Сердечно-сосудистая система животных
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Сердечно-сосудистая система состоит из сердца, кровеносных и лимфатических сосудов. В функциональном отношении эта система обеспечивает движение по организму крови и лимфы, создавая эффективную систему доставки к органам и тканям кислорода, питательных и биологически активных веществ и удаления продуктов метаболизма.
СЕРДЦЕ
Сердце тАФ основной орган, приводящий в движение кровь. У млекопитающих оно состоит из двух соединенных половин тАФ правой и левой. Как правая, так и левая половины имеют два отдела тАФ предсердие и желудочек, сообщающиеся через отверстия, на границе которых расположены открывающиеся в сторону желудочков клапаны. В стенке предсердий и желудочков различают три оболочки: внутреннюю тАФ эндокард, среднюю тАФ миокард и наружную тАФ эпикард.
В эмбриогенезе оболочки сердца начинают формироваться в очень ранний период развития, когда зародыш имеет вид трехВнслойной пластинки. Вначале из мезенхимных клеток между эндодермой и висцеральным листком несегментированной мезодермы развиваются две отдельные трубочки, которые выпячиваются в целомическую полость тела. В дальнейшем, в связи с увеличением туловищной складки, мезенхимные трубки сближаются, срастаются и из них образуется одна, которая становится эндокардом.
Участки висцерального листка мезодермы, прилежащие к эндокарду, получили название миоэпикардиальных пластинок. Внутренняя пластинка превращается в миокард, а из наружной образуется эпикард. Дальнейшее формирование сердца связано с неравномерным разрастанием отдельных участков сердечной трубки. Клапаны сердца развиваются как складки эндокарда, в которые позднее врастает соединительная ткань миокарда и эпикарда.
Эндокард. Эта оболочка представляет непрерывную выстилку предсердий, желудочков и покрывает все структурные образования, выступающие в их просвет, тАФ клапаны, сосочковые мышцы. По строению и происхождению эндокард соответствует стенке кровеносных сосудов. В области предсердий и желудочков в его составе различают три слоя. Самый внутренний образован эндотелием и расположенными под ним элементами соединительной ткани. Средний тАФ мышечно-эластический слой имеет наибольшую толщину и состоит из плотной соединительной ткани с многочисленными эластическими волокнами, располагающимися параллельно поверхности. В наружной части этого слоя имеются клетки гладкой мышечной ткани. Третий слой тАФ наружный соединительнотканный тАФ граничит с миокардом, состоит из рыхлой соединительной ткани, которая переходит в эндомизий миокарда. Этот слой содержит кровеносные сосуды, а в отдельных участках тАФ атипичные клетки проводящей мышечной ткани.
Между предсердиями и желудочками, а также между желудочками и выходящими из них сосудами располагаются клапаны. Предсердно-желудочковый (атриовентрикулярный) клапан в правой половине сердца состоит из трех створок, поэтому его называют трехстворчатым, а в левой половине тАФ клапан двустворчатый, или митральный. Створки обоих атриовентрикулярных клапанов обладают сходной гистологической структурой. Они покрыты с обеих сторон эндотелием и имеют средний слой плотной соединительной ткани, переходящей в основании створки в плотную соединительную ткань колец, окружающих отверстия. Предсердная сторона створок тАФ гладкая, а желудочковая с выступами, от которых начинаются сухожильные нити, противоположными концами прикрепляющиеся к выступающим на стенках желудочков сосочковым мышцам. Благодаря сухожильным нитям при сильных сокращениях желудочков не происходит выворачивания створок клапанов в сторону предсердий. Клапаны аорты и легочной артерии (полулунные) несколько тоньше, чем предсердно-желудочковые, но гистологически имеют сходное с ними строение. У полулунных клапанов нет сухожильных нитей.
Миокард образован сердечной мышечной тканью, в которой различают две разновидности тАФ рабочую и проводящую. Основная масса миокарда представлена рабочей мышечной тканью, состоящей из сократительных клеток тАФ сердечных миоцитов, важнейшей морфологической особенностью которых являются совершенные в структурном и функциональном отношении аппараты крепления их друг с другом. Вследствие того что миоциты прочно соединены своими концами и образуют многочисленные анастомозы, в миокарде сформирована единая структурно-функциональная клеточная сеть. При световой микроскопии зоны контакта миоцитов имеют вид одиночных темноокрашивающихся прямолинейных или ступенчатых полосок, расположенных перпендикулярно длинной оси клетки, которые получили название вставочных дисков (рис. 198).
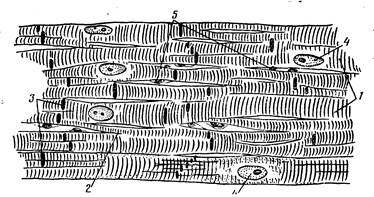
Рис. 198. Схема строения участка рабочей мышечной ткани миокарда:
1 тАФ миокардиоциты; 2 тАФ анастомозы; 3 тАФ вставочные участки; 4 тАФ ядра миокардиоцитов; 5 тАФ ядра эндотелия капилляров.
При электронной микроскопии в области вставочных дисков границы соседних клеток неровные: одна клетка вдается в другую пальцевидными выступами, что обеспечивает достаточную площадь сцепления клеток. По длине вставочного диска имеются различные по строению участки. Здесь много мест, содержащих волокнистое электроноплотное вещество, в которое вплетены концы тонких актиновых микрофиламентов (рис. 199). В зонах, не занятых микрофиламентами, расположены десмосомы и щелевые контакты. Считают, что в щелевых контактах происходит быстрая передача волны возбуждения от клетки к клетке без участия медиатора. Каждый сердечный миоцит содержит 1тАФ2 ядра, расположенные в центре клетки, миофибриллы занимают периферическую часть цитоплазмы. Между миофибриллами одиночно, группами или цепочками расположены митохондрии, для которых характерно большое количество крист. Миофибриллы окружены системой трубочек и канальцев саркоплазматической сети. Развита Т-система, образованная трубчатыми впячиваниями плазмолеммы миоцита, причем
Т-трубочки расположены на уровне Z-полоcок миофибрилл. В околоядерной зоне capкоплазмы расположены скопления митохондрий, комплекс Гольджи, лизосомы, гранулы гликогена и пигмента липофусцина.
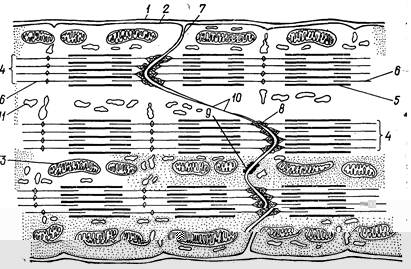
Рис. 199. Схема ультраструктурной организации миокардиоцитов в области контакта двух клеток (вставочного участка):
1 тАФ сарколемма; 2 тАФ плазмолемма; 3 тАФ митохондрии; 4 тАФ миофибрилла; 5 тАФ миозиновые филаменты; б тАФ актиновые филаменты; 7 тАФ граница между миокардиоцитами; 8 тАФ зона вплетения актиновых миофиламентов; 9 тАФ десмосома; 10 тАФ щелевой контакт; 11 тАФ канальцы саркоплазматической сети.
Миоциты окружены тонковолокнистой соединительной тканью (эндомизий миокарда), в которой содержатся многочисленные капилляры, обеспечивающие процессы микроциркуляции. В сердце имеется опорный скелет, состоящий из фиброзных колец на границе между предсердиями и желудочками и в устьях сосудов, выходящих из желудочков. В составе фиброзных колец находятся пучки плотной коллагеновой соединительной ткани, а в сердце животных хрящевая и даже костная ткань (крупный рогатый скот). Фиброзно-хрящевые кольца препятствуют растяжению отверстий и обеспечивают прикрепление свободных концов волокон миокарда.
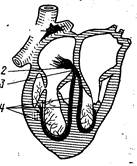 Автоматизм сокращения сердца, закономерная последоваВнтельность сокращений предсердий и желудочков на протяжении сердечного цикла обусловлены деятельностью атипичной мышечной ткани, входящей в состав проводящей системы сердца. Эта система состоит из синусно-предсердного узла, располоВнженного в устье краниальной полой вены, предсердно-желудочкового узла, лежащего в межпредсердной перегородке вблизи места прикрепления створки трехстворчатого клапана, предсердно-желудочкового ствола (пучка Гиса) и его разветвлений, расположенных под эндокардом межжелудочковой перегородки и в соединительнотканных прослойках миокарда тАФ волокна Пуркине (рис. 200). Все эти компоненты проводящей системы образованы атипичными мышечными клетками, которые в функциональном отношении специализированы или на выработке импульса распространяющегося по всему сердцу и вызывающего сокращение его отделов в необходимой последовательности и с определенной частотой (клетки узлов), или на его проведении и передаче сократительным миоцитам.
Автоматизм сокращения сердца, закономерная последоваВнтельность сокращений предсердий и желудочков на протяжении сердечного цикла обусловлены деятельностью атипичной мышечной ткани, входящей в состав проводящей системы сердца. Эта система состоит из синусно-предсердного узла, располоВнженного в устье краниальной полой вены, предсердно-желудочкового узла, лежащего в межпредсердной перегородке вблизи места прикрепления створки трехстворчатого клапана, предсердно-желудочкового ствола (пучка Гиса) и его разветвлений, расположенных под эндокардом межжелудочковой перегородки и в соединительнотканных прослойках миокарда тАФ волокна Пуркине (рис. 200). Все эти компоненты проводящей системы образованы атипичными мышечными клетками, которые в функциональном отношении специализированы или на выработке импульса распространяющегося по всему сердцу и вызывающего сокращение его отделов в необходимой последовательности и с определенной частотой (клетки узлов), или на его проведении и передаче сократительным миоцитам.
Рис. 200. Схема проводящей системы сердца:
1 тАФ синусно-предсердный узел; 2 тАФ предсердно-желудочковый узел; 3 тАФ предсердно-желудочковый ствол (пучок Гиса); 4 тАФ его ножки и разветвления.
Атипичные миоциты проводящей системы имеют характерные микроскопические и ультраструктурные признаки, отличающие их от сократительных миоцитов. При обычной гематоксилиновой окраске они более светлые, имеют неправильно овальную форму и, как правило, поперечный диаметр их больше, чем диаметр сократительных миоцитов, в 2тАФ3 раза. Однако в составе синусно-предсердного узла обнаружены мелкие клетки округлой формы. В функциональном отношении это водители ритма тАФ пейсмекеры. Весьма характерным для атипичных миоцитов являются большой объем саркоплазмы и слабое развитие миофибриллярного аппарата. Миофибриллы занимают самую периферическую часть в цитоплазме клеток, не имеют параллельной ориентации, вследствие чего для атипичных миоцитов несвойственна поперечная исчерченность. У них слабо развит саркоплазматический ретикулум, отсутствует система Т-трубок, а в саркоплазме мало митохондрий, но имеется большое количество гранул гликогена. В этих клетках много гликолитических ферментов и уменьшенное количество ферментов аэробного окисления (сукцинатдегидрогеназы и цитохромоксидазы), что свидетельствует о преобладании в них анаэробного гликолиза. Клетки проводящей системы значительно более устойчивы к кислородному голодаВннию, чем сократительные миоциты.
Эпикард тАФ наружная оболочка сердца. По строению представляет тонкую серозную оболочку, состоящую из соединительно-тканной основы, содержащей разнообразно ориентированные коллагеновые и эластические волокна, и поверхностного слоя тАФ плоского однослойного эпителия (мезотелия). В соединительной ткани эпикарда проходят крупные кровеносные сосуды и имеется жировая ткань.
Эпикард является висцеральным листком перикарда, плотно срастающимся с миокардом и в области корней крупных сосудов сердца переходящим в париетальный листок. Между этими листками имеется щелевидная перикардиальная полость, содержащая серозную жидкость, выделяемую клетками мезотелия. Постоянное увлажнение поверхности эпикарда жидкостью уменьшает его трение о париетальный листок перикарда и обеспечивает лучшую подвижность сердца.
Кровоснабжение и иннервация сердца. Кровь доставляется к стенке сердца по правой и левой венечным (коронарным) артериям, ответвляющимся от аорты вблизи ее клапана. По строению они относятся к артериям мышечно-эластического типа. Венечные артерии разветвляются на ряд мелких артерий, снабжающих кровью оболочки сердца. Между мелкими ветвями артерий и вен имеются анастомозы. В створках клапанов сердца кровеносных сосудов нет. В миокарде большое количество капилляров густой сетью оплетают волокна, образуя узкопетлистую сеть, обеспечивающую процессы микроциркуляции. Капиллярные сети вытянуты вдоль мышечных волокон. Показано, что каждый сократительный миоцит находится в контакте не меньше чем с двумя капиллярами. Кровь из капилляров собирается в коронарные вены, впадающие в правое предсердие.
Проводящая система, в клетках которой происходит генерация возбуждающих импульсов, способна обеспечить автоматические ритмические сокращения сердца лишь в покое. В условиях деятельности организма работа сердца находится под непрерывным воздействием нервной системы. Иннервация сердца осуществляется волокнами симпатического и блуждающего нервов, образующих в оболочках нервные сплетения с интрамуральными ганглиями. В составе постганглионарных симпатических волокон находятся аксоны клеток звездчатого ганглия и клеток передних грудных симпатических узлов. Концевые утолщения аксонов образуют в сердце двигательные нервные окончания. Парасимпатические волокна содержат аксоны клеток, тела их располагаются в ядре блуждающего нерва в продолговатом мозгу. В сердце они образуют синапсы на нейронах внутрисердечного ганглия, аксоны которых заканчиваются на мышечных клетках.
Афферентная иннервация осуществляется дендритами нервных клеток, тела которых находятся в узловатом ганглии блуждающего нерва и спинномозговых узлах первых шести грудных сегментов спинного мозга. Концевые веточки дендритов в миокарде формируют многочисленные чувствительные нервные окончания, которые можно разделить на две группы. Одна группа тАФ механорецепторы, расположенные в соединительнотканных прослойках и вокруг артериол. В них возникает сигнал при изменениях просвета кровеносных сосудов и растяжении соединительной ткани. Центростремительные импульсы от этих рецепторов вызывают рефлекторное ускорение ритма сердца. Другая группа тАФ мышечные рецепторы, имеющие вид спирали. Они специализированы для сигнализации о сокращении миоцитов (рис. 201). Кроме того, с участием различных нервных клеток, сосредоточенных в интракардиальных ганглиях, образуются местные рефлекторные дуги.
Рис. 201. Схема иннервации сердца:
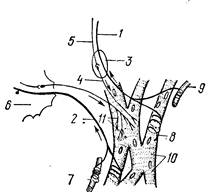 1 тАФ афферентное волокно блуждающего нерва;
1 тАФ афферентное волокно блуждающего нерва;
2 тАФ афферентное волокно, проходящее через узел;
3 тАФ внутрисердечный парасимпатический узел;
4 тАФ постганглионарное волокно;
5 тАФ преганглионарное волокно;
б тАФ звездчатый симВнпатический узел;
7 тАФ механорецепторы;
8 тАФ мышечные рецепторы;
9 тАФ кровеВнносный сосуд;
10 тАФ миокардиоциты;
11 тАФ двигательные нервные окончания.
КРОВЕНОСНЫЕ СОСУДЫ
Кровеносные сосуды представляют замкнутую систему разветвленных трубок разного диаметра, входящих в состав большого и малого кругов кровообращения. В этой системе различают: артерии, по которым кровь течёт от сердца к органам и тканям, вены тАФ по ним кровь возвращается в сердце, и комплекс сосудов микроциркуляторного русла, обеспечивающих наряду с транспортной функцией обмен веществ между кровью и окружающими тканями.
Кровеносные сосуды развиваются из мезенхимы. В эмбриогенезе наиболее ранний период характеризуется появлением многочисленных клеточных скоплений мезенхимы в стенке желточного мешка тАФ кровяных островков. Внутри островка образуются кровяные клетки и формируется полость, а расположенные по периферии клетки становятся плоскими, соединяются между собой при помощи клеточных контактов и формируют эндотелиальную выстилку образующейся трубочки. Такие первичные кровеносные трубочки по мере образования соединяются между собой и формируют капиллярную сеть. Окружающие клетки мезенхимы превращаются в перициты, гладкие мышечные клетки и адвентициальные клетки. В теле зародыша кровеносные капилВнляры закладываются из клеток мезенхимы вокруг щелевидных пространств, заполненных тканевой жидкостью. Когда по сосудам усиливается кровоток, эти клетки становятся эндотелиальными, а из окружающей мезенхимы формируются элементы средней и наружной оболочки.
Сосудистая система обладает очень большой пластичностью. Прежде всего, отмечается значительная изменчивость густоты сосудистой сети, так как в зависимости от потребностей органа в питательных веществах и кислороде в широких пределах колеблется количество приносимой ему крови. Изменение скорости кровотока и кровяного давления ведет к образованию новых сосудов и перестройке имеющихся сосудов. Происходит превращение мелкого сосуда в более крупный с характерными особенностями строения его стенки. Наибольшие изменения возникают в сосудистой системе при развитии окольного, или коллатеральВнного, кровообращения.
Артерии и вены построены по единому плану тАФ в их стенках различают три оболочки: внутреннюю (tunica intima), среднюю (tunica media) и наружную (tunica adventicia). Однако степень развития этих оболочек, их толщина и тканевый состав тесно связаны с функцией, выполняемой сосудом и гемодинамическими условиями (высотой кровяного давления и скоростью кровотока), которые в различных отделах сосудистого русла неодинаковы.
Артерии. По строению стенок различают артерии мышечВнного, мышечно-эластического и эластического типов.
К артериям эластического типа относятся аорта и легочная тАв артерия. В соответствии с высоким гидростатическим давлением (до 200 мм ртутного столба), создаваемым нагнетательной деятельностью желудочков сердца, и большой скоростью кровотока (0,5 - 1 м/с) у этих сосудов резко выражены упругие свойства, которые обеспечивают прочность стенки при ее растяжении и возвращении в исходное положение, а также способствуют превращению пульсирующего кровотока в постоянный непрерывный. Стенка артерий эластического типа отличается значительной толщиной и наличием большого количества эластических элементов в составе всех оболочек.
Внутренняя оболочка состоит из двух слоев тАФ эндотелиального и подэндотелиального. Эндотелиальные клетки, формирующие сплошную внутреннюю выстилку, имеют различную величину и форму, содержат одно или несколько ядер. В их цитоплазме немногочисленные органеллы и много микрофиламентов. Под эндотелием находится базальная мембрана. Подэндотелиальный слой состоит из рыхлой тонковолокнистой соединительной ткани, в составе которой наряду с сетью эластических волокон присутствуют малодифференцированные клетки звездчатой формы, макрофаги, гладкие мышечные клетки. В аморфном веществе этого слоя, имеющем большое значение для питания стенки, соВндержится значительное количество гликозаминогликанов. При повреждении стенки и развитии патологического процесса (атеросклерозе) в подэндотелиальном слое накапливаются липиды (холестерин и его эфиры). Клеточные элементы подэндотелиального слоя играют важную роль в регенерации стенки. На границе со средней оболочкой располагается густая сеть эластических волокон.
Средняя оболочка состоит из многочисленных эластических окончатых мембран, между которыми располагаются косо ориентированные пучки гладких мышечных клеток. Через окна (фенестры) мембран осуществляется внутристеночный транспорт веществ, необходимых для питания клеток стенки. Как мембраны, так и клетки гладкой мышечной ткани окружены сетью эластических волокон, формирующих вместе с волокнами внутренней и наружной оболочек единый каркас, обеспечивающий. высокую эластичность стенки.
Наружная оболочка образована соединительной тканью, в которой преобладают пучки коллагеновых волокон, ориентированных продольно. В этой оболочке расположены и ветвятся сосуды, обеспечивающие питание как наружной оболочки, так и наружных зон средней оболочки.
Артерии мышечного типа. К разным по калибру артериям этого типа относится большинство артерий, доставляющих и регулирующих приток крови к различным частям и органам организма (плечевая, бедренная, селезеночная и др.). При микроскопическом исследовании в стенке хорошо различимы элементы всех трех оболочек (рис. 202).
Внутренняя оболочка состоит из трех слоев: эндотелиального, подэндотелиального и внутренней эластической мембраны. Эндотелий имеет вид тонкой пластинки, состоящей из вытянутых вдоль сосуда клеток с овальными, выступающими в просвет ядрами. Подэндотелиальный слой более развит в крупВнных по диаметру артериях и состоит из клеток звездчатой или веретенообразной формы, тонких эластических волокон и аморфного вещества, содержащего гликозаминогликаны. На границе со средней оболочкой лежит внутренняя эластическая мембрана, хорошо заметная на препаратах в виде блестящей, окрашенной эозином в светло-розовый цвет волнистой полоски. Эта мембрана пронизана многочисленными отверстиями, имеюВнщими значение для транспорта веществ.
Средняя оболочка построена преимущественно из гладкой мышечной ткани, пучки клеток которой идут по спирали, однако при изменении положения артериальной стенки (растяжении) расположение мышечных клеток может изменяться. Сокращение мышечной ткани средней оболочки имеет значение в регулировании притока крови к органам и тканям в соответствии с их потребностями и поддержании кровяного давления. Между пучками клеток мышечной ткани расположена сеть эластических волокон, которые вместе с эластическими волокнами подэндотелиального слоя и наружной оболочки формируют единый эластический каркас, придающий стенке упругость при ее сдавливании. На границе с наружной оболочкой в крупных артериях мышечного типа имеется наружная эластическая мемВнбрана, состоящая из плотного сплетения продольно ориентированных эластических волокон. В более мелких артериях эта мембрана не выражена.
Наружная оболочка состоит из соединительной ткани, в которой коллагеновые волокна и сети эластических волокон вытянуты в продольном направлении. Между волокнами располагаются клетки, преимущественно фиброциты. В наружной оболочке находятся нервные волокна и мелкие кровеносные сосуды, питающие наружные слои стенки артерии.

Рис. 202. Схема строения стенки артерии (А) и вены (Б) мышечного типа:
1 тАФ внутренняя оболочка; 2 тАФ средняя оболочка; 3 тАФ наружная оболочка; а тАФ эндотелий; б тАФ внутренняя эластическая мембрана; в тАФ ядра клеток гладкой мышечной ткани в средней оболочке; г тАФ ядра клеток соединительной ткани адвентиции; д тАФ сосуды сосудов.
Артерии мышечно-эластического типа по строению стенки занимают промежуточное положение между артериями эластического и мышечного типа. В средней оболочке в равном количестве развиты спирально ориентированная гладкая мышечная ткань, эластические пластины и сеть эластических волокон.
Сосуды микроциркуляторного русла. На месте перехода артериального русла в венозное в органах и тканях сформирована густая сеть мелких прекапиллярных, капиллярных и посткапиллярных сосудов. Этот комплекс мелВнких сосудов, обеспечивающий кровенаполнение органов, транссосудистый обмен и тканевый гомеостаз, объединяют термином микроциркуляторное русло. В его состав входят различные артериолы, капилляры, венулы и артериоло-венулярные анастомозы (рис. 203).
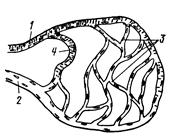
Рис. 203. Схема сосудов микроциркуляторного русла:
1 тАФ артериола; 2 тАФ венула; 3 тАФ капиллярная сеть; 4 тАФ артериоло-венулярный анастомоз
Артериолы. По мере уменьшения диаметра в артериях мыВншечного типа истончаются все оболочки и они переходят в артериолы тАФ сосуды диаметром менее 100 мкм. Внутренняя оболочка их состоит из эндотелия, расположенного на базальной мембране, и отдельных клеток подэндотелиального слоя. В некоторых артериолах может быть очень тонкая внутренняя эластическая мембрана. В средней оболочке сохраняется один ряд спирально расположенных клеток гладкой мышечной ткани. В стенке конечных артериол, от которых ответвляются капилляры, гладкомышечные клетки не образуют сплошного ряда, а расположены разрозненно. Это прекапиллярные артериолы. Однако в месте ответвления от артериолы капилляр окружен значительным количеством гладкомышечных клеток, которые образуют своеобразный прекапиллярный сфинктер. Вследствие изменения тонуса таких сфинктеров регулируется кровоток в каВнпиллярах соответствующего участка ткани или органа. Между мышечными клетками имеются эластические волокна. Наружная оболочка содержит отдельные адвентициальные клетки и коллагеновые волокна.
Капилляры тАФ важнейшие элементы микроциркуляторного русла, в которых осуществляется обмен газами и различными веществами между кровью и окружающими тканями. В большинстве органов между артериолами и венулами образуются ветвящиеся капиллярные сети, расположенные в рыхлой соединительной ткани. Плотность капиллярной сети в разных органах может быть различной. Чем интенсивнее обмен веществ в органе, тем гуще сеть его капилляров. Наиболее развита сеть капилляров в сером веществе органов нервной системы, в органах внутренВнней секреции, миокарде сердца, вокруг легочных альвеол. В скеВнлетных мышцах, сухожилиях, нервных стволах капиллярные сети ориентированы продольно.
Капиллярная сеть постоянно находится в состоянии переВнстройки. В органах и тканях значительное количество капилляров не функционирует. В их сильно уменьшенной полости циркулирует только плазма крови (плазменные капилляры). Количество открытых капилляров увеличивается при интенсифиВнкации работы органа.
Капиллярные сети встречаются и между одноименными сосудами, например венозные капиллярные сети в дольках печени, аденогипофизе, артериальные тАФ в почечных клубочках. Кроме образования разветвленных сетей, капилляры могут иметь форму капиллярной петли (в сосочковом слое дермы) или формировать клубочки (сосудистые клубочки почек).
Капилляры тАФ наиболее узкие сосудистые трубочки. Их калибр в среднем соответствует диаметру эритроцита (7тАФ8 мкм), однако в зависимости от функционального состояния и органной специализации диаметр капилляров может быть различным Узкие капилляры (диаметром 4 тАУ 5 мкм) в миокарде. Особые синусоидные капилляры с широким просветом (30 мкм и более) в дольках печени, селезенке, красном костном мозге, органах внутренней секреции.
Стенка кровеносных капилляров состоит из нескольких струкВнтурных элементов. Внутреннюю выстилку формирует слой эндотелиальных клеток, расположенных на базальной мембране, в последней содержатся клетки тАФ перициты. Вокруг базальной мембраны располагаются адвентициальные клетки и ретикулярные волокна (рис. 204).
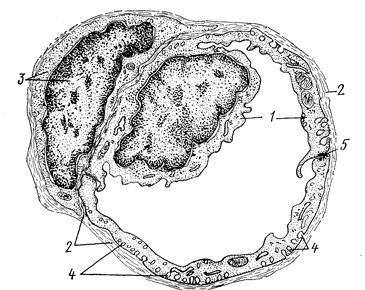
Рис. 204. Схема ультраструктурной организации стенки кровеносного капилВнляра с непрерывной эндотелиальной выстилкой:
1 тАФ эндотелиоцит: 2 тАФ базальная мембрана; 3 тАФ перицит; 4 тАФ пиноцитозные микропузырьки; 5 тАФ зона контакта между эндотелиальными клетками (рис. Козлова).
Плоские эндотелиальные клетки вытянуты по длине капилляра и имеют очень тонкие (менее 0,1 мкм) периферические безъядерные участки. Поэтому при световой микроскопии поперечного среза сосуда различима только область расположения ядра толщиной 3тАФ5 мкм. Ядра эндотелиоцитов чаще овальной формы, содержат конденсированный хроматин, сосредоточенный около ядерной оболочки, которая, как правило, имеет неровные контуры. В цитоплазме основная масса органелл расположена в околоядерной области. Внутренняя поверхность эндотелиальных клеток неровная, плазмолемма образует различные по форме а высоте микроворсинки, выступы и клапанообразные структуры. Последние особенно характерны для венозного отдела капилляров. Вдоль внутренней и наружной поверхностей эндотелиоцитов располагаются многочисленные пиноцитозные пузырьки, свидетельствующие об интенсивном поглощении и переносе веществ через цитоплазму этих клеток. Эндотелиальные клетки благодаря способности быстро набухать и затем, отдавая жидкость, уменьшаться по высоте могут изменять величину просвета капилляра, что, в свою очередь, влияет на прохождение через него форменных элементов крови. Кроме того, при электронной микроскопии в цитоплазме выявлены микрофиламенты, обусловливающие сократительные свойства эндотелиоцитов.
Базальная мембрана, расположенная под эндотелием, выявляется при электронной микроскопии и представляет плаВнстинку толщиной 30тАФ35 нм, состоящую из сети тонких фибрилл, содержащих коллаген IV типа и аморфного компонента. В последнем наряду с белками содержится гиалуроновая кислота, полимеризованное или деполимеризованное состояние которой обусловливает избирательную проницаемость капилляров. Базальная мембрана обеспечивает также эластичность и прочность капилляров. В расщеплениях базальной мембраны встречаются особые отросчатые клетки тАФ перициты. Они своими отростками охватывают капилляр и, проникая через базальную мембрану, формируют контакты с эндотелиоцитами.
В соответствии с особенностями строения эндотелиальной выстилки и базальной мембраны различают три типа капилляров. Большинство капилляров в органах и тканях принадлежит к первому типу (капилляры общего типа). Они характеризуются наличием непрерывных эндотелиальной выстилки и базальной мембраны. В этом сплошном слое плазмолеммы соседних эндотелиальных клеток максимально сближены и образуют соединения по типу плотного контакта, который непроницаем для макромолекул. Встречаются и другие виды контактов, когда края соседних клеток налегают друг на друга наподобие черепицы или соединяются зубчатыми поверхностями. По длине капилляров выделяют более узкую (5 - 7 мкм) проксимальную (артериолярную) и более широкую (8 - 10 мкм) дистальную (венулярную) части. В полости проксимальной части гидростатическое давление больше коллоидно-осмотического, создаваемого находящимися в крови белками. В результате жидкость фильтруется за стенку. В дистальной части гидростатическое давление становится меньше коллоидно-осмотического, что обусловливает переход воВнды и растворенных в ней веществ из окружающей тканевой жидВнкости в кровь. Однако выходной поток жидкости больше входного, и избыточная жидкость в качестве составной части тканевой жидкости соединительной ткани поступает в лимфатическую систему.
В некоторых органах, в которых интенсивно происходят процессы всасывания и выделения жидкости, а также быстрый транспорт в кровь макромолекулярных веществ, эндотелий капилляров имеет округлые субмикроскопические отверстия диаметром 60тАФ 80 нм или округлые участки, затянутые тонкой диафрагмой (почки, органы внутренней секреции). Это капилляры сфенестрами (лат. fenestrae тАФ окна).
Капилляры третьего типа тАФ синусоидные, характеризуются большим диаметром своего просвета, наличием между эндотелиальными клетками широких щелей и прерывистой базальной мембраной. Капилляры этого типа обнаружены в селезенке, красном костном мозге. Через их стенки проникают не только макромолекулы, но и клетки крови.
Венулы тАФ отводящий отдел микропиркуляторного русла и начальное звено венозного отдела сосудистой системы. В них соВнбирается кровь из капиллярного русла. Диаметр их просвета боВнлее широкий, чем в капиллярах (15тАФ50 мкм). В стенке венул, так же как и у капилляров, имеется слой эндотелиальных клеВнток, расположенных на базальной мембране, а также более выраженная наружная соединительнотканная оболочка. В стенках венул, переходящих в мелкие вены, находятся отдельные гладкие мышечные клетки. В посткапиллярных венулах тимуса, лимфатических узлов элдотелиальная выстилка представлена высокими эндотелиальными клетками, способствующими избирательной миграции лимфоцитов при их рециркуляции. В венулах вследствие тонкости их стенки, медленного кровотока я низкого кровяного давления может депонироваться значительное количество крови.
Артериоло-венулярные анастомозы. Во всех органах обнаружены трубочки, по которым кровь из артериол может направляться непосредственно в венулы, минуя капиллярную сеть. Особенно много анастомозов в дерме кожи, в ушной раковине, гребне птиц, где играют определенную роль в терморегуляции.
По строению истинные артериоло-венулярные анастомозы (шунты) характеризуются наличием в стенке значительного количества продольно ориентированных пучков из гладких мышечных клеток, расположенных или в подэндотелиальном слое интимы (рис. 205), или во внутренней зоне средней оболочки. В некоторых анастомозах эти клетки приобретают эпителиоподобный вид. Продольно расположенные мышечные клетки находятся и в наружной оболочке. Встречаются не только простые анастомозы в виде единичных трубочек, но и сложные, состоящие из нескольких ветвей, отходящих от одной артериолы и окруженных общей соединительнотканной капсулой.
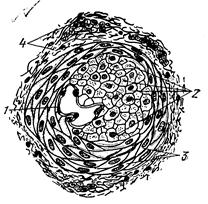
Рис. 205. Артериоло-венулярный анастомоз:
1 тАФ эндотелий; 2 тАФ продольно расположенные эпителиоидно-мышечные клетки; 3 тАФ циркулярно расположенные мышечные клетки средней оболочки;
4 тАФ наружная оболочка.
При помощи сократительных механизмов анастомозы могут уменьшить или полностью закрыть свой просвет, в результате чего течение крови через них прекращается и кровь поступает в капиллярную сеть. Благодаря этому органы получают кровь в зависимости от потребности, связанной с их работой. Кроме того, высокое давление артериальной крови через анастомозы передается в венозное русло, способствуя этим лучшему пере движению крови в венах. Значительна роль анастомозов в обогащении венозной крови кислородом, а также в регуляции кровообращения при развитии патологических процессов в органах.
Вены тАФ кровеносные сосуды, по которым кровь из органов и тканей течет к сердцу, в правое предсердие. Исключение составляют легочные вены, направляющие кровь, богатую кислородом, из легких в левое предсердие.
Стенка вен, так же как и стенка артерий, состоит из трех оболочек: внутренней, средней и наружной. Однако конкретное гистологическое строение этих оболочек в различных венах очень разнообразно, что связано с различием их функционирования и местными (в соответствии с локализацией вены) условиями кровообращения. Большинство вен одинакового диаметра с одноименными артериями имеет более тонкую стенку и более широкий просвет.
В соответствии с гемодинамическими условиями тАФ низким кровяным давлением (15тАФ20 мм рт. ст.) и незначительной скоростью кровотока (около 10 мм/с) тАФ в стенке вен сравнительно слабо развиты эластические элементы и меньшее количество мышечной ткани в средней оболочке. Эти признаки обусловливают возможность изменения конфигурации вен: при малом кровенаВнполнении стенки вен становятся спавшимися, а при затруднении оттока крови (например, вследствие закупорки) легко происхоВндят растяжение стенки и расширение вен.
Существенное значение в гемодинамике венозных сосудов имеют клапаны, расположенные таким образом, что, пропуская кровь по направлению к сердцу, они преграждают путь ее обратному течению. Число клапанов больше в тех венах, в которых кровь течет в направлении, обратном действию силы тяжести (например, в венах конечностей).
По степени развития в стенке мышечных элементов различают вены безмышечного и мышечного типов.
Вены безмышечного типа. К характерным венам данного типа относят вены костей, центральные вены печеночных долек и трабекулярные вены селезенки. Стенка этих вен состоит только из слоя эндотелиальных клеток, расположенных на базальной мембране, и наружного тонкого слоя волокнистой соединительной ткани С участием последней стенка плотно срастается с окружающими тканями, вследствие чего эти вены пассивны в продвижении по ним крови и не спадаются. Безмышечные вены мозговых оболочек и сетчатки глаза, наполняясь кровью, способВнны легко растягиваться, но в то же время кровь под действием собственной силы тяжести легко оттекает в более крупные венозные стволы.
Вены мышечного типа. Стенка этих вен, подобно стенке артерий, состоит из трех оболочек, однако границы между ними меВннее отчетливы. Толщина мышечной оболочки в стенке вен разной локализации неодинаковая, что зависит от того, движется кровь в них под действием силы тяжести или против нее. На основании этого вены мышечного типа подразделяют на вены со слабым, средним и сильным развитием мышечных элементов. К венам первой разновидности относят горизонтально расположенные вены верхней части туловища организма и вены пищеварительВнного тракта. Стенки таких вен тонкие, в их средней оболочке гладкая мышечная ткань не образует сплошного слоя, а расположена пучками, между которыми имеются прослойки рыхлой соединительной ткани.
К венам с сильным развитием мышечных элементов относят крупные вены конечностей животных, по которым кровь течет вверх, против силы тяжести (бедренная, плечевая и др.). Для них характерны продольно расположенные небольшие пучки клеток гладкой мышечной ткани в подэндотелиаль
Вместе с этим смотрят:
20 мешков картошки с 1 сотки
Cовершенствование организации производства молока на КПУП "ГМЗ № 3"
Cостояние полезащитных лесных полос в северном Приднестровье
РЖнтегрований захист озимоi пшеницi
РЖнтенсифiкацiя та ii економiчна ефективнiсть у сiльському господарствi