Эволюция биологических механизмов запасания энергии
Страница 4
Перенос по цепи электрона, отнятого от возбужденного хлорофилла, завершается по-разному в зависимости от типа фотосинтеза. У зеленых бактерий, использующих комплекс хлорофилла и белка, называемый фотосистемой 1 (рис. 5, а), продуктом оказывается НАДН, то есть восстановленная форма НАД+. Восстанавливаясь, то есть присоединяя два электрона, НАД+ связывает также один Н+. В дальнейшем образованный таким образом НАДН окисляется, передавая свой водород на различные субстраты биосинтезов.
Что касается хлорофилла, окисленного цепью, то у зеленых серных бактерий он получит недостающий электрон от сероводорода (H,S). В результате образуются также элементарная сера и ион Н+. Белок, окисляющий H2S, расположен на внешней поверхности бактериальной мембраны, а белок, восстанавливающий НАД+, — на внутренней ее поверхности. Вот почему оказывается, что запускаемый светом перенос электронов от H,S к НАД"1" образует ионы Н+ снаружи и потребляет их внутри бактерии. При этом внутренний объем клетки заряжается отрицательно относительно внешнего. Тем самым создается ∆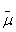 Н+, которая потребляется Н+ - АТФ-синтазой (комплексом факторов F0 и F,), образующей АТФ при переносе ионов Н "под гору", то есть снаружи внутрь.
Н+, которая потребляется Н+ - АТФ-синтазой (комплексом факторов F0 и F,), образующей АТФ при переносе ионов Н "под гору", то есть снаружи внутрь.
Другой тип бактериального фотосинтеза обнаружен у пурпурных бактерий (рис. 5. б). Здесь действует набор ферментов, отличающихся от ферментного комплекса зеленых бактерий. Это несущая хлорофилл фотосистема 2 и комплекс III. Как и в предыдущем случае, процесс начинается с поглощения кванта хлорофиллом. Первоначатьно перенос электронов происходит по фотосистеме 2.
Затем вступает комплекс III, способный транспортировать электроны сопряженно с откачкой ионов Н+ из бактерии. Процесс завершается возвращением электрона с комплекса III на хлорофилл. Что касается ионов Н+, то они возвращаются в клетку через Н+-АТФ-синтазу, образуя АТФ.
Отличительная черта фотосинтеза у пурпурных бактерий состоит в том, что система не нуждается во внешнем доноре электронов. Откачка ионов Н+ осуществляется путем циклического переноса электронов, поддерживаемого энергией света. Данное обстоятельство можно отнести, по-видимому, на счет эволюционного усовершенствования фотосинтеза пурпурными бактериями, которые по многим признакам являются эволюционно более продвинутой группой, чем зеленые серные бактерии.
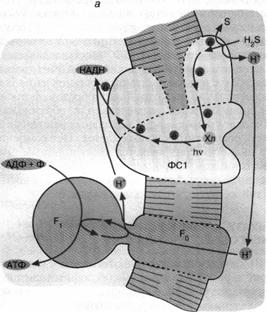
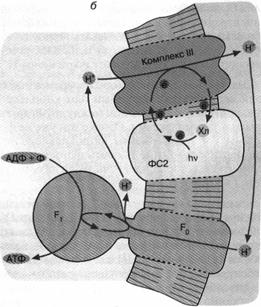
Рис. 5. Хлорофилльный фотосинтез зеленых серных (а) и пурпурных (б) бактерий: а - хлорофилл, связанный с особым белковым комплексом - фотосистемой 1 (ФС1), возбуждается квантом света и отдает электрон по цепи электронных переносчиков на НАД+. Восстанавливаясь, НАД* связывает внутриклеточный ион НГ. Потеря электрона на хлорофилле компенсируется окислением сероводорода до серы и иона ИГ снаружи бактериальной клетки. Движение Н+ внутрь клетки через комплекс F0 F1 дает АТФ; б - хлорофилл, связанный с белком фотосистемы 2 (ФС2), поглощает квант света и запускает циклический перенос электронов. В этом процессе участвуют переносчики электронов ФС2 и дополнительного белкового комплекса III. Перенос электронов комплексом III сопряжен с откачкой ионов Н+ из клетки. Откачанные ионы I-Г возвращаются через комплекс F0 F, с образованием АТФ
Следующим шагом в эволюции фотосинтеза стали, по-видимому, цианобактерии. Цепь переноса электронов в этом случае представляет собой комбинацию: а) фотосистемы 1 зеленых бактерий, б) фотосистемы 2 и комплекса III пурпурных бактерий и в) дополнительного комплекса, расщепляющего воду на О2 и Н+ (рис. 6). Фактически донором электронов вместо сероводорода (встречающегося в достаточных количествах лишь в некоторых приданных нишах) служит вездесущая вода, запасы которой практически неограниченны. В результате конечный акцептор электоров — НАДФ+ восстанавливается, а вода окисляется. Образующийся НАДФН окисляется затем сложной системой восстановления углекислого газа до глюкозы. Таким образом, фотосинтез цианобактерии параллельно с образованием АТФ дает углевод — одно из главных резервных веществ современных живых клеток. Нет сомнений, что цианобактерия является эволюционным предшественником хлоропластов — органелл зеленых растений, энергетика которых устроена в основном по той же схеме, что показана на рис. 6.
ДЫХАТЕЛЬНЫЙ МЕХАНИЗМ ЭНЕРГООБЕСПЕЧЕНИЯ
Побочным продуктом фотосинтеза у цианобактерий и растений служит молекулярный кислород. Нарастание его концентрации в атмосфере привело к появлению ферментов, убирающих этот сильный окислитель, опасный для жизнедеятельности. Вероятно, первой функцией ферментов, восстанавливающих О2 до Н2О, было снижение внутриклеточной концентрации кислорода. Однако в дальнейшем аэробная клетка научилась извлекать пользу из этого процесса, создав дыхательную цепь электронного транспорта, сопряженного с откачкой ионов Н+.
Дыхательная цепь некоторых современных бактерий включает уже знакомый нам комплекс III, служащий связующим звеном между двумя другими белковыми комплексами. Однако это уже не фотосистемы 1 и 2, а ферменты, выполняющие функции, противоположные таковым фотосистем 1 и 2. Ферменты, о которых идет речь, были названы комплекс I и комплекс IV.
Комплекс I не восстанавливает никотинамидный нуклеотид, а окисляет его. Комплекс IV не окисляет воду до О2, а восстанавливает О2 до воды. В итоге мы имеем сложную цепь реакций, начинающихся с окисления НАДН и кончающихся восстановлением О2. Все три комплекса дыхательной цепи способны откачивать из клетки ионы Н+ сопряжено с переносом электронов (рис. 7).
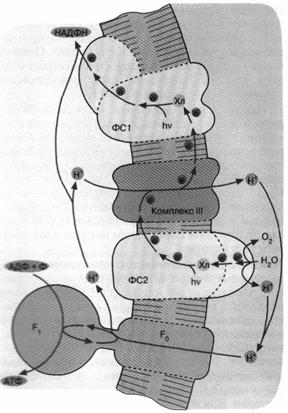
Рис. 6. Хлорофилльный фотосинтез цианобактерии. Квант света, поглощенный хлорофиллом фотосистемы 1, возбуждает перенос электронов по цепи, что завершается восстановлением НАДФ+ до НАДФН. Окисленный хлорофилл фотосистемы 1 восстанавливается комплексом III, который, в свою очередь, получает электрон от фотосистемы 2. Донирование электрона фотосистемой 2 требует еще одного кванта света (поглощаемого хлорофиллом этой фотосистемы). Потеря электрона на хлорофилле фотосистемы 2 компенсируется за счет окисления молекулы воды до О2 и Н+. Ферменты, катализирующие всю цепь реакций переноса электрона от Н2О до НАДФ*, расположены в мембране таким образом, что ионы Н+ откачиваются из бактериальной клетки, чтобы затем войти внутрь через FCF. и сделать АТФ. В хлоро-пластах зеленых растений происходят те же события, но ориентация всех ферментов противоположна той, которая имеет место у цианобактерии и показана на рис. 6. Соответственно у хлоропластов фотосинтетическая цепь накачивает ионы Н+ внутрь, а комплекс F0F, переносит их наружу
Подобно тому как хлоропласты произошли от цианобактерии, митохондрии животных, растений и грибов ведут свое происхождение от аэробных бактерий. Поэтому неудивительно, что митохондриальная дыхательная цепь описывается той же схемой, что изображена на рис. 7.
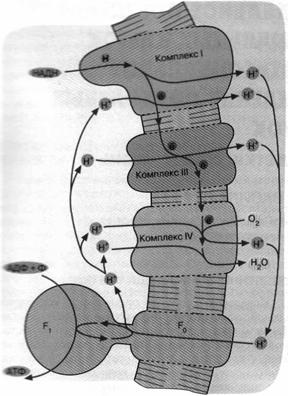
Рис.7. Механизм дыхательного фосфорилирования в аэробных бактериях и митохондриях. Ферментные комплексы I, III и IV катализируют перенос электронов от НАДН к О2 с образованием воды. Перенос электронов сопряжен с откачкой ионов Н*. Возвращение ионов Н+ через F0F1 приводит к синтезу АТФ
ФОТОСИНТЕЗ И БИОСФЕРА
Основным и практически неиссякаемым источником энергии на поверхности Земли является энергия солнечного излучения, постоянным потоком поступающая из космоса благодаря протеканию термоядерных реакций на ближайшем к нам светиле — Солнце. Как показано на рис. 1, спектр поступающего на Землю солнечного излучения соответствует спектру излучения абсолютно черного тела, нагретого до 5900 К. Полный поток солнечного излучения (измеренный за пределами земной атмосферы), приходящийся на единицу поверхности, нормальной к направлению на Солнце, близок к 1400 Вт/м2. Значительная часть этой энергии приходится на область видимого и ближнего инфракрасного излучения (0,3 - 1,0 мкм) — фотосинтетически активную радиацию, эффективно поглощаемую пигментами, участвующими в фотосинтезе растений и фотосинтезирующих бактерий.