Эукариотическая клетка
МИНИСТЕРСТВО НАУКИ И ПРОСВЕЩЕНИЯ РЕСПУБЛИКИ КАЗАХСТАН
КАФЕДРА БИОЛОГИИ
Контрольная работа
по предмету ВлМолекулярная биологияВ»
тема ВлЭукариотическая клеткаВ»
Выполнила ст-ка
гр. ЗБХ-31
КОТ Татьяна
г.Павлодар, 2002г.
План
- Структурно-функциональная организация эукариотической клетки.
а) биологическая мембрана;
б) транспорт через мембраны малых молекул;
в) мембранный транспорт молекул и частиц (экзоцитос и эндоцитоз);
2. Преобразование энергии (митахондрии и хлоропласты).
Эукариоты появились среди обитателей планеты около 1,5 млрд. лет назад. Отличаясь от прокариот более сложной организацией, они используют в своей жизнедеятельности больший объем наследВнственной информации. Так, общая длина молекул ДНК в ядре клетки млекопитающего составляет примерно 5 тАв 109 пар нуклеотидов, т. е. в 1000 раз превосходит длину молекулы ДНК бактерии.
Первоначально эукариоты имели одноклеточное строение. ДоВнисторические одноклеточные эукариоты послужили основой для возникновения в процессе эволюции организмов, имеющих многоВнклеточное строение тела. Они появились на Земле около 600 млн. лет назад и дали широкое разнообразие живых существ, расселивВншихся в трех основных средах: водной, воздушной, наземной. Полезно заметить, что многоклеточность возникла в эволюции в период, когда атмосфера планеты, обогатившись, приобрела устойчивый окислительный характер.
Структурно-функциональная организация эукариотической клетки
Эукариотический тип клеточной организации представлен двумя подтипами. Особенностью организмов простейших (рис. 2.2) являетВнся то, что они (исключая колониальные формы) соответствуют в структурном отношении уровню одной клетки, а в отношении физиологическом тАФ полВнноценной особи. В связи с этим одной из черт клеток части простейших является наличие в цитоплазме миВнниатюрных образований, выполняющих на клеточВнном уровне функции жизВнненно важных органов, апВнпаратов и систем органов многоклеточного организВнма. Таковы (например, у инфузорий) цитостом, цитофарингс и порошица,
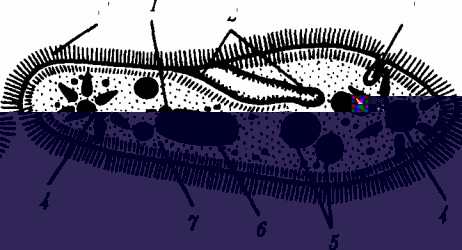
Структурная организация одноклеточВнного организма (инфузория):
/тАФ генеративное ядро,
2тАФ цитостом с цитофарингсом,
3тАФпорошица,
4тАФсократительные вакуоли,
5тАФ пищеварительные вакуоли,
6тАФ вегетативное ядро,
7тАФ гиалоплазма,
8тАФреснички
аналогичные пищеварительной системе, и сократительные вакуоли, аналогичные выделительной системе.
В традиционном изложении клетку растительного или живоВнтного организма описывают как объект, отграниченный оболочкой, в котором выделяют ядро и цитоплазму. В ядре наряду с оболочкой и ядерным соком обнаруживаются ядрышко и хроматин. ЦитоплазВнма представлена ее основным веществом (матриксом, гиалоплазмой), в котором распределены включения и органеллы.
Принцип компартментации. Биологическая мембрана
Компартментация объема клетки с помощью мембран:
/тАФядро,
2тАФшероховатая цитоплазматическая сеть,
3тАФмитохондрия,
4тАФтранспортный цитоплазматический пузырек,
5тАФлизосома,
6тАФпластинчатый компВнлекс,
7тАФ гранула секрета
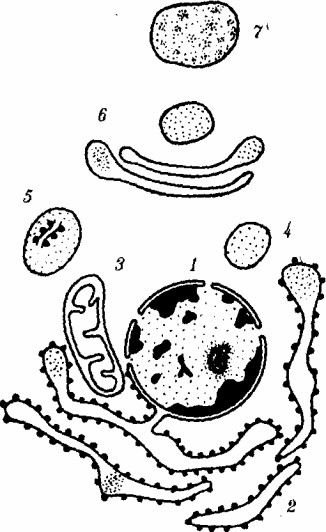
Высокая упорядоченность внутреннего содержимого эукариотической клетки достигается путем компартментации ее объема тАФ подразделения на ВлячейкиВ», отличающиеся деталями химического (ферментного) состава. Компартментация способствует пространственному разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой (лизосома) или ее частью (пространство, отграниченное внутренней мембраной митохондрии).
Молекулярная организация биологиВнческой мембраны:
IтАФ бимолекулярный слой липидов,
2тАФ белки
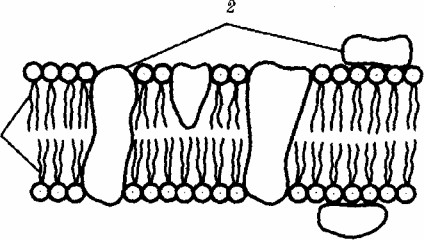
Важная роль в осуществлении компартментации принадлежит биологическим мембранам. Они выполняют ряд функций: отграниВнчивающую (барьерную), регуляции и обеспечения избирательной проницаемости веществ, образования поверхностей раздела между водной (гидрофильной) и неводной (гидрофобной) фазами с разВнмещением на этих поверхностях ферментных комплексов. БлагодаВнря присутствию липидов (жировых веществ) мембраны образуют гидрофобную внутриклеточную фазу как компартмент для химичеВнских реакций в неводной среде. Молекулярный состав мембран, набор соединений и ионов, размещающихся на их поверхностях, различаются от структуры к структуре. Этим достигается функциоВннальная специализация мембран клетки. Включение в мембрану клетки молекул рецепторов делает ее восприимчивой к биологичеВнски активным соединениям, например гормонам.
Предложено несколько схем взаимоотношения в мембране осВнновных химических компонентов тАФ белков и липидов, а также веществ, размещаемых на мембранной поверхности. В настоящее время большей популярностью пользуется точка зрения, согласно которой мембрана составлена из бимолекулярного слоя липидов. ГидВнрофобные участки их молекул повернуты друг к другу, а гидрофильВнные находятся на поверхности слоя. Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхностях.
Благодаря компартментации клеточного объема в эукариотической клетке наблюдается разделение функций между разными структурами. Одновременно различные структуры закономерно взаВнимодействуют друг с другом.
Транспорт через мембраны Жизненно важен по ряду причин. Он должен обесВнпечить поддержание в клетке соответствующего рН и надлежащей ионной концентрации, необходимых для эффективной работы клеточных ферментов; он оставляет питательные вещества, которые служат источником энергии, а также ВлсырьемВ» для образования клеточных компонентов; от него зависят выведение из клетки токсичных отходов, секреция различных полезных веществ и, наконец, создание ионных градиентов, необходимых для нервной и мышечной активности. Мы обсудим здесь трансВнпорт веществ через плазматическую мембрану, отВнметив, что аналогичный характер носит и транспорт через мембраны клеточных органелл. Существует четыре основных механизма для поступления веВнществ в клетку или выхода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, т. е. не требуют затрат энергии; два посВнледних - активные процессы, связанные с потреблеВннием энергии.
Диффузия
Газы, например кислород, потребляемый клетками при дыхании, и образующаяся в процессе дыхания СО2, в растворе быстро диффундируют через мемВнбраны, перемещаясь по диффузионному градиенту, т. е. из области с высокой концентрацией в область с низкой концентрацией. Ионы и малые полярные молекулы, такие, как глюкоза, аминокислоты, жирВнные кислоты и глицерол, обычно диффундируют через мембраны медленно. Гораздо более быстро проходят через мембраны незаряженные и жироВнрастворимые (липофильные) молекулы, о чем мы уже говорили выше.
Модификацией этого механизма является так наВнзываемая облегченная диффузия, при которой веВнществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть особый канал, пропускающий вещества только одного определенного типа. Примером такого пеВнремещения служит поступление глюкозы в эритВнроциты; оно не нарушается ингибиторами дыхания и, следовательно, не является активным процессом.
Осмос
Диффузия воды через полупроницаемые мембраны называется осмосом .
Активный транспорт - это сопряженный с потребВнлением энергии перенос молекул или ионов через мембрану против градиента концентрации. Энергия требуется потому, что вещество должно двигаться, вопреки своему естественному стремлению диффунВндировать в противоположном направлении. ДвиВнжение это обычно однонаправленное, тогда как диффузия обратима.
Для ионов направление диффузии определяется двумя факторами: один из этих факторов - концентВнрация, а другой - электрический заряд. Ионы обычВнно диффундируют из области с высокой их конВнцентрацией в область с низкой концентрацией. КроВнме того, они обычно притягиваются областью с противоположным зарядом и отталкиваются обВнластью с одноименным зарядом. Поэтому мы говоВнрим, что они движутся по электрохимическим граВндиентам, в которых объединяется эффект электриВнческого и концентрационного градиентов. Строго говоря, активный транспорт ионов - это их переВнмещение против электрохимического градиента.
Показано, что в клетках между двумя сторонами плазматической мембраны поддерживается разВнность потенциалов, иными словами, электрический заряд, и что почти во всех изученных клетках внутВнреннее содержимое клетки заряжено отрицательно по отношению к внешней среде. Поэтому катионы (положительно заряженные ионы) обычно стремятВнся в клетку, тогда как анионы клеткой отталкиваВнются. Однако их относительные концентрации внутВнри и вне клетки также играют роль, т.е. и от концентраций зависит, в каком направлении в дейстВнвительности диффундируют ионы.
Во внеклеточных и внутриклеточных жидкостях из ионов преобладают ионы натрия (Nа+), ионы калия (К+ и хлорид - ионы (С1-).
Ионный состав в клетках обоих этих типов резко отличается от состава окруВнжающего их наружного раствора. У них, например, как и у большинства клеток, концентрация калия внутри значительно выше, чем снаружи. Другая характерная особенность заключается в том, что внутриклеточная концентрация калия превышает концентрацию натрия.
Если каким-либо специфическим воздействием, например, с помощью цианида, подавить дыхание эритроцитов, то их ионный состав начнет постеВнпенно меняться и в конце концов сравняется с ионным составом плазмы крови. Это показывает, что данные ионы могут пассивно диффундировать через плазматическую мембрану эритроцитов, но что в норме за счет энергии, поставляемой процесВнсом дыхания, идет их активный транспорт, благоВндаря которому и поддерживаются концентрации. В клетках двух типов натрий активно выкачиВнвается из клетки, а калий активно накачивается в нее. Путем расчета можно показать, что реальный поток хлорид - ионов из плазмы в эритроциты отВнсутствует, несмотря на их, более высокую концентВнрацию в плазме крови. Объясняется это тем, что содержимое клетки отталкивает хлорид - ионы, посВнкольку оно заряжено отрицательно по отношению к внешней среде; иначе говоря, движение этих ионов определяется электрохимическим градиентом, что справедливо для всех клеток.
Сравнительно недавно выяснилось, что у большей части клеток в плазматической мембране действует натриевый насос, активно выкачивающий натрий из клетки. Обычно, хотя и не всегда, натриевый насос сопряжен с калиевым насосом, активно поглощаюВнщим ионы калия из внешней среды и переносящим их в клетку. Такой объединенный насос называют натрий - калиевым насосом (Nа+, К+ - насос).
Поскольку этот насос имеется в большинстве клеток и выполняет в них ряд важных функций, он представляет собой хороший пример механизма активного транспорта.
Nа+, К+-насос изучен в животных клетках и установлено, что его Влприводит в движениеВ» АТФ. О его физиологическом значении свидетельствует тот факт, что более трети АТФ, потребляемого животной клеткой в состоянии покоя, расходуется на перекачивание натрия и калия. Это необходимо для сохранения клеточного объема (осморегуляция), для поддержания электрической активности в нервВнных и мышечных клетках и, наконец, для активного транспорта некоторых других веществ, например Сахаров и аминокислот. Высокие концентрации каВнлия требуются также для белкового синтеза, глико-лиза, фотосинтеза и для некоторых других жизненно 3 важных процессов.
Насос-это особый белок, локализующийся в мембране таким образом, что он пронизывает всю ее толщу. С внутренней стороны мембраны к нему поступают натрий и АТФ, а с наружной тАУ калий.
Перенос натрия и калия через мембрану совершаетВнся, как полагают, в результате конформационных изменений, которые претерпевает этот белок. Белок действует и как АТФаза, катализируя гидролиз АТФ с высвобождением энергии, которая и привоВндит в движение насос. Обратите внимание, что на каждые два поглощенных иона калия из клетки выводится три иона натрия. ВследстВнвие этого содержимое клетки становится более отВнрицательным, по отношению к внешней среде, а между двумя сторонами мембраны возникает разВнность потенциалов.
Выкачиваемый из клетки натрий обычно пассивно диффундирует обратно в клетку. Однако мембрана мало проницаема для натрия, и потому эта диффуВнзия в обратном направлении происходит очень медВнленно. Для ионов калия мембраны приблизительно в 100 раз более проницаемы, чем для натрия; соотВнветственно и диффундирует калий гораздо быстрее.
Активный транспорт осуществляется всеми клетВнками, но в некоторых физиологических процессах он играет особо важную роль. Именно так обстоит дело в клетках эпителия, выстилающего кишечник и активный транспорт в кишечнике. Всасываясь в тонВнком кишечнике, продукты переваривания пищи должны пройти через клетки эпителия, выстилаюВнщего стенку кишки. Затем глюкоза, аминокислоты и соли через клетки, образующие стенки кровеносных сосудов, поступают в кровь и доставляются кровью в печень. Вскоре после приема пищи концентрация продуктов ее переваривания достигает в кишечнике довольно высокого уровня, так что всасывание в какой-то мере является и результатом диффузии. Однако диффузия происходит здесь очень медленно, и ее должен дополнять активный транспорт. Активный транспорт соВнпряжен с работой Nа+, К+ - насоса.
Натрий, выкачиваемый из клетки натрий - калиеВнвым насосом, стремится диффундировать обратно в клетку. В мембране находится белок, которому для выполнения его функции требуются натрий и глюВнкоза. Они транспортируются в клетку вместе пасВнсивно. Таким образом, натрий ВлтянетВ» глюкозу вмеВнсте с собой в клетку. Активный транспорт аминоВнкислот совершается при участии аналогичного белВнкового Влнатрий - аминокислотногоВ» переносчика; активной частью этого процесса является выкачиВнвание натрия обратно, наружу. При отсутствии градиента концентрации натрия оба эти переносчика тоже могут работать, если только наружная концентрация глюкозы или аминокислот превышает их внутреннюю концентрацию, т. е. в таких случаях имеет место облегченная диффузия.
Активный транспорт в нервных и мышечных клетках.
В нервных и мышечных клетках натрий - калиевый насос обеспечивает возникновение в плазматической мембране разности потенциалов, называемой поВнтенциалом покоя. В мембранах саркоплазматического ретикулума мышечных клеток действует насос, анаВнлогичный Nа+, К+ - насосу; в этом случае за счет энергии АТФ в саркоплазматический ретикулум активно накачивается кальций.
В почках также имеет место активный транспорт: из проксимальных изВнвитых канальцев почки активно транВнспортируются натрий и глюкоза, а в корковом веществе почки - натрий.
Эндоцитоз и экзоцитоз
Эндоцитоз и экзоцитоз - это два активных процесса, посредством которых различные материалы транВнспортируются через мембрану либо в клетки (эндоцитоз), либо из клеток (экзоцитоз).
При эндоцитозе плазматическая мембрана обраВнзует впячивания или выросты, которые затем, отшнуровываясь, превращаются в пузырьки или ваВнкуоли. Различают два типа эндоцитоза:
1. Фагоцитоз-поглощение твердых частиц. СпеВнциализированные клетки, осуществляющие фагоцитоз, называются фагоцитами; эту функцию выполВнняют, например, некоторые виды лейкоцитов. МемВнбранный мешочек, обволакивающий поглощаемую частицу, называют фагоцитозной вакуолью.
2. Пиноцитоз - поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Часто при этом образуются очень мелкие пузырьки. В таком случае говорят о микропиноцитозе и пузырьки называют микропиноцитозными.
Пиноцитоз характерен для амебоидных простейВнших и для многих других (часто амебоидных) клеВнток, таких, как лейкоциты, клетки зародыша, клетки печени и некоторые клетки почек, участвующие в водно-солевом обмене. Удается наблюдать пиноцитоз также и в клетках растений.
Экзоцитоз - процесс, обратный эндоцитозу. ТаВнким способом различные материалы выводятся из клеток: из пищеварительных вакуолей удаляются оставшиеся непереваренными плотные частицы, а из секреторных клеток путем Влпиноцитоза наоборотВ» выводится их жидкий секрет.
Все процессы жизнедеятельности клеток происходят с затратой энергии, которая поступает извне и преобразовывается специальными органалами клетки: в растительной клетке тАУ это хлоропласт, в животной тАУ митахондрии. Процессы преобразования энергии называется фотосинтез (растительная клетка), синтез АТФ тАУ животная клетка.
Хлоропласты
У эукариот фотосинтез происходит в особых органеллах, называемых хлоропластами. Хлоропласты рассеяны в цитоплазме, их число варьирует от одного примерно до ста. У высших растений Хлоропласты на срезе обычно имеют двояковыпуклую форму, а при взгляде сверху выВнглядят округлыми. Диаметр хлоропластов около 3-10 мкм (в среднем 5 мкм), так что они хорошо видны в световой микроскоп. У водорослей форма хлоропластов более разнообразна.
Хлоропласты образуются из небольших недифВнференцированных телец, называемых иропластидами; такие тельца имеются в растущих частях растеВнния (в клетках меристемы), они окружены двойной мембраной - будущей оболочкой хлоропласта. В хлоропластах всегда содержатся хлорофилл и друВнгие фотосинтетические пигменты, локализованные в системе мембран, которые погружены в основное вещество хлоропластастрому.
Детали строения хлоропластов выявляются с помощью электронного микроскопа. Мембранная система тАУ это то место, где протекают световые реакции. Темновые реакции фотосинтеза происходят в строме.
Биохимия фотосинтеза
Процесс фотосинтеза обычно описывают уравнеВннием:
Энергия света
6CO2 + 6Н2О тАФтАФтАФтАФ> С6Н12О6 + 6O2.
Хлорофилл
Им удобно пользоваться, когда надо показать, что образуется одна молекула сахара, но при этом не следует забывать, что это всего лишь суммарное отображение многих событий. Несколько лучший вариант:
Энергия света
СО2 + Н2О тАФтАФтАФтАФтАФ> [СН2О] + О2.
Хлорофилл
Такого соединения, как СН2О, не существует, но эта формула отражает состав углевода.
Источник кислорода
Глядя на приведенное выше уравнение, химик сразу же задумается о том, к какому типу относится эта реакция, а ответить на этот вопрос нельзя, если не знать, из чего - из двуокиси углерода или воды - обВнразуется выделяющийся кислород. Казалось бы, ответ ясен: из двуокиси углерода; в таком случае осталось бы только присоединить углерод к воде, и получился бы углевод. Но прямо ответить на этот вопрос удалось только после того, как в 40-е годы в биологических исследованиях начали применять.
Массовое число обычного изотопа кислорода равВнно 16, поэтому его обозначают lб0 (8 протонов, 8 нейтронов). А один из редких изотопов имеет масВнсовое число 18 (18О). Это стабильный изотоп, но его можно обнаружить благодаря его несколько больВншей массе. Для этого используют масс-спектроВнметр - очень важный аналитический прибор, способВнный улавливать разницу между массами отдельных атомов и молекул. В 1941 г. был поставлен экспериВнмент, результаты которого можно выразить слеВндующим образом:
СО2 + Н218О -> [СН2О] + 18O2.
Так было установлено, что источником кислорода служит вода. Из уравнения видно, что из каждой молекулы воды выделяется один атом кислорода. В сбалансированном виде уравнение должно выгляВндеть так:
Энергия света
СО2 + 2H218O тАФтАФтАФтАФ> [СН2О] + 18О2 + Н2О.
Хлорофилл
Это самое точное итоговое уравнение фотосинтеВнза; к тому же из него дополнительно вытекает, что вода в процессе фотосинтеза не только испольВнзуется, но и образуется. Рассмотренный выше экспеВнримент косвенно подтверждал полученные примерВнно в это же время данные Ван-Нила о том, что фотосинтезирующие бактерии совсем не выделяют кислорода, хотя и используют СО2. Ван-Нил приВншел к выводу, что всем фотосинтезирующим оргаВннизмам необходим источник водорода; у растений это вода, причем выделяется кислород; а, например, у серобактерий это сероводород, и вместо кислорода выделяется сера:
Энергия света
CO2 + 2H2S тАФтАФтАФтАФтАФтАФ> [СН2О] + 2S + Н2О.
Хлорофилл
Это уравнение для серобактерий полностью аналоВнгично уравнению для растений.
Упомянутые эксперименты позволили глубже поВннять природу фотосинтеза. Они показали, что фотоВнсинтез включает две стадии, первая из которых состоит в получении водорода. У растений водород получается путем расщепления воды на кислород и водород; для этого расщепления нужна энергия, которую и дает свет (отсюда и сам процесс стали называть фотолизом (греч. photos-свет, lysis-расВнщепление). Кислород выделяется как ненужный поВнбочный продукт. Во второй стадии водород соедиВнняется с СO2 и образуется углевод. Присоединение водорода - это один из примеров химической реакВнции, называемой восстановлением.
Тот факт, что фотосинтез является двухстадийВнным процессом, был впервые установлен в 20-е-30-е годы. Для первой стадии характерны так назыВнваемые световые реакции, для которых нужен свет. На второй стадии свет не нужен, и поэтому соотВнветствующие реакции, хотя они тоже происходят на свету, назвали темповыми реакциями. Сейчас выясВннено, что это два отдельных набора реакций, котоВнрые к тому же разделены и в пространстве: световые реакции происходят в мембранах хлоропластов, а темновые - в их строме.
Когда было установлено, что фотосинтез складыВнвается из световых реакций и следующих за ними темновых реакций, к концу 50-х годов осталось только выяснить, что же это за реакции.
Световые реакции
В 1958 г. Арнон и его сотрудники показали, что на свету изолированные хлоропласты могут синтезироВнвать АТФ из АДФ и фосфата (фосфорилирование), восстанавливать НАДФ до НАДФ-Н2 и выделять кислород.
Арнон показал также, что СО2 можно восстаноВнвить до углевода даже в темноте, при условии что в среде присутствуют АТФ и НАДФ тАв Н2. Это позволяло думать, что роль световых реакций состоит лишь в образовании АТФ и НАДФ-Нд. Арнон обратил внимание на сходство этого процесВнса с дыханием, при котором тоже происходит фосВнфорилирование АДФ. Для фосфорилирования нужВнна энергия. При дыхании энергия высвобождается в результате окисления питательных веществ пищи (чаще всего глюкозы), и потому этот процесс назыВнвают окислительным фосфорилированием. При фотосинтезе источником энергии служит свет, и соответствующий процесс назвали фотофосфорили-рованием. Таким образом, окислительное фосфориВнлирование - это превращение АДФ и Фн в АТФ за счет химической энергии, получаемой из пищи в процессе дыхания, а фотофосфорилирование-это такое же превращение с использованием энергии света в процессе фотосинтеза (Фн - неорганический фосфат).
Арнон совершенно верно предсказал, что фото-фосфорилирование, как и окислительное фосфориВнлирование, должно быть сопряжено с переносом электронов в мембранах. Перенос электронов - это основа для понимания как фотосинтеза, так и дыВнхания.
Процесс преобразования энергии в животных клетках происходит с помощью митахондрий.
Митохондрии
Митохондрии имеются во всех эукариотических клетках. Эти органеллы - главное место аэробной дыхательной активности клетки. Впервые наблюдал Митохондрии в виде гранул в мышечных клетках Кёлликер в 1850 г. Позднее, в 1898 г., Михаэлис показал, что они играют важную роль в дыхании: в его опытах митохондрии вызывали изменеВнние цвета окислительно-восстановительных индикаВнторов.
Число митохондрии в клетке очень непостоянно; оно зависит от вида организма и от природы клетВнки. В клетках, в которых потребность в энергии велика, содержится много митохондрии (в одной печеночной клетке, например, их может быть около 1000). В менее активных клетках митохондрии гоВнраздо меньше. Чрезвычайно сильно варьируют такВнже размеры и форма митохондрии. Митохондрии могут быть спиральными, округлыми, вытянутыми, чашевидными и даже разветвленными; в более активных клетках они обычно крупнее. Длина митоВнхондрии колеблется в пределах 1,5-10 мкм, а шиВнрина - в пределах 0,25-1,00 мкм.
Митохондрии способны изменять свою форму, а некоторые могут также перемещаться в особо активные участки клетки. Такое перемещение (коВнторому способствует ток цитоплазмы) позволяет клетке сосредоточивать большее число митохондВнрии в тех местах, где выше потребность в АТФ. В других случаях положение митохондрии более постоянно (как, например, в летательных мышцах насекомых).
Синтез АТФ
Изучением этого вопроса активно занимаются больше 30 лет однако четкого представления о механиз мах синтеза АТФ нас пока еще нет. В последнее время усиленно обсуждаются главным образом две гипотезы: гипотеза химического сопряжения и хемиосмотическая гипотеза.
Гипотеза химического сопряжения
Согласно этой гипотезе, синтез АТФ сопряжен с переносом электронов при посредстве одного или нескольких ВлвысокоэнергетическихВ» промежуточных Энергия, высвобождаемая при переносе в окислительно-восстановительных реакциях дыхательной цепи, используется в нескольких ее звеньях для образования высокоэнергетической связи в одном из таких продуктов. Затем при фосфорилировании АДФ эта энергия переходит к высокоэнергетической связи АТФ. До сих пор, однако, обнаружить подобные промежуВнточные продукты не удалось, и до тех пор, пока их существование не подтвердится, эту гипотезу нельзя считать убедительной.
Хемиосмотическая гипотеза
Большее признание завоевала гипотеза, выдвинутая Митчеллом в 1961 г. Он полагал, что синтез АТФ находится в тесной зависимости от того, каким образом электроны и протоны передаются по дыхаВнтельной цепи. Ниже перечислены условия, соблюдеВнния которых требует эта гипотеза.
1. Внутренняя митохондриальная мембрана должна быть интактна и непроницаема для протонов (ионов водорода), направляющихся снаружи внутрь.
2. В результате активности дыхательной (электрон-транспортной) цепи ионы водорода поступают в нее изнутри, из матрикса, а освобождаются на наружной стороне мембраны.
3. Движение ионов водорода, направленное изнутри наружу, приводит к их накоплению, вследствие чего между двумя сторонами митохондриальной мембраны возникает градиент рН. Это может быть связано с тем, что ферменты, принимающие и отдающие ионы водорода, расположены в мембране определенным образом и поэтому могут принимать ионы водорода только изнутри и отдавать их только наружу.
4. Сам по себе градиент рН не мог бы поддерживаться, так как ионы водорода диффундировали бы обратно в митохондрию. Поддержание такого градиента требует затраты энергии. Предполагается, что энергию поставляет перенос электронов по электронтранспортной (дыхательной) цепи.
5. Эта энергия используется затем для синтеза АТФ. Синтез АТФ, таким образом, поддерживается наличием градиента рН.
6. АТФ образуется в результате фосфорилирования АДФ:
АДФ + ФН = АТФ + Н2O.
По закону действующих масс удаление воды должно ускорять реакцию, идущую слева направо, т. е. благоприятствовать образованию АТФ. Согласно теории Митчелла, фермент, ответственный за образование воды при синтезе АТФ, ориентирован в мембране таким образом, освобождаются с внутренней стороны мембраны, где значение рН выше (т.е. концентрация Н'1'меньше), а гидроксильные ионы (ОН")-с наружной стороны, где рН ниже (т.е. концентрация Н4" больше). Таким образом, вода, образующаяся при синтезе АТФ, быстро удаляется, и это стимулирует синтез,
Из гипотезы Митчелла ясно, почему мембрана должна быть интактной (в моменты, ответственные за прохождение ионов водорода и за образование воды, так что любое изменение структуры мембраны неминуемо сказалось бы также на расположении ферментов и на их структуре). Объяснимо и требование непроницаемости мембраны для ионов водорода (в направлении снаружи внутрь): в основе гипотезы лежит представление о трансмембранном градиенте рН, атакой градиент не мог бы поддерживаться, если бы мембрана была полностью проницаемой. Следует отметить также, что гипотеза обходится без каких бы то ни было промежуточных продуктов в процессе синтеза АТФ.
Список использованной литературы.
- Зегбуш П. ВлМолекулярная и клеточная биологияВ»
- Грин Н., Стаут У., Тейлор Д. ВлБиологияВ» 1, 2 том.
Вместе с этим смотрят:
Адсорбенты и ионные обменники в процессах очистки природных и сточных водАзот 2Алкалоиды и история их открытияАлюминий