Закономірності успадкування ознак при статевому розмножені
PAGE 6
Лекція 5. Закономірності успадкування ознак
при статевому розмножені
1. Відкриття Г.Менделем законів спадковості
2. Символіка позначення генетичних ознак
3. Правила спадковості – закони Менделя Успадкування ознак при моно-, ди- та полігібридних схрещуваннях.
4. Види домінування
Термінологічний словник: домінантність (лат.) – пануючий; рецесивність (лат.) – відступ, віддалення:, альтернатива (лат.) – один із двох; гомозиготність (гр.) – гомо – однаковий, та зигота з'єднаний докупи; гетерозиготність (гр.) – гетеро – інший та зигота; генотип (гр.) – ген – рід, походження та тип; фенотип (гр.) – фено - показую, виявляюсь, проявляюсь та тип; моногібрид (гр.) – моно – один, гібрид (лат.) – помісь, реципрокний (лат.) – зворотний, взаємний; леталь (лат.) – смерть;
1. Відкриття Г.Менделем законів спадковості
Вперше у 1886 році Грегор Мендель – чеський вчений, монах, використав системний науковий підхід у вивченні механізму успадкування ознак. Для досліджень він вибрав 22 сорти садового горошку, які у вихідних формах відрізнялися за сімома парами контрастних ознак. Мендель вперше використав алгебру в генетиці, позначивши гени (ознаки) у генетичних схемах буквами латинського алфавіту. Він запропонував гібридологічний метод, до основних особливостей якого відносяться:
1. Використання у вигляді основних форм для схрещування альтернативних властивостей із порівняно невеликою кількістю ознак (1,2,3) і ретельне врахування характеру успадкування кожної із них.
2. Точний кількісний облік гібридних особин, які різняться за окремими ознаками в ряді наступних поколінь.
3. Індивідуальний аналіз потомства від кожної батьківської особини в ряді наступних поколінь.
Гібридологічний метод, пізніше одержав назву генетичного аналізу, який і тепер використовується у селекційній практиці.
2. Символіка позначення генетичних ознак
Символи, які використовуються при генетичному аналізі:
1. Процес схрещування позначають знаком Х.
2. При написанні схеми схрещування на першому місці ставлять жіночу стать, яка позначається знаком (дзеркало богині краси Венери), чоловічу стать позначають знаком (щит і спис бога війни Марса).
3. Батьківські організми, взяті для схрещування, позначаються буквою Р (перша літера латинського слова Pacenta – батьки).
4. Потомство від цих батьків позначають для скорочення літерою F (перша літера латинського слова Filli – діти) з цифровим нижнім індексом, який відповідає порядковому номеру гібридного покоління (F1; F2; F3 і так далі).
Для успішного засвоєння основ генетики слід добре знати вихідні поняття:
Алель – один з двох чи більше альтернативних форм гена, кожний з яких характеризується унікальною послідовністю нуклеотидів.
Алельні гени позначають однією і тією ж буквою – домінантний великою, рецесивний – малою. Наприклад, А і а.
Гетероалель – алель, який відрізняється від інших алелів того ж гена за нуклеотидною послідовністю в різних ділянках гена.
Гомозиготним – називають організм, в гомологічних хромосомах якого одночасно присутні однакові алелі однієї чи кількох пар алельних генів (АА, ВВ, аа, bb).
Гетерозиготним – називають стан гібридного організму, гомологічні хромосоми якого мають різні алелі тієї чи іншої пари алельних генів (Аа, Вв та ін.).
Гетерозиготними можуть бути організми не тільки за однією, але, як правило за багатьма ознаками. Наприклад, за двома – Аа; Вb; за трьома – Аа; Вb; Сс і т.д.
У гетерозигот кількість гамет відповідає формі 2n, де 2n – кількість (двох) типів за будь-якою ознакою, а n – число альтернативних ознак.
Генотип – сума всіх спадкових властивостей організму.
Фенотип – реалізація генотипу в конкретних умовах зовнішнього середовища.
Решітка Пеннета – досить зручний засіб записування результатів – схрещування пар, запропонований на початку ХІХ ст. англійським вченим Пеннетом.
3.Правила спадковості – закони Менделя. Успадкування ознак при моно-, ди- та полігібридних схрещуваннях
Г.Мендель, схрещуючи форми гороху, які розрізнялися за одною або двома вибраним ознакам, проаналізував І і ІІ покоління потомства, а також результати схрещування гібридів з вихідними формами (зворотне схрещування). Чітка повторюваність числових співвідношень форм з різними ознаками у всіх варіантах схрещування дозволили йому сформулювати три правила успадкування, які в подальшому одержали назву законів успадкування Менделя.
Вони є наступними:
1.Закон домінування, або одноманітності гібридів першого покоління. Він стверджує: при схрещуванні гомозигот домінантної і рецесивної (незалежно від альтернативних ознак) у першому поколінні все потомство має ознаки домінантного батька. Розщеплення за фенотипом 1 : 1.
Зразок запису:
PP AA x aa
Типи гамет а
|
Aa |
Типи гамет А
2.Закон розщеплення, відповідно до якого при схрещуванні гібридів І покоління, гетерозиготних за кожним алелем, що визначається, відбувається розщеплення за фенотипом (3 : 1)n, а за генотипом (1 : 2 : 1)n.
Зразок запису:
F1 Aa x Aa
Чоловічі типи гамет
А а
|
АА |
Аа |
|
Аа |
аа |
А
Жіночі типи
гамет
а
3.Правило незалежного успадкування ознак. В основі цього явища лежить вільне поєднання генів, які зумовлюють різні ознаки гібридів. Їх Мендель одержав при схрещуванні гороху з двома парами алельних ознак у другому поколінні. Це правило називають також законом чистоти гамет – явище не змішування генів кожної пари алелів нативних ознак.
Нативний – який перебуває в природному стані, не модифікований, який зберіг структуру, властиву йому в живій клітині.
В основі даного явища лежить цитологічний механізм мейозу. Дійсно, якщо розглянути гетерозиготу за однією ознакою Аа або за двома, трьома і більше ознаками, то очевидно, що кожна гетерозигота створює “чисті” гамети за формулою 2n, а саме
А
Аа Два типи гамет
а
Оскільки закони генетики ґрунтуються на схрещуванні, яке є основним методом створення нових порід тварин і сортів рослин, розглянемо його детальніше.
Схрещування – це природне або штучне сполучення двох спадково різних статевих клітин при запліднені. Нащадків, що виникли від схрещування двох особин, називають гібридами.
Залежно від кількості пар альтернативних спадкових ознак, за якими різняться батьківські форми, схрещування буває моногібридним, дигібрид-ним і полігібридним.
Моногібридне схрещування. Називають таке схрещування, при якому батьківські форми різняться лише за парою альтернативних (контрастуючих, протилежних) ознак. Це найпростіше схрещування, розглянуте на прикладі законів домінування.
Для успішного визначення особливостей успадкування ознак, локалізованих у не гомологічних хромосомах, необхідно необхідно пам’ятати про загальну форму розщеплення у F2 за фенотипом і генотипом. При повному домінуванні розщеплення за фенотипом від схрещування гетеро-зиготних за всіма генами ознак становить (3 : 1)n, за генотипом – (1 : 2 : 1)n. Виходячи з цього кількість фенотипових класів становить 2n, поряд з тим як генотипових 3n.
Найбільш показовим прикладом моногібридного схрещування у ВРХ є спадковість чорної і червоної масті. Чорна масть домінує (В), а червона – рецесивна (b).
Гетерозигота за однією ознакою (Аа) утворює два типи гамет А та а, в той час як домінантна гомозигота утворює лише тип гамет А, а рецесивна – тільки тип а. (Рис. вище: типи гамет)
Слід відмітити, що відбір у селекції за однією ознакою є найефек-тивнішим, наприклад мазаєвська порода довгововнових тонкорунних овець, американські рисаки за жвавістю, молочна худоба Голландії. Проте тривалий односторонній відбір веде до селекційної депресії і насамперед впливає на показники життєдіяльності та плодючості.
Дигібридне схрещування. Схрещування організмів, які відрізняються двома парами неалельних генів називають дигібридним.
Використовуючи цей метод схрещування Мендель відкрив третій закон незалежного успадкування ознак. Так при схрещуванні гладенького сорту гороху, який мав жовте забарвлення насіння (ААВВ), з сортом, що мав зелене зморшкувате насіння (ааbb), гібриди F1 мали жовте гладеньке насіння, так як гладенька форма домінує над зморшкуватою, а жовте забарвлення над зеленим.
У F2 серед 556 одержаного насіння Мендель виявив 315 гладеньких жовтих, 101 зморшкуватих жовтих, 108 гладеньких зелених і 32 зморшкуватих зелених, що склало співвідношення за фенотипом 9 : 3 : 3 : 1. При розгляді цього співвідношення видно, що поряд з насінням, яке має поєднання ознак, що характерне для вихідних форм (гладенькі жовті і зморшкуваті зелені), у F2 з’явилося насіння з новими поєднаннями ознак – гладенькі зелені та зморшкуваті жовті, тобто виникла перекомбінація ознак вихідних форм.
Наприклад, якщо уявити, що домінантні фактори комолості та чорного забарвлення К та А у бугая абердин-ангуської породи розміщені у різних негомологічних хромосомах (а незалежний розподіл факторів має місце лише при цій умові), а фактори рогатості і червоної масті k та a у корів, то весь процес розщеплення можна описати за допомогою решітки Пеннета, дивись рис. 1 та 2.
Для зручності пара хромосом, у яких розміщені гени К мають вигляд чорних і зігнутих паличок, гени А – чорні і прямі палички, тоді як алелі а і k мають вигляд аналогічних паличок, але вони білі.
Тобто, хромосоми домінантних форм пофарбовані у чорний колір, а хромосоми рецесивних у білий.
Так як після мейозу число хромосом зменшується удвоє, то гамети домінантного батька мають генотип АК (чорні комолі), а гамети рецесивного – генотип аb (червоні рогаті).
Поєднання гамет дає зиготу F1 з генотипом АаКk ( за фенотипом - чорні комолі).
Під час мейозу у гібридів F1 батьківські і материнські хромосоми розходяться у дочірні клітини незалежно одна від одної, тому гаплоїдні статеві клітини з однаковою вірогідністю можуть мати як дві хромосоми одного із батьків (АК або аk), так і по одній хромосомі від кожного із них (Аk або аK).
Чоловічі та жіночі гамети цих чотирьох груп незалежно з’єднуються між собою, з однаковою вірогідністю створюючи зиготи, які виникають в результаті поєднання чоловічих і жіночих гамет з будь-яким із чотирьох генотипів: АK, Аk, аK та аk.
Решітка Пеннета дає уяву про вільне поєднання гамет і генотипів зигот, які виникають в результаті поєднання цих гамет. Зиготи виписані в нутрі решітки, створюють комбінаційний ряд, який складається із дев’яти членів, які відрізняються один від одного за генотипом. Частоту їх зустрічі можна записати таким чином: ААKK+ААkk+ааKK+ааkk+2ААKk+2ааKk+2АаKK+ 2Ааkk+4АаKk.
За фенотипом зиготи поділяються на чотири феногрупи: АK – чорні комолі, Аk – чорні рогаті, аK – червоні комолі, аk – червоні рогаті.
Після підрахунку і об’єднання у групи фенотипово подібних зигот одержуємо співвідношення 9АK : 3Аk : 3аK : 1аk.
Аналогічний приклад з горошком. Так як після мейозу число хромосом зменшується удвоє, то гамети домінантного батька мають генотип АВ (гладенькі жовті), а гамети рецесивного – генотип аb (зелені зморшкуваті).
Поєднання гамет дає зиготу F1 з генотипом АаВb (гладенькі жовті).
Під час мейозу у гібридів F1 батьківські і материнські хромосоми розходяться у дочірні клітини незалежно одна від одної, тому гаплоїдні статеві клітини з однаковою вірогідністю можуть мати як дві хромосоми одного із батьків (АВ або аb), так і по одній хромосомі від кожного із них (Аb або аВ).
Чоловічі та жіночі гамети цих чотирьох груп незалежно з’єднуються між собою, з однаковою вірогідністю створюючи зиготи, які виникають в результаті поєднання чоловічих і жіночих гамет з будь-яким із чотирьох генотипів: АВ, Аb, аВ та аb.
Решітка Пеннета дає уяву про вільне поєднання гамет і генотипів зигот, які виникають в результаті поєднання цих гамет. Зиготи виписані в нутрі решітки, створюють комбінаційний ряд, який складається із дев’яти членів, які відрізняються один від одного за генотипом. Частоту їх зустрічі можна записати таким чином: ААВВ+ААbb+ааВВ+ааbb+2ААВb+2ааВb+2АаВВ+ 2Ааbb+4АаВb.
За фенотипом зиготи поділяються на чотири феногрупи: АВ – гладенькі жовті, Аb – гладенькі зелені, аВ – зморшкуваті жовті, аb – зморшкуваті зелені.
Після підрахунку і об’єднання у групи фенотипово подібних зигот одержуємо співвідношення 9АВ : 3Аb : 3аВ : 1аb.
Найбільш загальну форму алгебраїчного виразу розщеплення при полігібридних схрещуваннях може представити в наступному вигляді:
Р AABBCCDDEE … x aabbccddee …
F1 AaBbCcDdEe …
F2 (A+a)2 (B+b)2 (C+c)2 (D+d)2 (E+e)2 …
Встановлені Менделем правила дозволили для кожного із найбільш складних випадків гібридизації визначити, яку кількість особин охоплює загальна формула, скільки у цій формулі членів і скільки із них будуть гомозиготні, а також число сортів гамет (рівне числу гомозиготних форм у F2), створених гібридами F1.
Згідно цих правил, якщо число пар ознак, за якими відрізняються вихідні форми, дорівнює n, то для F2 число особин у загальній формулі дорівнює 4n (4 у моногібридів, 16 у дигібридів, 64 у тригібридів, 256 у тетрагібридів і т.д.).
Число членів (число різних генотипів) дорівнює 3n (3 у моногібридів, 9 у дигібридів, 27 у тригібридів, 81 у тетрагібридів і т.д.).
Число гомозиготних форм (і число фенотипів при повному домінуванні) дорівнює 2n (2 у моногібридів, 4 у дигібридів, 8 у тригібридів, 16 у тетрагібридів і т.д.).
Число сортів гамет дорівнює 2n (2 у моногібридів, 4 у дигібридів, 8 у тригібридів, 16 у тетрагібридів і т.д.).
4. Види домінування
Домінантний ген у генетичному розумінні означає перевагу дії одного гена з пари алелів. В інтерпретації молекулярної генетики під домінантним геном розуміють такий структурний стан гена, який дозволяє йому контролювати синтез функціонально найбільш надійного білка, що забезпечує високу життєздатність і плодючість організму. У випадку зміни структури гена, як наслідок його мутації, під його контролем синтезується аномальний білок, не здатний забезпечувати високі фундаментальні можливості обміну речовин. Сформована таким чином ознака є рецесивною.
Домінантна – це ознака, що з’являється у гібрида і пригнічує розвиток іншої альтернативної ознаки.
Рецесивна ознака – пригнічується тільки в гомозиготному стані.
Явище домінантності й рецесивності універсальне для рослин, тварин і людини. Повна домінантність і рецесивність існують тільки як вкрай рідкі випадки, між якими існують проміжні форми домінантності.
1. Неповне домінування – результат внутрішньої взаємодії, що призводить до формування проміжної між домінантною і рецесивною формами ознаки.
2. Нестійка домінантність – прояв відповідних алелів у гетерозигот-ному стані мінливий. В одному генотиповому середовищі і при одних зовнішніх умовах він рецесивний, в іншому і при інших зовнішніх умовах – домінантний.
3. Альтернуюча домінантність – у процесі онтогенезу проявляється спочатку один, потім інший алель з гетерозиготної пари алелів.
4. Умовна домінантність – домінування гена, який проявляється у гетерозиготному стані. Гомозиготний фенотип його невідомий.
5. Зворотне домінування – домінування, що змінюється під дією зовнішніх умов.
Співвідношення при розщеплені гібридів ІІ покоління, встановлені Менделем, спостерігалося при повному домінуванні, тобто коли один алель повністю подавляє дію другого. Надалі були знайдені й інші види домінування: неповне, кодомінантність і зверхдомінування.
Наприклад, при схрещуванні безвухих вівцематок і баранів з нормально розвинутими вухами одержували коротковусе потомство. Таке явище називають неповним домінуванням.
При кодомінуванні у гібридів першого покоління проявляються ознаки обох предків, які виражені однаково і незалежно одна від одної. Так, при схрещуванні червоних шортгорнських корів з білими шортгорнськими бугаями одержують телят чалої масті (помісь білого і червоного волося), або симентальських корів палевої масті з чорно-рябими гоштинськими бугаями, перше покоління буде буро-сірим.
Зверхдомінування характеризується тим, що у гібридів першого покоління спостерігається значно сильніший розвиток ознаки, ніж у вихідних батьківських форм. Таке явище називається гетерозисом.
Множинний алелізм. До цих пір ми розглядали гени, які існують в двох алельних станах, - домінантному і рецесивному, проте багато генів мають більше двох алелей. Серією множинних алелей називають три і більше станів одного локусу, які обумовлюють різні фенотипи.
Лекція 6 Взаємодія генів
1. Взаємодія генів
2. Методи генетичного аналізу. Аналізуюче та реціпрокне схрещування
3. Фактори, які впливають на розщеплення ознак у гібридів
1. Взаємодія генів
Внаслідок недостатнього вивчення явищ спадковості в ранньому періоді розвитку генетики викорінилася уява про те, що формування кожної ознаки зумовлене лише одним спадковим фактором (геном), який діє тільки на одну ознаку незалежно від інших генів. Подальші дослідження показали, що взаємодія генів і ознак досить складна. Доведено, що одна ознака в своєму розвитку може залежати від багатьох генів і що один ген може діяти на багато ознак. На перший погляд здавалося, що ці явища суперечать законам Менделя, але завдяки цим законам їх можна пояснити. Відкриття і вивчення різних типів взаємодії генів стало другим етапом розвитку генетики.
У сучасній генетиці розрізняють більше 20 різних типів взаємодії генів, основними з яких є: комплементарна, епістатична, полімерна, модифікуючи, плейотропна, летальна.
Комплементарним типом називається така взаємодія двох і більше неалельних генів (генів розміщених в різних локусах хромосом), при якій вони разом у домінантному стані визначають одну ознаку, але окремо її викликати не можуть. Крім того у ІІ поколінні можуть змінюватися характерні для менделівського успадкування співвідношення розщеплення за генотипами.
Наприклад, при схрещуванні курей із стрічкоподібним гребенем (rrPP) з півнями, що мають трояндоподібний гребінь (rrPP), у першому поколінні (F1) все потомство має нову форму гребня – горіхоподібну (RrPp). При схрещуванні гібридів F1 між собою серед покоління F2 відбувається таке розщеплення: 9 горіхоподібних, 3 трояндоподібних, 3 стручкоподібних і один простий гребінь.
Епістатичний тип взаємодії генів – це тип при якому алель одного із генів блокує дію алелів інших генів.
Гени, що пригнічують дію інших генів, називаються супресорами або інгібіторами, епістатичними.
Гени, дія яких пригнічена, називаються гіпостатичними.
Наприклад, у курей породи леггорн є ген забарвлення (С). Проте в нормі він пригнічений домінантним супресором І, тому пір’яний покрив у такої птиці білий. При схрещуванні білих леггорнів (ІІСС) з білими курми інших порід (іісс) І покоління ІіСс має біле пір’я. У ІІ поколінні три із 16 особин виявляються кольоровими, так як ген забарвлення у них звільнився від дії супресора (замість його присутній рецесивний алель і).
Полімерний тип взаємодії – це взаємодія генів, при якій формування ознаки відбувається під одночасним контролем ряду еквівалентних генів, що мають назву полімерні.
За типом полімерії успадковуються кількісні ознаки (інтенсивність росту, жива маса, несучість курей, молочність, жирномолочність, вовновість овець тощо).
У разі, коли дія кількох полімерних генів сумується, виникає кумулятивний ефект. Взаємодію такого типу називають кумулятивною полімерією (адитивним ефектом). Прикладом полімерної дії є формування забарвлення зерна у пшениці. Так у ІІ поколінні від кількості домінантних генів інтенсивність забарвлення зерна міняється від червоного (ААВВ), з різним ступенем послаблення відтінків у варіантах АаВВ, ааВВ, аааВ до білого у рецесивній формі – аавв.
При модифікуючій дії гени не проявляють власної дії на ознаку, але зумовлюють вплив на ефект дії других генів, таких генів називають генами-модифікаторами. Вважають, що гени-модіфікатори відіграють велику роль у формуванні у особини резистентності до інфекцій та інших хвороб, а також стрес-факторам. Прикладом дії генів-модифікаторів, які послаблюють або посилюють дію основного гена, може бути варіація білої строкатості у тварин айрширської або гоштинської порід. Обидві породи гомозиготні за геном, який визначає виникнення білих плям, але гени-модифікатори визивають варіацію строкатості від майже повної пігментації всього тіла до майже повної її відсутності.
Плейотропна (множинна) дія генів – полягає у дії одного гена на кілька ознак. Розуміння плейотропії важливе у зв’язку з тим, що це явище включає ті ознаки, які мають важливе еволюційне значення, наприклад, плодючість, тривалість життя, здатність виживати в несприятливих умовах. Нині плейотропізм настільки широко обговорюється, що викликає сумнів існування (у вищих організмів) генів, які не мають плейотропної дії.
Летальна дія генів. Серед найбільш яскравих прикладів доведення взаємодії генів у розвитку організму є приклади своєрідної «відмови» спільного функціонування генома – загибель організму, так названа летальність. З позиції розуміння фізико-хімічних механізмів генетичних процесів дію летальних генів необхідно розцінювати як результат блокування утворення головного метаболіту. Наприклад, забезпечення клітини енергією у вигляді АТФ.
Гени, які викликають загибель гомозигот, називають рецесивними леталями, а які діють на гетерозиготи – домінантними леталями.
Прикладом летальності є гомозиготний стан гена сірого забарвлення у каракульських овець. При схрещуванні сірих овець з сірими барана ми виявилось, що вони завжди гетерозиготні, так як у їх потомстві завжди було 25% чорних ягнят. У той же час 25% ягнят сірого забарвлення гинули від хронічної тимпанії при переході на грубі корми. Коли схрещували сірих баранів з чорними матками або навпаки, то у потомстві виявлялось 50% сірих і 50% чорних ягнят, причому сірі ягнята не хворіли. Виявилось, що у гомозиготному стані ген, що контролює розвиток сірого забарвлення, мав рецесивну летальну дію, яка основана на порушенні функцій парасимпатичної нервової системи.
При штучному осіменінні , коли від одного плідника, гетерозиготного по летальному гену, можна одержати від декількох десятків до сотень тисяч телят, можна уявити собі небезпечність такого явища.
До нинішнього часу у ВРХ визначено 46 летальних генів, у коней 10, у свиней 18, у овець 15, у курей 45, у індичок 6, у качок 3. Різниця у кількості леталій, знайдених у тварин різних видів, залежить не стільки від біологічних особливостей виду, скільки від ступеня його вивченості, кількості і матеріальної цінності одержаного потомства.
1. Методи генетичного аналізу.
Аналізуюче та реціпрокне схрещування
Залежно від мети, якої бажають досягти, завдання генетичного аналізу можуть бути різними. У селекційній практиці, як правило, користуються такими основними типами схрещувань.
Аналізуюче схрещування. У практиці часто виникає необхідність визначити генотип родоначальника лінії, родини тощо на наявність у ньому патологічних або, навпаки, дуже цінних генів. Для вирішення такого типу завдань можна використати один із важливих методів гібридологічного аналізу – аналізуюче схрещування.
Цей метод, названий ще тест-крос, дозволяє визначити генотип особини з домінантною ознакою (АА і Аа). Особину з домінантною ознакою схрещують з гомозиготною за рецесивним алелем (аа).
У цьому випадку рецесивна форма утворює тільки один сорт гамет з алелем а, що дає можливість проявитися будь якому з двох алелів генотипу, який тестують.
Реципрокним називають таку пару схрещувань, в якій організми з домінантними і рецесивними ознаками використовуються і як материнські, і як батьківські. Проте схрещування сприяє більш глибокому аналізу механізму успадкування і прояву явища гетерозису при формуванні кількісної ознаки. Дуже часто використання цього методу в селекції називають реципрокною селекцією.
У реципрокних схрещуваннях розрізняють пряме, наприклад, якщо АА х аа і зворотне, якщо аа х АА.
Широке використання реципрокного схрещування пов’язане з перевагами його над методами гібридизації інбредних ліній і топ кросів, оскільки дає можливість повністю використати гетерозис на основі виявлення найкращих поєднань батьківських пар і не потребує тих величезних витрат, які необхідні для ведення інбредної селекції.
Зворотне, або бекрос- схрещування – це використання одного із батьків в послідовній серії схрещувань з його безпосереднім потомством.
Мета зворотного схрещування – передати кілька ознак одного із батьків потомству, не викликаючи змін генотипу останнього (за винятком ознаки, яку треба ввести).
3 Фактори, які впливають на розщеплення ознак у гібридів
Дискретний характер генів призводить до чітких значень співвід-ношеннь розщеплення ознак в потомстві гібридів. Однак на практиці фактично ця закономірність не завжди спостерігається. Тому вивчаючи це питання, слід зупинитися на умовах, при яких відбувається незалежне комбінування ознак.
Умови, при яких зберігається закон розщеплення, такі:
перебування генів, які вивчають, у негомологічних хромосомах, кількість їх пар при цьому перевищує гаплоїдну кількість хромосом даного виду;
рівноймовірне утворення гамет усіх типів на основі випадкового розходження хромосом у мейозі;
рівноймовірне дозрівання гамет усіх типів;
рівноймовірна зустріч гамет при заплідненні:
рівноймовірне виживання зигот і дорослих організмів;
відносна стабільність розвитку ознак, які вивчають.
Розглянемо основні причини, які впливають на відхилення від класичних методів розщеплення у F2.
Недостатній об’єм вибірки. Для підтвердження менделівських законів необхідна велика статистична вибірка матеріалу, який вивчають, оскільки в процесі формування ознаки на організм діють різні фактори, які визивають зміни в розщепленні.
Взаємодія неалельних генів. Важливим фактором, що впливає на очікуване розщеплення ознак у гібридів, є феномен різних типів взаємодії генів, особливо множинного плейотропного впливу гена на багато ознак, насамперед на життєздатність і плодючість, летальні гени, зчеплене зі статтю успадкування, а також аутосомне зчеплення, мутаційний процес.
Штучний добір. Як було зазначено, важливою умовою виконання законів Менделя є рівноймовірність зустрічі усіх типів гамет різних гібридних форм. В умовах селекційно-племінної роботи це порушується свідомо, бо проводиться добір тих форм, які відповідають вимогам бажаного типу за екстер’єром, конституцією, продуктивністю.
Таким чином, добір, який широко застосовують у зоотехнічній діяльності – антагоніст вільного схрещування що збільшує швидкість мікроеволюційних процесів у стадах, які селекціонують.
Міграція генів. У племінних і товарних стадах значне місце належить міграції генів шляхом завезення більш цінних плідників цієї ж породи або її сперми з інших зон чи країн.
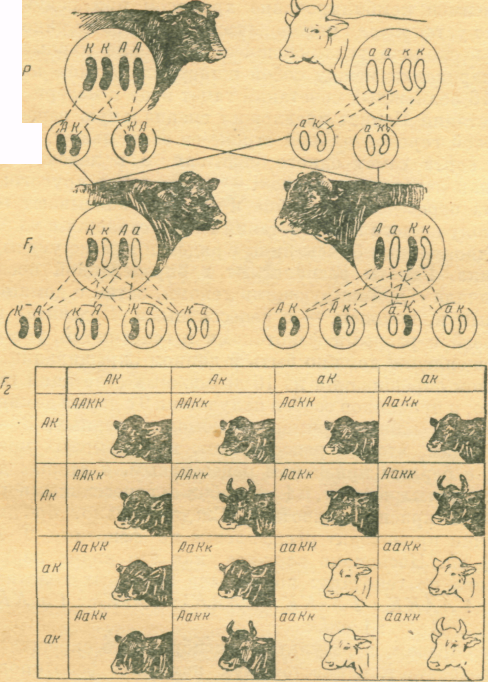
Рис. 1. Схема схрещування чорного комолого бугая
з червоними рогатими коровами
Рис. 2.Схема незалежного успадкування факторів при дигібридному схрещуванні
Закономірності успадкування ознак при статевому розмножені