Гаметициды и их применение в селекции
Страница 6
На основании исследований конкуренции между пантоте-натом и далапоном появилась возможность предсказать новые аналоги пантоата в виде хлорзамещенных алифатаче-ских кислот. Были синтезированы 4 соединения, биологическая активность которых (в данном случае гербицидная) варьировала в зависимости от степени хлорирования и место
положения хлора: (далапон); при кон
центрации 0,005 М активность далапона составляла 76%, при 0,05 М—99%.
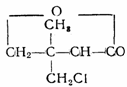
концентрациях составляла соответственно 77 и 100%. 19
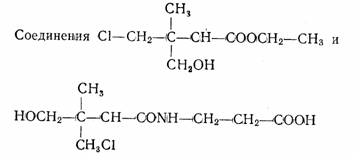
проявляли фитотоксический эффект: при концентрации 0,05 М он был равен 97 и 12%, при 0,05 М — соответственно 100 и 83%.
Таким образом, степень биологической активности препарата не имеет прямо пропорциональной зависимости от концентрации вещества, что свидетельствует о сугубо физиологической активности соединения, связанной с особенностями метаболизма растения.
В исследованиях по биохимизму действия ряда гербицидов установлено существенное влияние их на трансформацию энергия в клетке, повышение интенсивности окислительных процессов и угнетение фосфорелирования в митохондриях [10, 14, 21, 23]. Нарушение сопряженности окисления и фосфорелирования — результат угнетения активности многочисленных ферментов цикла Кребса и дыхательной цепи мито-хондрий. Получены дополнительные сведения о гербицидах, обладающих одновременно и гаметоцидной активностью. В частности, при нанесении далапона на растения люпина .изменялось соотношение сульфгидрильных и дисульфидных групп, входящих в состав активных центров многочисленных энзимов, участвующих в разнообразных ферментативных комплексах [13]. Кроме того, установлено повышенное содержание изофлавоновых глюкозидов и изменение их состава при o6pai6oTKe растений 2,4Д [22].
Появление хинонов — продуктов окислительного превращения фенольных соединений с высокой биологической активностью и их взаимодействие с амино- и сульфгидрильны-ми группами белков, сульфгидрильными группами аскорбиновой кислоты и другими SH-содержащими компонентами клетки обусловливают блокирование целых систем энзимати-чески взаимосвязанных комплексов. От окислительно-восстановительных условий и энергетических возможностей ткани, особенно спорогенной, зависят синтез и обмен важнейших органических соединений.
20
Характерными признаками ЦМС у сорго являются угнетение окислительно-восстановительных процессов и снижение энергетического обмена [41]. Различия в активностях АТФ'азы обнаруживались у стерильных аналогов уже в фазе тетрад и сохранялись в дальнейшем на всех фазах развития микроопор [16]. Среди соединений с гаметоцидными свойствами 2,4Д снижает содержание АТФ и АДФ — адениннук-леотидов, основных аккумуляторов энергии в клетке. Установлено, что 2,4Д ингибирует активность аденилаткиназы — фермента, осуществляющего равновесное соотношение компонентов аденилатного пула: 2 АДФ ^ АМФ+ДТФ [21].
Растительные гормоны, проявившие гаметоцидные свойства (2,4Д, ИУК, НУК, ГКз и т. п.), могут индуцировать мужскую стерильность на тех уровнях метаболических процессов, на которых они оказывают свое регуляторное действие:
на уровне генома, мембран, аллостерического эффекта. Возможно и одновременное влияние их на разные уровни, но во всех случаях отмечена взаимосвязь физиологически активных веществ, к которым относятся гаметоциды, с изменениями в энергетическом обмене клетки.
Существование специфических рецепторов в клеточных структурах и мембранах, способных обратимо связывать ауксины [25], может служить молекулярно-биологической интерпретацией действия ряда соединений, проявивших гаметоцид-ную активность на различных культурах и относящихся к ауксинам (ИУК, НУК, 2,4Д, Г.Кз, кинетин и др.) [11, 12, 14, 17, 58, 135].
Отмечено, что растительные гормоны (2,4Д, ИУК, ГКз), вызывающие при определенных концентрациях различную степень индукции мужской стерильности, влияют на активность энзимов, связанных с метаболизмом углеводов, определяющих структуру клеточных оболочек, с такими как р-1,4-глюканаза, р-1,3-глюканаза, (3-1,6-глюканаза я гемицеллю-лаза, а также а-1,3- и а-1,6-глюканазы [73, 99, 136]. Повышение активности глюканазных энзимов взаимосвязано с процессами деструкции их субстратов, а следовательно, и с изме-нениями в каллозной оболочке материнской клетки пыльцы и формирующихся тетрад, так как она является Р-1,3-свя-занным полимером глюкозы. Установлено, что ИУК и 2,4Д способствуют увеличению р-1,3-глюканаз'ной активности, в результате чего разрываются перекрестные связи в пределах клеточных стенок и оболочек, что обусловливает возрастание их эластичности и проницаемости [55].
Введение ИУК в растительную клетку повышает утилизацию глюкозы путем активации энзима УДФ-зависимой глю-кансинтетазы, локализованной в пределах аппарата Гольд-21
жи, что способствует формированию и повышенному содержанию глюканов, галактанов и пентозанов [42]. Подобным образом 2,4Д включается в один из уровней метаболизма клетки (через углеводы, путем активации плазменной, связанной с мембранами глюкансинтетазы), что способствует утилизации УДФ-арабинозы я увеличению количества связанных остатков арабинозы с галактаном [136]. Повышение числа сшивок в молекулах галактана изменяет пластичность клеточных стенок. Вместе с тем аккумуляция 2,4Д в мембранах вызывает нарушение комплекса связанного с мембранами белкового фактора, который обусловливает активность PHiK полимеразы, транскрибирующей определенные мРНК [66].
В опытах по конкурентному вытеснению связанных эффек-торов (производные феноксиуксусной кислоты и ИУК) наглядно продемонстрировано, что связывание биологически активных хлорированных производных мембранами (эффектор-рецептор) носит специфический характер [14]. Изменяя функциональную активность мембран и связанных с ними энзи-мов, ауксины с гаметоцидными свойствами могут вызывать индукцию синтеза определенных мРНК, ответственных за •продуцирование ряда энзимов, среди которых имеются ферменты, преобразующие углеводные компоненты мембран и клеточных оболочек. Возрастающая при этом проницаемость может вызывать нарушение селективной изоляция формирующихся тетрад с последующим их деградированием. Предполагают, что каллозная оболочка функционирует как «молекулярный фильтр», позволяющий проникать внутрь материнских клеток пыльцы основным питательным элементам, за исключением больших молекул. Последние в эту раннюю фазу могут помешать установлению автономии ядра гаплоид-ной споры в пределах собственной цитоплазмы [98].
Химическая изоляция материнских клеток пыльцы в стадии тетрад от окружающей диплоидной цитоплазмы является необходимой предпосылкой нормального развития пыльцы [94]. Установлено, что меченый тимидин поступает в материнские клетки пыльцы только до формирования каллозной оболочки, но не проникает, если они заключены в каллозу [72]. При изменении последней и освобождении тетрад метка свободно поступала в микроспоры. Эти наблюдения позволили сделать вывод о функционировании каллозной оболочки как «молекулярного фильтра».
Каллозное покрытие материнских клеток пыльцы функционировало как молекулярное сито: каллоза пропускала глюкозу и углекислый натрий, но задерживала фенилаланин, размер молекулы которого гораздо меньше глюкозы и он 22
должен был бы легко проникать в материнские клетки пыльцы [123]. Выводы J. Heslop-iHarrison и A. Mckenzie [72] также сомнительны, так как метка могла не включаться ввиду отсутствия синтеза ДНК. В других исследованиях показано, что роль каллозной оболочки значительно сложнее, чем «простого молекулярного фильтра» [4, 130].