними пучками, які складають судини, ситоподібні трубки (елементи ксилемита флоеми – для переміщення води, розчинних солей та цукрів) та механічні елементи.
Продихи – спеціалізовані комплекси клітин, переважно розташованих з нижньої поверхні листків, через які відбувається випарування води (транспірація) та газообмін.
Як і в усіх інших органів, будова листка та його функції тісно пов'язані між собою. Зверху і знизу листкова пластинка покрита епідермісом. Зовнішні стінки клітин епідерміса потовщені та вкриті кутикулою (особливо у ксероморфнихрослин), але можуть бути й відносно тонкими у рослин вологих місцезростань. Бічні стінки клітин звивисті, що сприяє міцному з'єднанню їх між собою.
Функції
Як було зазначено вище, листок виконує три основні функції:
- фотосинтезу – в клітинах мезофілу листка знаходяться хлоропласти – органели фотосинтезу, в хлоропластах енергія світла перетворюється на енергію хімічних зв'язків і як побічний продукт виділяється кисень;
- транспірації – кутикула на поверхні епідерми листка сприяє зменшенню випаровування води з одиниці поверхні, а продихи дозволяють регулювати цей процес, тому транспірація на відміну від випаровування процес фізіологічний;
- газообміну – через продихи, по міжклітинному простору (міжлітинники) до клітин надходить вуглекислий газ (для фотосинтезу), кисень (для дихання), а також виводиться зайва водяна пара та кисень, що утворився в процесі фотосинтезу.
Фотосинтез – процес синтезу органічних сполук з вуглекислого газу та води з використанням енергії світла й за участю фотосинтетичних пігментів (хлорофіл у рослин, хлорофіл, бактеріохлорофіл і бактеріородопсин у бактерій), часто з виділенням кисню як побічного продукту. Це надзвичайно складний процес, що включає довгу послідовність координованих біохімічних реакцій. Він відбувається у вищих рослинах, водоростях, багатьох бактеріях, деяких археях і найпростіших – організмах, відомих разом як фототрофи. Сам процес відіграє важливу роль у кругообігу вуглецю у природі.
Фотосинтез – єдиний процес у біосфері, який призводить до засвоєння енергії Сонця і забезпечує існування як рослин, так і всіх гетеротрофних організмів.[1]
Рослина поглинає не всю сонячну енергію, а лише її фотосинтетичну активну радіацію, під впливом якої проходить фотосинтез, що впливає на майбутній врожай культур.[2]
Узагальнене рівняння фотосинтезу (брутто-формула) має вигляд:
6СО2 + 6Н2О = С6Н12О6 + 6О2
Типи фотосинтезу
Розрізняють оксигенний і аноксигенний типи фотосинтезу. Оксигенний найбільш поширений, його здійснюють рослини, ціанобактерії і прохлорофіти. Аноксигенний фотосинтез проходить у пурпурних, деяких зелених бактеріях та геліобактеріях.
Виділяють три етапи фотосинтезу: фотофізичний, фотохімічний та хімічний. На першому етапі відбувається поглинання фотонів світла пігментами, їх перехід в збуджений стан і передача енергії до інших молекул фотосистеми. На другому етапі відбувається розділення зарядів в реакційному центрі, перенесенняелектронів по фотосинтетичному електронотранспортному ланцюзі, що закінчується синтезом АТФ і НАДФН. Перші два етапи разом називають світлозалежною стадією фотосинтезу. Третій етап відбувається вже без обов'язкової участі світла і включає біохімічні реакції синтезу органічних речовин з використанням енергії, накопиченої на світлозалежній стадії. Найчастіше в якості таких реакцій розглядається цикл Кальвіна і глюконеогенез, утворення цукріві крохмалю з вуглекислого газу повітря.
Просторова локалізація
Фотосинтез рослин здійснюється в хлоропластах, відособлених двомембранних органелах клітини. Хлоропласти можуть бути в клітинах плодів, стебел, проте основним органом фотосинтезу, анатомічно пристосованим до його здійснення, є листя. У листку найбагатша хлоропластами тканина – палісадна, або фотосинтезуюча/стовпчаста/хлорофілоносна, паренхіма. У деяких сукулентів з виродженим листям (наприклад, кактусів) основна фотосинтетична активність пов'язана із стеблом.
Світло для фотосинтезу захоплюється повніше завдяки плоскій формі листка, що забезпечує велике відношення поверхні до об'єму. Вода доставляється зкореня розвиненою мережею судин (прожилків листка). Вуглекислий газ надходить частково за допомогою дифузії через кутикулу і епідерміс, проте велика його частина дифундує в листя через продихи і по листку через міжклітинний простір. Рослини, що здійснюють С4 і CAM фотосинтез, сформували особливі механізми для активної асиміляції вуглекислого газу.
Внутрішній простір хлоропласта заповнений безбарвною речовиною (стромою) і пронизаний мембранами (ламелами), які, з'єднуючись один з одним, утворюютьтилакоїди, які у свою чергу групуються в стопки, так звані грани. Внутрішньотилакоїдний простір відокремлений і не сполучається з рештою строми, передбачається також, що внутрішній простір всіх тилакоїдов сполучений між собою. Світлові стадії фотосинтезу повязані з мембранами, автотрофна фіксаціявуглекислого газу відбувається в стромі.
У хлоропластах є свої ДНК, РНК, рибосоми (70S типу), йде синтез білка (хоча цей процес і контролюється з ядра). Вони не синтезуються знов щоразу, а утворюються шляхом поділу попередніх. Все це дозволило вважати їх попередниками вільних ціанобактерій, що увійшли до складу еукаріотичної клітини в процесі симбіогенезу.
Ціанобактерії та інші фотосинтетизуючі бактерії та археї, таким чином, самі виконють функції хлоропластів рослин, і фотосинтетичний апарат їх клітин не винесений в особливу органелу. Їхні тилакоїди, проте, не утворюють стопок, а формують різні складчасті структури (у однієї ціанобактерії Gloeobacter violaceus тилакоїди відсутні зовсім, а весь фотосинтетичний апарат знаходиться на цитоплазматичній мембрані, що не створює заглибин). У них і в рослин також є відмінності в світлозбираючому комплексі (див. нижче) і в складі пігментів.
Світлова (світлозалежна) стадія
В ході світлової стадії фотосинтезу утворюються високоенергетичні продукти: аденозинтрифосфат, що служить в клітині джерелом енергії, і НАДФН, що використовується як відновник. Як побічний продукт виділяється кисень
Фотохімічна суть процесу

Двостадійний фотосинтез («Z-схема»)
Хлорофіл має два рівні збудження (з цим пов'язана наявність двох максимумів на його спектрі поглинання): перший пов'язаний з переходом на вищий енергетичний рівень електрона системи зв'язаних подвійних зв'язків, другий, – із збудженням неспарених електронів азоту і кисню порфіринового ядра. При незмінному спіні електрона формуютьсясинглетні перший і другий збуджений стан, при зміненому – триплетні перший і другий.
Другий збуджений стан найбільш високоенергетичний, нестабільний, і хлорофіл за 1012 с переходить з нього на перший, з втратою 100 кДж/моль енергії тільки у вигляді тепла. З першого синглетного і триплетного станів молекула може переходити в основний з виділенням енергії у вигляді світла (флуоресценція) або тепла, з перенесенням енергії на іншу молекулу, або, оскільки електрон на високому енергетичному рівні слабко зв'язаний з ядром, з переносом електрона на іншу сполуку.
Друга можливість реалізується в світлозбиральних комплексах, перша – в реакційних центрах, де переведений в збуджений стан при поглинанні кванта світла хлорофіл стає донором електрона (відновлювачем) і передає його на первинний акцептор. Щоб запобігти поверненню електрона на позитивно заряджений хлорофіл, первинний акцептор передає його вторинному. Крім того, час життя отриманих сполук вищий, ніж у збудженої молекули хлорофілу. Відбувається стабілізація енергії і розділення зарядів. Для подальшої стабілізації вторинний донор електронів окислює позитивно заряджений хлорофіл, первинним донором же у разі оксигенного фотосинтезу є вода.
Проблемою, з якою стикаються при цьому організми з оксигенним типом фотосинтезу, є відмінність окислювально-відновних потенціалів води (E0=+0,82 В) і НАДФ+ (E0=-0,32 В). Хлорофіл при цьому повинен мати в основному стані потенціал, більший за +0,82, В щоб окиснювати воду, але при цьому мати у збудженому стані потенціал менший, ніж 0,32, В щоб відновлювати НАДФ+. Одна молекула хлорофілу не може відповідати обом вимогам. Тому сформувалися дві фотосистеми і для повного проведення процесу необхідно два кванти світла і два хлорофіли різних типів.
Світлозбиральні комплекси
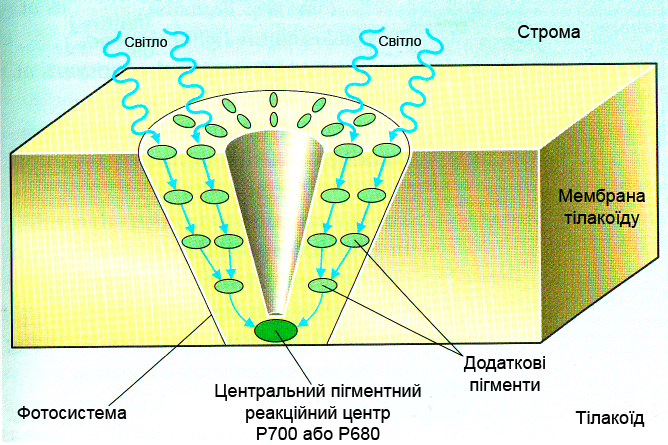
Світлозбиральний кластер фотосинтетичних пігментів на поверхні тилакоїдної мембрани хлоропластів
Хлорофіл виконує дві функції: поглинання і передачу енергії. Більше 90% всього хлорофілу хлоропластів входить до складу світлозбиральних комплексів (СЗК), що виконують роль антени, яка передає енергію до реакційного центру фотосистем I або II. Крім хлорофілу, в СЗК є каротиноїди, а у деяких водоростей і ціанобактерій – фікобіліни, роль яких полягає в поглинанні світла тих довжин хвиль, які хлорофіл поглинає порівняно слабо.
Передача енергії йде резонансним шляхом (механізм Ферстера) і займає для однієї пари молекул 1010-1012 сек., відстань, на яку здійснюється перенесення, становить близько 1 нм. Передача супроводжується деякими втратами енергії (10% від хлорофілу а до хлорофілу b, 60% від каротиноїдів до хлорофілу), через що можлива тільки від пігмента з максимумом поглинання при меншій довжині хвилі до пігмента з більшою довжиною хвилі в максимумі поглинання. Саме у такому порядку взаємно локалізуються пігменти СЗК, причому найбільш довгохвильові хлорофілли знаходяться в реакційних центрах. Зворотний перехід енергії неможливий.
СЗК рослин розташований в мембранах тилакоїдів, у ціанобактерій основна його частина винесена за межі мембран у прикріплені до них фікобілісоми – паличкоподібні поліпептидно-пігментні комплекси, в яких знаходяться різні фікобіліни: на периферії фікоеритрини (з максимумом поглинання при 495–565 нм), за ними фікоціаніни (550–615 нм) і алофікоціаніни (610–670 нм), що послідовно передають енергію на хлорофіл а (680–700 нм) реакційного центру.
Основні компоненти електронно-транспортного ланцюжка
Фотосистема II
Фотосистема – сукупність СЗК, фотохімічного реакційного центру і переносників електрона. Світлозбиральний комплекс II містить 200 молекул хлорофілу а, 100 молекул хлорофілу b, 50 молекул каротиноїдів і 2 молекули феофітину. Реакційний центр фотосистеми II є пігмент-білковим комплексом, розташованим в тилакоїдних мембранах і оточеним СЗК. У ньому знаходиться димер хлорофілу а з максимумом поглинання при 680 нм (П680). На нього кінець-кінцем передається енергія кванта світла з СЗК, внаслідок чого один з електронів переходить у вищий енергетичний стан, зв'язок його з ядром ослаблюється, і збуджена молекула П680 стає сильним відновником (E0=-0,7 В).
П680 відновлює феофітин, надалі електрон переноситься на хінони, що входять до складу ФС II, і далі на пластохінони, транспортується у відновленій формі до b6f комплексу. Одна молекула пластохинона переносить 2 електрони і 2 протони, які беруться зі строми. Заповнення електронної вакансії в молекулі П680 відбувається за рахунок води. До складу ФС II входить водоокислюючий комплекс, що містить в активному центрі іони марганцю в кількості 4 штук. Для утворення однієї молекули кисню потрібно дві молекули води, що дають 4 електрони. Тому процес проводиться в 4 такти, і для його повного здійснення потрібно 4 кванти світла. Комплекс розташований з боку внутрішньотилакоїдного простору, і отримані 4 протони викидаються в нього.
Таким чином, сумарний результат роботи ФС II – це окислення 2 молекул води за допомогою 4 квантів світла з утворенням 4 протонів у внутрішньотилакоїдному просторі і 2 відновлених пластохинонів в мембрані.
b6f або b/f-комплекс
b6f комплекс є насосом, що перекачує протони із строми у внутрішньотилакоїдний простір і створює градієнт їхньої концентрації за рахунок енергії, що виділяється в окисно-відновних реакціях електронно-транспортного ланцюжка. 2 пластохинони забезпечують перекачування 4 протонів. Надалі трансмембранний протонний градієнт (pH строми, близький до 8, внутрішньотилакоїдного простору – 5) використовується для синтезу АТФ трансмембраннимферментом АТФ-синтазою.
Фотосистема I
Світлозбиральний комплекс I містить приблизно 200 молекул хлорофілу.
У реакційному центрі першої фотосистеми знаходиться димер хлорофілу а з максимумом поглинання при 700 нм (П700). Після збудження квантом світла він відновлює первинний акцептор – хлорофіл а, той – вторинний (вітамін K1), після чого електрон передається на філохінон, від нього на ферредоксин, який і відновлює НАДФ за допомогою ферменту ферредоксин-НАДФ-редуктази. Білок пластоціанін, окислений в b6f комплексі, транспортується до реакційного центру першої фотосистеми з боку внутрішньотилакоїдного простору й відновлює окислений П700.
Циклічний і псевдоциклічний транспорт електрона
Крім повного нециклічного шляху електрона, описаного вище, виявлено циклічний і псевдоциклічний. Суть циклічного шляху полягає в тому, що феридоксин замість НАДФ відновлює пластохинон, який переносить його назад на b6f комплекс. В результаті утворюється більший протонний градієнт і більше АТФ, але не виникає НАДФН. При псевдоциклічному шляху феридоксин відновлює кисень, який надалі перетворюється на воду і може бути використаний у фотосистемі II. При цьому також не утворюється НАДФН.
Темнова стадія

Схематичне зображення циклу Кальвіна
У темновій стадії за участю АТФ і НАДФН відбувається відновлення CO2 до глюкози. Хоча світло не потрібне для здійснення даного процесу, воно бере участь у його регуляції.
С3 – фотосинтез, цикл Кальвіна
Цикл Кальвіна, або відновлювальний пентозо-фосфатний цикл, складається з трьох стадій:
- Карбоксиляція
- Відновлення
- Регенерація акцептора CO2
На першій стадії до рибулозо-1,5-бісфосфату приєднується CO2 під дією ферменту рибулозобісфосфат-карбоксилаза/оксигенази (Rubisco). Цей білок складає основну фракцію білків хлоропласту і, ймовірно, найбільш поширений фермент в природі. В результаті утворюється проміжне нестійке з'єднання, що розпадається на дві молекули 3-фосфогліцеринової кислоти (ФГК). У другій стадії ФГК в два етапи відновлюється. Спочатку вона фосфорилюєтся АТФ під дією фосфорогліцерокінази, потім НАДФН при дії тріозофосфатдегідрогенази, її карбоксильна група окислюється до альдегідної і вона стає вуглеводом (ФГА).
У третій стадії беруть участь 5 молекул ФГА, які через утворення 4-, 5-, 6- і 7-вуглецевих зв'язків об'єднуються в 3 5-вуглецевих рибулезо-1,5-біфосфати, для чого необхідні 3 АТФ. Нарешті, дві ФГА необхідні для синтезу глюкози. Для утворення однієї молекули глюкози потрібно 6 обертів циклу, 6 CO2, 12 НАДФН і 18 АТФ.
С4 – фотосинтез, цикл Хетча-Слека-Карпілова
Див. також: Цикл Хетча-Слека-Карпілова
При низькій концентрації розчиненого в стромі CO2 Rubisco каталізує реакцію окислення рибулезо-1,5-біфосфату і його розпад на 3-фосфогліцеринову кислоту і фосфогліколеву кислоту, яка вимушено використовується в процесі фотодихання. Для збільшення концентрації CO2 рослини С4 типу змінили анатомію листка. Цикл Кальвіна у них локалізується в клітинах обкладання провідного пучка, в клітинах мезофілу ж під дією ФЕП-карбоксилазифосфоенолпіруват карбоксилуєтся з утворенням щавелеоцетової кислоти, яка перетворюється на малат або аспартат і транспортується в клітини обкладання, де декарбоксилуєтся з утворенням пірувату, що повертається в кліини мезофіла.
С4 фотосинтез практично не супроводжується втратами рибулезо-1,5-біфосфату з циклу Кальвіна, тому ефективніший. Проте він вимагає не 18, а 30 АТФ на синтез 1 молекули глюкози. Це виправдовує себе в тропіках, де жаркий клімат вимагає тримати продихи закритими, що перешкоджає надходженню CO2 в листок, а також при рудеральній життєвій стратегії.
САМ фотосинтез
При CAM (Crassulaceae acid metabolism) фотосинтезі відбувається розділення асиміляції CO2 і циклу Кальвіна не в просторі, як у С4, а в часі. Вночі у вакуолях клітин по аналогічному описаному вище механізму при відкритих продихах накопичується малат, вдень при закритих продихах йде цикл Кальвіна. Цей механізм дозволяє максимально економити воду, проте поступається в ефективності і С4, і С3. Він виправданий при стрестолерантній життєвій стратегії.
Аноксигенний фотосинтез
Аноксигенний фотосинтез властивий деяким бактеріям та археям (наприклад, пурпурним, деяким зеленим бактеріям та геліобактеріям тощо). Ці організми не використовують води у якості відновника, тому кисень (O2) не є побічним продуктом синтезу. Замість води використовуються як сірководень (H2S) або йони двовалентного заліза (Fe++), унаслідок чого на виході виникають елементарна сірка (S) і тривалентні іони заліза (Fe+++), відповідно, або молекулярний водень (H2). Наприклад, фотосинтез з використанням H2S у якості відновника проходить такі стадії:
CO2 + 2 H2S (CH2O) + 2 S + H2O
Тут першим продуктом фотосинтезу виступає фіктивна хімічна сполука CH2O.
Значення фотосинтезу
Фотосинтез є основним джерелом біологічної енергії, фотосинтезуючі автотрофи використовують її для утворення органічних речовин з неорганічних, гетеротрофи існують за рахунок енергії хімічних зв'язків, запасеної автотрофами, вивільняючи її в процесах аеробного та анаеробного дихання. Енергія, отримувана людством при спалюванні викопного палива (вугілля, нафта, природний газ, торф), також є запасеною в процесі фотосинтезу.
Фотосинтез є головним методом залучення неорганічного вуглецю в біологічний цикл. Весь кисень атмосфери біогенного походження є побічним продуктом фотосинтезу. Формування кисневої атмосфери повністю змінило стан земної поверхні, зробило можливою появу дихання, а надалі, після утворення озонового шару, дозволило життю вийти на сушу.
Листопад