Кожа животных и её производные
КОЖА И ЕЕ ПРОИЗВОДНЫЕ
Кожа тАФ сложный и многофункциональный орган. Главная функция наружного покрова позвоночных тАФ защита организма от вредВнных воздействий окружающей среды. Сформировавшиеся в процесВнсе эволюции модификации эпидермиса, который находится в прямом контакте с окружающей средой, существенны для поддержаВнния постоянства внутренней среды в воде и в воздушной среде и защиты от опасностей окружения. Развитие кератинизированных придатков кожи, таких как ороговевшие ВлзубыВ» у миноговых, чешуя, панцирь и коготки рептилий, чешуйки, коготки, перья и клюв птиц, чешуйки, когти, копыта, ногти, шерсть, волосы и рога млекопитающих, необходимо не только для защиты, но и для полового поведения, локомоции, хищничества, поддержания постоянной температуры тела и др. Пигментация, иммунные механизмы, механо-, хемо- и терморецепторы также образуют существенные компоненты защитной системы и опосредуют приток информации из окружающей среды к телу. Защита обеспечивается также некератинизирующимися придатками кожи, такими как железы, отводящими через выводные протоки на поверхность кожи ряд веВнществ. У птиц и млекопитающих такими веществами являются липиды, чтобы смазывать перья, шерсть и поверхность кожи, а у млекопитающих, кроме того, пот для регуляции температуры тела. Кожа участвует в обмене веществ, в процессах теплорегуляции организма, выделения, синтеза витаминов (витамин D) и др.
Хотя строение эпидермиса и его придатков различно у различВнных классов позвоночных, они обладают общими свойствами: 1) состоят из эпителиальных клеток, происходящих из эктодермы, а под ними располагается дерма, происходящая из мезодермы; 2) содержат популяцию герминативных клеток, дающих начало клеткам, дифференцирующимся непрерывно или циклами; 3) дерма влияет на все виды кератинизирующихся тканей, индуцируя и направляя ход дифференцировки.
В соответствии с видовыми особенностями животных кожа хаВнрактеризуется рядом специфических производных кожного покроВнва: копыта травоядных животных, гребень птицы, рога, волосяной покров, молочные железы млекопитающих, перья у птиц и др.
КОЖА
Кожа развивается из двух эмбриональных зачатков. Из эктодермы зародыша развивается наружный слой кожи тАФ эпидермис (рис. 235). Глубокие слои кожного покрова тАФ дерма и подкожная клетчатка тАФ формируются мезенхимой, продуктом дифференцировки дерматомов зародыша.
Эпидермис кожи представлен многослойным плоским ороговевающим эпителием. Толщина его и степень ороговения специфичны для вида животного, области тела и развития волосяного покрова. Наиболее полно представлен эпидермис кожи в областях, не покрытых волосами. Процесс ороговения связан с накоплением клетками специфических фибриллярных белков тАФ кератинов и вторичным их преобразованием. В эпидермисе можно выделить пять слоев: базальный, шиповатый, зернистый, блестящий и роговой. Слои эпидермиса построены в основном из клеток, связанных с процессом ороговения,тАФ кератоцитов (эпидермоцитов).
Базальный слой расположен непосредственно на базальной мембране. Состоит в основном из камбиальных эпидермоцитов и единичных меланоцитов (число меланоцитов относится к числу эпидермоцитов как 1:10). Граница эпидермиса с подлежащей соединительной тканью дермы кожи неровная, что увеличивает площадь их контакта (рис. 236).
Эпидермоциты базального слоя цилиндрической формы. С клетВнками своего слоя и расположенного выше шиповатого слоя они связаны десмосомами, а с подлежащей базальной мембраной полудесмосомами. Клетки этого слоя содержат многочисленные органеллы, в том числе и свободные рибосомы, о чем свидетельствует базофилия их цитоплазмы. Они составляют камбий эпидермиса, обеспечивающий митотическим делением пополнение его клеточного состава в соответствии с интенсивностью ороговения клеток вышележащих слоев, Второй вид клеток тАФ меланоциты (рис. 237) имеют длинные ветвящиеся отростки, лежат свободно. Ядра клеток неправильной формы. В цитоплазме содержатся промеланосомы и меланосомы. Меланоциты синтезируют пигмент меланин, формируют меланосомы и поредают их кератиноцитам (цитокринная секреция). Поэтому по присутствию меланосом отличить меланоциты от кератиноцитов нельзя. Достоверным отличием меланоцитов на светооптическом уровне является положительная реакция на ДОФА-оксидазу (тирозиназу).
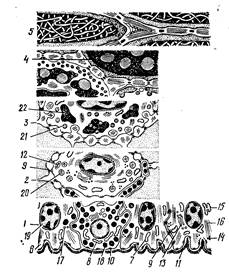
Рис. 237. Схема ультраструктурной организации отдельных слоев эпидермиса:
1тАФ базальный; 2 тАФ шиповатый; 3 тАФ зернистый, 4 тАФ блестящий и 5 тАФ рогоВнвой слои: б тАФ соединительная ткань; 7 тАФ кератоцит; 8 тАФ меланоцит; 9 тАФ десмосома; 10 тАФ полудесмосома; 11 тАФ базальная мембрана; 12 тАФ митохондрии; 13 тАФ комплекс Гольджи; 14 тАФ свободВнные рибосомы; 15 тАФ зернистая эндоплазматическая сеть; 16 тАФ тонофибриллы; 17 тАФ пиноцитозные пузырьки; 18 тАФ гранулы меланина; 19 тАФ ядра; 20 тАФ кератосомы; 21, 22 тАФ гранулы кератогиалина (по Заварзину).
Шиповатый слой представлен несколькими слоями крупных клеток полигональной формы. Многочисленные остистые отростки смежных клеток, связанные десмосомами, образуют межклеточные мостики. Цитоплазма клеток базального и особенно шиповатого слоев содержит многочисленные тонофибриллы, пучки которых заканчиваются в остистых отростках в области десмосом. Пучки тонофибрилл более плотно концентрируются вокруг ядра. В этом слое, кроме эпидермоцитов, встречаются клетки Лангерганса тАФ беспигментные гранулярные дендроциты. Клетка имеет 2 - 5 отростков, распространяющихся до зернистого слоя и базальной мембраны, сильно идентированное или лопастное ядро и светлую цитоплазму. От кератиноцитов их отличает отсутствие тонофибрилл и десмосом, от меланоцитов тАФ отсутствие меланосом и отрицательная реакция на ДОФА-оксидазу. Наиболее характерный признак тАФ наличие в их цитоплазме палочковидных структур и гранул в виде теннисной ракетки. Существует мнение, что эти клетки являются эпидермальными макрофагами.
Зернистый и блестящий слои эпидермиса представлены клетками в последующих стадиях дифференцировки эпителия, в процессе которой клетки постепенно утрачивают органеллы и ядро. Для зернистого слоя характерно присутствие зерен кератогиалина тАФ не ограниченных мембраной участков электроноплотного и базофильного вещества. В цитоплазме присутствуют также сферические ламеллярные гранулы тАФ кератосомы. Они содержат липиды и гидролитические ферменты. Последние активируются в верхних слоях эпидермиса и, возможно, помогают слущиванию роговых чешуек. Липиды выделяются в межклеточные пространства, обеспечивая наземным животным защиту от диффузии воды через кожу и потери жидкости тела. Блестящий слой на светооптическом уровне выглядит гомогенным, на уровне электронной микроскопии видны 2 - 4 слоя плоских клеток, почти лишенных органелл, с разрушающимися ядрами. В цитоплазме присутствуют многочисленные кератиновые фибриллы.
Роговой слой образован многими слоями клеток, завершивВнших процесс ороговения, тАФ роговыми чешуйками. Роговая чешуйка имеет толстую оболочку и заполнена кератиновыми фибриллами, спаянными аморфным кератиновым матриксом. Между чешуйками располагаются сильно измененные десмосомы. Вместо двух утолВнщенных областей противоположных мембран, разделенных менее плотным межклеточным промежутком, десмосома представлена теперь плотным тяжем, располагающимся экстрацеллюлярно в межклеточном пространстве. Таким образом, каждая клетка эпидермиса кожи превращается в роговую чешуйку. Корреляция интенсивности размножения камбиальных клеток базального слоя, ороговения и отторжения поверхностных клеток взаимообусловлена.
В эпидермисе нет кровеносных сосудов. Питательные вещества и кислород в него поступают из капилляров дермы. Последняя образует большую площадь контакта с эпидермисом благодаря обилию сосочков и высокой степени их развития.
Собственно кожа, или дерма, тАФ производное мезенхимы. СоВнстоит из двух слоев: наружного тАФ сосочкового и внутреннего тАФ сетчатого (см. рис. 235).
Сосочковый слой образован рыхлой неоформленной соеВндинительной тканью. Богат аморфным веществом. Содержит тонкие пучки коллагеновых волокон, эластическую сеть и значительное количество клеток: фиброцитов, гистиоцитов, ретикулярных клеток, тканевых базофилов и др. Степень развития сосочков коррелирует с толщиной эпителиальной ткани тАФ эпидермиса.
Сетчатый слой состоит из плотной неоформленной соединительной ткани. Для него характерна регионарная специфичность строения. В частности, в сетчатом слое кожи спины имеются толстые пучки коллагеновых волокон, плотно прилегающих друг к другу. Петли вязи волокон ромбовидной формы, ориентированы преимущественно вертикально. Здесь этот слой толще, чем на животе, где ткань более рыхлая с преимущественно горизонтальной ориентацией волокон, что обеспечивает ей большие возможности при растяжении.
Подкожная клетчатка тАФ слой рыхлой неоформленной соединиВнтельной ткани с преимущественным содержанием жировых клеток. Она подвижно соединяет кожу с подлежащими тканями: обеспечивает подвижность кожного покрова, предохраняет подлежащие ткани от механических повреждений, участвует в теплорегуляции.
Железы кожи. Сальные железы распространены по всему кожному покрову млекопитающих животных. Они отсутствуют в коже сосков вымени, коже носо-губного зеркала, пятачка свиней и мякишей конечностей. По форме сальные железы простые, разветвленные, альвеолярные (рис. 241), их выводные протоки, выстланные многослойным эпителием, открываются в воронку волоса.
Концевые отделы железы образованы многослойным эпителием, клетки которого находятся на различных стадиях процесса голокриновой секреции.
Непосредственно на базальной мембране ацинуса железы леВнжит слой мелких камбиальных клеток. В них различают округлые или овальные ядра и характерные для цитоплазмы органеллы. В цитоплазме выявляют одиночные или собранные в пучки тонофибриллы. По мере размножения камбиальные клетки дифференВнцируются и смещаются в центральную зону концевого отдела. Они при этом увеличиваются в размерах. Их ядра сморщиваются и распадаются. В цитоплазме появляются жировые включения. ПостеВнпенно смещаясь к выводному протоку, клетки железы перерождаются и, распадаясь, образуют секрет.
У крупного рогатого скота наиболее крупные сальные железы лежат в преанальной области, около носо-губного зеркала, у корня рогов и в области венчика копыта. Их нет в коже, не содержащей волос. Выводной проток открывается в волосяной канал. У овец сальные железы состоят из двух и более долек.
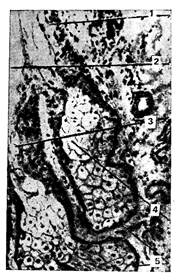
Рис. 241. Сальные железы кожи:
1 тАФ эпителий стенки выводного протока железы; 2 тАФ волосяная воронка; 3 тАФ просвет выводного протока железы; 4 тАФ разрушающиеся клетки концевого отдела железы; 5 тАФ малодифференцированные клетки концевого отдела железы.
Потовые железы простые, трубчатые, неразветвленные, их концевые секреторные отделы расположены в глубокой зоне сетчатого слоя дермы и образуют более или менее плотные клубки. В концевом отделе потовых желез два вида клеток: кубические тАФ железистые и отростчатые тАФ миоэпителиальные. Последние охватывают отростками концевой отдел железы и, сокращаясь, регулируют эвакуацию секрета через выводные протоки. У лошади, овцы, свиньи и кошки концевой отдел железы образует компактный клубок, тогда как у крупного рогатого скота, коз, собак он только извилист. Различают два вида потовых желез тАФ эккринные (мерокринные) и апокринные. Первые типичны для безволосых участков кожи. Их протоки открываются непосредственно на поверхности рогового слоя кожи. Апокринные потовые железы связаны с волосяным покровом. Их выводные протоки впадают в волосяные фолликулы, несколько выше сальных желез. Секрет апокринных желез богат белками.
ВОЛОС
Кожа домашних животных покрыта волосами. Волосяной покров отсутствует на носо-губном зеркале крупного рогатого скота, носоВнвом зеркале мелкого рогатого скота, пятачке свиней, мякише стопы плотоядных и в местах перехода кожи в слизистые оболочки. ВоВнлосы тАФ продукт дифференцировки эпидермиса кожи.
В волосе различают корень волоса (часть, погруженную в кожу) и стержень, свободно находящийся над ее поверхностью. Утолщенный конец корня волоса называется волосяной луковицей. Ее неороговевающие эпителиальные клетки составляют камбий волоса. В волосяную луковицу погружен соединительнотканный сосочек с кровеносными сосудами, обеспечивающими обмен веществ тканей луковицы (рис. 242).
Корень волоса заключен в волосяной мешочек тАФ фолликул. Стенка фолликула состоит из непосредственно прилегающего к волосу внутреннего корневого влагалища, наружного корневого влагалища и волосяной сумки. Внутреннее корневое влагалище развивается вместе с волосом из эпителия луковицы волоса. НаВнружное корневое влагалище, сформированное погружением эпиВндермиса кожи, соответствует его неороговевшем слоям. В области воронки фолликула оно переходит в ростковый слой эпидермиса. Волосяная сумка образуется соединительной тканью дермы кожи.
Строение. Волос состоит из корня, погруженного в кожу, и стержня, находящегося свободно над ее поверхностью. В нем различают мозговое вещество, корковое вещество и кутикулу.
Мозговое вещество имеется только в длинных волосах и в щетине. Его клетки полигональной формы, находящиеся на различных стадиях ороговения, образуют центрально-расположенный тяж. Они ацидофильны, содержат гранулы продукта ороговеВнния тАФ трихогиалина, пузырьки воздуха и зерна пигмента (рис.243-А).
Корковое вещество тАФ основная часть волоса. Интенсивно размножающиеся эпителиальные клетки луковицы по мере смещеВнния их от камбия быстро ороговевают и в составе коркового вещества представляют собой массу плотно упакованных роговых чешуек. Последние содержат продукт ороговения тАФ твердый кератин, зерна пигмента, определяющие цвет волоса, и пузырьки воздуха.
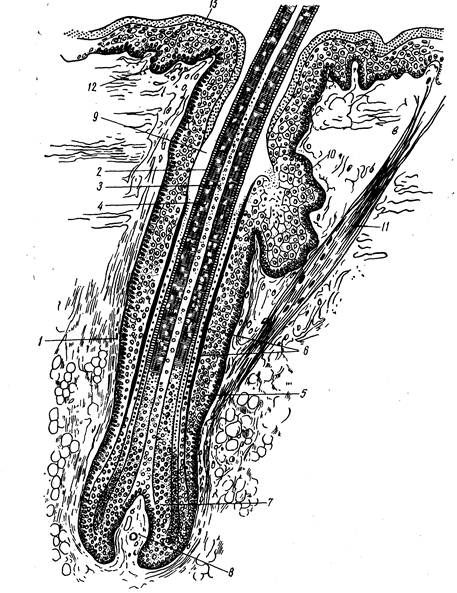
Рис. 242. Схема строения волоса: 1 тАФ волосяная сумка; 2 тАФ корковое и 3 тАФ мозговое вещество; 4 тАФ кутикула; 5 тАФ наружное эпителиальное влагалище; 6 тАФ два слоя внутреннего эпителиального влагалища; 7 тАФ волосяная луковица; 8 тАФ волосяной сосочек; 9 тАФ волосяная воронка; 10 тАФ сальная железа; 11 тАФ мышца, поднимающая волос; 12 тАФ ростковый слой эпидермиса; 13 тАФ роговой слой эпидермиса.
Кутикула тАФ наружный слой волоса. Форма и состояние клеВнток в различных зонах корня соответствуют степени их дифференцировки. Непосредственно вблизи луковицы кутикула представлеВнна одним слоем призматических клеток. По мере роста волоса, а соответственно и перемещения клеток кутикулы к поверхности кожи они, меняя форму и положение, формируют один слой плоских черепицеобразно расположенных чешуек. Форма и положение последних специфичны для различных видов животных.
Фолликул волоса (волосяной мешочек) состоит из внутреннего и наружного корневого влагалища и волосяной сумки.
Внутреннее корневое влагалище формируется размножением клеток периферической зоны волосяной луковицы. ПоВнкрывает корень волоса до протоков сальной железы. Состоит из трех слоев: кутикулы, гранулосодержащего эпителиального слоя (слой Гексли), бледного эпителиального слоя (слой Генле). Различия этих слоев определяются формой клеток и степенью их ороВнговения. Они хорошо видны лишь в глубокой трети корня (рис. 243тАФБ). Кутикула внутреннего корневого влагалища обраВнзована одним слоем плоских ороговевших черепицеобразно расВнположенных клеток. К ним прилежат 2тАФ3 слоя гранулосодержащих клеток. Бледный слой представлен одним слоем клеток на боВнлее глубокой стадии ороговения.
Наружное корневое влагалище, распространяясь от луковицы волоса, где оно образовано 1тАФ2 слоями клеток, постеВнпенно утолщается и переходит в области воронки волоса в росткоВнвый слой эпидермиса кожи. В нем можно выделить характерные для многослойных плоских эпителиев три слоя клеток: базальный шиповатый и поверхностный (плоские клетки).
Волосяная сумка образована базальной мембраной и двуВнмя слоями соединительной ткани. Внутренний, примыкающий к наружному корневому влагалищу, характеризуется преимущественно циркулярной ориентацией волокон межклеточного вещества, а наружный тАФ их продольным расположением. Волосяная сумка и волосяной сосочек богато васкуляризированы. В области губ, век, ноздрей находятся осязательные (синусоидные) волосы, волосяная сумка которых содержит особые полоститАФсинусоиды, выстланные эндотелием и наполненные кровью.
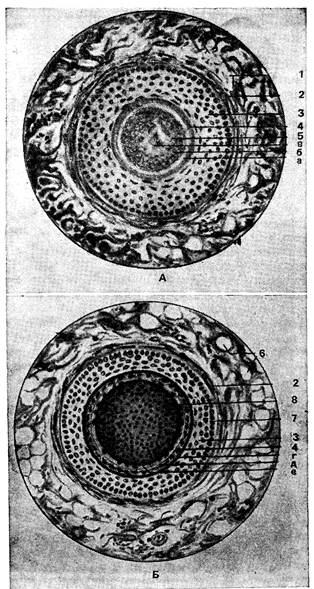
Рис. 243. Поперечные срезы корня волоса на уровне дна сальной железы (4) и на уровне нижней части корня (Б);
1 тАФ сетчатый слой кожи; 2 тАФ волосяная сумка; 3тАФ наружное корВнневое влагалище; 4 тАФ внутреннее корневое влагалище; 5 тАФ корень волоса (а тАФ кутикула волоса, б тАФ корковое вещество, в тАФ мозговое вещество); 6 тАФ жировые клетки; 7 тАФ базальная (стекловидная) мембрана; 8 тАФ волос (г тАФ бледный эпителиальный слой тАФ слой Генле, д тАФ гранулосодержащий эпителиальный слой тАФ слой Гексли, е тАФ кутикула волоса).
Развитие волоса. Волос тАФ продукт дифференцировки эпидермиса кожи. Его формирование начинается с локальной репродукции клеток эпидермиса и погружения их в виде тяжа в соединительную ткань дермы. Интенсивный процесс пролиферации эпителия коррелирует с активизацией смежных участков соединительной ткани дермы и ее кровоснабжения. Глубокий конец эпителиальной закладки волоса утолщается в виде колбы. Окружающая ее соединительная ткань формирует волосяную сумку и волосяной сосочек, погружающийся в расширенный конец эпителиального тяжа. Формируется волосяная луковица. Клетки центральной части эпителия луковицы, размножаясь, образуют ткань волоса тАФ его мозговое и корковое вещество и кутикулу. Смежная, периферическая, зона клеток луковицы продуцирует клетки внутреннего корневого влагалища волоса. Растущий от луковицы волос проВндвигается по каналу в центре эпиВнтелиальной закладки, образовавВншемуся в связи с ороговением и последующим отторжением клеток ее центральной зоны. Наружные, неороговевающие слои эпителиальной закладки волоса образуют наружное корневое влагалище, которое непосредственно контактирует с соединительнотканной волосяной сумкой.
Смена волоса. Для каждого вида животных характерны определенные закономерности роста волос и их смены. Различают ювенильную, периодическую и перманентную смену волос.
При ювенильной смене волосяной покров новорожденного животного заменяется дефинитивным. Эта линька не зависит от сезона года. Периодическая (сезонная) смена волос преимущественно характерна для диких животных. Она соответствует весеннему периоду года. Для большинства домашних животных, в том числе и для крупного рогатого скота, типична постепенная линька, не связанная с сезонами года. Перманентная линька тАФ смена волос в течение года, характерна для свиней, шерстяных волос овец культурных пород.
При линьке редуцируется волосяной сосочек. В клетках лукоВнвицы волоса исчезают митозы. Луковица в своей большей части ороговевает, отходит от соединительнотканного сосочка и смещаетВнся по волосяному каналу к поверхности кожи. Некоторое время волос остается в воронке волосяного фолликула, затем выпадает. По мере регенерации луковицы формируется новый волос (рис. 244).
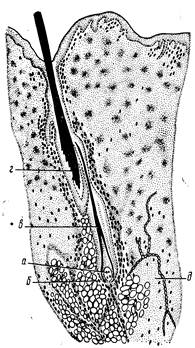
Рис. 244. Смена волос в коже свиньи (по Ржаницкой):
а тАФ волосяной сосочек; б тАФ волосяная луковица; в тАФ новый волос; г тАФ старый воВнлос; д тАФ нервные волокна.
МОЛОЧНАЯ ЖЕЛЕЗА
Молочные железы тАФ специализированные апокриновые железы кожного покрова, функционально органически связанные с эндоВнкринной регуляцией женской половой системы организма. Они формируются у зародышей на ранней стадии эмбрионального раз вития в виде двух продольных утолщений эпидермиса тАФ молочных линий вдоль туловища зародыша. Разрастание эпидермиса образует млечные бугорки, количество и местоположение которых соответствует количеству и расположению молочных желез у отдельных видов животных.
По морфологии выводных протоков и секреторных отделов молочная железа относится к сложным разветвленным трубчато-альвеолярным железам. Паренхима железы состоит из системы разВнветвленных выводных протоков, концевых секреторных отделов и соединительной ткани, богатой жировыми клетками. Последняя разделяет паренхиму железы на дольки различных размеров. В междольковой соединительной ткани проходят кровеносные сосуды, нервы и междольковые выводные протоки, заканчивающиеся в паренхиме долек концевыми железистыми отделами альвеолярной или трубкообразной формы. Крупные выводные протоки впадают в молочную цистерну. Из цистерны молоко поступает в сосковый канал (рис. 248).
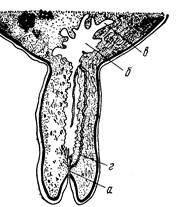
Рис. 248. Схема продольного разреза соска 6-месячной телки:
а тАФ сосковый канал; б тАФ молочная цистерна; в тАФ ее боковые выпячивания; д тАФ дополнительные боковые канальцы.
Первичные дольки железы содержат от 158 до 226 (по Техверу) концевых отделов тАФ молочных альвеол, концевые выводные протоки и соединительнотканную строму. Они разграничены междольковой соединительной тканью, богатой жировыми клеткаВнми. Стенка концевых отделов железы состоит из однослойного секреторного эпителия и миоэпителиальных корзинчатых клеток, охватывающих своими отростками концевые отделы.
В период лактации концевые отделы характеризуются кубичеВнской или призматической формой секреторных клеток (лактоцитов), связанных между собой системой десмосом. Апикальная поВнверхность железистых клеток неровная. Она снабжена микроворсинками. Цитоплазма клеток содержит гладкую и гранулярную эндоплазматическую сеть, комплекс Гольджи, микротрубулы и микрофибриллы. В период, предшествующий выведению секрета, клетки высокие. На их свободной поверхности образуются куполоВнобразные выпячивания, содержащие капли жира. Они достигают значительных размеров, отрываются и вместе с покрывающей их плазмолеммой поступают в полость альвеол (рис. 249). В альвеолы выводятся и другие продукты жизнедеятельности секреторных клеток: козеин, лактоза, соли и др.
Второй слой клеток стенки секреторной альвеолы, расположенной непосредственно на базальной мембране железы, образуют миоэпителиальные клетки, участвующие в выведении секрета в выводные протоки.
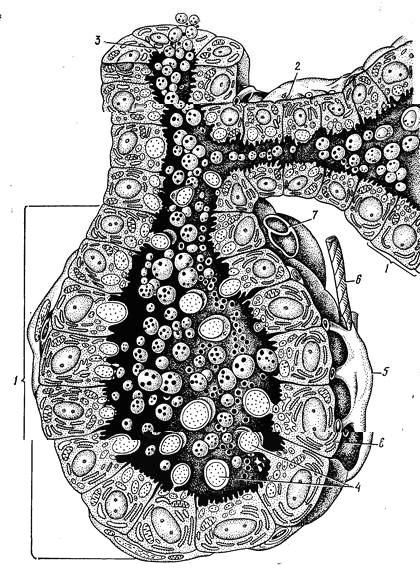
Рис. 249. Схема строения ацинусов молочной железы:
1 тАФ ацинусы; 2 тАФ молочный ход; 3 тАФ внутридольковый проток; 4 тАФ апокриновая секреция; 5 тАФ миоэпителиальные клетки; б тАФ нервное волокно; 7 тАФ гемокапилляр; 8 тАФ лактоцит.
Выводные протоки. Мелкие и средние выводные протоки молочной железы выстланы однослойным кубическим эпителием. Высота клеток эпителия увеличивается по мере увеличения калибра протока. В крупных протоках он высокий призматический однослойный и двухслойный. Вторым слоем в мелких и средних проВнтоках лежат миоэпителиальные клетки, которые в крупных протоках замещаются гладкими мышечными. В молочной цистерне эпителий двухслойный, а в соске многослойный ороговевающий.
Молочная цистерна выстлана двухслойным призматическим эпителием, частично в некоторых участках поверхности цистерны переходящим в многослойный плоский. Собственный слой слизистой оболочки образован рыхлой неоформленной соединительной тканью, богатой эластическими волокнами.
Сосок вымени коровы образуется как складка кожи, соответстВнвенно сосочковый канал выстилается многослойным плоским эпиВнтелием. Между двумя эпителиальными слоями (эпидермисом кожа соска и эпителием его канала) залегает слой соединительной ткани и гладких мышечных клеток. Эпителий сосочкового канала ороговевает, что предупреждает сужение или замыкание сосочкового канала. Соединительная ткань соска содержит гладкие мышечные клетки, образующие четыре слоя: 1) продольный, хорошо развиВнтый в области молочной цистерны; 2) кольцевой, образующий сфинктер соска; 3) слой мышечных клеток, переплетающихся между собой, и 4) радиальные пучки клеток.
Молочная железа богато иннервирована. От крупных нервных стволов отдельные волокна направляются по кровеносным сосудам в железистую паренхиму, где образуют чувствительные и эффекторные нервные окончания, участвующие в рефлекторных процессах молокоотделения и молокоотдачи.
Вместе с этим смотрят:
20 мешков картошки с 1 сотки
Cовершенствование организации производства молока на КПУП "ГМЗ № 3"
Cостояние полезащитных лесных полос в северном Приднестровье
РЖнтегрований захист озимоi пшеницi
РЖнтенсифiкацiя та ii економiчна ефективнiсть у сiльському господарствi