Рекомбинантные вакцины (Генная инженерия)
Страница 5
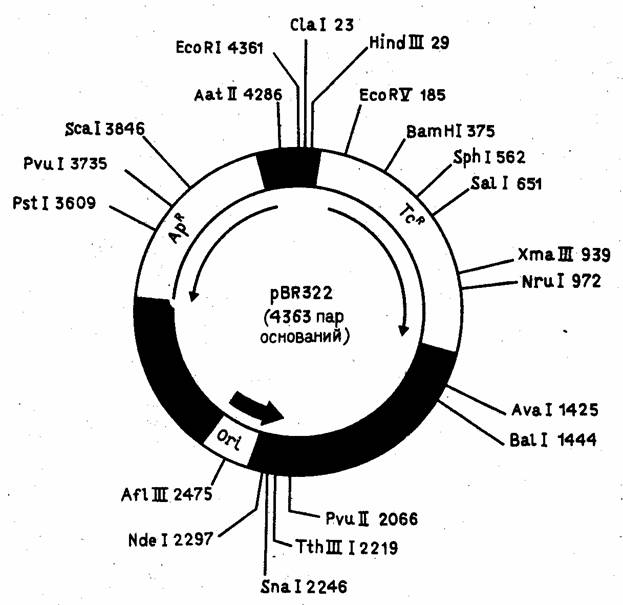
Рисунок 4. Детальная рестрикционная карта плазмиды pBR322.
вый геном, который на 10% больше или на 75% меньше генома дикого l фага, уже не может быть упакован в фаговые частицы.
Таких ограничений теоретически не существует для векторов, сконструированных на базе нитчатого бактериофага М13. Описаны случаи, когда в геном этого фага была встроена чужеродная ДНК длиной около 40 тыс. нуклеотидных остатков. Известно, однако, что фаг М13 становится нестабильным, когда длина чужеродной ДНК превышает 5 тыс. нуклеотидных остатков. Фактически же векторы, полученные из ДНК фага М13, используются главным образом для секвенирования и мутагенеза генов, и размеры встраиваемых в них фрагментов намного меньше.
Эти векторы конструируются из реплекативной (двутяжевой) формы ДНК фага М13, в которую встроены "полилинкерные" участки (пример такой конструкции показан на рис. 5). В фаговую частицу ДНК включается в виде однотяжевой молекулы. Таким образом, этот вектор позволяет получать клонированный ген или его фрагмент как в двутяжевой, так и в однотяжевой форме. Однотяжевые формы рекомбинантных ДНК широко используются в настоящее время при определении нуклеотидной последовательности ДНК методом Сэнгера и для олигодезоксинуклеотид-направленного мутагенеза генов.
Перенос чужеродных генов в клетки животных осуществляется с помощью векторов, полученных из ДНК ряда хорошо изученных вирусов животных - SV40, некоторых аденовирусов, вируса папиломы быка, вируса оспы и так далее. Конструирование этих векторов проводится по стандартной схеме: удаление "лишних" сайтов для рестриктаз, введение маркерных генов в области ДНК, не существенные для ее репликации (например, гена тимидин-киназы (tk) из HSV (вируса герпеса)), введение регуляторных районов, повышающих уровень экспрессии генов.
Удобными оказались так называемые "челночные векторы", способные реплицироваться как в клетках животных, так и в клетках бактерий. Их получают, сшивая друг с другом большие сегменты векторов животных и бактерий (например, SV40 и pBR322) так, чтобы районы, ответственные за репликацию ДНК, остались незатронутыми. Это позволяет проводить основные операции по конструированию вектора в бактериальной клетке (что технически намного проще), а затем полученную рекомбинантную ДНК использовать для клонирования генов в животной клетке.
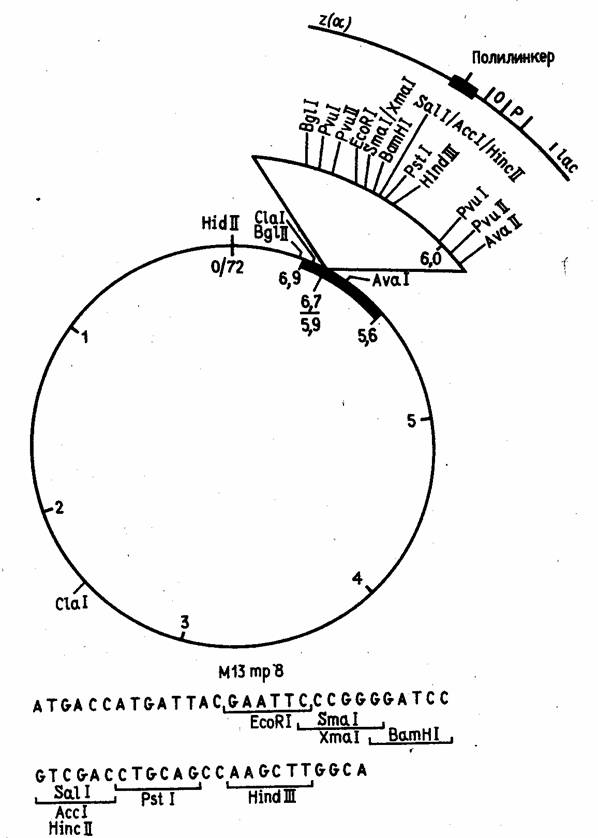
Рисунок 5. Рестрикционная карта вектора М13 mp8.
1.3. ВЫБОР СИСТЕМЫ ЭКСПРЕССИИ КЛОНИРОВАННОГО ГЕНА, СПОСОБНОЙ ОБЕСПЕЧИТЬ МАКСИМАЛЬНЫЙ ВЫХОД И ФУНКЦИОНАЛЬНУЮ ПОЛНОЦЕННОСТЬ ПРОДУКТА
Полученные рекомбинантные молекулы переносятся в определенные группы клеток, которые должны обеспечить экспрессию этих генов, то есть синтез соответствующих белков в количествах, экономически рентабельных по сравнению с обычной технологией их производства.
Обычно для данной цели используют бактериальные или дрожжевые культуры клеток, а также системы экспрессии на основе эукариотических клеток.
Из бактериальных клеток наиболее изученной в молекулярно-генетическом отношении является грамотрицательная бактерия Escherichia coli, поэтому для нее можно с наибольшей определенностью планировать генноинженерные конструкции. Однако E. coli слабо освоена промышленностью. Кроме того, она относится к условно-патогенным для человека микроорганизмам, что может создать трудности при получении на ее основе фармацевтических препаратов.
Отмеченные недостатки E. coli легко преодолеваются при конструировании методами генной инженерии штаммов-продуцентов на основе клеток Bacillus subtilis. Данная почвенная бактерия безопасна для человека и животных и прекрасно освоена микробиологической промышленностью. Бактерия B. subtilis по степени изученности следует за E. coli. Важное отличие ее от E. coli - способность эффективно секретировать во внешнюю среду целый ряд белков, поэтому особенно интересны работы по созданию штаммов-продуцентов B. subtilis, секретирующих чужеродные белки из клеток. Однако данная бактерия имеет свои недостатки: рекомбинантные плазмиды в B. subtilis характеризуются нестабильностью, выражающейся в перестройках и делециях ДНК; бациллы секретируют в культуральную среду большое количество протеаз, что существенно усложняет вопрос максимализации генноинженерного получения целевого белка на основе бацилл-продуцентов.
Среди эукариотических микроорганизмов наиболее изученным является низший эукариот Saccharomyces cerevisiae. Одно из преимуществ S. cerevisiae как экспериментальной системы - простота и надежность ее генетического анализа. В клетках дрожжей имеется ферментативная система гликозилирования белков, которая обеспечивает возможность синтеза в них полноценных белков высших эукариот. Аналогичных систем процессинга белков в бактериальных клетках нет. Многие штаммы дрожжей освоены микробиологической промышленностью. Доказана их безвредность для человека и животных. Именно на S. cerevisiae создан первый штамм-продуцент поверхностного антигена вируса гепатита Б, позволивший получить и испытать вакцину против данного вирусного заболевания человека.
С появлением генной инженерии внимание многих исследователей привлекла система культивируемых клеток животных. Особый интерес к культурам клеток животных стал проявляться после обнаружения того, что часть эукариотических генов раздроблена и лишь в системе клеток высших эукариот можно достичь правильной экспрессии таких генов. Кроме того, многие белки животных и их вирусов синтезируются первоначально в виде более высокомолекулярных предшественников, которые в результате специфического протеолитического процессинга переходят в так называемую зрелую форму. Такой процессинг данных белков, по-видимому, можно ожидать лишь в системе клеток животных. Все это убедительно доказывает важность разработки экспрессирующей системы на основе клеток животных.
Для обеспечения наиболее эффективной экспрессии клонированных генов в векторные молекулы встраивают определенные фрагменты ДНК, позволяющие увеличить выход чужеродного белка. Так, для достижения более высокого уровня экспрессии гена HBsAg в клетках E. coli были использованы различные по силе промоторы (промоторы генов cat, kan, bla, trp и тандемно расположенных промоторов генов kan и trp). Уровни синтеза последовательностей HВsAg (нативного и в составе химерных белков) составляли в зависимости от используемых конструкций векторов от 100 до 100000 молекул на клетку.
1.4. СОЗДАНИЕ ДОСТАТОЧНО УДОБНЫХ И ПО ВОЗМОЖНОСТИ
УНИВЕРСАЛЬНЫХ ВЕКТОРОВ ДЛЯ ЦЕЛЕВОЙ ДОСТАВКИ ГЕНОВ В
КЛЕТКИ И ТКАНИ ОРГАНИЗМА
Важным моментом при конструировании ДНК-вакцин является проблема целенаправленной доставки генов в необходимые клетки и защиты вводимых ДНК от действия нуклеаз крови. В результате экспериментальной работы были созданы разнообразные конструкции, позволяющие доставлять целевые гены в клетки-мишени.
Одной из подобных конструкций является модель молекулярного вектора для доставки генов в такие клетки, как лимфоциты и кераноциты. В качестве модельного был использован ген, кодирующий гибридный белок: фактор некроза опухолей-альфа - интерферон-гамма. В центре вектора находится интактная плазмидная ДНК, содержащая доставляемый ген, а на поверхности располагаются антитела к клеткам-мишеням. Конъюгат полиглюкина со спермидином и антителами применяется для связи компонентов (положительно заряженный спермидин обеспечивает связывание конъюгата с плазмидной ДНК). Описанный молекулярный вектор позволяет целенаправленно доставлять гены в клетки-мишени, сводя до минимума их попадание в другие виды клеток, защищать доставляемые гены от нуклеаз крови и использовать положительно заряженный комплекс спермидин-полиглюкин в качестве стимулятора проникновения ДНК в клетки.
В настоящее время также создана векторная модель для доставки в клетки костного мозга гена, кодирующего гранулоцитарный колониестимулирующий фактор человека (чГ-КСФ). Данный белок относится к семейству гемопоэтических факторов роста и является одним из физиологических регуляторов, специфически и высокоэффективно стимулирующих пролиферацию и дифференцировку гемопоэтических предшественников нейтрофилов. чГ-КСФ увеличивает продолжительность жизни клеток костного мозга, усиливает функциональную активность зрелых нейтрофилов. Созданный вектор представляет собой многослойную конструкцию. "Центральным ядром" конструкции является плазмида pGGF8, содержащая ген чГ-КСФ. Ее окружает полисахаридная оболочка, которая состоит из полиглюкина и спермидина. Внешний белковый слой содержит смесь сывороточного альбумина и белка доставки - трансферина. Эффективность описанной векторной модели была доказана опытным путем.