Актинобациллезная плевропневмония и гемофилёзный полисерозит свиней
МИНИСТЕРСТВО ОБЩЕГО И ПРОФЕССИОНАЛЬНОГО
ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
Московский Государственный университет
прикладной биотехнологии
На правах рукописи
УДК 619.4 : 616.9
СКОРОДУМОВ
Дмитрий Иванович
АКТИНОБАЦИЛЛЁЗНАЯ (ГЕМОФИЛЁЗНАЯ)
ПЛЕВРОПНЕВМОНИЯ И ГЕМОФИЛЁЗНЫЙ ПОЛИСЕРОЗИТ СВИНЕЙ ( ЭТИОЛОГИЯ, ЛАБОРАТОРНАЯ ДИАГНОСТИКА, ОСНОВЫ СПЕЦИФИЧЕСКОЙ ПРОФИЛАКТИКИ АКТИНОБАЦИЛЁЗНОЙ ПЛЕВРОПНЕВМОНИИ )
16.00.03. тАУ ветеринарная микробио-
логия, вирусология, эпи-
зоотология и микология
АВТОРЕФЕРАТ
диссертации на соискание
учёной степени доктора
ветеринарных наук
Москва тАУ 1997
- Общая характеристика работы
Актуальность проблемы. Болезни свиней представляют одну из значимых проблем ветеринарной науки и практики. Перевод свиновод-ства на промышленную технологию выявил значение ряда инфекционных болезней, которые до этого оставались за пределами внимания специалистов. К этой категории бактериозов свиней можно отнести актинобациллёзную ( гемофилёзную ) плевропневмонию и гемофилёзный полисерозит свиней ( Сидоров М.А., 1987, 1988; J. Nikolet, 1976, 1983, 1986; Schultz R. A., 1985; Schope R. E. et al, 1964; Sebunya T.N.K. et al, 1983; Rsing H. J., 1979,1980, 1991; Rosendal S. et al, 1983; Nielsen R., 1982,1995; Kielsten P. и.а., 1980, 1190, 1992, 1994; и другие)
Актинобациллёзная ( гемофилёзная ) плевропневмония приобрела повсеместное распространение, наносит значительный экономический ущерб, с большим трудом поддаётся лечению и специфической профилактике ( Beaudet R, et al, 1994; Byrd W. et al 1992; Fedorka тАУ Grey P. S. et al., 1990; Beskow P. et al .,1989; Bigbee H. C. et al, 1986; и др. )
Гемофилёзный полисерозит ( болезнь Глессера ) в связи с внедрением в технологию свиноводства использования СПФ тАУ животных неожиданно проявил себя как заболевание, способное поражать все возрастные группы свиней, в отличие от сложившегося представления как о болезни, к которой чувствительны поросята- -отъёмыши, подвергшиеся воздействию стресстАФфакторов ( Amano H. et al., 1987; Bachler J. F. et al., 1974; и др. ).
В России исследования по указанным вопросам были начаты в 1974г. ВНИИЭВ им. Я. Р. Коваленко под руководством профессора Сидорова М. А. Имевшиеся к этому периоду публикации в отечественной литературе по биологии гемофильных бактерий, патогенных для свиней, были недостаточны для эффективной лабораторной диагностики инфекций, вызываемых Actinobacillus ( Haemophilus ) A.pleuropneumoniae и H.parasuis. Разработки по диагностике и специфической профилактике перечисленных болезней практически отсутствовали.
Цели и задачи исследований.
Изучить биологические свойства НАДтАФзависимых бактерий, вызывающих генерализованные серозиты и фибринозно - -геморрагическую плевропневмонию у свиней. На основе результатов таксономического анализа разработать схему идентификации указанных возбудителей. Исследовать возможность специфической профилактики актинобациллёзной плевропневмонии свиней. Для достижения сформулированных целей были поставлены следующие задачи:
- Собрать коллекцию штаммов НАД--зависимых бактерий, ассоциирующихся с серозитами и фибринознотАФгеморрагическими пневмониями свиней, провести сравнительное изучение их фенотипических и генотипических характеристик. Определить критерии их дифференциации от таксономически близких и сходных видов бактерий.
- Изучить чувствительность к H.parasuis и A.pleuropneumoniae различных видов лабораторных животных с целью выбора лабораторной модели для диагностики, выяснения вопросов патотАФи иммуногенеза.
- Исследовать патогенность A.pleuropneumoniae и H.parasuis для свиней.
- Изучить методы серологической идентификации выделенных культур H.parasuis и A.pleuropneumoniae и их серовариантную структуру.
- Разработать и апробировать ускоренные методы обнаружения антигенов A.pleuropneumoniae.
- Изучить возможность специфической профилактики актинобациллезной плевропневмонии свиней при помощи инактивированной вакцины.
Научная новизна работы.
Установлена этиологическая роль H.parasuis в развитии генерализованных серозитов и Actinobacillus ( Haemophilus ) pleuropneumoniae в заболевании свиней фибринознотАФгеморрагической плевропневмонией в условиях отечественных свиноводческих хозяйств промышленного типа. Эти нозологические единицы впервые диагностированы в свиноводческих хозяйствах России.
Проведено комплексное изучение фенотипических и генотипических характеристик НАД--зависимых бактерий, выделенных от свиней при указанной патологии. Подтверждена принадлежность бактерий классифицируемых ранее как вид Haemophilus pleuropneumoniae к роду Actinobacillus. На основании нумерического анализа фенотипических признаков НАД--зависимых культур собранной коллекции выделены феноны, соответствующие видам H.parasuis, A.pleuropneumoniae и таксону Влмалая группаВ» и определены дифференцирующие признаки между указанными видами и другими таксонами семейства Pasteurellaceae. Определены критерии дифференциации культур НАД-- независимого биовара A.pleuropneumoniae от сходных бактерий.
Экспериментально обоснованы методы серологической идентификации культур A.pleuropneumoniae и H.parasuis, а также обнаружения антигенов A.pleuropneumoniae в тканевом материале. Определена серотиповая структура НАД--зависимых культур A.pleuropneumoniae и H.parasuis, выделяемых при соответствующей патологии свиней.
Изучена патогенность A.pleuropneumoniae и H.parasuis для лабораторных животных и свиней. Показано значение гемолизинов и экзотоксинов A.pleuropneumoniae для вирулентности возбудителя и очерчена их роль в патогенезе актинобациллёзной плевропневмонии свиней.
Дано научное обоснование методов лабораторной диагностики гемофилёзов свиней и технологии изготовления инактивированной эмульгированной вакцины против актинобациллёзной плевропневмонии свиней ( авторское свидетельство № 907899 от 21.10.1981г. ) , доказана эффективность активной иммунизации против актинобациллезной плевропневмонии свиней в условиях неблагополучного хозяйства.
Практическая ценность работы.
Разработаны и используются в ветеринарных диагностических лабораториях и свиноводческих хозяйствах:
-- Временные методические указания по лабораторной диагностике гемофилёзного полисерозита поросят. Рекомендованы МСХ СССР 17.10.1978г. № 116-18.
-- Временные методические указания по лабораторной диагностике гемофилёзного плевропневмонии свиней. Утверждены ГУВ МСХ СССР 16.04.1981г.
-- Методические указания по диагностике гемофилёзов свиней. Утверждены ГУВ МСХ СССР 02.11.1985г. №115-6а.
-- Методические указания на изготовление и применение коагглютинирующего антительного диагностикума для экспресс тАУ идентификации Гемофилюс плевропневмоние и индикации возбудителя в патологическом материале. Утверждены отделением ветеринарии РАСХН 16.02.1993г.
Временная инструкция о мероприятиях по борьбе с гемофилёзами свиней. Утверждена ГУВ МСХ СССР. 02.11.1985г.
№ 115-6а.
Разработаны и утверждены:
- Инструкция по изготовлению и контролю вакцин против гемофилёзной плевропневмонии свиней. Утверждена ГУВ МСХ СССР 11.07.1990г.
Вакцина против гемофилёзной плевропневмонии свиней эмульгированная. Технические условия ТУтАФ10-09-53-90. Утверждены ГУВ МСХ СССР 11.07.1990г.
- Наставление по применению вакцины против гемофилёзной плевропневмонии свиней. Утверждено ГУВ МСХ СССР 11.07.1990г.
Основные положения, выносимые на защиту:
- Результаты комплексного изучения биологических свойств и таксономического положения возбудителей актинобациллёзной плевропневмонии и гемофилёзного полисерозита свиней, критерии их дифференциации по фенотипическим свойствам от таксономически и экологически родственных видов бактерий.
- Методы серологической идентификации культур A.pleuropneumoniae и обнаружения антигенов возбудителя в тканевом материале.
- Принципы изготовления и лабораторного контроля инактивированной вакцины против актинобациллёзной плевропневмонии свиней, результаты испытания эффективности вакцины в производственных условиях.
Апробация работы.
Материалы диссертации доложены на заседаниях учёного и методического советов Всероссийского научно-исследовательского института экспериментальной ветеринарии им. Я. Р. Коваленко
(1975-1983г.г.), учёного совета ветеринарно - санитарного факультета Московского Государственного университета прикладной биотехнологии ( 1984-1995г.г.), ветеринарной секции Научно-
-технического совета Министерства сельского хозяйства СССР (23.11.1983г.), на семинаре бактериологов республиканских ветеринарных лабораторий (г.Тбилиси,1980г.), на семинаре бактериологов областных ветеринарных лабораторий РiСР (НПВЛ РiСР, 1981г.), на совещании специалистов свиноводческих комплексов СССР (г.Москва, ВДНХ, 1980г.), на научных конференциях ВНИИЭВ им. Я.Р. Коваленко (1993г.), МГАВМиБ им. К. И. Скрябина (1991,1996г.г.)
По материалам диссертации опубликована 41 научная работа,
методические указания и наставления; получены два авторских свидетельства на изобретения. Опубликована в соавторстве с
М.А.Сидоровым монография ВлГемофилёзы животныхВ» (М.,Колос,1986).
Объём и структура диссертации. Диссертация изложена на 552 страницах машинописного текста и состоит из выведения, двух глав обзора литературы, материалов и методов исследований, двух глав собственных исследований, обсуждения результатов, выводов, практических предложений и приложения. Диссертация содержит 82 таблицы, 30 рисунков. Список литературы содержит 472 источника, в том числе 61 отечественных и 411 иностранных авторов.
Собственные исследования.
- Материалы и методы исследований.
Работа выполнена во ВНИИЭВ им. Я. Р. Коваленко и МГУПБ в период 1974тАФ1995г.г. Выполненные исследования являлись частью плановой научной тематики ВНИИЭВ и МГУПБ. Отдельные этапы исследований проведены совместно с Сидоровым М.А., Шубиным В.А., Гумбатовым Ю.К., Мицаевым Ш.Ш., Блему Ж., Силла Ф., Лаврентьевым Н.И., Романовой Л.Я., Субботиным В.В., ПруссактАФГлотовым В.Э., Логиновым И.А., Корнелаевой Р.П.
Ряд комплексных исследований проведен совместно с сотрудниками лаборатории патологической анатомии ВНИИЭВ проф. Шубиным В.А., Татришвили И.К., Андрышиным А.Н., лаборатории молекулярной биологии и биохимии ВНИИЭВ Артюхиным С.К. и Фоминым Б.А.
В работе использовали 30 штаммов типовых культур бактерий семейства Pasteurellaceae: Actinobacillus ( Haemophilus ) pleuropneumoniae
( серовары 1,2,3,4,5,6,7,8,9,10,12, ) , Pasteurella тАУ haemolytica тАУ подобные бактерии, гемофильные бактерии Влмалой группыВ», таксона ВлСВ», H.influenzae, H.parainfluenzae, H.parasuis ( серовары А,В,С,Д ), P.multocida ( серовары А.В.Д ). Также использовали культуры S.aureus (шт.№СТС 8530, 209 р.), S.epidermidis (шт.АТСС14990),S.saprophyticus (шт.2284 ИГ),
В.megatherium (шт. №654), B.subtilis, S.cholerasuis (шт.№370), S.typhimurium (шт.№415).
Подробному изучению включая нумерический анализ, подвергнут 141 штамм НАД--зависимых эпизоотических культур бактерий, выделенных нами от свиней, и 160 штаммов подвергнуты серологической идентификации. Бактериологически или патологоанатомически исследованы материалы от более 2200 свиней.
При изучении вопросов пато- и иммуногенеза использовали 157 свиней, 210 морских свинок, 546 белых мышей.
Для культивирования НАД--зависимых бактерий применяли ВлшоколадныйВ» агар, бульон и агар Левинталя, сывороточно-
-дрожжевой агар и бульон, кровяной агар и бульон на основе МПБ, МПА, бульона и агара Хоттингера. В качестве источников специфических ростовых факторов использовали кристаллический гемин (ВлРеахимВ»), НАД (ВлРеахимВ»), дрожжевой экстракт, кровь различных видов животных, питающие культуры бактерий (ВлбаккормилкиВ»). Культивирование бактерий в жидких питательных средах осуществляли в стационарных условиях и в режиме аэрирования (лабораторный ферментер). Морфологические, тинкториальные, культуральные и ферментативные свойства бактерий
исследовали общепринятыми методами. В дифференциальнотАФдиагностические среды при определении ферментативной активности добавляли НАД и гемин, а также использовали СИБ и ПВДЭтАФдиагностические наборы производства Горьковского ИЭМ. Особенности колоний изучали в косопадающем пучке света при помощи стереоскопического микроскопа МБСтАФ1, (Акатова Н.С.,1966).
При проведении нумерического анализа у каждого исследованного штамма бактерий определяли 57 физиологических признаков, показатели преобразовывали в числовую форму, затем при помощи ЭВМ вычисляли коэффициент несоответствия по формуле
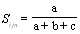 где:
где:
a - количество несовпадающих признаков,
b - количество совпадающих положительных признаков,
c - количество совпадающих отрицательных признаков.
Полученную матрицу коэффициентов подвергали кластерному анализу с представлением конечных данных в виде дендрограммы, отображающей распределение штаммов по кластерам ( фенонам ).
Для генетической характеристики ( исследования проведены совместно с Артюхиным С.К. и Фоминым В.А. ) эпизоотических и референтных штаммов ДНК выделяли по методу Marmur (1961 ). Нуклеотидный состав определяли спектрофотометрически по тепловой денатурации с помощью спектрофотометра ВлСпекорд М-40В».
Меченые  препараты ДНК получали с помощью реакции НИК - трансляции ( Маниатис и соавт.,1985 ). Реакцию ДНК тАУ ДНК гибридизации проводили по методу Денхардта ( 1966 ). Степень подобия нуклеоидных последовательностей ДНК определяли, принимая за 100% радиоактивность гомологической реакции.
препараты ДНК получали с помощью реакции НИК - трансляции ( Маниатис и соавт.,1985 ). Реакцию ДНК тАУ ДНК гибридизации проводили по методу Денхардта ( 1966 ). Степень подобия нуклеоидных последовательностей ДНК определяли, принимая за 100% радиоактивность гомологической реакции.
При изучении антигенной структуры бактерий использовали кроличьи гипериммунные сыворотки на цельные микробные клетки. Пробирочную и пластинчатую РА ставили по Mittal K. и соавт. ( 1984 ),
при постановке РА с 2 - меркаптоэтанолом ( 2МЭ ) серийные разведения сыворотки делали в растворе с 0,1М -2МЭ. Эритроцитарные антигенные диагностикумы для РНГА готовили по методу Сидорова М.А. и Агаевой Э.М. Ставили РНГА и учитывали результаты общепринятым способом. Антительные диагностикумы для реакции коагглютинации готовили по Mittal K. и соавт. ( 1987 ). Реакцию иммунофлуоресценции в двух ступенчатом варианте проводили с использованием коммерческих меченых ФИТЦ, антикроличьих сывороток и люминесцентного микроскопа МЛ - 2.
Реакцию ставили в классическом варианте, при исследовании свиных сывороток крови тАУ по Nicolet J. (1971 ) с добавлением в комплемент сыворотки крови крупного рогатого скота.
При оценке вирулентности бактериальных культур определяли
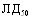 по Риду и Менну. Остаточную токсичность инактивированных вакцин определяли, используя тест прироста живой массы мышей. При определении специфической активности экспериментальных вакцин рассчитывали
по Риду и Менну. Остаточную токсичность инактивированных вакцин определяли, используя тест прироста живой массы мышей. При определении специфической активности экспериментальных вакцин рассчитывали 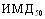 , индекс и коэффициент эффективности препарата ( Безденежных И.С., Леонтьева Л.Г.,1969 ).
, индекс и коэффициент эффективности препарата ( Безденежных И.С., Леонтьева Л.Г.,1969 ).
Цифровой материал обрабатывали, используя общепринятые методы математической статистики ( Ашмарин И.П., Воробьёв А.А., 1962; Терентьев П.В., Ростова Н.С.,1977 ).
2. Результаты исследований.
2.1. Свойства возбудителей, таксономическое положение и разработка лабораторной диагностики актинобациллёзной (гемофилёзной) плевропневмонии и гемофилёзного полисерозита свиней .
2.1.1. Питательные среды и культивирование A.pleuropneumoniae и H.parasuis.
Конструирование питательных сред для культивирования A.pleuropneumoniae и H.parasuis предполагает учёт потребности данных видов в НАД. Минимальный уровень зоны оптимума у обоих видов бактерий близок и составляет 1,8 - 3,6 НАД в 1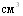 среды. Верхний пороговый уровень имеет видовые и штаммовые различия, причём он ниже у H.parasuis (15,62 тАУ 31,25 мкг/
среды. Верхний пороговый уровень имеет видовые и штаммовые различия, причём он ниже у H.parasuis (15,62 тАУ 31,25 мкг/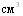 ), чем у A.pleuropneumoniae (250мкг/
), чем у A.pleuropneumoniae (250мкг/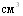 ) и, в свою очередь, он меньше в пределах вида H.parasuis у свежевыделенных эпизоотических штаммов, нежели у адаптированных музейных культур. Полученные данные позволили обоснованно конструировать питательные среды свиноводства учётом возможных штаммовых потребностей в НАД. Из естественных источников НАД наиболее доступен и эффективен дрожжевой экстракт. Переживающие животные ткани также содержат достаточное для роста V - зависимых бактерий количество НАД. Наиболее удобно в этом случае использовать кровяной сгусток на агаре Цинссера или сывороточном агаре.
) и, в свою очередь, он меньше в пределах вида H.parasuis у свежевыделенных эпизоотических штаммов, нежели у адаптированных музейных культур. Полученные данные позволили обоснованно конструировать питательные среды свиноводства учётом возможных штаммовых потребностей в НАД. Из естественных источников НАД наиболее доступен и эффективен дрожжевой экстракт. Переживающие животные ткани также содержат достаточное для роста V - зависимых бактерий количество НАД. Наиболее удобно в этом случае использовать кровяной сгусток на агаре Цинссера или сывороточном агаре.
Испытание различных видов питающих культур бактерий как продуцентов НАД показало, сто ростовой фактор экскретируют в достаточных количествах интенсивно растущие виды бактерий (эшерихии, сапрофитные бациллы, стафилококки), которые обеспечивают зону от 16,8 В± 0,89 мм до 31,6 В± 1,14мм в зависимости от вида бактерий и типа питательной среды.
Однако, при использовании культур стафилококков, установили, что некоторые из них ингибируют сателлитный рост A.pleuropneumoniae. Мы не обнаружили описание подобного феномена, хотя он известен у P.multjcida. Но в литературе есть сообщения о наличии гиалуроновой кислоты в составе капсулы некоторых штаммов A.pleuropneumoniae. Следовательно, причиной ингибиции роста может быть синтез культурой стафилококка гиалуронидазы. Мы также не исключаем вероятность образования продуцентом НАД соединений типа НАД-азы. В любом случае, питающая культура бактерий должна быть предварительно проверена по указанному критерию.
Определение НАД - зависимости тАУ важный этап в идентификации A.pleuropneumoniae (1-й биовар) и H.parasuis. Наши данные о том, что референтные и эпизоотические культуры A.pleuropneumoniae периодически могут утрачивать НАД- зависимость, причём это касается любого штамма 1-го биовара. Явление НАД -зависимости может быть восстановлено у культур, утративших это свойство, путём культивирования на ВлобеднённойВ» питательной среде с локальным источником НАД. Следовательно на обычных питательных средах животного происхождения культуры, потерявшие НАД зависимость, утилизируют какие-то предшественники НАД, отсутствующие, например, в глюкозо-казеиновом агаре. На последней питательной среде их рост возможен только в присутствии НАД, зависимость от которого они приобретают вновь.
Результаты исследований по изучению НАД - зависимости гемофильных бактерий позволили нам дать оценку и рекомендовать для практических целей сочетание питательных сред, позволяющих в условиях диагностической лаборатории тестировать V - и Х - зависимость.
Исследования показали отсутствие СО2 - зависимости у изученных культур H.parasuis и A.pleuropneumoniae, а также не удалось обнаружить существенных различий в частоте изоляции культур из патологического материала в условиях обычной атмосферы и повышенного содержания СО2 . Оба вида бактерий показали сильную зависимость от наличия в питательной среде сыворотки крови. У H.parasuis эта потребность выражена сильнее, чем у A.pleuropneumoniae. При отсутствии в питательном субстрате сыворотки крови оба вида бактерий быстро диссоциируют.
Количественные показатели, характеризующие рост A.pleuropneumoniae и H.parasuis в жидкой питательной среде (бульон Хоттингера, 120мг % аминного азота, рН 7,6,5,% сыворотки крови крупного рогатого скота, 10% дрожжевого экстракта), свидетельствуют о более интенсивном росте первого вида, в частности, период генерации соответственно составил 40,8 и 67 минут, удельная скорость роста 1,008 и 0,622, прирост бактериальных клеток за 1 час культивирования 0,44 и 0,27 log.
Оценка питательных сред для первичной изоляции культур H.parasuis и A.pleuropneumoniae (1-й биовар) показала, что для этой цели могут быть использованы агар и бульон Левинталя, ВлшоколадныйВ» агар, сывороточно-дрожжевой бульон и агар, сывороточный и кровяной агар, с локальными источниками V - ростового фактора. Предпочтительнее использование прозрачных питательных сред, позволяющих получать более полную информацию об особенностях колонии. Использование сред с локальными источниками НАД даёт важную информацию на первом этапе бактериологического исследования о НАД -зависимости, а в случае кровяного агара - также о гемолитической активности бактерий. Применение питающих бактерий как источника НАД требует отвивки культур в течении 24-48 часов из-за быстрой их гибели под влиянием метаболитов ВлбаккормилокВ». Подращивание материала в течении 6-8 часов в сывороточно - дрожжевом бульоне, содержащем бацитрацин, с последующим рассевом на среды увеличивает вероятность выделения культур НАД--зависимых бактерий.
Для поддержания культур H.parasuis и A.pleuropneumoniae при текущей работе в наибольшей степени подходят оптимальные плотные питательные среды с заливкой культур вазелиновым маслом при температуре хранения 5-8В°С. Наиболее длительно сохраняют жизнеспособность (6-7 недель) культуры, выращенные в сгустках крови и сохраняемые в замороженном состоянии.
2.1.2. Методы и результаты идентификации НАД--зависимых бактерий, выделенных от свиней, по культурально - морфологическим, ферментативным и генетическим свойствам.
Исследование ферментативных свойств НАД тАУ и гемин тАУ зависимых культур бактерий проводили на рутинных дифференциально-диагностических средах с добавлением ростовых факторов, а также на жидких средах Гисса в микрообъёмах и диагностических системах ПБДЭ и СИБ Горьковского НИЭМ. Максимальное совпадение результатов было получено на общепринятых средах, в ПБДЭ и микрообъёмах.
Таксономическое положение 165 культур НАД--зависимых бактерий, выделенных от клинически здоровых свиней, а также животных с явлениями фибриозно-геморрагической плевропневмонии или генерализованных серозитов, было исследовано при помощи нумерического анализа. В качестве опорных в данную коллекцию культур были включены типовые штаммы Haemophilus (Actinobacillus) pleuropneumoniae, H.parasuis, H.parainfluenzae, H.influenzae, A.ligieresii, A.suis. У каждого исследованного штамма были определены 57 фенотипических признаков, характеризующих их культуральные и ферментативные свойства. Все полученные данные были преобразованы в числовую форму, подвергнуты нумерическому анализу свиноводства конечным представлением полученных результатов в виде дендрограммы.
Анализ дендрограммы показал, что все культуры объединяются в единый массив при уровне сходства 75,02%, после чего они подразделяются на два больших фенона: один с уровнем сходства 81,87%, включающий типовые штаммы A.lignieresii, A.suis, Haemophilus, (Actinobacillus), pleuropneumoniae и типовой штамм Влмалой группыВ» (фенон ВлАВ»), второй с уровнем подобия 81,997% включает типовые штаммы H.influenzae, H.parainfluenzae и H.parasuis (фенон ВлНВ»). Из фенона ВлАВ», объединяющего культуры актинобацилл на уровне подобия 91,23%, выделяется фенон №1, включающий виды A.lignieresii и A.suis, и на уровне сходства 95,00% выделяется фенон №2, объединяющий эпизоотические культуры вокруг типового штамма Влмалой группыВ». При уровне подобия 89,48% формируется фенон №3, объединяющий эпизоотические штаммы вокруг типовых культур Haemophilus (Actinobacillus) pleuropneumoniae.
В феноне ВлНВ» на уровне сходства 81,997% выделяется фенон №4-
-H.influenzae, на уровне 87,63% - фенон №5, объединяющий штаммы H.parainfluenzae и на уровне подобия 89,60% вокруг типовых штаммов H.parasuis концентрируется другая группа эпизоотических штаммов -
-фенон №6.
Следовательно, массив эпизоотических штаммов дифференцируется на три фенотипические группы, соответствующие видам Haemophilus (Actinobacillus) pleuropneumoniae, H.parasuis и Влмалой группеВ» гемофильных бактерий. Полученные данные свидетельствуют о тяготении культур Haemophilus pleuropneumoniae не к роду Haemophilus, а к роду Actinobacillus.
С целью уточнения таксономического положения эпизоотических штаммов, классифицированных по фенотипическим свойствам как Haemophilus (Actinobacillus) pleuropneumoniae, НАД--зависимых бактерий, обозначаемых как Pasteurella-haemolytica- подобные бактерии, был определён нуклеотидный состав и уровень гомологии ДНК культур этих групп свиноводства представителями рода Haemophilus (H.parasuis, шт.J 95-таксон ВлСВ»), Pasteurella (P.multocida), Actinobacillus (A.suis), а также с типовыми культурами Haemophilus (Actinobacillus) pleuropneumoniae.
Результаты показали, что все исследованные штаммы по ГЦ -составу образуют довольно однородную группу свиноводства крайними значениями 39,8 - 43,2 (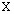 =41,3В±0,2). Такие значения вполне соответствуют приводимым в Руководстве Берги для семейства Pasteurellaceae. Несколько более высокий ГЦ состава обнаружен у P.haemolytica - подобных бактерий (42,5В±0,5) по сравнению с типовыми штаммами A.pleuropneumoniae, (40,7В±0,5). Эпизоотические штаммы Haemophilus (Actinobacillus) pleuropneumoniae имели показатель 41,2В±0,1 мол% ГЦ.
=41,3В±0,2). Такие значения вполне соответствуют приводимым в Руководстве Берги для семейства Pasteurellaceae. Несколько более высокий ГЦ состава обнаружен у P.haemolytica - подобных бактерий (42,5В±0,5) по сравнению с типовыми штаммами A.pleuropneumoniae, (40,7В±0,5). Эпизоотические штаммы Haemophilus (Actinobacillus) pleuropneumoniae имели показатель 41,2В±0,1 мол% ГЦ.
Уровень гомологии ДНК внутри группы P.haemolytica - подобных штаммов составил 100%; ДНК бактерий этой группы (шт.2288/77) имела гомологию с ДНК типовых штаммов A.pleuropneumoniae - 75%; с ДНК A.suis - 33%, с ДНК P.multocida и H.parasuis не более 9%; с ДНК эпизоотических штаммов Haemophilus (Actinobacillus) pleuropneumoniae - 75тАФ83%.
Эпизоотические штаммы Haemophilus (Actinobacillus) pleuropneumoniae по данным гибридизации ДНК образуют генетически взаимосвязанную группу с уровнем подобия нуклеотидных последовательностей тАУ 75-100%. В то же время бактерии этой группы тесно связаны с типовыми штаммами Haemophilus (Actinobacillus) pleuropneumoniae (78-98%), на уровне 33-38% гомологичных нуклеоидных последовательностей с ДНК A.suis , и только на уровне гомологии ДНК 5-9% с H.parasuis и 3-10% c P.multocida.
При интерпретации полученных результатов мы исходили из критериев генетического анализа, предложенных Медниковым и соавт.(1974), согласно которым культуры P.haemolytica - подобных бактерий не имеющие НАДтАФзависимости, могут рассматриваться как биовар Haemophilus (Actinobacillus) pleuropneumoniae. В целом эпизоотические штаммы, выделенные при фибринозно - геморрагических пневмониях, P.haemolytica - подобные бактерии и типовые штаммы Haemophilus (Actinobacillus) pleuropneumoniae предстают как видовая общность с заметной внутривидовой гетерогенностью. Подобный внутривидовой уровень гетерогенности не исключает выделения дополнительных внутривидовых таксонов, кроме НАДтАФзависимого и - независимого биоваров.
Полученные данные свидетельствуют, что культуры A.pleuropneumoniae, обозначаемые в 9-м издании Руководства по систематике бактерий Берги как видов рода Haemophilus, не относятся к нему, а также штамм J95 (таксон ВлСВ»), упоминаемый в числе видов рода Haemophilus, по данным гибридизации также не относится к роду Haemophilus.
На основании результатов нумерического анализа, с учётом частоты встречаемости тех или иных положительных ферментативных реакций, для идентификации НАД--зависимых бактерий, выделяемых при пневмониях и генерализованных серозитах свиней, рекомендованы в качестве основных дифференциальных признаков (кроме культуральных): НАДтАФзависимость, образование уреазы, щелочной фосфатазы, гемолизина, кислоты из ксилозы, маннита и маннозы. Остальные ферментативные признаки можно рассматривать как дополнительные.
Потеря культурами A.pleuropneumoniae НАДтАФзависимости и признание существования НАДтАФнезависимого биовара этого вида бактерий делает неприемлемой схему идентификации, предлагаемую для НАД--зависимых культур. Неизбежно расширяется число бактериальных видов, от которых необходимо дифференцировать НАД - независимые культуры A.pleuropneumoniae. Ситуация осложняется тем, что сходные патологоанатомические изменения в лёгких могут вызвать другие виды бактерий. В опытах экспериментального (интраназального) заражения свиней сходные изменения в легких были обнаружены при инокуляции культур A.pleuropneumoniae НАДтАФзависимого и - независимого биоваров, A.suis и бактерий Влмалой группыВ». Данное обстоятельство увеличивает значение бактериологического исследования в диагностике болезни. Сравнительное изучение физиологических свойств A.pleuropneumoniae (1 и 2 биовары), A.suis, P.multocida и B.bronchiseptica как видов сходных и достаточно часто обнаруживаемых при пневмониях свиней позволило нам отобрать для их дифференциации следующий минимальный набор признаков: липкость колоний и вязкость бульона, наличие капсулы и жгутиков, гемолитическая и уреазная активность, образование щелочной фосфатазы, индола, кислоты из маннита, сорбита, салицина, мелибиозы, утилизации цитратов.
- Патогенные свойства A.pleuropneumoniae.
Исследовали патогенность данного вида бактерий для лабораторных животных и свиней.
При испытаниях различных способов заражения морских свинок культурами A.pleuropneumoniae животные этого вида оказались наиболее чувствительны культур интраназальной инокуляции (ЛД50=125млн.м.к.), менее чувствительны тАУ к внутрибрюшинному (ЛД50=1,585 млрд.м.к.) и малочувствительны к подкожному заражению. Сравнительная оценка вирулентности девяти штаммов A.pleuropneumoniae при интраназальном способе инокуляции по критерию объёма поражённой лёгочной ткани показала наибольшую вирулентность культур 1,4,5 сероваров и существенно меньшую у культуры серовара 3; культуры серовара 2 показали штаммовые различия. Патологоанатомические изменения при интраназальном заражении морских свинок характеризовались развитием фиброзно-геморрагической пневмонии.
В отличие от морских свинок белые мыши оказались более чувствительны к внутрибрюшинному заражению: ЛД50 составила для культуры 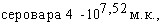
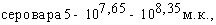
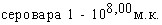 изменения в лёгких у заражённых мышей характеризовались развитием геморрагической пневмонии. Отличительной особенностью явилось практически полное отсутствие отложений фибрина на плевре и перикарде. В целом прослеживается видовые отличия изменений в лёгких при актинобациллёзной пневмонии: в воспалительном экссудате максимальное количество фибрина отмечается у свиней, меньше - у морских свинок и практически отсутствует у мышей.
изменения в лёгких у заражённых мышей характеризовались развитием геморрагической пневмонии. Отличительной особенностью явилось практически полное отсутствие отложений фибрина на плевре и перикарде. В целом прослеживается видовые отличия изменений в лёгких при актинобациллёзной пневмонии: в воспалительном экссудате максимальное количество фибрина отмечается у свиней, меньше - у морских свинок и практически отсутствует у мышей.
Полученные результаты свидетельствуют, что морские свинки и белые мыши могут быть использованы в качестве лабораторной модели при изучении вирулентности штаммов возбудителя, вопросов пато - и иммуногенеза.
Испытание различных способов заражения 2-2,5 - месячных свиней показало их чувствительность к интраназальному и трахеальному способам введения возбудителя при устойчивости к подкожной инокуляции культур. Заболевание удалось также воспроизвести контактным путем. При интраназальном и трахеальном заражении (доза 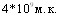 ) инкубационный период составил 3 - 6 часов, контактном - 72 часа. При всех способах заражения однотипные изменения в виде фибринозно - геморрагической пневмонии отмечали в лёгких. Это свидетельствует, что естественными входными воротами возбудителя являются дыхательные пути.
) инкубационный период составил 3 - 6 часов, контактном - 72 часа. При всех способах заражения однотипные изменения в виде фибринозно - геморрагической пневмонии отмечали в лёгких. Это свидетельствует, что естественными входными воротами возбудителя являются дыхательные пути.
При оценке патогенности для свиней культур возбудителя различных сероваров (сер. 1,2,3,4,5) установили, что колебания вирулентности больше отражают штаммовые различия или состояние культуры на момент исследования, чем серовариантную принадлежность. Только культура серовара 3 в опытах на свиньях и лабораторных животных закономерно показывала меньшую вирулентность, чем штаммы других сероваров. Титрация культуры эпизоотического штамма возбудителя на свиньях (интраназальное заражение) показала ЛД50 на уровне 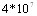 м.к.
м.к.
На результаты заражения свиней влияет возраст культуры бактерий. При интраназальном заражении свиней одинаковой дозой 6 - и 18 - часовых культур (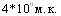 ) инкубационный период, соответственно, составил 3 и 7 часов, среднее время от заражения до гибели животных тАУ 48,5 и 103 часа, количества летальных исходов тАУ100% и 50%, объём поражённой лёгочной ткани - 70% и 47,5%, что свидетельствует о разной вирулентности культуры одного штамма на различных стадиях развития популяции.
) инкубационный период, соответственно, составил 3 и 7 часов, среднее время от заражения до гибели животных тАУ 48,5 и 103 часа, количества летальных исходов тАУ100% и 50%, объём поражённой лёгочной ткани - 70% и 47,5%, что свидетельствует о разной вирулентности культуры одного штамма на различных стадиях развития популяции.
Помимо свойств заражающей культуры возбудителя на течение и исход болезни влияют факторы внешней среды. В эксперименте показано значение факторов микроклимата. Двенадцать заражённых одной культурой свиней содержали в условиях микроклимата, соответствующего зоогигиеническим нормам, (группа А) и свиноводства отклонениями (группа Б). В группах А и Б соответственно всего пало 66,6% и 100% свиней, из них в течении 24 часов тАУ 33,3% и 66,6%. Среднее время от заражения до гибели в группах составило 67,5 и 27,3 часа. Объём поражённой лёгочной ткани - 32,2В±12,1% и 59,5В±5,9%. Различия по последнему показателю в группах А и Б статистически достоверны (р<0,01).
В опытах экспериментального воспроизведения актинобациллёзной пневмонии на морских свинках, свиньях, а также при исследовании животных, павших в условиях неблагополучного хозяйства, исследована диссеминация возбудителя.
При летальном исходе у свиней наиболее часто культуры возбудителя изолировали из поражённых участков лёгочной паренхимы (100%), бронхиальных и средостенных лимфатических узлов (63,33 - 100%), реже тАУ из печени, селезёнки, почек, костного мозга, крови, головного мозга. Полученные данные определяют характер материала, отбираемого для бактериологического исследования.
Исследования экспериментально заражённых морских свинок (интраназальная инокуляция) показало, что изменения в лёгких в виде очаговой гиперемии в зоне ветвлений крупных и средних бронхов развиваются уже через час после инокуляции культуры. Возбудитель на это стадии реизолировали из лёгочной ткани, бронхов, трахеи. Через три часа в лёгких развивались типичные изменения. На этой стадии в 25% случаев культуры возбудителя реизолировали из крови и паренхиматозных органов. По истечении 6 часов изменения в лёгких получили полное развитие. Таким образом, бактериемия и диссеминация возбудителя происходит на фоне уже имеющихся изменений в лёгких.
Наблюдение за экспериментально заражёнными больными свиньями в условиях неблагополучного хозяйства позволяют выделить три варианта течения болезни: сверхострое, острое и подострое. При сверхостром течении на фоне жёсткого респираторного синдрома летальный исход наступает в течении 6-24 часов. В грудной полости, где расположены поражённые доли лёгкого, обнаруживали 150-300см3 красноватой жидкости, изменённая доля лёгкого увеличена в объёме, ткань на разрезе тёмно-красного цвета, интерлобулярные септы отёчны, консистенция поражённой ткани плотная, с разреза стекает красноватая жидкость, бронхи и трахея заполнены аналогичной пенистой жидкостью. Отложения фибрина на пульмональной и костальной плевре отсутствуют.
У животных с острым течением болезни отмечен прогрессирующий респираторный синдром с повышением температуры до 40,6тАФ41В°C, завершающийся летальным исходом в течении 6тАФ7 суток. У павших свиней в грудной полости находили до 200 см3 красноватой жидкости с хлопьями фибрина. Поражённые лёгкие тёмно-красные, плотные, ломкие, с выраженным отёком междольковой соединительной ткани. Костальная и лёгочная плевра покрыты плёнками фибрина.
У свиней с подострым течением регистрировали симптомы пневмонии, лихорадку ремитирующего типа, плохую поедаемость корма. Поражённые доли лёгких увеличены, бугристые, плотные, неравномерно окрашены: участки тёмно-красного, серо-коричневого, грязно-бурого цвета. Через 15-20 дней в лёгочной ткани обнаруживали очаги уплотнения, окружённые соединительной тканью. Отложения фибрина на лёгочной и костальной плевре пронизаны соединительной тканью.
Гистологические изменения в лёгких на начальной стадии болезни могут быть охарактеризованы как бактериальный токсический шок (Гистологические исследования проведены д.в.н. Шубиным В.А.) Типичным для органопатологии бактериального шока считается парез и стаз артериол, мелких артерий, набухание и фибриноидный некроз стенок. Присоединение внутрисосудистого свёртывания крови придаёт шоку необратимый характер и приводит культур возникновению некрозов ткани. Дальнейшие изменения развиваются на указанном фоне. На 2-3 сутки начинается инфильтрация поражённых тканей мононуклеарными клетками, в основном лимфоцитами, при отсутствии или малом количестве нейтрофилов, эти клетки формируют своеобразный демаркационный вал. Позднее в поражённых тканях развиваются некротические очаги, подвергающиеся инкапсуляции. В зависимости от течения болезни и сроков гибели животного обнаруживается та или иная стадия патологического процесса.
Подтверждением вывода о ведущей роли токсинов в патогенезе актинобациллёзной плевропневмонии являются следующие результаты наших исследований. Установлено, сто в периодической аэрируемой культуре возбудитель в первые три часа культивирования синтезирует термолабильные, чувствительные к трипсину гемолизин и экзотоксин. Динамика их накопления и инактивации в культуральной жидкости, содержащей гемолизин и экзотоксин, вызывает у морских свинок в лёгких изменения типичные для актинобациллёзной пневмонии. По нашему мнению, 6-8- часовые агаровые и 3- часовые бульонные аэрируемые культуры в физиологическом отношении близки, поскольку находятся в начальной стадии логарифмического роста. Поэтому мы считаем, что на первом этапе развития изменений в лёгких определяющую роль играют гемолизин и экзотоксин. По нашему мнению, 6-8- часовые агаровые культуры возбудителя содержат не некоторое количество в связанном виде гемолизина и экзотоксина, что определяет большую вирулентность молодых культур.
- Патогенные свойства H.parasuis.
Исследовали чувствительность в возбудителю морских свинок, белых мышей, поросят-отъёмышей.
ЛД50 культуры H.parasuis штамм №1 для морских свинок при интраназальном заражении составила 354 млн.м.к., внутрибрюшинном - 594 млн.м.к., подкожном - 1,414 млд.м.к. При всех способах заражения с различной частотой обнаруживали развитие серозно-фибринозного плеврита, перитонита, реже перикардита. После интраназальной инокуляции у 100% животных отмечали развитие катаральной пневмонии. У павших животных культуры возбудителя наиболее часто выделяли из поражённых серозных оболочек и реже - из паренхиматозных органов.
ЛД50 культуры H.parasuis при внутрибрюшинном введении штамма серовара С составила 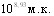 ,
, 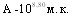 ,
, 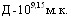 ,
, 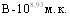 . Белые мыши, как и морские свинки были более чувствительны к внутрибрюшинному (ЛД50=1,414 млрд.м.к.) и интраназальному (ЛД50=1,549 млрд.м.к.) способам заражения при устойчивости к подкожному введению культуры. Патологоанатомические изменения у мышей и морских свинок были однотипны.
. Белые мыши, как и морские свинки были более чувствительны к внутрибрюшинному (ЛД50=1,414 млрд.м.к.) и интраназальному (ЛД50=1,549 млрд.м.к.) способам заражения при устойчивости к подкожному введению культуры. Патологоанатомические изменения у мышей и морских свинок были однотипны.
С нашей точки зрения, морских свинок можно рассматривать как наиболее удобную модель для биопробы и изучения пато - и иммуногенеза инфекции, вызываемой H.parasuis.
В опытах на поросятах-отъёмышах показано, что гемофилёзный полисерозит может быть воспроизведён при интраназальном, трахеальном, внутрибрюшинным и внутривенном введении культуры H.parasuis. Учитывая экологию возбудителя, можно констатировать, что естественными входными воротами инфекции являются дыхательные пути. При трахеальном заражении первые клинические симптомы в виде угнетения и повышения температуры тела отмечаются через 6-8 часов, позднее (20-24 часа) у абсолютного большинства животных развивается катаральная пневмония. В 75% случаев уже на этой стадии развивается серозно-фибринозный плеврит и в 50% случаев - перитонит, что свидетельствует о высоких инвазивных свойствах возбудителя. У животных, убитых через 24 часа после заражения, на фоне описанных патологоанатомических изменений культуры возбудителя были выделены в 100% случаев из лёгких и бронхиальных лимфатических узлов и в 25% случаев - из крови. В более поздние сроки существенных изменений в характере патологоанатомических изменений не наступало, за исключением резорбции экссудата из полостей и появления у отдельных животных симптомов поражения суставов конечностей и головного мозга. Гистологические исследования (выполнены д.в.н., профессором Шубиным В.А.) позволяют охарактеризовать изменения у экспериментально заражённых и естественно больных свиней как септико-токсический процесс с поражением серозных покровов, паренхиматозных органов, лимфатических узлов и центральной нервной системы.
Исследования показали, что типичные патоморфологические изменения могут вызвать как капсулообразующие, так и бескапсульные формы возбудителя при большей вирулентности первых.
По нашим данным изменения в лёгких, плевре, перикарде и перитонеуме ассоциируются с локализацией в них возбудителя. Частота изоляции культур H.parasuis из тканей и органов зависит от характера болезни и давности процесса. При остром течении в результате экспериментального (трахеального) заражения через 48-72 часа из лёгких культуры возбудителя выделили в 100% случаев, плевры - 75%, перитонеума и перикарда -25%, бронхиальных лимфатических узлов - 100%, селезёнки - 50%, крови - 25% случаев. При бактериологическом исследовании поросят, павших с картиной острого полисерозита, в естественных условиях культуры возбудителя выделили из лёгких у 42,85%, перикарда - 92,2%, плевры - 71,42%, печени - 57,14%, селезёнки - 66,66% животных. В более отдалённые сроки после экспериментального заражения и от животных, павших в естественных условиях с картиной подострого полисерозита, возбудитель выделяли реже и преимущественно из плевры, лёгких, перикарда, перитонеума и средостенных лимфоузлов.
Анализ возрастной заболеваемости поросят гемофилёзным полисерозитом показал, что пик отхода приходится на возраст 35-60 дней. Появление отхода поросят с явлениями гемофилёзного полисерозита совпадает со снижением титра колостральных антител к H.parasuis (серовар Д), увеличением уровня колонизации возбудителя в респираторном тракте. Согласно полученным данным, H.parasuis является достаточно обычным компонентом нормальной микрофлоры слизистых путей и на фоне описанных изменений, наряду с другими этиологическими агентами, участвует в возникновении пневмоний у поросят данной возрастной группы и ряде случаев это сопровождается развитием септической стадии с поражением серозных оболочек, суставов и головного мозга.
2.1.5.Обоснование критериев отбора материала для бактериологического исследования на актинобациллёзную пневмонию и гемофилёзный полисерозит.
Гемофилёзный полисерозит. Данные о диссеминации возбудителя свидетельствуют о наибольшей частоте его выделения из поражённых серозных оболочек, серозно-фибринозного экссудата. Выделение чистой культуры возбудителя наиболее вероятно при исследовании пунктата из поражённых суставов и перикарда. В замороженном патологическом материале H.parasuis сохраняет жизнеспособность до 30 суток (режим хранения -18В°С).
Актинобациллёзная плевропневмония. Независимо от характера течения болезни возбудитель закономерно обнаруживается в поражённой ткани лёгких и регионарных лимфатических узлах, что определяет характер материала, отбираемого для бактериологического исследования. В патологическом материале, заключённом в водно-глицериновую смесь ( температура хранения 4 --6В°С), возбудитель сохраняет жизнеспособность в течение 15 суток, в замороженном тканевом материале тАУ до 45 суток.
- Серологическая идентификация культур H.parasuis A.pleuropneumoniae.
- Методы и результаты серологической идентификации культур H.parasuis.
Использовали кроличьи гипериммунные сыворотки против типовых штаммов сероваров А, В, С, Д.
Дифференциация сероваров H.parasuis в пробирочной РА показала её малую специфичность. Более эффективной для указанной цели оказалась РСК с растворимыми антигенами.
Исследование в РСК с типовыми сыворотками 46 эпизоотических культур H.parasuis, выделенных при генерализованных серозитах, позволило 47,82% культур отнести к известным сероварам Бакоса. Штаммы серовара А составили 15,2%, В - 17,39%, С - 2,17%, Д-13,04%. В группе штаммов, выделенных из лёгких при пневмониях (18шт), культуры серовара А составили 16,6%, В - 11,1%, С - 5,56%, Д -33,3%, нетипируемые -33,3%. В числе культур, выделенных из носовой слизи клинически здоровых свиней (14 шт.), 35,7% отнесены к серовару А, 14,2% -- к серовару Д, не удалось типировать 50% штаммов.
В целом по всей группе из 78 штаммов H.parasuis было отнесено к серовару А - 19,23%, В - 12,82%, С - 2,56%, Д - 17,95%, нетипируемые культуры составили 47,44%.
Исследование нетипируемых культур в РСК свиноводства атисыворотками против изоляторов этой же категории показало наличие ещё трёх серологических вариантов и часть культур осталась за пределами этих групп. Полученные данные позволяют заключить, что необходимо расширение числа сероваров на базе схемы Бакоса, либо требуется создание новой схемы серовариантной дифференциации с новыми типовыми штаммами.
Анализ серотиповой структуры культур, выделенных в пределах одного хозяйства показал ассоциацию с генерализованными серозитами свиней H.parasuis различной серовариантной принадлежности.
2.1.6.2. Методы и результаты серологической идентификации культур A.pleuropneumoniae.
Для серовариантной дифференциации культур A.pleuropneumoniae испытывали РА пробирочную, РА с 2-МЭ, РА на стекле, РНГА, РНГА с 2-МЭ, РКА пробирочную на стекле, двухступенчатую РИФ. При внутривидовой дифференциации в различных серологических реакциях типовых культур A.pleuropneumoniae (серовары 1,2,3,4,5,6,7,8, 10,12) установлены перекрёстные реакции между сероварами 3,6 и 8, 2 и 3, 1 и 2, 4 и 7. Исследование двухсторонних антигенных связей в большинстве случаев позволяет определить серовариантную принадлежность культур. Из группы ускоренных серологических реакций для серологической идентификации наиболее перспективна РКА с растворимыми антигенами.
При серологической идентификации 82 эпизоотических НАД--зависимых культур бактерий, выделенных при фиброзно-геморрагических пневмониях свиней и классифицированных как A.pleuropneumoniae отнесены к серовару 2 -12,9%, 3 -14,53%, 4 -8,53%, 6 -3,65%, бактериям Влмалой группыВ» -7,31%. Большую группу штаммов серологически идентифицировать не удалось. Культуры последней группы имели антигенное родство с сероварами 3,6,8,10, но при исследовании в перекрёстной РА с адсорбцией были не идентичны им.
2.1.7. Результаты изучения широты распространения сероваров A.pleuropneumoniae на основании серологических исследований.
Была предпринята попытка дать оценку циркуляции различных сероваров A.pleuropneumoniae в свиноводческих хозяйствах одной из областей центральной России по результатам серологических исследований. В РНГА исследовали 639 сывороток крови свиней из десяти хозяйств. Установлено преобладание антител к сероварам 1,4,6,3,2,12. При бактериологическом исследовании в данном регионе культур сероваров 1 и 12 не выделяли. Причины такой ситуации требуют дальнейшего изучения. В сыворотках крови некоторых серопозитивных свиней удалось выявить нейтрализующие антитела к гемолизинам A.pleuropneumoniae, что свидетельствует о наличии определённого иммунитета к A.pleuropneumoniae.
2.1.8. Разработка и апробация ускоренных серологических методов обнаружения антигенов A.pleuropneumoniae в патологическом материале.
Для обнаружения антигенов A.pleuropneumoniae непосредственно в тканевом материале были испытаны реакции коагглютинации (РКА) и двухступенчатой иммунофлуоресценции (РИФ).
Опыты показали, что РКА может быть использована для видовой и серотиповой идентификации A.pleuropneumoniae. Серотиповую специфичность результатов РКА удаётся повысить путём уменьшения дозы сыворотки для сенсибилизации носителя и использования в реакции растворимых капсульных антигенов. Более активные видовые диагностикумы удалось сконструировать при использовании поливалентных сывороток, полученных от кроликов, иммунизированных не моноантигенами, а смесью антигенов A.pleuropneumoniae различных сероваров. Использование РКА как метода видовой идентификации потребовало выяснения межвидовых антигенных связей A.pleuropneumoniae. Изучали антигенное родство данного вида бактерий с A.suis, A.lignieresii, P.multocida (А,В,Д), H.parasuis (A,B,C,Д), B.bronchiseptica, S.cholerasuis, S.tiphimurium. Наиболее выраженное антигенное родство установлено с A.suis и A.lignieresii, наличие которых в исследуемом материале может обусловить ложноположительные результаты РКА.
Серовариантную дифференциацию типовых культур A.pleuropneumoniae проводили в РИФ с предельными разведениями антисывороток первой ступени, обеспечивающими свечение клеток свиноводства гомологичными реагентами на четыре креста. Обнаружили перекрёстные реакции сероваров 1 и 2, 2,3 и 6, 3,2,6 и 8, 4 и 7, 10 и 12. Таким образом, надёжной серовариантной дифференциации в ряде случаев добиться не удалось. Слабые перекрёстные реакции интенсивностью на два креста получены с культурами A.suis, A.lignieresii. Не отмечено перекрёстных реакций с P.multocida (А,В,Д), H.parasuis (А,В,С,Д), E.coll, S.cholerasuis и S.tiphimurium.
При изучении специфичности РКА как метода обнаружения антигенов возбудителя в органах и тканях исследовали супернатанты тканевых (лёгочных) гомогенатов клинически здоровых свиней, морских свинок и получили неспецифические замедленные положительные реакции. Для устранения неспецифической аглютинации стафилококковые антительные диагностикумы дополнительно обрабатывали нормальной кроличьей сывороткой, увеличивали концентрацию альбумина в диагностикуме, прогревали исследуемый материал при 100В°С (10,60 мин.) 120В°С (15 мин.). Эффективной оказалась термическая обработка материала. Необходимость прогревания исследуемого материала заставила рассмотреть влияние этого фактора на антигены возбудителя. Известно, что клетки A.pleuropneumoniae содержат специфические термостабильные и тАУ лабильные антигены (K.Mittal et al., 1987). Изучение этого вопроса показало, что по термостабильности антигенов культуры можно подразделить на группу термоустойчивых (сер.2,3,6,8,10) и термолабильных (сер.1,7, штамм 202). У культур последней группы при кипячении (60 мин.) снижалась активность антигенов в серологических реакциях, особенно антигенов серовара 1 (шт.ССМ5869). Прогревание антигенов A.pleuropneumoniae при 100В°С в течении 10 минут на серологической активности антигенов возбудителя, кроме серовара 1 не сказывается.
Возможность выявления антигенов возбудителя в тканевом материале (лёгкие), при помощи РКА и РИФ первоначально изучали на морских свинках, белых мышах и свиньях, заражённых культурами A.pleuropneumoniae, а также A.suis и гемофильными бактериями Влмалой группыВ».
Результаты исследований показали совпадение данных бактериологии РИФ и РКА. Слабые положительные реакции были получены с тканевым материалом, содержащим A.suis.
При исследовании лёгочной ткани от 78 свиней с явлениями пневмонии из 29 материалов были выделены микоплазмы, из 15 -стафилококки или стрептококки, изменений 16 - P.multocida, из 2 - P.multocida и гемофильные бактерии Влмалой группыВ», из 7 - гемофильные бактерии Влмалой группыВ», из 9 - S.cholerasuis. При наличии в патологическом материале P.multocida в двух случаях были получены позитивные результаты РКА с диагностикумом против штамма №202 (Влмалая группаВ»), не подтверждённые данными бактериологии. В остальных случаях при исследовании материалов, содержащих указанные выше гетерологичные виды бактерий результаты РКА и РИФ были отрицательными. В девяти случаях изоляции культур Влмалой группыВ» данные бактериологических исследований, РКА и РИФ совпали.
Полученные данные позволяют оценить РКА и РИФ как достаточно специфические и чувствительные методы обнаружения антигенов A.pleuropneumoniae. Преимуществом РКА является возможность исследования длительно хранившегося материала и выявления растворимых антигенов.
2.2. Разработка и испытание экспериментальных образцов инактивированной вакцины против актинобациллёзной плевропневмонии свиней.
Возрастная заболеваемость свиней актинобациллёзной плевропневмонией диктует необходимость вакцинации просят-отъёмышей и проблема реактогенности препарата актуальна. Исследование остаточной токсичности инактивированных цельноклеточных суспензий A.pleuropneumoniae в тесте прироста массы мышей показало высокие её уровни. Из испытанных способов инактивации наиболее эффективным оказалась обработка формальдегидом (0,2%) с последующей выдержкой бактериальных взвесей при 4-6В°С в течении 30-45 суток. Различия сравниваемых показателей по этому препарату, а также инактивированных 0,1% формальдегида и тиомерсалом были достоверны, на уровне значимости от Р<0,01 тАУ до Р<0,05 (в зависимости от сроков хранения). Полученные данные были учтены в технологии изготовления инактивированной эмульгированной вакцины. Испытание вакцины такого типа на 21,38 тАУ 40 - дневных поросятах и супоросных свиноматках показало её безвредность.
В качестве лабораторных моделей для испытания специфической активности инактивированной вакцины были испытаны морские свинки живой массой 200тАФ250г и белые мыши живой массой 14-16г. Мышей вакцинировали подкожно, двукратно с интервалом 10 суток в дозе 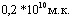 с последующим определением ЛД50 заражающей культуры на вакцинированных и контрольных животных (Влвнутрибрюшинная модельВ»). ЛД50 культуры, установленная на вакцинированных животных пятикратно, превышала тот же показатель, полученный на контрольных животных.
с последующим определением ЛД50 заражающей культуры на вакцинированных и контрольных животных (Влвнутрибрюшинная модельВ»). ЛД50 культуры, установленная на вакцинированных животных пятикратно, превышала тот же показатель, полученный на контрольных животных.
Морских свинок акционировали однократно различными дозами вакцины с последующим интраназальном заражении гомологичной культурой в дозе 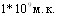 (Вллёгочная модельВ»). Подобный методический подход позволил определить ИМД50 вакцины. По нашему мнению, Вллёгочная модельВ» предпочтительнее, поскольку этот метод соответствует естественным входным воротам возбудителя инфекции. С другой стороны, об эффективности иммунизации можно дополнительно судить по наличию поражений в лёгких их массивности. Определённую информацию дают показатели серологического ответа - между объёмом поражённой лёгочной ткани и среднегрупповыми титрами антител (РСК) установлена функциональная обратная зависимость
(Вллёгочная модельВ»). Подобный методический подход позволил определить ИМД50 вакцины. По нашему мнению, Вллёгочная модельВ» предпочтительнее, поскольку этот метод соответствует естественным входным воротам возбудителя инфекции. С другой стороны, об эффективности иммунизации можно дополнительно судить по наличию поражений в лёгких их массивности. Определённую информацию дают показатели серологического ответа - между объёмом поражённой лёгочной ткани и среднегрупповыми титрами антител (РСК) установлена функциональная обратная зависимость 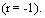
Исследовали влияние типа адъюванта на иммунологическую эффективность инактивированной вакцины. В качестве адъювантов использовали гидрат окиси алюминия и масляный адъювант на основе лёгкого минерального масла. Эмульгированной вакциной привили 315 поросят (полсектора) в неблагополучном по актинобациллёзной плевропневмонии хозяйстве, 317 свиней этого же сектора служили контролем. Гидроокись -алюминиевую вакцину ввели 647 поросятам, контролем служили 652 животных. Наблюдение за животными вели в течении 86 дней, заболевание свиней актинобациллёзной плевропневмонией наблюдали во всех группах. Индекс и коэффициент эффективности эмульгированной вакцины в этом опыте составил соответственно 7,95 и 8,42%. Аналогичные показатели в группе свиней привитых РОА вакциной были соответственно 3,12 и 67,98%, что свидетельствует о большей эффективности эмульгированной вакцины.
При конструировании инактивированной вакцины, исходя из полученных данных о накоплении в культуральной жидкости экзотоксина, гемолизина и капсульной субстанции, исследовали влияние растворимых антигенов бульонных культур на её иммуногенность. В опыте на белых была определена ИМД50 эмульгированной вакцины приготовленной на основе бульонных культур и отмытых клетках возбудителя. ИМД50 вакцины первого типа составила 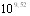 микробных клеток, у вакцины второго типа ИМД50 не удалось рассчитать, так как процент защиты при избранных дозах вакцины не превысил 36,36%. Следовательно, включение растворимых компонентов реакторных культур возбудителя в состав инактивированных вакцин необходимо.
микробных клеток, у вакцины второго типа ИМД50 не удалось рассчитать, так как процент защиты при избранных дозах вакцины не превысил 36,36%. Следовательно, включение растворимых компонентов реакторных культур возбудителя в состав инактивированных вакцин необходимо.
При отработке оптимальной схемы иммунизации свиней инактивированной эмульгированной вакциной исследовали влияние кратности её введения на уровень иммунитета у свиней. В одном случае свиньям (8 голов) вводили 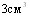 вакцины однократно, в другом случае тАУ дробно 1 и2
вакцины однократно, в другом случае тАУ дробно 1 и2 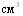 интервалом 10 суток (8 голов). Свиней заразили через 20 дней после вакцинации интраназально в дозе
интервалом 10 суток (8 голов). Свиней заразили через 20 дней после вакцинации интраназально в дозе 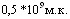 Установлено, что среди свиней, вакцинированных двукратно, летальных исходов не было. Из числа привитых однократно погибло 50% животных. Между животными этих групп также выявлена достоверная разница по высоте титров антител (РА). Исходя изменений полученных данных можно заключить, что результаты серологических реакций могут служить косвенным групповым показателем формирования иммунитета.
Установлено, что среди свиней, вакцинированных двукратно, летальных исходов не было. Из числа привитых однократно погибло 50% животных. Между животными этих групп также выявлена достоверная разница по высоте титров антител (РА). Исходя изменений полученных данных можно заключить, что результаты серологических реакций могут служить косвенным групповым показателем формирования иммунитета.
Исследовали влияние суммарной дозы антигена при двукратной (интервал - 15 суток) вакцинации свиней эмульгированной вакциной на состояние иммунитета. Установлено, что среди привитых различными дозами вакцины свиней (16 голов) при контрольном интраназальном заражении 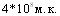 летальных исходов не было, в контроле погибло 50% животных. Среди животных, привитых вакциной в суммарных дозах 10,6,4 и 2
летальных исходов не было, в контроле погибло 50% животных. Среди животных, привитых вакциной в суммарных дозах 10,6,4 и 2 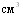 , объём поражённой лёгочной ткани в среднем по группам соответственно составил 5,25В±6,70, 5,94В±8,01, 5,69В±10,24, 10,50ВнВ±8,57%. У контрольных свиней было поражено 25,1% лёгочной ткани. Различия в титрах комплементсвязывающих антител у вакцинированных животных перечисленных групп перед заражением были статистически не достоверны, также не достоверны различия по объёму поражённой лёгочной ткани. Свиноводства учётом заболеваемости, количества летальных исходов и объёма поражённой лёгочной ткани мы считаем оптимальной дозу вакцины 4
, объём поражённой лёгочной ткани в среднем по группам соответственно составил 5,25В±6,70, 5,94В±8,01, 5,69В±10,24, 10,50ВнВ±8,57%. У контрольных свиней было поражено 25,1% лёгочной ткани. Различия в титрах комплементсвязывающих антител у вакцинированных животных перечисленных групп перед заражением были статистически не достоверны, также не достоверны различия по объёму поражённой лёгочной ткани. Свиноводства учётом заболеваемости, количества летальных исходов и объёма поражённой лёгочной ткани мы считаем оптимальной дозу вакцины 4 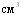
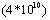 , введённую по схеме 1+3
, введённую по схеме 1+3 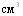 .
.
Исследовали сроки формирования и длительность иммунитета у свиней, привитых однократных эмульгированной вакциной в дозе 5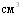 с контрольным интраназальным заражением через 10, 30 и 60 дней после иммунизации. Исходя из уровня защиты вакцинированных животных указанных групп и площади поражённой лёгочной ткани, близкие показатели состояния иммунитета имели свиньи через 10 и 30 дней со снижением к 60 дню после вакцинации. Между показателями защиты (%) и площади поражения лёгочной ткани выявлена обратная функциональная зависимость (r= -1). Значительная заболеваемость свиней актинобациллёзной плевропневмонией в первый период откорма в неблагополучном хозяйстве, с учётом полученных данных, свидетельствует о необходимости ревакцинации свиней в период передачи на откорм.
с контрольным интраназальным заражением через 10, 30 и 60 дней после иммунизации. Исходя из уровня защиты вакцинированных животных указанных групп и площади поражённой лёгочной ткани, близкие показатели состояния иммунитета имели свиньи через 10 и 30 дней со снижением к 60 дню после вакцинации. Между показателями защиты (%) и площади поражения лёгочной ткани выявлена обратная функциональная зависимость (r= -1). Значительная заболеваемость свиней актинобациллёзной плевропневмонией в первый период откорма в неблагополучном хозяйстве, с учётом полученных данных, свидетельствует о необходимости ревакцинации свиней в период передачи на откорм.
Определение иммуногенности вакцины в опытах на белых мышах в процессе хранения при 4-6В°С показало, что в интервале 1-11 месяцев ИМД50 препарата колебалась в пределах 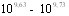 микробных клеток, что позволяет говорить о малом изменении активности препарата в указанные сроки.
микробных клеток, что позволяет говорить о малом изменении активности препарата в указанные сроки.
С учётом результатов испытания различных типов инактивированных вакцин на лабораторных животных и свиньях была проведена апробация экспериментальных образцов вакцин против актинобациллёзной плевропневмонии свиней в производственных условиях.
Первоначально эмульгированная вакцина была испытана на безвредность и иммунологическую эффективность на ограниченном контингенте свиней различного возраста в хозяйстве, неблагополучном по актинобациллёзной плевропневмонии свиней. В последующем в этом же хозяйстве проводилась поголовная вакцинация животных по следующей схеме: внутримышечно свиноматкам за 20-25 дней до ожидаемого опороса в дозе 1 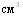 , аналогичную дозу вводили ремонтным свинкам, поросят вакцинировали в дозе 1
, аналогичную дозу вводили ремонтным свинкам, поросят вакцинировали в дозе 1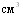 в возрасте 35 - 40 дне й. В период проведённых исследований вакцинации было подвергнуто 1677,4 тысячи свиней.
в возрасте 35 - 40 дне й. В период проведённых исследований вакцинации было подвергнуто 1677,4 тысячи свиней.
Исследования показали, что до начала вакцинации падёж свиней с диагнозом актинобациллёзной плевропневмонии составил 30,22% от общего количества павших животных, при частичной вакцинации поголовья -12,28%, при полной вакцинации - 8,89%. Между уровнем вакцинации и падёжом свиней от актинобациллёзной плевропневмонии установлена достаточно сильная обратная связь ( r= -0,71). Применение вакцины обеспечило снижение отхода свиней по причине актинобациллёзной плевропневмонии, в зависимости от анализируемого периода в 3 - 5 раз и увеличение живой массы поросят на 7,42 - 8,29 кг при передаче на откорм. Частота терапевтического вмешательства ветеринарного персонала в группах вакцинированных поросят была в 2,11 раза реже, чем среди не вакцинированных животных.
Выводы
- Исследование таксономического положения эпизоотических штаммов НАД--зависимых бактерий, выделенных при пневмониях и генерализованных серозитах свиней методом нумерического анализа, позволило дифференцировать их по фенотипическим свойствам на кластеры, соответствующие видам Actinobacillus (Haemophilus) pleuropneumoniae, Haemophilus parasuis и таксону гемофильных бактерий Влмалая группаВ». Культуры кластера Actinobacillus (Haemophilus) pleuropneumoniae имеют большее фенотипическое сходство с видами рода Actinobacillus (91,23) и меньшее с представителями рода Haemophilus (81,99%). На уровне подобия 95% бактерий кластера Влмалая группаВ» выделяются в самостоятельный таксон.
- Генетический анализ таксономического положения референтных и эпизоотических НАД--зависимых штаммов Actinobacillus (Haemophilus) pleuropneumoniae и культур P.haemolytica - подобных бактерий (2-ой биовар A.pleuropneumoniae) показал, что они образуют генетическую общность с уровнем гомологии ДНК 70-98%. ДНК культур 2-го биовара и эпизоотических штаммов A.pleuropneumoniae имеют гомологию с ДНК A.suis тАУ 33 - 38%, с ДНК H.parasuis и P.multocida тАУ не более 9%, что свидетельствует о принадлежности эпизоотических и референтных штаммов к роду Actinobacillus, но не Haemophilus и Pasteurella. Референтный штамм J 95 гемофильных бактерий таксона ВлСВ» по результатам гибридизации ДНК к роду Haemophilus не относится.
- Результаты изучения таксономического положения НАД--зависимых бактерий, выделенных при пневмониях и серозитах свиней, позволили определить в качестве основных дифференцирующих фенотипических признаков: НАДтАФзависимость, образование щелочной фосфатозы, уреазы, гемолизина, кислоты из ксилозы, маннита, маннозы. Для дифференциации культур НАДтАФнезависимого биовара A.pleuropneumoniae от сходных бактерий минимальный набор определяемых признаков включает продукцию индола, уреазы, утилизацию цитратов, наличие жгутиков. Для установления ферментативных свойств НАД--зависимых бактерий можно использовать дифференциально - диагностические среды в микрообъёмах и диагностический набор ПБДЭ Горьковского НИЭМ, что исключает использование чистого НАД или других его источников как факторов роста.
- К A.pleuropneumoniae и H.parasuis чувствительны при внутрибрюшинном и интраназальном заражении морские свинки (живая масса 200-250г) и белые мыши (живая масса 14-16г), которые могут быть использованы для определения патогенных свойств культур, изучения вопросов пато - и иммуногенеза при этих инфекциях.
- Входными воротами возбудителя инфекции при гемофилёзном полисерозите являются органы дыхания. При экспериментальном (трахеальном) заражении свиней H.parasuis инкубационный период составляет 6-8 часов. У заражённых свиней развивается катаральная пневмония и, в результате диссеминации возбудителя, генерализованные серозиты с одновременным поражением серозных оболочек грудной, брюшной полостей, пери тАУ и эпикарда.
- В естественных условиях H.parasuis вызывает у поросят-отъёмышей пневмонию, у части животных развивается септическая стадия с поражением серозных оболочек, суставов, головного мозга. У отдельных поросят встречается классическая форма болезни Глессера - генерализованные серозиты без микроскопически выраженной пневмонии. У экспериментально заражённых и естественно больных свиней наиболее часто удаётся изолировать H.parasuis из поражённых серозных оболочек.
- Входными воротами возбудителя инфекции при актинобациллёзной плевропневмонии свиней являются органы дыхания. Инкубационный период при экспериментальном интраназальном заражении составляет 2-4 часа, контактном тАУ до 3 суток, в естественных условиях тАУ 2-8 суток. Диссеминация возбудителя в организме заражённых животных происходит на фоне имеющихся изменений в лёгких. Наиболее постоянно культуры возбудителя удаётся изолировать от экспериментально заражённых и естественно больных животных из поражённой ткани лёгких и регионарных лимфатических узлов.
- Культуры A.pleuropneumoniae на начальной стадии логарифмического роста синтезируют термолабильные гемолизин и экзотоксин, которые определяют повышенную вирулентность молодых (6-8- часовых) культур возбудителя и играют существенную роль в развитии патологических изменений в лёгких заражённых животных.
- Макроскопически сходные изменения в лёгких свиней могут вызвать A.pleuropneumoniae, гемофильные бактерии Влмалой группыВ» и A.suis , поэтому решающее значение в диагностике актинобациллёзной плевропневмонии имеют результаты бактериологического исследования.
- При серологической идентификации эпизоотических культур H.parasuis установлено наличие сероваров А,В,С,Д, Бакоса, а также выявлены четыре серологические группы штаммов, не укладывающиеся в известную схему Бакоса, которая не охватывает всего антигенного многообразия вида.
- Серологическая идентификация эпизоотических культур A.pleuropneumoniae, выделенных от свиней в хозяйствах Российской Федерации, выявила наличие сероваров 2,3,4,6 и группы нетипируемых штаммов. Исследования в РНГА сывороток крови свиней из различных хозяйств показало доминирование антител к сероварам 1,2,4,6,12.
- На основании проведённых исследований разработана и используется в практике схема лабораторной диагностики актинобациллёзная ( гемофилёзная ) плевропневмонии и гемофилёзного полисерозита свиней, включающая: правила взятия материала для бактериологического исследования, методы выделения и идентификации культур возбудителя по биологическим свойствам. Также разработаны и рекомендованы методы видовой и серовариантной идентификации культур A.pleuropneumoniae и обнаружения антигенов этого вида бактерий в патологическом материале при помощи реакции коагглютинации.
- Разработана и предложена технология изготовления, контроля использования вакцины против гемофилёзной (актинобациллёзная) плевропневмонии свиней, что нашло отражение в соответствующих нормативных документах: ВлИнструкция по изготовлению и контролю вакцины против гемофилёзной плевропневмонии свинейВ», ВлВакцина против гемофилёзной плевропневмонии свиней эмульгированная. Технические условия ТУ 10-09-53-90В». ВлНаставление по применению вакцины против гемофилёзной плевропневмонии свиней эмульгированнойВ», утверждённых ГУВ ГК Совмина СССР по продовольствию и закупкам 11.07.1990г.
- Инактивированные вакцины из клеток A.pleuropneumoniae обладают значительной остаточной токсичностью. Эффективное снижение токсичности готовых препаратов достигается инактивацией бактериальных суспензий 0,2% формальдегида с последующим выдерживанием до употребления при 4-6В°С в течение 45 суток.
- В качестве лабораторных моделей при оценке иммуногенности инактивированных актинобациллёзных вакцин могут быть использованы белые мыши тАУ подкожная двукратная иммунизация с последующим внутрибрюшинном заражением различными дозами вирулентной культуры, или морские свинки в варианте подкожной вакцинации с последующим интраназальном заражением.
- Максимальной иммуногенностью для свиней обладают инактивированные эмульгированные вакцины из клеток A.pleuropneumoniae с включением в их состав растворимых антигенов, накапливающихся в реакторных культурах в процессе роста.
- В опытах прямого заражения при испытании эффективности иммунизации в условиях неблагополучного хозяйства установлено, что разработанная инактивированная вакцина не защищает полностью иммунизированных свиней от заражения, но обеспечивает более лёгкое течение болезни, в частности снижается отход в 3-5 раз, частота терапевтического вмешательства снижается в 2,11 раза, увеличивается живая масса поросят при передаче на откорм на 7,42 тАУ 8,29кг, что позволяет рассматривать вакцинацию как экономически целесообразный составной компонент комплекса лечебно-профилактических мероприятий по борьбе с актинобациллёзной плевропневмонией свиней.
Практические предложения.
- Временные методические указания по лабораторной диагностике гемофилёзного полисерозита поросят. Рекомендованы МСХ СССР 17.01.1978г. № 116-18.
- Временные методические указания по лабораторной диагностике гемофилёзной плевропневмонии свиней. Утверждены ГУВ МСХ СССР 16.04.1981г.
- Методические указания по диагностике гемофилёзов свиней. Утверждены ГУВ МСХ СССР 2.11.1985г. № 115-6а.
- Методические указания на изготовление и применение коагглютинирующего антительного диагностикума для экспресс тАУ идентификации Гемофилюс плевропневмоние и индикации возбудителя в патологическом материале. Утверждены отделением ветеринарии РАСХН 16.02.1993г.
- Временная инструкция о мероприятиях по борьбе с гемофилёзами свиней. Утверждена ГУВ МСХ СССР 2.11.1985г. № 115--6а.
- Инструкция по изготовлению и контролю вакцины против гемофилёзной плевропневмонии свиней. Утверждена ГУВ МСХ СССР 11.07.1990г.
- Вакцина против гемофилёзной плевропневмонии свиней эмульгированная. Технические условия ТУ 10-09-53-90. Утверждены ГУВ МСХ СССР 11.07.1990г.
- Наставление по применению вакцины против гемофилёзной плевропневмонии свиней. Утверждено ГУВ МСХ СССР 11.07.1990г.
Список опубликованных работ.
- Сидоров М.А., Скородумов Д.И., Тарасенок Н.И. и др. -
Инфекционный полисерозит поросят // Свиноводство, 1976 №11,35-36.
- Сидоров М.А., Скородумов Д.И., Гумбатов Ю.К. - Патогенность возбудителя гемофилёзного полисерозита для лабораторных животных и поросят // Труды ВИЭВ, 1977, Т.45,50-54.
- Скородумов Д.И., Гумбатов Ю.К. - Выделение возбудителя гемофилёзного полисерозита и изучение некоторых его свойств // Бюл. ВИЭВ, 1977, вып.21,14-17.
- Сидоров М.А., Скородумов Д.И., Гумбатов Ю.К. - Роль бактерий рода Гемофилюс в инфекционной патологии животных // Ветеринария, 1978, №5,102-108.
- Временные методические указания по лабораторной диагностике гемофилёзного полисерозита свиней (ГУВ МСХ СССР. 17.10.1978) / Сидоров М.А., Скородумов Д.И.
- Шубин В.А., Сидоров М.А., Андрыишин.., Скородумов Д.И. - Патоморфологические изменения у больных полисерозитом поросят // Труды ВИЭВ, 1978, т.47,135-141.
- Сидоров М.А., Скородумов Д.И., Шубин В.А., Тарасенок Н.И., Гумбатов Ю.М. - Экспериментальный гемофилёзный полисерозит поросят // Труды ВИЭВ, 1979, т.49,90-95.
- Сидоров М.А., Скородумов Д.И., Тарасенок Н.И., Гумбатов Ю.К. - Иммуногенность вакцины против гемофилёзного полисерозита поросят // Доклады ВАСХНИЛ, 1980, №8,30.
- Сидоров М.А., Скородумов Д.И., Гумбатов Ю.К., Тарасенок Н.И. - Серологические варианты гемофильных бактерий выделенных от свиней // Ветеринария, 1980, №4,66-67.
- Скородумов Д.И., Сидоров М.А. - Характеристика гемолитических гемофильных бактерий выделенных от больных пневмонией свиней // Труды ВИЭВ, 1980, т.52.,13-18.
- Шубин В.А., Сидоров М.А., Скородумов Д.И. - Динамика изменений в лёгких поросят при экспериментальном инфекционном полисерозите // Сборник ВлПатоморфология, патогенез и диагностика болезней с\х животныхВ». Научные труды ВАСХНИЛ, Москва,1980, 128-131.
- Сидоров М.А. Скородумов Д.И. - Свойства Гемофилюс плевропневмоние выделенных от больных свиней // Ветеринария, 1981, №1,45-47.
- Сидоров М.А., Скородумов Д.И., Лаврентьев Н.И., Мицаев Ш.Ш., Романова Л.Я., Юдин А.И. - Диагностика гемофилёзной плевропневмонии свиней // Ветеринария, 1981, №12, 66-68.
- Временные методические указания по лабораторной диагностике гемофилёзной плевропневмонии свиней (ГУВ МСХ СССР.1981 / Сидоров М.А., Скородумов Д.И.).
- Сидоров М.А., Скородумов Д.И., Панов В.С., Гушин В.Н., Седов В.А. - Вакцина против гемофилёзной плевропневмонии свиней, способ её изготовления и способ профилактики гемофилёзной плевропневмонии свиней. Авт. свидетельство на изобретение № 907899 от 21.10.81г.
- Сидоров М.А., Скородумов Д.И., Тарасенок Н.И., Шубин В.А. - Способ приготовления вакцины против гемофилёзного полисерозита поросят. Авт. свидетельство на изобретение № 957472 от 07.05.1982г.
- Сидоров М.А., Скородумов Д.И., Гущин В.А. - Не допускайте заболевания свиней гемофилёзами // Газета - плакат МСХ СССР, Москва, Колос, 1982, № 7.
- Сидоров М.А., Скородумов Д.И., Лаврентьев Н.И. - Специфическая профилактика гемофилёзной плевропневмонии свиней // Сборник ВлВетеринарные проблемы в промышленном свиноводствеВ», Киев, 1983, 97-98.
- Лаврентьев Н.И., Скородумов Д.И., Романова Л.Я., Финенко Р.Ф. - Применение РА в диагностике гемофилёзной плевропневмонии свиней // Сборник научных трудов ВлИнфекционные болезни с\х животныхВ», Омск, 1983.
- Скородумов Д.И., Гумбатов Ю.К., - Потребность бактерий рода Гемофилюс в ростовых факторах. // Бюллетень ВИЭВ, 1983, вып.45, 6-9.
- Сидоров М.А., Скородумов Д.И., Лаврентьев Н.И. - Гемофилёзная плевропневмонии свиней // Свиноводство, 1984, №4,14-15.
- Временная инструкция о мероприятиях по борьбе с заболеваниями свиней гемофилёзами (ГУВ МСХ СССР 2.11.1985) / Сидоров М.А.
- Методические указания по диагностике гемофилёзов свиней (ГУВ МСХ СССР 2.11.1985) / Сидоров М.А., Скородумов Д.И.
- Сидоров М.А., Скородумов Д.И., Логинов И.А., Лаврентьев Н.И. - Профилактика гемофилёзной плевропневмонии свиней // Ветеринария, 1985, № 12,37-38.
- Шубин В.А., Татришвили И.П., Сидоров М.А., Скородумов Д.И. - Патоморфологические изменения при гемофилёзной плевропневмонии поросят // Ветеринария, 1985, № 11, 40-43.
- Сидоров М.А., Скородумов Д.И. - Гемофилёзы животных. Агропромиздат.М.1986.
- Сидоров М.А., Мицаев Ш.Ш., Скородумов Д.И. - Патогенность H.pleuropneumoniae для животных // Ветеринария, 1989, № 11, 39-41.
- Вакцина против гемофилёзной плевропневмонии свиней эмульгированная. Технические условия ТУ 10-09059-90 (ГУВ МСХ СССР 11.07.1990) / Сидоров М.А., Скородумов Д.И., Логинов И.А., Субботин В.В., Мохина Т.Н.
- Инструкция по изготовлению и контролю вакцины против гемофилёзной плевропневмонии свиней эмульгированной (ГУВ МСХ СССР 11.07.1990) / Сидоров М.А., Скородумов Д.И., Логинов И.А., Субботин В.В.
- Наставление по применению вакцины против гемофилёзной плевропневмонии свиней эмульгированной (ГУВ МСХ СССР, 11.07.1990) / Сидоров М.А. Скородумов Д.И., Логинов И.А. Субботин В.В.
- Сидоров М.А., Скородумов Д.И., Столяренко В.Т., Субботин В,В. - Специфическая профилактика гемофилёзного полисерозита поросят // ВАСХНИЛ, НТЦ. Научно-технической информационный сборник. Москва,1991, вып.9.
- Сидоров М.А., Лаврентьев Н.И., Субботин В.В., Логинов И.А., Скородумов Д.И. - Эффективность вакцинации против гемофилёзной плевропневмонии свиней // Ветеринария, 1991, № 4,25-27.
- Скородумов Д.И., Бурлаков В.А., Костенко Т.С. - Таксономия бактерий семейства Pasteurellaceae // Методические указания. Москва, 1991.
- Скородумов Д.И., Сидоров М.А., Пруссак-Глотов В.Э. - Возбудители фибринозно-геморрагической плевропневмонии свиней // Ветеринария, 1992, №6,21-24.
- Фомин Б.А., Коромыслов Г.Ф., Артюшин С.К., Скородумов Д.И., Сидоров М.А. - Гомология ДНК H.pleuropneumoniae и некоторых видов бактерий семейства Pasteurellaceae // Ветеринария, 1992, № 6,28-30.
- Скородумов Д.И., Сидоров М.А., Блему Ж. - Методические указания на изготовление и применение коагглютинирующего диагностикума для экспресс - идентификации Гемофилюс плевропневмонии и индикации возбудителя в патологическом материале // Отделение ветеринарии РАСХН. 13.02.1993.
- Скородумов Д.И. - Свойства гемолизинов возбудителя актинобациллёзной плевропневмонии свиней // Сборник научных трудов ВлАктуальные вопросы инфекционных и инвазионных болезней животныхВ». Москва, 1995, 50-52.
- Сидоров М.А., Скородумов Д.И., Федотов В.Б. - Определитель зоопатогенных бактерий (справочная книга) М., Колос, 1995.
- Сидоров М.А., Скородумов Д.И., Блему Ж. - Применение реакции коагглютинации для идентификации возбудителя актинобациллёзной плевропневмонии свиней // Материалы Всероссийской научной конференции ВлИнфекционные болезни молодняка с\х животныхВ». М., 1996, 47-49.
- Скородумов Д.И., - Критерии дифференциации гемофильных бактерий патогенных для свиней // Сборник научных трудов ВлАктуальные вопросы инфекционных и инвазионных болезней животныхВ», Москва, 1996, 106-109.
- Скородумов Д.И. - Патогенные свойства культур возбудителя гемофилёзной плевропневмонии свиней // Сборник научных трудов ВлАктуальные вопросы инфекционных и инвазионных болезней животныхВ», Москва, 1996, 104-106.
Вместе с этим смотрят:
Алиментарная анемияАнализ ресурсов и оценка качества ремонта сельскохозяйственной техникиАнализ финансового состояния предприятия АПКАнализ финансовых результатов от реализации продукции растениеводства