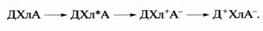Эволюция биологических механизмов запасания энергии
Страница 6
ВЫДЕЛЕНИЕ МОЛЕКУЛЯРНОГО КИСЛОРОДА
Приобретенная в процессе эволюции (более 2 млрд. лет назад) способность фотосинтезирующих растений к выделению молекулярного кислорода в результате окисления воды (см. рис. 2) привела к поистине революционным преобразованиям на Земле, из которых необходимо отметить следующие.
1. Фотосинтезирующие организмы, а через их посредство и вся живая природа, получили доступ к практически неиссякаемому и возобновляемому источнику электронов, участвующих во всех биоэнергетических процессах, — воде, что, естественно, привело к резкому возрастанию масштабов фотосинтеза и поступления энергии в биосферу. Ранее фотоокислению могли подвергаться лишь восстановленные вещества типа H2S, представленные на Земле в довольно ограниченном количестве.
2. Продукт фотосинтетического окисления воды — молекулярный кислород, содержание которого в атмосфере возросло в результате фотосинтеза практически от нуля в древней атмосфере до 21% в настоящее время, вызвал значительные изменения во всей живой природе. Появление в атмосфере свободного О2 привело к массовой гибели существовавших тогда организмов, для которых такой сильный окислитель как кислород оказался сильнейшим ядом, поскольку он резко нарушал протекание биоэнергетических процессов. В то же время в результате длительной эволюции на фоне возрастающей концентрации О2 в атмосфере появились новые, аэробные организмы, обратившие наличие О2 себе на пользу. Они сумели включить его в свой метаболизм в качестве эффективного конечного акцептора электронов в дыхательной цепи. Это позволило поднять биоэнергетику на новый, значительно более высокий уровень, так как в этом случае происходит более эффективное "сжигание" органики. Так, например, выход макроэргического эквивалента — аденозинтрифосфата (АТФ) на одну молекулу исходной гексозы составляет две молекулы при молочнокислом брожении и 36 — 38 молекул (то есть в 18—19 раз выше) при аэробном дыхании.
Фотосинтетическое образование О2, сопровождающееся поглощением СО2, в сочетании с появившейся возможностью аэробного дыхания, при котором происходит обратный процесс — потребление 02, сопровождающееся освобождением СО2, привело к образованию замкнутых циклов О2 и СО2, имеющих глобальные масштабы. Сочетание этих циклов позволяет восполнять содержание как СО2, так и О2, а их участие в едином цикле "фотосинтез-дыхание" обеспечивает постоянное поступление энергии в биосферу (см. рис. 2).
3. Появление кислородвыделяющих организмов привело к тому, что практически все процессы на поверхности Земли приняли биогеохимический характер. Именно в этот период значительного повышения О2 в атмосфере произошло окисление соединений железа, серы, марганца, причем на это ушло более 95% кислорода, выделенного фотосинтезирующими организмами за всю историю биосферы. Атмосфера Земли до появления кислородвыделяющих фотосинтезирующих организмов была разреженной и состояла главным образом из СО2 и аммиака. По мере возрастания О2 в атмосфере значительно увеличивается и содержание молекулярного азота благодаря окислению аммиака. Сам процесс фотосинтеза, сопровождающийся образованием и запасанием органического вещества, привел к значительному, более чем в 100 раз, уменьшению содержания СО2 в атмосфере.
В результате первичная разреженная атмосфера превратилась во вторичную плотную азотокислородную оболочку Земли. Теперь солнечная и другие виды космической радиации, прежде чем достичь поверхности, должны были фильтроваться через мощный слой атмосферы. Кроме того, под действием фотохимического процесса в верхних слоях атмосферы двухатомный молекулярный кислород превращается в трехатомную молекулу озона, имеющего интенсивное поглощение в области жесткого ультрафиолета (0,2 — 0,3 мкм). В результате образовавшийся озоновый экран стал задерживать наиболее опасную для живых организмов ультрафиолетовую радиацию.
Таким образом, фотосинтез, основанный на окислении воды, благодаря возможности использовать воду в качестве субстрата стал мощным поставщиком энергии в биосферу, революционным образом обогатил биоэнергетику благодаря включению О2 в метаболизм живых организмов, защитил живую материю от гибельного действия космической радиации. Все это индуцировало мощное развитие биологической материи на Земле, приведшее в конечном счете к появлению человека. И в настоящее время фотосинтез лежит в основе биологического круговорота энергии и веществ на Земле, от масштабов которого зависит и жизнь на нашей планете, и ее разнообразие и возможности.
МОЛЕКУЛЯРНЫЕ ОСНОВЫ ПРЕОБРАЗОВАНИЯ И ЗАПАСАНИЯ ЭНЕРГИИ СВЕТА ПРИ ФОТОСИНТЕЗЕ
Живая природа настолько рационально решает проблемы эффективного улавливания солнечной энергии, ее преобразования и запасания в удобной для биологического потребления форме, что до сих пор не удается воспроизвести основные этапы фотосинтеза в искусственных системах. Первичное улавливание и преобразование энергии света при фотосинтезе происходит с непременным участием хлорофилла, что было выявлено уже в классических работах К.А. Тимирязева, показавшего, что именно та часть солнечного света, которая поглощается хлорофиллом, эффективно инициирует процесс фотосинтеза.
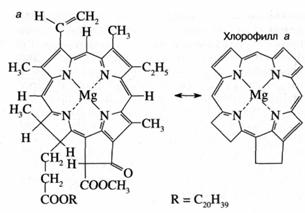
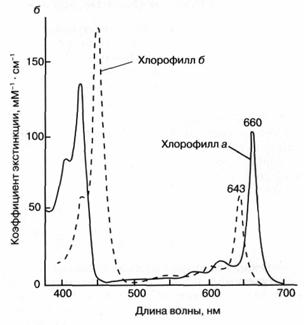
Как показано на рис. 3а, молекула хлорофилла состоит из порфиринового ядра, образованного тетрапиррольным кольцом с расположенным в центре атомом магния, и фитольного хвоста. Хлорофилл относится к классу веществ, называемых пигментами, то есть сильноокрашенных соединений, характеризующихся интенсивным поглощением видимого света. Было установлено, что система "легко возбудимых" сопряженных связей порфиринового кольца обусловливает высокую способность хлорофилла к поглощению света (рис. 36). Так, миллиметровый слой 1 мМ раствора хлорофилла поглощает около 90% падающего на него красного или синего света (зеленый свет поглощается менее эффективно, что и определяет зеленый цвет растений). Для хлорофилла, как и для других пигментов, характерно относительно длительное время, около 1(Г8 сек, нахождения в возбужденном состоянии, что важно для участия хлорофилла в фотохимических реакциях. У многих других пигментов, например, у производных хлорофилла, у которых центральный атом магния заменен на медь или марганец, это время гораздо ниже вследствие чрезвычайно быстрого перехода энергии электронного возбуждения в тепло.
Для выяснения молекулярных основ участия хлорофилла в первичных процессах преобразования солнечной энергии при фотосинтезе важнейшее значение имело открытие, сделанное нашим соотечественником, академиком А.А. Красновским, который показал, что при переходе в возбужденное состояние в результате поглощения кванта света молекула хлорофилла приобретает свойство вступать в такие окислительно-восстановительные (то есть идущие с переносом электронов) реакции, к которым она не была способна в темноте. При этом молекула хлорофилла (Хл) может потерять свой электрон, то есть окислиться, отдав его другому веществу, называемому в данном случае акцептором (А) электрона, тогда как хлорофилл выступает в данной реакции как донор электрона. Эта реакция получила название реакции Красновского. Возбужденная молекула хлорофилла (Хл*) может также восстанавливаться, то есть принять "лишний" электрон, отобрав его у другого вещества, служащего в данном случае донором (Д) электрона. Если помимо Хл в системе присутствуют Д и А, то в результате сочетания процессов окисления Хл и его восстановления свет, поглощаемый хлорофиллом, будет индуцировать перенос электрона от Д к А с образованием Д+ (окисленного Д) и А- (восстановленного А). Хлорофилл при этом, несмотря на участие в двух ступенях переноса электрона, возвращается в исходное состояние готовности к новому фотохимическому циклу переноса электрона, согласно реакции