Волновой генетический код
СОДЕРЖАНИЕ
Актуальность темы
Пересмотр модели генетического кода
Расширение модели волнового генетического кодирования
Экспериментальные подтверждения существования волновых генов
Теоретические модели волновых генов
Математическое моделирование солитонов на ДНК
Экспериментальные доказательства солитонообразования на информационных биополимерах тАЬin vitroтАЭ
Запись ИК-лазерного сигнала на уровне нелинейной динамики ДНК
О возможности создания лазера на информационных биомакромолекулах
Теоретическое исследование возможности создания лазера на Фрелиховских модах
Антенная модель, физико-математический формализм
Конверсия эпигеносигналов в электромагнитных солитонных структурах, их транспозиция в геном биосистем-акцепторов
Генератор пакетов уединенных волн (солитонов) в форме возврата Ферми-Паста-Улама
Единство фрактальной структуры ДНК-тАЬтекстовтАЭ и текстов на естественных языках
О возможности создания биокомпьютера на генетических структурах
Явление перехода света в радиоволны применительно к биосистемам и биокомпьютерам
Основные результаты
Литература
АКТУАЛЬНОСТЬ ТЕМЫ
В последние десятилетия начали постепенно выявляться некоторые кризисные явления в молекулярной биологии и биологии развития. После открытия структуры ДНК и детального рассмотрения участия этой молекулы в генетических процессах основная проблема феномена жизни - механизмы ее воспроизведения - осталась в своей сути не раскрытой. Отсюда ограниченность арсенала технических и биотехнических средств управления ростом и развитием биосистем. Наметился явный разрыв между микроструктурой генетического кода и макроструктурой биосистем. Даже открытие гомеобоксов ДНК, кардинально влияющих на формообразовательные акты эмбриогенеза, лишь более ярко высветили то, о чем в свое время предупреждал А. Г. Гурвич, считая, что нагрузка на гены слишком высока, и поэтому необходимо ввести понятие биологического поля, как пространственно-временной разметочной структуры, биологического поля, тАЬ.. свойства которого.. формально заимствованы.. из физических представленийтАЭ. Таким элементарным полем, по Гурвичу, будет являться тАЬ.. поле эквивалента хромосомы..тАЭ. И далее: тАЬ.. хроматин сохраняет свою тАЬактивностьтАЭ, т. е. является носителем активного поля, только в неравновесном состояниитАЭ. Здесь видно предвидение лазерной накачки хромосом как типично неравновесного состояния, полученного нами in vitro спустя 50 лет для ДНК и нуклеогистона [18].
Близкие идеи мы видим и у А. А. Любищева в его работе 1925 г. тАЬО природе наследственных факторовтАЭ . Он пишет: тАЬГены не являются ни живыми существами, ни кусками хромосомы, ни молекулами автокаталитических ферментов, ни радикалами, ни физической структурой, ни силой, вызываемой материальным носителем; мы должны признать ген как нематериальную субстанцию (выделено П.Г.), подобную эмбриональному полю Гурвича, но потенциальнуютАЭ (выделено П.Г.). И далее: тАЬ.. взаимодействие наследственности и хромосом подобно отношению материи и памяти по Бергсону.. Гены в генотипе образуют не мозаику, а гармоническое единство, подобное хорутАЭ (выделено П.Г.). Через 3 года другой наш русский научный предшественник - В. Н. Беклемишев приходит к тем же идеям в своей работе, выполненной также в Перми, тАЬМетодология систематикитАЭ. Чтобы приблизиться к реальному морфопроцессу (эмбриогенезу), - необходимо принять идею музыки и речи как неких моделей векторов генетических актов. И в музыке, и в речи тАЬ.. существуют тАЬанатомическиетАЭ свойства (возможны признаки стадий) - высота, интенсивность звука, обертоны и пр., а следовательно, возможно и описание отдельных стадий, и формальное описание процесса в его целостности.. Музыкальная вещь аналогична морфопроцессу гораздо глубже, чем с первого взгляда кажется. Между обоими процессами бросается в глаза различие: изменения в развивающемся теле накапливаются, изменения в потоке музыки сменяются бесследно. Но истинным субъектом развития в музыке является эстетическое впечатление; оно растет и развивается под влиянием процесса звучания. Это морфопроцесс сложного духовного организма.. Что является аналогом этого последнего в животных и растительных организмах? Не поток ли формативных раздражений, регулируемый индивидуальностью целого и направляющий морфогенез частей?тАЭ. Идеи русских биологов Гурвича, Любищева и Бекле-мишева - гигантское интеллектуальное достижение, намного опередившее свое время. Суть их мыслей в триаде:
1. Гены дуалистичны - они вещество и поле одновременно.
2. Полевые эквиваленты хромосом размечают пространство-время организма и тем самым управляют развитием биосистем.
3. Гены обладают эстетически-образной и речевой регуляторными функциями.
Современные молекулярная биология, генетика и эмбриология, проделав большой путь развития, завершили определенный виток в понимании сущности жизни. Оно было сугубо материалистичным, точнее, вещественным. Гены в этом смысле - только вещество. И когда это вещество - ДНК - детально изучили, открыв так называемый генетический код, то оказалось, что этого явно мало. Ключевая проблема биологии - преемственность поколений, наследственность, эмбриогенез - не раскрыта, более того, в тупике, правда более высокого ранга. Ситуация сейчас напоминает положение в классической физике начала xx века, когда с открытием элементарных частиц материи вещество вроде бы исчезло, осталось нечто, которое назвали неопределенным термином "энергия". Вот и в биологии, чем точнее понимание ДНК по части повсеместно принятой центральной догмы ДНК-РНК-Белок, тем дальше мы уходим от стратегии генома в построении биосистемы. Но если физика с достоинством приняла как реальность парадоксы: тАЬздесь и там одновременнотАЭ, тАЬволна и частица совмещенытАЭ, тАЬэлектрон резонирует со всей ВселеннойтАЭ, тАЬвакуум - ничто, но он порождает всетАЭ и т. д., то биологии только предстоит пройти сходный путь (Дао Биологии), и он будет гораздо тяжелее. Фактически мы уже вышли на него, вовремя вспомнив мысли Гурвича, Любищева и Беклемишева.
Наша задача состояла и состоит в том, чтобы развить их концептуальную триаду в контексте современных знаний и полученных нами результатов по теории и практике разработки и использования механизмов и технических средств волновой коррекции биосистем.
Цель работы: показать возможность дуалистической трактовки работы генома эукариот на уровнях вещества и поля в рамках физико-математических моделей, соединяющих формализм явления солито-нообразования в ДНК на примере явления возврата Ферми-Паста-Улама, а также голографической и иной памяти хромосомного континуума как биокомпьютера, связанного с волей Творца.
Показать возможность обычных и тАЬаномальныхтАЭ режимов работы генома эукариот с использованием волновых образно-знаковых матриц, а также эндогенной и экзогенной (зависящей от Творца) семиотико-лингвистической компоненты.
Найти экспериментальные доказательства правильности пред-лагаемой теории волновых образных и образно-лингвистических матриц генома как структур стратегического управления метаболизмом высших биосистем.
Практическая направленность настоящего исследования:
в теоретико-экспериментальном обосновании феномена свертки, транспозиции и резонансного введения супергенетической информации от биодонора к биоакцептору; при этом передаваемые эпигеносигналы могут существовать как акусто-электромагнитные солитоны в рамках явления возврата Ферми-Паста-Улама и входить в семантические знаковые ряды генетических структур, также реализующиеся в форме солитонных возбуждений;
в теоретико-экспериментальном обосновании единства фрактальной структуры человеческой речи и текстовых структур генетических молекул ДНК и РНК; это положение заложило основу для разработки начальных основ методологии введения регуляторных квази-вербальных структур в виде модулированных электромагнитных солитонов непосредственно в геном растений;
в теоретико-экспериментальном обосновании возможности создания искусственных ДНК-логических устройств (биокомпьютеров) с исполь-зованием волновых (голографических и иных) принципов памяти, сравнимой по механизмам и возможностям с генетической;
в создании технических средств мягкого регуляторного вхождения в неизвестные ранее семиотические ареалы генома высших биосистем с целью лечения, создания гибридов, продления жизни людей, форми-рования организма человека как гармоничной и устойчивой к неблагоприятным факторам структуры.
Ранее нами предложена гипотеза эпигенетической кодовой иерархии уровней организации хромосомной ДНК, рибосом и внеклеточных матриксов высших биосистем и участия их в синтезе волновых образных фрактальных построений, используемых высшими биосистемами для собственной самоорганизации [25]. Нелинейная динамика (акустика) и связанные с ней электромагнитные излучения указанных биоструктур in vivo не случайны, взаимно коррелированы, носят биознаковый (в частности, речеподобный) характер, изоморфно отображают структурно-функциональные состояния каждой из обменивающихся волновыми сигналами организменных клеточно-тканевых подсистем. В пространстве-времени организмов в эпигенетическом режиме происходит обмен информацией по физическим каналам нелинейных акустическими электромагнитных колебаний. при этом стратегической компонентой рассматриваемых волновых знаковых рядов является акустическое и электромагнитное излучение совокупного генетического материала (генома) биосистем. В настоящей работе выдвинутые положения развиваются как трактовка волновых состояний (собственных физических полей) организма и попытка понимания биологического смысла явления генерации внутри и межклеточных полевых сигналов в качестве основы волновой и, вслед за этим, вещественной самоорганизации живых систем.
ПЕРЕСМОТР МОДЕЛИ ГЕНЕТИЧЕСКОГО КОДА
В настоящее время создалась парадоксальная ситуация с моделью генетического кода - вершиной достижений молекулярной биологии 60-х годов. Точность кодирования последовательностей аминокислот белков в этой модели странным образом уживается с двойной вырожденностью предлагаемого тАЬкодатАЭ по линиям избытка транспортных РНК (тРНК) по сравнению с числом аминокислот и неоднозначного соответствия кодон-антикодон, когда только двум (а не трем) нуклеотидам триплетов иРНК необходимо точное спаривание c антикодоновой парой нуклеотидов тРНК, а по третьему нуклеотиду природой допускается неверное спаривание, так называемое тАЬвоблированиетАЭ (от англ. слова тАЬwobbleтАЭ- качание) по гипотезе Ф.Крика [4]. Это означает, что некоторые антикодоны могут тАЬузнаватьтАЭ более одного кодона в зависимости от того, какое основание находится в 1-м положении антикодона, соответствующем 3-му положению нуклеотида с учетом их антипараллельного комплементарного взаимодействия. тАЬУзнаваниетАЭ такого рода тАЬнеправильноетАЭ, если следовать парадигме генетического кода, поскольку возникают неканонические пары оснований тАЬАденин-ГуанинтАЭ, тАЬУрацил-ЦитозинтАЭ и другие с энергетически невыгодными водородными связями. тАЬКодтАЭ, особенно митохондриальный, становится настолько вырожденным, и логически следующий отсюда произвол включения аминокислот в пептидную цепь столь велик, что как бы исчезает само понятие генетического кодирования.
Процитируем высказывание из книги Альбертса, Уотсона и др. тАЬМолекулярная биология клеткитАЭ [20] (глава с характерным названием тАЬГеном митохондрий имеет ряд поразительных особенностейтАЭ): тАЬ..в митохондриях обычные правила спаривания кодонов с антикодонами соблюдаются менее строго, и многие молекулы тРНК способны узнавать любой из четырех нуклеотидов в третьей (неоднозначной) позициитАЭ. Вот эта тАЬменьшая строгостьтАЭ, как будто бы несовместимая с реально существующим метаболическим контролем порядка чередования амино-кислот в белках, заслуживает пристального внимания. тАЬМеньшая строгостьтАЭ не случайна, более того, она для чего-то нужна биосистемам.
Точность белкового синтеза эволюционно консервативна и высока, но может ли она достигаться такого рода тАЬтайнописьютАЭ, когда тАЬзнактАЭ (кодон) и тАЬобозначаемоетАЭ (аминокислота) не всегда изоморфны, не однозначны? Если придерживаться старой догмы генетического кода, логично думать, что две разные аминокислоты, шифруемые двумя одинаковыми (третий не важен) нуклеотидами кодонов иРНК, будут с равной вероятностью включаться в пептидную цепь, т.е. случайно. И таких парных неоднозначностей даже в немитохондриальном коде насчитывается шесть, если не считать еще две по стоповым кодонам (они же тАЬнонсенстАЭ или бессмысленные). Так что же, существует тАЬиндульгенция разрешениятАЭ частых и случайных замен аминокислот при синтезе белков? Однако, известно, что такие случайные замены в большинстве случаев имеют самые отрицательные последствия для организма (серповидная анемия, талассемии и т.д.). Налицо явное противоречие: нужна точность (однозначность) отношений тАЬзнак-обозначаемоетАЭ (кодон-аминокислота), а придуманный людьми код ее не обеспечивает.
Поэтому существующее и общепринятое представление о ключевых (знаковых) механизмах синтеза белков нуждается в дополнительном анализе. В связи с этим более подробно рассмотрим предложенные в 60-х годах принципы генетического кодирования. Как оценили перечисленные и очевидные странности ведущие авторы теории и экспериментов в этой области - Ф.Крик, М.Ниренберг и их последователи? Основной узел противоречий - неоднозначные соответствия (кодон-аминокислота) приведены в таблице:
НЕОДНОЗНАЧНЫЕ СООТВЕТСТВИЯ И СИНОНИМО-ОМОНИМИЧЕСКАЯ ДВУМЕРНОСТЬ ГЕНЕТИЧЕСКОГО КОДА
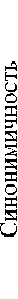
Омонимичность
Видно, что пары разных аминокислот шифруются одинаковыми значимыми дублетами кодоновых нуклеотидов (тАЬвоблирующиетАЭ мало значимые, по Крику [4], и вообще нечитаемые, по Лагерквисту [11], нуклеотиды смещены в индекс). В терминах лингвистики это явление носит название омонимия, когда одни и те же слова имеют разный смысл (например, русские слова тАЬлуктАЭ, тАЬкосатАЭ или английские тАЬboxтАЭ, тАЬringтАЭ и т.п.). С другой стороны, избыточные различающиеся кодоны, обозна-чающие одни и те же аминокислоты, уже давно рассматривают как синонимичные.
В отношении омонимии генетического кода высказывания в литературе нам не известны. Таким образом, если считать дуплетно-триплетные кодоны тАЬсловамитАЭ, то сам код является, кроме прочего, двумерным, то есть омонимо-синонимичным. По этим измерениям код распадается, как это видно из таблицы, в основном, на парные семейства, избыточно, но не однозначно, шифрующие разные аминокислоты. И только в двух случаях из шести омонимичные дублеты обозначают близкие по структуре и функции аминокислоты (аспарагиновая-глутаминовая и аспарагин-лизин). Следовательно, при неоднозначном (ошибочном) выборе аминокислот высока вероятность синтеза аномальных белков, если следовать логике общепринятой модели кода. Большинство этих сомнений и наметок на будущее в мягкой форме уже прозвучало в обобщающей статье Ф.Крика и М.Ниренберга тАЬГенетический кодтАЭ. Процитируем авторов дословно ввиду стратегической важности обсуждаемых принципов генетического кодирования:
с.133: тАЬбелок .. является как бы длинным предложением, записанным с помощью двадцати буквтАЭ. Вот одно из первых и плодотворных сравнений белков, а затем и ДНК, с текстами естественных языков, сравнений, повсеместно принятых на первых порах лишь как метафора, а затем развитых и формализованных нами в качестве квази-речевых образований [14,25,26,29]. В этой замечательной аналогии зачаток будущего выхода из плоского и тупикового понимания природы генов, предтеча понятия образных кодов (слово как образ), а это согласуется с идеями Гурвича, Любищева и Беклемишева, которые также видели в хромосомах потенциальные волновые образные и даже эстетические структуры в качестве организующих биосистему начал. А.А.Любищев еще в 1925 году высказал предположение, что гены образуют не мозаику, а гармоническое единство, подобное хору [47]. Вслед за ним в 1928г. В.Н.Беклемишев [21] развил это, хотя и афористично, однако, с огромным предвидением, на десятилетия опередившим аналогию Крика и Ниренберга о белках как тАЬпредложенияхтАЭ. Эмбриогенез он сравнил одновременно с музыкой и речью, в которых как и в дифференцирующихся тканях существуют тАЬанатомическиетАЭ свойства - признаки стадий: высота, интенсивность звука, обертоны и пр., и онтогенетические тАЬэмбриологическиетАЭ свойства - признаки хода процесса: ритм, мелодия и пр. Изменения в развивающемся теле накапливаются, а изменения в потоке музыки сменяются бесследно. Но истинным субъектом развития в музыке является эстетическое впечатление; оно растет и развивается под влиянием процесса звучания. Это морфопроцесс сложного духовного организма. Придя к этому, В.Н.Беклемишев спрашивает: что является аналогом этого последнего в животных и растительных организмах? Не поток ли формативных раздражений, регулируемый индивидуальностью целого и направляющий морфогенез частей?
Продолжим анализ основополагающей работы Крика и Ниренберга, постулирующей понятие генетического кода.
С.142 -143: тАЬ .. до сих пор все опытные данные хорошо согласовывались с общим предположением о том, что информация считывается тройками оснований, начиная с одного конца гена. Однако, мы получили бы те же результаты, если бы информация считывалась группами в четыре или даже более основанийтАЭ или тАЬ..группами, содержащими кратное трем число основанийтАЭ. Это положение почти забыто или не понято, но именно здесь видно сомнение, обязательно ли код триплетный. И не менее важно, что предугадано будущее понимание текстов ДНК и РНК как смысловых фрактальных образований, родственных естественным языкам, что продемонстрировано в наших исследованиях [25,26,29].
С.153: тАЬ .. одна аминокислота шифруется несколькими кодонами. Такой код называется вырожденным .. такого рода вырождение не говорит о какой-то неопределенности в построении молекулы белка .. оно лишь обозначает, что определенная аминокислота может быть направлена в соответствующее место цепи молекулы белка с помощью нескольких кодовых словтАЭ.
Авторы видят, что синонимия еще не нарушает однозначности кода.
С.153 -154: Но дальше следует тАЬ..однако, все же имеется одна реальная возможность появления неопределенности при синтезе белка. Эта неопределенность могла бы возникнуть, если бы одно кодовое слово соответствовало нескольким аминокислотам. До настоящего времени был отмечен только один случай такой неопределенности. Белок, син-тезируемый поли-U, состоит не только из лейцина, но и из фенилаланина, причем на каждую молекулу лейцина приходится 20-30 молекул фенилаланина. При отсутствии в растворе фенилаланина поли-U использует лейцин в количестве, равном половине обычно используемого количества фенилаланина. Молекулярное объяснение этой неопределенности неизвестнотАЭ. Это первая и четкая констатация логического несовершенства предлагаемой модели кодирования, ее противоречия фактам. Затем, сомнения еще более усиливаются.
С.155: тАЬнекоторые кодовые слова почти наверняка состоят из трех оснований. Однако, 18 из 20 аминокислот могут быть закодированы словами, содержащими только два различных основания. Если же код все-таки троичный, то возможно, что в некоторых случаях правильное кодирование будет иметь место при условии, что из трех оснований считывается только два. Возможно, что такое несовершенство случается более часто в синтетических РНК-полимерах, содержащих одно или два основания, чем в естественных РНК-посредниках, которые всегда состоят из смеси всех четырех оснований. Поэтому результаты, полученные с помощью искусственных РНК, свидетельствуют лишь о кодовых возможностях клетки..тАЭ
Явно просматривается неуверенность, что код только триплетный, он может быть и дуплетным, и тетраплетным и даже гетеромуль-типлетным. Нам же представляется, в развитии этих сомнений, что кодовые возможности клетки, хромосом, ДНК не исчерпываются знаковыми тройками нуклеотидов. Как речеподобные структуры, нуклеиновые кислоты в составе хроматина способны к образованию in vivo метаязыков методом фрактализации, и поэтому кодирование белкового континуума может проходить через крупные блоки, шифрующие не только порядок включения отдельных аминокислот в пептид, но и последовательность создания белковых доменов, субъединиц и даже структурно-функциональных ансамблей ферментов, например, дыхательной цепи. Фрактальность в данном случае может пониматься и так: ДНК, РНК и белки - это разноязыкие тексты и то, что было в одном масштабе тАЬфразойтАЭ или тАЬпредложениемтАЭ в другом, более крупном, будет тАЬсловомтАЭ. Если еще укрупнять - тАЬсловотАЭ превращается в тАЬбуквутАЭ. При более общем подходе можно рассматривать такие разномасштабные смысловые построения как знаки (иероглифы), являющиеся субстратом своего рода тАЬинформационного метаболизматАЭ клеток. Такой путь образования метаязыков свойствен математике. У нас нет оснований думать, что геном не пользуется этим тАЬматематическим приемомтАЭ в полной мере, строя все новые усложняющиеся семиотико-семантические ареалы с их постоянными переобозначениями на разных уровнях организации биосистемы в процессе ее развития. При этом роль основной массы синтезирующихся в организме белков заключается в реализации метаболических конструкций, неявно закодированных в ДНК и имеющих квази-вербальную составляющую. Биосистему можно рассматривать как совокупность таких конструкций, и это находит определенное подтверждение в работах [25,26,29]. Такой ход рассуждений хорошо соответствует представлениям В.В.Налимова, считающего все живое частью Семантической Вселенной [49]. Человек, в соответствии с такой логикой, есть многообразие текстов, грамматику и семантику которых мы хотим охватить единым, вероятностно задаваемым взглядом. В.В.Налимов полагает, что личность является самочитаемым текстом - текстом, способным самоизменять себя. Уменьшая масштаб рассмотрения человека как самоорганизующейся системы и учитывая фрактальность (переходящую иногда в голографичность) его хромосомного континуума, можно считать, что обратное отображение человека в его собственный геном, как и отображения любого организма в его хромосомы, носит изоморфный текстово-образный характер [25,29]. Предлагаемый способ рассуждений призван показать прием логического выхода из ограничений первичной модели генетического кода, остановившейся в фазе слабого понимания правил орфографии "записи" белковых тАЬсловтАЭ из аминокислотных "букв". Если же рассматривать идею фрактальности смысловых (текстовых) конструкций генома и принять их божественное начало, то подчеркнем, что эта идея восходит к VI в. и предложена Дионисием Ареопагитом в его труде тАЬО божественных именахтАЭ. Он говорит, что печать Божественности (читай слова) лежит на каждом из нас, и тАЬ..оттиски Печати имеют много общего с ее оригиналом: оригинал присутствует в каждом из отпечатков весь, и ни в одном из них - лишь какой-то своей частьютАЭ. Частичность Печати определяется свойствами воспринимающего материала - конкретной личности, т.е. потенциально в каждого привносится все, идущее свыше, но расслышать, увидеть и понять это все целиком не под силу никому.
Неспособность ранней концепции генетического кода быть непротиворечивой, казалось, должна была побудить к поиску новых идей. Вместо этого предпочтение было отдано анализу механизмов точности белкового синтеза, но без главного мотива этой точности - механизмов выбора однозначностей из кодирующих дуплетов-омонимов. Вот образец этих, в данном аспекте бесполезных, описаний и рассуждений, но необходимых нам для иллюстрации псевдологики в оценке главного в генокоде [20]: тАЬ .. точность белкового синтеза зависит от надежности двух адапторных механизмов: от связывания каждой аминокислоты с соответствующей молекулой тРНК и от спаривания кодонов в иРНК с антикодонами тРНК. Два механизма, действующие на этих этапах, совершенно различны. У многих аминоацил-тРНК-синтетаз имеется два отдельных активных центра: один ответственный за реакцию присоединения аминокислоты к тРНК, и другой, распознающий тАЬнеправильнуютАЭ аминокислоту и удаляющий ее путем гидролиза. Точность спаривания кодона с антикодоном обеспечивается более тонким механизмом тАЬкинетической коррекциитАЭ. После того как молекулы тРНК присоединят соответствующую аминокислоту, они образуют комплекс с особым белком, т.н. фактором элонгации (ФЭ,EF), который прочно связывается с аминоацильным концом молекулы тРНК и с молекулой GTP. Именно этот комплекс, а не свободная тРНК спаривается с надлежащим кодоном в молекуле иРНК. Связанный таким образом ФЭ обеспечивает возможность правильного спаривания антикодона с кодоном, но при этом препятствует включению данной аминокислоты в растущий пептид. Начальное узнавание кодона служит для ФЭ сигналом к гидролизу связанного с ним GTP до GDP+P, после чего ФЭ отделяется от рибосомы без тРНК и синтез белка продолжается. Благодаря ФЭ возникает короткий разрыв во времени между спариванием кодона с антикодоном и элонгацией пептида, что позволяет тРНК отделиться от рибосомы. тАЬНеправильнаятАЭ молекула тРНК образует в паре кодон - антикодон меньше водородных связей, чем правильная; поэтому она слабее удерживается на рибосоме и значит за данный промежуток времени имеет больше шансов отделитьсятАЭ.
Комментируя эту, важную для нас, длинную выдержку, можно сказать, что акцент в ней сделан на взаимном узнавании тРНК и аминокислот через посредство аминоацил-тРНК-синтетаз. Механизм его не ясен. Что касается точности узнавания кодоном антикодона, то она иллюзорна в силу тАЬвоблированиятАЭ третьего нуклеотида, что уже обсуждалось. Представляется, что выбор из дуплетных кодонов-омонимов реализуется по резонансно-волновым и контекстным (ассоциативным, голографическим) и так называемым "фоновым механизмам" (см. ниже). До сих пор они находились вне экспериментов и рассуждений, но в настоящее время необходимость в этом очевидна. Омонимичность (неоднозначность) кода может быть преодолена точно так же, как это происходит в естественных языках ,- путем помещения омонима, как части, в целое, т.е. в законченную фразу, контекст которой дешифрует омоним и присваивает ему единственное значение, создавая однозначность. Поэтому иРНК в качестве своего рода тАЬфразытАЭ или тАЬпредложениятАЭ должна работать в белковом синтезе как функциональное кодирующее целое, задающее последовательность аминокислот на уровне ассоциатов аминоацилированных тРНК, которые комплементарно взаимодействуют со всей молекулой иРНК. При этом роль А,Р-участков рибосомы, если они реальны, заключается в акцепции таких ассоциатов - предшественников белка с последующей энзиматической сшивкой аминокислот в пептидную цепь. В этом случае будет происходить контекстно-ориентированный однозначный подбор бывших омонимичных дуплет-кодонов. Можно предсказать в связи с этим, что взаимодействие аминоацилированных-тРНК с иРНК носит коллективный фазовый характер по типу реассоциации (тАЬотжигатАЭ) однотяжных ДНК при понижении температуры после тАЬплавлениятАЭ нативного полинуклеотида. Существуют ли экспериментальные данные, которые можно было бы трактовать в таком духе? Их немало и они сведены в обзорно-аналитическом исследовании [52]. Приведем некоторые из них.
Известно, что правильность узнавания молекулами тРНК терми-нирующих кодонов зависит от их контекстного окружения, в частности, от наличия за стоповым кодоном уридина и, кроме того, в работе убедительно показано следующее. Вставка строки из девяти редко используемых CUA-лейциновых кодонов после 13-го в составе 313 кодонов тестируемой мРНК сильно ингибируют их трансляцию без явного влияния на трансляцию других мРНК, содержащих CUA-кодоны. Напротив, строка из девяти часто используемых CUG-лейциновых кодонов в тех же позициях не имела выраженного эффекта на трансляцию. При этом ни редко, ни часто используемые кодоны не влияли на этот процесс, когда были введены после кодона 223 или 307. Дополнительные эксперименты продемонстрировали, что сильный позиционный эффект редко используемых кодонов не может быть объяснен различиями в стабильности иРНК или в степени строгости выбора соответствующих тРНК. Позиционный эффект становится понятным, считают авторы, если допустить, что транслируемые последовательности менее стабильны вблизи начала считывания: замедленность трансляции реализуется посредством малого использования кодонов, которые раньше следуют в сообщении, и это приводит к распаду продуктов трансляции, раньше чем осуществится полная трансляция. Как видим, для трактовки собственных экспериментов привлекаются громоздкие допущения о распаде продуктов трансляции, допущений, никак не следующих из их работы, и которые требуют специальных и тонких исследований. В этом смысле наша идея контекстных ориентаций в управлении синтезом белков проста, хотя экспериментально доказать ее непросто. Цитируемая работа хорошо высвечивает стратегическую линию влияния строго определенных и далеко расположенных от места образования пептидной связи кодоновых вставок в иРНК на включение или невключение конкретной аминокислоты в состав синтезируемого белка. Это именно дистантное влияние, но в цитируемой работе оно просто констатируется, оставаясь для исследователей непонятным и, видимо, поэтому даже не обсуждается. Таких работ становится все больше. В той, что мы обсуждаем, ссылаются, к примеру, на полдюжины аналогичных результатов, где трактовка в этом смысле также затруднена. Причиной этому является несовершенство общепринятой модели генетического кода. Это верно и потому, что имеются данные о существовании так называемого протяженного (swollen) антикодона [52]: во взаимодействии тРНК с иРНК в А-сайте рибосомы участвуют не три, а большее количество пар оснований. Это означает, что принятый пов-семестно постулат триплетности кода нарушается и здесь. Там же, в [52], приводятся результаты работы по взаимодействию тРНК-тРНК на рибосоме, и это соответствует нашей идее об ассоциате аминоацилированных тРНК как предшественнике белка. В [52] высказана мысль, что эффект действия контекста иРНК на однозначное включение аминокислот в пептид является отражением неких фундаментальных и пока плохо изученных закономерностей декодирования генетической информации в процессе белкового синтеза. В работе Ульфа Лагерквиста [11] тАЬwobbleтАЭ- гипотеза Крика получила расширенную трактовку и крайнее выражение, согласно которому нуклеотид в третьем положении кодона иРНК является лишним, бессмысленным, избыточным, его присутствие игнорируется, и поэтому чтение антикодоном кодона производится по правилу тАЬдва из трехтАЭ. Отсюда логично следует массированная неоднозначность прочтения иРНК и некорректность трансляции белковых молекул, что противоречит экспериментам, и это констатируется в [52], равно как и в других исследованиях. Вместе с тем, отмечается, что существует определенный уровень неоднозначности трансляции иРНК в клетке, но он слабо поддается осмыслению. Помимо ошибочной трансляции значащих кодонов и считывания стоп-кодонов как аминокислотных, в процессе белкового синтеза могут происходить многочисленные нормальные и редко ошибочные сдвиги и перекрытия рамок трансляции. Ошибки возникают в результате считывания дуплетов или квадриплетов оснований как кодонов. Механизмы сдвигов рамки считывания практически не изучены. Во многих работах показано, что ошибочная трансляции белков рибосомой вызывается разнообразными неблагоприятными факторами - антибиотиками, изменением температуры, созданием определенных концентраций катионов, аминокислотным голоданием и другими условиями внешней среды. Повышенная неоднозначность трансляции кодонов, локализованных в особом контексте, имеет биологическое значение и приводит к неслучайному распределению тАЬошибочныхтАЭ аминокислот по длине синтезируемого полипептида, приводящему к модификациям функций белков с выходом на механизмы клеточных дифференцировок, и поэтому контексты иРНК являются субстратом естественного отбора. Оптимальный уровень тАЬошибоктАЭ трансляции (если это действительно ошибки) регулируется неизвестными механизмами, и он онтогенетически и эволюционно оправдан [52]. Этому соответствуют и наши экспериментально-теоретические данные [8-18] о волновых знаковых взаимодействиях в водно-жидкокристаллической среде клетки, в которые вовлечен белок-синтезирующий аппарат. Нами обнаружены резонансные частоты, общие для ДНК, рибосом и коллагена, и имеющие, вероятно, биознаковую природу, а также открыта способность хромосом и ДНК быть лазеро-активной средой [18].
Вернемся вновь к общепринятым поначалу основным положениям генетического кода: он является триплетным, неперекрывающимся, вырожденным, не имеет тАЬзапятыхтАЭ, т.е. кодоны ничем не отделены друг от друга. И наконец, он универсален. Что осталось от этих положений? Фактически ничего. В самом деле, код, видимо, является двух-, трех-, четырех-, .. n-буквенным как фрактальное и гетеромультиплетное образование. Он перекрывающийся. Он имеет запятые, поскольку гетерокодоны могут быть отделены друг от друга последовательностями с иными функциями, в том числе с функциями пунктуации. Код не универсален - в митохондриях он приобретает специфические черты. Как понимать генетический код с учетом приведенных противоречий и предлагаемой нами логики рассуждений?
Для снятия этих противоречий можно постулировать качественную, упрощенную, первичную версию вещественно-волнового контроля за порядком выстраивания аминокислот в ассоциате аминоацилированных тРНК как предшественнике белка. С этой позиции легче понять работу генетического, а точнее белкового, кода как одной из множества иерар-хических программ вещественно-волновой самоорганизации биосистемы. В этом смысле такой код - первый этап хромосомных планов построения биосистемы, поскольку язык генома многомерен, плюралистичен и не исчерпывается задачей синтеза протеинов. Более детальное, физико-математически формализованное и экспериментально подтверждаемое, изложение новой версии работы белок-синтезирующего аппарата разра-батывается нами в настоящее время, хотя надо признать, что это задача xxI-xxII веков.
Основные положения предлагаемой ориентировочной модели вещественно-волновых знаковых процессов при биосинтезе белков сводятся к следующему:
1. Многокомпонентный рибонуклеопротеидный белоксинтезирую-щий аппарат является системой генерации высокоорганизованных знаковых семиотико-семантических излучений акустико-электромагнитных полей, стратегически регулирующих его самоорганизацию и порядок включения аминокислот в полипептидную цепь.
2. Аминоацилированные пулы тРНК ассоциируют в последователь-ности - предшественники синтезируемых белков до контакта с А-P участком рибосомы. При этом континуум антикодонов пула комп-лементарен всей иРНК, за исключением дислокаций, определяемых наличием неканонических нуклеотидных пар.
3. Порядок чередования аминоацилированных тРНК в ассоциатах-предшественниках белков определяется знаковыми коллективными резонансами всех участников синтеза аминокислотных последовательностей. Ключевые волновые матрицы здесь пре-иРНК, а также иРНК, работающие как целостный континуум разномасштабных по длине гетерополикодонов, включая интронную фракцию пре-иРНК как возможных макроконтекстов. Главная функция волновых матриц - ассоциативно-контекстная ориентация последовательности аминоацилированных тРНК, ориентация, в большей степени, чем воблгипотеза, игнорирующая правила канонических спариваний нуклеотидов в пространстве иРНК-тРНК.
4. На рибосоме, в дополнение и (или) наряду с резонансными регуляциями взаимного расположения кодон-антикодоновых континуумов функционируют лазероподобные излучения участников данного процесса, корригирующие порядок включения аминокислотных остатков в пептид.
5. Рибосома энзиматически ковалентно фиксирует тАЬде-юрэтАЭ пептидные связи аминокислотных последовательностей, наме
Вместе с этим смотрят:
"Инкарнация" кватернионов
* Алгебры и их применение
*-Алгебры и их применение
10 способов решения квадратных уравнений
Bilet