Бактеріофаги. Зборка частин бактеріофага. Розвиток бактеріофага в клітині
РЕФЕРАТ
з дисципліни “Біологія клітини”
на тему «Бактеріофаги. Зборка частин бактеріофага. Розвиток бактеріофага в клітині»
Зміст
- Вступ. ..................................................................................................3с.
- Віруси, природа вірусів, загальна характеристика......................... 4с.
- Бактеріофаги....................................................................................... 6с.
3.1. Відкриття бактеріофагів. ............................................................6с.
3.2. Загальна характеристика............................................................. 7с.
3.3. Будова. ..........................................................................................9с.
3.4. Хімічний склад............................................................................ 11с.
3.5. Проникненння в клітину. ...........................................................12с.
3.6. Розвиток в клітині. ......................................................................14с.
3.6.1. Вірулентні фаги. Літичний цикл. ..................................15с.
3.6.2. Латентні фаги. Лізогенія.................................................17с.
3.7. Зборка частин бактеріофага, вихід з клітини............................ 20с.
- Походження бактеріофагів. ...............................................................23с.
- Значення бактеріофагів. ....................................................................25с.
5.1. Роль у природі............................................................................. 25с.
5.2. Вплив на розвиток бактерій....................................................... 27с.
5.3. Використання бактеріофагів у діяльності людини.................. 27с.
- Висновки. ...........................................................................................29с.
- Використана література.................................................................... 30с.
- Вступ
Через 25 років після відкриття вірусів канадський вчений Фелікс д'Ерель за допомогою методу фільтрації виявив нову групу вірусів, які уражають бактерії – бактеріофаги. Це найбільша група вірусів. Не існує жодної бактерії, яка не могла б бути атакована фагом, в свою чергу кожен фаг є високоспецифічним та уражає лише певну бактерію – ці особливості зробили їх цікавими для медицини, адже вони здатні знищувати патогенні мікроорганізми, які набули стійкості до антибіотиків. Антибіотики мають ряд недоліків: знищують корисну мікрофлору організму, мають побічні ефекти на різні органи, можуть втрачати свою ефективність внаслідок пристосування до них бактерій. З фагами такого не відбувається, тож почалися активні спроби їх використання для лікування, та виявилось що вони мають свої недоліки: висока специфічність вимагає, щоб збудник хвороби був точно відомий, а на затрачаються додаткові ресурси, вони потребують особливих, складніших умов для розведення. Та їх не переставали вивчати й знаходити все нові способи їх застосування: як нешкідливий консервант для продуктів, що запобігає розмноженню в них хвороботворних бактерій; як єдиний спосіб боротьби зі стійкими до антибіотиків мікроорганізмами; так звані «наноін'єкції», коли бактеріофаги за допомогою спеціальних механізмів вводять в клітину свою генетичну інформацію – вчені прагнуть використовувати цей механізм для введення в бактеріальні клітини традиційного антибіотика. Вони є важливими об'єктами для генетики й молекулярної біології, стали інструментами генної інженерії та біотехнології. Це доводить, що бактеріофаги – доволі перспективний напрям, вже відкрито й знайдено практичне застосування багатьом їх властивостям, продовжуються активні дослідження фагів, вчені шукають нові способи їх використання і покращення вже відомих.
2. Віруси. Загальна характеристика вірусів
Назву «вірус» (лат. virus – отрута) застосовували спочатку для позначення різних маловивчених хвороботворних агентів. Пізніше вона закріпилася за групою збудників, відкритих в 1892 році Івановським, які виявились здатними проходити через бактеріальні фільтри. Їх стали називати «вірусами, що фільтруються» або просто «вірусами».[1]
Хоча вдалося досягти значних успіхів в отриманні високоочищених проб вірусів і було встановлено, що за хімічною природою це нуклеопротеїди (нуклеїнові кислоти, пов'язані з білками), самі частинки все ще залишалися невловимими і загадковими, тому що вони були занадто малі, щоб їх можна було побачити за допомогою світлового мікроскопа. Тому віруси і опинилися в числі перших біологічних структур, які були досліджені в електронному мікроскопі відразу ж після його винаходу в 30-і роки нашого століття.[2]
Віруси відрізняються від мікроорганізмів такими особливостями: 1) вони містять нуклеїнову кислоту тільки одного типу – або ДНК, або РНК; 2) для їх репродукції необхідна тільки нуклеїнова кислота; 3) вони не здатні розмножуватись поза живою клітиною.[1]
Віруси не розмножуються на штучних живильних середовищах - вони надто розбірливі в їжі. Звичайний м'ясний бульйон, який влаштовує більшість бактерій, для вірусів не годиться. Їм потрібні живі клітини, і не будь-які, а чітко визначені. Як і інші організми, віруси здатні до розмноження. Віруси мають спадковість. Спадкові ознаки вірусів можна визначити за спектром заражених і симптомами, що викликають захворювання, а також специфічністю імунних реакцій природних господарів або штучно імунізованих експериментальних тварин. Сума цих ознак дозволяє чітко визначити спадкові властивості будь-якого вірусу, і навіть більше - його різновидів, що мають чіткі генетичні маркери, наприклад: нейротропність деяких вірусів грипу і т.п. Мінливість є іншою стороною спадковості, і в цьому відношенні віруси подібні всім іншим організмам, що населяють нашу планету. При цьому у вірусів можна спостерігати як генетичну мінливість, пов'язану зі зміною спадкової речовини, так і фенотипічну мінливість, пов'язану з проявом того самого генотипу в різних умовах. [3]
На відміну від клітинних організмів, у вірусів відсутня власна система метаболізму , в тому числі і білок синтезуюча система. Віруси вносять у клітину тільки свою генетичну інформацію. з матриці вірусної ДНК і РНК синтезується інформаційна РНК, яка і слугує основою для утворення вірусних білків рибосомами інфікованої клітини. Молекула ДНК вірусів, або їх геном, може вбудовуватися в геном клітини хазяїна і існувати в такому вигляді , будучи як би додатковим геном, і навіть не проявляти себе невизначено довгий час.[4] Потім, все ще під впливом вірусу клітина утворює ферменти , які, зрештою, її ж і руйнують , вивільняючи при цьому безліч, іноді сотні вірусних частинок. Так розвиваються події при різних вірусних захворюваннях, наприклад, риніті, інфекційному гепатиті, поліомієліті, віспі,грипі,сказі. Симптоми цих хвороб обумовлені дією вірусних білків на організм. Деяким вірусам приписують роль збудників раку. Таким чином , паразитизм вірусів носить особливий характер це паразитизм на генетичному рівні.[5]
Поза клітиною вірус існує в вигляді вірусної частинки (віріона),яка складається з нуклеїнової кислоти та білкової оболонки - капсиду. Тому вірусну частинку називають також нуклеокапсидом. Віруси розпізнаються за наслідками свого розвитку в клітинах господаря. Вони руйнують цілі комплекси клітин і викликають ураження тканин, що веде до появи некротичних плям або зон лізису. Звичайні господарі вірусів - це рослини,тварини і мікроорганізми.[1]
Походження вірусів в процесі еволюції залишається незрозумілим. Їх залежність від живих клітин, що створюють те середовище, в якому вони ростуть і розмножуються, свідчить про те, що вони не могли з'явитися до того, як з'явилися клітинні організми. Це міркування поряд з відсутністю у них звичайних клітинних механізмів дозволяє припускати, що віруси являють собою, можливо, сильно дегенеровані клітини або фрагменти клітин, які якимось чином звільнитися від усього, без чого можна обійтися, за винятком свого спадкового апарату у вигляді нуклеїнової кислоти і захисного та інфекційного апарату у формі білка. Так чи інакше, але вірусні частинки це дуже своєрідно упаковані одиниці генетичної інформації, і більша частина даних про інформацію, що міститься в клітині, вперше була виявлена і зрозуміла на цих простих об'єктах. [6]
3. Бактеріофаги
3.1. Відкриття бактеріофагів
Вивчення бактеріофагів почалося в Індії з експериментів Ернеста Ханкіна (1896 рік), який вивчав цілющі властивості річки Ганг. Священну річку вважали продовженням волосся бога Шиви, і вона успішно виліковувала навіть від прокази. Тому Ханкін шукав у ній щось дуже маленьке, але мабуть дуже цілюще. Він виявив, що води річок Гангу і Джамна в Індії мають значну антибактеріальну активність, яка зберігалася після проходження через фарфоровий фільтр з порами дуже малого розміру, але усувалася при кип'ятінні. Найбільш докладно вивчав він дію невідомої субстанції на Vibrio Сholerae і припустив, що вона відповідальна за попередження поширення епідемій холери, викликаних вживанням води цих річок. Однак, в подальшому, він не пояснив цей феномен. 1898 року російський мікробіолог Н. Ф. Гамалія вперше спостерігав переривний лізис бактерій. 1915 року англійський учений Ф. Туорт описав це ж явище у гнійного стафілокока і відкрив перший вірус, що «пожирає бактерії» , коли він спостерігав лізис в культурах стафілококів з лімфи теляти. З його ім'ям пов'язано назву «феномен Туорта». 1917 року Фелікс д’Ерель робить аналогічне відкриття. Саме Фелікс д’Ерель, канадський співробітник Інституту Пастера в Парижі, дав їм назву «бактеріофаги» використовуючи суфікс «фаг» не в його прямому сенсі «є», а в сенсі розвитку за рахунок чогось, вони стали головною частиною всього його життя.
Мікробіолог, переважно самоучка, який провів 10 років у Гватемалі, Мексиці та Аргентині, там він мав справу з епідеміями дизентерії, жовтої лихоманки і грибків кавових дерев, а також дослідив декілька цікавих загадок бродіння. Все це було хорошою підготовкою до його роботи з фагами. В Інституті Пастера він проводив кропітке дослідження методу приготування вакцини за допомогою модельної системи «B. Typhimurium» на її природному господарі миші, оскільки був твердо переконаний в тому, що значущі дані про імунітет і патогенності можна отримати лише при використанні природних господарів. У вільний час він продовжував обстежувати дизентерійних хворих. З фекалій декількох з цих хворих він виділив анти-шигеловий «мікроорганізм», який був розмножений шляхом численних серій пасажів на бактерії-господарі і міг утворювати невеликі чисті кола на газоні цих бацил шигели.
Д’Ерель підсумовував початковий етап роботи з фагами у книзі «Бактеріофаг» (1922). Він провів класичні описи утворення бляшок, інфекційних центрів, літичних процесів, специфічної адсорбції на бактерії і розмноження. Все своє життя він присвятив, розробці можливості застосування фагів, отриманих шляхом належної селекції, як засоби для лікування захворювань. [3]
3.2. Загальна характеристика
Фаги, подібно всім іншим вірусам, є абсолютними клітинними паразитами. Вони несуть інформацію, необхідну для власної репродукції і водночас не мають власних енергосинтезуючих систем та білок-синтезуючих механізмів. Залежно від типу взаємодії з мікробною клітиною розрізняють вірулентні та помірні, помірно-вірулентні та вірулентно-помірні бактеріофаги. Помірні фаги можуть мати неінфекційну форму - форму профага. Більшість фагів мають ДНК-геном, але є і РНК-геномні фаги. За формою віріона їх поділяють на 4 групи:
1)фаги, що мають ізометричну голівку і відросток;
2) ізометричні фаги;
3) ниткоподібні фаги;
4) плеоморфні фаги.
Згідно останньої класифікації віруси бактерій виділені в 1 порядок, 13 родин та 31 рід. При їх класифікації використовують близько 40 критеріїв. Генетична інформація фагів закодована в його нуклеїновій кислоті, білок захищає геном від дії зовнішніх факторів та виконує структурну функцію, а також забезпечує проникнення нуклеїнової кислоти в чутливу клітину, відповідає за антигенні та імуногенні властивості вірусу.
Як і будь-які інші віруси, фаги є носіями усіх основних вірусних ознак, що
чітко відрізняє їх від інших об’єктів живої природи:
- Наявність одного типу нуклеїнової кислоти (ДНК чи РНК).
- Відсутність власного білок-синтезуючого апарата.
- Неспроможність до росту та розмноження на поживних середовищах.
- Диз’юнктивна (розрізнена) збірка віріонів.
- Здатність до кристалізації.
- Неінфекційність поза клітиною.
Більшість фагів складаються із двох структурних компонентів – головки і відростку, кожний з яких виконує певну функцію в залежності з їх хімічною природою. Головка фага представляє собою молекулу нуклеїнової кислоти –ДНК або РНК, оточена білковою оболонкою, відросток–стрижень, який оточений білковим чохлом. Нерідко фаговий відросток виражений дуже слабко, а іноді і зовсім відсутній. Крім того зустрічаються фаги, які мають паличковидну форму - іноді довгих і зігнутих. Генетичну функцію у фагів виконує нуклеїнова кислота, фаговий білок механічно захищає нуклеїнову кислоту від зовнішніх дій і приймає активну участь в процесі проникнення нуклеїнової кислоти в чутливу клітину. З білком пов’язані ферментні системи фагів. Фагова оболонка (капсид) складається з окремих структурних компонентів – субодиниць, які представляють собою поліпептидні ланцюги. Нуклеїнова кислота фагів характеризується різноманіттям структури та молекулярних мас. Основна функція фагів репродукційна, відбувається шляхом інфікування бактеріальних клітин зрілими (активними) фаговими часточками. Даний процес отримав назву фагової інфекції.[7]
3.3 Будова бактеріофагів
Застосування сучасних електронних мікроскопів, а також удосконалення методів приготування препаратів для електронної мікроскопії дозволили більш детально вивчити тонку структуру фагів. Виявилося, що вона досить різноманітна і у багатьох фагів складніша, ніж структура вірусів рослин і ряду вірусів людини і тварин.
Різні фаги відрізняються один від одного не тільки за формою, величині і складності своєї організації, але і за хімічним складом. Виявилося, що фаги, що лізують мікроорганізми різних груп, можуть бути цілком ідентичними за своєю морфологією. Водночас фаги, активні проти однієї і тієї ж культури, можуть різко відрізнятися за своєю структурою.[3]
Переважна більшість фагів має гексагональні голівки, що містять тільки один тип нуклеїнової кислоти. У деяких фагів від головки відходить більш або менш довгий відросток («хвіст»), який служить для адсорбції фага і впорскування в клітину бактерій нуклеїнової кислоти. Є так само довгі ниткоподібні фаги. На підставі морфологіїчних ознак і типу нуклеїнової кислоти Д. Бредлі (D.Bradley, 1967) розділив всі фаги на 6 груп (табл. 3.1) [9]
Таблиця 3.1
Групи фагів
|
Група
|
Морфологічні особливості
|
Тип НК
|
|
А
|
Відросток, що скорочується
|
Дволанцюгова ДНК
|
|
В
|
Довгий відросток, що не скорочується
|
Дволанцюгова ДНК
|
|
С
|
Короткий відросток, що не скорочується
|
Дволанцюгова ДНК
|
|
D
|
Замість відростка вирости на вершинах граней головки
|
Одноланцюгова ДНК
|
|
E
|
Шестигранник без виростів
|
Одноланцюгова РНК
|
|
F
|
Довга нитка (без головки і хвоста)
|
Одноланцюгова ДНК
|
Будову бактеріофагів в основному вивчали на прикладі фагів серії Т Escherichia coli. (рис. 3.1)
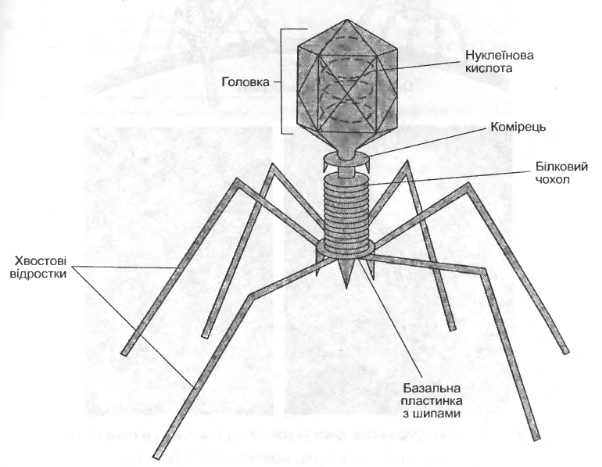
Рис. 3.1. Будова бактеріофага
Коліфаг Т2 складається з поліедричної головки довжиною 100 нм і відростка приблизно такої ж довжини. Тому говорять про «збірні» віруси. Головка складається із капсомерів і містить ДНК. Кількість білків і ДНК приблизно однакова. Відросток має складну будову, в ньому можна вирізнити не менше трьох частин: порожній стержень, який оточує чохол, що має здатність скорочуватись, базальна пластинка з шипами і нитками, від яких залежить специфічна адсорбція на клітині.[1]
3.4 Хімічний склад фагів
Вивчення хімічного складу фагів стало можливим лише тоді, коли були вдосконалені методи отримання великих кількостей очищених препаратів фага. В даний час вивчений хімічний склад фагів, що належать до різних морфологічних типів і вражають мікроорганізми майже всіх систематичних груп.[3]
Вивчення хімічного складу фагів показало, що він досить простий; по суті фаги є нуклеопротеїдами, тобто складаються в основному з білка і нуклеїнової кислоти. Фагові частинки мають кілька різних білків, насамперед структурних, які становлять капсид головки і елементи відростка (чохол, стрижень, базальну пластинку і нитки). У головці булавоподібних фагів є також внутрішній білок (3-7 % загального вмісту білка). У фагів виявлено ферменти лізоцим, фосфатазу та деякі інші. Білки виконують різні функції: захищають нуклеїнову кислоту від пошкоджень і дії ферментів нуклеаз, беруть участь у тісному контакті фага з бактеріальною клітиною, забезпечують через ферментативну дію процес зараження тощо. Другою важливою складовою частиною фагів є нуклеїнові кислоти. У фагів, як і в інших вірусів, е тільки один тип нуклеїнової кислоти — ДНК або РНК. Цією властивістю віруси відрізняються від інших мікроорганізмів, в клітинах яких є обидва типи нуклеїнових кислот. У фагів виявлено дволанцюгову ДНК (найчастіше) і одно-ланцюгові ДНК та РНК. Залежно від типу своєї нуклеїнової кислоти фаги поділяють на ДНК-вмісні і РНК-вмісні. Нуклеїнова кислота щільно упакована у головці фага. У деяких фагів знайдено невеличку кількість ліпідів (2,5-10,5 %), переважно жирних кислот і фосфоліпідів, а також сліди вуглеводів. Значення цих компонентів поки-що недостатньо вивчено. Вважають, що ліпіди та інші компоненти (подібно до інших вірусів) мають клітинне походження і фаговий геном не кодує їхнього синтезу.[8]
3.5 Проникнення в клітину
Найкраще вивчено першу стадію розмноження фагів адсорбцію. Шлях проникнення в клітину у бактеріофагів відрізняється від інших вірусів. Товсті клітинні стінки не дозволяють білкам-рецептора разом з приєднаним до нього вірусом занурюватися в цитоплазму, як це відбувається при інфікуванні клітин тварин. Тому бактеріофаг вводить порожнистий стрижень в клітку і заштовхує через неї ДНК (або РНК), що знаходиться у його голівці. Геном бактеріофага потрапляє в цитоплазму, а капсид залишається зовні. [4]
Не кожен фаг адсорбується на будь-якій бактерії. Специфічність відносин господаря і фага визначається специфічністю адсорбції, яка залежить від рецепторів, наявних у стінці клітини. Рецептори для одних фагів знаходяться в ліпопротеїновому шарі, для інших - у ліпополісахаридному. При надлишку бактеріофага на одній клітині може адсорбуватися 200-300 фагових частинок.[1]
Адсорбція фага на клітиніі реакція вельми специфічна. В клітинній стінці бактерій є особливі структури (рецептори), до яких можуть прикріпитися фаги. Адсорбуються на рецепторах тільки ті фаги, до яких чутлива клітина. [8]
Фаги, які мають відростки, адсорбуються на поверхні фагочутливих бактерій дистальним кінцем цих відростків, а базальна пластинка з шипами і нитками забезпечує тісний контакт. (рис. 3.2)
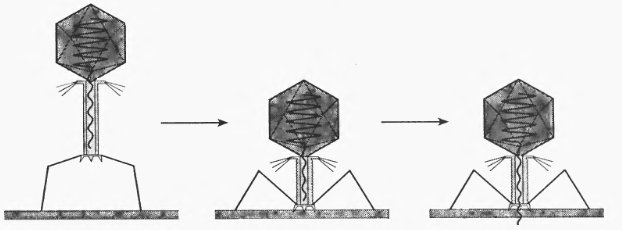
Рис. 3.2. Взаємодія фага з клітинною стінкою
Фаги можуть прикріплятися до різних ділянок клітини джгутиків, ворсинок чи інших виростів. Адсорбція фагів на клітинах - специфічна реакція. Вона зумовлена утворенням тісного зв'язку між спеціальним рецепторним апаратом фага і специфічними рецепторами клітини. Фагорецептори бактеріальної клітини є складними антигенними комплексами або структурами, які розташовані в різних ділянках і шарах клітинної стінки.[10]
Адсорбцію фагів на сприйнятливих до них бактеріях можна спостерігати в електронний мікроскоп. (рис. 3.3)
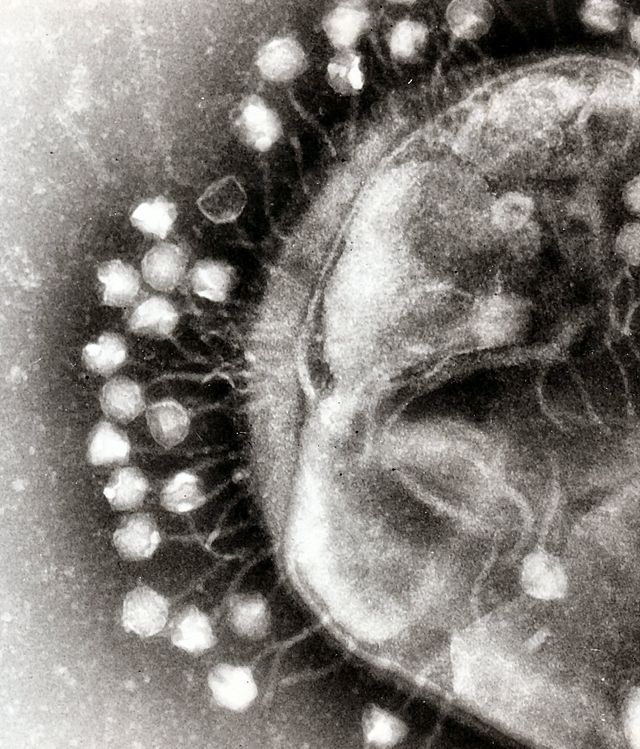
Рис. 3.3. Адсорбція фагів під мікроскопом
Вона залежить від фізичних і хімічних властивостей середовища, температури, природи фага, фізіологічного стану бактерій, а також від їх антигенних структур.[7] Ниткоподібні фаги, а також фаги, що не мають відростків, адсорбуються не так на мікробній стінці, як на ниткоподібних структурах, що оточують стінку фімбріях. Описано фаги, які прикріплюються відростком до бактеріальних жгутиків. У деяких фагів процес адсорбції може здійснюватися лише в тому випадку, коли в середовищі є певні речовини кофактори: амінокислоти (триптофан, тирозин і ін.) Або солі (кальцієві, магнієві).
На кінці фагового відростка є особливий фермент типу лізоциму. Після адсорбції фага під впливом цього ферменту відбувається розчинення стінки мікробної клітини і вміст головки фага - нуклеїнова кислота - перекачується в мікробну клітину. [3]
Слід зазначити, що до сих пір детально не з'ясовано механізм введення нуклеїнової кислоти у фагочутливу клітину фагами, які не мають відростків, а також тими фагами, в яких відростки не скорочуються. [7]
3.6 Розвиток у клітині
Взаємини між фагом і чутливої до нього клітиною дуже складні і не завжди завершуються лизисом клітини і розмноженням в ній фага. Одні бактеріофаги дуже специфічні і здатні лизувати клітини тільки одного якогось виду мікроорганізмів (монофаги), інші клітини різних видів (полифаги).[2]
Бактеріофаг може існувати в трьох станах – профага, вегетативного фага, зрілого фага. У вільному стані фаги інертні. Після адсорбції на клітині нуклеїнова кислота фага проникає всередину клітини, де починає реплікативний цикл. Від зрілого фага така частинка відрізняється в багатьох відношеннях, її називають вегетативним фагом, внаслідок притаманної їй необмеженої здатності до відтворення. Здатністю існувати в третьому стані, а саме в стані профага, характеризуються помірні фаги. Інфікуючи чутливу бактеріальну клітину, помірний фаг може переходити у інтегрований стан, який входить в спадково закріплені симбіотичні взаємовідносини з клітиною-господарем, відомі під назвою лізогенія. Фаги можна також розділити на 2 класи за характером їх життєвої стратегії: вірулентні та помірні. [7]
3.6.1 Вірулентні фаги. Літичний цикл
Літичний тип взаємодії фагів з бактеріями часто ще називають (як і для інших вірусів) продуктивною інфекцією. При такому типі взаємодії фага з клітиною хазяїна розрізняють чотири стадії або етапи: 1) адсорбцію фагів на поверхні бактеріальних клітин; 2) проникнення активного вмісту (нуклеїнової кислоти) в бактеріальну клітину; 3) латентний період (екліпс) Внутрішньо –клітинного розвитку фага; 4) руйнування (лізис) клітини і вихід з неї новоутворених фагів. (рис. 3.4)

Рис. 3.4. Життєвий цикл бактеріофага
Вірулентні фаги можуть репродукуватися лише шляхом літичного циклу; віріон фагу адсорбується на поверхні бактеріальної клітини та впорскує всередину НК, що призводить в кінцевому результаті до припинення всіх метаболічних процесів клітини та встановлення вірусних репродуктивних систем, що розпочинають синтезувати вірус-специфічні білки. Клітини господаря лізуються через певний проміжок часу, вивільняючи нове покоління фагів. [7]
Репродукція вірусу в клітині-господарі процес дуже складний. Його окремі етапи від зародження до звільнення зрілих інфекційних частинок, досить добре вивчені з біохімічного, генетичного та морфологічного боку на прикладі фагів серії Т ( Т2,Т4,Т6). Сприятливою передумовою для таких досліджень стало те, що у фаговій ДНК замість цитозину міститься 5-гідроксиметилцитозин, і тому її синтез легко простежити за появою цієї ознаки. Крім того можна отримати мутантні форми фага, у яких та чи інша стадія процесу репродукції блокована або ж протікає тільки в певних умовах. З допомогою таких мутантів вдалося з'ясувати, як відбувається всередині клітини-господаря морфологічний розвиток фага, тобто в якій часовій послідовності синтезуються різні компоненти фагових частинок.
Подібно до інших вірусів, фаги нерухомі. При змішуванні суспензії вільних фагів з суспензією бактерій фагові частинки в результаті випадкових зіткнень з клітинами прикріплюються до поверхні останніх (адсорбція) [10]
Після адсорбції фага на поверхні бактерій за допомогою ферменту типу лізоцима, який міститься в нижній частині відростка, відбувається розчинення клітинної стінки, і в цей невеличкий отвір кінець відростку, втискуючись (завдяки енергії АТФ), як «шприц», впорскує нуклеїнову кислоту із головки фага в бактеріальну клітину. З моменту проникнення геному фага в бактерію починається третя стадія його взаємодії з клітиною латентний (прихований) період внутрішньоклітинного розвитку фага. Тривалість цього періоду в різних фагів триваєвід 15-40 хв до 5 год і більше. На цій стадії нуклеїнова кислота фага, завдяки закодованій у ній інформації, спричиняє швидку перебудову внутрішніх біосинтезуючих процесів у бактеріальній клітині, повністю спрямовуючи їх на утворення нових частинок фага. На початку третьої стадії розмноження, у екліпс-фазі, виявити в зараженій клітині вегетативний фаг не вдається. Проте саме в цей час під його впливом відбувається пригнічення функцій до синтезу низки клітинних ферментів і водночас індукується процес утворення фагових ферментів або так званих ранніх білків, які каталізують процеси реплікації фагової ДНК з використанням нуклеотидів нуклеїнових кислот самої бактеріальної клітини. Дещо пізніше в клітині починається синтез пізніх білків, які є структурними білками фагів. У результаті агрегації таких білків відбувається побудова окремих елементів нових фагів: головок, відростків, базальних пластинок тощо. Після утворення всіх компонентів фага здійснюється складання дозрілих віріонів фага відповідної форми. Залежно від виду фага, стану бактеріальної клітини та інших чинників у одній клітині може утворитися від кількох десятків до кількох сотень фагових частинок.Отже, в результаті взаємодії вегетативного фага з чутливою клітиною у зараженій бактерії з'являється значна кількість нових корпускул фагів і ми говоримо про репродукцію фагів бактеріальною клітиною на основі генетичної інформації, заданої нуклеїновою кислотою батьківського фага. Саме в цьому й виявляється своєрідна форма паразитизму фагів на субклітинному молекулярному рівні. [7]
3.6.2 Латентні фаги і лізогенія
Помірні фаги мають вибір репродуктивної стратегії. Іноді помірний фаг ініціює літичний процес, що теж приводить до утворення багатьох нових віріонів паралельно з лізисом.
Альтернативним варіантом є лізогенний шлях, при якому замісить реплікації геном фагу переходить в стан спокою, часто інтегруючись з геномом клітини чи зберігаючись у вигляді плазміди. В цьому стані профаг, так ще називають інтегрований фаг, може знаходитися невизначено довгий час, при цьому при реплікації клітини передається дочірнім клітинам. Ці клітини стають лізогенізованими, або/і набувають властивість лізогенних. При певних умовах профаг може вийти зі стану спокою та піти по літичном шляху. До найбільш досліджених помірних фагів відносяться фаг , Р1, Р22, u. Варто відмітити, що мутації деяких генів, що відповідають за лізогенний тип розвитку, можуть приводити до появи вірулентних нащадків, які все-таки відносяться до групи помірних фагів. [7]
Лізогенний стан бактерій характеризується трьома особливостями:
- Кожна клітина потенційно здатна утворювати фаг, але за звичайних умов у популяції лізогенних бактерій лише невелика частина клітин знаходиться в стані продуктивної інфекції (за рахунок спонтанної індукції).
- В стані профага більшість генів фага виключено активністю репресора.
- Перехід клітини-лізогена в стан продуктивної інфекції проходить при інактивації репресора в клітині.
Найліпше вивченим помірним фагом є фаг (рис. 3.5). Він може або передаватися з клітини у клітину в прцесі інфекції, або від одного покоління до наступного в ході розмноження цього штаму бактерій. В останньому випадку латентний геном фага названо профагом, а клітини, які містять такий профаг — лізогенними.

Рис. 3.5. Фаг
Наявний геном фага в лізогенній культурі можна виявити при спонтанному звільненні фага з невеликої частини клітинної популяції, в якій відбувався розвиток фага.
Стабільність підтримання лізогенного стану бактерій забезпечується синхронністю реплікації профага і генома бактерій та наявністю білка-репресора, який виключає всі гени профага, що мають відношення до літичного циклу. Синхронність реплікації геномів бактерій і профага досягається у різних фагів по-різному. Профаги деяких фагів (, Р2) за звичайних умов інтегровані в бактеріальну хромосому і реплікуються як її невід’ємна частина.
У лізогенних клітинах профаг міцно зв’язаний з хромосомою клітини-господаря. Під час кон’югації клітин профаг разом з хромосомою господаря передається із клітини-донора у клітину-реципієнт. Фаг приєднується до хромосоми господаря в певному місці ( між галактозним і біотиновим оперонами) (рис. 3.6)
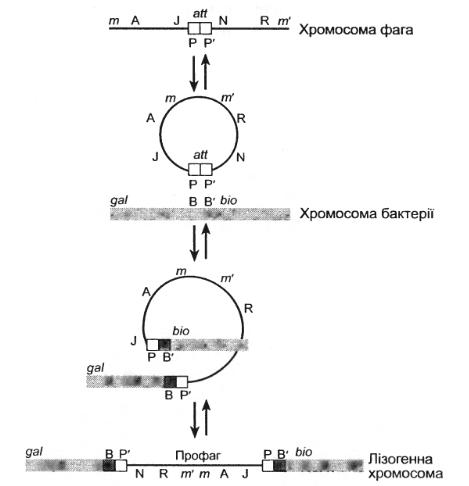
Рис. 3.6. Інтеграція фага в хромосому бактерії
Спочатку вважали, що ДНК бактеріофага тільки прикріплюється до хромосоми бактерії в цій ділянці. Пізніше було довелено, що фагова ДНК при лізогенізації не просто прикріплюється до бактеріальної ДНК, а включається в неї (специфічна інтеграція).
Включення ДНК профага в хромосому клітини відбувається в результаті розриву і перехресного з'єднання. За цю реакцію відповідає фермент лямбда-інтеграза.
Після інфікування клітини помірним фагом відбувається вибір шляху його розвитку: лізогенний чи літичний.
В інтегрованому стані фагова ДНК реплікується разом з бактеріальною, піддається тим самим регуляторним впливам, що і бактеріальна хромосома. Інформація, що міститься у фаговій ДНК, у цей час не реалізується. Тільки після переходу профага у вегетативний стан відновлюється автономність фагової ДНК і починається розмноження фага. Цей зворотний процес може відбуватися спонтанно або в результаті індукції ( наприклад, під дією УФ-променів). Вилучення фагової ДНК із бактеріальної хромосоми проходить, ймовірно, шляхом зворотних процесів, які зумовлюють її включення, і здійснюється дуже точно: більше 99% фагових часток, які звільняються з лізогенних клітин, ідентичні з вихідними ( інфікуючим фагом). [10]
Як тільки профаг перейшов у вегетативний стан, він знову стає автономним і може розмножуватись в бактеріальній клітині як вірулентний фаг, що призводить до лізису бактерії і вивільнення фага лямбда. [1]
3.7 Зборка частин бактеріофага
На відміну від ранніх етапів розвитку фага хід збирання капсидів і повних віріонів не програмується послідовною експресією фагових генів. Всі білки віріона синтезуються і нагромаджуються в клітині, утворюючи “фонд попередників”. Звідси вони виділяються щляхом прямої специфічної дії з іншими білковими молекулами, в результаті чого утворюють субструктури, які потім перетворюються в цілі віріони. Збирання віріонів фага Т4 протікає у трьох напрямках (рис. 3.7):
- Базальна пластинка фагового відростка побудована з 15 білків, в синтезі яких, окрім основних, беруть участь і інші гени. Пластинка містить декілька молекул ферментів, які кодуються фагом, - дигідрофолатредуктазу і тимідилатсинтетазу, а також деяку кількість фолієвої кислоти.
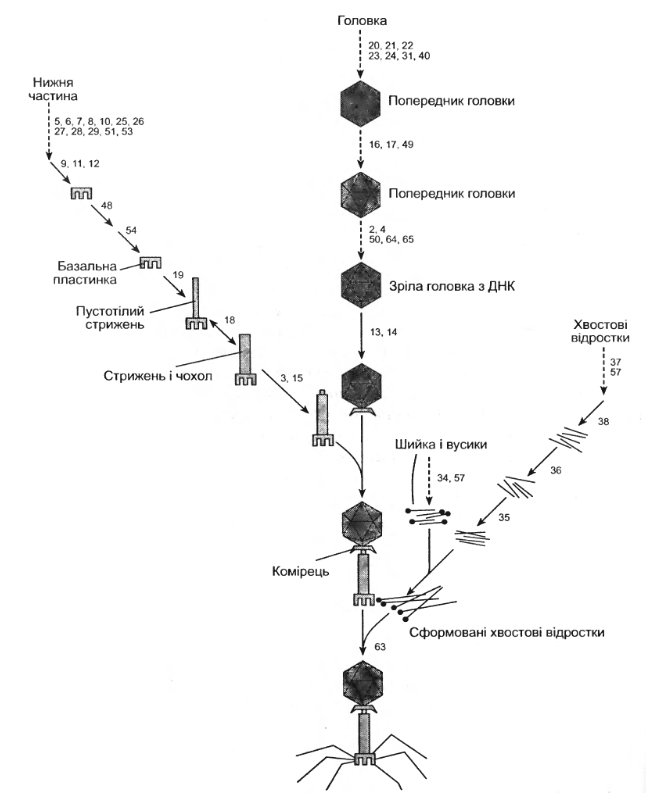
Рис. 3.7. Схема збирання фага
- Зібрана базальна пластинка після прєднання до неї білка продукту гена 54 слугує заправкою для збирання стрижня відростка, який складається із 144 молекул продукту гена 19. Навколо стрижня проходить збирання чохла — полімера, що побудований із 144 молекул продукту гена 18. Продукти двох інших генів стабілізують цю структуру.
- Оболонка фагової головки, що побудована з більш як 10 білків, утворюється у результаті активності багатьох генів. Основний із них — це продукт гена 23, який входить до складу вершини головки лише після відщеплення від основного поліпептиду фрагмента з молекулярною масою 10 кДа. Протеоліз здійснюється продуктом гена 22, а також 21, якого бракує в зрілому віріоні. Білок гена 22 — це внутрішній білок, який у кінці процесу перетворюється у дрібні пептиди, причому деякі з них залишаються в головці фага. Тут наявні й інші внутрішні білки, що піддаються частковому перетворенню білком гена 22.
- Після закінчення роздільного збирання головки і відростка, вони об’єднуються як in vitro, так і in vivo.
- Нитки відростка складаються із продуктів чотирьох генів. Їх збирання відбувається незалежно, але прикріплюються вони до базальної пластинки лише після об’єднання головки і хвостового відростка. Для цієї реакції потрібний білок гена 63, а також взаємодія з “вусиками”, прикріпленими до комірця, розміщеного між головкою і хвостом. Головка фага має специфічну форму, що визначається білком гена 23. Її будова змінюється в результаті мутацій певних генів. Сформована головка фага Т4 має форму неправильного ікосадельтаедра, по повній осі якого розміщений додатковий ряд субодиниць, що складається із 840 копій білка гена 23. Субодиниці білка гена 20 розміщені на вершинах. Така форма головки відображає наявність певних просторових обмежень. Якщо немає обмежень, будова фага дуже змінюється.
На заключному етапі внутрішньоклітинного розвитку фага вмикається функція гена, яка відповідає за руйнування бактеріальної оболонки і звільнення дозрілого фага (рис. 3.8)
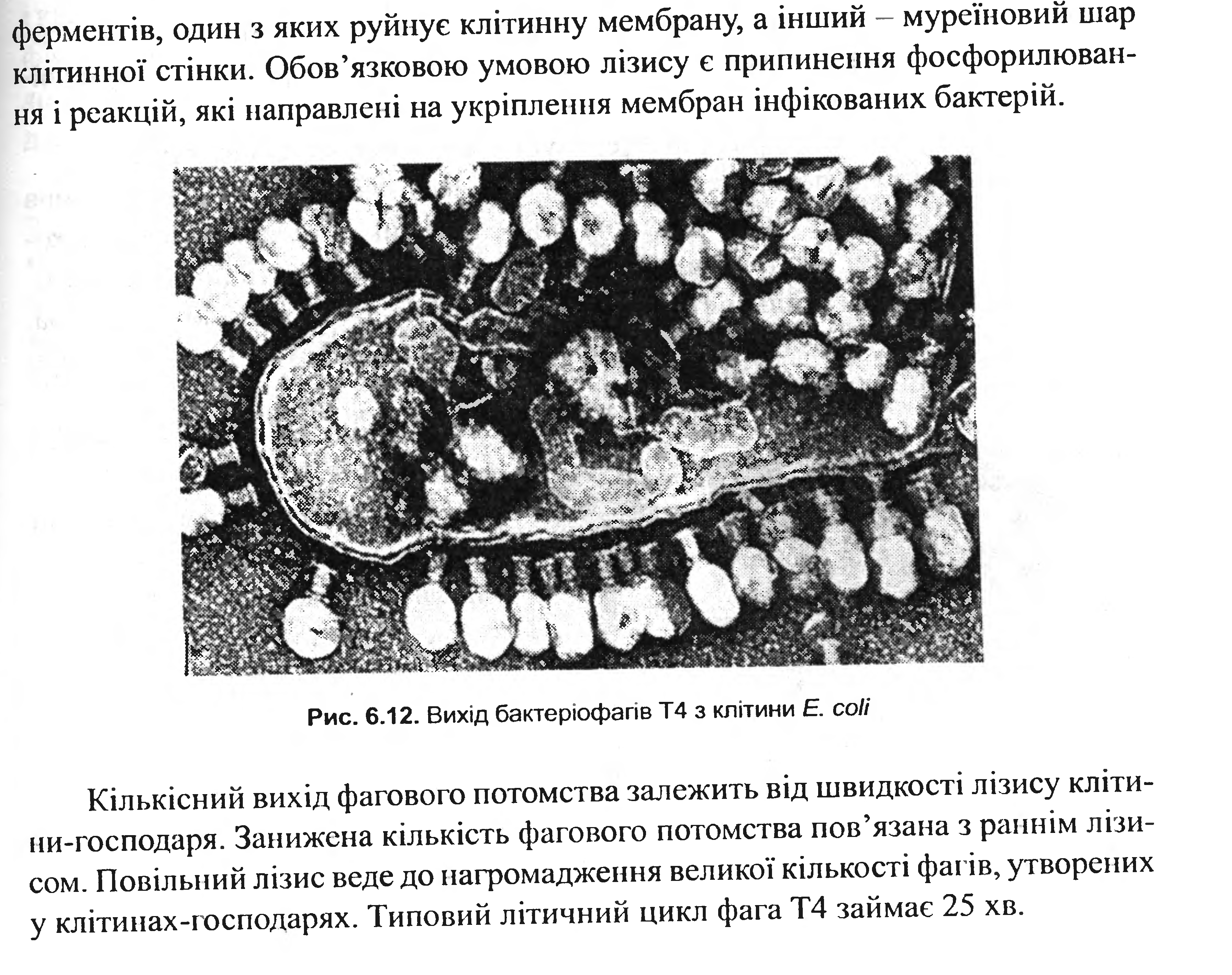
Рис. 3.8. Вихід бактеріофагів з клітини
У багатьох фагів механізми лізису функціонують по-різному. Т-парні фаги контролюють утворення двох різних літичних ферментів, один з яких руйнує клітинну мембрану, а інший – муреїновий шар клітинної стінки. обов’язковою умовою лізису є припинення фосфорилювання і реакцій, які направлені на укріплення мембран інфікованих бактерій.
Кількісний вихід фагового потомства залежить від швидкості лізису клітини-господаря. Занижена кількість фагово потомства пов’язана з раннім лізисом. Повільний лізис веде до нагромадження великої кількості фагів, утворених у клітинах-господарях. [10]
4. Походження бактеріофагів
Незважаючи на те, що явище бактеріофагії інтенсивно вивчається більше п'ятдесяти років, на природу фагів немає єдиної точки зору, і це питання до сих пір залишається спірним. В спеціальній пресі та на конференціях неодноразово виникали гарячі дискусії на цю тему. І це не випадково. Питання про походження фагів, як і в інших вірусів, має велике значення, так як з ним тісно пов'язане вирішення багатьох актуальних завдань сучасної біології: походження життя, можливі форми існування живого; існування живих істот, що не мають клітинної структури; походження клітинних форм життя; розвиток, мінливість і видоутворення у мікроорганізмів та ін. До теперішнього часу все ще існують діаметрально протилежні точки зору на природу вірусів, в тому числі і фагів. На думку одних вчених, фаги відносяться до живих організмів; інші розглядають їх як особливі речовини типу ферментів.
Важливо відзначити, що ті дослідники, які відносять фаги до живих організмів, по-різному трактують питання про їх походження. Одні дослідники вважають, що фаги, як і віруси людини, тварин і рослин, походять від найдавніших доклітинних форм, які в процесі еволюції пристосувались до паразитування у первинних одноклітинних організмах та у подальшому еволюціонували разом зі своїми господарями. Таким шляхом, як вважають ці вчені, виникли фаги мікроорганізмів, які по своєму відношенню до клітини-господаря є паразитами екзогенного, тобто зовнішнього походження. Інші ж вважають, що походження фагів пов'язано тим чи іншим чином з клітиною свого теперішнього господаря (ендогенне походження). На думку учених, які розглядають фаг як фермент ендогенного походження, фагова частка є продуктом життєдіяльності мікробної клітини. При попаданні в клітину фаги викликають процеси утворення активного фага, здатного руйнувати мікробну клітину. А розмноження фага в клітині відбувається приблизно так само, як утворення активного ферменту з його неактивного попередника - профермента. За останні роки завдяки застосуванню новітніх сучасних методів дослідження (електронна мікроскопія, мічені атоми) знання про структуру фагів, їх хімічний склад, особливості розмноження значно розширилися. Фагова частка виявилася досить складно організованою. Вона містить основні хімічні сполуки, не властиві живому організму,- нуклеїнові кислоти та білок. Подібно іншим живим істотам, фаги здатні змінювати всі свої властивості. Тому розглядати їх як ферменти немає ніяких підстав. Вірно, вони не володіють власним обміном речовин. Вони є абсолютними паразитами, що живуть повністю за рахунок клітини-господаря.
Отже, фаги логічно розглядати як особливі форми живих істот. Проте питання про походження фагів поки ще не можна вважати остаточно вирішеним. [3]
5. Значення бактеріофагів
5.1 Роль у природі
Фаги є найбільш численною групою вірусів на Землі. Їх можна знайти у всіх можливих еконішах, що зайняті їх господарями –бактеріями. Бактеріофаги широко розповсюджені в природі. [8]
На даний момент не існує жодної відомої бактерії, яка б не могла бути ураженою бактеріофагом чи не несла у своєму геномі профаг (інтегровану нуклеїнову кислоту фага). Бактеріофаги відомі для усіх типів бактерій без виключення: від бактеріальних паразитів роду Dellovibrios і до порівняно крупних синьо-зелених водоростей. [10]
Еволюційна роль полягає у тому, що вони здатні включатися у генетичну рекомбінацію і давати початок новим спадковим композиціям. [11]
5.2 Вплив на бактерії
Помірні фаги можуть „захищати” своїх господарів проти інфекції спорідненими фагами та викликати досить значні зміни властивостей своїх господарів, включаючи зміни в системі рестрикції-модифікації, стійкості до антибіотиків та інших стресових чинників зовнішнього середовища. Деякі помірні фаги навіть можуть перетворювати своїх господарів до патогенного фенотипу, як наприклад, дифтерійний чи ентерогеморагічні (EHEC) штами E. coli. [7]
Важливою особливістю багатьох фагів є здатність переносити бактеріальні гени – трансдукція. Розрізняють специфічну і загальну трансдукцію.Специфічна трансдукція характерна для помірних фагів. Це пов'язано з тим, що їх профаг існує в складі хромосоми бактерій і при індукції, тобто при перетворенні профага в зрілий фаг, може захоплювати бактеріальні гени, що розташовуються з одной чи іншої сторони від місця включення профага в хромосому («незаконна рекомбінація»). Тоді в популяції бактерій з’являються клітини з незвичайним фенотипом трансдуктантом. Механізм їх виникнення полягає у вбудовуванні генома дефектного фага в хромосому нового господаря і експресії генів, захоплених при індукції профага у колишнього господаря. При загальній (іноді її називають також неспецифічною) трансдукції частинки фага являють собою восновному шматок бактеріальної ДНК, упакований у фаговую оболонку. Це відбувається в процесі зборки капсида фага,коли вже починається дезінтеграція клітини господаря і формуються зрілі частки фага. При загальній трансдукції одним і тим же «фаголізомтом » можуть переноситися будь-які гени клітини донора, в чому легко переконатись шляхом використання належних методів селекції трансдуктантів (звідси й назва - «неспецифічна» трансдукция). З викладеного випливає, що нові властивості, що з'являються у бактерій при трансдукції, зобов'язані генам бактерій, які приносяться фагами. Однак відомо чимало випадків, коли властивості бактерій визначаються тільки фаговими генами. Придбання нових властивостей бактеріями виключно в результаті фагової інфекції називається фаговою конверсією, типовим прикладом якої служить токсигенність збудника дифтерії. Цей мікроб утворює токсин тільки тоді, коли він заражений фагом, що несе tox ген. Таким чином, конверсія принципово відрізняється від специфічної трансдукції, оскільки в останньому випадку фаг переносить гени, які в нормі виявляються в хромосомі клітини донора. Питання про походження конвертуючих фагів залишається відкритим, хоча є припущення, що в процесі еволюції хромосоми бактерій і фагів мали спільних попередників.
Роль фагів в еволюції бактерій явно недооцінювали. Тепер же можна сміливо стверджувати, що в горизонтальному перенесенні генів вони мають не менше, а можливо, й більше значення, ніж кон'югація і трансформація. Усвідомленню цього факту сприяли, зокрема, численні дані про здатність фагів
тривалий час зберігатися у навколишньому середовищі (в грунті, воді) без втрати інфекційності. Втім, те ж стосується інших вірусів, що служить підставою для відродження гіпотези про давнє походження вірусів та їх ролі в еволюції клітини. [9]
5.3 Використання бактеріофагів у діяльності людини
Дослідження бактеріофагів сприяло розумінню багатьох генетичних явищ. Швидке розмноження бактеріофагів дає можливість за одну добу здійснювати схрещування у потомстві двох послідовних поколінь. Для аналогічних схгещувань на профілі потрібно 3,5 тижнів, а на кукурудзі — щонайменше рік. Крім того велика кількість фагових популяцій, які містяться в кількох мілілітрах культуральної рідини , дає змогу спостерігати дуже рідкісні генетичні явища. Малий розмір геномів багатьох фагів, порівняно з геномом бактерій, дає змогу ідентифікувати всі або більшість фагових генів і досить детально уявити собі генетичну організацію і регулювання генома. Геном фага Х174 містить лише 11 генів, фага — менше 60, тоді як геном Е. coli нараховує декілька тисяч генів. Ці переваги зробили бактеріофаги незамінними генетичними об’єктами. Вони можуть служити моделями під час аналізу будови і роботи більш складних геномів. Ф. Туорт і Ф. Д’Ерель першими, і незалежно один від одного, виявили трансмісивні репродукуючі агенти, здатні проходити через бактеріальні фільтри і спричиняти лізис бактерій. З моменту відкриття бактеріофагів були зроблені спроби використати їх для боротьби з патогенними бактеріями. Бактеріофаги є ідеальним об’єктом для біохіміків, генетиків і мікробіологів, котрі вивчають фундаментальні механізми життєвих процесів. [10]
Практичне застоcування бактеріофагів:
- Епідеміологічне фаготипування.
- Фагоіндикація бактеріальних культур.
- Використання в молекулярній біології.
- Використання бактеріофагів для експресії білків (phage display).
- Екологічний моніторинг.
- Біоконтроль харчової продукції за допомогою фагів.
- Фагова терапія.
- Боротьба з патогенними бактеріями.Використання вірулентних фагів в якості лікувальних засобів проти захворювань, викликаних особливо стійкими патогенними бактеріями.[7]
- Висновки
Бактеріофаги – особлива група вірусів, що уражає бактерії. Вони були відкриті канадським дослідником Феліксом д'Ерелем у 1917 році. Фаги подібно до всіх інших вірусів не мають клітинної будови та є обов'язковими внутрішньоклітинними паразитами. Залежно від типу взаємодії з мікробною клітиною розрізняють вірулентні та помірні, помірно-вірулентні та вірулентно-помірні бактеріофаги. Більшість фагів складається з головки, що являє собою молекулу ДНК або РНК, оточену білковим капсидом та відростку. Будова фагів досить різноманітна і у багатьох складніша, ніж структура вірусів рослин і ряду вірусів людини і тварин. Хімічний склад фагів досить простий – вони є нуклеопротеїдами, тобто складаються з білка і нукленових кислот. Найкраще вивчено першу стадію розмноження фагів адсорбцію: фаг приєднується до клітинної стінки і, проколюючи її, мов шприцом, вводить свою НК. Це специфічна реакція, зумовлена взаємодією спеціальних рецепторів.
Взаємодія віруса з клітиною хазяїна починається після випадкового зіткнення у середовищі. Життєвий цикл бактеріофагів, які потрапили до клітини, може проходити двома шляхами, які суттєво відрізняються:
- Літичний шлях – вірус, потрапивши в клітину, стимулює синтез білків, необхідних для утворення нових вірусів.
- Лізогенний шлях – вірус у клітині включається у ДНК господаря.
Збирання віріонів фага протікає у трьох напрямках і не програмується послідовною експресією генів. Білки синтезуються, нагромаджуються і шляхом самозбирання утворюють віріони, які руйнують бактеріальну клітину і виходять з неї.
Походження фагів остаточно не з'ясоване. Еволюційна роль полягає у тому, що вони здатні включатися у генетичну рекомбінацію і давати початок новим спадковим композиціям. Бактеріофаги є зручними об'єктами для генної інженерії, також знайшли застосування в медицині, біології, для контролю харчової продукції, розпізнавання бактерій.
7. Список літератури
- Шлегель Г. Общая микробиология/ Шлегель Г.; пер. с нем. М.: Мир, 1987.-576с.
- Тейлор Д. Биология в трех томах. Т. 1/ Тейлор Д., Грин Н., Стаут У.; пер. с англ. Ю. Л. Амченкова, М. Г. Дуниной, Н. Ю. Замаэвой, Л. Г. Тер-Саркисян, Н. О. Фоминой М.: Мир, 2004.-454 с.
- Природа, морфологія і основні властивості бактеріофагів. Механізм дії на бактеріальну клітину. Застосування їх у діагностиці [Електронний ресурс]/ Генріх О. В. 2008. Режим доступу uadok.zawantag.com/text/40492/index-1.html
- Мамонтов С. Г. Биология для поступающих в вузы/ Мамонтов С. Г. Биология: Справ. издание. М.: Высш. шк. 1992.- 478с.:ил.
- Кемп П. Введение в биологию/ Кемп П., Армс К.; пер. с англ. М. Мир, 1980.- 303с.
- Свенсон К. Клетка/ Свенсон К., Уебстер П.; пер. с англ. Т. Днепровской. М.: Мир, 1980.- 303с.
- Андрійчук О. М. Методичні рекомендації до спецпрактикуму «Віруси мікроорганізмів»/ Андрійчук О. М., Харіна А. В., Поліщук В. П. К.: КНУ ім. Т. Шевченка,- 2011.- 45с.
- Векірчик К. М. Мікробіологія з основами вірусології/ Векірчик К. М. К.: Либідь, 2001.- 312с.
- Домарадский И. В. Основы вирусологии для екологов/ Домарадский И. В. М.: ЛексЭст, 2007.- 80с.
- Гудзь С. П. Загальна вірусолоогія/ Гудзь С. П., Перетятько Т. Б., Павлова Ю. О. Львів: Видавничий центр ЛНУ ім. І. Франка, 2010.- 264с.
- Біологія/ Шелест З. М., Войціцький В. М., Гайченко В. А., Байрак О. М. К.: Кондор, 2007.- 760с.
Бактеріофаги. Зборка частин бактеріофага. Розвиток бактеріофага в клітині