Усовершенствование метода окраски вкусовых луковиц ионофорезом
Вкус наряду со зрением, осязанием, слухом и обонянием является одним из пяти основных чувств, участвующих в восприятии информации об окружающем нас мире. Основной функцией вкусовой системы является оценка качества пищи, проверка ее съедобности. Так, в ходе эволюции млекопитающими была развита высокая чувствительность к горькому, так как большинство горьких веществ являются ядовитыми, с другой стороны, имеющие привлекательный (т.е., сладкий) вкус вещества являются высококалорийными. Считается, что человек способен различать пять базовых вкусов: кислый, соленый, сладкий, горький и умами (umami), который вызывается глютаматом и некоторыми другими аминокислотами. Поведенческие эксперименты, а также регистрации активности вкусовых нервных волокон свидетельствуют о том, что вкусовой анализатор животных так же способен различать эти базовые вкусовые модальности. Это предполагает существование специфических молекулярных структур на рецептирующей поверхности вкусовых клеток, которые отвечают за отдельное распознавание каждого вкусового стимула. Исследования вкусовой системы методами молекулярной биологии подтверждают, что это действительно так. В частности, в последние годы идентифицированы мембранные рецепторы для сладких и горьких веществ и аминокислот, вызывающих умами (umami) вкус. Тем не менее, молекулярные механизмы вкуса во многом не ясны, поскольку во вкусовой клетке ни для одного из вкусовых стимулов не прослежена вся последовательность событий от взаимодействия с молекулярным рецептором до выброса нейромедиатора.
Одной из задач физиологии вкусового органа, которую предстоит решить, является исследование межклеточных коммуникации во вкусовой почке. Популяция вкусовых клеток гетерогенна. Во вкусовой почке идентифицировано три морфологически различных типа веретенообразных клеток, функциональная роль которых доподлинно неизвестна, но которые, по-видимому, выполняют рецепторную, поддерживающую и/или секреторную функции. Эти клетки обмениваются примерно раз в двадцать дней, развиваясь из клеток предшественников и в конце жизни подвергаясь апоптозу. Поскольку вкусовые клетки устанавливают афферентные синапсы с вкусовыми нервами, то их непрерывное обновление требует постоянного установления новых синаптических связей во вкусовой почке. Кроме того, подобно тому, как это происходит в сетчатке или обонятельной луковице, сенсорная информация также может, подвергается первичной обработке во вкусовой почке. Протекание всех этих гетерогенных, но синхронизированных процессов, несомненно, требует хорошо отлаженных коммуникаций между клетками вкусовой почки. "Ряд фактов, установленных в последние годы, свидетельствуют в пользу подобной точки зрения. В частности, присутствие сигнальных молекул нескольких типов (серотонин, ацетилхолин, ГАМК, субстанция Р) во вкусовой почке было показано иммуногистохимически, в то время как присутствие во вкусовых клетках рецепторов к ним было показано методами молекулярной биологии и электрофизиологии.
2. Вкусовая сенсорная система
Качества и концентрации веществ, поступающих в ротовую полость, осуществляется с помощью вкусовой сенсорной системы. Следовательно, вкусовая сенсорная система является одним из каналов связи организма с внешней средой. При участии данной сенсорной системы не только потребляется или отвергается организмом различные вещества, но и обеспечиваются некоторые вегетативные реакции (слюноотделение, секреция желудочного сока, изменение частоты дыхания и сердечных сокращений и и.д.), а также сложные поведенческие реакции, связанные с поиском и потреблением питательных веществ.
Периферическую часть вкусовой сенсорной системы составляют специфические образования: вкусовые луковицы, локализированные в определенных местах в толще эпителия слизистой оболочки начального отдела пищеварительного тракта тАУ в грибовидных, листовидных и желобоватых сосочках языка, в слизистой губ, слизистой твердого и мягкого неба, задней стенке глотки, небных дужек, а также в начальных отделах дыхательных путей тАУ в эпителии надгортанника, в слизистой области черпаловидных хрящей гортани и даже в эпителии голосовых связок. Общее число вкусовых луковиц у взрослого человека достигает 9 тАУ 10 тыс. с возрастом часть вкусовых луковиц атрофируется. Основную роль во вкусовой рецепции выполняют вкусовые луковицы сосочков языка.
Вкусовую луковицу можно рассматривать как сложный рецепторный прибор, состоящий из рецепторных и опорных клеток, а также концевых ветвлений нервных волокон. Установлена тесная связь вкусовых луковиц и нервных элементов. При перерезке нервных волокон вкусовые луковицы дегенерируют. Каждая вкусовая почка представляет собой округлое, эллипсоидное или колбообразное образование диаметром около 70 мкм, состоящее из 30-80 уплощенных веретенообразных клеток, которые своими основаниями располагаются на базальной мембране эпителия, а апикальными частями выстоят во вкусовую ямку. Сама вкусовая ямка или вкусовой канал располагается во вкусовой почке ближе к свободной поверхности эпителия и заполнена жидкостью, в которой содержаться белки, мукопротеиды и определяется высокая активность щелочной фосфатазы. Вкусовая ямка открывается через вкусовую пору в ротовую полость. Через эту пору во вкусовую ямку попадают уже растворенные в слюне пищевые вещества. Полагают, что растворенные пищевые вещества образуют комплексы с белками и мукопротеидами жидкости вкусовой ямки, что способствует их адсорбции на мембранах рецепторных клеток. Следовательно, жидкость вкусовой ямки принимает определенное участие в рецепции пищевых стимулов.
Что касается клеток вкусовых луковиц, то большинство исследователей этой области выделяют две их разновидности тАУ рецепторные и опорные. Некоторые авторы описывают и третий тип тАУ переходные клетки. Наконец, приводятся описание четырех типов клеток вкусовой луковицы: рецепторные, опорные, базальные и перигемальные. Эти расхождения в описании разновидностей клеток вкусовых луковиц обусловлены тем, что вкусовые клетки являются одними из самых быстро сменяющихся клеток в организме. Продолжительность их жизни составляет 10-12 дней. На периферии вкусовых луковиц отмечается интенсивное митотическое деление клеток и перемещение вновь образованных клеток к центру луковиц со скоростью 0,06 мкм в час. Вследствие этого клетки вкусовых луковиц находятся в разных стадиях развития тАУ пролиферирующие, дифференцирующие, зрелые, дегенерирующие. Вместе с тем в составе вкусовых луковиц четко определяются два различных по морфологии, а возможно и по функции, типа клеток тАУ темные и светлые, первые из которых расцениваются как рецепторные, вторые тАУ опорные.
Рецепторные клетки имеют призматическую форму высотой 10-20 мкм и шириной 3-4 мкм, более темные при окраске и содержат на апикальных поверхностях 30-40 микроворсин, выступающих в полость вкусовой ямки. Предполагается, что именно на мембранах микроворсин, находятся рецепторные тАУ стереоспецифические участки, образованные рецепторными белками или простетическими группами, избирательно адсорбирующие вещества, попадающие во вкусовую ямку. Установлено, что мембраны микроворсин тоньше (9нм) мембран самой рецепторной клетки (12-13 нм). Ядра рецепторных клеток располагаются в базальных отдела, т. е. ближе к базальной мембране. В цитоплазме апикальных отделов клеток содержится много митохондрий, хорошо развитая гладкая эндоплазматическая сеть. Здесь же определяется высокая активность некоторых дегидрогеназ и фосфатаз. На боковые поверхности клеток выявляется многочисленные афферентные нервные окончания. Эфферентных синапсов на вкусовых клетках не обнаружено.
Опорные клетки располагаются между рецепторными и изолируют друг от друга. Цитоплазма опорных клеток вакуолизирована и поэтому при окраске более светлая. В ней содержится хорошо развитая гладкая и зернистая эндоплазматическая сеть, комплекс Гольджи. Ядра клеток крупные. Активность дегидрогеназ и фосфатаз невысокая. Апикальные отделы опорных клеток также выступают во вкусовые ямки и несут на поверхности микровыросты, которые значительно короче микроворсин рецепторных клеток. Считается, что эти клетки, помимо опорной, выполняют секреторную функцию. По-видимому, жидкость ямок вкусовых луковиц частично является продуктом деятельности опорных клеток, а так как ее компоненты принимают участие во вкусовой рецепции, то, следовательно, и опорные клетки косвенно принимают участие в восприятии вкуса.
Несмотря на указанные морфологические различия, пока е решен вопрос тАУ являются опорные и рецепторные клетки дефинитивными специфическими клетками или же они представляют собой только разные стадии развития единой эпителиальной клетки тАУ предшественницы. R. Murray (1967) рассматривал светлые клетки с вакуолизированной цитоплазмой как дегенерирующие рецепторные клетки. Однако, не вызывает сомнений тот факт, что рецепторные клетки развиваются из эпителиальных клеток и, следовательно. Относятся к группе вторичночувствующих сенсоэпителиальных рецепторов.
Как известно, вторичночувствующие рецепторы воспринимают специфические раздражения, генерируют рецепторный потенциал и передают его на нервные окончания, где и осуществляется кодирование в виде нервных импульсов. Поэтому представляет интерес рассмотреть взаимоотношение клеток и нервных элементов во вкусовых луковицах. В области основания вкусовых луковиц, т. е. под базальной мембраной эпителия, в котором заложены луковицы, располагается хорошо выраженное нервное сплетение, образованное тонкими миелиновыми и безмиелиновыми волокнами. Из этого сплетения во вкусовые луковицы проникают безмиелиновые, тонкие и более толстые миелиновые нервные волокна. Каждое тонкое волоконце заканчивается на одной рецепторной клетке, а более толстые разветвляются и заканчиваются на нескольких рецепторных клетках. Каждая рецепторная клетка получает иннервацию от нескольких (до 30) нервных волокон, т. е. отмечается перекрестная иннервация одной рецепторной клетки из нескольких нервных волокон. Кроме того, толстые миелиновые волокна, разветвляясь, иннервируют не только несколько рецепторных клеток в одной вкусовой луковице, но и несколько вкусовых луковиц. Что касается синоптических приборов на клетках вкусовых луковиц, то здесь нет полной ясности. Пока не определено, находятся они только на рецепторных или же на опорных клетках, так как на основании строения синоптических контактов нельзя отдифференцировать, какие клетки вкусовой луковицы являются рецепторными, а какие тАУ опорными.
Механизмы восприятия вкусовых стимулов рецепторными клетками, т. е. процессы трансформации химической энергии в нервные импульсы, также не вполне выяснены. Предположены несколько теорий вкусовой рецепции, одна из которых приводиться ниже. Различают четыре основных вкусовых ощущений: сладкого, кислого, соленого и горького. Достоверно установлено, что рецепторные клетки, так же как и вкусовые луковицы не обладают моноспецифичностью вкусового восприятия, т. е. являются для вкусовых стимулов полимодальными. Каждая вкусовая клетка и каждая луковица воспринимает любой из четырех вкусовых стимулов, но с предпочтением одного из них. Вместе с тем выявлена определенная предпочтительность восприятия отдельных вкусовых стимулов вкусовыми луковицами, локализированными в разных отделах слизистой оболочки ротовой полости. Так, ощущение сладкого и соленого с наибольшей чувствительностью воспринимается рецепторами вкусовых луковиц слизистой языка, а кислого и горького тАУ рецепторами вкусовых луковиц области неба.
Предполагается, что процесс восприятия вкусовых стимулов является многоступенчатым. Вначале пищевые вещества адсорбируются на мембранах микроворсин рецепторных клеток, предварительно образуя комплексы с белками и мукопротеидами жидкости вкусовых ямок. Активные группировки пищевых веществ вступают в межмолекулярные, атомарные или ионные взаимодействия с рецепторными участками мембран. Количество рецептивных участков в мембранах микроворсин и качество белков-рецепторов в разных вкусовых клетках неодинаково. Вкусовые вещества вступают во взаимодействие с белками-рецепторами нескольких рецептивных участков данной клетки. В результате такого взаимодействия наступают конформационные изменения рецепторных белков, что сопровождается нарушением ионной проницаемости мембран рецепторных клеток и возникновением в них рецепторных потенциалов. По другим данным, конформационные изменения рецепторных белков включает цепь ферментативных реакций, которые приводят к метаболическим сдвигам в рецепторных клетках и возникновению в них рецепторного потенциала. Следует отметить, что разные пищевые вещества взаимодействуют не только с разными рецептивными участками мембран, но и с разным их количеством. Так, для молекул, обладающих горьким вкусом, установлено наличие 3-х реакционноспособных группировок, вступающих во взаимодействие с определенными рецептивными участками мембран микроворсин рецепторных клеток. Подтверждением наличия специфических рецепторных белков в мембранах вкусовых клеток является факт выделения из слизистой языка особых сладкочувствительных и горькочувствительных белков. Однако в последнее время специфичность таких белков подвергается сомнению, так как они выделены не только из вкусовых клеток, но и из обычных покровных эпителиальных клеток языка.
Третьим этапом вкусового восприятия можно рассматривать процесс передачи рецепторного потенциала на чувствительные нервные окончания, т. е. процесс кодирования специфического стимула в нервные импульсы. Механизм этого процесса еще менее ясен. Даже морфология синапсов на вкусовых клетках недостаточно выяснена.
Последнее тем более важно, что, как указывалось ранее, продолжительность жизни вкусовых клеток незначительна, и, следовательно, должны наблюдаться процессы дегенерации синапсов и формирование новых на дифференцирующихся клетках.
Заканчивая описание периферической части вкусовой сенсорной системы, можно констатировать, что во вкусовых луковицах происходит восприятие вкусовых стимулов ( будь то ионы натрия и хлора тАУ для кислых веществ ионы водорода тАУ для соленых, а также более сложные молекулы большинства пищевых веществ) и кодирование их в нервные импульсы.
Прежде чем перейти к изложению промежуточной чести вкусовой сенсорной системы следует четко уяснить, что различают два вида чувствительной иннервации слизистой оболочки ротовой полости, глотки и гортани тАУ обычная чувствительность (тактильная, температурная, болевая) и специфическая (вкусовая), которая иннервирует только вкусовые луковицы. Предметом изучения в данном случае является только второй вид чувствительности. Однако вкусовая чувствительность сочетается с общей чувствительность сочетается с общей чувствительностью и придает определенный оттенок вкусовым ощущением. Наконец, описаны случаи врожденного отсутствия вкусовых луковиц, однако вкусовые ощущения у таких людей имеются. Полагают, что вкусовые восприятия в этих случаях обеспечиваются кустиковидными рецепторами.
Понимание и запоминание промежуточного отдела вкусовой сенсорной системы вызывает определенные затруднения в связи с тем, что вкусовые луковицы определенных областей ротовой полости, глотки и гортани иннервируются несколькими чувствительными нервами, вернее их ветвями: лицевого (промежуточного), языкоглоточного и блуждающего, а, следовательно, и чувствительные нейроны, обеспечивающие вкусовую иннервацию, расположены в разных узлах.
Вкусовые луковицы передних двух третей языка получают афферентную вкусовую иннервацию из чувствительных нейронов узла коленца (gangl.geniculi) , который расположен в канале лицевого нерва в пирамиде височной кости. Дендриты этих нейронов идут на периферию в составе барабанной струны, которая затем примыкает к язычному нерву достигает вместе с ним вкусовых луковиц передних двух третей языка. Здесь концевые ветвления дендритов и образуют синапсы на телах вкусовых клеток. Центральные отростки этих чувствительных нейронов входят в продолговатый мозг в составе промежуточного нерва и заканчивается в ростральной части ядра солитарного пучка (nucl. Tractus solitarius). Поскольку промежуточный нерв топографически связан с лицевым нервом, то и барабанную струну часто рассматривают как ветвь лицевого нерва. Барабанная струна, помимо чувствительных вкусовых волокон, содержит также преганглиозные секреторные волокна, прерывающиеся в подъязычную и подчелюстную железы. Неспецифическая чувствительность слизистой оболочки передних двух третей языка обеспечивается язычным нервом тАУ чувствительной ветвью тройничного нерва.
Вкусовая и общая чувствительная иннервация задней трети языка, твердого и мягкого неба, миндалин и задней глотки обеспечивается чувствительными волокнами языкоглоточного нерва (IX пара). Чувствительные нейроны, обеспечивающие эту область иннервации, локализованы в нижнем узле этого нерва. Их дендриты заканчиваются синапсами на вкусовых клетках луковиц и в эпителии слизистой, а аксоны входят в продолговатый мозг в составе ствола языкоглоточного нерва и заканчиваются в средней части ядра солитарного (одиночного) пучка. Вкусовая и общая чувствительная иннервация гортани и частично глотки осуществляется чувствительными волокнами блуждающего нерва (Х пара), достигающих этой области в составе верхнего гортанного нерва. Тела чувствительных нейронов локализуются в нижнем узле блуждающего нерва. Дендриты их заканчиваются на телах рецепторных клеток вкусовых луковиц и в слизистой оболочке названных отделов, а аксоны в составе ствола блуждающего нерва достигают каудальной части все того же ядра одиночного пучка.
Таким образом, вкусовая иннервация слизистой оболочки ротовой полости, глотки и гортани обеспечивается ветвями трех пар черепно-мозговых нервов тАУ VII, IX, X, а первые нейроны вкусового пути локализированы в узле коленца, в нижних узлах блуждающего и языкоглоточного нервов. Чувствительные ветви блуждающего и языкоглоточного нервов, как отмечалось выше, обеспечивают в указанных областях не только вкусовую, но и специфическую иннервацию. При этом пока не установлено, осуществляются разные виды иннервации одним или разными типами чувствительных клеток нижних узлов блуждающего и языкоглоточного нервов.
Интерес представляет тот факт, что вкусовая чувствительность из разных зон слизистой оболочки по разным нервам сходится (конвергирует) в одном ядре тАУ ядре одиночного пучка, в котором и расположены 2-е нейроны вкусового пути. Кроме того, в это же ядро по чувствительным волокнам блуждающего и языкоглоточного нервов поступает сенсорная информация почти из всех органов пищеварительной и дыхательной систем, а по волокнам тройничного нерва тАУ от головы и органов ротовой полости. Следовательно, ядро солитарного пучка получает разные виды чувствительности из разных органов и поэтому достигает больших размеров. Оно простирается по дорзальной поверхности продолговатого мозга м моста и состоит из мелких клеток, напоминающих по своей морфологии и расположению желатинозную субстанцию спинного мозга. Вкусовые волокна заканчиваются на нейронах передней части ядра, а волокна общей чувствительности - в каудальной части. Электрофизиологически установлено, что нейроны вкусовой части ядра неоднородны по своей функции. Одни из них отвечают импульсацией только на раздражение строго определенными вкусовыми раздражителями, другие реагируют на несколько из 4-х классических вкусовых раздражителей. Солитарное ядро является первичным центром, в котором осуществляется обработка вкусовой сенсорной информации.
Аксоны 2-х нейронов по выходе из ядра солитарного пучка идут через ретикулярную формацию на противоположную сторону, входят в состав медиальной петли и оканчиваются, главным образом, на нейронах аркуатного ядра зрительного бугра, в непосредственной близости с центрами соматосенсорной системы лица и ротовой полости. Часть аксонов этого пути заканчивается в сосочковых (мамиллярных) телах. Аксоны 3-х нейронов (главном образом аркуатного ядра) достигают корковых вкусовых центров. По данным Н.А. Ибадова (1985), часть аксонов 1-х нейронов не переключаясь достигает корковых вкусовых центров и даже полушарий и червя мозжечка.
Относительно локализации в коре вкусового центра в литературе приводятся противоречивые данные тАУ в гиппокампе, в парагиппокамповой извилине, в крючке, в нижних отделах пре- и постцентральной извилин, в параинсулярной и оперкулярной (покрышковой) областях (после 43). В последние годы предпочтение отдается последним двум областям. Основанием этому служат экспериментальные исследования с разрушением инсуло-оперкулярной области коры, в результате чего установлена дегенерация вентромедиальных ядер зрительного бугра. В клинических условиях установлено нарушение вкусовой чувствительности у больных после удаления оперкулярной области коры.
Однако не вся вкусовая сенсорная информация передается в корковые центры. Часть ее по выходе из ядра солитарного пучка передается, по видимому, через ретикулярную формацию на пищевой, дыхательной и сосудодвигательный центры, в частности, на ядра блуждающего и языкоглоточного нервов, и обуславливает такие акты, как рвоту, глотание, изменение частоты дыхания и сердцебиений.
Что касается саморегуляции вкусовой сенсорной системы, Ио она несомненно имеется, но механизмы ее почти не выяснены. Вообще, белых пятен в структурно-функциональной организации вкусовой сенсорной системы немало и потому объяснение многих сторон ее деятельности строится на предположениях. Достоверно установлено, что острота вкусовых восприятий зависит от состояния рецепторов желудка. Доказано, что число активно функционирующих вкусовых луковиц у людей в голодном состоянии значительно больше, чем в сытом. Эти данные свидетельствуют о наличии центробежных механизмов регуляции вкусовой сенсорной системы, в том числе регуляции ее периферической части. Однако, как передаются эти влияния на периферическую часть, пока не ясно, так как в составе барабанной струны не обнаружено эфферентных нервных волокон, передающих импульсы на рецепторные вкусовые клетки, а на самих рецепторных клетках не обнаружены эфферентные синапсы. Высказываются предположения, что эфферентные влияния на вкусовые рецепторы осуществляются через симпатическую нервную систему, но передаются непосредственно на вкусовые клетки, а на нервное сплетение под основаниями вкусовых луковиц.
В периферической части обонятельной сенсорной системы имеет место латеральное торможение тАУ при возбуждении одной рецепторной клетки часть других клеток тормозится. Это обусловлено тем, что одно чувствительное волокно, разветвляясь на многочисленные терминали, вступает в синаптические связи с большим числом рецепторных клеток. При возбуждении одной рецепторной клетки в данном нервном окончании возникают нервные импульсы, которые проводятся антидромно по всем разветвлениям этого волокна и блокируют образование импульсов во всех других терминалях. Механизмы латерального торможения более обстоятельно будут рассмотрены при изложении кожной части соматосенсорной системы.
Механизмы центрального торможения во вкусовой сенсорной системе практически не выяснены. В литературе указывается наличие центробежных эфферентных влияний из коры на клетки аркуатного ядра зрительного бугра. Что касается эфферентных влияний на нейроны ядра солитарного пучка, а тем более на чувствительные вкусовые нейроны (в частности узла коленца), то они пока не обнаружены.
Наконец, имеются данные, что некоторые пищевые вещества, например глюкоза, всосавшись в кровь в ротовой полости, буквально за несколько секунд достигают вкусовых центров и оказывают на их клетки определенное влияние, а исходящие из этих центров эфферентные импульсы влияют на промежуточные центры, а возможно и на периферическую часть (через симпатическую нервную систему), изменяя порог чувствительности вкусовых клеток и их адаптации к данному раздражителю.
В заключение следует отметить. Что ощущение вкуса вызывается не только действием четырех основных вкусовых стимулов на вкусовые клетки, но также действием на рецепторы прикосновения, температуры, давления, запаха. Следовательно, целостное ощущение вкуса является результатом совместного функционирования нескольких сенсорных систем вкусовой, экстероцептивной, интероцептивной и обонятельной.
3. Трансдукция вкусового сигнала
Основополагающей функцией вкусовых рецепторных клеток является распознавание вкусовых веществ (ионов и молекул, содержащихся в пище) - трансдукция - и кодирование информации об их концентрации и вкусовой модальности для дальнейшего анализа в соответствующих структурах мозга (Gilbertson et al., 2000; Herness and Chen, 2000). Для трансдукции вкусовых стимулов в настоящий момент общепризнанна множественность механизмов, которые можно разбить на две условные группы. Во-первых, предполагается, что при воздействии солёных и кислых стимулов для образования и передачи сигнала во вкусовой клетке достаточно модуляции ионных потоков через апикальную мембрану, обусловленной изменением ионного состава среды (Na+, в случае солёного; Н*, в случае кислого стимула) во вкусовой поре (Lindemann, 1996; Herness and Gilberson, 1999). Во-вторых, большинство сладких и горьких веществ (а так же ряд аминокислот) активируют хеморецепторный путь передачи сигнала. Предполагается, что в этом случае внешний стимул связываясь с трансмембранным G-белок связывающимся рецептором (GPCR, G-protein coupled receptor) (Lindemann, 2001), переводит его в возбужденное состояние, находясь в котором рецептор катализирует диссоциацию нескольких сотен гетеротримерных G-белков на а-субъединицу и комплекс Ру-субъединиц. Каждый из них может регулировать активность ионных каналов и/или эффекторных ферментов, которые генерируют внутриклеточные сигналы, что в свою очередь приводит к передаче вкусового сигнала далее от вкусовой клетки. (Herness and Gilberson, 1999; Lindemann 2001; Margolskee 2002). Следует отметить, что прогресс последнего десятилетия в области молекулярной биологии, иммуногистохимии и электрофизиологии вкусового органа позволил идентифицировать многие эффекторные молекулы его сигнальных каскадов. Так, одними из первых были идентифицированы восемь а-субъединиц G-белков, которые являются своего рода визитной карточкой сигнального каскада. Наиболее представлено семейство Gi/Go, четыре представителя которого экспрессируются во вкусовых клетках: а-гастдуцин, а-трансдуцин. О гастдуцине: поведенческие эксперименты и регистрации от вкусового нерва показывают, что мыши, у которых была подавлена (knock out) экспрессия гастдуцина, оказались на два порядка менее чувствительны к горьким и сладким (но не к соленым и кислым) веществам по сравнению с мышами дикого типа (Wong et al., 1996). Это однозначно говорит о принципиальной роли гастдуцина в формировании горького и сладкого вкуса. Методом single cell RT-PCR анализировалась экспрессия субъединиц G-белков, и было показано, что гастдуцин, скорее всего, состоит из а-гастдуцина и Ру-субъединиц (Huang et al., 1999), причем а-субъединица, скорее всего, активирует фосфодиэстеразу (PDE), а Ру-комплекс контролирует фосфолипазу С (PLC) (Margolskee, 2002). Гастдуцин экспрессируется преимущественно в клетках типа II (Boughter
et al., 1997; Sbarbati et al., 1999; Smith et al., 1999; Yang et al., 2000b), что является необходимым условием для нормального рецептирования сладких и горьких стимулов (Wong et al., 1995; Ruiz-Avila et al., 2001). Поэтому гасдуцин-положительные клетки типа II могут быть хеморецепторными, однако они не формируют синапсы с эфферентными волокнами; возможно, они секретируют сигнальные молекулы, чтобы, например, модулировать активность соседних вкусовых клеток.
Эти и перечисленные выше данные говорят о том, что потенциально возбуждение вкусовых клеток может происходить путем модуляции активности ионных каналов G-белками или в результате изменения внутриклеточной концентрации циклических нуклеотидов, или за счет генерации инозитолтрифосфата (1Р3) и мобилизации Са2+ (Ogura et al., 2002). Действительно, многие элементы циклонуклеотидного и фосфоинозитидного каскадов найдены во вкусовых клетках, и в частности: аденилатциклаза (AC) (Abaffy, et al., 2003), PDE (Kinnamon and Margolskee, 1996), циклонуклеотид-зависимые каналы (Kaupp and Seifert, 2002, Wei et al., 1998), PLC (Asano-Miyoshia et al., 2000, Rossler et al., 1998), 1Р3-рецепторы (Yan et al., 2001) и TRP-каналы (Zhang, et al., 2003). Однако вклад этих каскадов в возбуждение вкусовых клеток химическими стимулами фактически не исследован и мало известно о том, какие рецепторы контролируют активность этих сигнальных систем.
В 2000г было идентифицировано семейство генов кодирующих так называемые T2R/TRB-penenTopbi (Adler et al, 2000), специфически экспрессирующиеся во вкусовых клетках, и, по-всей видимости, функционирующие как вкусовые рецепторы (Lindemann, 2001). Со структурной точки зрения, T2R/TRB рецепторы лишь отдаленно связаны с другими GPCR, такими как VIR (рецепторы на феромоны), и даже внутри семейства степень гомологии составляет 30-70%. Эти рецепторы имеют высококонсервативные области в цитоплазматических петлях и прилежащих к ним трансмембранных сегментах (предположительно они выполняют функцию взаимодействия с G-белками) и сильно дивергирующие экстраклеточные области (потенциальные сайты связывания с лигандами) (Gilbertson, et al., 2000). У крыс и мышей T2R/TRB рецепторы экспессируются в 15 - 20% вкусовых рецепторных клеток желобоватых и листовидных сосочков и в очень малом количестве вкусовых рецепторных клеток грибовидных сосочков (Margolskee, 2002). Основываясь на данных in situ гибридизации, показано, что T2R/TRB рецепторы экспрессируются в определенных типах вкусовых рецепторных клеток (Adler et al., 2000). В частности, T2R/TRB рецепторы экспрессируются в клетках экспрессирующих гастдуцин (Gilbertson, et al., 2000; Margolskee, 2002). В биохимических экспериментах было показано, что T2R5 - рецептор, распознающий горькое вещество циклогексимид, селективно активирует гастдуцин, но не другие G-белки, экспрессируемые во вкусовых клетках (Chandrachekar et al., 2000). Это дает основание думать, что именно гастдуцин связывает T2R5 и возможно другие рецепторы к горьким веществам с эффекторными ферментами. Было также показано, что горькие вещества стимулируют синтез 1Р3 и что этот ответ блокируется антителами к Ру-субъединицам гастдуцина и к РЬСРг в. то время как контрольные антитела и антитела к а-гастдуцину были неэффективны (Rossler et al., 2000). Это говорит о том, что после рецептор-стимулируемой диссоциации гастдуцина именно ру-комплекс активирует PLCp2, запуская тем самым образование 1Р3 и диацилглицерола (DAG).
Методами биоинформатики в геноме мыши был найден новый рецептор, T1R3, который был идентифицирован как продукт гена из локуса sac на четвертой хромосоме мыши (Max et al., 2001). Поскольку мутации в этом гене ведут к потере чувствительности к сахарину и не затрагивают чувствительность к веществам других вкусовых модальностей, было предположено, что T1R3 - сладко-чувствующий рецептор. Он имеет гомологию порядка 30% с T1R1 и T1R2, GPCR, идентифицированными ранее (Hoon et al., 1999), которые структурно составляют еще одно семейство вкусовых рецепторов (T1R1, T1R2 и Т1Ю). Интересно отметить, что методом двойной in situ гибридизации было показано, что T1R1 и T1R2 эксперессируются в разных вкусовых клетках, но некоторые вкусовые клетки экспрессируют комбинации T1R1 и T1R3 или T1R2 и T1R3 (Hoon et al., 1999; Nelson et al, 2001). Ha основании этих данных было сделано предположение, что функциональный рецептор, скорее всего, является гетеродимером (T1R1/T1R3 и T1R2/T1R3), что нашло подтверждение в следующих экспериментах. Оказалось, что при экспрессии комбинации T1R2 + T1R3 в клетках НЕК-293 (которые в контроле не реагировали на вкусовые вещества) таковые отвечали на сладкие вещества (Nelson et al., 2001). При экспрессии комбинации T1R1 + T1R3 индуцировалась чувствительность к аминокислотам (Nelson et al., 2001). Это дает основание полагать, что гетеродимер T1R2/T1R3 является сладко-чувствующим рецептором, а T1R2/T1R3 - функционирует как рецептор аминокислот.
Вкус умами вызывается L-глутаматом в миллимолярных концентрациях (Lindemann, 2001). Во многих тканях глутамат идентифицирован как возбуждающий нейротрансмиттер, где в микромолярных концентрациях он активирует ионотропные и метаботропные (т.е. G-белок сопряжённые) рецепторы (Brown and Dale, 2002). Классические глутаматные рецепторы также найдены во вкусовых клетках (Chaudhari et al., 1996; Lin and Kinnamon, 1999), но они вряд ли обеспечивают рецепцию глутамата как вкусового вещества, поскольку афинность этих рецепторов к глутамату характеризуется микромолярными константами диссоциации. Недавно был идентифицирован подтип метаботропного рецептора (mGlu4t (truncated)), специфически экспрессируемый во вкусовых клетках (Chaudhari et al., 2000). Его особенность в том, что по сравнению с классическим mGlu4-рецептором у него не хватает 130 аминокислот на N-конце (расположен экстраклеточно и несет сайт связывания глутамата), что сдвигает афинность mGlu4t рецептора в область миллимолярных концентраций (Chaudhari et al., 2000). Это обстоятельство, а также апикальная локализация mGlu4t, дают основание считать, что данная изоформа метаботропного рецептора, специфическая для вкусовых клеток, обеспечивает детекцию глутамата в пище (Chaudhari et al., 2000).
Итак, во вкусовых клетках найдены многие элементы внутриклеточных сигнальных систем - рецепторы, G-белки, эффекторные ферменты, ионные каналы. На этом основании до недавнего времени доминировала гипотеза, предполагающая одновременное существование множественных механизмов, использующих cGMP/cAMP и IP3/DAG/Ca в качестве вторичных мессенджеров для трансдукции вкусовыми клетками стимулов разных вкусовых модальностей (Lindemann, 1996; Herness and Gilberson, 1999; Gilberson et al., 2000; Lindemann 2001; Margolskee 2002). Здесь следует отметить сложность изучения молекулярных механизмов вкуса, которая состоит в том, что многочисленные модели вкусовой трансдукции трудно проверить экспериментально, поскольку в электрофизиологических экспериментах не более 5% вкусовых клеток отвечает на адекватные вкусовые стимулы. Однако недавно появившиеся данные ставят под сомнение эту традиционную точку зрения. Согласно поведенческим экспериментам и регистрации от вкусового нерва, у животных с нокаутированными PLC ((32 изоформой) или ионным каналом TRPM5 (семейства TRP-каналов; Runnels et al., 2002, Vazquez et al., 2001, Venkatachalam et al., 2001), практически исчезала чувствительность к сладким, горьким стимулам и аминокислотам при неизменной чувствительности к кислым и солёным стимулам (Zhang et al., 2003). Следовательно, PLCP2 и TRPM5 являются общими и ключевыми элементами сигнальных каскадов, обеспечивающих трансдукцию сладких, горьких стимулов и аминокислот.
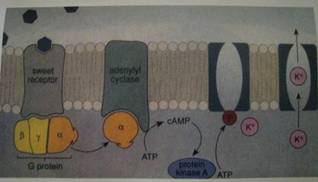
Рис.1 Трансдукция веществ, обладающих сладким вкусом
4. Вкусовые нарушения
вкусовой анализатор трансдукция окраска
4.1 Общие принципы
Анатомическое разнообразие системы вкуса, а также присутствие разнообразных резервных механизмов способствует значительной защите системы от полной и постоянной потери вкусового восприятия. Например, потеря нескольких периферических вкусовых очагов не вызовет изменений в способности ротовой полости в целом воспринимать вкус (Mott, Grushka and Sessle 1993). Система восприятия вкуса в гораздо большей степени восприимчива к вкусовым искажениям или восприятию фантомного вкуса. Например, дисгевзия является заболеванием намного более распространенным в условиях профессиональных воздействий, чем потеря вкусовых ощущений. Хотя способность восприятия вкуса рассматривается в качестве более устойчивой к возрастным изменениям, чем обоняние, специалистами были описаны случаи ослабления вкусовых ощущений с возрастом. При раздражении слизистой рта может иметь место временная потеря вкуса. Теоретически, это может быть вызвано воспалением вкусовых клеток, закупоркой вкусовых пор или функциональными изменениями на поверхности вкусовых клеток. Ожог может повлиять на кро
Вместе с этим смотрят:
Анализ медицинских информационных систем для лечебно-профилактических учреждений санаторного типа
Анатомо-физиологические особенности детей и подростков
Ассортимент лекарственных средств для лечения аллергии
Взаимосвязь нарушения осанки в следствии плоскостопия
Возникновение, лечение и профилактика нозокомиальных инфекций, вызванных грибами рода Candida