ГЕНЕТИКА ПОПУЛЯЦІЙ
Лекція 13 ГЕНЕТИКА ПОПУЛЯЦІЙ
Поняття «популяція» і «чиста лінія» запропоновані В. Йоганнсеном у 1907р. Популяція — сукупність тварин одного виду, породи, які вільно схрещуються між собою, що населяє конкретну територію і достатньо ізольована від інших груп. Популяцією можна вважати кожне стадо однієї породи, якщо тварин розводять без схрещування з іншими породами .чи іншими стадами. Тому в одному господарстві можуть бути кілька популяцій одного виду.
Метою генетики популяцій є вивчення закономірностей розподілу співвідношень генотипів () у популяціях, визначення концентрації домінантних і рецесивних генів, оскільки біологічне й господарсько корисне значення гомозиготних домінантних, гетерозиготних і гомозиготних рецесивних тварин різне. Вважають, що найкращі гетерозиготні форми ті, серед, яких може виникати гетерозис. На другому місці — гомозиготні домінантні і на останньому — гомозиготні рецесивні, як найменш пристосовані до умов зовнішнього середовища, часто мають низьку життєздатність. Тобто генетично популяція — це сукупність тварин з різними генотипами.
На противагу популяції чиста лінія — це потомки однієї гомозиготної самозапильної рослини, і природно, що ці потомки гомозиготні. Однак гомозиготність чистих ліній також відносна, тому що самозапилення не виключає можливість перехресного запилення і, тим більше, появи мутацій. У тварин чистих ліній взагалі не буває, хоч близькоспоріднене схрещування веде до збільшеная гомозиготності, проте вона стопроцентною не буває.
1. ВІДМІННОСТІ В ЕФЕКТИВНОСТІ ДОБОРУ В ПОПУЛЯЦІЯХ І ЧИСТИХ ЛІНІЯХ
В. Йоганнсен, вивчаючи мінливість квасолі за розміром, масою насіння і впливом добору на ці показники, встановив, що добір у чистах лініях неефективний. Він підбирав і висівав окремо великі і маленькі насінини, а потім проаналізував одержаних потомків і переконався в тому, що за розмірами і масою вони в середньому однакові в обох групах. Це пояснюється тим, що всі потомки чистої лінії першого і наступних поколінь однакові за генотипом, тобто генетичні можливості їх рівні, і відмінності щодо фенотипу пов'язані з модифікаціями, які виникли під впливом умов зовнішнього середовища.
У популяції, що є сукупністю особин з різними генотипами, генетичні можливості яких різні, добір ефективний. Серед відібраних великих квасолин переважатимуть генотипи , а серед маленьких генотипи . Тому посаджені великі квасолини дадуть у середньому більші за розмірами квасолини, ніж маленькі.
2. ЗАКОНОМІРНОСТІ ГЕНЕТИЧНОЇ СТРУКТУРИ ПОПУЛЯЦІЙ
Кожна популяція має свій генофонд, тобто сукупність алелів, що входять до її складу, і своє співвідно відношення генотипів (). Виникає питання: чи співвідношення генотипів стале і підкоряється якимось закономірностям, чи залежить від випадкових факторів?
Припустимо, що в популяції кількість особин з генотипами однакова, а звідси гамет з домінантним А і рецесивним а генами в такій популяції порівну: 0,5А і 0,5а. При вільному схрещуванні особин можливі такі комбінації гамет:
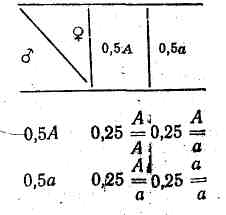
Звідси видно, що домінантних гомозигот буде. 0,25, гетерозигот — 0,50 і гомозигот рецесивних — 0,25. При подальшому схрещуванні таких потомків між собою співвідношення зазначених генотипів має бути таким самим, тому що гамет із домінантним геном А утвориться 0,5 (0,25 від гомозигот +0,25 від гетерозигот ), гамет з рецесивним геном а також 0,5 (0,25 від гомозигот +0,25 від гетерозигот ). Тобто в такій популяції співвідношення генотипів має бути 1 : 2 : 1 протягом усього часу існування популяції, якщо ніякі фактори на зміну цього співвідношення не впливатимуть.
Візьмемо іншу популяцію, в якій кількість особин з генотипами і неоднакова. Наприклад, якщо особин з генотипом 80, а з генотипом 20, тоді відповідно гамет з домінантним геном А буде 0,8, а з рецесивним геном а — 0,2 і при їх вільному схрещуванні будуть такі комбінації гамет:
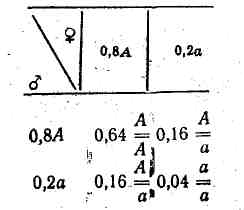
Співвідношення генотипів буде таке: 0,64 : 0,32 : 0,04 . У наступних поколіннях таке співвідношення генотипів має залишитись, оскільки гамет із домінантним геном А буде утворено 0,8 (0,64 від +0,16 від ), а з рецесивним геном а — 0,2 (0,04 від +0,16).
Як видно, співвідношення генотипів у кожній популяції має бути на характерному для неї рівні, тобто кожна популяція перебуває у певній рівновазі за парою алельних. Цю закономірність помітили англійський математики Г. Харді і німецький Лікар В.Вайнберг незалежно один від одного.Їх можна виразити у вигляді формули
р2 + 2 рq + q2 , звідки р = А; q = а.
Із цієї самої формули випливає, що кількість гомозиготних домінантних особин дорівнює квадрату частоти домінантного гена (р2), кількість гомозиготних рецесивних особин дорівнює квадрату частоти рецесивного гена (q2) і кількість гетерозиготних особин дорівнює подвоєному добутку частот обох алелів (2 рq).
Однак постійно однакове співвідношення генотипів могло б бути тільки в ідеальних неіснуючих популяціях, де не було б впливу різних селективних переваг, тобто факторів, що порушують ідеальність популяцій. До них належать: мутаційний тиск, який виникає внаслідок того, що частота прямих Аа і зворотних аА мутацій різна, звідки й зрушення у бік збільшення концентрації одного гена і зменшення — іншого; вплив природного і штучного добору на структуру популяцій.
Як уже зазначалось, різні генотипи мають різне біологічне і господарсько корисне значення. Для них також характерні неоднакові плодючість і живучість. Проводячи штучний добір, людина відбирає кращі фенотипи, серед яких найчастіше будуть домінантні гомозиготні і гетерозиготні генотипи, а гомозиготні рецесивні, як правило, вибраковуються. Вони () найчастіше нівелюються природним добором, бо мало пристосовані до умов зовнішнього середовища і мають знижену плодючість. Природно, що популяція зміщуватиметься у бік збільшення кількості особин з генотипами і ; вільне схрещування, яке деякою мірою обмежене навіть у диких популяціях, а в культурних зовсім неможливе при штучному осіменінні; генетичний дрейф генів — різка зміна концентрації окремих генів завдяки випадковим змінам чисельності популяції; міграція, тобто недостатня ізоляція популяцій, пов'язана із завезенням плідників або їхньої сперми з інших зон країни або з-за кордону; інбридинг — близькоспоріднене схрещуваний, що веде до збільшення гомозиготності.
Крім цих, можна виділити ще ряд факторів, які можуть порушувати співвідношення генотипів у популяціях. Слід підкреслити, що чисельність популяції має бути великою, щоб не позначився вплив випадкових відхилень. Тому про постійну рівновагу генотипів у популяціях як природних, так і штучних не може бути й мови.
Закон Харді — Вайнберга, який можна назвати законом рівноваги генних концентрацій у вільно схрещуваних (панміктичних) популяціях, має велике значення для розуміння механізму еволюційних змін. Крім того, користуючись формулою Харді — Вайнберга, можна вирішувати практичні завдання. Наприклад, спадкове захворювання контрактура (вкорочення) м'язів у великої рогатої худоби, яке призводить до загибелі телят, зумовлене рецесивним геном а. Хворі телята мають генотип , здорові—, а здорові і носії рецесивного гена — . Важливо знати кількість гетерозиготних особин, бо за фенотипом вони не відрізняються від гомозиготних . За допомогою формули Харді—Вайнберга можна визначити співвідношення генотипів і концентрацію окремо кожного гена — А і а. Наприклад, якщо з 350 новонароджених телят 7 мали контрактуру м'язів, то частота виникнення захворювання в даній популяції дорівнює 7/350=0,02, а це і є величина q2 з формули закону Харді — Вайнберга. Потім обчислюють значення q===0,14. Оскільки р+ q= 1, то р=1- q = 1—0,14==0,86. Знаючи величини q =0,14 і р=0,86, визначають співвідношення генотипів у популяції:
0,86+2*0,86*0,14 + 0,142 = 0,7396 + 0,2404 + 0,02 ,
тобто гомозиготних домінантних особин 74 %, що становить 259 особин, гетерозиготних 24 % — 84 особини і гомозиготних рецесивних 2 % — 7 особин. Далі можна обчислити концентрацію генів А і а за формулами
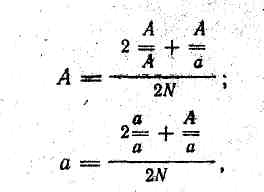
де — кількість гомозиготних домінантних особин; — кількість гетерозиготних особин; — кількість гомозиготних рецесивних особин;
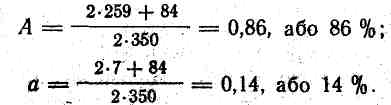
Однак у зв'язку з вибракуванням хворих гомозиготних рецесивних тварин співвідношення генотипів змінюється, оскільки зменшується кількість гамет з рецесивним геном. Так, гамет із геном А залишиться та сама кількість (0,86), а з рецесивним а — 0,12 замість 0,14 і при вільному схрещуванні вони дадуть такі комбінації:
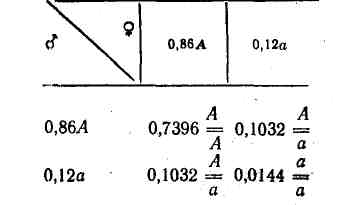
Співвідношення генотипів таке: 0,7396, або 73,96 %, тобто однакова кількість; 0,2064, або 20,64 % замість 0,2404, або 24,04 % : 0,144, або 1,44 % замість 0,02, або 2,0 % в попередньому поколінні.
Генетична структура популяції змінилась, проте при панміксії (вільному схрещуванні) уже в наступному поколінні, якщо вибракування немає, вона стабілізується на тому рівні, який встановився після вибракування. Схрещування, яке відновлює генетичну рівновагу, називається стабілізуючим.
Як уже зазначалось, ідеальних популяцій не існує, тому що багато факторів порушують співвідношення генотипів. Проте кожна біологічна система, в тому числі й популяція, має певні адаптивні генетичні механізми, за допомогою яких підтримує своє існування. Механізм підтримання генетичної рівноваги популяцій пов'язаний з утворенням постійної кількості гамет з домінантним і рецесивним генами в кожній конкретній популяції і явищами гетерозиготності і поліморфізму. Якщо гетерозиготність забезпечує кращу пристосовуваність до зміни умов середовища, то відбувається добір на користь гетерозигот, а це, в свою чергу, веде до збалансованого поліморфізму, тобто певного співвідношення генотипів, певного темпу і напрямку мутаційного процесу. Зростання рівня природної радіації, збільшення хімічних мутагенів викликає збільшення темпів мутаційного процесу, завдяки якому краще працюють механізми пристосування — гомеостазу.
3. ЕФЕКТИВНІСТЬ ДОБОРУ ДОМІНАНТНИХ І РЕЦЕСИВНИХ ОЗНАК
Якщо залишати особини з домінантними ознаками і видаляти з рецесивними, то частка гомозигот домінантних збільшиться, а рецесивних зменшиться. При цьому гетерозиготи все-таки залишаться і протягом тривалого часу відбуватиметьея «вищеплення» гомозиготних рецесивних генотипів . Якщо видаляти особини з домінантними ознаками, а залишати з рецесивними, то такий добір приводить до майже повного видалення домінантних гомозигот і гетерозигот та різкого збільшення гомозигот рецесивних . Такий напрямок добору використовується в хутровому звірівництві для одержання бажаного забарвлення хутра. Добір, спрямований на збільшення частий гетерозиготних особин, дає деяке збільшення їх у перших поколіннях, а потім залишається .на одному рівні. Наприклад, у каракульських сірих овець гомозиготні домінантні особини нежиттєздатні, тому схрещують гетерозиготних сірих з гомозиготними чорними й одержують знову гетерозиготних сірих і гомозиготних рецесивних чорних.
За даними С. Райта і Д. Лаша, на ефективність добору впливає частота алелю. Так, якщо частота бажаного алелю в популяції становить 0,3—0,7, то ефективність селекції висока, а якщо вона менша за 0,3 або більша за 0,7, то ефективність селекції значно нижча.
Ефективність добору значною мірою залежить від впливу факторів зовнішнього середовища.
Певна частина кількісних ознак, з якими має справу зооінженер (надій молока, вміст жиру в молоці, несучість курей, скороспілість свиней, настриг вовни від вівці, здатність до відгодівлі тощо), залежить від умов зовнішнього середовища. Тому виявити потенціальну можливість генотипу (норму реакції генотипу) можна тільки за оптимальних умов годівлі й утримання. За.даними багатьох дослідників, при недостатній годівлі й утриманні в поганих умовах корови високопродуктивної породи дають молока стільки, скільки корови низькопродуктивної породи, а то й менше. Прикладом впливу зовнішнього середовища на ефективність добору є досвід роботи вирощування молодняка на холоді у племзаводі «Караваєво». Встановлено, що телички, які народжувались узимку й утримувались при мінусових температурах, після того як стали коровами давали молоко з вищою жирністю. Запровадження методу вирощування молодняка на холоді сумісно з інтенсивним добором дало змогу за 15 років підвищити на 0,43 % жирність молока в стаді, тоді як за звичайних умов утримання вона підвищилась тільки на 0,1 %.
4. ВИКОРИСТАННЯ У ТВАРИННИЦТВІ ДОСЯГНЕНЬ ПОПУЛЯЦІЙНОЇ ГЕНЕТИКИ
Знання ролі різних факторів (мутацій, природного і штучного добору, міграцій, дрейфу генів, інбридингу тощо), які впливають на генетичну структуру популяцій, слід використовувати для досягнення бажаних співвідношені, генотипів. Треба вчасно виявити шкідливі мутації і проводити належну генетико-селекційну роботу (підбір батьківських пар, суворе й регулйрне вибраковування та ін.) протягом кількох поколінь, щоб очистити популяцію від шкідливих мутацій, і вести планомірну роботу, спрямовану на виявлення корисних мутацій і розмноження їх. Особливий генетичний нагляд треба встановити за плідниками, адже в умовах штучного осіменіння шкідливі мутації можуть швидко поширюватися.
Враховуючи те, що гетерозис може проявитись найчастіше у гетерозиготних за генотипом особин, потрібно спрямовувати селекційну роботу на збільшення гетерозиготності популяцій. У селекційній роботі треба звертати увагу на вплив факторів зовнішнього середовища, а також на здатність окремих тварин пристосовуватись до нової промислової технології і вести правильний добір у цьому напрямку.
Контрольні питання
- Що таке популяція і які бувають популяції?
- Поняття чистої лінії.
- Закономірності генетичної структури популяцій і чистих ліні.
- Як. визначити співвідношення генотипів у популяції і частоти генів?
- Яке схрещування називають стабілізуючим?
6. Використання у тваринництві популяційної генетики.
7. Ефективність добору домінантних і рецесивних ознак.
PAGE 8
ГЕНЕТИКА ПОПУЛЯЦІЙ