Биогеоценоз
Страница 2
Цепи питания. Перенос энергии от ее источника (растений) через ряд организмов называют пищевой цепью. Все живые организмы связаны между собой энергетическими отношениями, поскольку являются объектами питания других организмов. Травоядные животные (потребители первого порядка) поедают растения, первичные хищники (потребители второго порядка) поедают травоядных, вторичные хищники (потребители третьего порядка) поедают хищников помельче. Таким образом, создаются пищевые цепи из продуцентов и консументов, которые на разных этапах, смыкаются с сообществом редуцентов (рис. 3).
Пищевые цепи разделяют на два типа. Один тип пищевой цепи начинается с растений и идет к растительноядным животным и далее к хищникам. Это так называемая цепь выедания (пастбищная). Другой тип начинается от растительных и животных остатков, экскрементов животных и идет к мелким животным и микроорганизмам, которые ими питаются. В результате деятельности микроорганизмов образуется полуразложившаяся масса — детрит. Такую цепь называют цепью разложения (детритной).
На суше пищевые цепи первого типа состоят обычно из 3— 5 звеньев, например: растения -> овца ->- человек — трехзвенная цепь; растения -*- кузнечики -»- ящерицы -»- ястреб — четырехзвенная цепь; растения -»- кузнечики -v лягушки-»- змеи -»--»- орел — пятизвенная цепь. Через пищевые цепи биогеоценозов суши подавляющее количество прироста растительной биомассы поступает через опад в цепи разложения.
В морях распространены такие типы цепей: фитопланктон ->- рыбы -»- хищные птицы; фитопланктон ->- мелкие ракообразные-»-рыбы, питающиеся мелкими рачками и ракообразными -»- хищные рыбы ->- хищные птицы. В водных сообществах большая часть биомассы, накопленной одноклеточными водорослями, проходит через цепь выедания и значительно меньшая включается в цепь разложения (рис. 4).
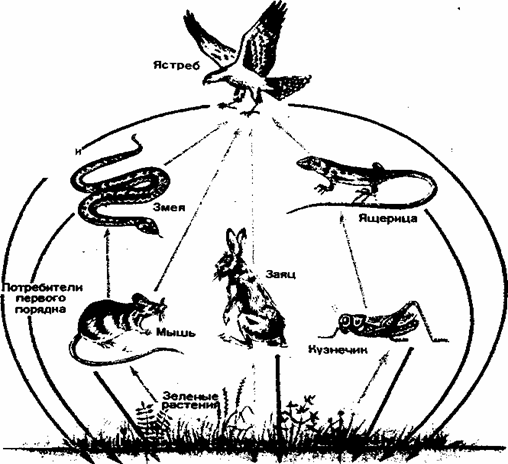
Рис. 3. Пищевые цепи в наземных экосистемах
Все типы пищевых цепей всегда существуют в сообществе таким образом, что член одной цепи является также членом другой. Соединение цепей образует пищевую сеть экосистемы. Угнетение или разрушение любого звена экосистемы с неизбежностью отразится на экосистеме в целом. Поэтому вмешиваться в жизнь экосистем надо с большой осторожностью и осмотрительностью.
Экологическая пирамида. Пищевые сети внутри каждой экосистемы имеют хорошо выраженную структуру. Она характеризуется количеством и размером организмов на каждом уровне цепи питания. При переходе с одного пищевого уровня на другой численность особей уменьшается, а их размер увеличивается. Например, в приведенной выше четырехзвенной цепи на 1 га травяной экосистемы насчитывается около 9 млн. растений (первый пищевой уровень), свыше 700 тыс. растительноядных насекомых (второй уровень), больше 350 тыс. хищных насекомых и пауков (третий уровень) и всего три птицы (четвертый уровень). Как мы видим, образуется пирамида чисел, основание которой в 3 млн. раз шире, чем вершина.
Только часть энергии, поступившей на определенный уровень биоценоза, передается организмам, находящимся на более высоком пищевом уровне. С уровня на уровень переходит около 10% энергии. Можно подсчитать, что энергия, которая доходит до пятого уровня (например, до орла в цепи: растения — кузнечики —»- лягушки - змеи -»- орел), составляет всего 0,01% энергии, поглощенной продуцентами. Таким образом, оказывается, что передача энергии с одного пищевого уровня на другой происходит с очень малым КПД. Это объясняет уменьшение числа и массы

Рис. 4. Пищевые цепи в океане
Способность организмов переносить неблагоприятные условия и высокий потенциал размножения обеспечивают сохранение популяций в экосистеме, что гарантирует ее устойчивость.
Саморегуляция . Поддержание определенной численности популяций основано на взаимодействии организмов в звеньях хищник — жертва, паразит — хозяин на всех уровнях пищевых цепей. Если по каким-либо причинам один из членов пищевых цепей исчезает, то виды, питавшиеся в основном исчезнувшим видом, начинают в большем количестве поедать ту пищу, которая раньше была для них второстепенной. Вследствие подобной замены пищи численность видов-потребителей сохраняется.
Массовое размножение вида в биогеоценозе регулируется прямыми и обратными связями, существующими в пищевых цепях. Нередко благодаря хорошим погодным условиям создается высокий урожай растений, которыми питается определенная популяция травоядных животных. В связи с хорошим питанием численность популяции возрастает. Травоядные сами могут быть пищей для хищников. Чем многочисленнее жертвы, тем более обеспечен едой хищник и тем интенсивнее он размножается. Следовательно, чем больше в нынешнем году жертв, тем больше на следующий год будет хищников. Возрастание количества хищников приводит к снижению численности жертв. Снижение численности жертв ведет к тому, что размножение хищника замедляется и количество хищника и жертвы возвращается к нормальному — исходному соотношению.
Колебания количества растительной пищи, травоядных животных и хищников, питающихся этими животными, сопряжены друг с другом. Классический пример — циклы изменения численности леммингов в тундре. Раз в несколько лет на огромной территории тундры их численность резко возрастает, вслед за тем, часто за один сезон, столь же резко падает. В соответствии с этим численность песцов, лис и сов, питающихся леммингами, либо увеличивается, либо уменьшается.
Колебания численности леммингов связаны с их кормовой базой. В годы повышения численности леммингов они сильно объедают растительность. Большое количество частей растений, содержащих питательные элементы, поступает в детрит. На следующий год из-за значительного повреждения растительного покрова пищи становится меньше и питательная ценность ее уменьшается. В связи с этим рост и выживание молодых леммингов снижается. Год становится малокормным для хищников, и они почти не размножаются.
В течение последующих лет растительные остатки, богатые питательными веществами, минерализуются; питательные элементы поглощаются растениями; количество пищи- леммингов и ее питательная ценность возрастают; численность леммингов вновь стремительно идет вверх; хищники, хорошо кормясь, начинают быстро размножаться. Таким образом, в биогеоценозе популяции организмов взаимно ограничивают свою численность, благодаря чему данная экосистема существует длительное время.
Каково значение саморегуляции численности, мы понимаем особенно хорошо, сталкиваясь с явлениями, когда саморегуляция нарушается. Это обычно происходит в тех случаях, когда человек нарушает сложившуюся структуру сообществ. Примером может служить история с кроликами в Австралии.
Когда человек из Европы стал переселятся на другие континенты, он повез с собой и домашних животных, в том числе кроликов. В 1859 г. на одной из ферм Австралии выпустили 12 пар привезенных зверьков. В биогеоценозах Австралии было слишком мало хищников, чьей пищей могли бы быть кролики. Через 40 лет численность кроликов достигла нескольких сот миллионов особей. Они расселились почти по всему континенту, разоряя луга и пастбища, выедая проростки местной сосны, и нанесли урон экономике страны.
Таким образом, численность особей в природных экосистемах саморегулируется. Нарушение естественных цепей питания под воздействием антропогенного фактора, неразумное вмешательство в экосистемы может привести к неконтролируемому росту численности особей отдельных популяций и к нарушению природных экологических сообществ.
СМЕНА ЭКОСИСТЕМ
Конкретный биогеоценоз не существует вечно. Рано или поздно он сменяется другим. Смены происходят под влиянием изменения среды самими живыми организмами, при смене климатических условий, в процессе эволюции жизни на Земле, под влиянием человека.