Регуляция клеточного цикла. Жизненный цикл клетки
СОДЕРЖАНИЕ
Введение……………………………………………………………...…3
1. Жизненный цикл клетки……………………………………….5
1.1. Этапы клеточного цикла и его особенности……………………5
1.2. Митоз. Стадии митоза. Амитоз………………………………….10
2. Регуляция клеточного цикла…………………………..…..17
Заключение……………………..………………………………….…22
Список использованных источников.…..………………..23
ВВЕДЕНИЕ
Клеточный цикл – это период существования клетки от момента её образования путем деления материнской клетки до собственного деления или смерти. Длительность клеточного цикла у разных клеток варьирует. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12-36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яиц иглокожих, земноводных и других животных [3, c. 209].
В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10-24 ч. Нарушение нормальной регуляции клеточного цикла является причиной появления большинства твердых опухолей. В клеточном цикле, как уже говорилось, прохождение контрольных пунктов его возможно только в случае нормального завершения предыдущих этапов и отсутствия поломок. Для опухолевых клеток характерны изменения компонентов сверочных точек клеточного цикла [27, c. 46].
Клеточный цикл включает периоды интерфазы и деления клетки. Различают несколько способов деления клеток, основными из которых являются митоз и мейоз. Помимо этих наиболее распространенных типов деления, встречаются амитоз (прямое деление клеток) и различные виды эндорепродукции, приводящие к увеличению количества ДНК в клетках. Исследования клеточного цикла и его этапов весьма актуальны и характеризуются теоретической и практической значимостью. Эти исследования способствуют пониманию особенностей, механизмов реализации и регуляции событий клеточного цикла, а также открывают перспективы целенаправленного управления клеточным циклом [2, c. 134].
Целью данной работы является рассмотрение особенностей жизненного цикла клетки и способов его регуляции.
1. ЖИЗНЕННЫЙ ЦИКЛ КЛЕТКИ
- Этапы клеточного цикла и его особенности
Изучение клеточного цикла базировалось на представ�лениях цитокинетики, которые к 60-м гг. XX в., несмотря на их новизну, уже вполне сформировались. Клетки многоклеточного организма чрезвычайно разнообразны по выполняемым функциям. В соответствии со специализацией клетки имеют разную продолжительность жизни. Так нервные клетки после завершения эмбриогенеза перестают делиться и функционируют на протяжении всей жизни организма [16, c. 88].
Клетки же других тканей (костного мозга, эпидермиса, эпителия тонкого кишечника) в процессе выполнения своей функции быстро погибают и замещаются новыми в результате клеточного деления. Деление клеток лежит в основе развития, роста и размножения организмов. Деление клеток также обеспечивает самообновление тканей на протяжении жизни организма и восстановление их целостности после повреждения [26, c. 167].
Существует два способа деления соматических клеток: амитоз и митоз. Преимущественно распространено непрямое деление клеток (митоз). Размножение с помощью митоза называют бесполым размножением, вегетативным размножением или клонированием. Жизненный цикл клетки (клеточный цикл) – это период существования клетки от деления до следующего деления или смерти. Продолжительность клеточного цикла в размножающихся клетках составляет 10-50 ч и зависит от типа клеток, их возраста, гормонального баланса организма, температуры и других факторов. Детали клеточного цикла варьируют среди разных организмов [18, c. 54].
У одноклеточных организмов жизненный цикл совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках клеточный цикл совпадает с митотическим циклом. Под митотический циклом понимают совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и период деления (табл. 1). В соответствие с приведенным выше определением митотический цикл подразделяют на интерфазу и митоз (греч. «митоз» – нить) [1, c. 25].
Таблица 1
Митотический цикл и митоз
Фазы
|
Процесс, происходящий в клетке
|
|
Интерфаза
|
Пресинтетический период (G1)
|
Синтез белка. На деспирализованных молекулах ДНК синтезируется РНК
|
|
|
Синтетический
период (S)
|
Синтез ДНК - самоудвоение молекулы ДНК. Построение второй хроматиды, в которую переходит вновь образовавшаяся молекула ДНК: получаются двухроматидные хромосомы
|
|
|
Постсинтетический период (G2)
|
Синтез белка, накопление энергии, подготовка к делению
|
|
Фазы митоза
|
Профаза
|
Двухроматидные хромосомы спирализуются, ядрышки растворяются, центриоли расходятся, ядерная оболочка растворяется, образуются нити веретена деления
|
|
|
Метафаза
|
Нити веретена деления присоединяются к центромерам хромосом, двухроматидные хромосомы сосредоточиваются на экваторе клетки
|
|
|
Анафаза
|
Центромеры делятся, однохроматидные хромосомы растягиваются нитями веретена деления к полюсам клетки
|
|
|
Телофаза
|
Однохроматидные хромосомы деспирализуются, сформировывается ядрышко, восстанавливается ядерная оболочка, на экваторе начинает закладываться перегородка между клетками, растворяются нити веретена деления
|
Интерфаза – период между двумя делениями клетки – подразделяется на фазы G1, S и G2 (рис. 1). По продолжительности интерфаза составляет большую часть митотического цикла клетки. Наиболее вариабельны по времени G1 и G2-периоды [15, c. 174].
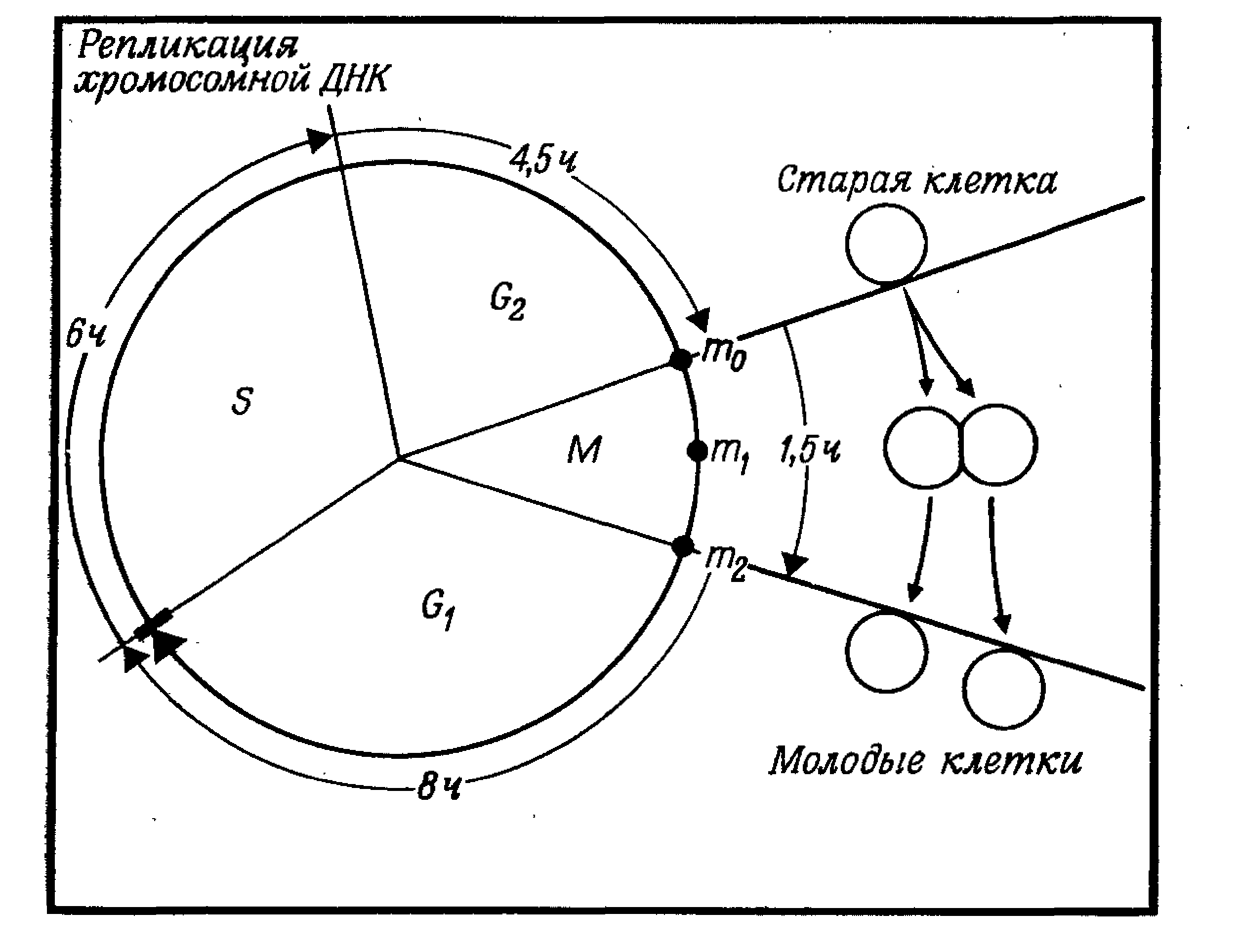
Рис. 1. Схема клеточного цикла и его фаз
G1 (от англ. grow – расти, увеличиваться). Продолжительность фазы составляет 4-8 ч. Это фаза начинается сразу после образования клетки. В этой фазе в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в синтезе ДНК. Если клетка в дальнейшем не делится, то переходит в фазу G0 - период покоя. С учетом периода покоя клеточный цикл может длиться недели или даже месяцы (клетки печени) [6, c. 104].
S (от англ. synthesis - синтез). Длительность фазы составляет 6-9 ч. Масса клетки продолжает увеличиваться, и происходит удвоение хромосомной ДНК. Две спирали старой молекулы ДНК расходятся, и каждая становится матрицей для синтеза новых цепей ДНК. В результате каждая из двух дочерних молекул обязательно включает одну старую спираль и одну новую. Тем не менее, хромосомы остаются одинарными по структуре, хотя и удвоенными по массе, так как две копии каждой хромосомы (хроматиды) все еще соединены друг с другом по всей длине. Это «сестринские хроматиды», то есть идентичные копии, образовавшиеся в процессе репликации ДНК за время С-фазы. Сестринские хроматиды соединяются друг с другом в особых участках - центромерах [24, c. 85-86].
В ядрах животных клеток имеются особые крошечные структуры, центриоли, которые начинают двигаться в противоположные стороны, образуя полюса деления. Между центриолями образуется структура, называемая веретеном деления; она состоит из многих нитей (скорее даже микротрубочек, образованных белком тубулином), которые распределяют хромосомы по двум ядрам. Некоторые из этих нитей тянутся от центриоли до центромеры хромосом, растаскивая их в разные стороны [11, c. 235].
Пока к центромерам прикрепляются нити веретена, хромосомы свободно движутся по клетке, но через некоторое время они останавливаются на полпути между полюсами (в экваториальной плоскости). Теперь клетка вступает в стадию метафазы, второй фазы митоза. Все сестринские хроматиды одновременно, словно по команде, начинают расходится к полюсам благодаря сокращению одних нитей веретена, в то же время другие нити веретена удлиняются и отодвигают полюса друг от друга. Эти события знаменуют собой анафазу, после которой клетка начинает сужаться в центре. После завершения фазы S митотического цикла клетка не сразу начинает делиться [23, c. 65-66].
В фазе G2 в клетке завершается процесс подготовки к митозу: накапливается АТФ, синтезируются белки ахроматинового веретена, удваиваются центриоли. Масса клетки продолжает увеличиваться до тех пор, пока она приблизительно вдвое не превысит начальную, а затем наступает митоз. Рассмотрим схему клеточного цикла, предложенную в 1953 г. А. Ховардом и С. Пелком. Сразу после митоза каждая дочерняя клетка является диплоидной и имеет нормальный набор хромосом (рис. 1). Эта клетка должна пройти фазу G1, чтобы быть в состоянии удвоить свою ДНК в течение фазы S жизненного цикла. После удвоения ДНК клетка переходит в фазу G2 и нахо�дится в тетраплоидном состоянии [5, c. 199].
В процессе митоза клетка снова делится, причем ее тетраплоидный набор хромосом распределяется поровну между двумя дочерни�ми клетками; в результате этого каждая из новых клеток: начинает цикл, имея диплоидный набор хромосом. Дочерняя клетка со�храняет в течение всей фазы G1 «старые», т. е. материнские, цепи ДНК. Дупликация этих цепей происходит в фазе S. В течение фазы G2 в клетке находятся и «старые» цепи ДНК, и «новые» (синтези�рованные в фазе S) [10, c. 203].
Во время фазы М (митоз, включающий стадии профазы, метафазы, анафазы и телофазы) материнская клетка передает каждой дочерней одну из старых цепей (имевшихся еще в фазе G1) и одну из новых цепей (построенных в фазе S). Фаза G1 у клеток разных типов имеет различную продолжительность (у некоторых клеток около 30 ч, а у некоторых может практически отсутствовать). Фазы S, G2 и М более стабильны по продолжительности. Длительность фа�зы М зависит от температуры. Для большинства клеток млекопитаю�щих при росте in vitro продолжительность цикла составляет пример�но 20 ч, при этом фазы G1, S, G2 и М длятся соответственно 8 и 6, 5 и 1 ч. [4, c. 34].
Установлено, что в нормальных тканях организма не�которые клетки, будучи дифференцированными, постоян�но делятся. Таковы, например, клетки кишечного эпите�лия, костного мозга, эпидермиса, эпителия семенных ка�нальцев («лабильные» клетки по терминологии Г. Биццоцеро). Другие же клетки по окончании дифференцировки больше не пролиферируют («вечные» клетки по Г. Биццоцеро). Лучшим при�мером клетки, не способной к делению, может служить нейрон, дифференцировка которого заканчивается сразу после рождения и который сохраняется на протяжении всей жизни индивидуума (условие, необходимое для со�хранения следов памяти) [28, c. 234].
В клетке ДНК имеется не только в хромосомах, но и в митохондриях. Для полного представления обо всем синтезе ДНК не�обходимо также учесть и процесс репликации митохондриальной ДНК на протяжении клеточного цикла и срав�нить его с синтезом, происходящим в ядре. В то время как синтез хромосом�ной ДНК в ядре происходит в фазе S митохондриальная ДНК синтезируется на двух различ�ных этапах клеточного цикла: в первый раз в той же фа�зе S одновременно с ядерной ДНК, а второй раз - в фа�зе G2, когда синтез ядерной ДНК уже закончен и идет только репаративный синтез. Когда это выяснилось, прежде всего, возникла мысль, что в клетке могут существовать два класса генетически различных митохондрий, каждый со своей особой ДНК и своим временем репликации [19, c. 264].
При изучении физико-хими�ческих свойств митохондриальной ДНК, выделенной в фазах S и G2, было установлено, что действительно существуют различия между этими двумя препаратами. Таким образом, гипотезу о том, что в эукариотической клетке существуют разные типы митохондриаль�ной ДНК и, следовательно, генетически различные митохондрии, можно считать правдоподобной [25, c. 209].
1.2. Митоз. Стадии митоза. Амитоз
Митоз условно разделяют на четыре фазы: профазу, метафазу, анафазу и телофазу (рис. 2).
Рис. 2. Митоз клетки: А – интерфаза; Б, В, Г, Д – профаза; Е, Ж –
метафаза; 3, И – анафаза; К, Л, М – телофаза
На стадии профазы в животных клетках две центриоли начинают расходиться к противоположным полюсам ядра. В клетках растений при этом образуются два полярных центра организации микротрубочек, лишенных центриолей. Ядерная мембрана разрушается; одновременно специальные белки объединяются, формируя микротрубочки в виде нитей. Центры организации микротрубочек, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). [3, c. 94].
Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя веретено деления. В это время хромосомы спирализуются и вследствие этого утолщаются. Они хорошо видны в световом микроскопе, особенно после окрашивания. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли [20, c. 44].
В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. В каждой хромосоме имеется область первичной перетяжки – центромера, к которой во время митоза присоединяются нить веретена деления и плечи. На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры [24, c. 84].
В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом (4n4с) [26, c. 203].
В телофазе хромосомы раскручиваются, деспирализуются. Из мембранных структур цитоплазмы образуется ядерная оболочка. В это время восстанавливается ядрышко. На этом завершается деление ядра (кариокинез), затем происходит деление тела клетки (или цитокинез) [2, c. 74].
При делении животных клеток на их поверхности в плоскости экватора появляется борозда, постепенно углубляющаяся и разделяющая клетку на две половины – дочерние клетки, в каждой их которых имеется по ядру. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму: она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки (т.е. растет изнутри кнаружи) [18, c. 70].
Клеточная пластинка формируется из материала, поставляемого эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточную мембрану и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки. Особенности протекания митоза у животных и растений приведены в таблице 2 [21, c. 54-55].
Таблица 2
Особенности митоза у растений и у животных
|
Растительная клетка
|
Животная клетка
|
|
Центриолей нет
Звезды не образуются
Образуется клеточная пластинка
При цитокенезе борозда не образуется
Митозы преимущественно
происходят в меристемах
|
Центриоли имеются
Звезды образуются
Клеточная пластинка не образуется
При цитокинезе образуется борозда
Митозы происходят в различных тканях организма
|
Цитокинез – разделение цитоплазмы на две обособляющиеся части. Процесс начинается в поздней анафазе или в телофазе. Плазмалемма втягивается между двумя дочерними ядрами в плоскости, перпендикулярной длинной оси веретена. Борозда деления углубляется, и между дочерними клетками остаётся мостик – остаточное тельце. Дальнейшее разрушение этой структуры приводит к полному разделению дочерних клеток. Так из одной клетки формируются две дочерние, в которых наследственная информация точно копирует информацию, содержавшуюся в материнской клетке. Начиная с первого митотического деления оплодотворенной яйцеклетки (зиготы) все дочерние клетки, образовавшиеся в результате митоза, содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз – это способ деления клеток, заключающийся в точном распределении генетического материала между дочерними клетками. В результате митоза обе дочерние клетки получают диплоидный набор хромосом. [22, c. 45-46].
Весь процесс митоза занимает в большинстве случаев от 1 до 2 ч. Частота митоза в разных тканях и у разных видов различна. Например, в красном костном мозге человека, где каждую секунду образуется 10 млн. эритроцитов, в каждую секунду должно происходить 10 млн. митозов. В нервной ткани митозы крайне редки: так, в центральной нервной системе клетки в основном перестают делиться уже в первые месяцы после рождения; а в красном костном мозге, в эпителиальной выстилке пищеварительного тракта и в эпителии почечных канальцев они делятся до конца жизни [15, c. 204].
Амитоз (от греч. а – отриц. частица и митоз) – прямое деление интерфазного ядра путем перешнуровывания без преобразования хромосом. При амитозе не происходит равномерное расхождение хроматид к полюсам. Этот способ деления не обеспечивает образование генетически равноценных ядер и клеток. По сравнению с митозом амитоз более кратковременный и экономичный процесс. Амитотическое деление может осуществляться несколькими способами. Наиболее распространенный тип амитоза – это перешнуровывание ядра на две части. Этот процесс начинается с разделения ядрышка (рис. 3). [23, c. 64].
Рис. 3. Амитоз. Эпителиальные клетки мочевого пузыря мыши:
I-IV – различные стадии непрямого деления ядра, 1 – ядро, 2 – ядрышко
Перетяжка углубляется, и ядро разделяется надвое. После этого начинается разделение цитоплазмы, однако это происходит не всегда. Если амитоз ограничивается только делением ядра, то это приводит к образованию дву- и многоядерных клеток. При амитозе может также происходить почкование и фрагментация ядер. Клетка, претерпевшая амитоз, в последующем не способна вступить в нормальный митотический цикл [17, c. 44].
Амитоз встречается в клетках различных тканей растений и животных. У растений амитотическое деление довольно часто встречается в эндосперме, в специализирующихся клетках корешков и в клетках запасающих тканей. Амитоз также наблюдается в высокоспециализированных клетках с ослабленной жизнеспособностью или дегенерирующих, при различных патологических процессах, таких как злокачественный рост, воспаление и т. п. [2, c. 164].
2. РЕГУЛЯЦИЯ КЛЕТОЧНОГО ЦИКЛА
Пролиферация клеток, происходящая путём митоза, жёстко регулируется множеством молекулярных сигналов. Регуляторы клеточного цикла и митоза подразделяют на внутриклеточные и межклеточные. Внутриклеточные молекулярные сигналы многочисленны, среди них в первую очередь следует назвать собственно регуляторы клеточного цикла (циклины, циклин-зависимые протеинкиназы, их активаторы и ингибиторы) и онкосупрессоры [9, c. 233].
Скоординированная деятельность этих многочисленных регуляторов клеточного цикла обеспечивает как переход клеток от фазы к фазе клеточного цикла, так и точное выполнение событий каждой фазы. Нарушения же такой скоординированной деятельности приводят как к искажениям прохождения клетки по фазам клеточного цикла (патология митоза), так и к потере контроля за пролиферативными потенциями клетки, что может привести к бесконтрольной её пролиферации – появлению неконтролируемого клона, т.е. к опухолевому росту [24, c. 209].
Клетки таких клонов называют трансформированными, или малигнизированными. Главная причина появления пролиферативно неконтролируемых клеток – мутации генов, кодирующих структуру собственных регуляторов клеточного цикла. Циклины – особые белки, которые играют важную роль в регуляции клеточного цикла. Идентифицировано шесть классов циклинов: A, B, C, D, E, F. Название этих белков отражает цикличность процессов сборки и разборки макромолекулярного комплекса в процессе каждого клеточного цикла. Циклины различаются экспрессией на определенных стадиях клеточного цикла и соответственно на различных стадиях регулируют деление клетки. К середине M-фазы концентрация циклинов в клетке резко уменьшается с последующим нарастанием синтеза на протяжении всего цикла. [13, c. 44]
Рис. 4. Концентрация циклинов на разных стадиях клеточного цикла
Cdk (cyclin dependent protein kinase) – циклин-зависимые протеинкиназы. Комплекс циклинов с циклин-зависимыми протеинкиназами играет центральную роль в клеточном цикле. Последовательная активация циклин-зависимых протеинкиназ и последующее фосфорилирование ими критических субстратов контролируют клеточный цикл, переключая его с одной фазы на другую (с G1 на S или G2 на M). Белки, связывающиеся с этим комплексом и ингибирующие его каталитическую активность, блокируют клеточный цикл в ответ на антипролиферативные сигналы (TGFb, факторы дифференцировки миогенных, миелоидных и нервных клеток). Интеграция вируса гепатита В в ген циклина А обнаружена при гепатоклеточной карциноме [8, c. 244].
Существуют специальные вещества, подавляющие развитие опухолей – онкосупрессоры, которые в нормальной клетке постоянно контролируют множество процессов, в том числе и пролиферацию клеток (точнее, онкосупрессоры блокируют клеточный цикл). Известно множество онкосупрессоров (например, Rb, p27, p53, PTEN), среди которых p53 известен как главный супрессор развития опухоли. Мутации генов онкосупрессоров неизбежно приводят к появлению бесконтрольно пролиферирующего клеточного клона [19, c. 145].
Белок p27 связывается с циклинами и Cdk и блокирует вхождение клетки в S-фазу цикла. Определение р27 используют в диагностике рака молочной железы. Снижение уровня р27 является плохим прогностическим признаком. Белок р53 – один из важнейших регуляторов клеточного цикла, специфически связывается с ДНК и подавляет рост клеток в фазе G1. р53 регистрирует различные сигналы при внешних воздействиях на клетку (вирусная инфекция, гипоксия) и состояние её генома (активация онкогенов, повреждения ДНК). [29, c. 254]
При неблагоприятной информации о состоянии клетки р53 блокирует клеточный цикл до тех пор, пока нарушения не будут устранены. В повреждённых клетках содержание р53 возрастает. Это даёт клеткам шансы восстановить ДНК путём блокирования клеточного цикла. При серьёзных повреждениях р53 инициирует самоубийство клетки – апоптоз. Опухоли (практически в 50%) сопровождаются мутациями гена р53. При этом, несмотря на возможные нарушения генома (включая изменения в количестве хромосом), клетки не входят в апоптоз и вступают в беспрерывный клеточный цикл. Репертуар мутаций гена р53 довольно широк. Они приводят к бесконтрольному размножению клеток при раке толстой кишки, печени, легкого, пищевода, молочной железы, глиальных опухолях мозга, опухолях лимфоидной системы. При синдроме Li Fraumeni врождённый дефект р53 является причиной высокой частоты поражения раком [11, c. 74-75].
На множество важных клеточных функций (экспрессия конкретных генов и специфическая дифференцировка, поддержание дифференцированного состояния и т.д.) влияют различные информационные сигналы, поступающие к клетке извне. Регуляция клеточного цикла и пролиферация (или блок пролиферации) клеток также регулируются внеклеточными сигналами в виде гормонов (например, гормон роста, эстрогены, фолликулостимулирующий гормон), цитокинов (например, интерлейкины и интерфероны), факторов роста (например, фактор роста эпидермиса - EGF). Некоторые из таких молекулярных сигналов расцениваются как стимулирующие митогенную активность факторы (митогены). Существенное влияние на пролиферативную активность клеток имеют и контакты с элементами межклеточного матрикса (например, с ламинином и фибронектином) [25, c. 56].
После взаимодействия с соответствующими рецепторами такие митогенные сигналы (в значительном числе случаев при помощи связанных с G-белками протеинкиназ) передаются на соответствующие участки генома, активируя транскрипцию контролирующих фазы клеточного цикла генов (например, циклин-зависимых протеинкиназ). Инфор�мационные РНК выходят из ядра в закончен�ном виде для выполнения своих функций в цитоплазме. После транскрипции, модификации и процессинга информационная РНК переходит из ядра в цитоплазму и нанизывает на себя определенное число рибосом, образуя полисому – своего рода конвейер, который на�чинает синтезировать белки [5, c. 104].
Соотношение «полисомы: мономеры цитоплазматических (SOS) рибосом: 40S- и 60S-субчастицы» в фазе G2 возрастает – почти все рибосомальные субчастицы объединяются в моносомы, а из моносом в то же время образуются полисомы. Высокая концентрация полисом в фазе G2 по сравне�нию с фазой G1 позволяет предполагать, что соответственно в исходной родительской клетке должен иметь место более интенсивный белковый синтез, чем в молодых дочерних клетках. В фазе G1 синтез полипептидов на поли�сомах идет гораздо активнее, чем в фазе G2. Таким образом, меньшему количеству полисом на одну клетку в фа�зе G1 соответствует более высокая скорость белкового синтеза, а большему количеству полисом в фазе G2 – меньшая скорость синтеза [29, c. 144].
В старых клетках происходит актив�ная мобилизация субчастиц рибосом для построения моносом с последующим быстрым образованием полисом. Поскольку в фазе G2 обнаруживаются гораздо более крупные полисомы, чем в фазе G1, возни�кает вопрос, одинакова ли скорость передвижения рибо�сом вдоль цепи мРНК в малых и больших полисомах. Считается, что эта скорость обратно пропорциональна длине мРНК. Биосинтез белков наиболее интенсивно протекает в двух фазах клеточного цикла – G1 и G2. Напротив, в фазе S, для которой ха�рактерен синтез ДНК, отмечается спад белкового синте�за. Таким образом, еще раз подтверждается правило «или ДНК, или белок» [21, c. 246].
ЗАКЛЮЧЕНИЕ
Клеточный цикл – это жизнь клетки от одного деления до другого. Про клетки, которые делиться больше не будут, обычно говорят, что
они вышли из клеточного цикла. Продолжительность клеточного цикла у разных организмов варьирует. Так, у бактерий она может составлять всего 20-30 мин, а у клеток эукариот цикл обычно длится не менее 10-12 ч, часто сутки и более. Исключение составляют быстро делящиеся клетки самых ранних зародышей, весь цикл у них может проходить за 15-20 мин. [13, c. 100].
Клетки взрослых многоклеточных организмов, как животных, так и растений, обладают разной способностью к делению. В одних тканях, например нервной, мышечной, клетки вообще не делятся. Другие ткани, напротив, постоянно обновляются. В этом случае существуют группы клеток, которые постоянно делятся, а их потомки перестают делиться, некоторое время функционируют и отмирают. Так происходит с клетками крови (делящиеся клетки находятся в костном мозге, а зрелые выходят в кровь), кожи, кишечника, в проводящей системе растений [20, c. 14].
Выход клеток из цикла может быть необратимым, но многие клетки, не размножающиеся в обычных условиях, могут приобрести эту способность вновь. Клетки печени, например, в норме почти не делятся, но после удаления части органа вступают в клеточный цикл и делятся один-два раза. Клетки коры некоторых многолетних растений способны, начав делиться, восстанавливать механические повреждения коры [1, c. 64].
Для правильного прохождения клеточного цикла у всех живых организмов существуют специальные системы регуляции. Систематическое изучение молекулярно-генетических основ клеточного цикла началось с середины 70-х гг. XX в. В результате наиболее ранних исследований была сформулирована гипотеза о том, что направленность процессов в клеточном цикле обусловлена сменой функционирования отдельных генов, ответственных за прохождение клеткой определенных его этапов. Дальнейшие исследования в этой области были направлены на выявление степени участия генома в регуляции отдельных процессов, развертывающихся на протяжении митотического цикла. [29, c. 159].
Согласно современным представлениям, в отличие от регуляции клеточного цикла в целом, которая в значительной степени базируется на дифференциальной активности генов, регуляция митоза осуществляется белковой «машиной» клеточного цикла на посттрансляционном уровне. Детальное исследование механизмов регуляции клеточного цикла и деления и путей воздействия на эти механизмы дает возможность целенаправленного управления судьбой клеток. Такой подход имеет огромное значение для медицины и биотехнологии. [7, c. 204]
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
- Абдукаева Н.С., Пашкина Н.С. И др. Биология клетки. - Спб.: Издательство СПбГПМА. 2002. - 108 с.
- Альбертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки. 2 том. - М.: Издательство Наука. 1997. - 540 c.
- Акуленко Л. В., Угаров И. В. Биология с основами медицинской генетики. - М.: Издательство ГЭОТАР-Медиа. - 2011. - 368 с.
- Асланян М. М., Солдатова О. П. Генетика и происхождение пола. - М.: Издательство Академия. - 2010. - 116 с.
- Дондуа А.К. Биология развития. Клеточные и молекулярные основы индивидуального развития. Т. 2. - Спб.: Издательство Санкт-Петербургского университета. - 2005. - 240 с.
- Ефремова В. В., Аистова Ю. Т. Генетика. - Ростов-на-Дону: Издательство Феникс. - 2010. - 256 с.
- Заяц Р. Г., Бутвиловский В. Э., Давыдов В. В. И др. Медицинская биология и общая генетика. - Минск: Вышэйшая школа. - 2011. - 496 с.
- Зенгбуш П. Молекулярная и клеточная биология. 3 т. - М.: Издательство Академия. - 2007. - 499 с.
- Клаг У., Каммингс М. Основы генетики. - М.: Издательство Техносфера. 2006. - 896 с.
- Кемп П., Армс К. Введение в биологию. - М.: Издательство АСТ. - 2007. - 675 с.
- Курчанов Н.А. Генетика человека с основами общей генетики. - М.: Издательство СпецЛит. - 2009. - 192 с.
- Лебедева Л.И., Федорова С.А., Трунова С.А. и др. Митоз. Регуляция и организация деления клеточного ядра // Генетика. – 2004. – т. 40. – № 12. – С. 1589-1608.
- Полевой В. В., Саламатова Т. С. Живое состояние клетки и биология старения. - Спб.: Издательство Санкт-Петербургского университета. 2004. - 136 с.
- Пехов А. П. Биология. - М.: Издательство ГЭОТАР-Медиа. - 2010. - 656 с.
- Пехов А. П. Биология и общая генетика. - М.: Издательство Российского Университета дружбы народов. - 2004. - 440 с.
- Пехов А. П. Биология. Медицинская биология, генетика и паразитология. - М.: Издательство Российского Университета дружбы народов. - 2007. - 668 с.
- Робертис Э., Новинский В., Саэс Ф. Биология клетки. - М.: Издательство Мир. - 1973. - 488 с.
- Репин В. С., Сухих Г. Т. Медицинская клеточная биология. - Спб.: Издательство Лань. - 2008. - 201 с.
- Рубан Э.Д. Генетика человека с основами медицинской генетики. - Ростов-на-Дону: Издательство Феникс. - 2012. - 320 с.
- Топорнина Н.А. Генетика человека. - М.: Издательство Академия. 2001.- 96 с.
- Уилсон Дж., Хант Т. Молекулярная биология клетки. - М.: Издательство Академия. - 2004. - 500 с.
- Фаллер Д. М., Шилдс Д. Молекулярная биология клетки. - М.: Издательство Бином. 2011. - 256 с.
- Харрисон Дж., Уайнер Дж., Таннер Дж. и др. Биология человека. - М.: Издательство Академия. - 2005. - 615 с.
- Хандорина Е.К., Терехова И. Д. Генетика человека с основами медицинской генетики. - М.: Издательство ГЭОТАР-Медиа. - 2011. - 232 с.
- Ченцов Ю.С. Введение в клеточную биологию. - М.: Издательство Академкнига. - 2005. - 496 с.
- Шапиро Н. Общая генетика. - М.: Издательство Академия. - 2004. - 305 с.
- Эдвард Алькамо И. Биология. - М.: Издательство Астрель. - 2002. - 216 с.
- Ярыгин В. Н., Васильева В. И., Волков И. Н. и др. Биология. Жизнь. Гены. Клетка. Онтогенез. Человек. 1 т. - М.: Издательство Высшая школа. - 1999. - 200 с.
- Lewis J., Raff M., Roberts K., Walter P. Molecular Biology of the Cell. - New York: Garland Science. - 2002. - 702 c.
Регуляция клеточного цикла. Жизненный цикл клетки