Физиология сенсорных систем
Страница 5
На поперечном разрезе улитковая часть перепончатого лабиринта имеет форму вытянутого треугольника. Его нижняя сторона, граничащая с лестницей барабана, образована основной пластинкой, которая состоит из погруженных в гомогенную массу тончайших эластических соединительнотканных волокон, натянутых между свободным краем спиральной костной пластинки и наружной стенкой канала улитки. Верхняя сторона треугольника граничит с лестницей преддверия, отходя под острым углом от верхней поверхности
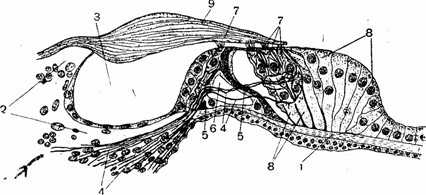
Рис. 4. Схема строения кортиева органа:
/ — основная пластинка; 2 — костная спиральная пластинка; 3 — спиральный канал;
4 — нервные волокна; S — столбовые клетки, образующие тоннель (6); 7 — слуховые, или волосковые, клетки; 8 — опорные клетки; 9 — покровная пластинка.
спиральной костной пластинки и направляясь, как и основная пластинка, к наружной стенке канала улитки. Третья, самая короткая сторона треугольника состоит из соединительной ткани, плотно сращенной с наружной стенкой костного канала. Функция кортиева органа. Рецепторный аппарат слухового анализатора, или спиральный кортиев орган, расположен внутри улитковой части перепончатого лабиринта на верхней поверхности основной пластинки (рис. 4). Вдоль внутренней части основной пластинки, на некотором расстоянии друг от друга, расположены два ряда столбовых клеток, которые, соприкасаясь своими верхними концами, отграничивают свободное треугольное пространство, или тоннель. По обе стороны от него находятся чувствительные к звуковым колебаниям см/ховые, или волосковые, клетки, каждая из которых на своей верхней свободной поверхности имеет 15—20 небольших тончайших волосков. Концы вслосков погружены в покровную пластинку, она укреплена на костной-спиральной пластинке и свободным концом покрывает кортиев орган. Волосковые клетки расположены кнутри от тоннеля в один ряд, а кнаружи—в три ряда. От основной пластинки они отделены опорными клетками.
К основаниям волосковых клеток подходят конечные разветвления волокон биполярных нервных клеток, тела которых расположены в центральном канале костного стержня улитки, где они образуют так называемый спиральный узел, гомологичный межпозвоночному узлу спинномозговых нервов. Каждая из трех с половиной тысяч внутренних волосковых клеток связана с одной, а иногда и с двумя отдельными нервными клетками. Наружные волосковые клетки, количество которых достигает 15—20 тысяч, могут быть соединены и с несколькими нервными клетками, но при этом каждое нервное волокно дает ответвления только к волоско-вым клеткам одного и того же ряда.
Перилимфа, окружающая перепончатый аппарат улитки, испытывает давление, которое и меняется соответственно частоте, силе и форме звуковых колебаний. Изменения давления вызывают колебания основной пластинки вместе с расположенными на ней клетками, волоски которых испытывают при этом изменения давления со стороны покровной пластинки. Это, по-видимому, и ведет к возникновению возбуждения в волосковых клетках, которое передается на конечные разветвления нервных волокон.
Резонансная теория слуха.
Среди различных теорий, объясняющих механизм периферического анализа звуков, наиболее обоснованной следует считать резонансную теорию, предложенную Гельмгольцем в 1863 году. Если около открытого рояля воспроизвести музыкальным инструментом или голосом звук определенной высоты, то начнет резонировать, т. е. звучать в ответ, струна, настроенная на тот же самый тон. Изучая структурные особенности основной пластинки улитки, Гельмгольц пришел к выводу, что звуковые волны, приходящие из окружающей среды, вызывают колебания поперечных волокон пластинки по принципу резонанса.
Всего насчитывают в основной пластинке около 24 000 поперечных эластических волокон. Они различны по длине и степени натяжения: самые короткие и сильнее натянутые расположены у основания улитки; чем ближе к ее вершине, тем они длиннее и слабее натянуты. Согласно резонансной теории, различные участки основной пластинки реагируют колебанием своих волокон на звуки разной высоты. Такое представление подтвердилось опытами Л. А. Андреева. После выработки у собак условных рефлексов на чистые тоны различной высоты улитку одного уха он полностью удалял, а улитку другого подвергал частичному повреждению. В зависимости от того, какой участок кортиева органа второго уха был поврежден, наблюдалось исчезновение ранее выработанных положительных и отрицательных условных рефлексов на звуки определенной частоты колебаний. 'При разрушении кортиева органа ближе к основанию улитки исчезали условные рефлексы на высокие тоны. Чем ближе к верхушке локализовалось повреждение, тем ниже были тоны, утратившие значение условных раздражителей.
Проводящие пути слухового анализатора.
Первый нейрон проводящих путей слухового анализатора — упомянутые выше биполярные клетки. Их аксоны образуют улитковый нерв,, волокна которого входят в продолговатый мозг и оканчиваются в ядрах, где расположены клетки второго нейрона проводящих путей. Аксоны клеток второго нейрона доходят до внутреннего коленчатого тела,
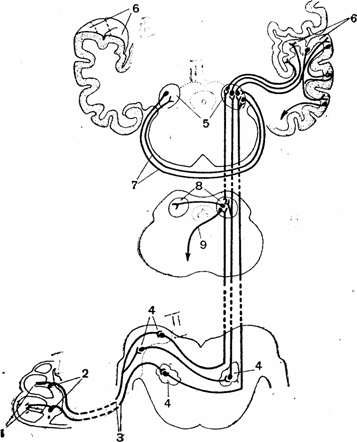
Рис. 5. Схема проводящих путей слухового анализатора:
1 — рецепторы кортиева органа; 2 — тела биполярных нейронов; 3 — улитковый нерв; 4 — ядра продолговатого мозга, где ' расположены тела второго нейрона проводящих путей; 5 — внутреннее коленчатое тело, где начинается третий нейрон основных проводящих путей; 6 •— верхняя поверхность височной доли коры больших полушарий (нижняя стенка поперечной щели), где оканчивается третий нейрон; 7 — нервные волокна, связывающие оба внутренних коленчатых тела; 8 — задние бугры четверохолмия; 9 — начало эфферентных путей, идущих от четверохолмия.
главным образом противоположной стороны. Здесь начинается третий нейрон, по которому импульсы достигают слуховой области коры больших полушарий (рис. 5).
Помимо основного, проводящего пути, связывающего периферический отдел слухового анализатора с его центральным, корковым отделом, существуют и другие пути, через которые могут осуществляться рефлекторные реакции на раздражение органа слуха у животного и после удаления больших полушарий. Особое значение имеют ориентировочные реакции на звук. Они осуществляются при участии четверохолмия, к задним и отчасти передним буграм которого идут коллатерали волокон, направляющихся к внутреннему коленчатому телу.
Корковый отдел слухового анализатора.
У человека ядро коркового отдела слухового анализатора расположено ^в височной, области коры больших,, полушарий. В той части поверхности височной' области, которая представляет собой нижнюю стенку поперечной, или сильвиевой, щели, расположено поле 41. К нему, а возможно и к соседнему полк» 42, направляется основная масса волокон от внутреннего коленчатого тела. Наблюдения показали, что при двустороннем разрушении указанных полей наступает полная глухота. Однако в тех случаях, когда поражение ограничивается одним полушарием, может наступить небольшое и нередко лишь временное понижение слуха. Это объясняется тем, что проводящие пути слухового анализатора неполностью перекрещиваются. К тому же оба внутренних коленчатых тела связаны между собой промежуточными нейронами, через которые импульсы могут переходить с правой стороны на левую и обратно. В результате корковые клетки каждого полушария получают импульсы с обоих кортиевых органов.
От коркового отдела слухового анализатора идут эфферентные пути к нижележащим отделам мозга, и прежде всего к внутреннему коленчатому телу и к задним буграм четверохолмия. Через них осуществляются корковые двигательные рефлексы на звуковые раздражители. Путем раздражения слуховой области коры можно вызвать у животного ориентировочную реакцию настораживания (движения ушной раковины, поворот головы и т. п.). Анализ и синтез звуковых раздражении. Анализ звуковых раздражений начинается в периферическом отделе слухового анализатора, что обеспечивается особенностями строения улитки, и прежде всего основной пластинки, каждый участок которой колеблется в ответ на звуки только определенной высоты.