Проводящая система листьев. Строение, типы жилкования
- Министерство образования Российской Федерации.
Башкирский Государственный Университет.
Биологический факультет
Кафедра ботаники
Курсовая работа по ботаники на тему:
Проводящая система листьев. Строение, типы
жилкования.
биологического
факультета
Уфа
- Содержание
1. Морфология, анатомия листа тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж... 3
2. Строение, типы жилкования тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж 4
3. Проводящая система листьев тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж. 7
4. Обкладка проводящих пучков тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж.. 17
5. Взаимосвязь между проводящими тканями стебля и листа тАж.. 19
6. Эволюция проводящей системы тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж.. 19
7. Заключение тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж. 21
8. Использованная литература тАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАжтАж. 22
Морфология, анатомия листа .
Разнообразие листьев цветковых растений богатство их форм поистине фантастическое морфологическое многообразие листьев цветковых несравнимо ни с одной другой группой высших растений , в том числе значительно превосходит многообразие листьев даже папоротников . Необычная эволюционная пластинчатость листа , удивительная его полиморфность в пределах часто одного семейства , а во многих случаях даже рода тАУ одна из характерных особенностей цветковых растений . Листья очень чутко реагируют на освещение и влажность и их колебания экологические условия отражаются не только на форме и размерах листа , но также на его строении , в частности на строении мезофилла , устьиц , кутикулы и характере жилкования . Поэтому изучение листьев имеет большое значение для экологии и физиологии растений.
Лист - часть побега орган высших растений, выполняющий функции фотосинтеза и транспирации, а также обеспечивающий газообмен с воздушной средой и участвующий в др. важнейших процессах жизнедеятельности растения. . Внешне листья разных растений сильно различаются, но между ними много общего. Листья большинства растений имеют зеленую окраску и состоят из листовой пластинки и черешка, которым они соединены со стеблем.
У двудольных лист обычно состоит из плоской пластинки, в которой происходят все основные физиологические процессы, связанные с фотосинтезом, и черешка тАУ суженной ножковидной части, прикрепляющей пластинку к стеблю. У многих растений листья не имеют выраженного черешка; такой лист называется сидячим. У большинства однодольных и многих двудольных основание листа расширено в так называемое влагалище более или менее охватывающие стебель. У основания листа многих растений имеются симметрично расположенные парные придатки, называемые прилистниками. Они бывают листовидные, чешуе видные, щетинновидные и прочие. По расположению на стебле различают очередные (спиральные), супротивные (в порах), и мутовчатое( по три или больше) листья.
Листья двудольных бывают простыми или сложными. Простой лист никогда не расчленяется на отдельные резко ограниченные сегменты, называемые листочками. Сложный лист, например лист конского каштана или большинства бобовых, напротив, разделён на листочки каждый из которых обычно снабжен собственным маленьким черешочком. Различают два основных типа сложных листьев тАУ перистосложные и пальчатосложные. В перистосложных листьях листочки расположены по обе стороны главной оси, или рахиса, представляющего собой продолжение черешка. Все листочки пальчатосложного листа отходят от верхушки черешка, и рахис у них отсутствует. Листочки типичных сложных листьев снабжены сочленением.
По форме пластинка простого листа, так же как пластинка листочков
сложного листа, бывает округлой, эллиптической, яйцевидной,
обратнояйцевидной ромбовидной, продолговатой, ланцетовидной, обратноланцетной, линейной, шиловидный, игловидной и т.д.(рис. 1)
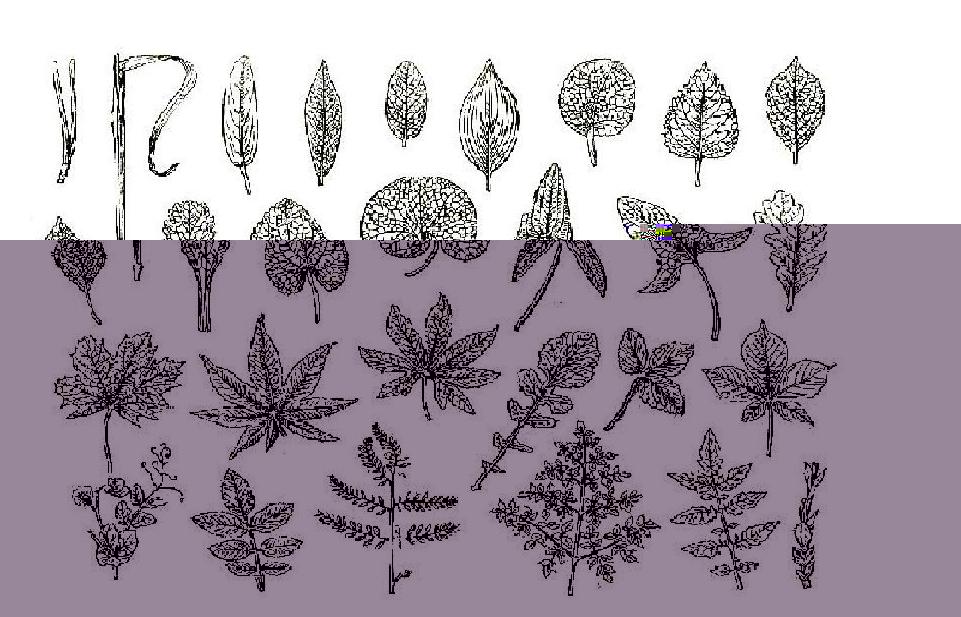
Рис. 1
Для характеристики пластинки большое значение имеет форма её основания которая может быть округлой, сердцевидной, почковидной, стреловидной, копьевидной, клиновидной, усеченной и т.д., а также верхушка, которая бывает острой, заострённой, остроконечной, тупой, вдавленной, выемчатой и прочие. Очень важны особенности края пластинки: она может быть цельнокрайней, городчатой, зубчатой, пильчатой, реснитчатой и т.д. В зависимости от характера и глубины расчленения пластинки листья бывают пальчатолопостными или перистолопостными, надрезанными, раздельными или рассеченными.
Строение, типы жилкования.
В листе проходят проводящие пучки, которые обычно различным образом ветвятся и анастомозируют. В большинстве случаев проводящая система листа выступает в виде так называемого жилкования, обычно особенно хорошо выраженного на нижней его стороне. Жилкование листьев отличается очень большим разнообразием и у разных групп цветковых растений имеет свои характерные признаки. Нередко, найдя на земле отдельный лист растения, можно определить по характеру жилкования, к какому роду, а иногда даже виду он относится. На это обстоятельство уже давно обратили внимание палеоботаники, которые, изучая детали жилкования на отпечатках, ископаемых листьев, часто безошибочно определяют если не видовую то родовую их пренадлежность. Поэтому не случайно, что классификацией типов жилкования и разработкой специальной терминологией для ее описания занялся австрийский палеоботаник К.фон Эттингсхаузен.
Проводяшая система листа цветковых растений обычно представляет собой довольно сложную картину, в которой более или менее ясно выделяются жилки разного порядка ветвления. Жилки первого порядка представляют собой самые толстые, главные жилки листа. У листьев с перистым жилкованием это средняя жилка, представляющая непосредственное продолжение черешка. В листьях с иным типом жилкования это несколько жилок, отходящих от верхушки черешка.
От первичных жилок отходят более тонкие вторичные жилки. Еще более тонкие жилки, ответвляющиеся обычно от вторичных жилок, а также
непосредственно от первичных, называются третичными жилками. Далее могут быть жилки четвертого, пятого и даже следующих порядков. Отходящие от первичных жилок ветви разных порядков обычно анастомозируют между собой и часто образуют сложную сеть, густо покрывающую всю пластинку листа. Рассматривая эту сеть при большом увеличении, мы заметим, что она состоит из более или менее ясно выраженных ячеек, или ареолей ( от латинского areola- площадка). Во многих случаях в этих ячейках можно видеть свободные концы отдельных тонких жилок, которые нередко ветвятся.
Жилки первого порядка ( первичные жилки) аналогичны магистральным линиям городской водопроводной сети, транспортирующим воду транзитом в удаленные районы снабжаемой территории, а все остальные жилки подобно распределительной сети, подающей воду к отдельным домовым ответвлениям. Бывают листья с одной, двумя или несколькими магистральными линиями. Задача транспорта воды и ассимилятов технически решается у них по- разному.
Листья большинства двудольных характеризуются жилкованием с одной главной магистральной линией. Это перистожилковатые или перистонервные листья (от лат. nervus тАУ жила, нерв). Боковые (вторичные) жилки отходят от средней (первичной). Жилки под более или менее острым, редко прямым углом. В зависимости от экологических условий средняя жилка, а также черешок бывают развиты в большей или меньшей степени. Усиление средней жилки связано с увеличением ее роли как главной магистральной линии, а усиление черешка связано с повышением его механических функций. Мощное развитие средней жилки и черешка особенно характерно для вечнозеленых листьев, деревьев тропических и субтропических дождевых лесов. Листья этих растений обычно крупные и тяжелые и поэтому снабжены мощным черешком, который имеет более или менее цилиндрическую форму. Как указывает в своей книге ВлАрхитектоника растенийВ» В.Ф.Раздорский , мощные черешки, так же как мощные средние жилки, являются хорошими пружинами, работающими на изгиб, что дает им возможность оказывать эффективное сопротивление таким динамическим воздействиям, как порывы ветра удары дождевых капель во время ливней и прочих погодных условий.
Боковые жилки у листьев с перистым жилкованием ведут себя по-разному. В некоторых случаях боковые жилки тянутся прямо до края пластинки и оканчиваются здесь в лопостях, концах зубчиков, выемках или даже выступают в виде щетинок или остей- это так называемое перистокраевое или краепедодромное ( от греческого Kraspedon-край, окраина и dromos- бег) жилкование. Оно встречается, например у каштана, бука, лещины, ольхи, березы, дзельквы, ильма, и многих других растений.Иначе ведут себя боковые жилки в листьях с перистопетлевидным или брохидодромным (от греч. вrochos тАУ петля) жилкованием. Они направляются здесь к краю пластинки, но еще не достигнув его заворачивают дугой вперед соединяясь со следующей передней боковой жилкой и образуя с ней петлю.Жилки образуют вдоль края листа все уменьшающиеся петли, которые ясно выделяются из остальной нежной сети более мелких жилок. Перистопетлевидное жилкование характерно для довольно большого числа двудольных, в том числе и для магнолии, лавра, ряда видов коричного дерева, камелии,миртовых и других растений.
Различают также перистосетчатое или диктиодромное (от греч. diktyon тАУсеть) жилкование, иногда неудачно называемое ВлретикулодромнымВ». В отличие от брохидодромного типа жилки второго порядка по направлению к краю листа в результате повторного ветвления постепенно образуют все более густую сеть, ясно выраженные густые петли здесь отсутствуют. Такое жилкование хорошо выражено у видов барбариса, ивы, радодендрона, груши, яблони, айвы. В тех случаях, когда боковые жилки свободно ветвятся по напрвлению к краю пластинки, не достигая его и не образуя петель, жилкование называется кладодромным (от греч. klados тАУ ветвь).
Еще более разнообразны типы жилкования с двумя и большим числом магистральных линий. Среди двудольных, широко распространены листья с так называемым пальчатым или дальневидным жилкованием. Это пальчатожилковатые или пальчатонервные листья. Первичные жилки отходят радиально от одной точки или у самого основания пластинки (базально) или несколько выше основания (супрабазально). Примером такого жилкования могут служить листья многих видов клена. В пальчатом жилковании повторяются те же основные типы, которые мы видели у перистого жилкования. Это пальчатокраевое или пальчатокраепедодромное жилкование (например у ликвидальбара, платана, клена, стеркумис, фастеин, виноград), пальчатопетлевидное или пальчатоброхидодромное жилкование например у церцидифиллума, пальчатосетчатое или пальчатодиктиодромное жилкование у тетрацентрона, лукосемянника и нудина дерева. В широком смысле слова к пальчатому типу можно отнести жилкование ряда щитовидных листьев например: лотоса, клещевины, настурции и других.
От типичного пальчатого жилкования отличается пальчатовершинобежное или пальчатоакродромное (от греч. akron тАУ вершина) жилкование. Из трех или больше первичных жилок расходящихся у основания пластинки (базально) или несколько выше (супрабазально) , боковые, которые иногда бывают развиты несколько слабее средней, направляются к краю листа, но недоходят до него, сворачивают в виде дуги к верхушке и там теряются. Такое жилкование характерно для многих двудольных (виды Cinnamomum, Cocculus, Melasbomataceal, Coriaria, Paliurus,Ziziphus, Rhamnus, Ceahothus, Baccharis и другие) и некоторых однодольных например Paris.
Особым типом жилкования, отличным как от пальчатого, так и от перистого, является дуговидно - кривобежное или кампилодромное (от греч. Campylos тАУ согнутый) жилкование. Первичные (главные) жилки вступают по нескольку и всегда более или менее отдельно в пластинку, причем внешние из них направляются параллельно краю пластинки другой к вершине листа. Многие из боковых первичных жилок не достигают вершины пластинки и присоединяются к соседней жилке. Вторичные жилки, также тонкие и нежные, что их часто нельзя заметить простым глазом, образуют всегда перемычки (так называемые комиссуральные жилочки), соединяющие в поперечном направлении соседние главные жилки. Подобное жилкование мы встречаем у многих однодольных, например у сусановых, водокрасовых, частуховых, рода хоста (Hosta), банановых, канновых.
К дуговидно тАУ кривобежному типу морфологически очень близко жилкование, известное под не вполне удачным названием параллельного, параллельнобежного или параллелодромного (от греч. parallelos тАУ рядом идущий). Первичные жилки (две или больше) входят самостоятельно из листового влагалища в основание пластинки и идут далее более или менее параллельно до верхушки листа, где смыкаются. Как и у листьев с дуговидно тАУ кривобежным жилкованием, параллельные первичные жилки соединены прямыми или косыми перемычками тАУ тонкими комиссуральными жилочками. Имеются переходы от дуговидно тАУ кривобежного к параллельному типу жилкования. Типичное параллельное жилкование можно видеть у многих лилейных, орхидных, осок и особенно злаков, а также у некоторых двудольных например у некоторых эпакрисовых.
Проводящая система листьев.
Размещение проводящих пучков то есть жилкование, придает листьям характерный вид. Слово ВлжилкованиеВ» происходит от термина жилка, который применяется в ботанике либо к проводящему пучку или группе тесно сближенных пучков, либо к пучкам совместно с пространственно объединенной с ними непроводящей тканью. В настоящей главе термином жилка мы обозначаем проводящий пучок или группу тесно сомкнутых пучков.
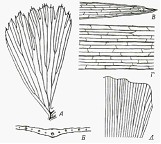
Для листа может быть характерно наличие одной, двух или более жилок. Примеры листьев с одной жилкой можно найти у хвойных и у Equisetum, тогда как листья со многими жилками обычно для высших папоротников и покрытосеменных. У покрытосеменных преобладают два типа жилкования тАУ сетчатое и параллельное. При сетчатом жилковании, которое широко распространено среди двудольных, различные по размерам жилки боразуют анастомозирующую сеть, причем мелкие жилки отходят от более крупных. У листьев с параллельным жилкованием, которые характерны для однодольных, жилки сравнительно одинакового размера располагаются продольно, но сходятся друг с другом и сливаются у верхушки листа или на обоих концах пластинки или у ее краев.(Рис. 2)
Рис. 2 Типы жилкования листа.
Просветлённые листья (А, В тАУ Д) и поперечный разрез (Б). А. Открытое дихотомическое жилкование у двудольного Kingdonia uniflora. Б тАУ Г. Параллельное жилкование у злака Avena.На рис. В и Г поперечные анастомозы между продольными жилками. Д. Открытое дихотомическое жилкование у Ginkgo.
На протяжении всей пластинки продольные жилки соединяются тонкими перемычками. Эти анастомозы часто располагаются наподобие ступеней лестницы (рис. 2, Г). Но могут располагаться также иным путем. Для некоторых однодольных характерен видоизмененный тип параллельного жилкования, при котором жилки на некотором протяжении располагаются продольно, а затем перисто раздваиваются ( рис. 3 Д). Параллельное жилкование наблюдается также у некоторых двудольных (Plantado,Tradopodon) и соответственно некоторые однодольные имеют сетчатое жилкование (Araceae, Smilacoideae, Taccaceae, Orchidaceae).
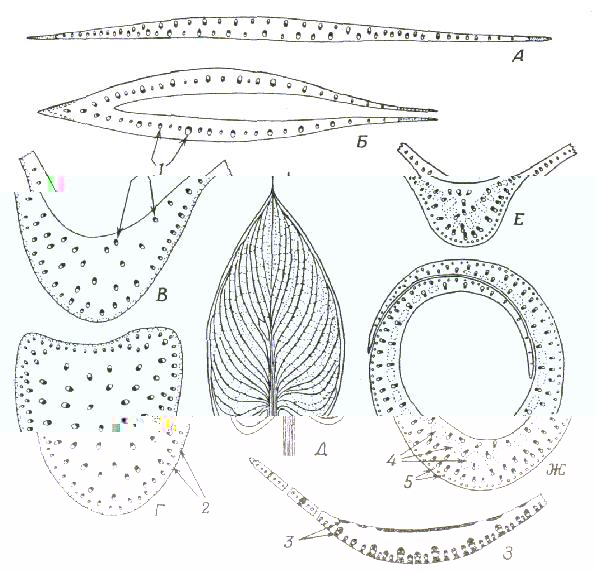
Рис. 3. Проводящая система в листьях однодольных.
А. Поперечный разрез пластинки листа Iris; Б. Поперечный разрез влагалища листа Iris; В. Поперечный разрез средней жилки листа Zantedeschia; Г. Поперечный разрез черешка листа Zantedeschia; Д. Вид листа Zantedeschia с поверхности; Е. Поперечный разрез средней жилки листа Canna; Ж. Поперечный разрез влагалища листа Canna; З. Поперечный разрез средней жилки и части пластинки Zea; В проводящих пучках ксилема обозначена черным, а флоэма тАУ белым.
1 тАУ проводящие пучки; 2 тАУ колленхима; 3 тАУ склеренхима; 4 тАУ лакуны; 5 тАУ пучки волокон.
Обе системы жилкования, как сетчатая так и параллельная, называются закрытыми так как жилки анастомозируют друг с другом. У представителей двух реликтовых родов двудольных тАУKinqdonia и Circaeaster тАУ было обнаружено, что листья их имеют открытое дихотомическое (многократно вильчатое) жилкование ( рис. 2 А ), напоминающее жилкование Ginnqo (рис. 2,Д) и некоторых папоротников. Открытым такое жилкование называется потому, что крупные подразделения системы жилок свободно оканчиваются в толще листа или на его краях.
В анастомозирующей проводящей системе листьев двудольных жилки имеют самые различные размеры. Наиболее крупная жилка часто занимает медианное положение и образует среднюю жилку, а несколько меньше по размерам жилки ответвляются от нее латерально ( рис.125 ; лист с перистым жилкованием). У других листьев может быть несколько крупных жилок, сходных по размерам и расходящихся от основания пластинки по направлению к краям. Крупные жилки обычно встречаются в утолщенных частях пластинки, которые имеют вид ребер на абаксиальной поверхности листа ( рис.4,126).
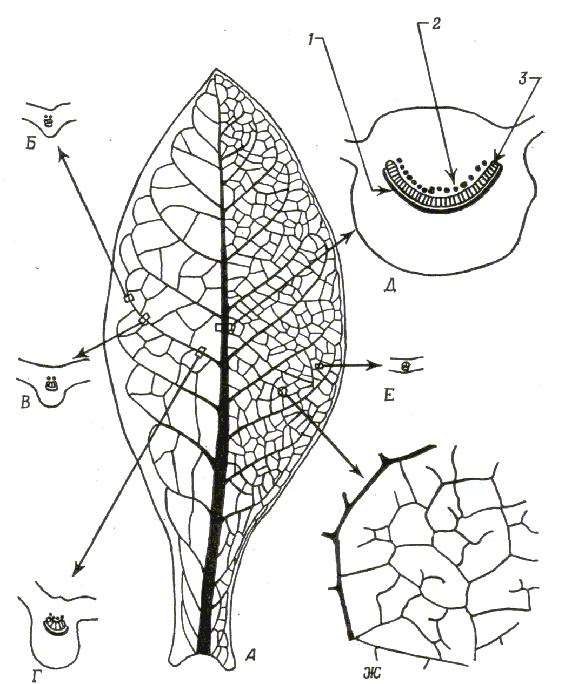
Рис. 4 Жилкование листа Nicotiana tabacum.
А. Зрелый лист со средней жилкой, основными боковыми жилками и крупной сетью проводящих пучков; Б тАУ Е. Поперечные срезы жилок (схема), ксилема на которых заштрихована, а флоэма (наружная и внутренняя) обозначена черным. Ж. Часть пластинки, на которой видны мельчайшие жилки и их свободные окончания в мезофилле; 1 тАУ наружная флоэма; 2 тАУ внутренняя флоэма; 3 тАУ ксилема.
Эти ребра состоят из паренхимы с относительно небольшим содержанием хролофилла и некоторого количества механической ткани, обычно представленной колленхимой. Проводящие пучки крупных жилок погружены в паренхиму и таким образом до известной степени отделены от самого мезофилла ( рис. 5).
Рис. 5 Строение листа Nicotiana tabacum на поперечном срезе.
1 тАУ палисадный слой; 2 тАУ губчатый мезофилл; 3 тАУ устьица; 4 тАУ ребро; 5 тАУ внутренняя флоэма; 6 тАУ ксилема; 7 тАУ наружная флоэма.
Мелкие жилки, напротив, проходят по самой толще мезофилла, образуя между крупными жилками густоразветвленную сеть. Они располагаются в средней части мезофилла, обычно, под палисадными клетками, то есть иными словами находятся в верхнем слое губчатой паренхимы ( рис. 6,7).
Рис. 6 Поперечный разрез листа груши.1 тАУ палисадная паренхима; 2 тАУ обкладка пучков; 3 тАУ верхний эпидермис; 4 тАУ продолжение обкладки пучка; 5 тАУ волокна; 6 тАУ ксилема; 7 тАУ флоэма; 8 тАУ губчатая паренхима; 9 тАУ нижний эпидермис.
Рис. 7 Поперечный разрез листьев 
А. Nerium oleander. Б. Lilium.
1 тАУ многослойный эпидермис; 2 тАУ друза; 3 тАУ палисадная паренхима; 4 тАУ губчатая паренхима; 5 тАУ устьице; 6 тАУ устьичная крипта; 7 тАУ трихома; 8 тАУ обкладка пучка.
В листьях двудольных распределение мелких жилок образует широкий ряд взаимосвязанных типов (рис.8). Ответвление этих жилок разделяют мезофилл на серии последовательно уменьшающихся многоугольников с конечными ответвлениями, то есть окончаниями пучков, или жилок, проникающими в мельчайшие подразделения мезофилла (areoles) и свободно там заканчивающимися. В этих небольших участках свободные окончания жилок могут и отсутствовать (рис.8,А). Тонкоразветвленное жилкование с параллельно расположенными мелкими жилками встречается у представителей Quiinaceae и Rubiaceae.
Рис. 8 Жилки продолжения обкладки пучков.
А. Tilia americana. Б. Quercus marcocarpa. В. Morus alba. Г. Ricinus communis. Двойными линиями обозначены линии жилки с продолжениями обкладки; сплошными черными линиями тАУ жилки без продолжений обкладки.
В листе однодольных продольные жилки могут иметь почти равную толщину или варьировать по своим размерам, причем крупные жилки в этом случае чередуются с мелкими. Средняя жилка может быть крупнее остальных и сочетаться с хорошо выраженным ребром (рис.2,Б). Боковые жилки иногда образуют ребра, а иногда и нет. У некоторых крупных злаков средняя часть пластинки утолщена в среднюю жилку за счет дифференциации массивной бесцветной паренхимы на адаксиальной стороне (рис.3,З). В такой средней жилке располагаются многочисленные проводящие пучки. У многих однодольных мельчайшие пучки простираются от одной крупной жилки к другой, а у других в мезофилле встречаются свободные окончания жилок.
Жилкование катафиллов, гипсофиллов и семядолей сходно с жилкованием вегетативных листьев того же растения, но является более простым, создается впечатление, что оно несколько недоразвито.
Проводящие пучки различных размеров по своему гистологическому составу обнаруживают количественные и качественные различия. Наиболее крупные пучки содержат ксилему и флоэму в количествах, сравнимых с теми, которые имеются в пучках черешка или листового следа. В коллатеральных пучках ксилема располагается на адаксиальной, а флоэма тАУ на абаксиальной стороне листа (рис.6). Если листовые следы являются биколлатеральными, адаксиальная флоэма встречается также и в листе, но в мелких жилках может отсутствовать (рис.4,5). В наиболеекрупных жилкахлистьев двудольных проводящая ткань образует либо один, либо несколько пучков (рис.9). Как видно на поперечных срезах жилок, проводящие пучки могут располагаться в виде кольца (рис.9,Д;Liriodendron, Vitis), полукольца (Ambrosia) или иметь беспорядочное распределение (Silphium, Helianthus). В тех случаях, когда имеется один проводящий пучок, у некоторых растений он имеет форму полумесяца (рис.9, В; Cercis, Ulmus, Tilia, Abutilon), а у других тАУ округлую форму (рис.9, Ж; Catalpa, Acer, Quercus).
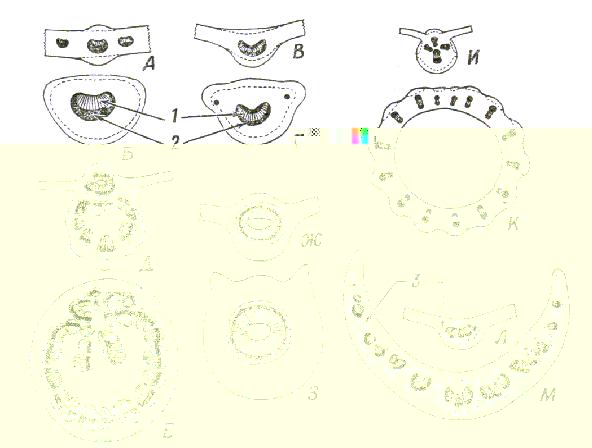
Рис. 9 Проводящая ткань на поперечных разрезах средних жилок и черешков листьев двудольных.
А,Б. Euonymus; В.Г. Nerium; Д,Е. Platanus; Ж,З. Citrus; И,К. Cucurbita; Л,М. Mahonia. В каждой паре рисунков верхний из них представляет собой поперечный разрез средней жилки, а нижний тАУ черешка.
1 тАУ ксилема; 2 тАУ флоэма; 3 тАУ склеренхима.
В крупных жилках листьев двудольных могут встречаться как первичные, так и вторичные ткани, более же мелкие обычно являются полностью первичными. Камбиальная активность в листьях представителей вечнозеленых видов выражена сильнее, чем у листопадных растений. В жилках различных размеров, за исключением самых мелких, в ксилеме имеются сосуды, а во флоэме тАУ ситовидные трубки. В мелких жилках трахеальные элементы представлены трахеидами. Вблизи окончаний ответвлений жилок флоэмная часть может содержать только паренхиму (рис.10, Ж) и окончания жилок у двудольных нередко состоят лишь из трахеид (рис.10,А,Б,Г,З,). Однако у представителей некоторых видов ситовидные элементы все время сопутствуют трахеидам, а иногда даже оказываются расположенными дальше их. Ситовидные элементы в окончаниях пучков часто ассоциированы с исключительно крупными клетками-спутниками. Трахеиды обычно имеют спиральные и иногда кольчатые утолщения. У самого конца жилки иногда встречается одна трахеида, в других случаях тАУ две трахеиды, располагающиеся параллельно друг другу, или наблюдаются расположенная без порядка группа трахеид. За последними трахеидами окончаний жилок, находясь в контакте с ними, могу дифференцироваться склереиды. У представителей некоторых родов жилки увенчиваются крупными, имеющими овальную форму или неправильно разветвленными трахеидами, нередко с пористыми стенками. Иногда эти клетки рассматривают как несущие функции накопления и хранения запасов воды и называют запасающими трахеидами.
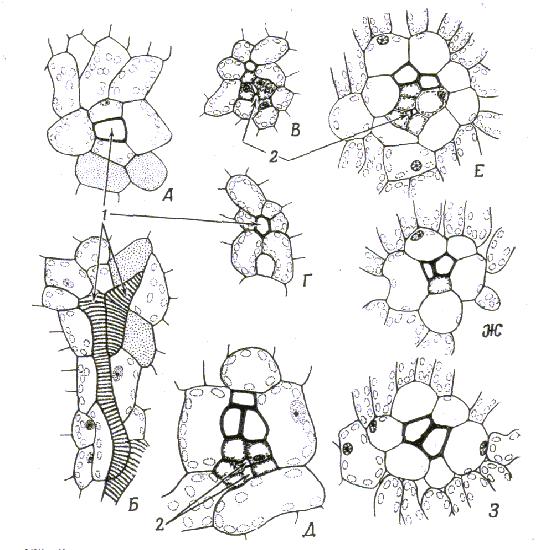
Рис. 10 Строение мелких жилок листа двудольных. Поперечный и продольный тангентальный разрезы мелкой жилки листа.
А, Б.Vitis vinifera. Окончания жилок состоят из трахеид, окруженных клетками обкладки; В, Г. Поперечный разрез мелкой жилки листа Humulus. Одна жилка содержит трахеиды, ситовидные элементы и некоторое количество паренхимных клеток, другая тАУ лишь одну трахеиду; Д. Поперечный разрез мелкой жилки листа Nicotiana tabacum; Е тАУ З. Поперечные разрезы мелкой жилки листа Prunus;
1 тАУ трахеиды; 2 тАУ ситовидные элементы.
В мелких жилках однодольных также имеется мало проводящих элементов. Поперечные анастомозы в листьях злаков могут состоять из одного ряда трахеальных и одного ряда ситовидных элементов (рис. 11 ,Б-Д). в небольших проводящих пучках однодольных, так же как и двудольных, ситовидные элементы могут располагаться рядом с трахеальными элементами (рис. 11).
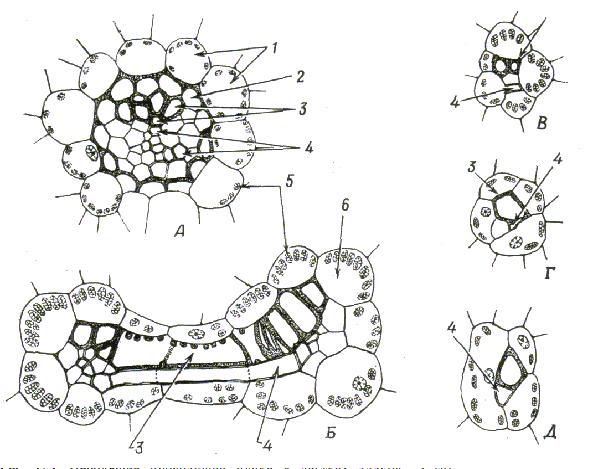
Рис. 11 Небольшие проводящие пучки в листьях злаков.
А. Продольный пучок на поперечном разрезе листа Tritticum. Б. Два продольных пучка, связанных поперечно расположенным пучком на поперечном разрезе листа Zea. В. Один из мельчайших продольных пучков Zea. Г,Д. Поперечные анастомозы у Zea, какими они представляются на срезах, сделанных параллельно длинной оси листа и перпендикулярно слоям эпидермальных клеток.
1 тАУ наружная обкладка пучка; 2 тАУ внутренняя, или местомная, обкладка пучка; 3 тАУ трахеальные элемент; 4 тАУ ситовидные элементы; 5 тАУ хлоропласты; 6 тАУ клетка обкладки пучка.
Особенно важным признаком характеризующим проводящую систему листа, является её тесная пространственная связь с мезофиллом. Измерения, проведенные на представителях шести видов двудольных тАУ травянистых кустарниковых и древесных, - показали, что общая длина жилок в среднем составляет 102см на 1см листовой поверхности. Интенсивное развитие проводящей ткани в мезофилле иллюстрируется небольшими размерами участков, свободных от жилок. Согласно некоторым данным, расстояние между жилками в листьях двудольных составляет в среднем около 130 мн. Между распределением жилок и теми структурными особенностями непроводящей тканей листа, которые могут оказывать влияние на проведении веществ, существует значимая корреляция. Так, чем больше объем ткани, у которой латеральный контакт между клетками относительно невелик тАУ размещение, которое определяет относительно низкую эффективность латерального проведения веществ, - тем ближе друг к другу располагаются проводящие пучки. Напротив, чем больше объем тканей, клетки которых имеют обширные латеральные контакты ( как, например, эпидермис и губчатая паренхима ), тем больше промежутки между жилками. Световые листья, у которых памиадная ткань обычно особенно сильно развита, имеют большую длину жилок, чем теневые листья.
Обкладка проводящих пучков.
В листьях двудольных крупные проводящие пучки окружены клетками паренхимы с незначительным количеством хлоропластов, тогда как небольшие пучки располагаются непосредственно в мезофилле. Эти небольшие пучки не соприкасаются с межклетными пространственными, но обычно окружены слоем плотно сомкнутых паренхимных клеток тАУ обкладкой пучков (рис.10). У двудольных паренхима обкладки пучков называется также окаймляющей паренхимой.
Обкладки пучков в листьях двудольных обычно состоят из клеток, удлиненных в направлении, параллельном ходу пучка, и имеют такие же тонкие оболочки, как и оболочка примыкающих к ним клеток мезофилла. У некоторых растений эти клетки содержат такое же количество хлоропластов, как и клетки мезофилла (рис.10. В-Д); у других же растений хлоропластов в них мало или они вовсе лишены их (рис.10,Е-З). Отдельные клетки обкладки могут содержать кристаллы. Обкладки простираются до окончания жилок и полностью окружают конечные трахеиды (рис.10, Б).
У многих двудольных тяжи клеток, сходных с клетками обкладки пучков, тянутся от обкладки по направлению к одному или к обоим эпидермисам, причем некоторые из них заканчиваются в мезафилле, другие же достигают эпидермиса ( рис.6 ). Этим продолжениям обкладки пучков уделялось в литературе большое внимание. Проведенные на листьях некоторых двудольных измерения показали, что жилки окружены паренхимой обкладки на расстоянии, составляющем 99 % общей длины. Если обкладки пучков и их продолжения связаны с проведением веществ, их наличие существенно увеличивает область контакта между мезофиллом и проводящими клетками.
Некоторые наблюдения дают основание полагать, что обкладки пучков и их продолжения принимают участие в проведении веществ. Оказалось, что введенный в листья раствор ферроцианида калия проходит по жилкам в обкладке пучков, а через продолжения обкладки тАУ в эпидермис и распространяется по всей его поверхности. Затем было установлено существование корреляции между распространением жилок и наличием продолжений обкладки пучков ( рис.8 ).
Наиболее распространены обкладки пучков, состоящие из паренхимных клеток, однако у некоторых двудольных различные по размерам пучки окружены склеренхимой (Winteraceae, Melaqtomaceae ). У некоторых представителей Winteraceae склеренхима облекает даже конечные ответвления жилок.
В листьях однодольных, также встречаются обкладки пучков. Они лучше всего изучены у представителей Gramineae. Листья злаков имеют два типа обкладок: полностью паренхиматические с хлоропластами и состоящие из относительно толстостенных клеток, не содержащие хлоропластов. Обкладку, состоящую из клеток с толстыми оболочками,Швенденер назвал местомной обкладкой, так как термин тАЬместомтАЭранее использовался для обозначения проводящих элементов пучка. Если местомная обкладка имеется, она располагается в непосредственном соседстве с проводящей тканью, а с наружной ее стороны находится вторая обкладка, состоящая из тонкостенных клеток, содержащих хлоропласты.
Среди Gramineae у многих представителей Panicoideae небольшие проводящие пучки окружены одной обкладкой из тонкостенных клеток ( рис.11,Б-Д обкладка вокруг более крупных пучков может быть составлена относительно толстостенными клетками), тогда как у представителей Festucoideae часто имеется две обкладки ( рис.11А ). Внутренняя или местомная обкладка состоит из удлиненных живых клеток с тупыми или заостренными концами. Утолщение оболочек варьируют даже в различных частях одной обкладки и часто являются простыми. Иногда внутренние стенки клеток оказываются толще, чем наружные. В небольших пучках внутренняя обкладка может быть приурочена только к флоэмной части пучка.
В листьях порытосеменных обкладка пучка представляет собой эндодерму. Хотя в большинстве случаев полосы Каспари неразличимы, оболочки и содержимое клеток обкладки могут реагировать на различные красители и индикаторы таким же образом, как оболочки и содержимые клеток типичной эндодермы других частей растения. Кроме того, полосы Каспари были обнаружены в местомных обкладках молодых листьев некоторых Gramineae и Cyperaceae.
Обкладка пучка может быть выражена также в виде крахмалоносного влагалища. В некоторых двудольных и у злаков, которые имеют однослойные обкладки пучков, в клетках паренхимной обкладки образуется крахмал. У представителей таких родов, как Zea и Sorghum, хролопласты в клетках обкладки особенно крупны ( рис.11Б-Д ) и, по тАУвидимому, представляют собой единственные пластиды листа, связанные с образованием крахмала во время активного фотосинтеза. При исследовании ультраструктуры этих хлоропластов было обнаружено, что у Zea они лишены гран. У представителей Festucoideae, имеющих двойные обкладки вокруг пучков ( рис.11 А), в клетках внутренней обкладки хлоропласты отсутствуют, а в клетках наружной имеют несколько меньшие размеры, чем в остальных клетках мезофилла. Крахмал образуется во всех зеленых клетках, так что в этом отношении обкладка не представляется сколько- нибудь заметно дифференцированной.
Обкладка пучков двудольных, единственная паренхиматическая обкладка представителей Panicoideae и наружная из двух обкладок представителей Festucoideae по своему происхождению, по-видимому, являются частью основной ткани. Внутренняя, или местомная, обкладка, возможно, имеет прокамбиальное происхождение.
Взаимосвязь между проводящими тканями
стебля и листа.
Расположение проводящих пучков в стебле указывает на тесную связь в строении и развитии между ним и его боковыми придатками, то есть листьями. Общий термин Вл побег В» служит не только удобным обозначением для комплекса надземных вегетативных органов, но и отражает эту взаимосвязь.
Прокамбиальные тяжи стебля начинаются сразу же за апикальной меристемой под развивающимися листовыми примордиями, а иногда под местом их будущего заложения еще до начала этого развития. По мере роста примордиев в длину тяжи дифференцируются по направлению к кончику листа. Таким образом, уже с самого начала прокамбиальная система последнего оказывается непрерывно связанной с прокамбием стебля.
В каждом узле один или более проводящих пучков идут от центрального цилиндра стебля через кору в лист ( или листья ) данного узла. В стебле пучок, направляющийся от продольного стеблевого пучка к основанию листа, где он связывается с проводящей системой последнего, называется листовым следом, а широкий промежуток, то есть участок основной ткани, находящийся в центральном цилиндре над местом отхождения листового следа, называется листовым прорывом. В один лист может входить из стебля один или более листовых следов. Число междоузлий, которые они при этом пересекают, различно, то есть длина листовых следов неодинакова.
Если проследить ход проводящего пучка вверх и вниз по стеблю, выяснится, что он связан с несколькими листовыми следами, образуя так называемый симподий. В одних стеблях некоторые или все симподии взаимосвязаны, в то время как в других каждый из них представляет собой независимый проводящий комплекс. В любом случае структура проводящей системы стебля отражает расположение на нем листьев.
Эволюция проводящей системы.
Проводящая система цветковых растений достигла наиболее высокого уровня эволюционного развития. Проводящая система у цветковых растений оказалась значительно более совершенной, чем у голосеменных, а тем более у папоротников и других групп высших растений. Некоторые из наиболее примитивных представителей ныне живущих двудольных, такие, как виды семейства винтеровых ( Winteraceae) и роды троходендрон ( Trohodendron ) и тетрацентрон ( Tetracentron ), по строению проводящей системы мало чем отличаются от примитивных представителей голосеменных типа современных саговниковых или вымерших беннеттитовых. У названных родов нет сосудов, а имеются только трахеиды. В протокселеме эти трахеиды с кольчатыми и спиральными утолщениями, в метаксилеме обычно лестничные. Иногда отсутствие сосудов у травянистых растений тАУ явление вторичное ( у рясковых ).
Лишь у относительно немногих цветковых растений сохранилась бессосудистая ксилема. У подавляющего их большинства наряду с трахеидами имеются также сосуды, являющиеся основными водопроводящими элементами. В отличие от трахеиды каждый членик сосуда имеет сквозные отверстия, называемые перфорациями. Наиболее примитивные членики сосудов очень похожи ветереновидной формой и заостренными концами на трахеиды. Они очень длинные, узкие, в поперечном сечении угловатые, тонкостенные, не имеют конечной стенки или с очень слабо выраженной и очень косой конечной стенкой. Боковые стенки таких сосудов имеют ещё лестничные окаймлённые поры, а перфорационная пластинка, то есть место соприкосновения и сообщения двух соседних члеников, состоит из многочисленных лестничных перекладин чередующихся с продолговатыми перфорациями. Немецкий ботаник Антон де Бари впервые высказал мысль, что лестничная перфорация члеников сосудов возникла из лестничной поровости трахеид в результате исчезновения замыкающих пленок пор в местах соприкосновения налегающих друг на друга стенок соседних трахеид. В эволюционном смысле от лестничной трахеиды до членика сосуда с лестничной перфорации лишь один шаг и неудивительно поэтому, что сосуды возникли независимо и гетерохронно в разных линиях развития цветковых растений. Они возникли не только совершенно у двудольных и однодольных, но появились независимо даже в разных группах как двудольных, так и однодольных. Превращение лестничных трахеид в членике сосудов тАУ один из ярких примеров параллельной эволюции.
Как и все другие структурные элементы, членики сосудов в процессе эволюции совершенствуются. Длина их постепенно уменьшается, они становятся шире и в большинстве случаев приобретают более толстые стенки. Сечение их на поперечном срезе становится округлым, лестничные боковые поры заменяются более или менее округлыми окаймлёнными порами, которые располагаются сначала в горизонтальных рядах ( супротивная поровость ), а затем расположение их становится очередным, в виде косых рядов. Возникают ясно выраженные конечные стенки, на первых этапах эволюции ещё очень косые. Постепенно они принимают поперечное положение, то есть располагаются под прямым углом к длине сосуда. По мере укорочения длины и увеличения диаметра члеников сосуда отверстия в лестничной перфорационной пластинке расширяются, число перекладин уменьшается, и в конце концов после исчезновения всех перекладин образуется одна большая перфорация, называемая простой, простая перфорация тАУ это наиболее совершенный тип сквозного отверстия между члениками сосудов, так как сопротивление току жидкости сведено здесь к минимуму. Высшим, наиболее совершенным типом членика сосуда является короткий бочонкообразный членик, ширина которого превосходит его длину. Эволюция члеников сосудов тАУ один из самых ярких и наиболее документированных примеров приспособительной эволюции. Это также пример эволюционного ряда, который завершается кульминационным типом, представляющим собой конечное звено в цепи структурных преобразований.
Эволюция ситовидных трубок цветковых растений также начинается с очень примитивных типов, близких к ситовидным клеткам голосеменных. Членики ситовидных трубок отличаются от ситовидных клеток голосеменных главным образом наличием ясно выраженных ситовидных участков, представляющих собой более тонкие места ( углубления ) первичной стенки, пронизанные порами, через которые протопласты соседних члеников сообщаются посредствам связующих тяжей. Предполагают, что поры обычно возникают из каналов плазмодесм путем их ферментативного тАЬрассверливаниятАЭ. Эти ситовидные участки представляют собой видоизменённые первичные поровые поля обыкновенных паренхимных клеток. Связующие тяжи ситовидных участков значительно толще плазмодесм первичных поровых полей, и, кроме того, каждая пора в ситовидном участке обычно содержит маленький каллозовый цилиндр, через который проходит тяж ( каллоза тАУ полисахарид, состоящий из остатков глюкозы, соединенных в спиральную цепочку ). У голосеменных связующие тяжи ещё тонкие и похожи на обыкновенные плазмодесмы, но у цветковых растений они достигают значительной толщины. В процессе эволюции происходит постепенное утолщение связующих тяжей и окружающих их каллозовых трубок.
На более ранних стадиях эволюции ситовидных трубок все ситовидные участки данного членика одинаковы, но затем начинают выделяться участки с более развитыми каллозовыми трубками. Такие, более специализированные ситовидные участки данного членика одинаковы, но затем начинают выделяться участки с более развитыми каллозовыми трубками. Такие, более специализированные ситовидные участки обычно локализуются на определенных стенках члеников, чаще всего на конечных. Части стенки, несущие такие, более специализированные ситовидные участки, называются ситовидными пластинками. Ситовидная пластинка может состоять из нескольких или многих ситовидных участков ( с лестничным, сетчатым или иным их расположением ). Такая пластинка носит название сложной. Если ситовидная пластинка состоит из одного ситовидного участка, её называют простой.
Наиболее примитивные формы ситовидных трубок состоят из довольно длинных узких и заостренных члеников с очень косыми конечными стенками и с более или менее одинаковыми ситовидными участками на конечных и боковых стенках. Подобно конечным стенкам члеников сосудов, конечные стенки члеников ситовидных трубок в процессе эволюции постепенно принимают все менее наклонное положение и в конце концов часто становится поперечным, то есть располагаются под прямым углом к боковым стенкам. Одновременно с этим происходит постепенная локализация ситовидных участков на конечных стенках. При этом сложные ситовидные пластинки переходят в простые, более приспособленные для транспорта ассимилятов в растении. Этот процесс аналогичен превращению лестничной перфорации члеников сосудов в простую. В обоих случаях совершенствуется механизм передвижения жидкостей.
Наконец, в процессе эволюции цветковых растений происходило уменьшение длины и увеличение диаметра члеников ситовидных трубок, что, однако не привело здесь к тем очень коротким и широким бочонкообразным отдельностям, которые встречаются у высокоспециализированных сосудов.
Наблюдается определенная корреляция в эволюции ситовидных трубок, и, как правило, уровень специализации ситовидных трубок соответствует уровню развития сосудов.
Заключение.
У большинства двудольных листья состоят из пластинки и черешка. Иногда пластинки разделены на листочки. Устьица обычно более многочисленны на нижней стороне листа. Основная ткань, или мезофилл, листа является специализированной фотосинтезирующей и у мезофитов дифференцирована на палисадную и губчатую паренхиму. Она обильно пронизана межклетниками и жилками, то есть проводящими пучками, состоящими из флоэмы и ксилемы и окруженными паренхиматозными обкладками. Ксилема обычно расположена на верхней стороне жилки, а флоэма на нижней.
У большинства однодольных, включая злаки, лист состоит из пластинки и влагалища, охватывающего стебель. Листья С3- и С4- злаков имеют существенные анатомические различия. Наиболее значительным из них является присутствие у С4- и отсутствие у С3- видов кранц-анатомии, то есть такого строения, при котором клетки мезофилла и обкладки располагаются вокруг проводящих пучков двумя концентрическими слоями.
Листья образуются в периферической зоне апекса побега, и их положение на стебле отражается в особенностях расположения в нем проводящей системы. Их рост детерминированный, то есть относительно непродолжителен, тогда как у вегетативных апексов побега он может быть неограниченным, или недетерминированным. У многих видов листья, выросшие при высокой освещенности, мельче и толще развившихся при относительном затенении. Первые называются световыми, вторые тАУ теневыми.
У многих растений опадению листьев предшествует формирование у основания черешка отделительной зоны.
Использованная литература.
- К. Эсау тАЬАнатомия растенийтАЬ. М : - тАЬМиртАЭ. 1969
- тАЬЖизнь растенийтАЭ (под ред. А.Л. Тахтаджян). М : тАЬПросвещениетАЭ. 1980
- тАЬЖизнь растенийтАЭ (под ред. М.М. Голлербах). М : тАЬПросвещениетАЭ.1977
- Богданова. Биология. М., 1991
- Серебряков И.Г., тАЬМорфология вегетативных органов высших растенийтАЭ. М., 1952
- П. Рейвн, Р. Эверт, С. Айкхорн. тАЬСовременная ботаникатАЭ
Вместе с этим смотрят:
Прогнозы ученых на ХХI векПрограмма эмуляции развития популяций животныхПрокариоты и вирусы, их строение и функционированиеПростейшие паразиты человека