Анаэробные сообщества микроорганизмов, разрушающих ароматические ксенобиотики
Страница 2
3.1.Ароматические субстраты и пути их разрушения в анаэробных условиях.
Микроорганизмы-денитрификаторы могут использовать п-крезол, ванилат, катехол, анилин, нитробензол, хлорбензоат, бензиловый спирт, толуол, этилбензол, а также аминокислоту фенилаланин.
Синтрофные метаногенные ассоциации (Syntrophus buswellii,S. gentianae) разлагают бензоат, кротонат, гентизат и гидрохиноны.
Thauera aromatica и Azoarcus evansii используют 2- и 4-аминобензойные кислоты и их тиоэфиры. Помимо этого, могут быть использованы толуол, фенол, фенилпируват, фторзамещённая ароматика, фенилацетальдегид, фенилглиоксилат, алкилбензолы, терефталат и др.( Kleerebezem,1999;Lochmeyer,1992).
4.Синтрофные ассоциации и консорциумы, разлагающие аминоароматические вещества.
Синтрофизмособый случай симбиотической кооперации между метаболически разными типами бактерий, которые зависят друг от друга при разрушении субстратов.
Термин “консорциум” используется для описания различных коопераций микроорганизмов.В примечании к Правилу 31 в “Международном кодексе номенклатуры бактерий”(1978) говорится:”Консорциумэто совокупность или ассоциация двух или более организмов”.Таким образом, в это понятие входят и такие формы сообществ микроорганизмов, как ассоциация и смешанная культура.
Примером консорциума может служить “Methanobacillus omelanskii”(2 организма-партнёра, штамм S и штамм М.о.Н.).:
Штамм S: 2 EtOH+ 2H2O = 2CH3COO--+ 2 H+ + 4 H2 ΔGº=+19 кДж/2 моль EtOH
Штамм M.о.Н.: 4H2 + CO2 =CH4 + 2 H2O ΔGº=-131 кДж/1 моль CH4
Сообщество:2 EtOH+ CO2 =2CH3COO--+ 2 H+ +CH4 ΔGº=-112 кДж/1 моль CH4
В данном случае 2 штамма кооперируются для превращения этанола в ацетат и
метан, при этом происходит межвидовой перенос водорода. Штамм S не может
расти на этаноле в отсутствие штамма M.o.H., потребляющего водород, т.к. реакция
при стандартных условиях эндотермична.Таким образом, ни один из партнёров не
может расти самостоятельно на этаноле, и деградация этилового спирта зависит от
кооперации двух штаммов(Schink,1997).
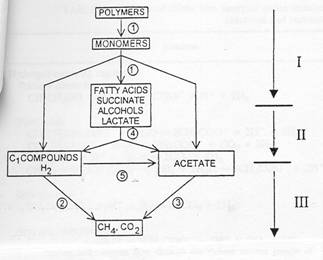
Рис.1. Общая схема потока углерода и электронов в трофических связях микроорганизмов, участвующих в метаногенной деградации органических веществ в анаэробных условиях.
1-первичные бродильщики; 2-водородокисляющие метаногены; 3-ацетатпотребляющие метаногены; 4-вторичные бродильщики; 5-ацетогенные бактерии. (Schink,1997).
Данная схема применима и к анаэробным сообществам микроорганизмов,
разлагающих аминоароматические субстраты.
Разделение метаболических функций и их распределение среди метаболически разных микроорганизмов является компенсацией отсутствия эффективных механизмов деградации в аэробных условиях. Метаболические взаимодействия кооперированных сообществ зависят от переноса метаболитов между партнёрами. Поток Н2 обратно пропорционален расстоянию между водородобразующими бактериями и водородпотребляющими метаногенами. Оптимальная скорость переноса метаболитов достигается в случае тесного контакта партнёров: непосредственного соседства, образования агрегатов или “флокков”. Подобные флокки образуются, например, у бактерий, разлагающих жирные кислоты.Образование таких структур требует времени (иногда даже месяцев) (Schink,1997).
В настоящее время ведутся работы по изучению анаэробных микробных сообществ, разлагающих аминобензойную и аминосалициловую кислоты и их изомеры ( Калюжный, 1998;Савельева и др., 1999; Тян и др.,1999). Эти вещества в окислительных условиях полимеризуются в трудноразлагаемые макромолекулы, что усложняет их разложение, а аэробная деградация приводит к образованию ряда токсических промежуточных продуктов.
Эти исследования проводятся на консорциумах, выделенных из мезофильного и термофильного илов. Получено стабильное метаногенное сообщество,способное потреблять 2-АВА(2-аминобензойная кислота) с высокой скоростью. В качестве промежуточных продуктов образуются бензоат, ацетат и СО2. Предполагается, что эта культура является первичным анаэробным деструктором 2-АВА в выделенном из мезофильного ила стабильном метаногенном сообществе (Савельева и др., 1999).
Из того же мезофильного ила получено стабильное анаэробное сообщество, способное разлагать 4-АВА с образованием метана. Промежуточные продукты деградации те же, что и в случае анаэробного разложения 2-АВА. Из этого консорциума выделена чистая культура первичного деструктора 4-АВА (Савельева и др.,1999). В настоящее время ведутся работы по получению стабильной накопительной культуры и выделению из неё чистой культуры микрорганизмов, разрушающих 5-ASA(5-амносалициловой кислоты) и 3-АВА в термофильных условиях.
Из стабильного консорциума были получены культуры факультативных анаэробов, окисляющих 5-ASA и салициловую кислоты в анаэробных условиях с образованием пропионата, ацетата, СО2 , иногда бутирата и водорода. Считается, что во всех этих сообществах первичным деструктором является 1 и тот же микроорганизм (Савельева и др., 1999; Тян и др., 1999).
Подобные взаимоотношения были описаны в статье N.C.G.Tan (Tan et al.,1999) для анаэробного гранулированного ила, разлагающего азокрасители.В процессе деградации азосоединений происходит восстановление азо-связи с образованием различных ароматических соединений (в т.ч. и с аминозаместителями).Данный консорциум использует другую стратегию деградации субстрата (в 2 этапа). Первый этап,анаэробный,восстановление азо-красителей до ароматических аминов, второй,аэробный,дальнейший распад ароматических соединений. Второй этап осуществляется аэротолерантными микроорганизмами, которые обитают на периферии анаэробного биотопа. Интеграция аэробных и анаэробных условий является благоприятной для биоминерализации азотсодержащих ксенобиотиков(Tan. et al.,1999).
Метаногенные консорциумы участвуют также в деградации такого соединения,как толуол. В состав такого сообщества входят представители царств Бактерий и Архей. Филогенетический анализ доминирующих организмов показал, что в состав данного консорциума входят два вида архей, относящихся к родам Methanosaeta, Methanospirillum,а также один из видов рода Desulfotomaculum.Остальные представители сообщества не были определены, но их вклад в общий метаболизм весьма важен, т.к. при отсутствии какого-либо компонента в сообществе изменяется количественный и качественный состав ( Ficker еt al.,1999).
Помимо этого, существуют сообщества микроорганизмов, состав которых ещё полностью не известен, но есть данные об их способности разлагать ароматические субстраты. На электронных микрофотографиях представителей таких сообществ видно тесное соседство крупных и мелких клеток. Они принадлежат разным организмам, но их расположение свидетельствует об их возможной связи через метаболизм. Обычно эти микроорганизмы трудно выделяются в чистые культуры (Ficker еt al.,1999).
Описана культура микроорганизмов, осуществляющую деградацию о-фталата(Kleerbezem,1999). В культуре присутствовали 2 основных типа микроорганизмов: короткие толстые палочки и маленькие палочки. Толстые ответственны за ферментацию. Маленькие палочки располагались очень близко к толстым и,возможно,являлись метаногенами, потребляющими водород (Methanobacterium). Иногда обнаруживаются споровые палочки, которые, скорее всего, выполняют роль деструкторов (первые стадии деградации). Идентификация до чистых культур не производилась (Kleerbezem et al.,1999).
Щербаковой В.А. было исследовано сообщество,выделенноез полупромышленного UASB-реактора.В разрушении бензолсульфоната и п-толуолсульфоната(ТС) принимают участие 9-10 микроорганизмов, 5 из которых удалось выделить в чистые культуры.Чтобы определить ключевые группы микроорганизмов,участвующие в разрушении ТС до метана, было изучено изменение микробной популяции во время процесса.В первые сутки роста основную часть популяции составлял спорообразующий организм штамм 14 (68.5 %). Во вторые сутки его численность уменьшилась, однако в сообществе преобладали клостридиальные штаммы (80.5%). В дальнейшем наблюдалось увеличение численности штамма SS(спорообразующий организм).С третьих суток наблюдалось активное спорообразование у клостридиальных штаммов, начинался рост численности у метанобразующих микроорганизмов,и к концу первой недели их суммарная численность составляла около 60 % общей популяции.Скачок в потреблении п-толуолсульфоната можно соотнести с ростом численности штамма SS в популяции микробов на третьи сутки процесса. Во время развития сообщества происходило, наряду с образованием метана, накопление летучих жирных кислот. Содержание ацетата в культуральной жидкости было на порядок выше, чем концентрация других. Так, на начальных этапах n- толуолсульфонат используется бактериями рода Clostridium как источник углерода и серы,бактериями рода Desulfovibrio как акцептор электронов.А сразу после появления ацетата толуолсульфонат используется метаносарциной в качестве стимулирующей органической добавки.Толуол является промежуточным продуктом деградации толуолсульфоната.Другим важным интермедиатом процесса является изобутират. Кроме выделенного в чистую культуру Methanobacterium formicicum штамм МН, образующего метан из СО2 и Н2, важную роль в исследуемом сообществе играет другой метаноген рода Methanospirillum.Это наиболее распространённые партнёры в синтрофных организациях, способные расти при низких парциальных давлениях водорода(Щербакова,2000).